
Cytosolic Triacylglycerol Biosynthetic Pathway in Oilseeds. Molecular Cloning and Expression of...
11
Cytosolic Triacylglycerol Biosynthetic Pathway in Oilseeds. Molecular Cloning and Expression of Peanut Cytosolic Diacylglycerol Acyltransferase 1[W] Saikat Saha, Balaji Enugutti, Sona Rajakumari, and Ram Rajasekharan* Department of Biochemistry, Indian Institute of Science, Bangalore 560012, India Triacylglycerols (TAGs) are the most important storage form of energy for eukaryotic cells. TAG biosynthetic activity was identified in the cytosolic fraction of developing peanut (Arachis hypogaea) cotyledons. This activity was NaF insensitive and acyl-coenzyme A (CoA) dependent. Acyl-CoA:diacylglycerol acyltransferase (DGAT) catalyzes the final step in TAG biosyn- thesis that acylates diacylglycerol to TAG. Soluble DGAT was identified from immature peanuts and purified by conventional column chromatographic procedures. The enzyme has a molecular mass of 41 6 1.0 kD. Based on the partial peptide sequence, a degenerate probe was used to obtain the full-length cDNA. The isolated gene shared less than 10% identity with the previously identified DGAT1 and 2 families, but has 13% identity with the bacterial bifunctional wax ester/ DGAT. To differentiate the unrelated families, we designate the peanut gene as AhDGAT. Expression of peanut cDNA in Escherichia coli resulted in the formation of labeled TAG and wax ester from [ 14 C]acetate. The recombinant E. coli showed high levels of DGATactivity but no wax ester synthase activity. TAGs were localized in transformed cells with Nile blue A and oil red O staining. The recombinant and native DGAT was specific for 1,2-diacylglycerol and did not utilize hexadecanol, glycerol-3-phosphate, monoacylglycerol, lysophosphatidic acid, and lysophosphatidylcholine. Oleoyl-CoA was the preferred acyl donor as compared to palmitoyl- and stearoyl-CoAs. These data suggest that the cytosol is one of the sites for TAG biosynthesis in oilseeds. The identified pathway may present opportunities of bioengineering oil-yielding plants for increased oil production. Oils and fats are glycerol triesters of fatty acids (triacylglycerols [TAGs]) and are mainly derived from plant and animal sources, respectively. Vegetable oils are the major source of edible lipids, accounting for more than 75% of the total lipids consumed across the world (Broun et al., 1999). The global demand for plant oils has intensified our efforts to genetically modify the organism to enhance oil yield. De novo biosynthesis of TAG has been shown to occur by the sequential acylation of glycerol-3-P (Kennedy, 1961; Ohlrogge et al., 1991). The first en- zyme in this pathway, glycerol-3-P acyltransferase, cat- alyzes the formation of lysophosphatidic acid (LPA) that can be acylated to give phosphatidic acid (PA) by LPA acyltransferase. PA is the precursor for diacyl- glycerol (DAG) and anionic phospholipids. PA phos- phatase catalyzes the dephosphorylation of PA to form DAG, which is an immediate precursor for TAG, phosphatidylcholine, and phosphatidylethanolamine. The possible routes of TAG formation are (1) acyl-CoA- dependent acylation of DAG by DAG acyltransferases (DGATs), (2) acyl-CoA-independent transacylation us- ing two molecules of DAG by DAG transacylase (Stobart et al., 1997), and (3) acyl-CoA-independent acylation using phospholipids as acyl donors and DAG as an acyl acceptor catalyzed by phospholipid:DGAT (Dahlqvist et al., 2000). DGAT1 encoding DGAT (EC 2.3.1.20) was identified in mice (Cases et al., 1998), and the gene is homologous to acyl-CoA:cholesterol acyltransferase (ACAT). The isolated gene shared significant sequence homology with an Arabidopsis (Arabidopsis thaliana) expressed sequence tag (EST), and the sequence information was used to isolate Arabidopsis (Hobbs et al., 1999; Zou et al., 1999; Bouvier-Nave et al., 2000) and rapeseed (Brassica napus) DGAT1 (Nykiforuk et al., 2002). ACAT- unrelated DGAT2 gene has been isolated from Saccha- romyces cerevisiae (Oelkers et al., 2000) and Mortierella ramanniana (Lardizabal et al., 2001). Recently, a novel bifunctional DGAT/wax ester synthase was described in Acinetobacter calcoaceticus (Kalscheuer and Steinbuchel, 2003). All the acyltransferases in these pathways are membrane bound. Here, we describe the identification and molecular cloning of a cytosolic DGAT from developing peanut (Arachis hypogaea) cotyledons. The gene encoding sol- uble DGAT was expressed in Escherichia coli and char- acterized. This study provides evidence for the presence of an additional TAG biosynthetic pathway in plants. 1 This work was supported by the Department of Biotechnology, Government of India, New Delhi. * Corresponding author; e-mail [email protected]; fax 91–80–23600814. The author responsible for distribution of materials integral to the findings presented in this article in accordance with the policy described in the Instructions for Authors (www.plantphysiol.org) is: Ram Rajasekharan ([email protected]). [W] The online version of this article contains Web-only data. Article, publication date, and citation information can be found at www.plantphysiol.org/cgi/doi/10.1104/pp.106.082198. Plant Physiology, August 2006, Vol. 141, pp. 1533–1543, www.plantphysiol.org Ó 2006 American Society of Plant Biologists 1533 www.plant.org on August 12, 2014 - Published by www.plantphysiol.org Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of Cytosolic Triacylglycerol Biosynthetic Pathway in Oilseeds. Molecular Cloning and Expression of...

Cytosolic Triacylglycerol Biosynthetic Pathway inOilseeds. Molecular Cloning and Expression of PeanutCytosolic Diacylglycerol Acyltransferase1[W]
Saikat Saha, Balaji Enugutti, Sona Rajakumari, and Ram Rajasekharan*
Department of Biochemistry, Indian Institute of Science, Bangalore 560012, India
Triacylglycerols (TAGs) are the most important storage form of energy for eukaryotic cells. TAG biosynthetic activity wasidentified in the cytosolic fraction of developing peanut (Arachis hypogaea) cotyledons. This activity was NaF insensitive andacyl-coenzyme A (CoA) dependent. Acyl-CoA:diacylglycerol acyltransferase (DGAT) catalyzes the final step in TAG biosyn-thesis that acylates diacylglycerol to TAG. Soluble DGAT was identified from immature peanuts and purified byconventional column chromatographic procedures. The enzyme has a molecular mass of 41 6 1.0 kD. Based on the partialpeptide sequence, a degenerate probe was used to obtain the full-length cDNA. The isolated gene shared less than 10%identity with the previously identified DGAT1 and 2 families, but has 13% identity with the bacterial bifunctional wax ester/DGAT. To differentiate the unrelated families, we designate the peanut gene as AhDGAT. Expression of peanut cDNA inEscherichia coli resulted in the formation of labeled TAG and wax ester from [14C]acetate. The recombinant E. coli showedhigh levels of DGAT activity but no wax ester synthase activity. TAGs were localized in transformed cells with Nile blue Aand oil red O staining. The recombinant and native DGAT was specific for 1,2-diacylglycerol and did not utilizehexadecanol, glycerol-3-phosphate, monoacylglycerol, lysophosphatidic acid, and lysophosphatidylcholine. Oleoyl-CoAwas the preferred acyl donor as compared to palmitoyl- and stearoyl-CoAs. These data suggest that the cytosol is one of thesites for TAG biosynthesis in oilseeds. The identified pathway may present opportunities of bioengineering oil-yieldingplants for increased oil production.
Oils and fats are glycerol triesters of fatty acids(triacylglycerols [TAGs]) and are mainly derived fromplant and animal sources, respectively. Vegetable oilsare the major source of edible lipids, accounting formore than 75% of the total lipids consumed across theworld (Broun et al., 1999). The global demand for plantoils has intensified our efforts to genetically modify theorganism to enhance oil yield.De novo biosynthesis of TAG has been shown
to occur by the sequential acylation of glycerol-3-P(Kennedy, 1961; Ohlrogge et al., 1991). The first en-zyme in this pathway, glycerol-3-P acyltransferase, cat-alyzes the formation of lysophosphatidic acid (LPA)that can be acylated to give phosphatidic acid (PA) byLPA acyltransferase. PA is the precursor for diacyl-glycerol (DAG) and anionic phospholipids. PA phos-phatase catalyzes the dephosphorylation of PA to formDAG, which is an immediate precursor for TAG,phosphatidylcholine, and phosphatidylethanolamine.
The possible routes of TAG formation are (1) acyl-CoA-dependent acylation of DAG by DAG acyltransferases(DGATs), (2) acyl-CoA-independent transacylation us-ing twomolecules ofDAGbyDAG transacylase (Stobartet al., 1997), and (3) acyl-CoA-independent acylationusing phospholipids as acyl donors and DAG asan acyl acceptor catalyzed by phospholipid:DGAT(Dahlqvist et al., 2000).
DGAT1 encoding DGAT (EC 2.3.1.20) was identifiedinmice (Cases et al., 1998), and the gene is homologousto acyl-CoA:cholesterol acyltransferase (ACAT). Theisolated gene shared significant sequence homologywith an Arabidopsis (Arabidopsis thaliana) expressedsequence tag (EST), and the sequence information wasused to isolate Arabidopsis (Hobbs et al., 1999; Zouet al., 1999; Bouvier-Nave et al., 2000) and rapeseed(Brassica napus)DGAT1 (Nykiforuk et al., 2002). ACAT-unrelated DGAT2 gene has been isolated from Saccha-romyces cerevisiae (Oelkers et al., 2000) and Mortierellaramanniana (Lardizabal et al., 2001). Recently, a novelbifunctionalDGAT/waxester synthasewasdescribed inAcinetobacter calcoaceticus (Kalscheuer and Steinbuchel,2003). All the acyltransferases in these pathways aremembrane bound.
Here, we describe the identification and molecularcloning of a cytosolic DGAT from developing peanut(Arachis hypogaea) cotyledons. The gene encoding sol-uble DGATwas expressed in Escherichia coli and char-acterized. This study provides evidence for thepresence of an additional TAG biosynthetic pathwayin plants.
1 This work was supported by the Department of Biotechnology,Government of India, New Delhi.
* Corresponding author; e-mail [email protected]; fax91–80–23600814.
The author responsible for distribution of materials integral to thefindings presented in this article in accordance with the policydescribed in the Instructions for Authors (www.plantphysiol.org) is:Ram Rajasekharan ([email protected]).
[W] The online version of this article contains Web-only data.Article, publication date, and citation information can be found at
www.plantphysiol.org/cgi/doi/10.1104/pp.106.082198.
Plant Physiology, August 2006, Vol. 141, pp. 1533–1543, www.plantphysiol.org � 2006 American Society of Plant Biologists 1533 www.plant.org on August 12, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.

RESULTS
Identification of a Cytosolic TAG Biosynthetic Pathwayin Developing Peanut Cotyledons
Subcellular distribution of DGAT in fresh cotyle-dons was obtained by differential centrifugation. TheTAG biosynthetic capacity was found to be high (70%)in membrane fraction, and 31% of the total activitywas found in cytosolic fraction (Fig. 1A). The effect of[14C]oleoyl-CoA concentration on the biosynthesis ofTAGwas studied in cytosolic andmembrane fractions.The maximum amount of TAG formation was ob-served with 10 mM oleoyl-CoA in both the fractions(Fig. 1B). Addition of 1,2-DAG in cytosolic fraction didnot alter the rate and the pattern of incorporation of[14C]oleoyl-CoA into TAG.
TAG can be synthesized either by dephosphoryla-tion of PA followed by acylation of DAG or by thesuccessive acylation of monoacylglycerol (MAG). Tofind out the contribution of each pathway to the totalTAG pool, peanut cytosolic fraction was treated with20 mM NaF to inhibit PA phosphatase activity, and theincorporation of [14C]oleoyl-CoA into TAG and itsbiosynthetic intermediates was studied in the presenceof NaF. As shown in Figure 1C, there was about 28% to34% decrease in the incorporation of [14C]oleoyl-CoAinto TAG, suggesting the presence of a PA dephos-phorylation-independent pathway. To rule out the hy-drolysis as well as transacylation of fatty acids eitherfrom TAG or from phosphatidylcholine, the cytosolicfraction was incubated either with [3H]phosphatidyl-choline or with [3H]TAG followed by separation oflipids by thin-layer chromatography (TLC). There wasno formation of labeled mono-, di-, and triacylglyc-erols from labeled phosphatidylcholine and therewas no mono- and diacylglycerol formed from labeledTAG, indicating the absence of transacylation and hy-drolysis reactions in the cytosol (data not shown).
Purification of Cytosolic DGAT from DevelopingPeanut Cotyledons
A summary of purification of DGAT is presented inTable I. Solid ammonium sulfate was added to bringcytosolic fraction to 1 M and then loaded onto an octyl-Sepharose column. The columnwas eluted with 1 to 0 M
linear-reversed gradient of ammonium sulfate. This wasan efficient step resulting in a 62-fold purification ofDGAT. The active fractions from the octyl-Sepharosewere loaded onto a blue-Sepharose column and elutedwith a linear NaCl gradient. The activity was eluted be-tween0.35and0.4MNaCl.The recoveryofDGATactivityfrom the blue-Sepharose column was nearly 48% of thatapplied. The pooled active fractions were applied to apreparative Superdex 75 column. TheDGATactivitywaseluted between 27 to 31 fractions. The active fractionswere pooled and applied to a heparin-agarose column asthe final step. As summarized in Table I, an overallpurification of 1,724-fold was obtained, and the specificactivity of acyltransferase was 7.1 nmol min21 mg21.
Figure 1. Generation of TAG. A, Fresh developing peanut cotyledons(50 g) were used for obtaining various subcellular fractions by differ-ential centrifugation. TAG formation was monitored using [14C]oleoyl-CoA in the absence of exogenously added acyl acceptors. Values aremean 6 SD of three independent determinations. Homo, Homogenate;Plas, plastidal fraction; Mito, mitochondrial fraction; Cyto, cytosolicfraction; Memb, membrane fraction. B, Acylation of [14C]oleoyl-CoAinto TAG was carried out for 15 min at 30�C in the presence of 50 mM
1,2-DAG in cytosolic (28 to 36 mg; d) and in membrane (40 to 45 mg;s) fractions of developing peanut cotyledons. Each point is the averageof two independent experiments. C, Time-dependent formation of TAG.Incorporation of [14C]oleoyl-CoAwas performedwith cytosolic fractionof developing peanut cotyledon into TAG in the absence of added acylacceptors. The formation of TAG was monitored either in the presence(s) or in the absence (d) of 20 mM NaF. Each point is mean6 SD of fourindependent determinations.
Saha et al.
1534 Plant Physiol. Vol. 141, 2006 www.plant.org on August 12, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.

The preparation from each step of purification wasresolved on a 12% (w/v) SDS-PAGE, which showedtwo major proteins (41 and 37 kD) upon silver staining(Fig. 2A). The final preparation was unstable. Despiteour effort, we could not purify the enzyme to homo-geneity. We therefore electroblotted the final enzymepreparation onto a nitrocellulose membrane and per-formed immunoblot analysis with peptide antibodiesraised against the oleaginous yeast cytosolic DGAT.The immunoblot revealed a strong cross-reactivitywith only 41 6 1.0 kD protein (Fig. 2B). It is possiblethat the internal amino acid sequence of Rhodotorulaglutinis DGAT (Gangar et al., 2001) may have a signif-icant similarity with soluble peanut DGAT, and thiscould be the reason for peptide antibody cross-reactivity.The reaction products formed at each step of purifica-
tion were analyzed on silica-TLC and autoradiographed(Fig. 2C). When the cytosol was incubated with labeledoleoyl-CoA in the presence of DAG, TAG and otherbiosynthetic intermediates were formed, suggesting thepresence of many different acylation activities.After electrophoresis proteins were electroblotted
onto a polyvinylidene difluoride membrane; the poly-peptide corresponding to molecular size of 41 kD wasexcised and treated with cyanogen bromide to cleaveMet residues. Themajor peptide was taken for sequenc-ing and the sequence (NH2-GNKCKKSGSIALLQE-FERVVGA) did not match with any known sequencein the database. This was suitable for designing a de-generate primer for gene isolation.
Cloning of a Gene Encoding DGAT
To obtain the full-length cDNA clone encoding pu-tativeDGAT, a seed-specific cDNAlibrarywas screenedwith the oligonucleotide primer 5#-GGCAACAAGTG-CAAGAAAAGCGGAAGTATCGCTTTATTGCAGGA-ATTCGAGAGGGTCGTGGGGGCC-3# (66-mer), basedon the internal amino acid sequence of the protein(KCKKSGSIALLQEFERVVGA). The nucleotide probewas radiolabeled at the 3# end using TdTand [g-32P]ATP.A large number of positive clones were isolated fromthe screen, and the plasmid from clones containing thelongest inserts (approximately 1.6 kb) was sequenced
on both strands using an automated sequencer. Thesequence (1,592 bp) was submitted to GenBank withthe identification as AhDGAT (accession no. AY875644).Analysis of the nucleotide sequence revealed an openreading frame of 345 amino acids with translationinitiating at the 360ATG362 codon (Supplemental Fig. 1).
Table I. Purification of cytosolic DGAT from developingpeanut cotyledons
The results are the summary of a typical purification of DGAT. Frozenimmature peanut cotyledons (100 g) were used for preparing thecytosol. Enzyme activity measurements and the purification steps aredescribed in ‘‘Materials and Methods.’’
Fraction Protein Total Activity Specific Activity Purification
mg nmol min21 pmol min21 mg21 -fold
Cytosol 1,604 6.58 4.1 1Octyl-Sepharose 19.42 4.93 253.7 62Blue-Sepharose 1.39 2.37 1,707.4 416Superdex 75 0.47 1.45 3,073.8 750Heparin-agarose 0.16 1.13 7,069.5 1,724
Figure 2. Purification of cytosolic DGAT from developing peanutcotyledons. A, SDS-PAGE protein profile. Samples from each purifica-tion step were separated on a 12% (w/v) SDS-PAGE and stained withsilver nitrate. Lane 1 is standard molecular mass marker, and lanes 2 to 6correspond to the pooled active fractions from steps 1 to 5 (Table I). B,Western-blot analysis. Samples from each step of purification wereseparated by 12% (w/v) SDS-PAGE and electroblotted onto a nitrocel-lulose membrane, followed by probing with anti-DGATantibodies fromR. glutinis. Lane 1, Prestained proteinmolecular mass marker. Lanes 2 to6 correspond to the pooled active fractions from steps 1 to 5 (Table I). C,Autoradiography of the reaction products formed at each step of thepurification. DGAT assays were performed using [14C]oleoyl-CoA andDAG for 15 min. The products formed were separated on a TLC usingpetroleum ether:diethyl ether:acetic acid (70:30:1, v/v). Lanes 1 to 5correspond to the active fractions from steps 1 to 5 of purification Table I.
Cytosolic Triacylglycerol Biosynthesis in Oilseeds
Plant Physiol. Vol. 141, 2006 1535 www.plant.org on August 12, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.

The 239-bp 3#-untranslated region contained anoncanonical polyadenylation signal (AATAT) nearthe poly(A) tail, indicating that the isolated clonerepresented full-length cDNA. The calculated molec-ular mass of the open reading frame was 37.5 kD.When the sequence of internal amino acid was aligned
with the deduced amino acid sequence, there was acomplete match, confirming that the cloned cDNAencoded DGAT. Hydropathy plot of the predictedprotein indicated the absence of any transmembranedomains (Fig. 3A). Absence of signal sequence alsoconfirms its cytosolic nature.
Figure 3. DGAT gene sequence analyses. A, Hydropathy plot of cytosolic peanut DGAT. No transmembrane domain waspredicted. B, Alignment of AhDGAT and homolog genes. Sequence alignment was performed using the http://www.ch.embnet.org/software/BOX_form.html program. AhDGAT, peanut DGAT (AY875644); G. max, soybean (BM187962; nucleotidesequence was retrieved from the National Center for Biotechnology Information (NCBI) and then translated; longest readingframe was selected for alignment); At1g48300, Arabidopsis 1g48300 (AAD49767); O. sativa, rice (XP_475575). Conservedamino acids are shaded black.
Saha et al.
1536 Plant Physiol. Vol. 141, 2006 www.plant.org on August 12, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.

Figure 4. Conserved acyltransferase domains across acyltransferase families and phylogenetic analysis. Sequence alignment ofpeanut DGAT (AY875644) and other known acyltransferase family members was performed using the http://www.ch.embnet.org/software/BOX_form.html sequence alignment program. Identical amino acid residues are highlighted in black. Conservedacyltransferase domains are underlined. Catalytic site conserved residues are marked with stars. A, Alignment of the catalyticdomains of members of the acyltransferase family. Sequences aligned are designated as follows: E. coli GPAT and LPEAT, EcGPATand EcLPEAT; Mus musculus GPAT and LPAAT, MmGPAT and MmLPAAT; S. cerevisiae GPAT1, GPAT2, and LPAAT are ScGAT1,ScGAT2, and ScLPAAT; Homo sapiens LPAAT1 to 6 and DHAPT are HsLPAAT1 to 6 and HsDHAPT; Arabidopsis GPAT, AtGPAT;A. calcoaceticus wax synthase/DGAT, AcWS/DGAT; and peanut DGAT, AhDGAT. B, Similar alignment of catalytic domain of
Cytosolic Triacylglycerol Biosynthesis in Oilseeds
Plant Physiol. Vol. 141, 2006 1537 www.plant.org on August 12, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.

BLAST analyses have identified likely orthologs inArabidopsis, rice (Oryza sativa), and soybean (Glycinemax), and closely related genes are represented in ESTcollections from diverse plant species. Soybeanshowed the identity of 50% (63% similarity) to peanutDGAT. Similarly, rice showed 44% identity (53% sim-ilarity) and Arabidopsis showed 33% identity (simi-larity 48%) to peanut DGAT (Fig. 3B). Therefore, theisolated DGAT gene is unique and might constitute adifferent family of cytosolic DGAT.
When the deduced amino acid sequence of 38 kDwas examined for a number of structural motifs, weidentified potential DGAT motifs at 54HVQYYGD60
(Fig. 4A) and 205HHNAVELFSRNND217 (Fig. 4B), anda partially conserved DGAT active site at81KKRVLFDDL89 (Fig. 4C), which matched with re-ported members of the acyltransferase family. Inser-tion of a few amino acids between critical His and Aspresidues has also been reported in Mycobacteriumtuberculosis bifunctional wax ester synthase/DGAT
Figure 4. (Continued.)known acyltransferase with yet another domain in peanut DGAT. C, Active site of catalytic residues of known DGATs aligned withAhDGAT. Designation of the sequences aligned is as follows: M. musculus DGAT1, MmDGAT1; HARGP1; Nicotiana tabacumDGAT1, NtDGAT1; Caenorhabditis elegansDGAT1, CeDGAT1; Arabidopsis DGAT1, AtDGAT1; and rapeseedDGAT1, BnDGAT1.D, Phylogenetic tree of different DGAT family members. Cytosolic DGAT from peanut (AhDGAT; AY875644) was added to theassembly and a similarity tree was constructed. G.max, soybean (BM187962); O. sativa, rice (XP_475575); At1g48300, Arabidopsis1g48300 (AAD49767); Rv3130c,M. tuberculosis H37Rv DGAT (NP_217646); AcDGAT, A. calcoaceticusWS/DGAT (AF529086);MmDGAT1, M. musculus DGAT1 (NP_034176); NtDGAT1, N. tabacum DGAT1 (AF129003); BnDGAT1, rapeseed DGAT1(AF164434); AtDGAT1, Arabidopsis DGAT1 (AF051849); ScDGAT2, S. cerevisiae DGAT2 (YOR245c); HsDGAT2, H. sapiensDGAT2 (NP_115953); MrDGAT2b, M. ramanniana DGAT2b (AF391090); MrDGAT2a, M. ramanniana DGAT2a (AF391089);CeDGATa, C. elegans DGATa (CAB04533); CeDAGT2b, C. elegans DGAT2b (AAB04969); AtPDGAT, Arabidopsis PDGAT(NP_171897); and ScPDGAT, S. cerevisiae PDGAT (CAA54576). GenBank/NCBI accession numbers are given (in parentheses).
Figure 5. Induction of TAG synthesis in E. colioverexpressing AhDGAT. A, Time course of ace-tate labeling. Vector-transformed and recombi-nant E. coli cells expressing soluble peanut DGATwere labeled with [14C]acetate for various timepoints. The labeling experiments were repeatedthree times and a typical phosphor image isshown. B, Time course of recombinant DGATactivity. Enzyme activity was done using 18,000gsupernatant of E. coli BL21 (DE3) cells overex-pressing AhDGAT. C and D, Immunoblot analysesof the expressed AhDGAT. Induced (1IPTG) anduninduced (2IPTG) recombinant cell lysateswere resolved in 12% (w/v) SDS-PAGE, trans-ferred onto a nitrocellulose membrane, andprobed with anti-His-tag (C) and peptide anti-RgDGAT (D) antibodies. E, Alignment ofR. glutinisDGAT peptide sequence with deducedamino acid sequence of peanut DGAT.
Saha et al.
1538 Plant Physiol. Vol. 141, 2006 www.plant.org on August 12, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.

(Daniel et al., 2004). In addition, Tyr kinase phosphor-ylation site (PCDOC0007) 183RKAETMIL190 showed 100%identity. There are several protein kinase C and caseinkinase 2 phosphorylation sites present in the pro-tein. Phosphopantetheine attachment site (PCDO-C0012) displayed 75% similarity between residues24RRGGDSCVTVPVRMRK39. Thiolase acyl enzymeintermediatesignature(PCDO0092)displayed68%sim-ilarity between residues 135TNPDCESSSSSSSSESESES154.An invariant Pro is present in the hydrophobic blockat position 34 in peanut DGAT between a putativephosphopantetheine attachment site and a thiolaseacyl enzyme intermediate signature and therefore maybe responsible for presenting the fatty acyl group tothe active site for esterification to the glycerol back-bone. Fatty-acid binding protein signature pattern(PCDOC00188) displayed 64% similarity between res-idues 255KSGSIALLQEFERVVGAEG273.An alignment with known acyltransferases con-
firmed the presence of several acyltransferase motifs,including the DGATcatalytic motif present in cytosolicpeanut DGAT. In addition, we used the ClustalWalgorithm to obtain an optimized multiple sequencealignment with known DGAT1 and 2 family membersand found the identity to be ,10% (,30% similarity)and 13% (53% similarity), respectively, with the bac-terial bifunctional wax ester synthase/DGAT. There-fore, the isolated DGAT gene is unique and mightconstitute a different family of cytosolic DGAT. Den-drogram representing the phylogenetic relationship ofAhDGAT with DGAT from many different organismsacross the species was performed. The analysis re-vealed that AhDGAT is closely related to bacterialbifunctional DGAT/wax ester synthase (Fig. 4D).
Expression of AhDGAT in E. coli
A pBluescript SK2 vector containing the peanutDGAT full length (345 amino acids) was subcloned inpRSET A by BamHI and HindIII digestion. The trans-formed E. coli BL21 (DE3) cells were induced with0.5 mM isopropyl-1-thio-b-D-galactopyranoside (IPTG),and the induced cells were used for preparing cellextracts for enzyme activity and the intact cells usedfor [14C]acetate labeling. Studies on the incorporationof [14C]acetate into lipids of transformed E. coli showeda time-dependent increase in the incorporation oflabel into TAG and wax ester as compared to vector-transformed cells (Fig. 5A). In vitro DGAT assay withAhDGAT-transformed cell lysates showed a time-dependent increase in incorporation of [1-14C]oleoyl-CoA into TAG (Fig. 5B). Immunoblot analysis of thetransformed cells with anti-His-tag (Fig. 5C) and pep-tide anti-DGAT (Fig. 5D) antibodies showed an intensesignal at around 42 kD, but no signal was detectedin the vector-transformed cells. The internal aminoacid sequence of R. glutinis (Gangar et al., 2001) onaligning with the deduced amino acid sequence ofAhDGAT gene revealed a significant homology
(Fig. 5E), which could be the reason for antibodycross-reactivity.
Total lipids extracted from E. coli cells expressingAhDGAT gene were separated by silica-gel column,and the purified TAG and wax ester were subjected tostructural analysis using 1H-NMR and infrared spec-troscopy. These analyses confirmed the molecularidentity of TAG (data not shown) and wax ester (Sup-plemental Fig. 2). Cells were viewed under confocalmicroscope after staining with Nile blue A and oil redO. Localization studies revealed that cells harboringAhDGAT gene showed intense Nile blue A and oil red
Figure 6. Purification of recombinant AhDGAT. A, SDS-PAGE profile ofrecombinant DGAT purification. Samples from each fraction wereseparated by 12% (w/v) SDS-PAGE and stained with Coomassie BlueR-250. Lane 1 represents vector IPTG uninduced; lane 2, vector IPTGinduced; lane 3, molecular mass marker; lane 4, AhDGAT IPTGuninduced; and lane 5, AhDGAT IPTG induced. Lanes 6 to 7 corre-spond to 250 mM imidazole eluted fractions from Ni-nitrilotriaceticacid agarose chromatography. B, Time course of DGAT activity usingpurified recombinant enzyme. Lane 1 represents no-enzyme control.Lanes 2 to 6 represent 0, 5, 10, 15, and 30 min, respectively.
Cytosolic Triacylglycerol Biosynthesis in Oilseeds
Plant Physiol. Vol. 141, 2006 1539 www.plant.org on August 12, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.
O staining as compared to vector-transformed cells(Supplemental Fig. 3).
The full-length AhDGAT cDNA was subcloned intopRSETA and expressed in E. coli BL21 (DE3) cells as anN-terminal fusion protein of oligo-His. The His-6-AhDGATwas purified by a nickel (Ni)-affinity columnand shown to have amolecularmass of 42 kD (Fig. 6A).Recombinant DGAT is of a higher molecular mass ascompared to the calculated molecular mass, and thiscould be due to the insertion of a few amino acids atthe N terminus from the parent vector.
Characterization of Recombinant AhDGAT
To determine the acylation kinetics, the purifiedrecombinant acyltransferase was incubated with DAG
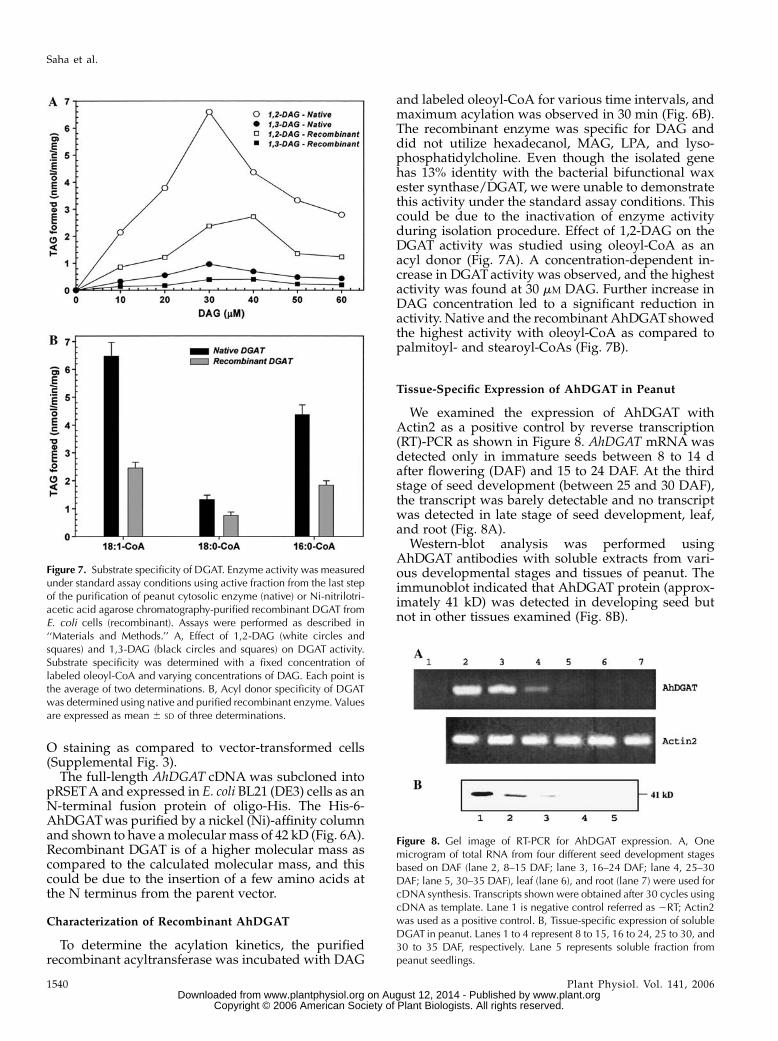
and labeled oleoyl-CoA for various time intervals, andmaximum acylation was observed in 30 min (Fig. 6B).The recombinant enzyme was specific for DAG anddid not utilize hexadecanol, MAG, LPA, and lyso-phosphatidylcholine. Even though the isolated genehas 13% identity with the bacterial bifunctional waxester synthase/DGAT, we were unable to demonstratethis activity under the standard assay conditions. Thiscould be due to the inactivation of enzyme activityduring isolation procedure. Effect of 1,2-DAG on theDGAT activity was studied using oleoyl-CoA as anacyl donor (Fig. 7A). A concentration-dependent in-crease in DGATactivity was observed, and the highestactivity was found at 30 mM DAG. Further increase inDAG concentration led to a significant reduction inactivity. Native and the recombinant AhDGATshowedthe highest activity with oleoyl-CoA as compared topalmitoyl- and stearoyl-CoAs (Fig. 7B).
Tissue-Specific Expression of AhDGAT in Peanut
We examined the expression of AhDGAT withActin2 as a positive control by reverse transcription(RT)-PCR as shown in Figure 8. AhDGAT mRNAwasdetected only in immature seeds between 8 to 14 dafter flowering (DAF) and 15 to 24 DAF. At the thirdstage of seed development (between 25 and 30 DAF),the transcript was barely detectable and no transcriptwas detected in late stage of seed development, leaf,and root (Fig. 8A).
Western-blot analysis was performed usingAhDGAT antibodies with soluble extracts from vari-ous developmental stages and tissues of peanut. Theimmunoblot indicated that AhDGAT protein (approx-imately 41 kD) was detected in developing seed butnot in other tissues examined (Fig. 8B).
Figure 7. Substrate specificity of DGAT. Enzyme activity was measuredunder standard assay conditions using active fraction from the last stepof the purification of peanut cytosolic enzyme (native) or Ni-nitrilotri-acetic acid agarose chromatography-purified recombinant DGAT fromE. coli cells (recombinant). Assays were performed as described in‘‘Materials and Methods.’’ A, Effect of 1,2-DAG (white circles andsquares) and 1,3-DAG (black circles and squares) on DGAT activity.Substrate specificity was determined with a fixed concentration oflabeled oleoyl-CoA and varying concentrations of DAG. Each point isthe average of two determinations. B, Acyl donor specificity of DGATwas determined using native and purified recombinant enzyme. Valuesare expressed as mean 6 SD of three determinations.
Figure 8. Gel image of RT-PCR for AhDGAT expression. A, Onemicrogram of total RNA from four different seed development stagesbased on DAF (lane 2, 8–15 DAF; lane 3, 16–24 DAF; lane 4, 25–30DAF; lane 5, 30–35 DAF), leaf (lane 6), and root (lane 7) were used forcDNA synthesis. Transcripts shown were obtained after 30 cycles usingcDNA as template. Lane 1 is negative control referred as 2RT; Actin2was used as a positive control. B, Tissue-specific expression of solubleDGAT in peanut. Lanes 1 to 4 represent 8 to 15, 16 to 24, 25 to 30, and30 to 35 DAF, respectively. Lane 5 represents soluble fraction frompeanut seedlings.
Saha et al.
1540 Plant Physiol. Vol. 141, 2006 www.plant.org on August 12, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.

DISCUSSION
The biosynthesis of TAG is shown to occur inmicrosomal membranes (Ohlrogge et al., 1991; Hobbset al., 1999; Zou et al., 1999; Bouvier-Nave et al., 2000;Oelkers et al., 2000; Lardizabal et al., 2001; Nykiforuket al., 2002; Kalscheuer and Steinbuchel, 2003). Wereport here the presence of TAG biosynthetic machin-ery in cytosol of developing peanut cotyledons. Theexistence of cytosolic enzymes that provide importantprecursors for TAG biosynthesis has been well docu-mented in plant systems. Soluble glycerol-3-P acyl-transferase has been identified and purified fromseveral sources (Murata and Tasaka, 1997). In devel-oping rapeseed, the formation of PA has been demon-strated in cytosolic fraction (Murphy, 1988), and it hasbeen proposed that the cytosolic PA phosphatasecould be metabolically inactive (Ichihara et al., 1988).Cytosolic LPA phosphatase (Shekar et al., 2002) andMAG acyltransferase (Tumaney et al., 2001) have beenreported in developing peanut cotyledons. Severalindependent lines of evidence indicate that the iden-tified DGAT is cytosolic in nature, and this is basedprimarily on the following observations. (1) The ac-tivity is associated with 150,000g supernatant. (2) Theenzyme is purified by successive column chromato-graphic separations without detergent. (3) The isolatedgene (AhDGAT) has neithermembrane spanning regionnor signal sequence (GenBank accession no. AY875644).Our results demonstrate that the isolated cDNA
encodes a DGAT. (1) Although E. coli cells do not syn-thesize TAG, AhDGAT-transformed cells showed sig-nificant amounts of TAG and wax ester accumulationas compared to vector-transformed cells. (2) E. colicells expressing AhDGAT gene were capable of incor-porating radiolabeled acetate into TAG and wax ester.(3) The expressed recombinant protein cross-reactedwith R. glutinis peptide DGAT polyclonal antibodiesand the same protein also cross-reacted with mono-clonal antibody against His-tag. (4) A higher level ofDGAT activity was obtained from cells expressing theisolated cDNA than from cell-free extract from vector-transformed E. coli cells. The acyltransferase activitydepended on the presence of oleoyl-CoA and wasspecific for DAG. We propose that the isolated enzymecould be involved in yet another TAG and wax esterbiosynthetic pathway in plants.Four independent gene families (DGAT1, DGAT2,
PDAT, and bifunctional WS/DGAT) have been con-ferred the ability to synthesize TAG. Recent reportssuggest, apart from unspecific bifunctional WS/DGAT(Uthoff et al., 2005), that DGAT1 has wax ester syn-thase and acyl CoA:retinol acyltransferase activitieswhen expressed in COS 7 cells (Yen et al., 2005). It hasbeen speculated that these gene families may playdifferent roles across species, in a tissue- and time-dependent manner during development (Cases et al.,1998; Hobbs et al., 1999; Zou et al., 1999; Bouvier-Naveet al., 2000; Dahlqvist et al., 2000; Oelkers et al., 2000;Lardizabal et al., 2001; Nykiforuk et al., 2002; Kalscheuer
and Steinbuchel, 2003). Therefore, the isolatedAhDGAT may be involved in a unique pathway ofTAG and wax ester synthesis. Further studies are re-quired to understand the roles of various gene familiesin various organisms.
DGAT1 mRNA from Arabidopsis was present athigh levels in many different tissues, including ger-minating seeds, young seedlings, roots, and leaves.However, our results from peanut indicated thatAhDGAT mRNA is detected only in immature seeds.During embryo development, AhDGAT protein ispresent at 8 DAF, increases dramatically at 15 to24 DAF, and then gradually declines in the matureseeds. A recent microarray study of tissue-specificArabidopsis ESTs has determined that Arabidopsishomolog of AhDGAT is highly (.2-fold) expressed inthe early stages of seed development (Yamada et al.,2003). Arabidopsis homolog (At1g48300) is highlyexpressed in cork, xylem, hypocotyl, and senescentleaf, and moderately expressed in pollen and stem(Supplemental Table I). DGAT homolog is shown to beubiquitously expressed in various developmentalstages, and very highly expressed in leaf and silique(Zimmermann et al., 2004). Therefore, it is likely thatAhDGAT and AtDGAT expression is regulated in atissue-specific and time-dependent manner. Hence,the identification of AhDGAT and AtDGAT has sig-nificant implications in understanding the regulationof TAG biosynthesis in plants.
In addition to the membrane-bound pathway forTAG synthesis, an alternate pathway was proposedthat involved the formation of MAG from LPA by theenzyme LPA phosphatase (Shekar et al., 2002), whichin turn gets converted to DAG by MAG acyltransfer-ase (Tumaney et al., 2001). The further acylation ofDAG to TAG involves another enzyme, DGAT, thuscompleting the biosynthesis of TAG in oilseeds. Genesinvolved in this pathway could find application inimproving the quantity of vegetable oils.
MATERIALS AND METHODS
Materials
[1-14C]Oleoyl-CoA (54 mCi mmol21), [1-14C]acetate (2 mCi mmol21), [9,10-3H(N)]trioleoylglycerol (10 Ci mmol21), [2-palmitoyl-9,10-3H]phosphatidyl-
choline (92.3 Ci mmol21), and [g-32P]ATP (3,000 Ci mmol21) were obtained
from Perkin Elmer Biosystems. Superdex 75 (26/60 prep grade) FPLC column,
octyl-Sepharose, blue-Sepharose, heparin-agarose, and gel-filtration molecular
mass standards were purchased fromAmersham. Silica-TLC plates were from
Merck. Hexadecanol, 1,2-dipalmitoyl-sn-glycerol (1,2-DAG), 1,3-dipalmitoyl-
sn-glycerol (1,3-DAG), Nile blue A, oil red O, and other chemicals were from
Sigma-Aldrich. Field-grown developing peanut (Arachis hypogaea; JL 24)
cotyledons were harvested at 20 to 24 DAF and used either fresh or stored
at 280�C until further use.
Subcellular Fractionation
Frozen immature seeds (50 g) were ground in a prechilled mortar and
pestle with 5 g of acid-washed sand and 125 mL of extraction buffer
containing 50 mM Tris-HCl, pH 8.0, 1 mM EDTA, 10 mM KCl, 1 mM MgCl2,
1 mM b-mercaptoethanol, 0.1 mM phenylmethylsulfonyl fluoride, 1 mg mL21
Cytosolic Triacylglycerol Biosynthesis in Oilseeds
Plant Physiol. Vol. 141, 2006 1541 www.plant.org on August 12, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.

leupeptin, and 0.25 M Suc. The extract was passed through two layers of
cheesecloth and the filtrate was differentially centrifuged to fractionate
intracellular components. The filtrate was centrifuged at 3,000g for 10 min
and the supernatant was centrifuged at 18,000g for 15 min. The 18,000g
supernatant was further centrifuged at 150,000g for 1.5 h. The pellet thus
obtained was resuspended in a small volume of buffer containing 20 mM Tris-
HCl, pH 7.0, and 1 mM b-mercaptoethanol. Cytosol (150,000g supernatant,
soluble fraction) was used as the enzyme source. All the operations were
performed at 4�C. Protein concentrations were determined by the bicincho-
ninic acid method (Smith et al., 1985) using bovine serum albumin (BSA) as
the standard.
DGAT Assay
The assaymixtures consisted of 50mM Tris-HCl, pH 7.5, 1 mMMgCl2, 10 mM
[1-14C]oleoyl-CoA (2 3 105 cpm), 15 to 40 mg of protein, and 30 mM sonicated
suspension of 1,2-DAG in a total volume of 100 mL. The incubation was
carried out at 30�C for 15 min and stopped by the addition of 400 mL of
CHCl3:CH3OH (1:2, v/v). Lipid extraction was carried out and chloroform-
soluble material was separated by silica-TLC using petroleum ether:diethyl
ether:acetic acid (70:30:1, v/v) as a solvent system. The lipids were visualized
with iodine vapor and identified by their migration with standards. The spots
of TAGwere scraped off from TLC for determination of radioactivity by liquid
scintillation counting. Control incubations were carried out for zero time, in
the absence of enzyme and heat-inactivated enzyme source. The control
values were subtracted from the actual assay value, and enzyme activity was
calculated after correction. Wax ester synthase activity was monitored under
the standard DGAT assay conditions. Acyl acceptor in this assay was hexa-
decanol instead of DAG.
Purification of DGAT
We previously reported the purification of MAG acyltransferase from
cytosol (soluble fraction) of immature peanut (Tumaney et al., 2001). During
purification, both MAG and DGAT activities coeluted, and the separation of
these activities was achieved in heparin-agarose column. The DGAT activity
was eluted at 0.25 M salt.
Screening of a cDNA Library
A seed-specific cDNA library was constructed in l-ZAP II (Rudrabhatla
and Rajasekharan, 2002). The library was screened using oligonucleotide
generated from the internal amino acid sequence of the purified peanut
DGAT. A total of 2 3 106 plaques were screened. Positive plaques were
purified by three additional screenings with a homologous oligonucleotide
probe.
Sequence Analysis
Plasmid DNA isolated from the positive clone was sequenced on both
strands using the Biotech Taq cycle sequencing kit on an automated sequencer
(Applied Biosystems 377). Sequence data were analyzed and compared using
the following programs and software. Sequence analysis was performed using
BLAST (http://www.ncbi.nlm.nih.gov/BLAST). Multiple sequence align-
ment was done with ClustalW (http://www.ebi.ac.uk). Pedro’s Biomolecular
Research Tool (http://www.public.iastate.edu/_pedro) and ExPASy Molec-
ular Biology Server (http://www.expasy.org) were used for domain andmotif
identification. Protein hydropathy plot was generated by http://www.bio.
davidson.edu/-courses/compbio/flc/home.html. The phylogenetic tree was
generated using bootstrapping for 10,000 trees, using ClustalW version 1.82.
The bootstrap tree was converted to figure format using NJplot (Perriere and
Gouy, 1996).
Expression of a His-6-AhDGAT Fusion Protein
The cDNA spanning region of AhDGAT was subcloned into His-tagged
fusion protein expression vector pRSETA (Invitrogen) by BamHI and HindIII
digestion. The constructs were expressed in Escherichia coli BL21 (DE3). The
fusion protein was induced with 0.5 mM IPTG for 12 h at 22�C and purified by
Ni-nitrilotriacetic acid agarose chromatography (Qiagen). Expression was
verified by immunoblotting with oleaginous yeast anti-DGAT and His-tag
antibodies.
Immunoblotting
Proteins were separated by 12% (w/v) SDS-PAGE and transferred onto a
nitrocellulose membrane by a semidry transfer apparatus. Upon transfer, the
membrane was blocked with 0.5% (w/v) BSA in phosphate buffered saline for
1 h. The peptide corresponding to Rhodotorula glutinis cytosolic DGAT was
conjugated to BSA and antiserumwas raised as described (Gangar et al., 2001).
Primary antibodies were diluted (1:500, v/v) in phosphate buffered saline
containing 0.1% (w/v) BSA, added to the blocked membrane, and incubated
for 1 h. The unbound antibodies were washed with phosphate buffered saline
containing 0.05% (v/v) Tween 20, and goat anti-rabbit IgG-alkaline phospha-
tase conjugate was added to the membrane and incubated for 45 min. The
membrane was washed and the immunoreactive bands were visualized using
5-bromo-4-chloro-3-indolyl phosphate/nitro blue tetrazolium substrate.
Nile Blue A and Oil Red O Staining
A smear of E. coli cells was prepared on a glass slide and heat fixed. The
slides were immersed in 1% (w/v) aqueous solution of Nile blue A stain for
10 min at 55�C. Excess stain was removed, air-dried, and visualized under
confocal microscope (Gangar et al., 2001). Oil red O was dissolved to a
stock solution by adding 500 mg oil red O to 100 mL of 60% (v/v) triethyl-
phosphate. For staining, 36% (v/v) triethyl-phosphate working solution,
containing 12 mL of oil red O stock solution and 8 mL of water, was prepared,
and cells were stained for 30 min and washed three times in water, air-dried,
and visualized under confocal microscope on 1003 magnification.
Incorporation of [1-14C]Acetate into TAG
E. coli cells (106 cells mL21) were incubated with [14C]acetate (1.5 mCi mL21
Luria-Bertani medium) and grown for 2 h at 37�C. Cells were harvested by
centrifugation, and the cell pellet was washed twice with ice-cold water. To the
pellet 0.5 mL of 10% (v/v) acetic acid in isopropyl alcohol was added and
boiled for 3 min. One milliliter of hexane was added to the mixture to extract
lipids. Hexane layer was removed and concentrated. The extracted lipids were
separated on a silica-TLC using petroleum ether:diethyl ether:acetic acid
(70:30:1, v/v) as the solvent system. Lipids were identified by their migration
with standards. The TLC plate was phosphorimaged, and individual spots
were scraped off from the plate and the radioactivity was measured in a liquid
scintillation counter.
RNA Isolation and RT-PCR
Total RNAwas isolated from four different seed development stages based
on DAF (stage 1, 8–15 DAF; stage 2, 16–24 DAF; stage 3, 25–30 DAF; stage 4,
30–35 DAF), leaf, and root from soil-grownwild-type plants using TRI reagent
(Sigma) as described by the manufacturer. One microgram of RNA was
reverse transcribed at 43�C using Moloney murine leukemia reverse tran-
scriptase and the RevertAid H Minus first-strand cDNA synthesis kit
(Fermentas GmbH) with oligo(dT) as primer according to the protocol
provided by the supplier. The resulting cDNA was diluted 10-fold and 1 mL
used as template for 30 cycles of PCR amplification using Taq DNA polymer-
ase with the specific oligonucleotides for AhDGAT (forward 5#-ATG-
GAGGTTTCAGGCGCCGTTC-3#; reverse 5#-TTCATTAGTACTTTCCTGATT-
CTCGCCCAAG-3#). Amplification of the Actin2 control transcript was
done using the primers Actin2-1 (5#-TCCCTCAATCTCATCTTCTTCC-3#)and Actin2-1 (5#-GACCTGCCTCATCAATCTTCTTCC-3#). The PCR products
were analyzed on ethidium bromide-stained agarose gels.
Other Methods
Samples were resuspended in CdCl3, andNMR spectra of the purified lipid
were recorded using the JEOL JNM-LA 300 FT NMR system. Infrared spectra
were recorded on the JASCO FT/IR-410 system with neat samples. Samples to
be analyzed were converted to fatty-acid methyl esters and purified by
preparative TLC, and then analyzed under GC-EIMS using theVGAutoSpecM
mass spectrometer equipped with HP 5890 series II gas chromatography fitted
with a HP-5 capillary column (Tumaney and Rajasekharan, 1999).
Saha et al.
1542 Plant Physiol. Vol. 141, 2006 www.plant.org on August 12, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.

Sequence data from this article can be found in the GenBank/EMBL data
libraries under accession number AY875644.
Received April 16, 2006; revised May 27, 2006; accepted May 27, 2006;
published June 23, 2006.
LITERATURE CITED
Bouvier-Nave P, Benveniste P, Oelkers P, Sturley SL, Schaller H (2000)
Expression in yeast and tobacco of plant cDNAs encoding acyl CoA:
diacylglycerol acyltransferase. Eur J Biochem 267: 85–96
Broun P, Gettner S, Somerville C (1999) Genetic engineering of plant
lipids. Annu Rev Nutr 19: 197–216
Cases S, Smith SJ, Zheng Y-W, Myers HM, Lear SR, Sande E, Novak S,
Collins C, Welch CB, Lusis AJ, et al (1998) Identification of a gene
encoding an acyl CoA:diacylglycerol acyltransferase, a key enzyme in
triacylglycerol synthesis. Proc Natl Acad Sci USA 95: 13018–13023
Dahlqvist A, Stahl U, Lenman M, Banas A, Lee M, Sandager L, Ronne H,
Stymne S (2000) Phospholipid:diacylglycerol acyltransferase: an en-
zyme that catalyzes the acyl-CoA-independent formation of triacylglyc-
erol in yeast and plants. Proc Natl Acad Sci USA 97: 6487–6492
Daniel J, Deb C, Dubey VS, Sirakova TD, Abomoelak B, Morbidoni HR,
Kolattukudy PE (2004) Induction of a novel class of diacylglycerol
acyltransferases and triacylglycerol accumulation in Mycobacterium
tuberculosis as it goes into a dormancy-like state in culture. J Bacteriol
186: 5017–5030
Gangar A, Karande AA, Rajasekharan R (2001) Isolation and localization
of a cytosolic 10 S triacylglycerol biosynthetic multienzyme complex
from oleaginous yeast. J Biol Chem 276: 10290–10298
Hobbs DH, Lu C, Hills MJ (1999) Cloning of cDNA encoding diacylglyc-
erol acyltransferase from Arabidopsis thaliana and its functional expres-
sion. FEBS Lett 452: 145–149
Ichihara K, Takahashi T, Fujii S (1988) Diacylglycerol acyltransferase in
maturing safflower seeds: its influences on the fatty acid composition of
triacylglycerol and on the rate of triacylglycerol synthesis. Biochim
Biophys Acta 958: 125–129
Kalscheuer R, Steinbuchel A (2003) A novel bifunctional wax ester
synthase/acyl-CoA: diacylglycerol acyltransferase mediates wax ester
and triacylglycerol biosynthesis in Acinetobacter calcoaceticus ADP1.
J Biol Chem 278: 8075–8082
Kennedy EP (1961) Biosynthesis of complex lipids. Fed Proc 20: 934–940
Lardizabal KD, Mai JT, Wagner NW, Wyrick A, Voelker T, Hawkins DJ
(2001) DGAT2 is a new diacylglycerol acyltransferase gene family:
purification, cloning, and expression in insect cells of two polypeptides
from Mortierella ramanniana with diacylglycerol acyltransferase activity.
J Biol Chem 276: 38862–38869
Murata N, Tasaka Y (1997) Glycerol-3-phosphate acyltransferase in plants.
Biochim Biophys Acta 1348: 10–16
Murphy DJ (1988) A highly active soluble diacylglycerol synthesizing
system from developing rapeseed, Brassica napus L. Lipids 23: 157–163
Nykiforuk CL, Furukawa-Stoffer TL, Huff PW, Sarna M, Laroche A,
MoloneyMM,Weselake RJ (2002) Characterization of cDNAs encoding
diacylglycerol acyltransferase from cultures of Brassica napus and su-
crose mediated induction of enzyme biosynthesis. Biochim Biophys
Acta 1580: 95–109
Oelkers P, Tinkelenberg A, Erdeniz N, Cromley D, Billheimer JT,
Sturley SL (2000) A lecithin cholesterol acyltransferase-like gene me-
diates diacylglycerol esterification in yeast. J Biol Chem 275: 15609–
15612
Ohlrogge JB, Browse J, Somerville C (1991) The genetics of plant lipids.
Biochim Biophys Acta 1082: 1–26
Perriere G, Gouy M (1996) WWW-query: an on-line retrieval system for
biological sequence banks. Biochimie 78: 364–369
Rudrabhatla P, Rajasekharan R (2002) Developmentally regulated dual-
specificity kinase from peanut that is induced by abiotic stresses. Plant
Physiol 130: 380–390
Shekar S, Tumaney AW, Rao TJVS, Rajasekharan R (2002) Isolation of
lysophosphatidic acid phosphatase from developing peanut cotyledons.
Plant Physiol 128: 988–996
Smith PK, Krohn RI, Hermanson GT, Mallia AK, Gartner FH,
Provenzano MD, Fujimoto EK, Goeke NM, Olsen BJ, Klenk DC (1985)
Measurement of protein using bicinchoninic acid. Anal Biochem 150:
76–85
Stobart K, Mancha M, Lenman M, Dahlqvist A, Stymne S (1997) Triac-
ylglycerols are synthesized and utilized by transacylation reactions in
microsomal preparations of developing safflower (Carthamus tinctorius
L.) seeds. Planta 203: 58–66
Tumaney AW, Rajasekharan R (1999) Synthesis of azidophospholipids and
labeling of lysophosphatidylcholine acyltransferase from developing
soybean cotyledons. Biochim Biophys Acta 1439: 47–56
Tumaney AW, Shekar S, Rajasekharan R (2001) Identification, purification,
and characterization of monoacylglycerol acyltransferase from devel-
oping peanut cotyledons. J Biol Chem 276: 10847–10852
Uthoff S, Stoveken T, Weber N, Vosmann K, Klein E, Kalscheuer R,
Steinbuchel A (2005) Thio wax ester biosynthesis utilizing the unspe-
cific bifunctional wax ester synthase/acyl coenzyme A:diacylglycerol acyl-
transferase of Acinetobacter sp. strain ADP1. Appl Environ Microbiol 71:
790–796
Yamada K, Lim J, Dale JM, Chen H, Shinn P, Palm CJ, Southwick AM, Wu
HC, Kim C, NguyenM, et al (2003) Empirical analysis of transcriptional
activity in the Arabidopsis genome. Science 302: 842–846
Yen CL, Monetti M, Burri BJ, Farese RV Jr (2005) The triacylglycerol
synthesis enzyme DGAT1 also catalyzes the synthesis of diacylglyc-
erols, waxes, and retinyl esters. J Lipid Res 46: 1502–1511
Zimmermann P, Hirsch-Hoffmann M, Hennig L, Gruissem W (2004)
GENEVESTIGATOR. Arabidopsis microarray database and analysis
toolbox. Plant Physiol 136: 2621–2632
Zou J, Wei Y, Jako C, Kumar A, Selvaraj G, Taylor DC (1999) The
Arabidopsis thaliana TAG1 mutant has a mutation in a diacylglycerol
acyltransferase gene. Plant J 19: 645–653
Cytosolic Triacylglycerol Biosynthesis in Oilseeds
Plant Physiol. Vol. 141, 2006 1543 www.plant.org on August 12, 2014 - Published by www.plantphysiol.orgDownloaded from
Copyright © 2006 American Society of Plant Biologists. All rights reserved.




















