
Immunological profiling of Hepatitis B virus Surface gene in Pakistan
10
Novus International Journal of Biotechnology & Bioscience Immunological profiling of Hepatitis B virus surface gene in Pakistan Abbas Raza a* , Kashif Mehmood a , Fakhira Begum a , Mustafeez Mujtaba Babar a , Suleman Riaz a , Umer Javed a , Zaira Rehman a , Sahar Sadaf Zaidi a , Aleem Siddiqui b and Ishtiaq Qadri c a NUST Center of Virology and Immunology, National University of Sciences and Technology, Islamabad, Pakistan b University of California, San Diago, USA c Husein Ebrahim Jamal Research Institute of Chemistry, University of Karachi, Karachi, Pakistan. ___________________________________________________________________________ ABSTRACT Hepatitis B virus (HBV) is the major etiological agent of chronic hepatitis infections worldwide. Pakistan alone stands as one of the world’s highest load of chronic hepatitis and mortality due to liver disease, cirrhosis and hepatocellular carcinoma. Universal vaccination strategy started by World Health Organisation as a means to reduce the large number of affected patients of hepatitis B during 1990-1991 but this seem not to be the solution as Pakistan still has 9 million people who are HBV positive. A large number of genomic variations have been reported in HBV over the past two decades and these induce a specific change in virus biology. It is believed that variants like those involved in host immune system evasion (S-mutants) dramatically modify the virus-host interactions when present and are a serious challenge to healthcare personnel. The altered immunogenicity of surface protein in HBV can make HBsAg undetectable (occult infections), or induces immune escape variants to be selected for and thus evade virus clearance (escape mutants). To investigate this kind of immune evasion operative in this part of the subcontinent, a prospective study was conducted in 100 HBV infected patients identified randomly in different tertiary care hospitals of Islamabad, which lead to the characterization of sequence variations in HBsAg. A total of 13 different TA-Cloned sequences of surface ORF were analysed and the mutants circulating in Pakistan include observed replacements in both humoral and/or cellular [major histocompatibility complex class I (MHC-I) and MHC-II] HBV mutated epitopes, such as Q 30K, T45S, T46P, L49R, S55P, T68I, I86T, Y100C, S114T, T115I, T118V, P120T, R122K, P127T, P127S, A128V, G130N, T131N Y134V Y134F S143T I152V G159A F161Y A168T, A168V, F170S, V 180A, V194A, M198V, S207N. A 3D model of small surface protein and HBIg is developed using computational biology tools and these are docked to identify important residues involved in immune evasion. The insilico analysis further confirms the presence of virus evaders circulating in Pakistani population. Development of a good vaccine able to neutralise all variants of HBV has been a challenge. Novel treatment options have to be employed to treat accumulated viral mutations as seen in this study. These may include a modified vaccine (incorporating the native strain of HBV) along with an enhancement of B and T cell immune responses e.g. a multiepitope vaccine, and an HBIG directed against a mutated cluster of alpha determinant to treat such patients. There is a dire need that pathologists, pharmaceutical and the healthcare industry augment their awareness of HBV Novus International Journal of Biotechnology & Bioscience 2012, Vol. 1, No. 2 www.novusscientia.org
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Immunological profiling of Hepatitis B virus Surface gene in Pakistan

Novus International Journal of Biotechnology & Bioscience�������������������������������������������������������������
�
�
�
�
Immunological profiling of Hepatitis B virus surface gene in Pakistan
Abbas Razaa*, Kashif Mehmooda, Fakhira Beguma, Mustafeez Mujtaba Babara, Suleman Riaza, Umer Javeda, Zaira Rehmana, Sahar Sadaf Zaidia, Aleem Siddiquib and Ishtiaq Qadric
a NUST Center of Virology and Immunology, National University of Sciences and Technology, Islamabad, Pakistan
b University of California, San Diago, USA c Husein Ebrahim Jamal Research Institute of Chemistry, University of Karachi, Karachi, Pakistan.
___________________________________________________________________________
ABSTRACT
Hepatitis B virus (HBV) is the major etiological agent of chronic hepatitis infections worldwide. Pakistan alone stands as one of the world’s highest load of chronic hepatitis and mortality due to liver disease, cirrhosis and hepatocellular carcinoma. Universal vaccination strategy started by World Health Organisation as a means to reduce the large number of affected patients of hepatitis B during 1990-1991 but this seem not to be the solution as Pakistan still has 9 million people who are HBV positive. A large number of genomic variations have been reported in HBV over the past two decades and these induce a specific change in virus biology. It is believed that variants like those involved in host immune system evasion (S-mutants) dramatically modify the virus-host interactions when present and are a serious challenge to healthcare personnel. The altered immunogenicity of surface protein in HBV can make HBsAg undetectable (occult infections), or induces immune escape variants to be selected for and thus evade virus clearance (escape mutants). To investigate this kind of immune evasion operative in this part of the subcontinent, a prospective study was conducted in 100 HBV infected patients identified randomly in different tertiary care hospitals of Islamabad, which lead to the characterization of sequence variations in HBsAg. A total of 13 different TA-Cloned sequences of surface ORF were analysed and the mutants circulating in Pakistan include observed replacements in both humoral and/or cellular [major histocompatibility complex class I (MHC-I) and MHC-II] HBV mutated epitopes, such as Q 30K, T45S, T46P, L49R, S55P, T68I, I86T, Y100C, S114T, T115I, T118V, P120T, R122K, P127T, P127S, A128V, G130N, T131N Y134V Y134F S143T I152V G159A F161Y A168T, A168V, F170S, V 180A, V194A, M198V, S207N. A 3D model of small surface protein and HBIg is developed using computational biology tools and these are docked to identify important residues involved in immune evasion. The insilico analysis further confirms the presence of virus evaders circulating in Pakistani population.
Development of a good vaccine able to neutralise all variants of HBV has been a challenge. Novel treatment options have to be employed to treat accumulated viral mutations as seen in this study. These may include a modified vaccine (incorporating the native strain of HBV) along with an enhancement of B and T cell immune responses e.g. a multiepitope vaccine, and an HBIG directed against a mutated cluster of alpha determinant to treat such patients. There is a dire need that pathologists, pharmaceutical and the healthcare industry augment their awareness of HBV
�
Novus International Journal of Biotechnology & Bioscience
2012, Vol. 1, No. 2�
www.novusscientia.org
� ����� �����

Novus International Journal of Biotechnology & Bioscience�������������������������������������������������������������
mutants and how these mutants may alter existing diagnostic and treatment options in the context of Pakistan.
KEYWORDS: Escape mutants, HBV, ‘a’ determinant, abnitio modelling
*Corresponding author: Abbas Raza Department of Biology, School of Science and Engineering (SSE),
Lahore University of Management Sciences, D.H.A. Lahore Cantt. 54792, Pakistan
Email address: [email protected]
INTRODUCTION
Hepatitis B virus (HBV) is the major etiological agent of chronic hepatitis infections, having 400 million carriers worldwide and is responsible for about a million deaths annually ( Liaw et al., 2010). Of the estimated 50 million new cases reported of HBV infection diagnosed annually, 5-10 % adults and up to 90 % infants shall become chronically infected, and this prolonged immunologic response to HBV infection leads to the development of cirrhosis, liver failure, or hepatocellular carcinoma (HCC) in 40 % of the patients (Rustgi et al., 2010). It is alarming to note that a country like Pakistan alone stands as one of the world’s highest load of chronic hepatitis and mortality due to liver disease (Alam et al., 2007; Mujeeb and Pearce, 2008; Noorali et al., 2008; Ali et al., 2011).
Hepatitis B virus is the prototype member of the family hepadnaviradae (Pungpapong et al., 2007), has a partially double stranded 3.2 kb DNA genome that codes for four Open Reading Frames translated into seven different proteins (Figure 1).
Figure 1. Schematic representation of hepatitis B virus genome.
Hepatitis B surface antigen (HBsAg) contains a vastly conformational epitope cluster called major hydrophilic region (MHR) extending from amino acids 99-169, where a common � determinant is located at amino acid 124-147 (see figure 2) (Xia et al., 2001; Lu et al., 2006). This region has important humoral and cellular viral epitopes like major histocompatibility complex, MHC-I and MHC-II (Tong et al., 2010). Antibodies produced by natural infection or by administration of vaccine are directed against � determinant and confer cross-protective immunity to all subtype of this virus. However, amino acid substitution, insertion or deletion in MHR may lead to the modification of conformational epitope and altered immunogenicity of HBsAg thus making HBsAg undetectable (occult infections), or induces immune escape variants to evade virus clearance (escape mutants).

Novus International Journal of Biotechnology & Bioscience�������������������������������������������������������������
Figure 2. Schematic representation of S protein (positions 99 to 210 amino acids), including the first and second
loops, according to the second model proposed by Carman et al., 1997 (see reference).
There is no study regarding the surface mutants circulating among the native population with reference to genome comparisons of surface gene variants so this study was designed to analyze the epitopes linked to immune evasion.
MATERIALS AND METHODS
Blood Sample Collection
A total of 13 HBsAg ELISA positive blood samples from HBV carrier subjects were processed at NUST Center of Virology and Immunology Diagnostic Center NUST Islamabad between 2009-2010 for detection of HBV DNA by qualititave PCR. A blood sample of 3ml was collected in a vacutainer from each patient and the serum was separated to be stored at -20°C until processed. Standard protocol for reducing contamination was strictly followed.
Extraction of Viral DNA and Polymerase Chain Reaction of Surface gene
HBV viral DNA was isolated using PureLink™ Viral RNA������������� 12280-050, Invitrogen, USA). Standard manufacturer’s protocol was followed. The extracted DNA was stored at -20°C. Polymerase Chain Reaction (PCR) was carried out to amplify HBV S gene using the sample serum DNA. The reaction mixture contained 800ng sample DNA template along with 10µM gene specific forward (5’ GGGCTCGAGAAAAGAATGGAGAACATCACATCAGGATTC 3’) and reverse primer (5’ GGGGCGGCCGCTTAAATGTATACCCAAAGACAAAAGAAAATTGG 3’), 2mM dNTPs, 2mM MgCl2, 1.5 units of Taq polymerase (Fermentas, USA) along with 1X Taq Polymerase Buffer and nuclease free water making a total reaction volume of 20µL. PCR mixture was then placed in Thermal Cycler (Swift™ MaxPro Thermal Cycler, Esco, Singapore). The amplification round consisted of one denaturation at 94ºC for 5 minute, followed by 35 cycles comprising a 45 second denaturation step at 94ºC, a 45 second annealing step at 67ºC, and a 1 minute extension step at 72ºC, each. The amplification was concluded by one final extension at 72ºC for 10 minutes.

Novus International Journal of Biotechnology & Bioscience�������������������������������������������������������������
TA cloning of Surface gene
The gel purified PCR amplified surface gene was T/A cloned using Dual Promoter TA cloning kit (Invitrogen # K2050-40) according to the manufacturer’s protocol. 10µL of ligation mixture was transformed into 200µL of DH5� Escherichia coli competent cells and the white colonies were selected for plasmid prep and restriction digestion analysis. Plasmid DNA was isolated using the GeneJETTM Plasmid Miniprep Kit (# K0502, Fermentas, USA) according to manufacturer’s protocol. The insertion of the gene into the plasmid was confirmed by subjecting the isolated plasmid to digestion with EcoR1 enzyme (Fermentas, USA) as the vector contains restriction sites on both sides of the cloned gene. The isolated plasmid (5µL or 150ng) was treated with 10 units of enzyme (1µL), 10 X Digest reaction buffer (1µL) and nuclease free water (3µL). The reaction mixture was incubated at 37oC for 3 hours and confirmation was made by resolving the digested product on 1% agarose gel. The gel was stained with ethidium bromide and visualized on gel documentation system (Wealtec Dolphin Doc, Sparks, USA) under UV light.
Sequencing of positive clones:
The plasmids isolated in section 2.3 were sent to Sure Bio-diagnostic Canada for sequencing with universal M13 forward and reverse primers on ABI sequencer.
In-Silico Studies of Surface protein:
Sequences of Hepatitis B Virus surface (S) gene globally reported were retrieved from NCBI nucleotide data bank. Using the CLC workbench 6.1 (http://www.clcbio.com), all the sequences were aligned and mutations were compared in the, B and T cell epitope involved in immune evasion.
RESULTS
PCR Amplification of HBV Surface gene
Using gene specific primers, HBV S gene was PCR amplified from DNA samples extracted of HBV patients. Figure 3 shows a band of size 681 bp that indicates successful amplification of small surface ORF.
Figure 3: Representative digital printout on 1% agarose gel of PCR amplified HBV S gene. Lane M: 100 bps ladder (Fermentas, USA). Lanes 1 – 13: HBV S gene amplified from different patients. The numbers indicate
different sample codes assigned to patients. PC means positive control.
���

Novus International Journal of Biotechnology & Bioscience�������������������������������������������������������������
Restriction Digests of TA clones The positive colonies were cultured and plasmid was isolated and then digested for the confirmation of cloning of gene of interest. Figure 4 shows the confirmatory digested product of 681 bp for all the digested clones. A total of 13 different patients had their surface gene cloned and processed using this approach.
Figure 4 :(A) Representative digital picture of surface gene T/A clones run on1% agarose gel and their restriction digestion with ECoR1 enzyme. Lane M: 1 kb ladder (Fermentas, USA). Lanes 1, 3, 5, 7, 9, 11, 13, 15 & 17: showing the uncut (UC) surface gene clones from different patients. Lanes 2, 4, 6, 8, 10, 12, 14, 16 & 18:
digested (C) (B) Representative digital picture of surface gene T/A clones run on 1% agarose gel and their restriction
digestion with ECoR1 enzyme. Lane M: 1 kb ladder (Fermentas, USA). Lanes 1, 3, 5 & 7: showing the uncut (UC) surface gene clones from different patients. Lanes 2, 4, 6 & 8: digested (C).
Sequencing Data
The plasmid was read in both the forward and reverse directions using M13 universal primers and the sequences were submitted in nucleotide database of NCBI and have been assigned the accession numbers JN684761, JN687948, JN558565, JN558566, JN132123, JN132122, JN132121, JN132120, JN132119, JN132118, HQ641713, HQ641712, and HQ641711.
Epitope Maps of Surface protein and the mutated version of Pakistani Isolates
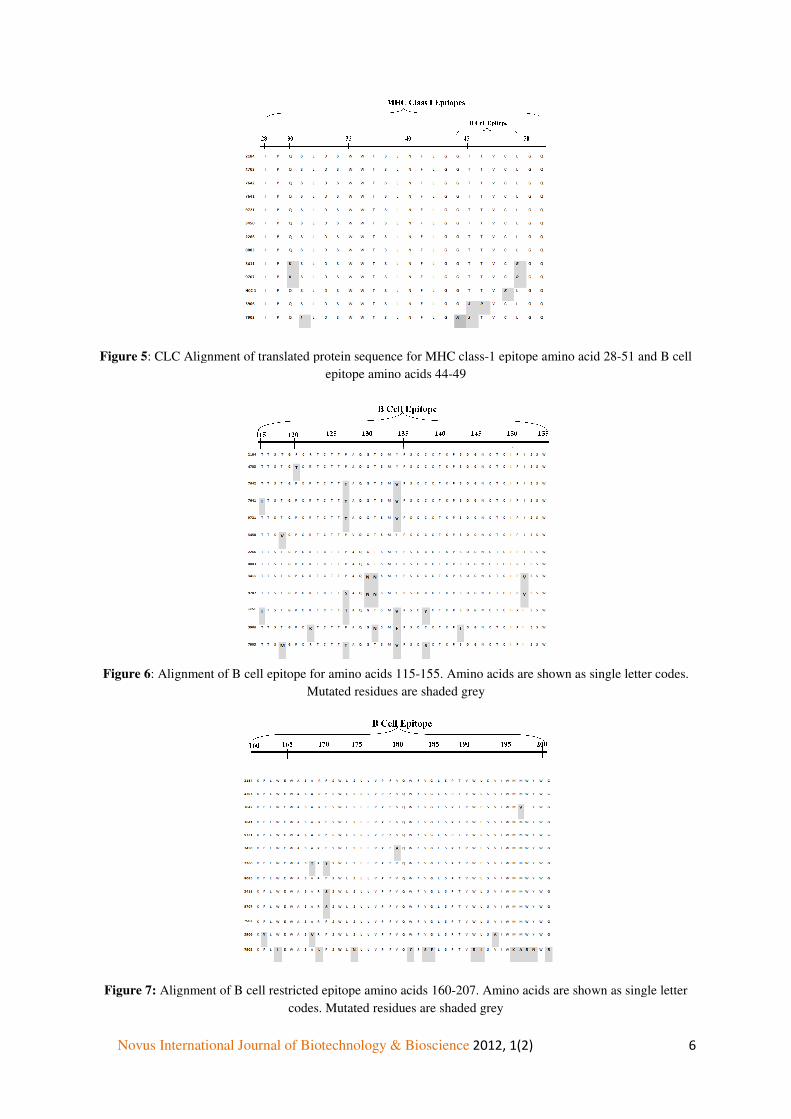
Alignment of translated protein sequence of MHC class-1 epitope amino acid 28-51, B cell epitope amino acids 44-49 (Figure 5), 115-155 (Figure 6) and 160-207 (Figure 7); MHC class-II restricted epitope amino acids 136-155 and 213-226 (Figure 8 and 9) was performed using CLC work bench 6.5.1. B cell epitope amino acid 44-49 and 115-155 is highly mutated. Patient 3906, 5294, 7903 show the maximum variance in MHC I and MHC II amino acids.
��
��
Novus International Journal of Biotechnology & Bioscience�
Figure 5: CLC Alignment of translated protein sequence for MHC class
Figure 6: Alignment of B cell epitope for amino acids 115
Figure 7: Alignment of B cell restricted epitope amino acids 160
Novus International Journal of Biotechnology & Bioscience����������������������������������������������������������
: CLC Alignment of translated protein sequence for MHC class-1 epitope amino acid 28epitope amino acids 44-49
: Alignment of B cell epitope for amino acids 115-155. Amino acids are shown as single letter codes.
Mutated residues are shaded grey
Alignment of B cell restricted epitope amino acids 160-207. Amino acids are shown as single letter codes. Mutated residues are shaded grey
��������������������������������������������������
1 epitope amino acid 28-51 and B cell
acids are shown as single letter codes.
207. Amino acids are shown as single letter

Novus International Journal of Biotechnology & Bioscience�������������������������������������������������������������
Figure 8: Alignment of MHC class-II restricted epitope amino acids 136-155.Amino acids are shown as single letter codes. Mutated residues are shaded grey
Figure 9: Alignment of MHC class-II restricted epitope amino acids 213-226. Amino acids are shown as single letter codes. Mutated residues are shaded grey.
DISCUSSION
A successful prophylactic program in Pakistan needs to be justified with the molecular dynamics of HBV circulating in the native population. Till date surface gene variants have never been characterized or given importance in the HBV infected pool of Pakistan. The alarming rise in HBV pool in Pakistan together with the extensive use of HBV vaccine are quite contrasting and debatable issues. There is little information regarding random population statistics for surface mutations in Pakistan. Added to the fact that in populations like Pakistan where the reverse transcriptase inhibitor lamivudine has been used widely to treat patients continuously for periods of several years, viruses with alterations in surface and pol ORF are likely to occur frequently (Khokhar et al., 2005). There are reports that these

Novus International Journal of Biotechnology & Bioscience�������������������������������������������������������������
resistant mutants can spread to and adversely affect the vaccinated individuals (Villet et al., 2008; Sayan and Akhan, 2011). This study was therefore designed with the specific aims to identify the status of variants in vaccine escape mutants of surface gene so that the information can be utilized by the personnel involved in pharmaceutical companies (development of drugs/inhibitors for reverse transcriptase of HBV), healthcare (diagnostic errors of HBV and occult infections) as well as the authorities (design future policies) for the proper management of Hepatitis B virus infections in Pakistan.
This study also identifies a cluster of mutations (Q 30K, Q30P, S31I, L39F, G44A, T45S, T46P, C48R, L49R) that are found between amino acid residues 20 and 50 of surface and lie within a region (residues 28–51) predicted to be a putative HLA class I restricted cytotoxic T cell epitope (Chisari and Ferrari, 1995). These mutations may help the virus evade cellular immunity and enable the virus to persist.
Five amino acid changes (Y72N, I86T, I86F, L97P, Y100C) was observed flanked by residues 73 and 103, a region shown to be a transmembrane domain(Stirk et al., 1992) and therefore improbable to be subject to immune pressure. Nevertheless, mutations have been reported in this region (Ogura et al., 1999; Liu et al., 2002); and there is also supporting evidence that a HLA-A2- restricted epitope is located in this domain (Chisari and Ferrari, 1995) and so may be subjected to selective pressure by cellular immune responses. Mutations at the C terminal end of the S protein between residues 184 and 216 that add in mainly HCC patients occur in a region that has been postulated to be part of a discontinuous B cell epitope (amino acid 187–207) (Chen et al., 1996), so mutations in this region may also be driven by immune pressure. This study reports an alarming number of amino acid changes (W191R, L192I, V194A, M197K, M198V, M198A, W199R, Y200N, G202R, S207N) among the Pakistani isolates and this shows the level of immune pressure observed among the genotype D strains in South Asia. This area of immune evasion has not been tested in-silico as this was beyond the scope of this study. However, the information is vital to identify other immune system evasion strategies operative in the native strain of HBV.
A large number of hepatitis B virus sequences (figure 10, 11, 12 and 13) show variations within B-cell epitopes [HBsAg 44 to 49: five sequences (38.4%), 115 to 155: ten sequences (77%) , 124 to 147: seven sequences (54%) and 160 to 207: seven sequences (54%)]. Five HBV sequences (38.4%) exhibited amino acid exchanges in class II T helper epitopes (positions 136 to 155), while four sequences (31%) showed the consecutive mutation in amino acid of another class II T-cell epitope (positions 213 to 226). This is indeed very alarming and the healthcare personnel’s are very much blinded regarding the mutated strains of HBV.
Qiu (2011) had reported two conserved residues, M75 and M103 with respect to HBsAg, mutations of which not only attenuated HBsAg secretion (M75 only), but also suppressed HBV genome replication without compromising the overlapping p-gene product (Qiu et al., 2011). In the present study we found conserved positions for the respective amino acids among the Pakistani isolates.

Novus International Journal of Biotechnology & Bioscience�������������������������������������������������������������
Toressi has described I195M, W196S, and the M198I mutations in surface ORF among lamivudine resistant individuals (Torresi et al., 2002). These mutations are found to show reduced binding to anti HBs antibody. Analysis of the pakistani isolates identified two isolates of M198V/M198I. This is a further confirmation of the extensive use of Lamivudine drug in treating HBV patients of Pakistan. The percentage of isolates may even be higher as this study didn’t target the specific non-responder group of lamivudine. The important point here is that if the random population has such lamivudine mutants of HBV, the idea of prescribing this drug for the pakistani population is quite alarming.
Surface gene mutants have also gained more importance recently among pegylated interferon users (Hsu and Yeh, 2011). The amino acid changes reported by Hsu and Yeh among non-responders of HBIG include psL30S, psE54D, psA55T, psG145S, sL97P, sT125A, and sN207H. Out of those 13 isolates of surface gene Q30K, Q30P, S55P, L97P and S207N were seen. It is postulated that such mutants may be the reason for the interferon non-responder pool in Pakistani HBV patients This is the first report of interferon immune patients in the native population. Further studies are needed to test these mutants among interferon non-responders to get more information in this regard.
Acknowledgement
We would like to thank members of the HBV lab at NCVI for their encouragement in preparing this manuscript. The work was supported by a H̀igher Education Commissiion’ and US Department of State joint funding in a Pak-US project entitled “HBV induced hepatocellular carcinoma developement between the NUST center of virology and Immunoogy & University of California, San Diago
The authors declare no conflict of interest
REFERENCES:
• Alam, M. M., Zaidi, S. Z., Malik, S. A., Naeem, A., Shaukat, S., Sharif, S., Angez, M., Khan, A., and Butt, J. A. (2007). Serology based disease status of Pakistani population infected with hepatitis B virus. BMC Infect Dis.7: 64.
• Chen, Y. C., Delbrook, K., Dealwis, C., Mimms, L., Mushahwar, I. K., and Mandecki, W. (1996).
• Discontinuous epitopes of hepatitis B surface antigen derived from a filamentous phage peptide library. Proc Natl Acad Sci U S A.93(5): 1997-2001.
• Chisari, F. V., and Ferrari, C. (1995). Hepatitis B virus immunopathogenesis. Annu Rev Immunol.13: 29-60.
• Hsu, C. W., and Yeh, C. T. (2011). Emergence of hepatitis B virus S gene mutants in patients experiencing hepatitis B surface antigen seroconversion after peginterferon therapy. Hepatology.54(1): 101-108.
• Khokhar, N., Gill, M. L., and Alam, A. Y. (2005). Treatment of chronic hepatitis B with lamivudine. J Coll Physicians Surg Pak.15(2): 78-80.

Novus International Journal of Biotechnology & Bioscience��������������������������������������������������������������
• Liu, C. J., Kao, J. H., Shau, W. Y., Chen, P. J., Lai, M. Y., and Chen, D. S. (2002). Naturally occurring hepatitis B surface gene variants in chronic hepatitis B virus infection: correlation with viral serotypes and clinical stages of liver disease. J Med Virol.68(1): 50-59.
• Lu, H. Y., Zeng, Z., Xu, X. Y., Zhang, N. L., Yu, M., and Gong, W. B. (2006). Mutations in surface and polymerase gene of chronic hepatitis B patients with coexisting HBsAg and anti-HBs. World J Gastroenterol.12(26): 4219-4223.
• Mujeeb, S. A., and Pearce, M. S. (2008). Temporal trends in hepatitis B and C infection in family blood donors from interior Sindh, Pakistan. BMC Infect Dis.8: 43.
• Noorali, S., Hakim, S. T., McLean, D., Kazmi, S. U., and Bagasra, O. (2008). Prevalence of Hepatitis B virus genotype D in females in Karachi, Pakistan. J Infect Dev Ctries.2(5): 373-378.
• Ogura, Y., Kurosaki, M., Asahina, Y., Enomoto, N., Marumo, F., and Sato, C. (1999). Prevalence and significance of naturally occurring mutations in the surface and polymerase genes of hepatitis B virus. J Infect Dis.180(5): 1444-1451.
• Qiu, J., Qin, B., Rayner, S., Wu, C. C., Pei, R. J., Xu, S., Wang, Y., and Chen, X. W. (2011). Novel evidence suggests Hepatitis B virus surface proteins participate in regulation of HBV genome replication. Virol Sin.26(2): 131-138.
• Rustgi, V., Carriero, D., Bachtold, M and Zeldin, G. (2010). Update on Chronic Hepatitis B. The Journal for Nurse Practitioners, 6(8): 631-639
• Sayan, M., and Akhan, S. C. (2011). Antiviral drug-associated potential vaccine-escape hepatitis B virus mutants in Turkish patients with chronic hepatitis B. Int J Infect Dis.
• Stirk, H. J., Thornton, J. M., and Howard, C. R. (1992). A topological model for hepatitis B surface antigen. Intervirology.33(3): 148-158.
• Tong, W., Sun, L., He, J., He, S., and Du, F. (2010). A novel nucleotide insertion in S gene of hepatitis B virus in a chronic carrier. Virol J.7: 104.
• Torresi, J., Earnest-Silveira, L., Deliyannis, G., Edgtton, K., Zhuang, H., Locarnini, S. A., Fyfe, J., Sozzi, T., and Jackson, D. C. (2002). Reduced antigenicity of the hepatitis B virus HBsAg protein arising as a consequence of sequence changes in the overlapping polymerase gene that are selected by lamivudine therapy. Virology.293(2): 305-313.
• Villet, S., Pichoud, C., Billioud, G., Barraud, L., Durantel, S., Trepo, C., and Zoulim, F. (2008). Impact of hepatitis B virus rtA181V/T mutants on hepatitis B treatment failure. J Hepatol.48(5): 747-755.
• Xia, G., Nainan, O. V., Jia, Z., Wang, J. J., Liu, H. B., Li, R. C., Cao, H. L., Liu, C. B., and Margolis, H. S. (2001). [Prevalence of mutants in the determinant region of hepatitis B surface antigen among Chinese carriers after receiving only active postexposure immunoprophylaxis]. Zhonghua Shi Yan He Lin Chuang Bing Du Xue Za Zhi.15(3): 212-215.




















