
Physical properties of ordered mesoporous SBA-15 silica as immunological adjuvant

lable at ScienceDirect
Fish & Shellfish Immunology xxx (2013) 1e10
Contents lists avai
Fish & Shellfish Immunology
journal homepage: www.elsevier .com/locate / fs i
Full length article
A redox active site containing murrel cytosolic thioredoxin: Analysisof immunological properties
Rajesh Palanisamy a, Prasanth Bhatt a, Venkatesh Kumaresan a, Mukesh Kumar Chaurasia a,Annie J. Gnanamb, Mukesh Pasupuleti c, Marimuthu Kasi d, Jesu Arockiaraj a,*aDivision of Fisheries Biotechnology & Molecular Biology, Department of Biotechnology, Faculty of Science and Humanities, SRM University, Kattankulathur,Chennai 603 203, Tamil Nadu, Indiab Institute for Cellular and Molecular Biology, The University of Texas at Austin, 1 University Station A4800, Austin, TX 78712, USAc Lab PCN 206, Microbiology Division, CSIR e Central Drug Research Institute, B.S. 10/1, Sector 10, Jankipuram Extension, Sitapur Road, Lucknow 226031,Uttar Pradesh, IndiadDepartment of Biotechnology, Faculty of Applied Sciences, AIMST University, Semeling Bedong, 08100 Bedong, Kedah, Malaysia
a r t i c l e i n f o
Article history:Received 16 September 2013Received in revised form21 October 2013Accepted 21 October 2013Available online xxx
Keywords:ThioredoxinChanna striatusHydrogen peroxideCell proliferationAntioxidant
* Corresponding author. Tel.: þ91 44 27452270; faxE-mail address: [email protected] (J. Arockiar
1050-4648/$ e see front matter � 2013 Elsevier Ltd.http://dx.doi.org/10.1016/j.fsi.2013.10.016
Please cite this article in press as: Palanisamproperties, Fish & Shellfish Immunology (20
a b s t r a c t
In this study, we have reported the immunological properties of cDNA encoding thioredoxin which isobtained from the database of Channa striatus (named as CsTRx) cDNA library. The analysis showed thatthe CsTRx polypeptide contains a thioredoxin domain between Val2 and Asn106. The domain possessed athioredoxin active family at 24e42 along with a redox active site (also known as catalytic center) at31WCGPC35. The analysis showed that the catalytic center is responsible for the control of proteinfunction. Phylogenetic study showed that CsTRx clustered together with vertebrate TRx-1. Based on thephylogenetic analysis and other bioinformatics analysis, it is confirmed that the characterized CsTRxbelongs to TRx-1 family. In addition, the sub-cellular localization prediction analysis showed that CsTRxis a cytosol thioredoxin. The highest gene expression was observed in gill (P < 0.05). Further, its tran-scriptional modulation was evaluated under fungal (Aphanomyces invadans), bacterial (Aeromonashydrophila) and H2O2 challenges. The recombinant CsTRx protein was over-expressed and purified usingan Escherichia coli expression vector system. We conducted a H2O2 peroxidase assay using recombinantCsTRx protein under various pH and temperature. Further, we studied the influence of recombinantCsTRx protein on C. striatus spleen leukocyte activation. The recombinant CsTRx protein enhanced thecell proliferation in a concentration dependant manner. The results of antioxidant analysis showed thatthe antioxidant capacity of recombinant CsTRx protein was determined to be 4.2 U/mg protein. Weconducted an insulin disulfides assay to study the enzymatic oxidoreductase activity of CsTRx and weobserved no activity in the control group. But the recombinant CsTRx protein addition rapidly increasedthe enzymatic oxidoreductase activity. Over all, the results showed that the CsTRx may contain potentialantioxidant properties, which could regulate the oxidative stress created by various biological pathogensas well as chemical stress in the immune system of C. striatus, thus protecting it.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction
The innate immune system in eukaryotes is a crucial hostdefense against invading pathogens, particularly in aquatic or-ganisms. One of the protective defense mechanism is the pro-duction of microbicidal reactive oxygen species (ROS) [1e3]. Atlow concentration, ROS have been shown to be beneficial inmany biological processes such as intracellular signaling, cell
: þ91 44 27453903.aj).
All rights reserved.
y R, et al., A redox active site13), http://dx.doi.org/10.1016
progression, apoptosis [4], immunity and defense against mi-crobial infection [5e7]. However, the high level of ROS andreactive oxygen intermediates (ROI) results in increased oxida-tive stress may lead to cell damage [8,9]. In order to detoxify oreliminate the raised levels of ROS/ROI, cells hold several anti-oxidants/reductants system derived either by intrinsically orextrinsically to maintain the intracellular environment in ahighly reduced state [10]. Redox regulation is vital for living or-ganisms, and many reactions are entailed in the regulation. As asmall redox protein that neutralizes ROS, thioredoxin (TRx) is oneof the key systems responsible for keeping the intracellularenvironment in a highly reduced state [11].
containing murrel cytosolic thioredoxin: Analysis of immunological/j.fsi.2013.10.016

R. Palanisamy et al. / Fish & Shellfish Immunology xxx (2013) 1e102
TRxs are small multi functional proteins (w12 kDa), ubiquitousin all forms of life [12]. TRxs are characterized by a conserved cat-alytic motif Trp-Cys-Gly-Pro-Cys (WCGPC) in their active sites [13].TRx was primarily discovered in Escherichia coli as a hydrogendonor for ribonucleotide reductase in 1964 by Laurent et al. [14].Since then, TRxs have been identified in many organisms [15]. Themain catalytic action of TRx is antioxidant-redox regulatory role, inwhich, it reduces disulfide bridge of substrate through the vicinalcysteine residue in its catalytic motif of WCGPC. In addition to itsredox-regulatory role, TRx also participates in regulation of growth[16], transcription factors [17], protein folding [18], scavengingradicals [19], signal transduction and DNA replication [20,21]. TRx issecreted by lymphocytes, hepatocytes, fibroblasts and cancer cells[16,22e24]. Recent report [25] says that antigen presenting cellssecrete TRx to activate T lymphocytes. TRxs have also been shownto be hiking the expression of a variety of cytokines, includinginterleukin-1, 2, 6 and 8 and tumor necrosis factor-a [26].
There are three different forms of TRx encoded by differentgenes that have been cloned and characterized so far [9]. TRx-1is a 12 kDa cytosolic protein existing primarily in cytosol, butalso present in nucleus and blood plasma. It is the most studiedTRx amongst the three forms [10,27]. It has also been cloned inboth prokaryotic and mammalian cells [15,28e30]. TRx-2 is amitochondrial protein that has a special 60 amino acid residuesas N-terminal translocation signal [31,32]. It contains aconserved catalytic site, but does not have other cysteine resi-dues that are present in Trx-1 [27]. The third isoform TRx-3 is avariant that is highly expressed in spermatozoa [33]. All forms ofTRx possess a conserved WCGPC active site, important for thefunction as a common and potent protein disulfide oxidoreduc-tase [34].
So far, only a few TRx have been reported from fin fishes such asblack rockfish Sebastes schlegelii [35] and rock bream (Oplegnathusfasciatus) [36]. However, these TRxs are varied. The knowledge onthe structural and functional role of fish TRx in the defense processis limited. Moreover, the information on TRx from snakehead fish(or striped murrel) Channa striatus is not available. Hence, it isnecessary to provide sound information on TRx from C. striatus.
C. striatus is a favorite freshwater food fish in many parts ofIndia due to its nutrient rich flesh, good market value and highmedicinal qualities. It is widely cultured in China and SoutheastAsian countries. With vast development of fish farming activities,a diverse group of biotic agents such as fungus, bacteria, andcutaneous ectoparasites may initiate skin lesions, which areconsequently colonized by fungus Aphanomyces invadans andeventually lead to epizootic ulcerative syndrome (EUS). Out-breaks of EUS had a drastic impact on striped murrel aquacultureindustry causing significant economic losses to fish farmers inrecent years [37e41]. Though effective use of antibiotics hasraised concerns such as the presence of antibiotic residues in thefish leading to the development of antibiotic resistance andenvironmental pollution. Thus, the enhancement of diseaseresistance in these fish using molecular techniques would be aviable alternative to address these problems.
To obtain a sound knowledge on the role of TRx from stripedmurrel immune system, we have characterized a full-length TRxcDNA from C. striatus (named as CsTRx) using various bioinfor-matics tools. Further, we reported the tissue-specific gene expres-sion of CsTRx and also reported the modulation of the geneexpression by biotic agents such as fungus A. invadans and bacteriaAeromonas hydrophila and abiotic agent hydrogen peroxide (H2O2).Moreover, we cloned the coding sequence of CsTRx in an E. colivector and purified its recombinant protein to study the functionalaspect of CsTRx including cell proliferation activity, antioxidantactivity and insulin disulfide reduction activity.
Please cite this article in press as: Palanisamy R, et al., A redox active siteproperties, Fish & Shellfish Immunology (2013), http://dx.doi.org/10.1016
2. Materials and methods
2.1. C. striatus cDNA library, CsTRx identification and bioinformaticsanalysis
A normalized C. striatus cDNA library was constructed usingtotal RNA isolated from spleen, liver, kidney, muscle and gills ofmurrel. The detailed procedure on C. striatus cDNA library estab-lishment was explained in our earlier studies [42,43]. A cDNAencoding CsTRx was identified as described in our earlier report[44,45] from C. striatus cDNA library during screening. The obtainedcomplete CsTRx cDNA sequence was analyzed on DNAssist 2.2 [46].Sequence similarities were searched on BLAST program (http://blast.ncbi.nlm.nih.gov/Blast) at NCBI Database. Domains and mo-tifs were analyzed on PROSITE Database (http://prosite.expasy.org/scanprosite/). The N-terminal transmembrane sequence wasdetermined by DAS Transmembrane Prediction Program (http://www.sbc.su.se/wmiklos/DAS). Signal peptide analysis was con-ducted using the SignalP (http://www.cbs.dtu.dk). CsTRx poly-peptide sequence was analyzed in CELLO (ver. 2.5) online programfor sub-cellular localization prediction. Multiple sequence align-ment was carried out on ClustalW (ver. 2) (http://www.ebi.ac.uk/Tools/msa/clustalw2/) and the alignment was edited on Bioedit(ver. 7.1.3.0). The phylogenetic tree was constructed usingNeighbor-Joining Method at MEGA 5.05. Further the genetic dis-tances were calculated using the Poisson Correction Method [47].Secondary structure of the CsTRx protein was constructed usingSOPMA Program and further the structure was analyzed on Poly-view Method (http://polyview.cchmc.org).
2.2. Fish, disease challenge and tissue collection
Healthy C. striatus (average body weight of 50 g) were obtainedfrom the Surya Agro Farms, Erode, Tamil Nadu, India. The fisheswere transported to Division of Fisheries Biotechnology & Molec-ular Biology, SRM University in oxygenated polythene bags. Theyweremaintained in 15 flat-bottomed plastic containers (150 L) withaerated and filtered de-chlorinated freshwater (water quality:dissolved oxygen, 5.8� 0.2 mg/L; water temperature, 28� 1 �C andpH, 7.2 � 0.2) in the laboratory. All fishes were acclimatized for 1week before being challenged to various immune stimulants. Amaximum of 15 fish per tank were maintained during the experi-ment. During acclimatization period, the fishes were fed ad libitumtwo times daily at 09:00 and 16:00 h with a commercially availablefish feed (Cargill Animal Nutrition, Andhra Pradesh, India).
To study the relative gene expression of CsTRx upon infection,C. striatuswere injected with fungus and bacteria intraperitoneally.PBS (1�) were prepared and served as control (100 mL/fish). Forfungus infection, 50 g fish were injected with 100 mL of A. invadans(a primary causative agent of EUS) at a concentration of 102 spores.In our earlier findings [42,43], we have clearly explained theisolation of A. invadans from the infected C. striatus muscle, culturein the laboratory, identification and injection to the fishes. Forbacterial challenge, the fish was injected with A. hydrophila (asecondary causative agent of EUS) (5 � 106 CFU/ml) suspended in1� phosphate buffer saline (100 mL/fish). A. hydrophila was alsoisolated and identified from the muscle sample of EUS infectedC. striatus as described by Dhanaraj et al. [48]. For H2O2 stress, thefish were injected with 100 mL H2O2 (0.5 g/mL).
Tissue samples (blood, liver, spleen, heart, intestine, kidney,head kidney, muscle, skin, gill and brain) were collected before(0 h), and after injection (3, 6, 12, 24, 48 and 72 h) and wereimmediately snap-frozen in liquid nitrogen and stored at �80 �Cuntil the total RNA was isolated. Using a sterilized syringe, blood(0.5e1.0 mL fish�1) was collected from the fish caudal fin and
containing murrel cytosolic thioredoxin: Analysis of immunological/j.fsi.2013.10.016

R. Palanisamy et al. / Fish & Shellfish Immunology xxx (2013) 1e10 3
immediately centrifuged at 4000 � g for 10 min at 4 �C to allowblood cell collection for total RNA extraction.
2.3. RNA extraction and cDNA synthesis
Total RNA from the control and infected fish were isolatedusing Tri Reagent� (Life Technologies), according to the manu-facture’s protocol with slight modifications [49,50]. Using 2.5 mgof RNA, first strand cDNA synthesis was carried out using aSuperScript� VILO� cDNA Synthesis Kit (Life technologies) withslight modifications [49,50]. The isolated cDNA was storedat �20 �C for further analysis.
2.4. Gene expression analysis by qRT-PCR
The gene expression of CsTRx in various tissues was determinedby quantitative real-time polymerase chain reaction (qRT-PCR).qRT-PCR was carried out using a ABI 7500 Real-time DetectionSystem (Applied Biosystems) in 20 mL reaction volume containing4 mL of cDNA from each tissue, 10 mL of Fast SYBR� Green MasterMix, 0.5 mL of each primer (20 pmol/ml) (CsTRx F1, CTG AAG GTGGAT GTGGAT GAG and CsTRx R2, CTG CAT CAG CAC CAGAGA A) and5 mL dH2O. The qRT-PCR cycle profile was 1 cycle of 95 �C for 10 s,followed by 35 cycles of 95 �C for 5 s, 58 �C for 10 s and 72 �C for 20 sand finally 1 cycle of 95 �C for 15 s, 60 �C for 30 s and 95 �C for 15 s.The same qRT-PCR cycle profile was used for the internal controlgene, b-actin. b-actin of C. striatus primers (b-actin F3, TCT TCC AGCCTT CCT TCC TTG GTA and b-actin R4, GAC GTC GCA CTT CAT GATGCT GTT) were designed from the sequence of GenBank AccessionNo. EU570219. After the PCR program, data were analyzed with ABI7500 SDS software (Applied Biosystems). To maintain consistency,the baseline was set automatically by the software. The compara-tive CT method (2�DDCT method) was used to analyze the expres-sion level of CsTRx [51]. All samples were analyzed in threeduplications and the results are expressed as relative fold of onesample as mean � standard deviation.
2.5. Cloning of CsTRx and recombinant expression and purification
All of the cloning experiments were carried out according toSambrook et al. [52] with slight modifications [53]. The primer setof CsTRx F5: (GA)3GAATTC CAA GTT AAG GAC AAG GCT GAT TTCEcoRI and CsTRx R6: (GA)3 AAGCTT TGC ATC AGC ACC AGA GAA CHindIII was designed with the corresponding restriction enzymesites at the N- and C-termini for CsTRx, in order to clone the codingsequence into the expression vector, pMAL-c2X (BioLabs Inc.,).Using plasmid DNA of CsTRx as a template and Taq DNA polymerase(Qiagen), PCR was carried out to amplify the coding sequence. ThePCR product was purified using the GeneJET� Gel Extraction Kit(Thermo Scientific). After PCR, the amplified product was purifiedby using the GeneJET� (Thermo Scientific). Further, the clonedproducts were sequenced using the ABI Prism-Bigdye TerminatorCycle Sequencing Ready Reaction kit by using the same primersused for the amplification and analyzed using an ABI 3730sequencer. Then, both insert and vector were digested with therespective restriction enzymes. The ligated product was trans-formed into XL1 blue cells and the correct recombinant product (asconfirmed by restriction enzyme digestion and sequencing) wastransformed into competent E. coli BL21 (DE3) cells for proteinexpression.
Transformed E. coli BL21 (DE3) cells were incubated in ampi-cillin (100 mg/mL) Luria broth (LB) overnight. This culture was thenused to inoculate 100 mL of LB broth in 0.2% glucose-rich mediumwith ampicillin at 37 �C until cell density reached 0.7 at OD600.E. coli BL21 (DE3) harboring pMAL-c2x-CsTRx was induced for over
Please cite this article in press as: Palanisamy R, et al., A redox active siteproperties, Fish & Shellfish Immunology (2013), http://dx.doi.org/10.1016
expression with 1 mM isopropyl-b-thiogalactopyranoside (IPTG)and incubated at 16 �C for 4 h. Cells were harvested by centrifu-gation (4000 � g for 20 min at 4 �C). E. coli BL21 (DE3) uninducedculture was used as a negative control. Then the cells were re-suspended in column buffer (TriseHCl, pH 7.4, 200 mM NaCl) andfrozen at �20 �C overnight. After thawing on ice, cells were dis-rupted by sonication. The crude CsTRx fusion protein fused withmaltose binding protein (MBP) was purified using pMAL� proteinfusion and purification system protocol (BioLabs). Further thefusion protein was digested and purified as described in our earlierreport [53]. The purified proteinwas subjected toMS/MS analysis todetermine the quality of recombinant CsTRx protein following themethodology of Mann and Aebersold [54]. Then the protein wasverified by 12% SDS-PAGE and the molecular weight of targetprotein was evaluated using protein molecular weight standards.Proteins were visualized by staining with 0.05% Coomassie blue R-250. The concentration of the purified protein was determined viathe Bradfordmethod [55], using bovine serum albumin (BSA) as thestandard. The purified protein was kept at �80 �C until determi-nation of biological activity.
2.6. Functional studies of recombinant CsTRx protein
2.6.1. Thermal stability and pH toleranceFor determination of optimum pH, the purified recombinant
CsTRx preparation was added to various reaction buffers with pHadjusted from 5.0 to 12.0, then incubated for 30 min at 30 �C asdescribed by Hansen et al. [56]. For determination of the optimumtemperature, aliquots of the purified recombinant CsTRx proteinwere incubated with reaction buffer and assayed as described byJiang et al. [57]. For both assay, after incubation, 50 mM H2O2 wasadded to the reaction mixture and again incubated for another30 min. After incubation, 0.9 mL (8% v/v) of trichloroacetic acid(TCA) was added to the reaction mixture. Finally, to terminate thereaction, 10 mM ferrous ammonium sulfate (0.2 mL) and 2.5 Mpotassium thiocyanate (0.1 mL) were added into the reactionmixture. Then, the relative activity of the reaction mixture wasmeasured at 480 nm.
2.6.2. Cell proliferation assayThe proliferation assay was performed as described by Kim et al.
[58] with slight modifications. The assay was performed usingspleen leucocytes which were prepared as described by Grahamand Secombes [59] and Montero et al. [60]. C. striatus spleen leu-cocytes (1 � 107 cells per well) were prepared by centrifugationprocess (500 � g for 30 min at 4 �C). Further, the proliferation ac-tivity was conducted using WST-1 cell proliferation assay kit(Clontech) as the manufacturer suggested. The cells were furtheradjusted to 2 � 104 cells per well and re-suspended in Roswell ParkMemorial Institute (RPMI) 1640 medium (Life Technologies) alongwith recombinant CsTRx protein at various concentrations (1, 5, 10,25, 50 and 100 mg/mL). Followed by, for the proliferation activity,10 mL of theWST-1 reagent (Clontech) was added into eachwell andincubated at 25 �C for 12 h. After incubation, the optical density wasmeasured in a microplate reader (Promega) at 450 nm. Blank withRPMI 1640 medium was treated as control. The assay was per-formed in three replications.
2.6.3. Antioxidant activityThe antioxidant capacity of the recombinant CsTRx protein
was performed as described by Mu et al. [61]. The activity ofrecombinant CsTRx protein was measured with antioxidant ca-pacity detection kit (Abcam). Briefly, 350 mL recombinant CsTRxprotein was added into the reaction mixture of 3.50 mL. Then, themixture was incubated at 37 �C for 30 min. After incubation, the
containing murrel cytosolic thioredoxin: Analysis of immunological/j.fsi.2013.10.016

Fig. 1. Nucleotide (above) and deduced amino acid (below) sequence of C. striatus TRx. A putative TRx domain between V2 and N106 is gray shaded. The catalytic center (31WCGPC35)is shaded in orange color. The active cysteine residues (Cys32 and Cys35) of the catalytic center are circled. The non-catalytic cysteine residues at Cys71 and Cys75 are shaded inyellow color. The start codon (ATG) and stop codon (TAA) are underlined. The asterisk (*) indicates the stop codon. The secondary structural elements of CsTRx are shown beneaththe amino acid sequence: blue line, coil; red wave, a-helix; green arrow, b-sheet. The structure was predicted using POLYVIEW 2D online tool. (For interpretation of the references tocolor in this figure legend, the reader is referred to the web version of this article.)
R. Palanisamy et al. / Fish & Shellfish Immunology xxx (2013) 1e104
reaction was terminated by adding 100 mL stop solution. Themixture was kept at room temperature for 10 min before A520was determined. An antioxidant capacity unit was calculated asan increase by 0.01 at A520 min�1 mg protein. The antioxidantcapacity of glutathione (GSH) was also calculated as that of re-combinant CsTRx protein.
2.6.4. Enzymatic oxidoreductase assayThe assay was performed using the modified procedure of
turbidimetric assay of insulin disulfide reduction as described byAispuro-Hernandez et al. [27] and Holmgren [62]. As suggested[61,27], the enzymatic oxidoreductase activity of recombinantCsTRx protein was determined based on the insulin disulfidereduction by thioredoxin. In brief, the reaction mixture was pre-pared in cuvettes by adding 23 mL of insulin (1.74 mM), 100 mL re-combinant CsTRx protein, 60 mL of EDTA (10 mM) and PBS (0.1 mM)to give a final volume of 300 mL. The reaction was initiated byadding 3 mL of dithiothreitol in all cuvettes, except the blank. Thecuvettes were then thoroughly mixed and placed in the microplatereader (Promega). The readings were taken at 650 nm. During thereaction, the non-enzymatic reduction of insulin by dithiothreitolwas recorded in a control cuvette without adding recombinantCsTRx protein. The specific activity was calculated as described byHolmgren [62].
2.7. Statistics
For comparison of CsTRx gene expression, cell proliferation ac-tivity and enzymatic oxidoreductase activity, statistical analysiswas performed using one-way ANOVA and mean comparisonswere performed by Tukey’s Multiple Range Test using SPSS 11.5 atthe 5% significance level.
Please cite this article in press as: Palanisamy R, et al., A redox active siteproperties, Fish & Shellfish Immunology (2013), http://dx.doi.org/10.1016
3. Result
3.1. Sequence characterization and homologous analysis
A full-length cDNA sequence encoded thioredoxin was iden-tified from the established C. striatus cDNA library using GS-FLX�technique. The sequence was deposited in EMBL GenBank data-base under the accession number HF674396. CsTRx cDNA iscomprised of 327 base pairs (bp) along with an open readingframe (ORF) of 324 bp. The ORF encodes a polypeptide of 108amino acid residues with predicted molecular weight of 12 kDaand theoretical isoelectric point (pI) of 4.7. The domain and motifanalysis showed that CsTRx contains a long thioredoxin domainbetween V2 and N106. The domain possessed a thioredoxin activefamily at 24e42 along with a redox active site (otherwise knownas catalytic center) at 31WCGPC35 along with a conserved disulfidebond (32CXXC35). The domain also contains another disulfidebond at 71CXXXC75, which is considered as non-catalytic cysteineresidue (Fig. 1). Further analysis showed that CsTRx contains sixdifferent amino acid residues at T31, W32, V61, D62, C75 and M77,which are found to be involved in the formation of the hydro-phobic dimerization interface. Among these, Asp62 plays a majorrole, since it maintains the pH of the dimerization interface.Moreover, the analysis showed that CsTRx neither have a trans-membrane region nor a signal sequence. Other than these domainand motifs, CsTRx also contains a casein kinase II phosphorylationsite at Ser92-Gly93-Ala94-Asp95. The sub-cellular localization pre-diction analysis showed that CsTRx is a cytosol thioredoxin. Thesecondary structural analysis showed that CsTRx contains 40.7%a-helix, 19.4% b-sheet and 39.8% random coil (Fig. 1). Furtheranalysis showed that CsTRx protein contains higher amount oflysine (13%), valine (10%) and aspartic acid (11%).
containing murrel cytosolic thioredoxin: Analysis of immunological/j.fsi.2013.10.016

R. Palanisamy et al. / Fish & Shellfish Immunology xxx (2013) 1e10 5
The homologous analysis showed that CsTRx shared significantsimilarity with TRx-1 from zebrafish Danio rerio (84%) followed byrock bream O. fasciatus and Korean rockfish S. schlegelii (81%) (datanot shown). Moreover, multiple sequence alignment showed thatCsTRx possessed many conserved region including redox active site(31WCGPC35) in the thioredoxin active family site, non-catalyticcysteine residues at Cys71 and Cys75, the amino acid residuesinvolved in the formation of the hydrophobic dimerization inter-face T31, W32, V61, D62, C76 and M77 and the casein kinase II phos-phorylation site (Fig. 2).
3.2. Phylogenetic analysis
A phylogenetic tree was constructed to find out the geneticdistance and class of CsTRx. The tree produced three distinct groupsincluding TRx-1, TRx-2 and TRx-3. Further, TRx-1 group formed astwo separate clusters such as vertebrate TRx-1 and invertebrateTRx-1. Our CsTRx was clustered together with vertebrate TRx-1(Fig. 3). Hence, based on the phylogenetic analysis as well asother analysis including homologous analysis by BLAST programand domain and motif search analysis by Prosite program, it ispredicted that CsTRx belongs to TRx-1 group.
3.3. Tissue-specific mRNA expression and transcriptional changes ofCsTRx upon infection
The relative gene expression of CsTRx in various tissues wasexamined by real-time PCR using cDNA (as template) synthesizedfrom the extracted total RNA of blood, liver, spleen, heart, kidney,head kidney, intestine, muscle, skin, gill and brain along with genespecific primers and other reaction mixtures. b-actin gene wasconsidered as a housekeeping gene. The tissue-specific mRNAtranscription was measured by comparing the expression quantitydiscovered in tissues taken for analysis with that from skin (Fig. 4A).The gene expression was noticed in all the tissues considered foranalysis, representing its important immunological function in allthe tissues. Significantly (P< 0.05) the highest gene expressionwasobserved in gill, spleen, kidney and head kidney, moderateexpression in heart, liver and blood and the lowest expression inmuscle, intestine, brain and skin.
10 20 30 ....|....|....|....|....|....|.C. striatus MVRQVKDKADFDNLLK-EAGDKLVVVDFTAS. schlegelii MVREVTSLAEFQSILKEEAGDRLVVVDFTAD. rerio MVLEIEDKAAFDNALK-NAGDKLVVVDFTAO. fasciatus MVREVENLDEFKAILK-EAGDRLVVVDFTAA. fimbria MIDDVKTLAEFKKILA-DAGDKLVVVDFTAE. sinensis MVYQVKDQEDFKKQLK-EAGQKLVVVDFYAG. gallus MVKSVGNLADFEAELK-AAGEKLVVVDFSAH. sapiens MVKQIESKTAFQEALD-AAGDKLVVVDFSA
70 80 90
....|....|....|....|....|....|.C. striatus DVDEADDVSSACKITCMPTFQFYKSGEKVDS. schlegelii DVDEADDVSSHCGINCMPTFHFYKNGEKVFD. rerio DVDDAQDVAALCGISCMPTFHFYKNGKKVDO. fasciatus DVDEASDVSEHCKISCMPTFHFYKNEAKVSA. fimbria DVDEADEVSKDCGISCMPTFHFYKNGIKVDE. sinensis DVDECEDVAVTYQISCMPTFLFFKEEQKIDG. gallus DVDDAQDVATHCDVKCMPTFQFYKNGKKVQH. sapiens DVDDCQDVASECEVKCTPTFQFFKKGQKVG
Fig. 2. The multiple sequence alignments of CsTRx amino acid sequence with other known Tzebrafish Danio rerio (NP_001002461), rock bream Oplegnathus fasciatus (BAK38716), sablefiJungle fowl Gallus gallus (NP_990784), Chinese mitten Crab Eriocheir sinensis (ACQ59118) andaccession number. Amino acids identical to CsTRx sequence are shown in black shade anhighlighted in a red color box. The residues involved in the formation of the hydrophobic diinterpretation of the references to color in this figure legend, the reader is referred to the
Please cite this article in press as: Palanisamy R, et al., A redox active siteproperties, Fish & Shellfish Immunology (2013), http://dx.doi.org/10.1016
In addition, CsTRx transcriptional modulation in gill was eval-uated under fungal, bacterial and H2O2 challenge by qRT-PCRanalysis. Immune challenge of C. striatus with A. invadans,A. hydrophila and H2O2 resulted in significant modulations of CsTRxgene expression in gill. The transcriptional changes of CsTRx mRNAexpression in the gills of C. striatus after challenged with fungus,bacteria and H2O2 showed time-dependent transcriptional patternsat various time points during the study. During the first 48 h afterA. invadans injection, CsTRx gene expression was significantly(P < 0.05) increased compared to their corresponding control, andthen slightly decreased at 72 h post-injection (p.i) (Fig. 4B). For thefishes injected with A. hydrophila, the CsTRx mRNA expression wassignificantly (P < 0.05) up-regulated until 24 h p.i, then theexpression decreased and finally at 72 h p.i the expression wasobserved nearly to the basal level (Fig. 4C). In H2O2 injectedC. striatus, the CsTRx gene expression was significantly (P < 0.05)up-regulated from 3 h p.i to 12 h p.i, then the expression wasdecreased and finally at 72 h p.i, the expression was observedexactly at the basal level (Fig. 4D).
3.4. Production of recombinant CsTRx protein
The recombinant CsTRx protein was over-expressed and puri-fied using an E. coli expression vector system. The recombinantCsTRx protein was purified from the supernatant of induced cells.The molecular weight of the recombinant CsTRx protein along withfusion protein (MBP) was found to be 54.5 kDa as shown by SDS-PAGE (42.5 kDa for MBP and 12 kDa for recombinant CsTRx pro-tein). In order to purify the recombinant CsTRx protein from theMBP fusion protein, DEA-sepharose ion exchange chromatographywas performed (data not shown).
3.5. Functional properties of recombinant CsTRx protein
3.5.1. Thermal stability and pH tolerance of recombinant CsTRxprotein
In order to characterize the biochemical properties of recom-binant CsTRx protein, we conducted a H2O2 peroxidase assay undervarious pH and temperature. The relative activity of recombinantCsTRx protein against various pH buffers is shown in Fig. 5. The
40 50 60 ...|....|....|....|....|....| TWCGPCRFIGPIFQKMSEDPENKNVVFLKV 59 TWCGPCKIIGPIFEQQAAKPENKNVLFLKV 60TWCGPCQTIGPYFKLLSEKPENKNVVFLKV 59 TWCGPCKMIGPEFERLSKLPENKNVIFLKV 59 TWCGPCKMISPVFQQEDGKPENKNVIFVKV 59 TWCGPCKMIAPKLQEMSSQ--MTDVVFLKV 57TWCGPCKMIKPFFHSLCDK--FGDVVFIEI 57 TWCGPCKMINPFFHSLSEK--YSNVIFLEV 57
100
...|....|....|.... Identity (%)EFSGADAEKLKETLEKNRT 108 100.0 EFSGANKDTLLEKLAALRT 109 68.8 EFSGSNQSKLEEKINSHK- 107 67.2 EFSGANKDTLAEKVEALRT 108 65.7 EFSGANKDTLLEKLVALRT 108 63.8 SFSGASEEKIRDYIAKYK- 105 57.0 EFSGANKEKLEETIKSLV- 105 56.7 EFSGANKEKLEATINELV- 105 37.2
Rx sequences. The sequences used for the multiple sequence alignments are as follows:sh Anoplopoma fimbria (ACQ58191), Korean rockfish Sebastes schlegelii (BAK82164), redhuman Homo sapiens (AAF86467). The number given in the parentheses are GenBank
d absent amino acids are indicated by dashes (�). The redox active motif WCGPC ismerization interface (T31, W32, V61, D62, C76 and M77) are indicated by arrow mark. (Forweb version of this article.)
containing murrel cytosolic thioredoxin: Analysis of immunological/j.fsi.2013.10.016

Oplegnathus fasciatus (BAK38716)
Sebastes schlegelii (BAK82164)
Anoplopoma fimbria (ACQ58191)
Trachidermus fasciatus (AFH68184)
Channa striatus TRx
Danio rerio (NP_001002461)
Gallus gallus (NP_990784)
Homo sapiens (AAF86466)
Tupaia chinensis (ELW70571)
Rattus norvegicus (AAH58454)
Mus musculus (AAH94415)
Mesobuthus gibbosus (CAE54157)
Litopenaeus vannamei (ACA60746)
Eriocheir sinensis (ACQ59118)
Scylla paramamosain (AGC96525)
Portunus trituberculatus (AFE88625)
Triticum aestivum (AAL67139)
Plantago major (CAH59452)
Arabidopsis thaliana (AAC49353)
Glycine max (ABV71992)
Xenopus laevis (AAH43794)
Oreochromis mossambicus (AAX61130)
Melopsittacus undulates (AAO72715)
Rattus norvegicus (AAC53008)
Homo sapiens (AAF86467)94
56
35
100
85
96
84
99
83
99
62
57
71
88
98
92
92
52
43
54
88
42
0.1
Inv
erte
bra
tes
V
erte
bra
tes
TRx-3
TRx-1
TRx-2
Fig. 3. Consensus Neighbor-Joining phylogenetic tree of CsTRx along with other homologous sequences constructed using Mega 5.05. The bootstrap confidence values given at theforks of the tree are based on 1000 bootstrap replications. The GenBank accession numbers are given in the parentheses.
R. Palanisamy et al. / Fish & Shellfish Immunology xxx (2013) 1e106
optimum pH for the relative activity of recombinant CsTRx proteinwas found to be 8. Similarly, the relative activity of recombinantCsTRx protein against various temperatures is given in Fig. 6. Theoptimum temperature for H2O2 reduction was examined at varioustemperatures, ranging between 10 �C and 70 �C. The results showedthat the recombinant CsTRx protein has the maximum relativeactivity at 35 �C.
3.5.2. Proliferation assayTo study the influence of recombinant CsTRx protein on
C. striatus spleen leukocyte activation, the leukocytes weretreated with recombinant CsTRx protein at various concentra-tions, and the cell proliferation was determined. We observedthat the spleen leukocytes treated with recombinant CsTRxprotein at various concentrations produced a greater cell prolif-eration activity (Fig. 7). Further analysis indicated that the re-combinant CsTRx protein was functioning in a concentrationdependant manner.
3.5.3. Antioxidant capacity and enzymatic oxidoreductase activityof CsTRx
Antioxidant capacity assay was conducted to study the ferricreducing power of recombinant CsTRx protein. The results showedthat the antioxidant capacity of recombinant CsTRx protein wasdetermined to be 4.2 U/mg protein (data not shown). Moreover, weconducted an insulin disulfides assay to study the enzymaticoxidoreductase activity of CsTRx. In this assay, we observed noactivity in the control group. But the recombinant CsTRx protein
Please cite this article in press as: Palanisamy R, et al., A redox active siteproperties, Fish & Shellfish Immunology (2013), http://dx.doi.org/10.1016
addition rapidly increased the enzymatic oxidoreductase activity(Fig. 8).
4. Discussion
Thioredoxins are 12 kDa redox proteins which are present in allforms of living organism. TRx plays an important role in themaintenance of cellular thiol redox balance [11,63,64], which isessential for the exact function of each single cell [31]. In the cur-rent investigation, a full-length CsTRx cDNAwas identified from theestablished cDNA library of C. striatus, further the sequence wasobtained by cloning. The obtained sequence belongs to a cytosolicTRx with a molecular mass of 12 kDa and further analysis showedthat it belongs toTRx-1 familymember. CsTRx polypeptide does nothave a signal sequence, thus Kanzok et al. [65] stated that thosepolypeptides without signal sequence are not likely to be disclosedto the N-glycosylation system and therefore the protein may not beglycosylated, but it contains essential motifs for glycosylation.However, Spyrou et al. [31] reported that translocation of thio-redoxin from the cytoplasm to the nucleus in response of stress,even in the absence of a recognized nuclear-translocation signal,has been demonstrated. Notably, the structurally essential redoxactive site Try-Cys-Gly-Pro-Cys (WCGPC) are strongly conserved inCsTRx similar to earlier reported cytosolic TRx [10,30]. The catalyticmotif WCGPC has been showed to be vital for the regulation ofprotein function through the redox structural state or catalytic thiolgroups [66]. Revathy et al. [67] reported that the cysteine residuesundergo reversible oxidation when thioredoxin reduces the
containing murrel cytosolic thioredoxin: Analysis of immunological/j.fsi.2013.10.016
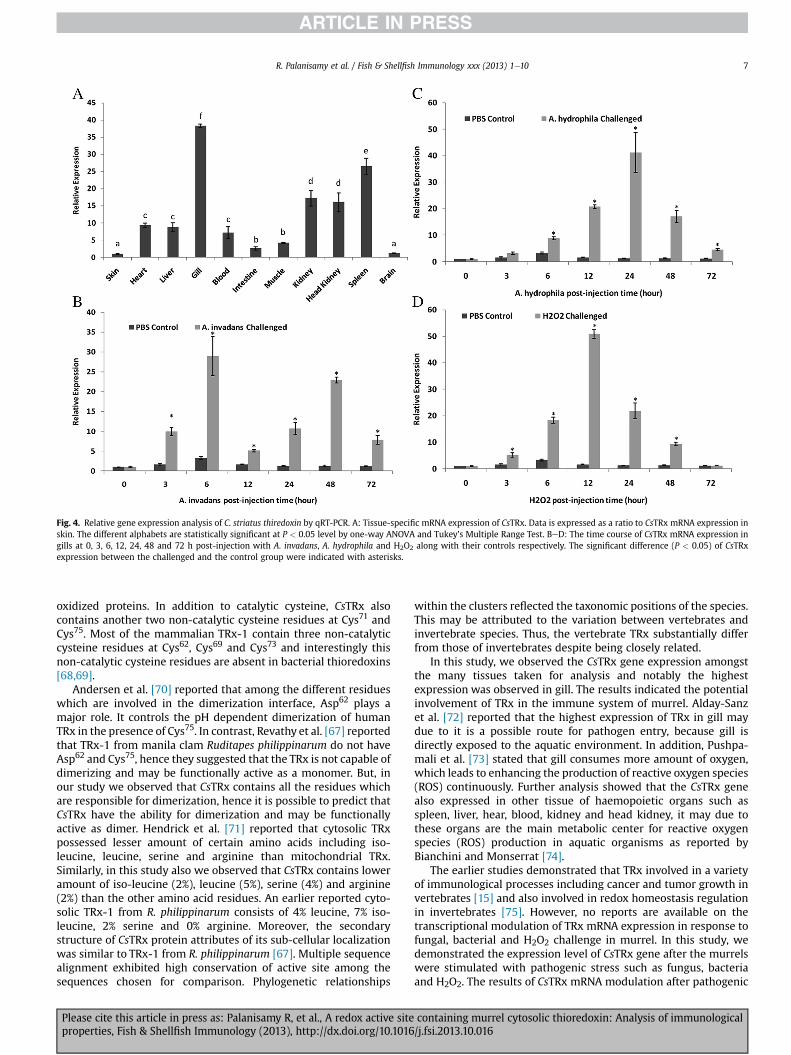
Fig. 4. Relative gene expression analysis of C. striatus thiredoxin by qRT-PCR. A: Tissue-specific mRNA expression of CsTRx. Data is expressed as a ratio to CsTRx mRNA expression inskin. The different alphabets are statistically significant at P < 0.05 level by one-way ANOVA and Tukey’s Multiple Range Test. BeD: The time course of CsTRx mRNA expression ingills at 0, 3, 6, 12, 24, 48 and 72 h post-injection with A. invadans, A. hydrophila and H2O2 along with their controls respectively. The significant difference (P < 0.05) of CsTRxexpression between the challenged and the control group were indicated with asterisks.
R. Palanisamy et al. / Fish & Shellfish Immunology xxx (2013) 1e10 7
oxidized proteins. In addition to catalytic cysteine, CsTRx alsocontains another two non-catalytic cysteine residues at Cys71 andCys75. Most of the mammalian TRx-1 contain three non-catalyticcysteine residues at Cys62, Cys69 and Cys73 and interestingly thisnon-catalytic cysteine residues are absent in bacterial thioredoxins[68,69].
Andersen et al. [70] reported that among the different residueswhich are involved in the dimerization interface, Asp62 plays amajor role. It controls the pH dependent dimerization of humanTRx in the presence of Cys75. In contrast, Revathy et al. [67] reportedthat TRx-1 from manila clam Ruditapes philippinarum do not haveAsp62 and Cys75, hence they suggested that the TRx is not capable ofdimerizing and may be functionally active as a monomer. But, inour study we observed that CsTRx contains all the residues whichare responsible for dimerization, hence it is possible to predict thatCsTRx have the ability for dimerization and may be functionallyactive as dimer. Hendrick et al. [71] reported that cytosolic TRxpossessed lesser amount of certain amino acids including iso-leucine, leucine, serine and arginine than mitochondrial TRx.Similarly, in this study also we observed that CsTRx contains loweramount of iso-leucine (2%), leucine (5%), serine (4%) and arginine(2%) than the other amino acid residues. An earlier reported cyto-solic TRx-1 from R. philippinarum consists of 4% leucine, 7% iso-leucine, 2% serine and 0% arginine. Moreover, the secondarystructure of CsTRx protein attributes of its sub-cellular localizationwas similar to TRx-1 from R. philippinarum [67]. Multiple sequencealignment exhibited high conservation of active site among thesequences chosen for comparison. Phylogenetic relationships
Please cite this article in press as: Palanisamy R, et al., A redox active siteproperties, Fish & Shellfish Immunology (2013), http://dx.doi.org/10.1016
within the clusters reflected the taxonomic positions of the species.This may be attributed to the variation between vertebrates andinvertebrate species. Thus, the vertebrate TRx substantially differfrom those of invertebrates despite being closely related.
In this study, we observed the CsTRx gene expression amongstthe many tissues taken for analysis and notably the highestexpression was observed in gill. The results indicated the potentialinvolvement of TRx in the immune system of murrel. Alday-Sanzet al. [72] reported that the highest expression of TRx in gill maydue to it is a possible route for pathogen entry, because gill isdirectly exposed to the aquatic environment. In addition, Pushpa-mali et al. [73] stated that gill consumes more amount of oxygen,which leads to enhancing the production of reactive oxygen species(ROS) continuously. Further analysis showed that the CsTRx genealso expressed in other tissue of haemopoietic organs such asspleen, liver, hear, blood, kidney and head kidney, it may due tothese organs are the main metabolic center for reactive oxygenspecies (ROS) production in aquatic organisms as reported byBianchini and Monserrat [74].
The earlier studies demonstrated that TRx involved in a varietyof immunological processes including cancer and tumor growth invertebrates [15] and also involved in redox homeostasis regulationin invertebrates [75]. However, no reports are available on thetranscriptional modulation of TRx mRNA expression in response tofungal, bacterial and H2O2 challenge in murrel. In this study, wedemonstrated the expression level of CsTRx gene after the murrelswere stimulated with pathogenic stress such as fungus, bacteriaand H2O2. The results of CsTRx mRNA modulation after pathogenic
containing murrel cytosolic thioredoxin: Analysis of immunological/j.fsi.2013.10.016
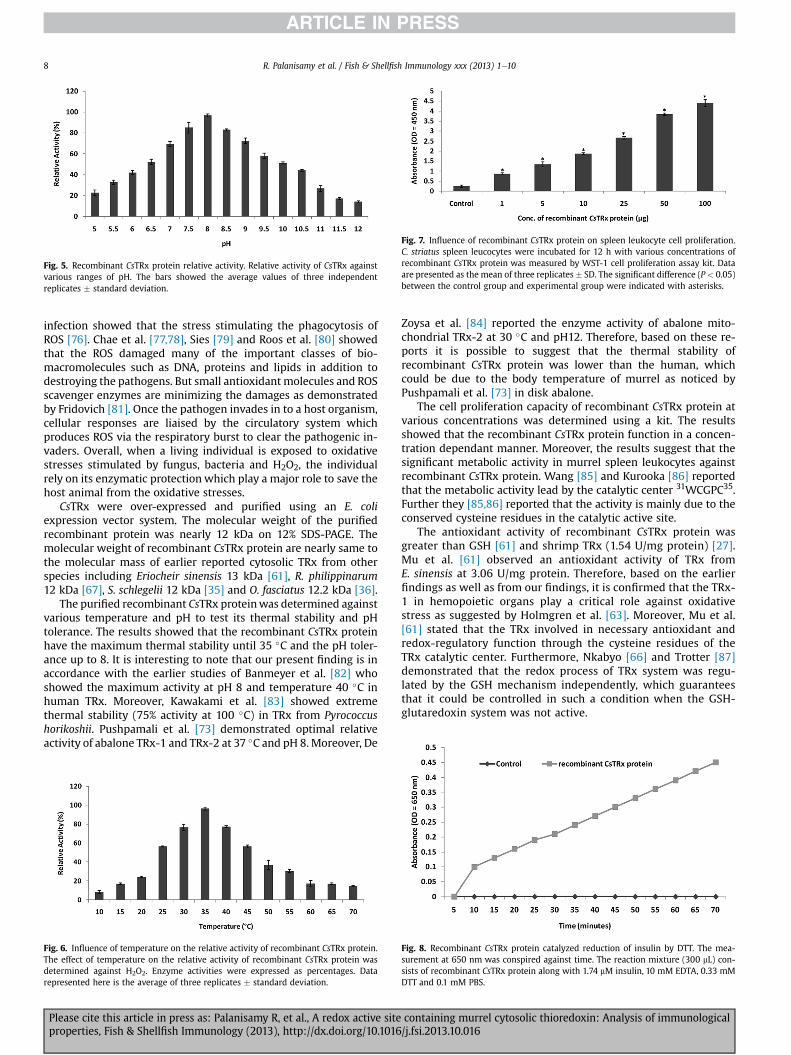
Fig. 5. Recombinant CsTRx protein relative activity. Relative activity of CsTRx againstvarious ranges of pH. The bars showed the average values of three independentreplicates � standard deviation.
Fig. 7. Influence of recombinant CsTRx protein on spleen leukocyte cell proliferation.C. striatus spleen leucocytes were incubated for 12 h with various concentrations ofrecombinant CsTRx protein was measured by WST-1 cell proliferation assay kit. Dataare presented as the mean of three replicates � SD. The significant difference (P < 0.05)between the control group and experimental group were indicated with asterisks.
R. Palanisamy et al. / Fish & Shellfish Immunology xxx (2013) 1e108
infection showed that the stress stimulating the phagocytosis ofROS [76]. Chae et al. [77,78], Sies [79] and Roos et al. [80] showedthat the ROS damaged many of the important classes of bio-macromolecules such as DNA, proteins and lipids in addition todestroying the pathogens. But small antioxidantmolecules and ROSscavenger enzymes are minimizing the damages as demonstratedby Fridovich [81]. Once the pathogen invades in to a host organism,cellular responses are liaised by the circulatory system whichproduces ROS via the respiratory burst to clear the pathogenic in-vaders. Overall, when a living individual is exposed to oxidativestresses stimulated by fungus, bacteria and H2O2, the individualrely on its enzymatic protection which play a major role to save thehost animal from the oxidative stresses.
CsTRx were over-expressed and purified using an E. coliexpression vector system. The molecular weight of the purifiedrecombinant protein was nearly 12 kDa on 12% SDS-PAGE. Themolecular weight of recombinant CsTRx protein are nearly same tothe molecular mass of earlier reported cytosolic TRx from otherspecies including Eriocheir sinensis 13 kDa [61], R. philippinarum12 kDa [67], S. schlegelii 12 kDa [35] and O. fasciatus 12.2 kDa [36].
The purified recombinant CsTRx proteinwas determined againstvarious temperature and pH to test its thermal stability and pHtolerance. The results showed that the recombinant CsTRx proteinhave the maximum thermal stability until 35 �C and the pH toler-ance up to 8. It is interesting to note that our present finding is inaccordance with the earlier studies of Banmeyer et al. [82] whoshowed the maximum activity at pH 8 and temperature 40 �C inhuman TRx. Moreover, Kawakami et al. [83] showed extremethermal stability (75% activity at 100 �C) in TRx from Pyrococcushorikoshii. Pushpamali et al. [73] demonstrated optimal relativeactivity of abalone TRx-1 and TRx-2 at 37 �C and pH 8. Moreover, De
Fig. 6. Influence of temperature on the relative activity of recombinant CsTRx protein.The effect of temperature on the relative activity of recombinant CsTRx protein wasdetermined against H2O2. Enzyme activities were expressed as percentages. Datarepresented here is the average of three replicates � standard deviation.
Please cite this article in press as: Palanisamy R, et al., A redox active siteproperties, Fish & Shellfish Immunology (2013), http://dx.doi.org/10.1016
Zoysa et al. [84] reported the enzyme activity of abalone mito-chondrial TRx-2 at 30 �C and pH12. Therefore, based on these re-ports it is possible to suggest that the thermal stability ofrecombinant CsTRx protein was lower than the human, whichcould be due to the body temperature of murrel as noticed byPushpamali et al. [73] in disk abalone.
The cell proliferation capacity of recombinant CsTRx protein atvarious concentrations was determined using a kit. The resultsshowed that the recombinant CsTRx protein function in a concen-tration dependant manner. Moreover, the results suggest that thesignificant metabolic activity in murrel spleen leukocytes againstrecombinant CsTRx protein. Wang [85] and Kurooka [86] reportedthat the metabolic activity lead by the catalytic center 31WCGPC35.Further they [85,86] reported that the activity is mainly due to theconserved cysteine residues in the catalytic active site.
The antioxidant activity of recombinant CsTRx protein wasgreater than GSH [61] and shrimp TRx (1.54 U/mg protein) [27].Mu et al. [61] observed an antioxidant activity of TRx fromE. sinensis at 3.06 U/mg protein. Therefore, based on the earlierfindings as well as from our findings, it is confirmed that the TRx-1 in hemopoietic organs play a critical role against oxidativestress as suggested by Holmgren et al. [63]. Moreover, Mu et al.[61] stated that the TRx involved in necessary antioxidant andredox-regulatory function through the cysteine residues of theTRx catalytic center. Furthermore, Nkabyo [66] and Trotter [87]demonstrated that the redox process of TRx system was regu-lated by the GSH mechanism independently, which guaranteesthat it could be controlled in such a condition when the GSH-glutaredoxin system was not active.
Fig. 8. Recombinant CsTRx protein catalyzed reduction of insulin by DTT. The mea-surement at 650 nm was conspired against time. The reaction mixture (300 mL) con-sists of recombinant CsTRx protein along with 1.74 mM insulin, 10 mM EDTA, 0.33 mMDTT and 0.1 mM PBS.
containing murrel cytosolic thioredoxin: Analysis of immunological/j.fsi.2013.10.016

R. Palanisamy et al. / Fish & Shellfish Immunology xxx (2013) 1e10 9
The disulfide reductase capacity of recombinant CsTRx proteinwas analyzed using the modified procedure of turbidimetric assay(insulin disulfide reductase assay) and we found to be 4.89. Similarcomparable results were reported from E. sinensis (5.03) [61], E. coli(4.93), calf thymus (6.50) and calf liver (5.09) [62]. Holmgren [13]reported that the TRx contains a generalized disulfide reductasesystem, thus catalyzes the NADPH-dependent reductions of theexposed disulfide bridges in different protein. Overall, the resultindicates that the CsTRx genewas a biologically activemolecule andplayed critical roles in the physiological and immunological pro-cesses through the catalytic active site (CXXC) of TRx, thus regulatesthe protein metabolism.
5. Conclusion
Conclusively, a cytosolic TRx-1 was identified from C. striatuscDNA library and the sequence was obtained by cloning. The ob-tained sequence was characterized at molecular level using variousbioinformatics tools. CsTRx was found to be expressed in manytissues of C. striatus, but the highest expressionwas observed in gilland other hemopoietic organs. Further the expression was up-regulated at various time points induced by different immunestimulants including fungus (A. invadans), bacteria (A. hydrophila)and H2O2. The recombinant product of CsTRx was over-expressedand purified using an E. coli expression vector system. The puri-fied recombinant CsTRx protein was further assayed to study itfunctional properties including thermal stability, pH tolerance, cellproliferation activity, antioxidant activity and oxidoreductase ac-tivity. All these activity assays showed the important role of CsTRxin oxidative stress. Further, we observed that the characterizedCsTRx gene contains a potential antioxidant property, which couldregulate the oxidative stress created by various biological patho-gens as well as chemical stress in the immune system of C. striatus,thus protecting it.
Acknowledgments
This research is supported by DBT’s Prestigious Ram-alingaswami Re-entry Fellowship (D.O.NO.BT/HRD/35/02/2006)funded by Department of Biotechnology, Ministry of Science andTechnology, Government of India, New Delhi.
References
[1] Nakano H, Nakajima A, Sakon-Komazawa S, Piao JH, Xue X, Okumura K.Reactive oxygen species mediate crosstalk between NF-kappa B and JNK. CellDeath Differ 2006;13:730e7.
[2] Kohchi C, Inagawa H, Nishizawa T, Soma G. ROS and innate immunity. Anti-cancer Res 2009;29:817e21.
[3] Molina-Cruz A, DeJong RJ, Charles B, Gupta L, Kumar S, Jaramillo-Gutierrez G,et al. Reactive oxygen species modulate Anopheles gambiae immunity againstbacteria and Plasmodium. J Biol Chem 2008;283:3217e23.
[4] Ghosh J, Myers CE. Inhibition of arachidonate 5-lipoxygenase triggers massiveapoptosis in human prostate cancer cells. Proc Natl Acad Sci U S A 1998;95:13182e7.
[5] Yin GY, Yin YF, He XF. Effect of zhuchun pill on immunity and endocrinefunction of elderly with kidney-yang deficiency. Zhongguo Zhong Xi Yi Jie HeZa Zhi 1995;15:601e3.
[6] Bae YS, Kang SW, Seo MS, Baines IC, Tekle E, Chock PB, et al. Epidermal growthfactor (EGF)-induced generation of hydrogen peroxide. Role in EGF receptor-mediated tyrosine phosphorylation. J Biol Chem 1997;272:217e21.
[7] Lee YJ, Galoforo SS, Berns CM, Chen JC, Davis BH, Sim JE, et al. Glucosedeprivation induced cytotoxicity and alterations in mitogen-activated proteinkinase activation are mediated by oxidative stress in multidrug-resistanthuman breast carcinoma cells. J Biol Chem 1998;273:5294e9.
[8] Griswold CM, Matthews AL, Bewley KE, Mahaffey JW. Molecular character-ization and rescue of acatalasemic mutants of Drosophila melanogaster. Ge-netics 1993;134:781e8.
[9] Maulik N, Das DK. Emerging potential of thioredoxin and thioredoxin inter-acting proteins in various disease conditions. Biochim Biophys Acta2008;1780:1368e82.
Please cite this article in press as: Palanisamy R, et al., A redox active siteproperties, Fish & Shellfish Immunology (2013), http://dx.doi.org/10.1016
[10] McEligot AJ, Yang S, Meyskens Jr FL. Redox regulation by intrinsic speciesand extrinsic nutrients in normal and cancer cells. Annu Rev Nutr 2005;25:261e95.
[11] Arner ES, Holmgren A. Physiological functions of thioredoxin and thioredoxinreductase. Eur J Biochem 2000;267:6102e9.
[12] Holmgren A. Thioredoxin. Annu Rev Biochem 1985;54:237e71.[13] Holmgren A. Thioredoxin and glutaredoxin systems. J Biol Chem 1989;264:
13963e6.[14] Laurent TC, Moore EC, Reichard P. Enzymatic synthesis of de-
oxyribonucleotides. IV. Isolation and characterization of thioredoxin, thehydrogen donor from Escherichia coli B. J Biol Chem 1964;239:3436e44.
[15] Powis G, Montfort WR. Properties and biological activities of thioredoxins.Annu Rev Pharmacol Toxicol 2001;41:261e95.
[16] Rubartelli A, Bonifaci N, Sitia R. High rates of thioredoxin secretion correlatewith growth arrest in hepatoma cells. Cancer Res 1995;55:675e80.
[17] Matthews JR, Wakasugi N, Virelizier JL, Yodoi J, Hay RT. Thioredoxin regulatesthe DNA binding activity of NF-kappa B by reduction of a disulphide bondinvolving cysteine 62. Nucl Acid Res 1992;20:3821e30.
[18] Berndt C, Lillig CH, Holmgren A. Thioredoxins and glutaredoxins as facilitatorsof protein folding. Biochim Biophys Acta 2008;1783:641e50.
[19] Schallreuter KU, Wood JM. The role of thioredoxin reductase in the reductionof free radicals at the surface of the epidermis. Biochem Biophys Res Commun1986;136:630e7.
[20] Kondo N, Nakamura H, Masutani H, Yodoi J. Redox regulation of human thi-oredoxin network. Antioxid Redox Signal 2006;8:1881e90.
[21] Yoshioka J, Schreiter ER, Lee RT. Role of thioredoxin in cell growth throughinteractions with signaling molecules. Antioxid Redox Signal 2006;8:2143e51.
[22] Rubartelli A, Bajetto A, Allavena G, Wollman E, Sitia R. Secretion of thio-redoxin by normal and neoplastic cells through a leaderless secretorypathway. J Biol Chem 1992;267:24161e4.
[23] Hori K, Katayama M, Sato N, Ishii K, Waga S, Yodoi J. Neuroprotection by glialcells through adult T cell leukemia derived factor/human thioredoxin (ADF/TRX). Brain Res 1994;652:304e10.
[24] Berggren M, Gallegos A, Gasdaska JR, Gasdaska PY, Warneke J, Powis G. Thi-oredoxin and thioredoxin reductase gene expression in human tumors andcell lines, and the effects of serum stimulation and hypoxia. Anticancer Res1996;16:3459e66.
[25] Angelini G, Gardella S, Ardy M, Ciriolo MR, Filomeni G, Di Trapani G. Antigen-presenting dendritic cells provide the reducing extracellular microenviron-ment required for T lymphocyte activation. Proc Natl Acad Sci U S A 2002;99:1491e6.
[26] Schenk H, Vogt M, Droge W, Schulze-Osthoff K. Thioredoxin as a potentcostimulus of cytokine expression. J Immunol 1996;156:765e71.
[27] Aispuro-Hernandez E, Garcia-Orozco KD, Muhlia-Almazan A, Del-Toro-Sanchez L, Robles-Sanchez RM, Hernandez J, et al. Shrimp thioredoxin is apotent antioxidant protein. Comp Biochem Physiol C 2008;148:94e9.
[28] Terashima H, Gotoh S, Yagi K, Mizoguchi T. cDNA sequence of bovine thio-redoxin. DNA Seq 1999;10:331e3.
[29] Yu GW, Xu JY, Xu L, Lee KS. cDNA sequence of porcine thioredoxin. DNA Seq2002;13:113e5.
[30] Hirota K, Nakamura H, Masutani H, Yodoi J. Thioredoxin superfamily andthioredoxin-inducing agents. Ann N Y Acad Sci 2002;957:189e99.
[31] Spyrou G, Enmark E, Miranda-Vizuete A, Gustafsson J. Cloning and expressionof a novel mammalian thioredoxin. J Biol Chem 1997;272:2936e41.
[32] Spyrou G, Miranda-Vizuete A, Damdimopoulos AE. The mitochondrial thio-redoxin system. Antioxid Redox Signal 2000;2:801e10.
[33] Wahl MC, Irmler A, Hecker B, Schirmer RH, Becker K. Comparative structuralanalysis of oxidized and reduced thioredoxin from Drosophila melanogaster.J Mol Biol 2005;345:1119e30.
[34] Nordberg J, Arner ESJ. Reactive oxygen species, antioxidants and themammalian thioredoxin system. Free Radic Biol Med 2001;31:1287e312.
[35] Park CI, Jung JH, Shim WJ, Kim JW, Kim EG, Jeong JM, et al. Molecular char-acterization, expression, and functional analysis of two thioredoxins in theblack rockfish (Sebastes schlegelii). Fish Shellfish Immunol 2012;32:808e15.
[36] Kim DH, Kim JW, Jeong JM, Park HJ, Park CI. Molecular cloning and expressionanalysis of a thioredoxin from rock bream, Oplegnathus fasciatus, and biologicalactivity of the recombinant protein. Fish Shellfish Immunol 2011;31:22e8.
[37] Haniffa MA, Marimuthu K, Nagarajan M, Arockiaraj J, Kumar D. Breedingbehaviour and parental care of the induced bred spotted murrel Channapunctatus under captivity. Curr Sci 2004;86:1375e6.
[38] Marimuthu K, Haniffa MA, Muruganandam M, Arockiaraj J. Low cost murrelseed production technique for fish farmers. Naga 2001;24:21e2.
[39] Arockiaraj J, Haniffa MA, Perumalsamy PRR, Marimuthu K, Muruganandam M.An effective treatment to the spotted murrel Channa punctatus for epizooticulcerative syndrome (EUS). Fishing Chimes 2003;23:27e8.
[40] Arthimanju R, Haniffa MA, Arunsingh SV, Ramakrishanan MC, Dhanaraj M,Xavier Innocent B, et al. Effect of dietary administration of Effinol� FG ongrowth and enzymatic activities of Channa striatus (Bloch, 1793). J Anim VetAdv 2011;10:796e801.
[41] Arockiaraj J, Muruganandam M, Marimuthu K, Haniffa MA. Utilization ofcarbohydrates as a dietary energy source by striped murrel Channa striatus(Bloch) fingerlings. Acta Zool Taiwan 1999;10:103e11.
[42] Abirami A, Venkatesh K, Akila S, Rajesh P, Prabha N, Prasanth B, et al. Fish lilytype lectin-1 contains b-prism 2 architecture: immunological characteriza-tion. Mol Immunol 2013;56:497e506.
containing murrel cytosolic thioredoxin: Analysis of immunological/j.fsi.2013.10.016

R. Palanisamy et al. / Fish & Shellfish Immunology xxx (2013) 1e1010
[43] Arockiaraj J, Annie JG, Dhanaraj M, Mukesh P, Milton J, Arun S. An upstreaminitiator caspase 10 of snakehead murrel Channa striatus, containing DED, p20and p10 subunits: molecular cloning, gene expression and proteolytic activity.Fish Shellfish Immunol 2013;34:505e13.
[44] Arockiaraj J, Annie JG, Dhanaraj M, Thirumalai MK, Mukesh P, Milton J, et al.Macrobrachium rosenbergii cathepsin L: molecular characterization and geneexpression in response to viral and bacterial infections. Microbiol Res2013;168:569e79.
[45] Arockiaraj J, Annie JG, Gopi P, Milton J, Mukesh P, Prasanth B, et al. A novelprophenoloxidase, hemocyanin encoded copper containing active enzymefrom prawn: gene characterization. Gene 2013;524:139e51.
[46] Patterton HG, Graves S. DNAssist: the integrated editing and analysis of mo-lecular biology sequences in windows. Bioinforma Appl Note 2000;16:652e3.
[47] Uinuk-Ool TS, Takezaki N, Kuroda N, Figueroa F, Sato A, Samonte IE, et al.Phylogeny of antigen-processing enzymes: cathepsins of a cephalochordate,an agnathan and a bony fish. Scand J Immunol 2003;58:436e48.
[48] Dhanaraj M, Haniffa MA, Ramakrishnan CM, Arunsingh SV. Microbial florafrom the Epizootic Ulcerative Syndrome (EUS) infected murrel Channa striatus(Bloch, 1797) in Tirunelveli region. Turk J Vet Anim Sci 2008;32:221e4.
[49] Arockiaraj J, Annie JG, Dhanaraj M, Ranganath G, Milton J, Arun S, et al.Crustin, a WAP domain containing antimicrobial peptide from freshwaterprawn M. rosenbergii: immune characterization. Fish Shellfish Immunol2012;34:109e18.
[50] Arockiaraj J, Puganeshwaran V, Sarasvathi E, Arun S, Othman RY, Subha B.Molecular functions of chaperonin gene, containing tailless complex poly-peptide 1 from Macrobrachium rosenbergii. Gene 2012;508:241e9.
[51] Livak KJ, Schmittgenm TD. Analysis of relative gene expression data usingreal-time quantitative PCR and the 2(�Delta Delta C(T)) method. Methods2001;25:402e8.
[52] Arockiaraj J, Sarasvathi E, Puganeshwaran V, Arun S, Othman RY, Subha B.Immunological role of thiol-dependent peroxiredoxin gene in Macrobrachiumrosenbergii. Fish Shellfish Immunol 2012;33:121e9.
[53] Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: a laboratory manual.2nd ed. New York: Cold Spring Harbor Laboratory, Cold Spring Harbor Lab-oratory Press; 1989.
[54] Mann M, Aebersold R. Mass spectrometry-based proteomics. Nature2003;422:198e207.
[55] Bradford MM. A rapid and sensitive method for the quantification of micro-gram quantities of protein utilizing the principle of protein-dye binding. AnalBiochem 1976;72:248e54.
[56] Hansen JM, Zhang H, Jones DP. Differential oxidation of thioredoxin-1, thio-redoxin-2, and glutathione by metal ions. Free Radic Biol Med 2006;40:138e45.
[57] Jiang Q, Yan YH, Hu GK, Shang YZ. Molecular cloning and characterization of aperoxiredoxin from Phanerochaete chrysosporium. Cell Mol Biol Lett 2005;10:659e68.
[58] Kim JW, Kim EG, Kim DH, Shim SH, Park CI. Molecular identification andexpression analysis of the CC chemokine gene in rock bream (Oplegnathusfasciatus) and the biological activity of the recombinant protein. Fish ShellfishImmunol 2013;34:892e901.
[59] Graham S, Secombes CJ. The production of macrophage-activating factor fromrainbow trout Salmo gairdneri leucocytes. Immunology 1988;65:293e7.
[60] Montero J, Ordas MC, Alejo A, Gonzalez-Torres L, Sevilla N, Tafalla C. CK12, arainbow trout chemokine with lymphocyte chemo-attractant capacity asso-ciated to mucosal tissues. Mol Immunol 2011;48:1102e13.
[61] Mu C, Zhao J, Wang L, Song L, Song X, Zhang H, et al. A thioredoxin withantioxidant activity identified from Eriocheir sinensis. Fish Shellfish Immunol2009;26:716e23.
[62] Holmgren A. Thioredoxin catalyzes the reduction of insulin disulfides bydithiothreitol and dihydrolipoamide. J Biol Chem 1979;254:9627e32.
[63] Holmgren A, Johansson C, Berndt C, Lonn ME, Hudemann C, Lillig CH. Thiolredox control via thioredoxin and glutaredoxin systems. Biochem Soc Trans2005;33:1375e7.
[64] Myers JM, Antholine WE, Myers CR. Hexavalent chromium causes theoxidation of thioredoxin in human bronchial epithelial cells. Toxicology2008;246:222e33.
[65] Kanzok SM, Fechner A, Bauer H, Ulschmid JK, Muller HM, Botella-Munoz J,et al. Substitution of the thioredoxin system for glutathione reductase inDrosophila melanogaster. Science 2001;291:643e6.
Please cite this article in press as: Palanisamy R, et al., A redox active siteproperties, Fish & Shellfish Immunology (2013), http://dx.doi.org/10.1016
[66] Nkabyo YS, Ziegler TR, Gu LH, Watson WH, Jones DP. Glutathione and thio-redoxin redox during differentiation in human colon epithelial (Caco-2) cells.Am J Physiol Gastrointest Liver Physiol 2002;283:G1352e9.
[67] Revathy KS, Umasuthan N, Lee Y, Whang I, Kim HC, Lee J. Cytosolic thio-redoxin from Ruditapes philippinarum: molecular cloning, characterization,expression and DNA protection activity of the recombinant protein. Dev CompImmunol 2012;36:85e92.
[68] Deiss LP, Kimchi AA. Genetic tool used to identify thioredoxin as a mediator ofa growth inhibitory signal. Science 1991;252:117e20.
[69] Gasdaska PY, Oblong JE, Cotgreave IA, Powis G. The predicted amino acidsequence of human thioredoxin is identical to that of the autocrine growthfactor human adult T-cell derived factor (ADF): thioredoxin mRNA is elevatedin some human tumors. Biochim Biophys Acta 1994;1218:292e6.
[70] Andersen JF, Sanders DA, Gasdaska JR, Weichsel A, Powis G, Montfort WR.Human thioredoxin homodimers: regulation by pH, role of aspartate 60, andcrystal structure of the aspartate 60-asparagine mutant. Biochemistry1997;36:13979e88.
[71] Hendrick JP, Hodges PE, Rosenberg LE. Survey of amino-terminal proteolyticcleavage sites in mitochondrial precursor proteins: leader peptides cleaved bytwo matrix proteases share a three-amino acid motif. Proc Natl Acad Sci U S A1989;86:4056e60.
[72] Alday-Sanz V, Roque A, Turnbull JF. Clearing mechanisms of Vibrio vulnificusbiotype I in the black tiger shrimp Penaeus monodon. Dis Aquat Org 2002;48:91e9.
[73] Pushpamali WA, De Zoysa M, Kang HS, Oh CH, Whang I, Kim SJ, et al.Comparative study of two thioredoxin peroxidases from disk abalone (Haliotisdiscus discus): cloning, recombinant protein purification, characterization ofantioxidant activities and expression analysis. Fish Shellfish Immunol2008;24:294e307.
[74] Bianchini A, Monserrat JM. Effects of methyl parathion on Chasmagnathusgranulatus hepatopancreas: protective role of sesamol. Ecotoxicol Environ Saf2007;67:100e8.
[75] Kanzok SM, Schirmer RH, Turbachova I, Iozef R, Becker K. The thioredoxinsystem of the malaria parasite Plasmodium falciparum. Glutathione reductionrevisited. J Biol Chem 2000;275:40180e6.
[76] Segal A. How neutrophils kill microbes. Annu Rev Immunol 2005;23:197e223.
[77] Chae HZ, Robison K, Poole LB, Church G, Storz G, Rhee SG. Cloning andsequencing of thiol-specific antioxidant from mammalian brain: alkyl hy-droperoxide reductase and thiol-specific antioxidant define a large family ofantioxidant enzymes. Proc Natl Acad Sci U S A 1994;91:7017e21.
[78] Chae HZ, Kim IH, Kim K, Rhee SG. Cloning, sequencing, and mutation of thiolspecific antioxidant gene of Saccharomyces cerevisiae. J Biol Chem 1993;268:16815e21.
[79] Sies H. Strategies of antioxidant defense. Eur J Biochem 1993;215:213e9.[80] Roos D, van Bruggen R, Meischl C. Oxidative killing of microbes by neutro-
phils. Microbes Infect 2003;5:1307e15.[81] Fridovich I. The biology of oxygen radicals. Science 1978;201:875e80.[82] Banmeyer I, Marchand C, Clippe A, Knoops B. Human mitochondrial perox-
iredoxin 5 protects from mitochondrial DNA damages induced by hydrogenperoxide. FEBS Lett 2005;579:2327e33.
[83] Kawakami R, Sakuraba H, Kamohara S, Goda S, Kawarabayasi Y, Ohshima T.Oxidative stress response in an anaerobic hyperthermophilic archaeon: pres-ence of a functional peroxiredoxin in Pyrococcus horikoshii. J Biochem2004;136:541e7.
[84] De Zoysa M, Pushpamali WA, Whang I, Kim SJ, Lee J. Mitochondrialthioredoxin-2 from disk abalone (Haliotis discus discus): molecular charac-terization, tissue expression and DNA protection activity of its recombinantprotein. Comp Biochem Physiol B 2008;149:630e9.
[85] Wang XW, Liou YC, Ho B, Ding JL. An evolutionarily conserved 16-kDathioredoxin-related protein is an antioxidant which regulates the NF-kBsignaling pathway. Free Radic Biol Med 2007;42:247e59.
[86] Kurooka H, Kato K, Minoguchi S, Takahashi Y, Ikeda J, Habu S, et al. Cloningand characterization of the nucleoredoxin gene that encodes a novel nuclearprotein related to thioredoxin. Genomics 1997;39:331e9.
[87] Trotter EW, Grant CM. Non-reciprocal regulation of the redox state of theglutathione-glutaredoxin and thioredoxin systems. EMBO Rep 2003;4:184e8.
containing murrel cytosolic thioredoxin: Analysis of immunological/j.fsi.2013.10.016
Copyright © 2022 FDOKUMEN




















