
Loss of D 3 receptors in the zitter mutant rat is not reversed by l-dopa treatment
12
Loss of D 3 receptors in the zitter mutant rat is not reversed by L-dopa treatment Jeffrey N. Joyce, a, * T.C. Der, a,b Lynn Renish, a Tracy Osredkar, a Diane Hagner, a Maria Reploge, a Shinichi Sakakibara, c and Shuichi Ueda c a Thomas H. Christopher Center for Parkinson’s Disease Research, Sun Health Research Institute, Sun City, AZ 85351, USA b Department of Anatomy and Neurosciences, Arizona College of Osteopathic Medicine, Midwestern University, Glendale, AZ 85308, USA c Department of Histology and Neurobiology, Dokkyo University School of Medicine, Mibu, Tochigi 321-0293, Japan Received 11 September 2003; revised 19 November 2003; accepted 13 January 2004 Abstract In Parkinson’s disease (PD) and animal models of parkinsonism the destruction of nigrostriatal (NSB) system results in a marked loss of the dopamine D 3 receptor and mRNA in the islands of Calleja (ICj) and the nucleus accumbens shell (NAS). In animal models, it has been reported that both measures are elevated by repeated intermittent administration of L-dopa. However, a large proportion of PD cases are resistant to L-dopa-induced elevation of D 3 receptor number. The zitter mutant (Zi/Zi) rat replicates the slow progressive degeneration of the NSB observed in PD and also exhibits a loss of D 3 receptor number in the NAS or ICj. To test if this could be reversed with subchronic L-dopa treatment, injections of carbidopa (10 mg/kg ip) were followed an hour later with injection of L-dopa (100 mg/kg ip) twice a day for 10 days. In control Sprague – Dawley (SD) and zitter heterozygote (Zi/) rats that do not show a loss of D 3 receptors with vehicle treatment, L-dopa produced no change in D 3 receptor number or in DA terminal density as measured by dopamine transporter (DAT) binding and tyrosine hydroxylase immunoautoradiography (TH-IR). There was a marked loss of DAT and TH-IR in caudate –putamen (CPu) and NA, as well as D 3 receptors in NAS and ICj in Zi/Zi rats but no further change with L-dopa treatment. To determine if the resistance to L-dopa- induced increase in D 3 receptor was due to a deficiency in expression of cortical BDNF or its receptor, TrkB, in CPu and NAS, we examined BDNF mRNA by ISHH in frontal cortex and TrkB mRNA in frontal cortex, CPu, and NA. The loss of the NSB in the Zi/Zi did not alter levels of BDNF or TrkB mRNA, nor did L-dopa administration alter levels BDNF or TrkB mRNA. Thus, unlike in 6-hydroxydopamine- treated rats, in Zi/Zi rats administered L-dopa does not reverse the loss of BDNF mRNA or lead to an elevation of D 3 receptor number. D 2004 Elsevier Inc. All rights reserved. Keywords: Parkinson’s disease; Striatum; Nucleus accumbens; Tyrosine hydroxylase; Substantia nigra; Ventral tegmental area Introduction Parkinson’s disease (PD) is a neurodegenerative disorder with an insidious onset and a prolonged course over many years. Symptoms are secondary to the death of nigrostriatal (NSB) DA neurons and the resultant depletion of DA in the striatum (Hornykiewicz, 1998). Symptomatic treatment includes levodopa (L-dopa, the most effective antiparkinso- nian drug), which is converted to DA, and direct-acting DA agonists (bromocriptine, pergolide, pramipexole, ropinirole) (Brooks, 2000). However, many patients lose their response to these antiparkinsonian drugs (APDs) and they are inef- fective for cooccurring dementia and depression (Fabbrini et al., 1988; Mouradian et al., 1988). Dementia is in fact highly correlated with the severity of parkinsonism, older age of onset, and declining response to APDs (Apaydin et al., 2002; Caparros-Lefebvre et al., 1995; Hughes et al., 2000; Hurtig et al., 2000; Marder et al., 1995; Portin and Rinne, 1987; Stern et al., 1993). Loss of response, and its correla- tion with dementia, is differentiated from a complicated response to APDs or dyskinesias, which can be managed by titrating the dosage of medication. Five distinct subtypes of G-protein coupled DA receptors mediate the actions of DA, three of which, D 2 ,D 3 , and D 4 , belong to the D 2 subfamily (Civelli et al., 1993). DA regulates output pathways of the striatum via interaction with different DA receptors that are, in turn, differentially 0014-4886/$ - see front matter D 2004 Elsevier Inc. All rights reserved. doi:10.1016/j.expneurol.2004.01.012 * Corresponding author. Thomas H. Christopher Center for Parkinson’s Disease Research, Sun Health Research Institute, 10515 West Santa Fe Drive, Sun City, AZ 85351. Fax: +1-623-876-5695. E-mail address: [email protected] (J.N. Joyce). www.elsevier.com/locate/yexnr Experimental Neurology 187 (2004) 178 – 189
Transcript of Loss of D 3 receptors in the zitter mutant rat is not reversed by l-dopa treatment

www.elsevier.com/locate/yexnr
Experimental Neurology 187 (2004) 178–189
Loss of D3 receptors in the zitter mutant rat is not reversed
by L-dopa treatment
Jeffrey N. Joyce,a,* T.C. Der,a,b Lynn Renish,a Tracy Osredkar,a Diane Hagner,a Maria Reploge,a
Shinichi Sakakibara,c and Shuichi Uedac
aThomas H. Christopher Center for Parkinson’s Disease Research, Sun Health Research Institute, Sun City, AZ 85351, USAbDepartment of Anatomy and Neurosciences, Arizona College of Osteopathic Medicine, Midwestern University, Glendale, AZ 85308, USA
cDepartment of Histology and Neurobiology, Dokkyo University School of Medicine, Mibu, Tochigi 321-0293, Japan
Received 11 September 2003; revised 19 November 2003; accepted 13 January 2004
Abstract
In Parkinson’s disease (PD) and animal models of parkinsonism the destruction of nigrostriatal (NSB) system results in a marked loss of
the dopamine D3 receptor and mRNA in the islands of Calleja (ICj) and the nucleus accumbens shell (NAS). In animal models, it has been
reported that both measures are elevated by repeated intermittent administration of L-dopa. However, a large proportion of PD cases are
resistant to L-dopa-induced elevation of D3 receptor number. The zitter mutant (Zi/Zi) rat replicates the slow progressive degeneration of the
NSB observed in PD and also exhibits a loss of D3 receptor number in the NAS or ICj. To test if this could be reversed with subchronic
L-dopa treatment, injections of carbidopa (10 mg/kg ip) were followed an hour later with injection of L-dopa (100 mg/kg ip) twice a day for
10 days. In control Sprague–Dawley (SD) and zitter heterozygote (Zi/�) rats that do not show a loss of D3 receptors with vehicle treatment,
L-dopa produced no change in D3 receptor number or in DA terminal density as measured by dopamine transporter (DAT) binding and
tyrosine hydroxylase immunoautoradiography (TH-IR). There was a marked loss of DAT and TH-IR in caudate–putamen (CPu) and NA, as
well as D3 receptors in NAS and ICj in Zi/Zi rats but no further change with L-dopa treatment. To determine if the resistance to L-dopa-
induced increase in D3 receptor was due to a deficiency in expression of cortical BDNF or its receptor, TrkB, in CPu and NAS, we examined
BDNF mRNA by ISHH in frontal cortex and TrkB mRNA in frontal cortex, CPu, and NA. The loss of the NSB in the Zi/Zi did not alter
levels of BDNF or TrkB mRNA, nor did L-dopa administration alter levels BDNF or TrkB mRNA. Thus, unlike in 6-hydroxydopamine-
treated rats, in Zi/Zi rats administered L-dopa does not reverse the loss of BDNF mRNA or lead to an elevation of D3 receptor number.
D 2004 Elsevier Inc. All rights reserved.
Keywords: Parkinson’s disease; Striatum; Nucleus accumbens; Tyrosine hydroxylase; Substantia nigra; Ventral tegmental area
Introduction to these antiparkinsonian drugs (APDs) and they are inef-
Parkinson’s disease (PD) is a neurodegenerative disorder
with an insidious onset and a prolonged course over many
years. Symptoms are secondary to the death of nigrostriatal
(NSB) DA neurons and the resultant depletion of DA in the
striatum (Hornykiewicz, 1998). Symptomatic treatment
includes levodopa (L-dopa, the most effective antiparkinso-
nian drug), which is converted to DA, and direct-acting DA
agonists (bromocriptine, pergolide, pramipexole, ropinirole)
(Brooks, 2000). However, many patients lose their response
0014-4886/$ - see front matter D 2004 Elsevier Inc. All rights reserved.
doi:10.1016/j.expneurol.2004.01.012
* Corresponding author. Thomas H. Christopher Center for Parkinson’s
Disease Research, Sun Health Research Institute, 10515 West Santa Fe
Drive, Sun City, AZ 85351. Fax: +1-623-876-5695.
E-mail address: [email protected] (J.N. Joyce).
fective for cooccurring dementia and depression (Fabbrini et
al., 1988; Mouradian et al., 1988). Dementia is in fact highly
correlated with the severity of parkinsonism, older age of
onset, and declining response to APDs (Apaydin et al.,
2002; Caparros-Lefebvre et al., 1995; Hughes et al., 2000;
Hurtig et al., 2000; Marder et al., 1995; Portin and Rinne,
1987; Stern et al., 1993). Loss of response, and its correla-
tion with dementia, is differentiated from a complicated
response to APDs or dyskinesias, which can be managed by
titrating the dosage of medication.
Five distinct subtypes of G-protein coupled DA receptors
mediate the actions of DA, three of which, D2, D3, and D4,
belong to the D2 subfamily (Civelli et al., 1993). DA
regulates output pathways of the striatum via interaction
with different DA receptors that are, in turn, differentially

J.N. Joyce et al. / Experimental Neurology 187 (2004) 178–189 179
expressed in these output pathways (Joyce, 2001b). How-
ever, to date, all effective DA agonists stimulate D2-like
receptors, and changes in expression of the D2 receptor have
been hypothesized to underlie alterations in the response to
APDs in PD (Pizzolato et al., 1995). In a recent review
(Joyce, 2001b), we discussed the clinical and preclinical
literature that supports the hypothesis that the initial expres-
sion of parkinsonian symptoms is not only correlated with
the loss of DA innervation and an elevation in receptor
number of the D2 subclass, but also a decrease D3 receptor
number. In animal models of PD, it has been demonstrated
that the reduction of D3 receptor number after the loss of
DA innervation can be reversed by L-dopa in DA denervated
animals (Bordet et al., 1997; Morissette et al., 1998; Quik et
al., 2000). However, we have found in PD that the mainte-
nance of response to APDs (and the absence of PD
dementia) is associated with maintenance or an increase in
D3 receptor number, and conversely that loss of response to
APDs (and presence of PD dementia) is associated with a
decrease in DA D3 receptor number (Joyce et al., 2002;
Ryoo et al., 1998). Interestingly, measurement of DA
terminals with tyrosine hydroxylase (TH) immunoautora-
diography and dopamine transporter (DAT) binding in
striatum, as well as unbiased stereological counts of TH-
IR neurons in the midbrain, showed that damage to the
nigrostriatal and mesolimbic DA system was not different in
PD cases who had loss of response to APDs (and dementia)
compared to PD cases who maintained response to APDs
(and who were free from clinical symptoms of dementia)
(Joyce et al., 2002).
In view of the above findings, it becomes important to
ask why L-dopa does not appear capable of normalizing
deficient D3 receptor expression in PD cases that are
unresponsive to APDs. Unlike that for the D2 receptor,
depletion of DA fibers by 6-hydroxydopamine (6-OHDA)
or 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)
leads to a reduction of D3 receptor number (Joyce, 1991;
Joyce et al., 1986; Levesque et al., 1995; Morissette et al.,
1998; Wade et al., 2001). The relationship between regional
loss of DA fibers and loss of postsynaptically located D3
receptors has not been clarified, but it is clear that regulation
of D3 receptors is not directed by levels of DA per se (Joyce,
2001a; Joyce et al., 2000; Levesque et al., 1995). Guillin et
al. (2001) have proposed that the D3 receptor is regulated by
brain-derived neurotrophic factor (BDNF). The primary
source of BDNF in striatum is from DA fibers (Altar et
al., 1997) and its reduction following lesions to the nigros-
triatal DA system is thought to correlate with reduced D3
receptor. In fact, it has been proposed that the elevation of
D3 receptor number in L-dopa-treated DA-denervated ani-
mals is regulated by an increased production of BDNF in
corticostriatal fibers (Guillin et al., 2001).
Based on those studies in animal models of PD, it is not
clear why L-dopa administration loses its ability to normal-
ize or elevate D3 receptor number in a subset of PD cases
(Joyce et al., 2002). One possibility is that animals used to
explore regulation of D3 receptor number and response to L-
dopa utilize rapid and massive depletions of DA neurons or
DA fibers. We have explored the regulation of the D3
receptor in a more natural animal model of PD that exhibits
a progressive degeneration of the DAergic and serotonergic
system over a 12-month period, the zitter (Zi/Zi) mutant rat
(Joyce et al., 2000). The Zi/Zi rat is an autosomal recessive
mutant derived from the Sprague–Dawley (SD) rat strain
(Ueda et al., 2002). The Zi/Zi rat exhibits several neurolog-
ical abnormalities that are age progressive. Abnormal me-
tabolism of H202 and catalase leads to an increase in oxygen
species in the brain that is correlated with a significant
reduction in DA and dihydroxyphenylacetic acid (Gomi et
al., 1994; Ueda et al., 2002). In the Zi/Zi rat, there is a
characteristic pattern of DA fiber and neuron degeneration
that is observed initially in the nigrostriatal DA system
followed by a slower degeneration to the mesolimbic DA
system (Ueda et al., 2000). However, even by 3 months of
age when DA levels and fiber density are normal in the
nucleus accumbens (NA) of the Zi/Zi rat, there is a pro-
nounced loss (>70%) of D3 receptor. In the present paper,
we show that the loss of D3 receptors in the Zi/Zi rat is not
normalized or elevated by subchronic L-dopa treatment.
Furthermore, subchronic L-dopa treatment does not produce
an increase in cortical BDNF mRNA, nor are there changes
in TrkB mRNA in striatum of Zi/Zi rats.
Materials and methods
Subjects
Zitter mutant (Zi/Zi), zitter heterozygote (Zi/+) rats were
raised and maintained in the Laboratory Animal Research
Center, Dokkyo University School of Medicine. For Exper-
iment 1, the Sprague–Dawley (SD) controls were also
raised and maintained at the same institution. For Experi-
ment 2, the SD controls were purchased from Charles River
(6 and 10 months old retired breeders) and maintained at
Sun Health Research Institute. All animals were housed in
groups of two or three with continuous access to food and
water and maintained on a 12:12-h light–dark cycle.
Drug treatment
Experiment 1
At 6 or 10 months age, four Zi/Zi and four Zi/+ zitter
male rats were injected with carbidopa (10 mg/kg ip; Sigma,
St Louis MO), a peripheral decarboxylase inhibitor. An hour
later, the animals were given an injection of L-dopa (100
mg/kg ip; Sigma). This procedure was followed twice a day
for 10 successive days. An additional four Zi/Zi, four Zi/+
zitter, and four SD male rats were treated with the vehicle
saline (1 ml/kg ip) twice daily for 10 successive days. At the
end of the 10th day, the rats were overdosed with pento-
barbital and the brain immediately frozen in isopentane

J.N. Joyce et al. / Experimental Neurology 187 (2004) 178–189180
(�30jC) and stored at �80jC and maintained at �45jCduring shipping to Sun Health Research Institute (Sun
City, AZ).
Experiment 2
In Experiment 1, we observed nonsignificant changes in
some analyses that indicated the need for further substan-
tiation of the data, additional analyses, and additional
protocols. An additional study was undertaken with four
female and two male Zi/Zi rats at 6 or 10 months age and
four female and four male Zi/+ zitter male rats. They were
treated with subchronic L-dopa/carbidopa (100:10 mg/kg
ip) twice a day for 10 days in exactly the same manner as
in Experiment 1. An additional four female Zi/Zi and four
female Zi/+ zitter rats were treated with the vehicle saline
(1 ml/kg) two times daily for 10 successive days. To
obtain SD controls, at Sun Health Research Institute, six
SD male rats at 6 months and six at 10 months age were
treated with subchronic L-dopa/carbidopa, (100:10 mg/kg
ip) twice a day for 10 days in exactly the same manner as
in Experiment 1. An additional five SD rats at 6 months
and 6 at 10 months age were treated with the vehicle
saline (1 ml/kg) two times daily for 10 successive days. At
the end of the 10th day, the rats were overdosed, the brains
removed and immediately frozen in isopentane (�30jC)and stored at �80jC. The Animal Welfare Committee of
Dokkyo University School of Medicine and the IACUC
committee of Sun Health Research Institute approved all
procedures.
Autoradiographic procedures
The brains were sectioned at 20 Am thickness in a Leica
CM 1900 cryotome at �20jC, thaw-mounted onto Probe
On Plus slides (Fisher, Pittsburgh, PA), dried on warm plate,
and stored at �70jC until further experiments. Sections
from the striatal complex were processed for D3 receptor
autoradiography, DAT autoradiography, and TH-immunoau-
toradiography per our standard procedures (Joyce et al.,
2002; Murray et al., 1995; Ryoo et al., 1998). Alternate
sections were used for radioligand binding and in situ
hybridization (ISHH) experiments. Sections from the mid-
brain were processed for TH-immunoautoradiography.
[125I](R)-trans-7-hydroxy-2-[N-propyl-N-(3V-iodo-2V-pro-penyl)-amino]tetralin ([125I]trans-7-OH-PIPAT) (NEN, Bos-
ton, MA) binding to D3 receptors was used with conditions
that allow selective labeling of D3 receptor as described
earlier (Gurevich and Joyce, 1999, Gurevich et al., 1999).
The sections were preincubated for 30 min at 30jC in buffer
containing 50 mmol/l Tris–HCl (pH 7.4), 100 mmol/l NaCl.
To each section, 500 Al of incubation buffer containing 50
mmol/l Tris–HCl (pH 7.4), 0.7 nmol/l [125I]trans 7-OH-
PIPAT (Dupont, New England Nuclear, Boston, MA), 40
mmol/l NaCl, and 100 Amol/l GTP was applied for 90 min
at room temperature. Nonspecific binding was defined as
the binding in the presence of 10 Amol/l 7-OH-DPAT. The
sections were then washed three times in the same buffer
without GTP and [125I]trans 7-OH-PIPAT at 4jC for 1 h,
briefly dipped in ice-cold deionized water and dried.
Dopamine transporter sites were labeled with [125I]RTI-
55 (3h-(4-iodophenyl)tropan-2 h-carboxylic acid methyl
ester) (Dupont) (Joyce et al., 2000; Ryoo et al., 1998).
The slide-mounted sections were incubated with 100 pmol/
l [125I] RTI-55 in 50 mmol/l Tris–HCL buffer (pH 7.4)
containing 150 mmol/l NaCl and 5 mmol/l KCl for 2 h at
room temperature. 100 nmol/l of paroxetine was added to
block 5-HT transporter sites. Nonspecific binding was
determined with 4 Amol/l benztropine. Sections were then
washed in the same Tris buffer twice for 20 min at 0jC and
rinsed in cold ddH20 for 30 s before dried on slide warmer.
Sections along with standards were apposed to 3H-Hyper-
film for 18 h.
For tyrosine hydroxylase (TH) immunoautoradiography
(Joyce et al., 2002; Ryoo et al., 1998), sections were cut and
immediately fixed in 4% paraformaldehyde in phosphate-
buffered saline (PBS) for 1 h. Tissues were rinsed three
times in PBS, dehydrated through 70%, 80%, 95%, and
100% ethanol, and stored at �70jC until further use.
Sections were first incubated in the primary antibody, mouse
anti-TH (Chemicon, Temecula, CA) at 1:500 dilution for 24
h in PBS with 0.1% bovine serum albumin (BSA) and 0.3%
Triton X-100. The following day, sections were rinsed three
times in 0.1% BSA-PBS solution and then incubated with
the secondary antibody, [125I] F(ab)2 sheep anti-mouse
immunoglobulin G (Amersham, Arlington Heights, Ill) at
1:200 dilution in 0.1% BSA-PBS for 1 h. Sections were then
washed three times in PBS, three times in deionized water,
and dried on warm plate. Sections along with standards
were apposed to 3H-Hyperfilm for 24 h.
In situ hybridization histochemistry
Adjacent sections were processed for TrkB and BDNF
ISHH per our routine procedures (Gurevich et al., 1999).
Antisense BDNF mRNAwas generated from the rat recom-
binant plasmid pR1112-8 with T3 RNA polymerase after
digestion with BamHI. This cRNA probe contains 384 bases
complementary to the mRNA encoding mature rat BDNF
protein (Conner et al., 1997; Isackson et al., 1991). The
cRNA probe for TrkB contains 483 bases complementary to
extracellular domain of the mouse TrkB protein (Klein et al.,
1990). Digestion was completed with HindIII and transcrip-
tion with T7. Sense probes for both BDNF and TrkB were
also produced. The probes were labeled in standard in vitro
transcription reaction with 33P-UTP (Perkin Elmer, Boston,
MA). Before hybridization, sections were treated with
proteinase K (10 min at 37jC, 1 Ag/ml), rinsed briefly in
water, and incubated in 0.1 M triethanolamine buffer con-
taining 0.25% acetic anhydride for 15 min at room temper-
ature. After brief wash in 2 � SSC, sections were
dehydrated in ethanols, dried, and approximately 1–2 �106 cpm of the probe in 25 Al of hybridization buffer

J.N. Joyce et al. / Experimental Neurology 187 (2004) 178–189 181
containing 75% formamide, 50 mM Tris–HCl, 2.5 mM
EDTA, 3 � SSC, 10% dextran sulfate, 25 mM DTT, 1 �Denhardt’s solution, 0.5 mg/ml tRNA, and 0.2 mg/ml
Salmon testes DNAwere applied per section. Sections were
coverslipped and placed in hybridization oven at 55jC for
20 h. Upon completion of hybridization, sections were
subjected to RNAse digestion and washes of increased
stringency culminating in 1 h wash in 0.1 � SSC at
60jC. Following dehydration in ethanols, sections were
exposed to Kodak Biomax MR film (Amersham) along
with standards for 10 days. Film was developed with Kodak
GBX developer and fixer. The sections were developed in
Kodak D-19 developer, fixed in 30% sodium thiosulfate,
washed extensively in water, dehydrated, counterstained
with Nuclear Fast Red, and mounted from xylene in
Permount. Four sections for antisense probe and two sec-
tions for sense probe hybridization were used for each
animal. Labeling of the sense probe used as control for
specificity of hybridization signal was negligible as was the
background labeling of the antisense probe (in such areas as
white matter). In addition, we utilized pre-proenkephalin
cRNA probe as a control for stringency of ISHH protocols.
Data analysis
For Experiment 1, the autoradiographs were analyzed
using a Macintosh computer-based image analysis system
(BRAIN version 2.0, Drexel University), which converts
gray values to the amount of radioligand bound in fmol/mg
of protein (Artymyshyn et al., 1990). For Experiment 2,
autoradiographs were analyzed using a PC computer-based
image analysis system (AIS, Imaging Research Inc., Ontario
Canada) that converts transmitted optical density to the
amount of radioligand bound in fmol/mg of protein. The
optical density values derived from the autoradiographs of
[125I]radioligand binding were converted to fmol/mg protein
using standards calibrated for 125I. For ISHH autoradiogra-
phy, optical density values were converted to dpm/cm2
using 14C standards calibrated for 33P and then converted
into femtomole of probe per gram of wet weight using
specific activity of the probe in each experiment. For
analysis of the autoradiographs, four sections for total and
Table 1
Effects of strain and drug treatment on the dopamine system
Dopamine transporter
CPu NAc NAs Olf Tub
SD with saline 45.7 F 4.7 28.9 F 3.1 25.8 F 5.8 27.9 F 3
Zi/+ with saline 45.3 F 5.3# 23.7 F 5.7 19.8 F 5.5 25.9 F 3
Zi/Zi with saline 33.1 F 3.1* 29.7 F 4.4 15.8 F 4.1* 18.6 F 4
Zi/+ with L-dopa 46.1 F 5.4y 24.7 F 4.0 13.8 F 4.4* 22.2 F 3
Zi/Zi with L-dopa 25.5 F 2.0* 21.5 F 3.7 10.4 F 3.9* 13.6 F 9
*Significant at P < 0.05 vs. SD with saline.# Significant at P < 0.05, Zi/+ with saline vs. Zi/Zi with saline.y Significant at P < 0.05, Zi/+ with L-dopa vs. Zi/Zi L-dopa.£ Significant at P < 0.05, Zi/+ with saline vs. Zi/+ with L-dopa.
two sections for nonspecific binding from each animal were
utilized. Optical densities in defined brain areas were
measured bilaterally, allowing for eight measurements for
total and four measurements for nonspecific binding. For
each animal, the total and nonspecific binding were aver-
aged and nonspecific binding was subtracted from total
binding to yield values for specific binding for each region
of each animal. For the present study, the striatum was
divided into subdivisions: caudate–putamen (CPu), nucleus
accumbens shell (NAs), nucleus accumbens core (NAc),
islands of Calleja (ICj), and olfactory tubercle (Olf Tub).
For ISHH autoradiography, optical density values were
converted to ACi/g using 14C standards (American Radio-
labeled Chemicals, Inc., St. Louis MO) utilizing the cali-
bration scale provided with each standard (Gurevich et al.,
1999). For analysis of the autoradiographs of the sections
processed for ISHH, four sections for the antisense probe
and two sections for the sense probe from each animal were
utilized. Optical densities in defined brain areas were
measured bilaterally, allowing for eight measurements for
the antisense probe and four measurements for the sense
probe. For each animal, the readings from the sense probe-
labeled sections were subtracted from antisense probe-la-
beled sections to yield values for specific labeling for each
region of each animal. For the ISHH analysis, readings were
taken from the cingulate cortex (CG), frontal cortex (FC),
and nucleus accumbens (NA) (Paxinos and Watson,1986).
For statistical analysis, a two-way ANOVA, with group and
region as main factors, and pairwise comparisons performed
using post hoc Dunnett with Bonferroni correction. For
Experiment 2, data from the zitter groups were analyzed
separately from the SD groups because the autoradiographic
procedures were completed independently.
Results
Experiment 1
The results of Experiment 1 (shown in Table 1) confirm
our previous results that the zitter homozygotic rats (Zi/Zi)
have significantly lower concentrations of DAT labeling of
D3 receptor binding
CPu NAc NAs ICj
.3 3.0 F 1.9 9.17 F 1.8 13.31 F 3.2 22.96 F 4.1
.5 5.6 F 3.3 13.37 F 2.9# 18.97 F 2.9*,# 23.75 F 3.6#
.1* 2.3 F 2.5 2.85 F 1.5* 3.25 F 2.5* 9.63 F 2.2*
.0y 4.0 F 1.5 8.21 F 1.6 8.83 F 2.1£ 17.69 F 5.1
.1* 1.1 F 3.6 2.04 F 2.1*,y 2.66 F 2.7*,y 13.14 F 5.0*

J.N. Joyce et al. / Experimental Neurology 187 (2004) 178–189182
DA fibers, a marker for the integrity of the DA fibers (Joyce
et al., 2002; Murray et al., 1995; Ryoo et al., 1998), as
compared to SD control rats (Joyce et al., 2000), and
intermittent L-dopa treatment nonsignificantly exacerbated
that effect. Analysis of the effects of group (SD, Zi/+ with
saline, Zi/Zi with saline, Zi/+ with L-dopa, and Zi/Zi with L-
dopa) and regions demonstrated group (P < 0.001, F =
21.42), region (P < 0.001, F = 83.68), and interaction
effects (P = 0.0012, F = 3.68) for DAT binding. The
concentrations of DAT binding sites were not different in
zitter heterozygotes (Zi/+) treated with saline or L-dopa as
compared to the SD controls (Table 1). Zi/Zi rats did have
significantly lower concentrations of DAT in the CPu
(�28%), NAs (�39%), and Olf Tub (�34%) as compared
to SD rats. Treatment of Zi/Zi rats with L-dopa nonsignif-
icantly enhanced the loss of DAT sites in the CPu (�45% of
Fig. 1. Low-power photomicrograph of the original autoradiograph of the cauda
binding from individual subjects of four groups: (A) Zi/Zi with saline, (B) Zi/Zi wit
saline, and (D) Zi/+ with L-dopa/carbidopa. Note the marked depletion of DAT site
The ROI used for analysis is depicted by dashed lines in panel D. Abbreviation
accumbens core; Olf Tub, olfactory tubercle.
SD), NAs (�60% of SD), and Olf Tub (�45% of SD) as
compared to the Zi/Zi treated with saline.
As reported previously (Joyce et al., 2000), Zi/Zi rats had
significantly lower concentrations of D3 receptors in striatal
regions as compared with SD rats, but intermittent L-dopa
treatment did not further alter the concentrations of D3
receptors in the Zi/Zi rats. Analysis of the effects of group
(SD, Zi/+ with saline, Zi/Zi with saline, Zi/+ with L-dopa, Zi/
Zi with L-dopa) and regions demonstrated group (P < 0.001,
F = 39.91), region (P < 0.001, F = 80.50), and interaction
effects (P = 0.0019, F = 3.094) for concentrations of D3
receptors. There were no differences in the concentration of
D3 receptors in the CPu, NAc, or ICj between SD and Zi/+
rats treated with saline or L-dopa, although the Zi/+ rats
treated with saline did have a somewhat greater concentra-
tion of D3 receptors in the NAs as compared to the SD rats.
te–putamen and nucleus accumbens of tissue sections processed for DAT
h L-dopa/carbidopa (100:10 mg/kg ip) twice a day for 10 days, (C) Zi/+ with
s from the caudate–putamen in Zi/Zi and lack of affect of L-dopa treatment.
s: CPu, caudate–putamen; NAs, nucleus accumbens shell; NAc, nucleus
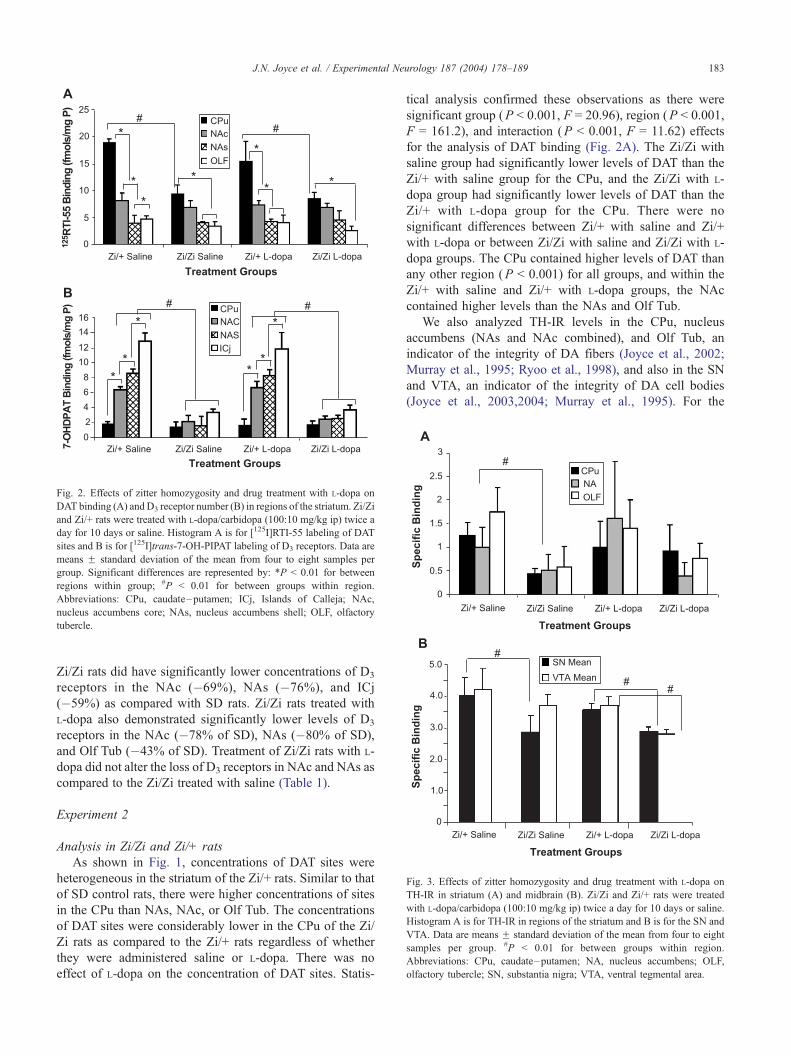
Fig. 2. Effects of zitter homozygosity and drug treatment with L-dopa on
DAT binding (A) andD3 receptor number (B) in regions of the striatum. Zi/Zi
and Zi/+ rats were treated with L-dopa/carbidopa (100:10 mg/kg ip) twice a
day for 10 days or saline. Histogram A is for [125I]RTI-55 labeling of DAT
sites and B is for [125I]trans-7-OH-PIPAT labeling of D3 receptors. Data are
means F standard deviation of the mean from four to eight samples per
group. Significant differences are represented by: *P < 0.01 for between
regions within group; #P < 0.01 for between groups within region.
Abbreviations: CPu, caudate–putamen; ICj, Islands of Calleja; NAc,
nucleus accumbens core; NAs, nucleus accumbens shell; OLF, olfactory
tubercle.
Fig. 3. Effects of zitter homozygosity and drug treatment with L-dopa on
TH-IR in striatum (A) and midbrain (B). Zi/Zi and Zi/+ rats were treated
with L-dopa/carbidopa (100:10 mg/kg ip) twice a day for 10 days or saline.
Histogram A is for TH-IR in regions of the striatum and B is for the SN and
VTA. Data are means F standard deviation of the mean from four to eight
samples per group. #P < 0.01 for between groups within region.
Abbreviations: CPu, caudate–putamen; NA, nucleus accumbens; OLF,
olfactory tubercle; SN, substantia nigra; VTA, ventral tegmental area.
J.N. Joyce et al. / Experimental Neurology 187 (2004) 178–189 183
Zi/Zi rats did have significantly lower concentrations of D3
receptors in the NAc (�69%), NAs (�76%), and ICj
(�59%) as compared with SD rats. Zi/Zi rats treated with
L-dopa also demonstrated significantly lower levels of D3
receptors in the NAc (�78% of SD), NAs (�80% of SD),
and Olf Tub (�43% of SD). Treatment of Zi/Zi rats with L-
dopa did not alter the loss of D3 receptors in NAc and NAs as
compared to the Zi/Zi treated with saline (Table 1).
Experiment 2
Analysis in Zi/Zi and Zi/+ rats
As shown in Fig. 1, concentrations of DAT sites were
heterogeneous in the striatum of the Zi/+ rats. Similar to that
of SD control rats, there were higher concentrations of sites
in the CPu than NAs, NAc, or Olf Tub. The concentrations
of DAT sites were considerably lower in the CPu of the Zi/
Zi rats as compared to the Zi/+ rats regardless of whether
they were administered saline or L-dopa. There was no
effect of L-dopa on the concentration of DAT sites. Statis-
tical analysis confirmed these observations as there were
significant group (P < 0.001, F = 20.96), region (P < 0.001,
F = 161.2), and interaction (P < 0.001, F = 11.62) effects
for the analysis of DAT binding (Fig. 2A). The Zi/Zi with
saline group had significantly lower levels of DAT than the
Zi/+ with saline group for the CPu, and the Zi/Zi with L-
dopa group had significantly lower levels of DAT than the
Zi/+ with L-dopa group for the CPu. There were no
significant differences between Zi/+ with saline and Zi/+
with L-dopa or between Zi/Zi with saline and Zi/Zi with L-
dopa groups. The CPu contained higher levels of DAT than
any other region (P < 0.001) for all groups, and within the
Zi/+ with saline and Zi/+ with L-dopa groups, the NAc
contained higher levels than the NAs and Olf Tub.
We also analyzed TH-IR levels in the CPu, nucleus
accumbens (NAs and NAc combined), and Olf Tub, an
indicator of the integrity of DA fibers (Joyce et al., 2002;
Murray et al., 1995; Ryoo et al., 1998), and also in the SN
and VTA, an indicator of the integrity of DA cell bodies
(Joyce et al., 2003,2004; Murray et al., 1995). For the

J.N. Joyce et al. / Experimental Neurology 187 (2004) 178–189184
striatum, there were significant group (P = 0.0002, F =
7.73) but no region or interaction effects (Fig. 3A). TH-IR
was significantly lower in the Zi/Zi with saline group as
compared to the Zi/+ with saline group, and the Zi/Zi with
L-dopa group was significantly lower than the Zi/+ with L-
dopa group. There were no significant differences between
Zi/+ with saline and Zi/+ with L-dopa groups or between Zi/
Zi with saline and Zi/Zi with L-dopa groups. Analysis of
TH-IR in the SN/VTA of the zitter groups revealed signif-
icant group (P < 0.001, F = 18.67), region (P < 0.05, F =
4.297), and interaction (P < 0.05, F = 3.669) effects (Fig.
3B). As would be expected from the previous study (Ueda et
al., 2000), TH-IR was significantly lower in the Zi/Zi with
saline group as compared to the Zi/+ with saline group for
the SN, and the Zi/Zi with L-dopa group was significantly
lower than the Zi/+ with L-dopa group for both the SN and
VTA. There were no significant differences between Zi/+
Fig. 4. Low-power photomicrograph of the original autoradiograph of the caud
receptor binding from individual subjects of four groups: (A) Zi/Zi with saline, (B)
Zi/+ with saline, and (D) Zi/+ with L-dopa/carbidopa. Note the marked reduction of
dopa treatment. The ROI used for analysis is depicted by dashed lines in panel D
with saline and Zi/+ with L-dopa groups or between Zi/Zi
with saline and Zi/Zi with L-dopa groups. Thus, as with
analysis of DAT binding, degeneration of DA fibers as
measured with striatal TH-IR and of TH-IR cell bodies in
SN/VTA was evident in the Zi/Zi rats but not enhanced by
treatment with L-dopa.
As shown in Fig. 4, the concentrations of D3 receptors
were heterogeneously distributed in the striatum of the Zi/+
rats, and similar to SD controls, there were higher concen-
trations of sites in the ICj than NAs, NAc, or the CPu. The
concentrations of D3 receptors were considerably lower in
the CPu of the Zi/Zi rats administered saline or L-dopa as
compared with the Zi/+ rats administered saline or L-dopa.
There was no effect of L-dopa on the concentration of D3
receptors. Statistical analysis confirmed these observations,
as there were significant group (P < 0.001, F = 154.9),
region (P < 0.001, F = 181.1), and interaction (P < 0.001, F
ate–putamen and nucleus accumbens of tissue sections processed for D3
Zi/Zi with L-dopa/carbidopa (100:10 mg/kg ip) twice a day for 10 days, (C)
D3 receptor sites from the caudate–putamen in Zi/Zi and lack of affect of L-
. Abbreviations as in Fig. 1.
J.N. Joyce et al. / Experimental Neurology 187 (2004) 178–189 185
= 27.12) effects for the analysis of D3 receptor binding (Fig.
2B). The Zi/Zi with saline group had significantly lower
levels of D3 receptors than the Zi/+ with saline group for the
NAc, NAs, and ICj; and the Zi/Zi with L-dopa group had
significantly lower concentrations of D3 receptors than the
Zi/+ with L-dopa group for the NAc, NAs, and ICj. There
were no significant differences between Zi/+ with saline and
Zi/+ with L-dopa groups or between Zi/Zi with saline and
Zi/Zi with L-dopa groups. The ICj contained higher con-
centrations of D3 receptors than any other region for all
groups, and within the Zi/+ with saline and Zi/+ with L-dopa
groups, the NAs contained higher levels than the NAc and
was lowest in the CPu.
Analysis in SD control rats
Analysis of DAT binding in the SD groups also demon-
strated significant group (P < 0.05, F = 6.701), region (P <
Fig. 5. Low-power photomicrograph of the original autoradiograph of the forebrai
individual subjects of four groups: (A) Zi/Zi with saline, (B) Zi/Zi with L-dopa/car
(D) Zi/+ with L-dopa/carbidopa. Methods for labeling of the tissue sections with th
used for analysis is depicted by dashed lines in panel A. Abbreviations: CG, cing
0.001, F = 157.5), and interaction (P < 0.05, F = 2.978)
effects (data not shown). The SD with L-dopa group had a
slightly greater level of DAT in the CPu as compared to the
SD with saline group, and the CPu contained higher levels
of DAT than any other region for both groups (P < 0.001).
Analysis of D3 receptor binding in the SD groups also
demonstrated significant region (P < 0.001, F = 121.3) but
not group or interaction effects (Fig. 5B). For both the SD
with saline and SD with L-dopa groups, the ICj contained
higher concentrations of D3 receptors than any other region
and the NAs contained higher levels than the NAc and
binding was lowest in the CPu.
BDNF and TrkB ISHH
Representative photomicrographs of sections for rat
forebrain of Zi/Zi and SD rats treated with L-dopa or saline
and prepared for ISHH for BDNF and TrkB are shown in
n of tissue sections processed for ISHH for antisense to BDNF mRNA from
bidopa (100:10 mg/kg ip) twice a day for 10 days, (C) Zi/+ with saline, and
e 33P-labeled riboprobe is described in the Materials and methods. The ROI
ulate cortex; FR, frontal cortex.

J.N. Joyce et al. / Experimental Neurology 187 (2004) 178–189186
Figs. 5 and 6. Analysis of BDNF ISHH was limited to the
CG and deep layers of the FC and for TrkB, the CG, FC,
CPu, and NA (NAs and NAc combined), as these were
found to be regions with significant alterations in 6-OHDA
lesioned and L-dopa-treated rats (Guillin et al., 2001). While
the levels of BDNF mRNA (Fig. 5) were higher in the CG
as compared to the FR (P < 0.001, F = 18.50), there were
not any differences between the four groups analyzed (Zi/Zi
saline, Zi/Zi L-dopa, SD saline, and SD L-dopa). Analysis of
the TrkB ISHH (Fig. 6) revealed significant group (P <
0.001, F = 10.13) and region (P < 0.001, F = 18.50) but
not interaction effects. There was a consistent pattern of
labeling with the CG higher than the FR, which was higher
than NA and the NA was higher than the CPu. The Zi/Zi
saline group had higher levels of TrkB labeling than the
other groups for the CG, but no other group differences
existed.
Fig. 6. Low-power photomicrograph of the original autoradiograph of the forebra
individual subjects of four groups: (A) Zi/Zi with saline, (B) Zi/Zi with L-dopa/car
(D) Zi/+ with L-dopa/carbidopa. Methods for labeling of the tissue sections with th
used for analysis is depicted by dashed lines in panel A. Abbreviations: CG, cing
Discussion
The results of Experiments 1 and 2 were largely in accord
that reduction of D3 receptors in Zi/Zi rats could not be
reversed with subchronic treatment with L-dopa. We utilized
a regimen of L-dopa treatment that in 6-OHDA-treated rats
does lead to the reversal of the loss and an elevation of D3
receptor binding (Bordet et al., 1997; Guillin et al., 2001).
One of the possible explanations we considered was that the
L-dopa treatment in the Zi/Zi rats might lead to further
damage to the presynaptic DA system, unlike in 6-OHDA-
treated rats, as L-dopa can elevate production of reactive
oxygen species (Blunt et al., 1993; Lai and Yu, 1997;
Naudin et al., 1995; Spencer et al., 1995), and the zitter
mutant rat has abnormal metabolism of oxygen species in
the brain (Gomi et al., 1994; Ueda et al., 2002). In fact, in
Experiment 1, we observed a nonsignificantly lower level of
in of tissue sections processed for ISHH for antisense to TrkB mRNA from
bidopa (100:10 mg/kg ip) twice a day for 10 days, (C) Zi/+ with saline, and
e 33P-labeled riboprobe is described in the Materials and methods. The ROI
ulate cortex; FR, frontal cortex; NA, nucleus accumbens.

J.N. Joyce et al. / Experimental Neurology 187 (2004) 178–189 187
DAT in the Zi/Zi rats treated with L-dopa as compared with
Zi/Zi rats treated with saline. To examine this in more detail,
an additional group of Zi/Zi, Zi/+, and SD rats were
analyzed for the effects of subchronic L-dopa on the pre-
synaptic DA system by measuring both DAT and TH-IR in
the striatum and TH in the midbrain. Those results were
confirmatory, as we did not identify any differences in the
degree of loss of DAT or TH-IR in the striatal regions of the
Zi/Zi rats treated with L-dopa as compared with Zi/Zi with
saline rats. The pattern of loss of DAT in the Zi/Zi rat
striatum observed here is very similar to that described for
degenerative changes in TH-IR fibers in histologically well-
preserved tissue (Ueda et al., 2000). These findings along
with our observations of the reduction of striatal levels of
TH-IR and TH-IR in the SN of the Zi/Zi rats support our
belief that we are accurately measuring any possible impact
of L-dopa on the presynaptic DA system in the Zi/Zi rats.
Not surprisingly, the subchronic treatment with L-dopa in
the SD rats did not lower DAT binding, and L-dopa
treatment has not been observed to alter levels of TH-IR
labeling of DA fibers in SD rats treated for longer periods
with L-dopa (Murer et al., 1998).
We believe our data demonstrate that the failure to
elevate D3 receptor number with L-dopa treatment in zitter
mutant rats does not occur because of further damage to
the DA innervation to the striatum since we did not find
that L-dopa treatment caused a further reduction of DAT
and TH in the striatum nor of TH-IR in SN/VTA. We also
eliminated the possibility that zitter mutation had effects
other than through the DA system, as the zitter hetero-
zygotes had normal levels of D3 receptors and DAT
binding to DA fibers. Another group has examined ex-
pression of a number of neurotrophic factors in the zitter
mutant rat (Muto et al., 1999). Quantification of mRNA
levels for NGF, BDNF, NT-3, ciliary neurotrophic factor,
and GDNF showed that only BDNF was down-regulated
in the cerebrum and brainstem of the adult Zi/Zi rat.
Furthermore, there was an age-related decrease in the
cerebrum that was evident at an early postnatal period in
the brainstem. The timing of this reduction of BDNF in
brainstem is consistent with the slowly progressive degen-
eration of the DA system in the Zi/Zi rat (Ueda et al.,
2000, 2002). Given the evidence for the direct effect of
BDNF on control of expression of the D3 receptor (Guillin
et al., 2001), we hypothesized that the failure for proper
adult expression of the D3 receptor in the Zi/Zi rat is due
to the combination of the lowered BDNF and degeneration
of DA innervation. Furthermore, we hypothesized that the
failure of L-dopa treatment to up-regulate D3 receptor
might be due to deficient cortical expression of BDNF.
In fact, in another study, we have confirmed the findings of
Guillin et al. (2001) that BNDF plays an important role in
the proper development of the D3 receptor as BDNF
heterozygote mice with 1/2 the levels of BDNF demon-
strate significantly lower levels of D3 receptors in forebrain
of adult mice (Joyce et al., 2004). While in this study we
did not observe lower cortical levels of BDNF mRNA in
the Zi/Zi rats as compared with the SD rats, treatment with
L-dopa failed to increase BDNF mRNA in the Zi/Zi or SD
rats. In the study by Guillin et al. (2001), it was also
demonstrated that 6-OHDA lesioned and L-dopa-treated
rats exhibited an elevation of TrkB receptor mRNA, and
L-dopa-induced up-regulation of D3 receptor mRNA was
antagonized by blocking the TrkB receptor with IgG-TrkB.
In this study, there was no difference in TrkB receptor
mRNA in the CPu or NA of the Zi/Zi rats as compared
with the SD rats, and treatment with L-dopa did not alter
TrkB receptor mRNA levels in the Zi/Zi or SD rats. Thus,
the absence of alterations in FR BDNF mRNA and striatal
TrkB receptor mRNA with L-dopa in the Zi/Zi rats could
underlie the resistance to the impact of L-dopa in regulating
expression of the D3 receptor in the Zi/Zi rats.
Interestingly, unlike what has been observed by another
group (Muto et al., 1999), we did not observe lower levels
of BDNF mRNA in FR of Zi/Zi rats. This could suggest that
the lowered D3 receptor number in the Zi/Zi rats is not due
to reduced BDNF availability. In another test of the role of
BDNF and DA fiber loss in the control of expression of D3
receptors in forebrain, we have examined the impact of
methamphetamine toxicity to DA fibers in combination with
lowered BDNF (BDNF +/� mice) on D3 receptors in
forebrain of adult mice (Joyce et al., 2004). We found that
in BDNF +/� mice, with 1/2 levels of BDNF, given a
methamphetamine regimen that produces 50% loss of DA
innervation, there was no greater reduction in the number of
D3 receptors than in the BDNF +/� mice treated with the
vehicle. In MPTP-treated mice with significantly greater
damage to the DA system than achieved with methamphet-
amine treatment, there is a reduction in striatal D3 receptor
number (Gross et al., 2003). In concert, these results would
suggest that reduced BDNF and reduced DA innervation are
not additive in the control of expression of D3 receptors. In
other models of DA denervation with 6-OHDA (Bordet et
al., 1997; Guillin et al., 2001) and MPTP (Morissette et al.,
1998; Wade et al., 2001) that lead to reduced expression of
forebrain D3 receptors, there is massive loss of DA fibers
that is not matched by the zitter mutation (Ueda et al., 2000,
2002). Thus, our current evidence does not support our
hypothesis that BDNF expression plays a predominant role
in regulating expression of the D3 receptor in the Zi/Zi rats.
We believe that there are other factors than the control of
BDNF release by DA fibers that regulate expression of the
D3 receptor. Perhaps additional specific cortical pathology
(Kondo et al., 1995), and/or loss of serotonergic afferents
(Ueda et al., 1998), or loss of cortical DA is required for
deficient control of expression of the D3 receptor in the
zitter rat.
The results of the current study are particularly relevant
for our studies in PD (Joyce et al., 2002; Ryoo et al., 1998),
where we find that a population of PD patients fail to show
normalization of D3 receptors with chronic L-dopa treatment
and this is related to loss of response to APDs and presence

J.N. Joyce et al. / Experimental Neurology 187 (2004) 178–189188
of dementia. Certainly, there is widespread changes in the
cortex of PD cases with dementia (Apaydin et al., 2002;
Hughes et al., 2000; Hurtig et al., 2000; Mattila et al., 2000).
While we believe that the loss of DA fibers and abnormal
morphology of the remaining DA fibers alters axonal flow
of a number of factors that regulate expression of the D3
receptor, other pathological substrates might play a contrib-
utory role. Perhaps additional specific cortical pathology
(Apaydin et al., 2002; Hurtig et al., 2000), and/or loss of
serotonergic afferents (Hornykiewicz, 1998; Jellinger,
1991), or loss of cortical DA (McRitchie et al., 1997) is
required for deficient control of expression of the D3
receptor in PD. Since all of these changes have been
observed to occur in the zitter mutant rat, it further sub-
stantiates the value of the ZI/Zi rat as a pathological model
of PD. Certainly, other models of PD than that caused by
rapid and extensive DA fiber or cell loss induced by 6-
OHDA and MPTP must be considered for exploring the
regulation of the D3 receptor.
Conclusions
The principle finding of the present study is that the zitter
mutant rat model of PD pathology, which demonstrates a
reduction of striatal D3 receptors as in the 6-OHDA and
MPTP models of PD, fails to exhibit a normalization of D3
receptor to L-dopa. Furthermore, we provide evidence that
this may be the result of the failure of L-dopa to regulate
forebrain BDNF and TrkB receptor mRNAs. This is differ-
ent from what is observed in the 6-OHDA and MPTP
models of PD but is comparable to what occurs in PD. In
a recent review (Joyce, 2001b), we discussed the clinical
and preclinical literature that supports the hypothesis that
the initial expression of parkinsonian symptoms is not only
correlated with the loss of DA innervation and an elevation
in receptor number of the D2 subclass, but also a decrease
D3 receptor number. Furthermore, a subgroup of PD cases
that become resistant to the clinical benefits of antiparkin-
sonian mediation also exhibit a sustained loss of D3 recep-
tors (Joyce et al., 2002; Ryoo et al., 1998). The role of the
D3 receptor in the therapeutic response to antiparkinsonian
drugs in PD remains unproven, but recent data support the
hypothesis that it plays an important contributory role as a
selective D3 receptor antagonist blocks the ability of L-dopa
to reverse the parkinsonian symptoms in MPTP-treated
monkeys (Bezard et al., 2003). Hence, identifying factors
that regulate the expression of the D3 receptor may be
important for developing therapeutic interventions for treat-
ing the clinical symptoms of PD.
Acknowledgments
Funded by Federal Grant NS40669. The technical
assistance of Han Ryoo is greatly appreciated.
References
Altar, C.A., Cai, N., Bliven, T., Juhasz, M., Conner, J.M., Acheson, A.L.,
Lindsay, R.M., Wiegand, S.J., 1997. Anterograde transport of brain-
derived neurotrophic factor and its role in the brain. Nature 389,
856–860.
Apaydin, H., Ahlskog, J.E., Parisi, J.E., Boeve, B.F., Dickson, D.W., 2002.
Parkinson disease neuropathology: later-developing dementia and loss
of the levodopa response. Arch. Neurol. 59, 102–112.
Artymyshyn, R., Smith, A., Wolfe, B.B., 1990. The use of 3H standards in
125I autoradiography. J. Neurosci. Methods 32, 185–192.
Bezard, E., Ferry, S., Mach, U., Stark, H., Leriche, L., Boraud, T.,
Gross, C., Sokoloff, P., 2003. Attenuation of levodopa-induced dys-
kinesia by normalizing dopamine D3 receptor function. Nat. Med. 9,
762–767.
Blunt, S.B., Jenner, P., Marsden, C.D., 1993. Suppressive effect of L-dopa
on dopamine cells remaining in the ventral tegmental area of rats pre-
viously exposed to the neurotoxin 6-hydroxydopamine. Mov. Disord. 8,
129–133.
Bordet, R., Ridray, S., Carboni, S., Diaz, J., Sokoloff, P., Schwartz, J.C.,
1997. Induction of dopamine D3 receptor expression as a mechanism of
behavioral sensitization to levodopa. Proc. Natl. Acad. Sci. U.S.A. 94,
3363–3367.
Brooks, D.J., 2000. Dopamine agonists: their role in the treatment of Par-
kinson’s disease. J. Neurol. Neurosurg. Psychiatry 68, 685–689.
Caparros-Lefebvre, D., Pecheux, N., Petit, V., Duhamel, A., Petit, H., 1995.
Which factors predict cognitive decline in Parkinson’s disease?. J. Neu-
rol. Neurosurg. Psychiatry 58, 51–55.
Civelli, O., Bunzow, J.R., Grandy, D.K., 1993. Molecular diversity of the
dopamine receptors. Annu. Rev. Pharmacol. Toxicol. 33, 281–307.
Conner, J.M., Lauterborn, J.C., Yan, Q., Gall, C.M., Varon, S., 1997.
Distribution of brain-derived neurotrophic factor (BDNF) protein and
mRNA in the normal adult rat CNS: evidence for anterograde axonal
transport. J. Neurosci. 17, 2295–2313.
Fabbrini, G., Mouradian, M.M., Juncos, J.L., Schlegel, J., Mohr, E., Chase,
T.N., 1988. Motor fluctuations in Parkinson’s disease: central patho-
physiological mechanisms: Part I. Ann. Neurol. 24, 366–371.
Gomi, H., Ueno, I., Yamanouchi, K., 1994. Antioxidant enzymes in the
brain of zitter rats: abnormal metabolism of oxygen species and its
relevance to pathogenic changes in the brain of zitter rats with genetic
spongiform encephalopathy. Brain Res. 653, 66–72.
Gross, C.E., Ravenscroft, P., Dovero, S., Jaber, M., Bioulac, B., Bezard, E.,
2003. Pattern of levodopa-induced striatal changes is different in normal
and MPTP-lesioned mice. J. Neurochem. 84, 1246–1255.
Guillin, O., Diaz, J., Carroll, P., Griffon, N., Schwartz, J.C., Sokoloff, P.,
2001. BDNF controls dopamine D3 receptor expression and triggers
behavioural sensitization. Nature 411, 86–89.
Gurevich, E.V., Joyce, J.N., 1999. Distribution of dopamine D3 receptor
expressing neurons in the human forebrain: comparison with D2 recep-
tor expressing neurons. Neuropsychopharmacology 20, 60–80.
Gurevich, E.V., Himes, J.W., Joyce, J.N., 1999. Developmental regulation
of expression of the D3 dopamine receptor in rat nucleus accumbens
and islands of Calleja. J. Pharmacol. Exp. Ther. 289, 587–598.
Hornykiewicz, O., 1998. Biochemical aspects of Parkinson’s disease. Neu-
rology 51, S2–S9.
Hughes, T.A., Ross, H.F., Musa, S., Bhattacherjee, S., Nathan, R.N., Mind-
ham, R.H., Spokes, E.G., 2000. A 10-year study of the incidence of and
factors predicting dementia in Parkinson’s disease. Neurology 54,
1596–1602.
Hurtig, H.I., Trojanowski, J.Q., Galvin, J., Ewbank, D., Schmidt, M.L.,
Lee, V.M., Clark, C.M., Glosser, G., Stern, M.B., Gollomp, S.M.,
Arnold, S.E., 2000. Alpha-synuclein cortical Lewy bodies correlate
with dementia in Parkinson’s disease. Neurology 54, 1916–1921.
Isackson, P.J., Huntsman, M.M., Murray, K.D., Gall, C.M., 1991. BDNF
mRNA expression is increased in adult rat forebrain after limbic seiz-
ures: temporal patterns of induction distinct from NGF. Neuron 6,
937–948.

J.N. Joyce et al. / Experimental Neurology 187 (2004) 178–189 189
Jellinger, K.A., 1991. Pathology of Parkinson’s disease. Changes other than
the nigrostriatal pathway. Mol. Chem. Neuropathol. 14, 153–197.
Joyce, J.N., 1991. Differential response of striatal dopamine and muscarinic
cholinergic receptor subtypes to the loss of dopamine: I. Effects of
intranigral or intracerebroventricular 6-hydroxydopamine lesions of
the mesostriatal dopamine system. Exp. Neurol. 113, 261–276.
Joyce, J.N., 2001a. D2 but not D3 receptors are elevated after 9 or 11
months chronic haloperidol treatment: influence of withdrawal period.
Synapse 40, 137–144.
Joyce, J.N., 2001b. Dopamine D3 receptor as a therapeutic target for anti-
psychotic and antiparkinsonian drugs. Pharmacol. Ther. 90, 231–259.
Joyce, J.N., Marshall, J.F., Bankiewicz, K.S., Kopin, I.J., Jacobowitz,
D.M., 1986. Hemiparkinsonism in a monkey after unilateral internal
carotid artery infusion of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine
(MPTP) is associated with regional ipsilateral changes in striatal dopa-
mine D-2 receptor density. Brain Res. 382, 360–364.
Joyce, J.N., Yoshimoto, K., Ueda, S., 2000. The zitter mutant rat exhibits
loss of D3 receptors with degeneration of the dopamine system. Neuro-
Report 11, 2173–2175.
Joyce, J.N., Ryoo, H.L., Beach, T.B., Caviness, J.N., Stacy, M., Gurevich,
E.V., Reiser, M., Adler, C.H., 2002. Loss of response to levodopa in
Parkinson’s disease and co-occurrence with dementia: role of D3 and
not D2 receptors. Brain Res. 955, 138–152.
Joyce, J.N., Presgraves, S., Renish, L., Borwege, S., Hagner, D., Osred-
kar, T., Replogle, M., Paz Soldan, M.M., Millan, M.J., 2003. Neuro-
protective effects of the novel D3/D2 receptor agonist and anitparkinson
agent S32504, in vitro against 1-methyl-4-phenylpyridinium (MPP+)
and in vivo against 1-methyl- 4-phenyl-1,2,2,6-tetrahydropyridine
(MPTP): a comparison to ropinirole. Exp. Neurol. 184, 393–407.
Joyce, J.N., Renish, L., Osredkar, T., Walro, J.M., Kucera, J., Dluzen, D.E.,
2004. Methamphetamine induced loss of striatal dopamine innervation
in BNDF heterozygote mice does not further alter D3 receptor concen-
trations. Synapse 52, 11–19.
Klein, R., Conway, D., Parada, L.F., Barbacid, M., 1990. The trkB tyrosine
protein kinase gene codes for a second neurogenic receptor that lacks
the catalytic kinase domain. Cell 61, 647–656.
Kondo, A., Sendoh, S., Miyata, K., Takamatsu, J., 1995. Spongy degener-
ation in the zitter rat: ultrastructural and immunohistochemical studies.
J. Neurocytol. 24, 533–544.
Lai, C.T., Yu, P.H., 1997. Dopamine- and L-beta-3,4-dihydroxyphenylala-
nine hydrochloride (L-dopa)- induced cytotoxicity towards catechol-
aminergic neuroblastoma SH-SY5Y cells. Effects of oxidative stress
and antioxidative factors. Biochem. Pharmacol. 53, 363–372.
Levesque, D., Martres, M.P., Diaz, J., Griffon, N., Lammers, C.H., Sokol-
off, P., Schwartz, J.C., 1995. A paradoxical regulation of the dopamine
D3 receptor expression suggests the involvement of an anterograde
factor from dopamine neurons. Proc. Natl. Acad. Sci. U.S.A. 92,
1719–1723.
Marder, K., Tang, M.X., Cote, L., Stern, Y., Mayeux, R., 1995. The fre-
quency and associated risk factors for dementia in patients with Parkin-
son’s disease. Arch. Neurol. 52, 695–701.
Mattila, P.M., Rinne, J.O., Helenius, H., Dickson, D.W., Roytta, M., 2000.
Alpha-synuclein-immunoreactive cortical Lewy bodies are associated
with cognitive impairment in Parkinson’s disease. Acta Neuropathol.
(Berl.) 100, 285–290.
McRitchie, D.A., Cartwright, H.R., Halliday, G.M., 1997. Specific A10
dopaminergic nuclei in the midbrain degenerate in Parkinson’s disease.
Exp. Neurol. 144, 202–213.
Morissette, M., Goulet, M., Grondin, R., Blanchet, P., Bedard, P.J., Di
Paolo, T., Levesque, D., 1998. Associative and limbic regions of mon-
key striatum express high levels of dopamine D3 receptors: effects of
MPTP and dopamine agonist replacement therapies. Eur. J. Neurosci.
10, 2565–2573.
Mouradian, M.M., Juncos, J.L., Fabbrini, G., Schlegel, J., Bartko, J.J.,
Chase, T.N., 1988. Motor fluctuations in Parkinson’s disease: central
pathophysiological mechanisms, Part II. Ann. Neurol. 24, 372–378.
Murer, M.G., Dziewczapolski, G., Menalled, L.B., Garcia, M.C., Agid, Y.,
Gershanik, O., Raisman-Vozari, R., 1998. Chronic levodopa is not toxic
for remaining dopamine neurons, but instead promotes their recovery, in
rats with moderate nigrostriatal lesions. Ann. Neurol. 43, 561–575.
Murray, A.M., Weihmueller, F.B., Marshall, J.F., Hurtig, H.I., Gottleib,
G.L., Joyce, J.N., 1995. Damage to dopamine systems differs between
Parkinson’s disease and Alzheimer’s disease with parkinsonism. Ann.
Neurol. 37, 300–312.
Muto, Y., Hayashi, T., Higashi, Y., Endo, T., Yamamoto, T., Sato, K., 1999.
Age-related decrease in brain-derived neurotrophic factor gene expres-
sion in the brain of the zitter rat with genetic spongiform encephalop-
athy. Neurosci. Lett. 271, 69–72.
Naudin, B., Bonnet, J.J., Costentin, J., 1995. Acute L-DOPA pretreatment
potentiates 6-hydroxydopamine-induced toxic effects on nigro-striatal
dopamine neurons in mice. Brain Res. 701, 151–157.
Paxinos, G., Watson, C., 1986. The Rat Brain in Stereotaxic Coordinates.
Academic Press, Sydney Australia.
Pizzolato, G., Chierichetti, F., Rossato, A., Cagnin, A., Fabbri, M., Dam, M.,
Ferlin, G., Battistin, L., 1995. Alterations of striatal dopamine D2 recep-
tors contribute to deteriorated response to L-dopa in Parkinson’s disease:
a [123I]-IBZM SPET study. J. Neural. Transm., Suppl. 45, 113–122.
Portin, R., Rinne, U.K., 1987. Predictive factors for cognitive deterioration
and dementia in Parkinson’s disease. Adv. Neurol. 45, 413–416.
Quik, M., Police, S., He, L., Di Monte, D.A., Langston, J.W., 2000. Ex-
pression of D3 receptor messenger RNA and binding sites in monkey
striatum and substantia nigra after nigrostriatal degeneration: effect of
levodopa treatment. Neuroscience 98, 263–273.
Ryoo, H.L., Pierrotti, D., Joyce, J.N., 1998. Dopamine D3 receptor is
decreased and D2 receptor is elevated in the striatum of Parkinson’s
disease. Mov. Disord. 13, 788–797.
Spencer, J.P., Jenner, P., Halliwell, B., 1995. Superoxide-dependent deple-
tion of reduced glutathione by L-DOPA and dopamine. Relevance to
Parkinson’s disease. NeuroReport 6, 1480–1484.
Stern, Y., Marder, K., Tang, M.X., Mayeux, R., 1993. Antecedent clinical
features associated with dementia in Parkinson’s disease. Neurology 43,
1690–1692.
Ueda, S., Aikawa, M., Ishizuya-Oka, A., Koibuchi, N., Yamaoka, S., Yosh-
imoto, K., 1998. Age-related degeneration of the serotoninergic fibers
in the zitter rat brain. Synapse 30, 62–70.
Ueda, S., Aikawa, M., Ishizuya-Oka, A., Yamaoka, S., Koibuchi, N.,
Yoshimoto, K., 2000. Age-related dopamine deficiency in the meso-
striatal dopamine system of zitter mutant rats: regional fiber vulnera-
bility in the striatum and the olfactory tubercle. Neuroscience 95,
389–398.
Ueda, S., Sakakibara, S., Watanabe, E., Yoshimoto, K., Koibuchi, N., 2002.
Vulnerability of monoaminergic neurons in the brainstem of the zitter
rat in oxidative stress. Prog. Brain Res. 136, 293–302.
Wade, T.V., Rothblat, D.S., Schneider, J.S., 2001. Changes in striatal do-
pamine D3 receptor regulation during expression of and recovery from
MPTP-induced parkinsonism. Brain Res. 905, 111–119.




















