
Structure of the Whole Cytosolic Region of ATP-Dependent Protease FtsH
11
Molecular Cell 22, 575–585, June 9, 2006 ª2006 Elsevier Inc. DOI 10.1016/j.molcel.2006.04.020 Structure of the Whole Cytosolic Region of ATP-Dependent Protease FtsH Ryoji Suno, 1,4 Hajime Niwa, 2,4 Daisuke Tsuchiya, 3,5 Xiaodong Zhang, 2 Masasuke Yoshida, 1, * and Kosuke Morikawa 3,6, * 1 Chemical Resources Laboratory Tokyo Institute of Technology 4259 Nagatsuta Yokohama 226-8503 Japan 2 Centre for Structural Biology Faculty of Life Sciences Imperial College London London SW7 2AZ United Kingdom 3 Department of Structural Biology Biomolecular Engineering Research Institute 6-2-3 Furuedai, Suita Osaka 565-0874 Japan Summary An ATP-dependent protease, FtsH, digests misas- sembled membrane proteins in order to maintain mem- brane integrity and digests short-lived soluble pro- teins in order to control their cellular regulation. This enzyme has an N-terminal transmembrane segment and a C-terminal cytosolic region consisting of an AAA + ATPase domain and a protease domain. Here we present two crystal structures: the protease domain and the whole cytosolic region. The cytosolic region fully retains an ATP-dependent protease activity and adopts a three-fold-symmetric hexameric structure. The protease domains displayed a six-fold symmetry, while the AAA + domains, each containing ADP, alter- nate two orientations relative to the protease domain, making ‘‘open’’ and ‘‘closed’’ interdomain contacts. Apparently, ATPase is active only in the closed form, and protease operates in the open form. The protease catalytic sites are accessible only through a tunnel fol- lowing from the AAA + domain of the adjacent subunit, raising a possibility of translocation of polypeptide substrate to the protease sites through this tunnel. Introduction ATP-dependent proteases are involved in various cellu- lar processes including cell division, cell differentiation, signal transduction, and stress response (Ogura and Wil- kinson, 2001; Hanson and Whiteheart, 2005). Escherichia coli contains five kinds of ATP-dependent proteases, that is, ClpAP, ClpXP, HslUV, Lon, and FtsH. The first three have a composite structure of the ATPase complex (ClpA, ClpX, and HslU) and the protease complex (ClpP and HslU). In the case of the last two, catalytic sites for ATPase and protease exist in different domains in the same subunit. In all cases, the subunits or domains re- sponsible for ATPase activity share common motifs of AAA + (ATPases associated with diverse cellular activi- ties) proteins. Among them, FtsH is unique because it is the only protease that is anchored to the cytoplasmic membrane (Tomoyasu et al., 1993), and it is the only es- sential protease for bacterial growth (Jayasekera et al., 2000). Some closely related homologs of FtsH have been found in eukaryotic mitochondria and chloroplast (Langer, 2000), and mutation of a human FtsH ortholog causes a neurodegenerative disease (Casari et al., 1998). FtsH digests not only misassembled subunits of membrane protein complexes for their quality control but also some short-lived cytosolic regulatory proteins for cellular regulation (Ito and Akiyama, 2005). FtsH is comprised of the N-terminal transmembrane segment and the C-terminal cytosolic region, which con- sists of the AAA + and protease domains (Tomoyasu et al., 1993). The AAA + domain is responsible for ATPase activity and has the conserved Walker A, Walker B, and second region of homology (SRH) motifs. The arginine residue at the C terminus of the SRH motif, the so-called ‘‘arginine finger,’’ is crucial for ATP hydrolysis (Ogura et al., 2004). The crystal structures of the isolated AAA + domain of FtsH from E. coli (Krzywda et al., 2002) and Thermus thermophilus (Niwa et al., 2002) suggested that native FtsH would form the hexameric ring structure. Analysis of the primary sequence revealed that FtsH carries a zinc binding sequence (HEXXH, where X is any residue) within the protease domain. Activity stimulation by zinc ion (Tomoyasu et al., 1995) indicates that FtsH could be categorized into the family of zinc metallopro- teases termed as ‘‘zincin.’’ Normally, zinc ion in zincins is coordinated by three residues: the two histidine resi- dues in HEXXH and either a histidine (metzincin) or a glu- tamic acid (gluzincin) as the third residue (Hooper, 1994). In general, ATP-dependent proteases degrade sub- strate proteins in a processive manner without releasing large intermediate polypeptides. Typically, hexameric AAA + complex can unfold a substrate protein and trans- locate a polypeptide through the central pore into the chamber of the protease complex where protease cata- lytic sites exist at the surface of the chamber wall (Wang et al., 1997; Bochtler et al., 1997). In this ‘‘pore/chamber’’ scheme, the central pore is formed by conserved resi- dues (MFVG in the case of FtsH), of which the aromatic residue has been shown to be particularly important for substrate translocation (Hinnerwisch et al., 2005; Park et al., 2005). The translocation is coupled with the ATPase cycle, accompanying large conformational change of the AAA + complex. Crystal structures of HslU indicated that four different conformational states of the AAA + complex exist in the catalytic cycle (Wang et al., 2001; Sousa et al., 2002; Burton et al., 2005). In *Correspondence: [email protected] (M.Y.); morikako@ protein.osaka-u.ac.jp (K.M.) 4 These authors contributed equally to this work. 5 Present address: Institute for Advanced Biosciences, Keio Uni- versity, 403-1 Nipponkoku, Daihoji, Tsuruoka, Yamagata 997-0017, Japan. 6 Present address: Institute for Protein Research, Osaka University, 6-2-3 Furuedai, Suita, Osaka 565-0874, Japan.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Structure of the Whole Cytosolic Region of ATP-Dependent Protease FtsH

Molecular Cell 22, 575–585, June 9, 2006 ª2006 Elsevier Inc. DOI 10.1016/j.molcel.2006.04.020
Structure of the Whole Cytosolic Regionof ATP-Dependent Protease FtsH
Ryoji Suno,1,4 Hajime Niwa,2,4 Daisuke Tsuchiya,3,5
Xiaodong Zhang,2 Masasuke Yoshida,1,*
and Kosuke Morikawa3,6,*1Chemical Resources LaboratoryTokyo Institute of Technology4259 NagatsutaYokohama 226-8503Japan2Centre for Structural BiologyFaculty of Life SciencesImperial College LondonLondon SW7 2AZUnited Kingdom3Department of Structural BiologyBiomolecular Engineering Research Institute6-2-3 Furuedai, SuitaOsaka 565-0874Japan
Summary
An ATP-dependent protease, FtsH, digests misas-
sembled membrane proteins in order to maintain mem-brane integrity and digests short-lived soluble pro-
teins in order to control their cellular regulation. Thisenzyme has an N-terminal transmembrane segment
and a C-terminal cytosolic region consisting of anAAA+ ATPase domain and a protease domain. Here we
present two crystal structures: the protease domainand the whole cytosolic region. The cytosolic region
fully retains an ATP-dependent protease activity andadopts a three-fold-symmetric hexameric structure.
The protease domains displayed a six-fold symmetry,while the AAA+ domains, each containing ADP, alter-
nate two orientations relative to the protease domain,making ‘‘open’’ and ‘‘closed’’ interdomain contacts.
Apparently, ATPase is active only in the closed form,and protease operates in the open form. The protease
catalytic sites are accessible only through a tunnel fol-lowing from the AAA+ domain of the adjacent subunit,
raising a possibility of translocation of polypeptidesubstrate to the protease sites through this tunnel.
Introduction
ATP-dependent proteases are involved in various cellu-lar processes including cell division, cell differentiation,signal transduction, and stress response (Ogura and Wil-kinson, 2001; Hanson and Whiteheart, 2005). Escherichia
*Correspondence: [email protected] (M.Y.); morikako@
protein.osaka-u.ac.jp (K.M.)4 These authors contributed equally to this work.5 Present address: Institute for Advanced Biosciences, Keio Uni-
versity, 403-1 Nipponkoku, Daihoji, Tsuruoka, Yamagata 997-0017,
Japan.6 Present address: Institute for Protein Research, Osaka University,
6-2-3 Furuedai, Suita, Osaka 565-0874, Japan.
coli contains five kinds of ATP-dependent proteases,that is, ClpAP, ClpXP, HslUV, Lon, and FtsH. The firstthree have a composite structure of the ATPase complex(ClpA, ClpX, and HslU) and the protease complex (ClpPand HslU). In the case of the last two, catalytic sites forATPase and protease exist in different domains in thesame subunit. In all cases, the subunits or domains re-sponsible for ATPase activity share common motifs ofAAA+ (ATPases associated with diverse cellular activi-ties) proteins. Among them, FtsH is unique because itis the only protease that is anchored to the cytoplasmicmembrane (Tomoyasu et al., 1993), and it is the only es-sential protease for bacterial growth (Jayasekera et al.,2000). Some closely related homologs of FtsH havebeen found in eukaryotic mitochondria and chloroplast(Langer, 2000), and mutation of a human FtsH orthologcauses a neurodegenerative disease (Casari et al.,1998). FtsH digests not only misassembled subunits ofmembrane protein complexes for their quality controlbut also some short-lived cytosolic regulatory proteinsfor cellular regulation (Ito and Akiyama, 2005).
FtsH is comprised of the N-terminal transmembranesegment and the C-terminal cytosolic region, which con-sists of the AAA+ and protease domains (Tomoyasuet al., 1993). The AAA+ domain is responsible for ATPaseactivity and has the conserved Walker A, Walker B, andsecond region of homology (SRH) motifs. The arginineresidue at the C terminus of the SRH motif, the so-called‘‘arginine finger,’’ is crucial for ATP hydrolysis (Oguraet al., 2004). The crystal structures of the isolated AAA+
domain of FtsH from E. coli (Krzywda et al., 2002) andThermus thermophilus (Niwa et al., 2002) suggestedthat native FtsH would form the hexameric ring structure.Analysis of the primary sequence revealed that FtsHcarries a zinc binding sequence (HEXXH, where X is anyresidue) within the protease domain. Activity stimulationby zinc ion (Tomoyasu et al., 1995) indicates that FtsHcould be categorized into the family of zinc metallopro-teases termed as ‘‘zincin.’’ Normally, zinc ion in zincinsis coordinated by three residues: the two histidine resi-dues in HEXXH and either a histidine (metzincin) or a glu-tamic acid (gluzincin) as the third residue (Hooper, 1994).
In general, ATP-dependent proteases degrade sub-strate proteins in a processive manner without releasinglarge intermediate polypeptides. Typically, hexamericAAA+ complex can unfold a substrate protein and trans-locate a polypeptide through the central pore into thechamber of the protease complex where protease cata-lytic sites exist at the surface of the chamber wall (Wanget al., 1997; Bochtler et al., 1997). In this ‘‘pore/chamber’’scheme, the central pore is formed by conserved resi-dues (MFVG in the case of FtsH), of which the aromaticresidue has been shown to be particularly important forsubstrate translocation (Hinnerwisch et al., 2005; Parket al., 2005). The translocation is coupled with theATPase cycle, accompanying large conformationalchange of the AAA+ complex. Crystal structures ofHslU indicated that four different conformational statesof the AAA+ complex exist in the catalytic cycle (Wanget al., 2001; Sousa et al., 2002; Burton et al., 2005). In

Molecular Cell576
Table 1. X-Ray Data Collection, Experimental Phasing, and Structure Refinement
Space group
A. aeolicus pFtsH T. thermophilus sFtsH (G399L)
P63 P3121
Unit cell
a = b = 116.8 [A], c = 63.5 [A] a = b = 146.2 [A], c = 349.1 [A]
HgCl2 SeMet Native HgCl2 SeMet
Data Collection
Beamline BL38B1 BL38B1 BL41XU BL38B1 BL38B1
Wavelength, A 1.00000 1.05000 1.00524 1.00714 0.970000 0.985000 0.977527 0.977719 1.00000 1.00000 0.97900
Resolution, A 100–2.79
(2.90–2.79)
100–3.50
(3.70–3.50)
100–3.90 100–6.00 100–4.50
(4.11–3.90) (6.33–6.00) (4.74–4.50)
Rmerge 0.094
(0.391)
0.092
(0.439)
0.114
(0.630)
0.110
(0.600)
0.108
(0.110)
0.107
(0.109)
0.104
(0.378)
0.104
(0.108)
0.077
(0.547)
0.101
(0.446)
0.141
(0.529)
Completeness,
%
99.3
(93.3)
99.6
(96.5)
99.7
(97.1)
99.6
(96.9)
100
(100)
100
(100)
100
(100)
100
(100)
99.9
(100)
100
(100)
99.8
(99.3)
I/s(I) 42.9 (9.9) 41.5 (8.4) 33.7 (5.9) 34.7 (5.9) 5.7 (2.4) 5.3 (2.0) 5.4 (1.9) 5.4 (1.9) 6.2 (1.4) 5.0 (1.6) 3.3 (1.3)
Experimental
Phasing
Number of sites 2 8 - 12 55
Figure of merit 0.51 0.17/0.21a
Refinement
Resolution, A 20.0–2.79
(2.86–2.79)b15.0–3.90
(3.99–3.90)c
Rwork 0.251
(0.305)
0.299
(0.334)
Rfreed 0.299
(0.347)
0.342
(0.377)
Rmsd in bond
length, A
0.010 0.016
Rmsd in bond
angle, º
1.36 1.69
Numbers in parentheses indicate the values for the highest resolution shell.a Values for centric/acentric reflections.b The data for the HgCl2 derivative (wavelength, 1.00000 A) were used.c The data for the native crystal were used.d Calculated with 7% of diffraction data not used in refinement.
contrast to the typical ATP-dependent proteases, themechanism of FtsH is poorly understood, mostly dueto insufficient structural information.
Here, we present the crystal structure of the proteasedomain of FtsH at 2.79 A resolution, as well as the struc-ture of the whole cytosolic region of FtsH at 3.9 A resolu-tion. The former exhibits a unique fold and forms a stablesix-fold-symmetric hexamer, which exhibits proteaseactivity in an ATP-independent manner. The latter retainsATP-dependent protease activity and forms a three-fold-symmetric hexamer with the alternately aligned ‘‘open’’and ‘‘closed’’ subunits. Interestingly, in this sFtsH struc-ture, a substrate polypeptide can reach the active prote-ase catalytic sites through a tunnel leading from theAAA+ domain of the adjacent subunit, but not from thecentral axial region. This raises a possibility of directdelivery of a polypeptide through this tunnel.
Results
Robust Hexameric Structure of FtsH ProteaseDomain
To crystallize the protease domain, we prepared asegment from Aquifex aeolicus, one of the most hyper-thermophilic bacteria available to date (Deckert et al.,1998). The sequence analyses among various FtsHprotease domains and limited proteolysis experiments
suggested that a fragment, covering the entire proteasedomain (residues 405–634, pFtsH), could be adequatefor crystallization (Figures 1A and 1B). Gel filtration anal-ysis revealed that the isolated protease domain formeda stable hexamer (see Figure S1 in the SupplementalData available with this article online). The correspond-ing region of pFtsH of T. thermophilus FtsH (residues404–593) also forms a hexamer (data not shown). Re-markably, pFtsH of A. aeolicus bears protease activity(Figure S2), in contrast to inactive protease activity ofthe isolated protease components of HslV, ClpP, andLon (Sauer et al., 2004; Rasulova et al., 1998).
Crystallographic analysis of pFtsH from A. aeolicus at2.79 A resolution (Table 1) allowed us to trace the mainchain from T406 to K607, except for some residues inthe N and C termini, residues from R443 to D457 (be-tween b6 and b7, corresponding to the lid helix a14 asmentioned later), and residues from V525 to M533 (be-tween a16 and a17, corresponding to the b hairpin b9–b10 as mentioned later) (Figures 1B and 1D). Each sub-unit adopts the shape of a flat, triangular wedge madeof seven tightly packed a helices (a13–a19) and sixb strands (b6–b11). Two subunits in the asymmetric unitare nearly identical (rmsd, 0.430 A for all Ca positions),and the three-fold rotational symmetry derived fromthe crystallographic 63 screw axis relates these subunitsto generate a hexamer with pseudo six-fold symmetry

Crystal Structure of FtsH577
Figure 1. FtsH Protease Domain
(A) Schematic drawing of the constructions
for FtsH. Loop regions between rigid do-
mains are colored red, and the large and
small subdomains of the AAA+ domain are
shown in cyan and green, respectively.
(B) Multiple sequence alignment of the prote-
ase domain. T. thermophilus FtsH (accession
code AB032368) aligned with A. aeolicus
FtsH (AE000714-12), E. coli FtsH (U01376),
and yeast Yme1 (L14616). Cylinders and ar-
rows indicate the secondary structure ele-
ments (a helices and b strands, respectively)
of T. thermophilus FtsH. All secondary struc-
tures in A. aeolicus pFtsH are the same as
those in T. thermophilus FtsH, except the dis-
ordered regions. The conserved glycine
(G399) in the loop between the AAA+ and pro-
tease domains is highlighted in white. Limited
proteolysis of A. aeolicus FtsH revealed that
trypsin preferably digested the loop region
(red arrowhead). The lid helix (helix 14) ap-
pears to be kinked at the conserved glycine
residue in green. The zinc binding motif
(HEXXH) and the third coordinating aspartic
acid residue are shown in red.
(C) Hexameric structure of A. aeolicus pFtsH
from the top (left) and side (right) views. In
the side view, the protease structure is super-
posed onto T. thermophilus sFtsH (transpar-
ent molecular surface).
(D) A stereopair diagram of the pFtsH mono-
mer. The catalytic residues are shown in stick
models. The Zn binding site (gray sphere)
was actually occupied by an Hg atom in the
crystal. The dotted lines indicate the disor-
dered regions.
(E) Stereo view of the catalytic residues in the
pFtsH (orange) superposed onto the corre-
sponding residues of thermolysin (cyan) and
tricorn interacting factor F3 (green).
(Figure 1C). The hexamer plate is clasped by six a17 he-lices on the bottom. Moreover, the large interface be-tween the two adjacent subunits (1203 A2), includinga hydrogen-bonding network between b7 of one subunitand b8 of the adjacent subunit, contributes to the stabil-ity of the plate. These structural features suggest thatthis hexamer should be robust and hardly influencedby the conformational change of the AAA+ domains.
DALI-server search (Holm and Sander, 1993) showedthat this architecture has no significant similarity toeven other zinc metalloproteases, displaying a uniqueprotein fold.
Structure of a Protease Catalytic SiteA catalytic site of the protease domain lies at a ratherperipheral position of the hexamer plate and is located

Molecular Cell578
Figure 2. Characterization of sFtsH
(A) The wild-type (thin line) and G399L mutant
(thick line) analyzed by gel filtration chroma-
tography (protein concentration, 7.5 mg/ml).
The molecular weight marker was purchased
from Amersham Biosciences.
(B) Protease activities of sFtsH (black arrow)
in the presence (lanes 1–6) or absence (lanes
7–12) of ATP, and the full-length FtsH (open
arrow) in the presence of ATP (lanes 13–18).
Substrate digestion (a-casein; asterisk) for
each reaction was monitored at the incubation
times of 0, 2, 4, 8, 16, and 32 min, respectively.
(C) ATP-dependent protease activities of the
wild-type (lanes 1–5) and the G399L mutant
(lanes 6–10) in the presence of ATP. These
enzymes (black arrow) were incubated with
a-casein (asterisk) for 0, 5, 10, 15, and 30
min, respectively. For reference, a-casein
(lane 11) and the G399L mutant (lane 12)
were loaded separately.
in the cleft composed of the three helices a13, a15, anda16, as well as b6 (Figures 1C and 1D). The structural el-ement (lid helix) covering the protease catalytic site inthe whole cytosolic region, as mentioned later, was dis-ordered in pFtsH structure. Probably for this reason,a polypeptide substrate could reach the catalytic siteof pFtsH and be degraded. The crystal structure con-tained mercury atoms at the predicted positions of thezinc ion at the catalytic sites (HEXXH). FtsH is a memberof the zinc-dependent zincin family in which zinc ion isliganded by three residues, although the third zinc-coor-dinating residue was yet unclear in FtsH. Comparison ofthe catalytic site of pFtsH with those of thermolysin (Pro-tein Data Bank, 1KEI) and tricorn interacting factor F3(1Z1W) (Figure 1E) reveals that, despite the limited struc-tural similarity among them, relative positions of the cat-alytic residues are remarkably conserved, suggestingthat FtsH hydrolyzes peptide bonds in the same manneras other zincins. This comparison also shows that thecatalytic metal ion in FtsH is coordinated with the imid-azole rings of H418 and H422 in the HEXXH motif ina13 and with the carboxyl group of D496 in a16. Amongknown zincins, FtsH protease is a distinct example thatadopts an aspartic acid, instead of a glutamic acid, forthe third coordinating residue. Saikawa et al. (2002) re-ported that E479 of E. coli FtsH (E480 in A. aeolicusFtsH) might be the third ligand for the metal ion. How-ever, this residue is too far from the zinc. Instead, theside chain of E480 makes a hydrogen bond with theside chain of H418 to stabilize indirectly the coordinationof the zinc.
Crystallization of Active sFtsH
We prepared a soluble, cytosolic region of FtsH (resi-dues 126–624, sFtsH) from T. thermophilus. The isolatedsFtsH formed an oligomeric structure (Figure 2A).
Although Akiyama and Ito (2000) reported that the trans-membrane segment was indispensable for both theATPase and protease activities, sFtsH showed nearlythe same ATP-dependent protease activity as that ofthe full-length FtsH (Figure 2B). The oligomeric structurebecomes unstable when the protein concentration islow in the high salt concentration buffer (data notshown). The sFtsH could be crystallized (P3121; a =b = 147.0 [A], c = 697.3 [A]), but the crystal diffractedonly to w7 A resolution. In an attempt to reduce themain chain flexibility, we found that a mutant in whichG399 in the linker region (between the AAA+ and prote-ase domain) was replaced by leucine yielded a differentcrystal form (Table 1) with better diffraction to 3.5 A res-olution. The function of FtsH was not impaired by thismutation (Figures 2A and 2C). Furthermore, ATPase ac-tivity of the G399L mutant was stimulated by the addi-tion of a substrate protein (a-casein) (data not shown),as observed for the full-length FtsH (Akiyama and Ito,2000; Asahara et al., 2000). Using this mutant, we solvedthe crystal structure of sFtsH to 3.9 A resolution.
Alternate Arrangement of Open and Closed Subunits
in sFtsHThe crystal of sFtsH contains one hexameric molecule inthe asymmetric unit. The hexameric structure (Figure 3A)adopts a flat-cylinder-like shape (135 A diameter, 65 Aheight) bearing a noncrystallographic three-fold rota-tional symmetry. As shown in the middle panel ofFigure 3A, the cylinder is divided into two disks. Thelower disk (the protease domain) forms the six-fold-sym-metric structure that is almost identical to that of pFtsH(Figure 3A, the bottom view), but with the apparently va-cant metal binding sites. The upper disk is composed ofsix AAA+ domains, and each AAA+ domain contains ADP.The folding topology of the AAA+ domain of sFtsH

Crystal Structure of FtsH579
Figure 3. Structure of the Hexameric sFtsH
(A) Entire structure of the ADP bound sFtsH hexamer. Cylinders and arrows represent a helices and b strands, respectively. An open (green) and
a closed (orange) subunit are surrounded by broken lines. These two structures are used for the following illustrations with the same coloring.
Bound ADP molecules and the arginine finger (R313) are depicted in blue and red space-filling models, respectively.
(B) Structural difference between the open and closed subunits, shown by the superposition at their protease domains (cyan). The mobile regions
in the protease domain are highlighted by the secondary-structure elements with the corresponding subunit color.
(C) Stereo view of the AAA+ domains in the open and closed subunits, superposed onto the structures observed in the nucleotide-free crystals
(Niwa et al., 2002) with P65 (red) and C2221 (gray) symmetries. The AAA+-domain structure complexed with AMPPNP is basically the same as that
for the nucleotide-free state in P65 symmetry, due to the same crystal packing (Niwa et al., 2002), and thus is not drawn for clarity. The blue stick
model represents the bound ADP molecule at the closed subunit.
resembles those of other AAA+ proteins (Hanson andWhiteheart, 2005). Remarkably, orientation of the AAA+
domain relative to the protease domain is significantlydifferent between adjacent subunits (Figure 3B). Onesubunit (orange in Figure 3B) exhibits extensive contactsbetween AAA+ and protease domains and thus is desig-nated as a closed subunit. In the adjacent subunit desig-nated as an open subunit (green in Figure 3B), the AAA+
domain rotates by 31º away from the protease domain.Consequently, the architecture of FtsH is a trimer of
dimers. The overall structure does not appear to be seri-ously affected by crystal packing, because each of thesix subunits makes distinct contacts within the crystallattice, due to the noncrystallographic three-fold axisirrelevant to any crystallographic ones (Figure S3).
Subdomains in the AAA+ Domain
The AAA+ domain consists of a large subdomain (resi-dues 143–324) and a small subdomain (residues 325–400). The ATPase catalytic site resides at the interface

Molecular Cell580
between the large and small subdomains. The large sub-domains assemble around the three-fold axis to formthe core of the disk, and the small subdomains are lo-cated on the rim of the disk. The small subdomain ofthe closed subunit tightly interacts with the large subdo-main of the adjacent open subunit (Figure 3A, top view).On the other hand, the small subdomain of the opensubunit protrudes into the solvent and appears to alloweasy access of nucleotide to the catalytic site from themedium. Therefore, exchange of nucleotide (ADP re-lease and ATP binding) in the ATPase cycle would occurin the open form but not the closed form. Despite the dif-ferent subdomain contacts, the structures of AAA+ do-mains by themselves in the open and closed subunits(Figure 3C) are essentially identical (rmsd value, 1.01 Afor all Ca positions of the AAA+ domain). This structuralsimilarity is partly due to the fact that all of the six sub-units harbor ADP. We previously reported the crystalstructures of the AAA+ domain free from nucleotideand complexed with AMPPNP (Niwa et al., 2002). Thesuperposition of these structures onto the presentstructure (Figure 3C) shows obvious differences in therelative orientation between the large and small subdo-mains. This difference is likely to reflect the nucleotide-dependent conformational changes, as observed ina number of other AAA+ proteins, where nucleotidebinding induces a more compact organization betweenthe two subdomains.
Catalytic Environments in the Open and Closed
Subunits
The arginine finger plays a critical role in ATP hydrolysisfor many ATPases (Ogura et al., 2004): the guanidinogroup of arginine of one subunit extends and interactswith g-phosphate oxygen of ATP bound to the adjacentsubunit. In the present hexameric structure, R313 is theresidue corresponding to the arginine finger, and R316,which is an alternative candidate for the arginine finger(Karata et al., 1999), is too far from the nucleotide(Figure 4A). In the closed subunit, the oxygen atom ofthe b-phosphate of the bound ADP is spatially separatedby 7.6 A from the Nh atom of R313 of the adjacent opensubunit (Figure 4A, lower). This distance would be closeenough for R313 to interact with the oxygen atom of theg-phosphate of ATP, when bound to the active site. Incontrast, the corresponding distance in the open subunitis 24.8 A, which is too far for ATP hydrolysis (Figure 4A,upper). Thus, only the ATPase catalytic site in the closedsubunit should be active. Therefore, in FtsH, ATPase-active and -inactive subunits are alternately arranged inone hexamer.
Likewise, the structure around the protease active siteshows a significant difference between the two sub-units. The interface between the AAA+ and protease do-mains in sFtsH structure is primarily formed by two mo-bile regions: a lid helix a14 (P442–E455) and a b hairpinb9–b10 (Y519–Q534). In the closed form, the AAA+ do-main pushes down the lid helix toward the protease do-main, inducing the kink at the position of G448 (Figures3B and 4B) that provides the conformational flexibility.As a result, the kinked helix occupies the space for thepolypeptide substrate just above the protease catalyticsite (Figure 4B, lower). On the contrary, in the open form,the lid helix becomes straight and moves away from the
active site (Figure 4B, upper). Then, the active site is ca-pable of accommodating the substrate. This structuraldifference suggests a possibility that protease may beactive only in the open form.
Crosslinking between Two Domains Inactivates
sFtsHOur sFtsH structure is indicative of dynamic open-closemotion in the hexameric structure coupled with theATPase cycle. To examine this contention, we carriedout a disulfide crosslinking experiment using the full-length T. thermophilus FtsH containing the transmem-brane region. Two residues, A310 (AAA+ domain) andL527 (protease domain), were simultaneously replacedwith cysteines. These two residues are close enoughto make a disulfide bond only in the closed subunit (Fig-ure 5A). The cysteine residue in the wild-type T. thermo-philus FtsH (C250) was replaced with alanine to avoidunwanted crosslinking, as done by Makyio et al.(2002). When the C250A/A310C/L527C triple mutantwas oxidized with CuCl2, a significant fraction, but notall, of the protein exhibited intrasubunit disulfide bond-ing (Figure 5B). The result is consistent with the viewthat the designed crosslinking is possible only for theclosed subunit (Figure 5A). The ATP-dependent proteol-ysis was lost for the oxidized mutant sFtsH but was re-covered by the treatment with a reducing reagent nearlyto the level of the wild-type (Figure 5C). Thus, crosslink-ing experiments strongly suggest not only the occur-rence of this structure for native FtsH but also the re-quirement of open-close motion for the FtsH function.
Discussion
Previous studies assumed that FtsH would take a six-fold-symmetric architecture (Niwa et al., 2002; Ya-mada-Inagawa et al., 2003). However, our experimentalresults strongly indicate that the three-fold-symmetricstructure (Figure 3A) plays crucial roles in the catalyticprocess. Based on this idea, we will discuss the struc-tural mechanism for the ATP-dependent processivedegradation by FtsH.
A Potential Role of N302Our previous study showed that N302 in the SRH motifcould discriminate the bound nucleotide (Niwa et al.,2002). Its side chain interacts with the g-phosphate ofAMPPNP, an ATP analog, via a water molecule. By con-trast, the side chain moves away from the active site inthe nucleotide-free state. In fact, a mutational analysissuggested that N302 plays a critical role in the ATP-de-pendent proteolytic function (Karata et al., 1999). Nota-bly, this residue is located at the interface between theAAA+ and protease domains. In the open subunit wheremost likely the ATPase is inactive and the nucleotide ex-change would occur, N302 occupies a proximal positionto the lid helix (Figure 4B, upper). Although we could notconclusively define the local structure due to the resolu-tion limit, N302 may play a modulatory role in the open-close transition. When ATP binds to the catalytic site, itsg-phosphate attracts N302 and alters structure aroundthe interface between the AAA+ and protease domains.This conformational change would eventually contributeto triggering the closing motion.
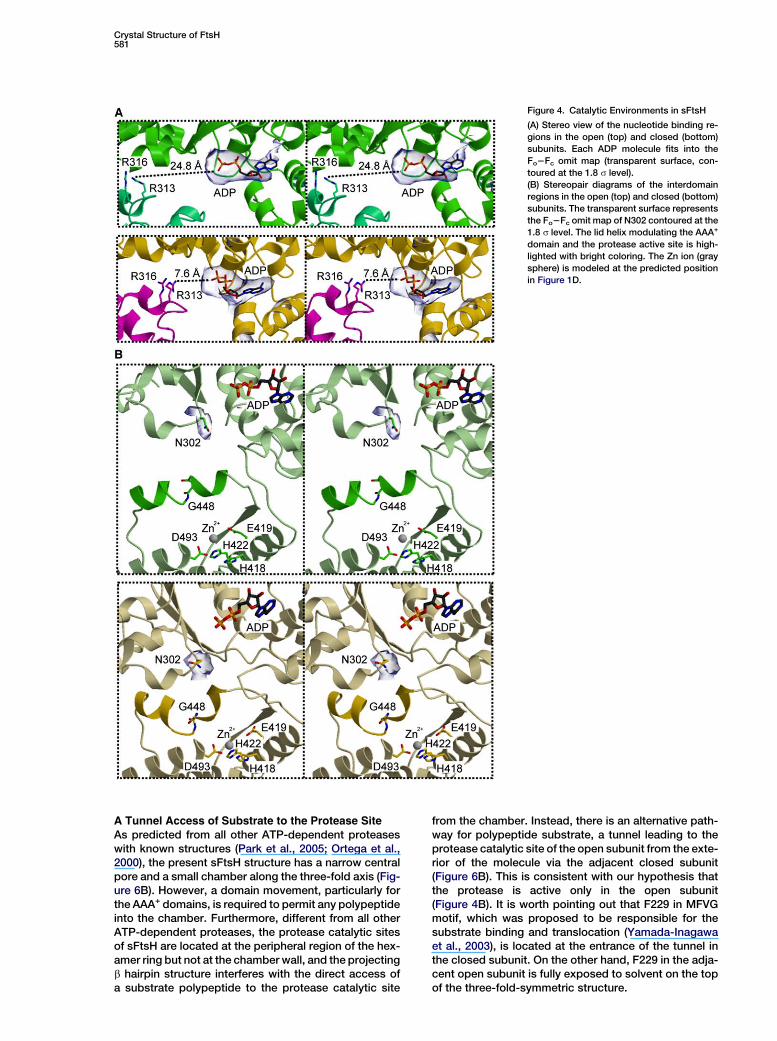
Crystal Structure of FtsH581
Figure 4. Catalytic Environments in sFtsH
(A) Stereo view of the nucleotide binding re-
gions in the open (top) and closed (bottom)
subunits. Each ADP molecule fits into the
Fo2Fc omit map (transparent surface, con-
toured at the 1.8 s level).
(B) Stereopair diagrams of the interdomain
regions in the open (top) and closed (bottom)
subunits. The transparent surface represents
the Fo2Fc omit map of N302 contoured at the
1.8 s level. The lid helix modulating the AAA+
domain and the protease active site is high-
lighted with bright coloring. The Zn ion (gray
sphere) is modeled at the predicted position
in Figure 1D.
A Tunnel Access of Substrate to the Protease Site
As predicted from all other ATP-dependent proteaseswith known structures (Park et al., 2005; Ortega et al.,2000), the present sFtsH structure has a narrow centralpore and a small chamber along the three-fold axis (Fig-ure 6B). However, a domain movement, particularly forthe AAA+ domains, is required to permit any polypeptideinto the chamber. Furthermore, different from all otherATP-dependent proteases, the protease catalytic sitesof sFtsH are located at the peripheral region of the hex-amer ring but not at the chamber wall, and the projectingb hairpin structure interferes with the direct access ofa substrate polypeptide to the protease catalytic site
from the chamber. Instead, there is an alternative path-way for polypeptide substrate, a tunnel leading to theprotease catalytic site of the open subunit from the exte-rior of the molecule via the adjacent closed subunit(Figure 6B). This is consistent with our hypothesis thatthe protease is active only in the open subunit(Figure 4B). It is worth pointing out that F229 in MFVGmotif, which was proposed to be responsible for thesubstrate binding and translocation (Yamada-Inagawaet al., 2003), is located at the entrance of the tunnel inthe closed subunit. On the other hand, F229 in the adja-cent open subunit is fully exposed to solvent on the topof the three-fold-symmetric structure.

Molecular Cell582
Figure 5. Disulfide Crosslinking Experiment
(A) Close-up view of A310 and L527 in the
open (left) and closed (right) subunits, from
the same direction as in Figure 3B, where
these residues are similarly illustrated with
spheres. The broken lines indicate the Ca dis-
tances between the two residues.
(B) Intrasubunit disulfide crosslinking of the
C250A/A310C/L527C triple mutant. The pro-
tein was prepared by gel filtration chromatog-
raphy under a denaturing condition in the ab-
sence of DTT (see Experimental Procedures).
The major peak was eluted at the corre-
sponding position to monomeric FtsH and
analyzed by SDS-PAGE under reducing
(lane 1) and nonreducing (lane 2) conditions.
In the latter, a new band (black arrow) ap-
peared in the higher molecular weight range
than the predicted position for monomeric
FtsH (open arrow).
(C) ATP-dependent protease activity of the
triple mutant in the presence and the absence
of DTT, as monitored by the fluorescence in-
crease ascribed to release of small proteo-
lytic fragments of BODIPY-labeled casein.
ATP was added at 100 s (open arrow) to start
the reaction.
Models for ATP-Dependent Processive Degradationby sFtsH
Combined with the data supporting the ATP-dependentopen-close domain arrangements, the present struc-tural features, including the intriguing tunnel and the cat-alytic environments, allow us to postulate the followingmodel for the ATPase cycle (Figure 6A). (1) The opensubunit exchanges its bound ADP with ATP, and theclosing motion is triggered. (2) The closing motion ac-companies rearrangement of the small subdomain,which leads to the conformational transition of the wholeFtsH molecule. (3) Upon completion of the closing mo-tion, the ATPase catalytic site becomes activated andATP is hydrolyzed. (4) The closed subunit can return tothe open form without releasing ADP, and the wholeFtsH structure also returns to the original state. TheATP binding to other open subunits in the same hexamermight help this transition. This ATPase cycle assumesthat the conformational change is apparently carriedout by a pair of subunits, although there remains an ex-treme possibility that only a single subunit can drive thestructural transition, as postulated for ClpXP (Martinet al., 2005).
The synchronized open-close motions of subunits inFtsH would drive the translocation of a substrate poly-peptide to the protease catalytic sites. As mentioned,translocation through the central pore/chamber couldoccur only if a drastic conformational change takes placeand the b hairpin completely dislocates. Although wecould not exclude a possibility of the conventionalpore/chamber mechanism in FtsH, the functional do-main arrangement in the present structure, particularlyfor the arginine finger (Figure 4A), necessitates an alter-native pathway for polypeptide substrate (Figure 6B).In accordance with the above ATPase cycle, (1) a sub-strate polypeptide binds to the MFVG region of the
open subunit. (2) It should be sent into the tunnel leadingto the protease catalytic site of the adjacent subunit, (3)in which it is degraded, and (4) the MFVG region relievesthe grasp on polypeptide and returns to the original po-sition. If F229 of the open subunit binds the substratepolypeptide and undergoes the open-close transition,it could drag polypeptide as long as w35 A (Figure 6B).This length corresponds to 12–13 residues of an ex-tended polypeptide that roughly coincides with our pre-vious estimation on the average size of the product oligo-peptides (Asahara et al., 2000). The prediction from thisinteresting possibility, for example, multiple substratebinding to a single FtsH molecule, should be worth test-ing by experiments.
Experimental Procedures
Plasmid Constructions, Overproductions, and Purifications
Plasmids for the recombinant FtsH proteins fused with glutathione
S-transferase (GST) at its C terminus were constructed using
pGEX-NdeI (Niwa et al., 2002). All mutations were introduced by
the PCR-based method with QuikChange Site-Directed Mutagene-
sis Kit (Stratagene) and were verified by DNA sequencing.
The fusion proteins were overproduced in E. coli strain BL21(DE3).
The cells carrying the plasmid proliferated at 37ºC in LB or 2 3 YT
medium. Addition of isopropyl-b-D-thiogalactopyranoside (0.5
mM) induced expression of the recombinant proteins. Those derived
from T. thermophilus were purified as described previously (Asahara
et al., 2000). The cells overproducing A. aeolicus pFtsH fused with
GST were lysed by sonication in the buffer containing 50 mM Tris-
HCl (pH 8.0), 1 mM ethylenediaminetetraacetic acid (EDTA), and 1
mM dithiothreitol (DTT). Unexpectedly, in the cell lysate the recombi-
nant protein carried only seven residues above the N terminus of
pFtsH. It would cleave the linker between GST and pFtsH because
the cleavage did not occur in a protease-inactive mutant, E419Q
(data not shown). After heat treatment (80ºC, 10 min), the superna-
tant was loaded onto a HiTrap Q column (Amersham Biosciences),
and the eluate containing pFtsH was dialyzed into 10 mM Tris-HCl
(pH 8.0). The selenomethionine-labeled proteins were produced in

Crystal Structure of FtsH583
the methionine auxotroph E. coli strain B384(DE3), cultured in methi-
onine assay medium (Difco) with 40 mg/ml L-selenomethionine
(Sigma). The labeled protein was purified under essentially the
same condition as that for the native protein.
Gel Filtration Analysis and Enzymatic Assays
The molecular size was estimated with Superdex 200 gel filtration
column (Amersham Biosciences) equilibrated with a buffer contain-
ing 0.1 M Tris-HCl (pH 8.0), 150 mM KCl, and 1 mM DTT. To analyze
intrasubunit disulfide crosslinking, the protein pretreated with 10
mM DTT was oxidized with 10 mM CuCl2, and the reaction was termi-
nated with 2 mM N-ethylmaleimide and 5 mM EDTA. The oxidized
sample was denatured by 10 min exposure to the 8 M urea buffer
containing 10 mM Tris-HCl (pH 8.0), 150 mM KCl, 10% glycerol,
and 1% 1-O-n-octyl-b-D-glucopyranoside, and subsequently
loaded onto Superdex 200 column equilibrated with the 4 M urea
Figure 6. The ATPase Cycle and Putative Polypeptide Translocation
Pathway
(A) Schematic diagram of the ATPase cycle in the hexameric FtsH.
The cycle for the blue subunit is described in the text, where step
one corresponds to the upper left. T, D, and R represent ATP,
ADP, and the arginine finger, respectively. The catalytic cycle for
the red subunit is supplemented, because the binding of ATP might
help transition from the lower right to the lower left.
(B) A diagram representing a cross-section of three-fold-symmetric
FtsH, including the three-fold axis (black line) and two protease ac-
tive sites (blue arrowheads). The axial area of the AAA+ domains is
closed by three D273 residues of the closed subunits (green). The
b hairpin structures composed of b9 and b10 (cyan) interfere with
the direct access to the open protease active site from the central
chamber. F229 (red) of the closed subunit is located at the entrance
of the putative polypeptide translocation tunnel (bold arrow in dark
magenta), whereas the residue in the open subunit is fully exposed
on the top of the AAA+-domain hexamer. The pathway connecting
the two F229 residues (light magenta) appears to reflect the unit
length of polypeptide substrate, which one open-to-close transition
can send into the protease active site.
buffer. The peak position for the monomeric FtsH was confirmed
by analyzing the denatured protein in the presence of 10 mM DTT.
The ATPase activity was determined by the amount of released inor-
ganic phosphate detected by malachite green. The activity for the 1
mg/ml enzyme was measured in the reaction buffer (for full-length
FtsH, 10 mM Tris-HCl [pH 8.0], 50 mM KCl, 10 mM MgCl2, 25 mM
Zn acetate, 0.3% 1-O-n-octyl-b-D-glucopyranoside; for sFtsH, 10
mM Tris-HCl [pH 8.0], 10 mM MgCl2, 10 mM Zn acetate, 1 mM
DTT). After preincubation at 55ºC for 5 min, the reaction started by
adding ATP (5 mM) and ended by adding perchloric acid on ice.
Protease activity was monitored by degradation of a-casein (Sigma)
analyzed by SDS-PAGE. The enzyme (1 mg/ml) was incubated with
the substrate (final 0.5 mg/ml) at 55ºC in the reaction buffer in the
presence or absence of 4 mM ATP. The reaction was terminated
by adding an equal volume of 23 sample buffer for SDS-PAGE.
For detecting the full-length FtsH activity with a fluorescent probe,
the enzyme (0.04 mg/ml) was preincubated with BODIPY-casein
(0.025 mg/ml; Molecular Probes) in the reaction buffer for 5 min at
55ºC. Then, addition of ATP (5 mM) triggered the substrate diges-
tion, which was monitored by fluorescence at 530 nm excited by
485 nm light.
X-Ray Crystallography
Crystals of A. aeolicus pFtsH were grown in a buffer solution con-
taining 0.1 M MES (pH 6.0) and 5% PEG8K, at 20ºC, using the va-
por-diffusion technique. The crystals diffracted at least 2.8 A resolu-
tion with synchrotron radiation at SPring-8 (BL38B1; Hyogo, Japan)
when they were flash frozen at 100 K in the buffer containing 0.1 M
MES (pH 6.0), 2.0 M (NH4)2SO4, and 25% glycerol. The diffraction im-
ages were processed with DENZO/SCALEPACK (Otwinowski and
Minor, 1997) or MOSFLM (Leslie, 1999)/SCALA (CCP4, 1994). The
primary phases were estimated by the multiple anomalous disper-
sion method using both the mercury derivative and the selenome-
thionine-labeled protein crystal. Density modification performed
solvent flattening, histogram mapping, and noncrystallographic
symmetry (NCS) averaging to improve the experimental electron
density map. The constructed atomic model was refined by using
CNS (Brunger et al., 1998) for refinement of the atomic coordinates
and by using REFMAC (Murshudov et al., 1997) for the TLS refine-
ment in the end.
T. thermophilus sFtsH was crystallized by the vapor diffusion
method with the buffer containing 0.1 M Tris-HCl (pH 7.2), 1%
PEG8K, 0.2 mM Zn acetate, 5 mM MgCl2, and 2.5 mM ADP at
20ºC. The diffraction data were collected at the beamline BL41XU
at SPring-8 at 100 K. The primary phases were initially estimated
by the molecular replacement (MR) and later recalculated by the
multiple isomorphous replacement supplemented with the anoma-
lous dispersion (MIRAS). The initial MR trial succeeded if the pFtsH
structure was used as the probe but failed if the structure of the
AAA+ domain (Niwa et al., 2002) was used. However, the density
modification with the 5 A resolution data barely enabled us to locate
the AAA+ domains. Validity of the phases was confirmed by the dif-
ference Fourier map between the native and the Hg-derivative. The
improved phases also visualized 55 of the 72 selenium atoms in the
anomalous difference Fourier map. These heavy atom positions al-
low us to perform the MIRAS phasing (Figure S4). To avoid the model
bias, the MR phases were not supplemented to the experimental
ones. Density modification, including the NCS averaging, solvent
flattening, and histogram mapping, yielded an interpretable map
with the 3.9 A resolution data. The molecular structure was built
with a guide of the structures of the AAA+ domain (Niwa et al.,
2002) and A. aeolicus pFtsH (the present study). The atomic model,
containing the whole sFtsH region except for some N- and C-termi-
nal residues and the residues 264–271 in the open subunits, was re-
fined by using REFMAC (Murshudov et al., 1997), whose TLS param-
eterization and the tight NCS restraint were critical to reduce the R
factor. Presence of ADP in the crystal (Figures 3A and 4A) was con-
firmed by the HPLC analysis (data not shown), even if crystals ob-
tained with commercially available ATPgS (Roche), in which a small
fraction of ADP contaminates, were used. Although the crystalliza-
tion buffer contained the Zn ion, it was invisible in the electron den-
sity map, probably due to the limited resolution.
All data collection and the refinement statistics are summarized in
Table 1.

Molecular Cell584
Supplemental Data
Supplemental Data include four figures and can be found with
this article online at http://www.molecule.org/cgi/content/full/22/5/
575/DC1/.
Acknowledgments
We are grateful to Drs. K. Miura, N. Shimizu, and K. Hasegawa at
SPring-8 for help during the synchrotron beam time. We also thank
Dr. T. Oyama for assistance in X-ray data collection. This research
was partially supported by a research grant endorsed by the New
Energy and Industrial Technology Development Organization
(NEDO), and the ‘‘National Project on Protein Structural and Func-
tional Analyses’’ of the Ministry of Education, Culture, Sports, Sci-
ence, and Technology of Japan.
Received: January 1, 2006
Revised: February 17, 2006
Accepted: April 18, 2006
Published: June 8, 2006
References
Akiyama, Y., and Ito, K. (2000). Roles of multimerization and mem-
brane association in the proteolytic functions of FtsH (HflB). EMBO
J. 19, 3888–3895.
Asahara, Y., Atsuta, K., Motohashi, K., Taguchi, H., Yohda, M., and
Yoshida, M. (2000). FtsH recognizes proteins with unfolded structure
and hydrolyzes the carboxyl side of hydrophobic residues. J. Bio-
chem. (Tokyo) 127, 931–937.
Bochtler, M., Ditzel, L., Groll, M., and Huber, R. (1997). Crystal struc-
ture of heat shock locus V (HslV) from Escherichia coli. Proc. Natl.
Acad. Sci. USA 94, 6070–6074.
Brunger, A.T., Adams, P.D., Clore, G.M., DeLano, W.L., Gros, P.,
Grosse-Kunstleve, R.W., Jiang, J.S., Kuszewski, J., Nilges, M.,
Pannu, N.S., et al. (1998). Crystallography & NMR system: a new
software suite for macromolecular structure determination. Acta.
Crystallogr. D. Biol. Crystallogr. 54, 905–921.
Burton, R.E., Baker, T.A., and Sauer, R.T. (2005). Nucleotide-depen-
dent substrate recognition by the AAA+ HslUV protease. Nat. Struct.
Mol. Biol. 12, 245–251.
Casari, G., De Fusco, M., Ciarmatori, S., Zeviani, M., Mora, M., Fer-
nandez, P., De Michele, G., Filla, A., Cocozza, S., Marconi, R.,
et al. (1998). Spastic paraplegia and OXPHOS impairment caused
by mutations in paraplegin, a nuclear-encoded mitochondrial metal-
loprotease. Cell 93, 973–983.
CCP4 (Collaborative Computational Project, Number 4) (1994). The
CCP4 suite: programs for protein crystallography. Acta Crystallogr.
D. Biol. Crystallogr. 50, 760–763.
Deckert, G., Warren, P.V., Gaasterland, T., Young, W.G., Lenox, A.L.,
Graham, D.E., Overbeek, R., Snead, M.A., Keller, M., Aujay, M., et al.
(1998). The complete genome of the hyperthermophilic bacterium
Aquifex aeolicus. Nature 392, 353–358.
Hanson, P.I., and Whiteheart, S.W. (2005). AAA+ proteins: have en-
gine, will work. Nat. Rev. Mol. Cell Biol. 6, 519–529.
Hinnerwisch, J., Fenton, W.A., Furtak, K.J., Farr, G.W., and Horwich,
A.L. (2005). Loops in the central channel of ClpA chaperone medi-
ate protein binding, unfolding, and translocation. Cell 121, 1029–
1041.
Holm, L., and Sander, C. (1993). Protein structure comparison by
alignment of distance matrices. J. Mol. Biol. 233, 123–138.
Hooper, N.M. (1994). Families of zinc metalloproteases. FEBS Lett.
354, 1–6.
Ito, K., and Akiyama, Y. (2005). Cellular functions, mechanism of ac-
tion, and regulation of FtsH protease. Annu. Rev. Microbiol. 59, 211–
231.
Jayasekera, M.M., Foltin, S.K., Olson, E.R., and Holler, T.P. (2000).
Escherichia coli requires the protease activity of FtsH for growth.
Arch. Biochem. Biophys. 380, 103–107.
Karata, K., Inagawa, T., Wilkinson, A.J., Tatsuta, T., and Ogura, T.
(1999). Dissecting the role of a conserved motif (the second region
of homology) in the AAA family of ATPases. Site-directed mutagen-
esis of the ATP-dependent protease FtsH. J. Biol. Chem. 274,
26225–26232.
Krzywda, S., Brzozowski, A.M., Verma, C., Karata, K., Ogura, T., and
Wilkinson, A.J. (2002). The crystal structure of the AAA domain of the
ATP-dependent protease FtsH of Escherichia coli at 1.5 A resolution.
Structure 10, 1073–1083.
Langer, T. (2000). AAA proteases: cellular machines for degrading
membrane proteins. Trends Biochem. Sci. 25, 247–251.
Leslie, A.G.W. (1999). Integration of macromolecular diffraction
data. Acta. Crystallogr. D. Biol. Crystallogr. 55, 1696–1702.
Makyio, H., Niwa, H., Motohashi, K., Taguchi, H., and Yoshida, M.
(2002). Stabilization of FtsH-unfolded protein complex by binding
of ATP and blocking of protease. Biochem. Biophys. Res. Commun.
296, 8–12.
Martin, A., Baker, T.A., and Sauer, R.T. (2005). Rebuilt AAA+ motors
reveal operating principles for ATP-fuelled machines. Nature 437,
1115–1120.
Murshudov, G.N., Vagin, A.A., and Dodson, E.J. (1997). Refinement
of macromolecular structures by the maximum likelihood method.
Acta. Crystallogr. D. Biol. Crystallogr. 53, 240–255.
Niwa, H., Tsuchiya, D., Makyio, H., Yoshida, M., and Morikawa, K.
(2002). Hexameric ring structure of the ATPase domain of the mem-
brane-integrated metalloprotease FtsH from Thermus thermophilus
HB8. Structure 10, 1415–1423.
Ogura, T., and Wilkinson, A.J. (2001). AAA+ superfamily ATPases:
common structure-diverse function. Genes Cells 6, 575–597.
Ogura, T., Whiteheart, S.W., and Wilkinson, A.J. (2004). Conserved
arginine residues implicated in ATP hydrolysis, nucleotide-sensing,
and inter-subunit interactions in AAA and AAA+ ATPases. J. Struct.
Biol. 146, 106–112.
Ortega, J., Singh, S.K., Ishikawa, T., Maurizi, M.R., and Steven, A.C.
(2000). Visualization of substrate binding and translocation by the
ATP-dependent protease, ClpXP. Mol. Cell 6, 1515–1521.
Otwinowski, Z., and Minor, W. (1997). Processing of X-ray diffrac-
tion data collected in oscillation mode. Methods Enzymol. 276,
307–326.
Park, E., Rho, Y.M., Koh, O.J., Ahn, S.W., Seong, I.S., Song, J.J.,
Bang, O., Seol, J.H., Wang, J., Eom, S.H., and Chung, C.H. (2005).
Role of the GYVG pore motif of HslU ATPase in protein unfolding
and translocation for degradation by HslV peptidase. J. Biol.
Chem. 280, 22892–22898.
Rasulova, F.S., Dergousova, N.I., Starkova, N.N., Melnikov, E.E.,
Rumsh, L.D., Ginodman, L.M., and Rotanova, T.V. (1998). The iso-
lated proteolytic domain of Escherichia coli ATP-dependent prote-
ase Lon exhibits the peptidase activity. FEBS Lett. 432, 179–181.
Saikawa, N., Ito, K., and Akiyama, Y. (2002). Identification of glutamic
acid 479 as the gluzincin coordinator of zinc in FtsH (HflB). Biochem-
istry 41, 1861–1868.
Sauer, R.T., Bolon, D.N., Burton, B.M., Burton, R.E., Flynn, J.M.,
Grant, R.A., Hersch, G.L., Joshi, S.A., Kenniston, J.A., Levchenko,
I., et al. (2004). Sculpting the proteome with AAA(+) proteases and
disassembly machines. Cell 119, 9–18.
Sousa, M.C., Kessler, B.M., Overkleeft, H.S., and McKay, D.B.
(2002). Crystal structure of HslUV complexed with a vinyl sulfone in-
hibitor: corroboration of a proposed mechanism of allosteric activa-
tion of HslV by HslU. J. Mol. Biol. 318, 779–785.
Tomoyasu, T., Yuki, T., Morimura, S., Mori, H., Yamanaka, K., Niki,
H., Hiraga, S., and Ogura, T. (1993). The Escherichia coli FtsH protein
is a prokaryotic member of a protein family of putative ATPases in-
volved in membrane functions, cell cycle control, and gene expres-
sion. J. Bacteriol. 175, 1344–1351.
Tomoyasu, T., Gamer, J., Bukau, B., Kanemori, M., Mori, H., Rutman,
A.J., Oppenheim, A.B., Yura, T., Yamanaka, K., Niki, H., et al. (1995).
Escherichia coli FtsH is a membrane-bound, ATP-dependent prote-
ase which degrades the heat-shock transcription factor s32. EMBO
J. 14, 2551–2560.
Wang, J., Hartling, J.A., and Flanagan, J.M. (1997). The structure of
ClpP at 2.3 A resolution suggests a model for ATP-dependent prote-
olysis. Cell 91, 447–456.

Crystal Structure of FtsH585
Wang, J., Song, J.J., Franklin, M.C., Kamtekar, S., Im, Y.J., Rho,
S.H., Seong, I.S., Lee, C.S., Chung, C.H., and Eom, S.H. (2001). Crys-
tal structures of the HslVU peptidase–ATPase complex reveal an
ATP-dependent proteolysis mechanism. Structure 9, 177–184.
Yamada-Inagawa, T., Okuno, T., Karata, K., Yamanaka, K., and
Ogura, T. (2003). Conserved pore residues in the AAA protease
FtsH are important for proteolysis and its coupling to ATP hydroly-
sis. J. Biol. Chem. 278, 50182–50187.
Accession Numbers
Atomic coordinates for pFtsH and sFtsH have been deposited with
the RCSB Protein Data Bank under accession codes 2DI4 (A. aeoli-
cus pFtsH) and 2DHR (T. thermophilus sFtsH).




















