
Electrostatic Charging of Nonpolar Colloids by Reverse Micelles
Upload
independentCategory
view
1download
0
TOPICAL PAPERLiquid-Liquid Extraction of Proteins with Reversed MicellesM. J. Pires, M. R. Aires-Barros,* and J. M. S. Cabral
Laboratorio de Engenharia Bioquımica, Instituto Superior Tecnico, Lisboa 1000, Portugal
The recovery of proteins using reversed micelles is a liquid-liquid extraction processthat has received increasing attention since proteins were shown to be solubilized inorganic solvents with surfactants, maintaining their functional properties, and to betransferred between an aqueous solution and a reversed micellar organic phase. Thisarticle reviews the application of reversed micellar systems as a bioseparationtechnique for isolation and purification of proteins. The parameters that affect proteinsolubilization into the reversed micelles and the equilibrium and kinetics aspects thatare involved in the extraction and back-extraction of proteins are discussed. Severalexamples are also described including the application of this technique for purificationof recombinant proteins: cytochrome b5 and a cutinase from Fusarium solani pisi.
Contents
1. Introduction 2902. Factors Affecting Protein Transfer 2912.1. pH 2912.2. Ionic Strength 2912.3. Type of Electrolyte 2922.4. Surfactant Concentration 2922.5. Charge Distribution 2922.6. Other Factors Affecting Reversed
Micelle Structure292
2.7. Use of Biospecific Ligands 292
3. Driving Forces Involved in theTransfer of Proteins into ReversedMicelles
293
3.1. Electrostatic Interactions 2933.2. Hydrophobic Interactions 294
4. Driving Forces for the Back-Extractionof Proteins in Reversed Micelles
295
5. Interfacial Transport of Proteins intoand out of Reversed Micelles
295
6. Applications 2966.1. Selective Separation of Mixtures of
Proteins296
6.2. Recovery of Enzymes fromFermentation Broth
296
6.3. Recovery of Intracellular Enzymes 2976.4. Continuous Extraction of r-Amylase 2976.5. Liquid-Liquid Extraction of
Recombinant Proteins297
6.5.1. Cytochrome b5 2976.5.2. Cutinase from Fusarium
solani pisi298
7. Liquid-Liquid Extraction of Proteinswith Reversed Micelles: Strategies ofOperation and Scale-up
299
8. Conclusions 300
1. Introduction
The production of foreign proteins using a selected hostwith the necessary post-translational modifications is oneof the key successes in modern biotechnology. Thismethodology allows the industrial production of proteinsthat were otherwise produced in a small scale. Theseparation and purification of these proteins from thefermentation medium constitutes a major bottleneck forthe widespread commercialization of proteins producedin this way (Van Brakel and Kleizen, 1990).Three types of situation might occur when producing
recombinant proteins: the protein is accumulated ineither the cytoplasm, the periplasm, or the extracellularmedium. The intracellular protein can also form ag-gregates with highly dense structures called inclusionbodies. The choice of the purification scheme will dependon these factors and on the degree of purity of theproduct, which depends on the further utilization of theprotein. As a first step, after cell lysis, if the protein isnot secreted, the cellular material is separated from theliquid medium, using a centrifugation or a filtration step,followed by chromatographic steps.It has been demonstrated that it is possible to use
liquid-liquid extraction technology to integrate the firsttwo steps into one to obtain separation of the productfrom the cellular material with simultaneous concentra-tion of the protein. Two different systems were proposedand are in study presently. The first one used twoimmiscible aqueous phases of simple electrolytes andwater soluble polymers (like polyethylene) or of incom-patible water soluble polymers (e.g., dextran/polyethyl-ene) (Kula and Kroner, 1982; Huddleston et al., 1991).The second one was developed more recently and uses awater in oil microemulsion (defined by the parameter w0
) [H2O/[surfactant]) with an excess of the aqueous phaseto achieve the desired separation/concentration of theproduct (Goklen and Hatton, 1985, 1987; Rahaman et al.,1988). In both cases, it has been claimed that the easeof scale up and the high partition coefficients obtainedallow its application in the downstream processing ofproteins produced by fermentation.
290 Biotechnol. Prog. 1996, 12, 290−301
S8756-7938(95)00050-6 CCC: $12.00 © 1996 American Chemical Society and American Institute of Chemical Engineers

This review deals with the liquid-liquid extraction ofproteins using microemulsions. The factors affectingprotein transfer and the interactions which are respon-sible for the extraction and back-extraction of proteins
from these systems are described, as well as the applica-tions of this new technology to the purification ofrecombinant proteins.
2. Factors Affecting Protein TransferThe distribution of proteins between a micellar organic
phase and an aqueous solution is largely determined bythe conditions in the aqueous bulk phase, namely pH,ionic strength, and type of salt. The parameters relatedto the organic phase also influence the partition of aprotein, such as the concentration and type of surfactant,presence of co-surfactant, and type of solvent. Changesin temperature can also affect the solubilization ofbiomolecules. The phase transfer depends on the specificcharacteristics of the proteins, namely the isoelectricpoint, size and shape, hydrophobicity, and charge distri-bution.2.1. pH. The pH of the aqueous solution determines
the net charge of proteins. Electrostatic interactionsbetween the protein and the surfactant head groups canfavor the transfer of protein into the organic phase. Thistrend has been observed using anionic (Goklen andHatton, 1987; Goklen, 1986) and cationic surfactants(van’t Riet and Dekker, 1984; Dekker et al., 1987),solubilization occurring only when the protein and thesurfactant head groups have opposite charges. However,this is not always observed, and Luisi et al. (1979)obtained identical pH profiles for R-chymotrypsin (pI )8.3) and pepsin (pI < 1.1) using TOMAC, a cationicsurfactant in cyclohexane. Similar results were reportedby Goklen (1986) with another cationic surfactant, di-dodecyldimethylammonium bromide (DDAB) and a mix-ture of proteins with isoelectric points ranging from 5.5to 7.8. In both studies using quaternary ammoniumsalts, the transfer of proteins into the reversed micellarphase is more effective in the alkaline pH range (lessprotonated residues on the protein), which could beexplained by ion pairing between the anionic surfaces ofthe protein and the positively-charged surfactant. Luisiet al. (1979) found sharp decreases in the transfer at pHhigher than 13 for R-chymotrypsin and pepsin, attributedto competition of the proteins with OH- ions. Proteindenaturation and changes in the ionization state of thesurfactant at extreme pH values make additionallydifficult the interpretation of the phase-transfer pattern.2.2. Ionic Strength. Increasing ionic strength of the
aqueous phase will reduce electrostatic interactions as aresult of the Debye screening effect, stronger for largerions. Thus, at a higher ionic strength, the interactionsbetween hydrophilic biomolecules and the surfactantpolar groups are reduced, smaller micelles being formed.As a consequence of this effect of increasing the ionicstrength of the aqueous solution, the solubilizationcapacity of the organic phase for water and biomoleculeswill decrease. The amount of water solubilized in thereversed micelles is commonly referred to as wo, themolar ratio of water and surfactant (wo ) [H2O]/[surfac-tant]). Complete solubilization of three proteins, cyto-chrome c, lysozyme, and ribonuclease A, using an AOT/isooctane system, was obtained at low ionic strength (0.1M KCl), and no solubilization occurred at high ionicstrength (1.0 M KCl) (Goklen and Hatton, 1987). Theinversion of the partition coefficients is achieved in a verynarrow range of pH, which depends on the protein speciesand does not correlate with the isoelectric point of theprotein. Ribonuclease A is the protein with the lowerisoelectric point, but a sharp change of solubilizationbehavior occurs at a KCl concentration (0.5 M) lower thanthat in the case of lysozyme (0.7 M) Goklen and Hatton,1987). The importance of the particular electrostatic
Mario Pires has worked as a Product Group Manager inthe bread ingredients business area, at Gist-Brocades in Lis-bon, since 1993. He received a B.Sc. in Biochemistry (1987)from the Science Faculty of Lisbon, Portugal, and an M.Sc.in Biochemical Engineering (1990) and a Ph.D. in ChemicalEngineering (1993) from the Technical University of Lisbon.
Joaquim M. S. Cabral is a Full Professor in the Depart-ment of Chemical Engineering, Instituto Superior Tecnico,Technical University of Lisbon, Portugal. He received a Di-ploma in Chemical Engineering and Ph.D. and D.Sc. degreesfrom the Technical University of Lisbon. He is the chairmanof the Working Party on Applied Biocatalysis of the Euro-pean Federation of Biotechnology and is a member of theSteering Committee of the European Science FoundationProgramme in Process Integration in Biochemical Engineer-ing. His research interests are in biocatalysis in noncon-ventional media, bioseparation, and process integration.
M. Raquel Aires-Barros has been an Associate Professorin the Department of Chemical Engineering Instituto Su-perior Tecnico, Technical University of Lisbon, Portugal,since 1987. She received a Diploma in Chemical Engineer-ing (1984), an M.Sc. in Biochemical Engineering (1987), anda Ph.D. in Chemical Engineering (1990) from the TechnicalUniversity of Lisbon. Her research interests are in the areasof enzymatic technology, biocatalysis in nonconventionalmedia, and recovery of biological products.
Biotechnol. Prog., 1996, Vol. 12, No. 3 291

characteristics on the protein surface is suggested bythese results. Participation of electrostatic interactionsin phase transfer is supported by the narrowing of thepH peaks when the ionic strength is increased, asobserved for the extraction of cytochrome c (Goklen,1986). These results, found with anionic surfactant AOT,are consistent with the phase-transfer behavior of R-amy-lase (Dekker et al., 1986) in a system including TOMAC,a cationic surfactant.Additional effects of the ionic strength in the solubi-
lization of proteins in reversed micellar biphasic systemsare competition with ionic species for transfer into thereversed micelles and changes in the electrostatic stateof the micelles and/or proteins, more likely to occur athigh ionic strength.It has been noticed that, when the ionic strength of
the aqueous phase is below a certain limit, reversedmicelles and phase separation do not occur, a stablemicroemulsion being formed. Therefore, the transfer ofproteins between phases requires a minimum value forthe ionic strength of the aqueous solution. A minimumconcentration of KCl around 0.1 M was found to achievequantitative transfer of cytochrome c into the AOT/isooctane system (Goklen and Hatton, 1985). However,if the aqueous solution is buffered, the buffer can supplyenough of the electrolyte.2.3. Type of Electrolyte. The salt composition of the
aqueous phase can also affect the efficiency of transferof proteins between phases, apart from the expecteddifferent contribution to the ionic strength of the solution.Specific interactions of the protein and/or the surfactantwith the ions in solution can account for a change in thesolubilization pattern of proteins at given conditions, inthe presence of different ionic species (Luisi et al., 1983;Leser et al., 1986; Goklen, 1986; Castro et al., 1988;Marcozzi et al., 1991). The buffering system itself caninfluence the solubilization behavior of proteins (Goklen,1986; Meier et al., 1984).2.4. Surfactant Concentration. Increasing the
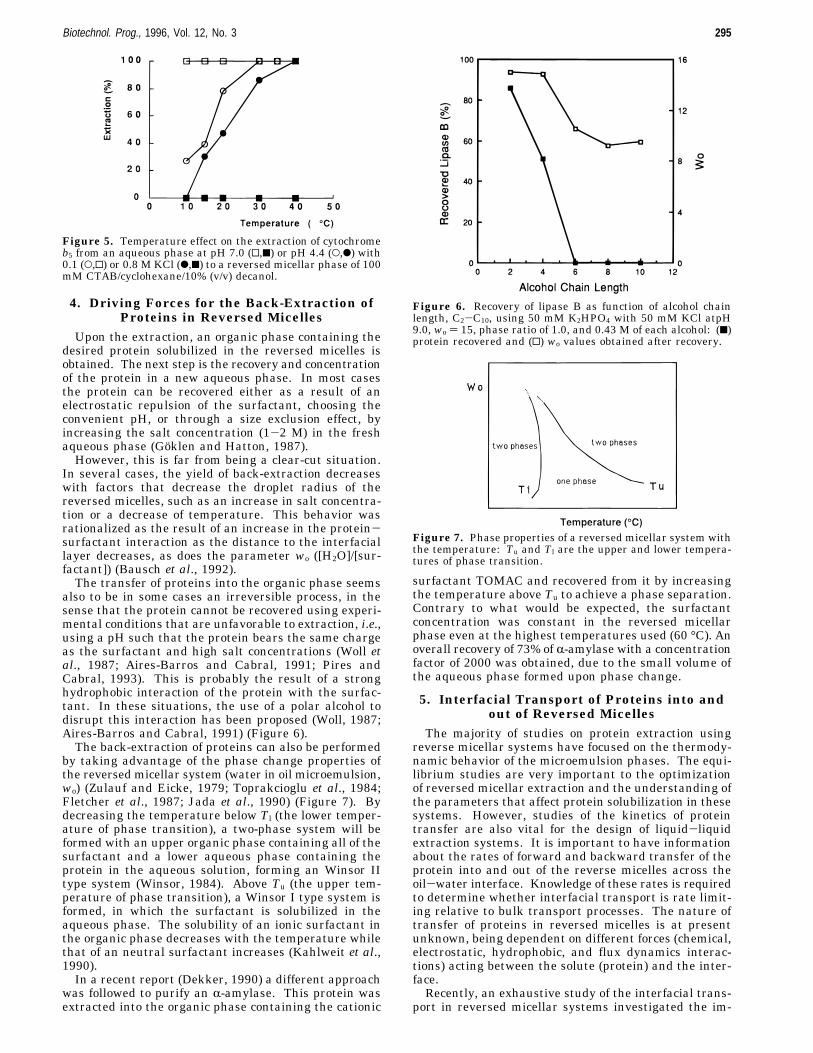
surfactant concentration favors protein solubilization inthe organic phase (Hentsch et al., 1992; Carneiro-da-Cunha et al., 1994a), resulting in broader pH-solubiliza-tion peaks (Goklen, 1986). On the other hand, highersurfactant concentrations make difficult the backwardtransfer of proteins into a second aqueous phase (Castroet al., 1988; Hentsch et al., 1992; Carneiro-da-Cunha etal., 1994a). Therefore, the optimum surfactant concen-tration in a double-phase transfer corresponds to theminimum limit for achieving maximum transfer into theorganic phase.The selectivity of the reversed micellar phase for a
given protein can thus be changed by choosing anappropriate surfactant concentration in the organicphase. For example, smaller proteins will be more easilyextracted into the organic phase than proteins withhigher molecular weight, at low surfactant concentra-tions.These results also point out to a limitation of the size
of the protein that can be extracted from the aqueousphase. Although one could solve this problem by increas-ing the surfactant concentration when dealing with largeproteins (>100 000 Da), this has a spurious effect on thephase separation by decreasing it (Goklen and Hatton,1986; Ishikawa et al., 1990).2.5. Charge Distribution. The yield of extraction
depends on the total charge present on the protein, sinceit depends strongly on the pH of the aqueous phase. Onemight ask if the charge distribution on the protein surfaceis also important for this matter. This question wasaddressed for the first time on a paper by Wolbert et al.
(1989). These authors found that proteins with highercharge asymmetry are more easily extracted into anorganic phase containing trioctylmethylammonium chlo-ride (TOMAC) surfactant and obtained a correlationbetween the yield of extraction and the degree of chargeasymmetry (calculated as Barlow and Thornton, 1986)of the protein. This was not a general observation, sincethe same was not valid when using AOT reversedmicelles.In another work (Pires et al., 1994), the solubilization
of native (pI ) 4.5) and modified (pI ) 4.7) cytochromesb5, by site-specific mutagenesis, with a substitution of aglutamic acid at positions 44, 56, and 92 of the polypep-tide chain for a lysine, to a reversed micelle phase of AOT/octane was studied. The percent area of positive poten-tial caused by basic amino acid residues was calculatedat the surface of the protein (Table 1). A protein titrationprogram, TITRA, developed by P. Martel and S. Petersen(results not published) was used to compute the averagecharge distribution of the protein at a given pH, using amodified Tanford-Kirkwood method to calculate theelectrostatic interactions between titrable groups in theprotein (Shire et al., 1974; Tanford and Roxby, 1972). Byplotting the electrostatic potential on the molecularsurface, the size of the negatively and positively chargedareas can be estimated. A positive correlation was foundwith this parameter (Table 1), i.e., higher extractionyields corresponded with higher positive surface charge.In this case, although the cytochrome was extracted atan unfavorable pH, it was nonetheless demonstrated thatthe extraction is governed by an electrostatic mechanism.2.6. Other Factors Affecting Reversed Micelle
Structure. The type of organic solvent affects the sizeof the reversed micelles and consequently the watersolubilization capacity (Goklen, 1986). A different mi-cellar structure resulting from the solvent effect can beexploited to achieve significant changes in the solubili-zation of R-chymotrypsin with different solvents (Luisiet al., 1979). When comparing the results obtained withseveral proteins, in a system including cyclohexane asorganic solvent, the transfer efficiencies were also verydifferent.The introduction of a cosurfactant can improve the
solubility capacity of the organic phase (Dekker et al.,1987), probably due to an increase in the micelle size,when using cationic surfactants.Changes in temperature have drastic effects on the
physicochemical properties of the reversed micellar sys-tem. Increasing the temperature can enhance the solu-bility of proteins in the organic phase. Luisi et al. (1979)obtained increases of 50% and 100% in the transfer yieldsof R-chymotrypsin and glucagon, respectively, into aNH4
+/chloroform phase.2.7. Use of Biospecific Ligands. Significant en-
hancements in the selectivity of the protein extractionprocess with reversed micellar systems can be achievedby introducing affinity ligands in the organic phase (Wollet al., 1989; Coughlin and Baclaski, 1990; Paradkar andDordick, 1991; Kelley et al., 1993).An increased partitioning in the transfer of concavalin
A into reversed micelles was obtained (Woll et al., 1989)
Table 1. Positive Electrostatic Surface Areas inCytochrome b5 and Mutants as a Function of pH
pH native E44K E56K E92K
4.0 57.9 61.6 62.7 645.0 36.7 41.1 42.7 476.0 31.4 35.8 37.6 417.0 26.7 31. 32.3 358.0 21.4 25.8 25.7 29.5
292 Biotechnol. Prog., 1996, Vol. 12, No. 3

if the biosurfactant octyl â-D-glycopyran (2-20% of totalsurfactant concentration) was included in the organicphase. The transfer of the control protein, ribonucleaseA was not affected by the presence of the biologicaldetergent, while glucose inhibited the solubilization ofconcanavalin A due to competition with the ligand fortwo binding sites in the protein. High values for protein/ligand binding constants in the organic phase, ligandpartition coefficient between the organic and the aqueousphases, and number of binding sites in the protein favorthe solubilization of proteins in reversed micelles, whereasa low protein/ligand constant in the aqueous solution isdesired.Increased selectivity for the extraction of avidin into
reversed micelles was also achieved by a using an affinityligand, biotinylated, incorporated into the micelle as acosurfactant (Coughlin and Baclaski, 1990).Liquid-liquid extraction coupling with an affinity
interaction can be used for selective and efficient puri-fication and separation of glycoproteins, namely horse-radish peroxidase (Paradkar and Dordick, 1991). Thebasis of the separation is the selective extraction ofglycoproteins from and aqueous solution into a reversemicellar organic phase by using concanavalin A (a sugar-binding lectin) as a facilitate carrier.Kelly and co-workers (1993) used affinity cosurfactants,
consisting of hydrophilic ligands derivatized with hydro-phobic tails, to purify concanavalin A, a membrane-associated protein and chymotrypsin. The addition of asmall concentration of the affinity cosurfactants to themicellar phase extends the operating range of pH andsalt concentration over which proteins can be extracted.
3. Driving Forces Involved in the Transfer ofProteins into Reversed Micelles
3.1. Electrostatic Interactions. The liquid-liquidextraction of proteins with reversed micelles has beenstudied with more than 30 proteins (Table 2). It can besaid that presently one does not have still a definitiveanswer on how the proteins are extracted and back-
extracted using these systems. In most cases it isaccepted that the driving force for this process is anelectrostatic interaction between the charged polar headsof the surfactant and opposite charged groups in theprotein (Luisi and Magid, 1986; Hatton, 1989; Dekker etal., 1989; Cabral and Aires-Barros, 1993). However,there are reported studies indicating that other forcesmay also play an important role in the solubilizationmechanism, such as hydrophobic interactions (Aires-Barros and Cabral, 1992; Pires and Cabral, 1992; Piresand Cabral, 1993) and solubilization by an ion-pairingmechanism (Hatton, 1989).The transfer of proteins to the reversed micellar phase
depends on several factors as pointed out in section 2.From the dependence of the yield of extraction on thepH and salt concentration of the aqueous phase, it wasdeduced that electrostatic interactions between thecharged groups in the protein and the ionic groups of thesurfactant play an important role in the solubilizationmechanism of proteins into the reversed micellar phase(Dekker et al., 1986; Goklen and Hatton, 1987; Marcozziet al., 1991).Figure 1 illustrates the extraction curves usually
obtained for protein solubilization in a micellar organicphase of anionic (curve A1) and cationic (curve C1)surfactants. However, it has been reported (Hatton,1989; Wolbert et al., 1989) that this behavior is observedonly for small proteins (around 20 000 Da) and at lowsalt concentrations (around 100 mM). In fact, a linearcorrelation of |pHext - pI| with the molecular weight ofthe extracted protein was obtained, where pHext is theoptimum of pH for maximum extraction and pI itsisoelectric point (Wolbert et al., 1989). The higher themolecular weight of the protein the greater the shift fromthe isoelectric point. The difference (pHext - pI) waspositive when using an anionic surfactant and negativewhen using a cationic surfactant. These results wereexplained in terms of a necessity for more electrostaticinteractions between the surfactant and the protein asit size increases (Wolbert et al., 1989). In the same way,
Table 2. Examples of Proteins Extracted into Reversed Micelles
protein pI surfactant MW (Da) refa
albumin 4.6 AOT 65 000 3, 5, 17alcohol dehydrogenase 5.7 TOMAC 68 000 5alkaline protease 9 TOMAC; AOT 33 000 4, 5R-amylase 4.6 TOMAC; CTAB; BDBAC; CPB; CPC; AOT 55 000 2, 5, 6, 10carbonic anhydrase 4.4 DDAB 31 000 2, 5R-chymotrypsin 6.0 AOT; TOMAC 25 000 3, 5, 9, 11, 12, 13, 18cytochrome b5 4.4 CTAB; AOT 13 600 19cytochrome c 9.4 DDAB; Tween 85/Span-85; AOT 12 000 1, 5, 7, 8cytochrome c3 5.2; 7.0; 10.2 AOT 13 000 16elastase 7.6 AOT 25 000 2flavodoxin 3.7 TOMAC 15 000 5hemoglobin 4.4-5.4 Tween 85/Span-85; TOMAC 60 000-64 000 5, 8hexokinase 4.6-4.0 TOMAC 100 000 5trypsin inhibitor 4.5 TOMAC 24 500 5insulin 4.6 TOMAC 12 000 5lipase A 3.6 AOT 120 000 14, 15lipase B 6.3 AOT 30 000 14, 15lysozyme 11.0 AOT; TOMAC 14 400 1, 5parvalbumin 4.3 TOMAC 12 000 5pepsin <1.0 TOMAC 34 000 5, 12peroxidase 5.6 TOMAC 40 000 5renin 4.6 AOT 43 000 2ribonuclease A 6.7 DDAB; TOMAC 13 700 1, 5rubredoxin 3.2 TOMAC 5 000 5superoxide dismutase 4.3 TOMAC 14 500 5trypsin 9.4 AOT; TOMAC 25 000 2, 5, 12a References: (1) Goklen and Hatton, 1987; (2) Goklen, 1986; (3) Goklen and Hatton, 1986; (4) Rahaman et al., 1988; (5) Wolbert et al.,
1989; (6) Krei and Hustedt, 1992; (7) Ishikawa et al., 1992; (8) Ayala et al., 1992; (9) Marcozzi et al., 1991; (10) Dekker et al., 1989; (11)Luisi et al., 1977; (12) Luisi et al., 1979; (13) Hentsch et al., 1992; (14) Aires-Barros and Cabral, 1991; (15) Camarinha-Vicente et al.,1990; (16) Castro et al., 1988; (17) Armstrong and Li, 1988; (18) Jolivalt et al., 1990; (19) Pires and Cabral, 1993.
Biotechnol. Prog., 1996, Vol. 12, No. 3 293
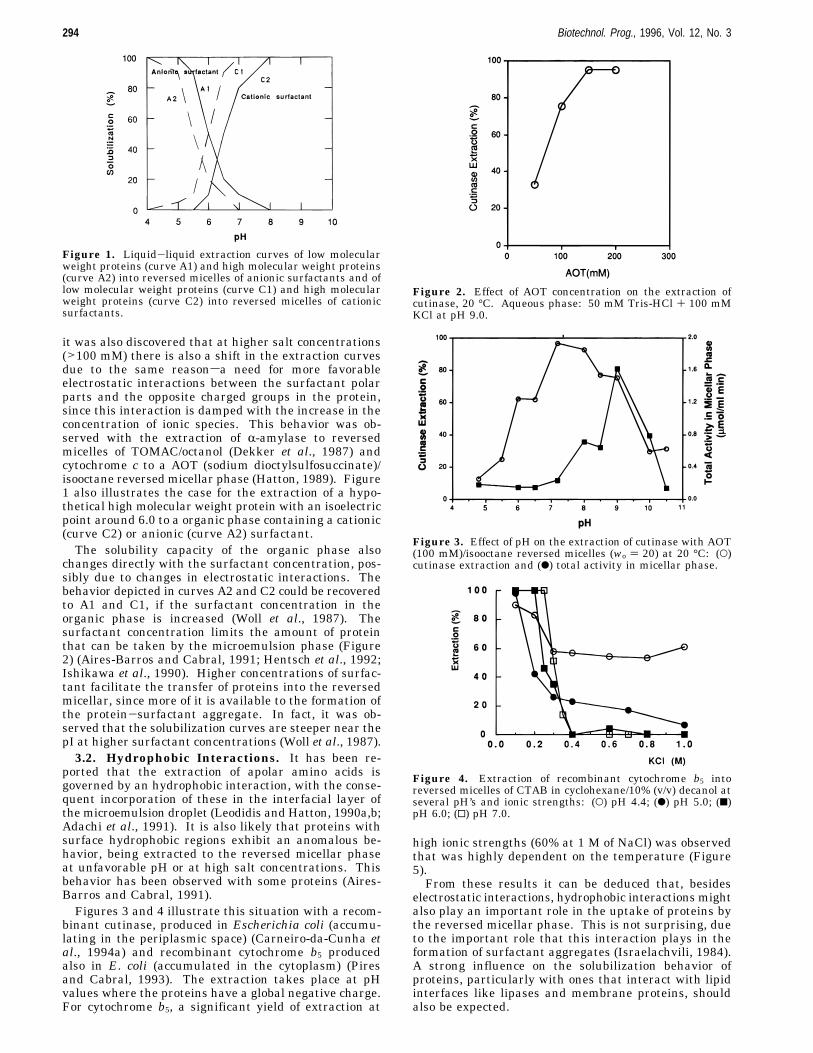
it was also discovered that at higher salt concentrations(>100 mM) there is also a shift in the extraction curvesdue to the same reasonsa need for more favorableelectrostatic interactions between the surfactant polarparts and the opposite charged groups in the protein,since this interaction is damped with the increase in theconcentration of ionic species. This behavior was ob-served with the extraction of R-amylase to reversedmicelles of TOMAC/octanol (Dekker et al., 1987) andcytochrome c to a AOT (sodium dioctylsulfosuccinate)/isooctane reversed micellar phase (Hatton, 1989). Figure1 also illustrates the case for the extraction of a hypo-thetical high molecular weight protein with an isoelectricpoint around 6.0 to a organic phase containing a cationic(curve C2) or anionic (curve A2) surfactant.The solubility capacity of the organic phase also
changes directly with the surfactant concentration, pos-sibly due to changes in electrostatic interactions. Thebehavior depicted in curves A2 and C2 could be recoveredto A1 and C1, if the surfactant concentration in theorganic phase is increased (Woll et al., 1987). Thesurfactant concentration limits the amount of proteinthat can be taken by the microemulsion phase (Figure2) (Aires-Barros and Cabral, 1991; Hentsch et al., 1992;Ishikawa et al., 1990). Higher concentrations of surfac-tant facilitate the transfer of proteins into the reversedmicellar, since more of it is available to the formation ofthe protein-surfactant aggregate. In fact, it was ob-served that the solubilization curves are steeper near thepI at higher surfactant concentrations (Woll et al., 1987).3.2. Hydrophobic Interactions. It has been re-
ported that the extraction of apolar amino acids isgoverned by an hydrophobic interaction, with the conse-quent incorporation of these in the interfacial layer ofthe microemulsion droplet (Leodidis and Hatton, 1990a,b;Adachi et al., 1991). It is also likely that proteins withsurface hydrophobic regions exhibit an anomalous be-havior, being extracted to the reversed micellar phaseat unfavorable pH or at high salt concentrations. Thisbehavior has been observed with some proteins (Aires-Barros and Cabral, 1991).Figures 3 and 4 illustrate this situation with a recom-
binant cutinase, produced in Escherichia coli (accumu-lating in the periplasmic space) (Carneiro-da-Cunha etal., 1994a) and recombinant cytochrome b5 producedalso in E. coli (accumulated in the cytoplasm) (Piresand Cabral, 1993). The extraction takes place at pHvalues where the proteins have a global negative charge.For cytochrome b5, a significant yield of extraction at
high ionic strengths (60% at 1 M of NaCl) was observedthat was highly dependent on the temperature (Figure5).From these results it can be deduced that, besides
electrostatic interactions, hydrophobic interactions mightalso play an important role in the uptake of proteins bythe reversed micellar phase. This is not surprising, dueto the important role that this interaction plays in theformation of surfactant aggregates (Israelachvili, 1984).A strong influence on the solubilization behavior ofproteins, particularly with ones that interact with lipidinterfaces like lipases and membrane proteins, shouldalso be expected.
Figure 1. Liquid-liquid extraction curves of low molecularweight proteins (curve A1) and high molecular weight proteins(curve A2) into reversed micelles of anionic surfactants and oflow molecular weight proteins (curve C1) and high molecularweight proteins (curve C2) into reversed micelles of cationicsurfactants.
Figure 2. Effect of AOT concentration on the extraction ofcutinase, 20 °C. Aqueous phase: 50 mM Tris-HCl + 100 mMKCl at pH 9.0.
Figure 3. Effect of pH on the extraction of cutinase with AOT(100 mM)/isooctane reversed micelles (wo ) 20) at 20 °C: (O)cutinase extraction and (b) total activity in micellar phase.
Figure 4. Extraction of recombinant cytochrome b5 intoreversed micelles of CTAB in cyclohexane/10% (v/v) decanol atseveral pH’s and ionic strengths: (O) pH 4.4; (b) pH 5.0; (9)pH 6.0; (0) pH 7.0.
294 Biotechnol. Prog., 1996, Vol. 12, No. 3
4. Driving Forces for the Back-Extraction ofProteins in Reversed Micelles
Upon the extraction, an organic phase containing thedesired protein solubilized in the reversed micelles isobtained. The next step is the recovery and concentrationof the protein in a new aqueous phase. In most casesthe protein can be recovered either as a result of anelectrostatic repulsion of the surfactant, choosing theconvenient pH, or through a size exclusion effect, byincreasing the salt concentration (1-2 M) in the freshaqueous phase (Goklen and Hatton, 1987).However, this is far from being a clear-cut situation.
In several cases, the yield of back-extraction decreaseswith factors that decrease the droplet radius of thereversed micelles, such as an increase in salt concentra-tion or a decrease of temperature. This behavior wasrationalized as the result of an increase in the protein-surfactant interaction as the distance to the interfaciallayer decreases, as does the parameter wo ([H2O]/[sur-factant]) (Bausch et al., 1992).The transfer of proteins into the organic phase seems
also to be in some cases an irreversible process, in thesense that the protein cannot be recovered using experi-mental conditions that are unfavorable to extraction, i.e.,using a pH such that the protein bears the same chargeas the surfactant and high salt concentrations (Woll etal., 1987; Aires-Barros and Cabral, 1991; Pires andCabral, 1993). This is probably the result of a stronghydrophobic interaction of the protein with the surfac-tant. In these situations, the use of a polar alcohol todisrupt this interaction has been proposed (Woll, 1987;Aires-Barros and Cabral, 1991) (Figure 6).The back-extraction of proteins can also be performed
by taking advantage of the phase change properties ofthe reversed micellar system (water in oil microemulsion,wo) (Zulauf and Eicke, 1979; Toprakcioglu et al., 1984;Fletcher et al., 1987; Jada et al., 1990) (Figure 7). Bydecreasing the temperature below Tl (the lower temper-ature of phase transition), a two-phase system will beformed with an upper organic phase containing all of thesurfactant and a lower aqueous phase containing theprotein in the aqueous solution, forming an Winsor IItype system (Winsor, 1984). Above Tu (the upper tem-perature of phase transition), a Winsor I type system isformed, in which the surfactant is solubilized in theaqueous phase. The solubility of an ionic surfactant inthe organic phase decreases with the temperature whilethat of an neutral surfactant increases (Kahlweit et al.,1990).In a recent report (Dekker, 1990) a different approach
was followed to purify an R-amylase. This protein wasextracted into the organic phase containing the cationic
surfactant TOMAC and recovered from it by increasingthe temperature above Tu to achieve a phase separation.Contrary to what would be expected, the surfactantconcentration was constant in the reversed micellarphase even at the highest temperatures used (60 °C). Anoverall recovery of 73% of R-amylase with a concentrationfactor of 2000 was obtained, due to the small volume ofthe aqueous phase formed upon phase change.
5. Interfacial Transport of Proteins into andout of Reversed Micelles
The majority of studies on protein extraction usingreverse micellar systems have focused on the thermody-namic behavior of the microemulsion phases. The equi-librium studies are very important to the optimizationof reversed micellar extraction and the understanding ofthe parameters that affect protein solubilization in thesesystems. However, studies of the kinetics of proteintransfer are also vital for the design of liquid-liquidextraction systems. It is important to have informationabout the rates of forward and backward transfer of theprotein into and out of the reverse micelles across theoil-water interface. Knowledge of these rates is requiredto determine whether interfacial transport is rate limit-ing relative to bulk transport processes. The nature oftransfer of proteins in reversed micelles is at presentunknown, being dependent on different forces (chemical,electrostatic, hydrophobic, and flux dynamics interac-tions) acting between the solute (protein) and the inter-face.Recently, an exhaustive study of the interfacial trans-
port in reversed micellar systems investigated the im-
Figure 5. Temperature effect on the extraction of cytochromeb5 from an aqueous phase at pH 7.0 (0,9) or pH 4.4 (O,b) with0.1 (O,0) or 0.8 M KCl (b,9) to a reversed micellar phase of 100mM CTAB/cyclohexane/10% (v/v) decanol.
Figure 6. Recovery of lipase B as function of alcohol chainlength, C2-C10, using 50 mM K2HPO4 with 50 mM KCl atpH9.0, wo ) 15, phase ratio of 1.0, and 0.43 M of each alcohol: (9)protein recovered and (0) wo values obtained after recovery.
Figure 7. Phase properties of a reversed micellar system withthe temperature: Tu and Tl are the upper and lower tempera-tures of phase transition.
Biotechnol. Prog., 1996, Vol. 12, No. 3 295

portance of the kinetics associated with the forward andbackward extraction by measuring the interfacial masstransfer coefficient and transport rates for two modelproteins, R-chymotrypsin and cytochrome c (Dungan etal., 1991; Bausch et al., 1992). The measurements weremade using a stirred cell technique that allows theimportance of the interfacial resistance to protein trans-fer in these systems to be established. The kinetics ofprotein transfer into the micellar phase were up to 3orders of magnitude (overall mass transfer coefficientsof 10-3 cm/s) faster than rates measured for small solutetransfer (Albery et al., 1987). This indicates that theprotein facilitates its own solubilization process by asignificant influence on the interfacial formation processdue to the large size of proteins. A strong decrease inforward transfer rates with an increase in the pH andionic strength of the aqueous phase was also observed(Dungan et al., 1991).Back-transfer rates were found to be 3 orders of
magnitude (2 × 10-5 cm/s for R-chymotrypsin and 6 ×10-6 cm/s for cytochrome c) slower than those for forwardtransfer, as well as being much lower than water bulktransfer rates. The protein desolubilization rates aredependent on aqueous pH, suggesting that coalescenceof the protein-filled micelle with the bulk interfacedominates the desolubilization kinetics (Dungan et al.,1991).
6. Applications
The selective solubilization of proteins in reversedmicelles aims the development and application of thistechnique to the recovery and purification of proteins inlarge scale. This depends on the ability of this techniqueto extract the protein of interest from the productionmedium without loss of its activity. Significant applica-tions, still at the experimental level, of reversed micellarsystems as a bioseparation technique for recovery andpurification of proteins are discussed below.6.1. Selective Separation of Mixtures of Proteins.
The selective separation of proteins with reverse micelleswas first demonstrated by Goklen and Hatton (1987).These authors used a mixture of pure proteinssribonuclease A, cytochrome c, and lysozyme (Goklen andHatton, 1987).More recently this technique was also demonstrated
on a crude lipolytic preparation from Chromobacteriumviscosum. This mixture contains two lipases of differentmolecular weight and isoelectric point (lipase A with amolecular weight of 120 000 and a pI of 3.7 and lipase Bwhich has a molecular weight of 30 000 and a pI of 7.3).The extraction was carried out using reversed micellarsystems (Camarinha-Vicente et al., 1990; Aires-Barrosand Cabral, 1991). Lipase B was completely solubilizedin AOT/isooctane reversed micellar solution, at pH 6.0and low ionic strength (50 mM KCl), while lipase Aremained in the aqueous solution (Figure 8). At this pH,lipase B is positively charged and is easily solubilized inthe organic phase while lipase A is excluded by sizeexclusion and electrostatic effects. The solubilizationcurve of lipase B in the organic phase containing theanionic surfactant AOT reveals that a significant transferof this protein occurs at pH values higher than its pI,indicating that this process is not only dependent on aelectrostatic driving force.Back-extraction of lipase B from the micellar phase was
only achieved by introducing a water-miscible organicsolvent. The best results were obtained by adding 2.5%(v/v of the total volume of the phases) ethanol to themicellar phase at pH 9.0, using the same ionic strength
as for extraction (Figure 6). Ethanol was used tominimize the hydrophobic interactions between the lipaseand the surfactant and/or organic solvent, allowing itsrecovery into the aqueous phase. A yield of 77% wasobtained for lipase B by analysis of the protein concen-tration in organic phase which revealed lipolytic activity.A FPLC gel filtration chromatogram of the aqueous phaseafter back-extraction confirmed that only lipase B waspresent in the aqueous phase (Camarinha-Vicente et al.,1990). The results of the purification procedure aresummarized in Table 3. Lipase A was purified 4.3-foldand 91% of lipase activity was recovered, and lipase Bwas purified 3.7-fold with a recovery of 76% from thecrude extracted.6.2. Recovery of Enzymes from Fermentation
Broth. This was demonstrated for the first time in apaper reporting the extraction of an alkaline proteasefrom Bacillus sp. from a complex fermentation brothusing an organic solution of AOT in isooctane (Rahamanet al., 1988). The yields obtained in a single extraction/stripping stage are low (on the order of 10-20% of thetotal active species). By using a multistage process theyield of the process was increased, leading to a total yieldof 42% with a purification factor of 2.2. This processcould be repeated as often as necessary to obtain thedesired yield. Maximal purification factors of 6 wereobtained with a yield of 56% if three cycles of extractionwere performed in cascade. The selectivity of the processwas improved as the aqueous/organic volume ratioincreased.More recently Krei and Hustedt (1992) applied also
this technique to the liquid-liquid extraction of anextracellular enzyme, R-amylase produced by fermenta-tion of Bacillus licheniformis. A reversed micellar systemof CTAB (hexadecyltrimethylammonium) in isooctane/hexanol (0.95:0.05) was used for the extraction step. Ina two-step extraction process, a maximum yield of 89%R-amylase activity was achieved, for pH values between8.8 and 10, and a purification factor of 8.9. Comparingthese results with data obtained using a syntheticaqueous phase (0.1 M ionic strength and pH 8.8) showsthat the simplified model system is well suitable todescribe the partitioning behavior of enzymes in WinsorII systems, where the reversed micellar system is inequilibrium with a real fermentation medium (Krei andHustedt, 1992). Medium components like biomass, otherproteins, extracellular metabolites or substrates do notaffect the enzyme partition in the aqueous fermentationbroth/reversed micellar organic biphasic system. How-
Figure 8. Effect of pH on the solubilization of lipase B fromC. viscosum in AOT (250 mM)/isooctane reversed micelles atwo ) 20: (0) lipase B extracted; (9) specific activity of lipase Bin the micellar phase.
296 Biotechnol. Prog., 1996, Vol. 12, No. 3

ever, contrary to the extraction of an alkaline proteasewith an anionic surfactant (AOT), with a cationic sur-factant, the biomass does not remain in the aqueousphase but aggregates at the phase boundary. Thisdrawback could be avoided by separating previously thebiomass from the fermentation medium, before theextraction step.6.3. Recovery of Intracellular Enzymes. The
separation and purification of intracellular enzymes wasalso tested. A reversed system of 0.2 M CTAB in octane/hexanol was used to liberate intracellular enzymesdirectly into the reversed micelles (Giovenco et al., 1987).The CTAB acts as a surfactant to desintegrate thebacterial cells and at the same time to solubilize theproteins which were liberated in the hexanol/octanesolution. Three dehydrogenases (â-hydroxybutyrate de-hydrogenase, isocitrate dehydrogenase, and glucose 6-phos-phate dehydrogenase) from Actobacter vinelandii werechosen as model systems. Glucose 6-phosphate dehy-drogenase did not solubilize in the reversed micelles,probably because of size exclusion (MW ) 200 000 Da).The selective back-transfer into a second aqueous phasewas performed. Purification factors ranging from 2.8 to7.6, depending of the water concentration in the organicphase, were achieved, indicating that recovery of intra-cellular enzymes using reversed micelles may provide analternative method to time-consuming conventional meth-ods that involve several purification steps.6.4. Continuous Extraction of r-Amylase. The
application of reversed micelles for continuous extractionof enzymes has been already demonstrated (Dekker etal., 1986). A double-extraction process for the continuousextraction of R-amylase using a reversed micellar systemof TOMAC in isooctane was developed. The extractionwas performed in two mixer-settler units, the reversedmicellar phase being recirculated through the extractionunits. The efficiency of the process dropped after threecirculation cycles, and it was necessary to add moresurfactant to the organic phase. During the extraction,surfactant was lost (<15% per circulation) and ac-cumulated in the interface together with denaturatedprotein (the mass balance for R-amylase shows a defi-ciency of 10-20%). R-Amylase was recovered with aconcentration factor of 8 and a recovery yield of 70%.6.5. Liquid-Liquid Extraction of Recombinant
Proteins. Two model recombinant proteins, cytochromeb5 and a cutinase from Fusarium solani pisi, were usedto exemplify the application of reversed micellar systemsto the recovery of these proteins and to study the differentparameters which affect the extraction and back-extrac-tion steps. These two recombinant proteins are two goodcase studies, since they are easily produced by fermenta-tion, their tertiary structure is known, and severalmutants obtained by site-directed mutagenesis are avail-able.6.5.1. Cytochrome b5. Cytochrome b5 from rat was
cloned into the plasmid pUC 13 and expressed in E. coliTB1 (von Bodman et al., 1986). Cytochrome b5 has amolecular weight of 13 600 Da and an isoelectric pointof 4.4. This recombinant protein was extracted into areversed micellar phase of a cationic surfactant, CTABin cyclohexane/decanol, and back-extracted to a fresh
aqueous solution (Pires and Cabral, 1993). Dependingon the experimental conditions (pH, ionic strength, andtemperature), the extraction/back-extraction steps arecontrolled by either hydrophobic or electrostatic interac-tions.The extraction of cytochrome b5 from an aqueous
solution to a reverse micellar phase of 100 mM CTAB incyclohexane/10% (v/v) decanol depends of three mainparameters: ionic strength, pH, and temperature. Bymanipulating these three parameters, it was possible toobserve a shift in the type of forces responsible for thesolubilization of cytochrome b5 in the micelle phase.Figure 4 shows the effect of pH and ionic strength on
protein extraction. At pH far above the pI (pH 6 and 7)the extraction is mainly governed by an electrostaticinteraction with the positive surfactant, since at highionic strength, the extraction was reduced to zero, dueto the screening of the electrostatic interaction of thenegative protein and the positive surfactant. At highionic strength this interaction is damped and the extrac-tion does not occur (Pires and Cabral, 1993). At pH closeto the pI the extraction seems to be the result ofhydrophobic interaction between the protein and thesurfactant, as above 0.4 M KCl, it is independent of theionic strength of the aqueous phase. The initial decreasein the extraction can be attributed to the dampening ofelectrostatic interactions that still exist at low ionicstrength.Since hydrophobic forces are strongly dependent on
temperature, the extraction of cytochrome b5 was testedat several temperatures, between 10 and 40 °C, at lowand high ionic strength, and at pH ) pI and a higher pH(Figure 5). As it can be seen the extraction is onlydependent on the temperature when the pH of theaqueous phase is close to the isoelectric point. As thetemperature is lowered, the extraction of cytochrome b5decreases. This is to be expected if the driving force forthe extraction, at a pH close to the pI, is a hydrophobicinteraction of the protein with the surfactant, as theresults shown in Figure 5 imply. The hydrophobicinteraction between an acidic protein such as cytochromeb5 and a cationic surfactant, CTAB, may be explainedby the existence of an hydrophobic region at the cyto-chrome surface that can interact with the micelle ag-
Table 3. Purification of Lipases from C. viscosum Using Reversed Micelles (Camarinha-Vicente et al., 1990)
purification stepstotal protein(mg/mL)
total activity(µmol/mL‚min)
specific activity(µmol/mg‚min)
purificationfactor
recovery yield(%)
crude extract (lipases A+B) 1.41 21.7 15.4 1.0 100aqueous phase after extraction-lipase A 0.299 19.8 66.2 4.3 91aqueous phase after back-extraction-lipase B 0.290 16.6 57.2 3.7 76
Figure 9. Liquid-liquid extraction of native and modifiedcytochromes b5 to an anionic reversed micellar phase: nativecytochrome b5 (O), E44k (0), E56K (b), and E92 (2).
Biotechnol. Prog., 1996, Vol. 12, No. 3 297

gregate under conditions of pH close to the pI and highionic strength (Pires and Cabral, 1993).Contacting the organic phase loaded with cytochrome
b5 with a new aqueous phase at several ionic strengths(up to 2 M salt) and using low temperatures to decreasethe hydrophobic interactions between the protein and themicelle aggregate did not lead to any transfer of theprotein to the fresh aqueous phase. The cytochromestayed in the organic phase, and most of the CTAB (70%)accumulated at the interface. However, by addition ofan alcohol to the biphasic system, part of the cytochromewas back-extracted to the aqueous phase (Aires-Barrosand Cabral, 1991; Pires and Cabral, 1993). This effectwas dependent on the type and concentration of alcohol,increasing with alcohols of short chain length. The addedalcohol also caused the solubilization of the CTAB in theaqueous phase, along with the back-extraction of thecytochrome, and as more surfactant is solubilized in theaqueous phase, more cytochrome is transferred. Thissuggests that the back-extracted cytochrome is stillassociated with the surfactant. It is possible that theback-extraction of the protein is due to a decrease of thedielectric constant of the aqueous medium caused by aresidual presence of alcohol in the aqueous phase. Theremaining protein in the organic phase is probably stillencapsulated inside reverse micelles, or associated withthe surfactant, but in a denatured form, since thespectrum of microencapsulated cytochrome b5 in thevisible wavelength range was markedly different fromthe native protein.It was observed that the back-extraction of cytochrome
b5 in the presence of 20% (v/v) hexanol was also depend-ent on pH and ionic strength (Pires and Cabral, 1993).It was only possible to back-extracted cytochrome b5 byaddition of an alcohol to disrupt the hydrophobic interac-tions, provided that the ionic strength was kept high(higher than 300 mM) and at a pH when the protein andthe surfactant have opposite charges (pH 5 and 7).Apparently the back-extraction process is controlled by
a size-exclusion effect at high ionic strength and by thecapability of the alcohol to disrupt the hydrophobicinteraction between the protein and the alcohol, whichincreases as the pI of the protein is approached. Thisduplicity of behavior could be a consequence of theambivalent character of the cytochrome, since it mayinteract with the surfactant through its apolar sidechains, in the hydrophobic surface region, or with itsioinic side chains, in the remaining surface region (Piresand Cabral, 1993).The mechanism of extraction of cytochrome b5 into a
reversed micellar phase of AOT in octane was studiedusing protein engineering of surface residues (Pires etal., 1994). The extraction behavior of native cytochromeand modified proteins with substitutions of glutamic acidby a lysine residue at positions 44 (E44K), 56 (E56K),and 92 (E92K) at different pH values is shown in Figure9. All the modified proteins have a net charge increaseof +1 to +2, depending on the pH. Since the organicphase contains micelles of an anionic surfactant, it wouldbe expected that an increase in the extraction of themodified cytochromes is due to the increase of the netpositive charge. This increase was indeed observed andwas most evident with the modified protein at position92. These results suggest that the driving force respon-sible for the extraction of the cytochromes is a electro-static interaction of the protein with the micellar aggre-gate and that it is possible to increase it by engineeringthe protein surface, making it more positive (Pires et al.,1994).6.5.2. Cutinase from Fusarium solani pisi. Cuti-
nase from Fusarium solani pisiwas cloned and expressedin recombinant E. coli WK-6 (Lauwereys et al., 1990).This enzyme has a molecular weight of 22 000 Da.The influence of the pH on the extraction of the
cutinase in reversed micelles of AOT (100 mM) inisooctane was studied. The pH of the aqueous phase wasvaried between pH 4.8 and 10.5, using different types ofbuffer systems, according to their pKa value. The ionicstrength of the aqueous phase was constant, the watercontent of the resulting micellar solution determined tobe wo ) 20. The activity and amount of cutinasetransferred toward the reversed micellar phase, as func-tion of the initial pH in the aqueous phase, are given inFigure 3.Extraction of the recombinant cutinase into reversed
micelles of AOT in isooctane was found at pH valuesabove the isoelectric point (pI ) 4.5-5.0), where both theprotein and the micellar interface are negatively charged.(Figure 3) (Carneiro-da-Cunha et al., 1994a). This indi-cated that cutinase is extracted to the reversed micellarphase essentially by hydrophobic interactions betweenthe protein and the apolar tails of the surfactant. Thecutinase could be extracted to the reversed micellar phasewith maximum activity (1.6 µmol/(mL‚min)) in the mi-cellar phase obtained at pH 9 and low ionic strength.The best extraction results were obtained at low ionic
strength (100 mM KCl) which corresponds to a wo of 20(Table 4). Increasing ionic strength led to a decrease ofthe amount of cutinase extracted due to size exclusionas the water pool radius approached the cutinase radius,and when rmicelle ) rcutinase, no extraction was achieved.As cutinase is essentially extracted by hydrophobic
interactions with the apolar tails of the anionic surfac-tant, a strong temperature effect on its solubilizationwould be expected. The hydrophobic forces are stronglydependent on temperature, becoming less important atlower temperatures, which led to a decrease of thecutinase extracted from 100 to 35.6% as the temperatureis lowered from 40 to 4 °C. However, the specificactivities of the cutinase in reversed micelles decreasewith increasing temperature due to partial denaturationof the protein (Carneiro-da-Cunha et al., 1994a).Back-extraction of cutinase from the reversed micellar
phase (100 mM AOT in isooctane, wo ) 20) to a newaqueous phase did not change significatively by increas-ing the ionic strength of the aqueous phase. Instead, theamount of precipitated cutinase at the interface in-creased, and with KCl concentrations lower than 0.5 M,the protein is mainly in the aqueous phase and precipi-tated in the interface. At these KCl concentrations (woe 11), the size of the water pool approached the size ofthe protein leading to its release from the reversedmicellar phase. Increasing the pH of the aqueous phaseled to an enhancement of the cutinase back-extraction,
Table 4. Effect of Ionic Strength of the Aqueous Phaseon Cutinase Extraction (wo) and Radius of the WaterPool (rmicelles) with AOT (100 mM)/Isooctane ReversedMicelles at pH 9.0 (Carneiro-da-Cunha et al., 1994a)
KCl(mM)
proteinextraction
(%) woa
rmicellesb
(Å)
rmicellesrcutinase
100 75.4 20.0 34.0 1.98150 13.6 16.5 29.0 1.69200 13.2 14.2 22.0 1.28300 6.0 12.8 22.0 1.28500 4.3 10.9 21.0 1.221000 0 8.1 15.7 0.91a wo ) [H2O]/[surfactant]. b rmicelles was calculated on the basis
of light scattering data and ultracentrifuge measurements (Eickeand Rehak, 1976). c rcutinase ) 17.2 Å (Martinez et al., 1992).
298 Biotechnol. Prog., 1996, Vol. 12, No. 3

reaching a maximum at pH 9.0 with 50% re-extraction.Addition of an alcohol to the back-extraction step led toan increased of the amount of cutinase precipitated atthe interface (about 80%), and no further increased oncutinase solubilization was achieved (Carneiro-da-Cunhaet al., 1994a).The recovery of cutinase from the reversed micellar
phase (100 mM AOT/isooctane, wo ) 20) into a newaqueous phase increased with the temperature reachinga maximum at 20 °C (Figure 10). This was accompaniedby an increase of the parameter wo from 12.4 to 20.9 withincreasing temperature from 4 to 40 °C. At low temper-atures (4 and 10 °C), the hydrophobic interactionsbetween the protein and the surfactant are reduced,which would facilitate the protein back-extraction. How-ever, at these low temperatures, cutinase solubility inthe aqueous phase decreased, which lead to 40-50%protein precipitation at the interface. At 30 and 40 °C,the protein remained essentially in the reversed micellarphase due to the increase of the hydrophobic interactionsbetween the protein and the surfactant with increasingtemperature. Also at these temperatures protein dena-turation occurred, leading to a lower protein back-extraction.Combination of the optimal conditions for extraction
and back-extraction steps led to protein and activityyields for the whole process of 38 and 45%, respectively.However, the specific activities obtained in the reversedmicellar phase are about 20-30 times lower than theinitial activities in the aqueous phase, suggesting aninhibitory effect of the surfactant. Stability studies alsoshowed a rapid deactivation of the cutinase in AOT/isooctane reversed micelles with half-life times of 1-6 hdepending of the wo used (Sebastiao et al., 1993), whichindicates that the enzyme is probably in contact with themicellar interface and inhibited by the surfactant. Al-though cutinase inhibition occurred in reversed micelles,after back-extraction to a new aqueous phase, 100% ofthe cutinase specific activity was recovered. This wasconfirmed by fluorescence studies of tryptophane andtyrosine residues of the cutinase (Melo et al., 1992).These results led to the possibility of utilizing as extrac-tant of the cutinase the AOT/isooctane reversed micellarphase, besides its toxicity.
7. Liquid-Liquid Extraction of Proteins withReversed Micelles: Strategies of Operation
and Scale-upThe process of recovery of a given protein produced by
fermentation using liquid-liquid extraction with re-
versed micelles is outlined in Figure 11. Two strategiescan be followed. In the usual situation (strategy 1), themethod involves two consecutive steps: (1) the proteinis extracted from the fermentation broth into the reversedmicellar phase through contact of the extract with theorganic solution and (2) the encapsulated protein is back-extracted to a fresh aqueous phase, by contact of theloaded reversed micellar phase with a new aqueoussolution. Alternatively (strategy 2), after the first step,the protein is recovered from the organic phase viainducing a phase change in the reversed micellar systemby decreasing the temperature.Most of the purification schemes of proteins are based
on multistep sequences of chromatographic methods.These procedures are sometimes troublesome and timeconsuming, usually resulting in low final yields. Alter-natively, reversed micellar systems can be used as a firststep of purification procedures because of their simplicityand ease to scale up. Therefore, similar recovery yieldsand purification factors may be achieved with a fewernumber of steps when combining this novel methodologywith chromatographic processes.The use of reversed micelles for the separation of
proteins has the potential to be applied in a continuousmode, at a large-scale operation, as in conventionalsolvent extraction in chemical engineering. Liquid-liquid extraction techniques are easy to scale up, mini-mizing the problems encountered with other separationtechniques when applied on a large scale, namely loss ofresolution capacity, complex equipment design, inabilityto operate in continuous mode, and economic limitations.The main equipment required for liquid-liquid extrac-
tion includes mixer-settler units, centrifugal extractors,and column extractors. The selection of the suitable typeof equipment will depend primarily on the characteristicsof reversed micellar systems. The agitation duringmixing, owing to the presence of surfactants, can causethe emulsification of the reaction mixture. This problemis particularly important with column extractions, where“floating” could be enhanced by the formation of emul-sions. The mixer-settler and the centrifugal contactors
Figure 10. Effect of temperature on (b) cutinase back-extraction and (0) wo, at pH 9.0 and low ionic strength.
Figure 11. Liquid-liquid extraction of proteins with reversedmicelles: M, reversed micellar phase; W, water phase; Tu andTl, upper and lower temperatures of phase transition. W1 isthe aqueous feed, containing the protein to be extracted intoM1. M2 is the collected reversed micellar phase containing theprotein to be back-extracted into W4, through the aqueous feedW3 (strategy 1), or to be collected in its microaqueous phaseW5, through a phase change, induced by a cooling step.
Biotechnol. Prog., 1996, Vol. 12, No. 3 299

seem to be appropriate for solvent extraction withreversed micelles.The studies conducted by Decker et al. (1986, 1987)
for the continuous extraction of R-amylase provide prom-ising results for a continuous-mode operation usingmixer-settler units. The extraction apparatus can bedesigned to provide maximal interface area and concen-tration of the bioproducts (ratio of volumes betweenaqueous and organic phases).A continuous perforated rotating disc contactor was
also used for the extraction of a recombinant cutinasefrom an aqueous solution to a reversed micellar phase ofAOT in isooctane. The cutinase was extracted to theorganic phase with a protein yield of 78% after 70 minof operation (Carneiro-da-Cunha et al., 1994b). Theresults obtained show that this type of equipment canbe successfully used as continuous extraction equipmentfor protein and enzyme extraction using reversed micellarsystems.
8. Conclusions
This review pointed out several aspects important forthe implementation of liquid-liquid extraction of proteinsby reversed micelles as a process in large-scale applica-tions.The equilibrium and kinetics aspects that govern the
solubilization and desolubilization of proteins were ana-lyzed, and it can be concluded that electrostatic andhydrophobic interactions are responsible for the microen-capsulation of protein, depending on the type and con-centration of surfactant used and the surface properties(charge distribution and hydrophobicity) of the proteinsunder study. Interfacial transport processes also affectthe encapsulation of proteins, since back-transfer ratesare slower than forward transfer.This technology has been applied to different proteins
and enzymatic systems, at laboratory scale, includingrecombinant proteins. With these it is possible to havea rational design of the process, as the tertiary structureof the protein used is known and mutatns are available.By selecting the right operational conditions, an improvedextraction process can be obtained.
Literature CitedAdashi, M.; Haada, M.; Shioi, A.; Sato, Y. Extraction of aminoacids to microemulsion. J. Phys. Chem. 1991, 95, 7925-7931.
Aires-Barros, M. R.; Cabral, J. M. S. Selective separation andpurification of two lipases from chromobacterium viscosumusing AOT reversed micelles. Biotechnol. Bioeng. 1991, 38,1302-1307.
Albery, W. J.; Choudhery, R. A.; Atay, N. Z.; Robinson, B. H.Rotating difusion cell studies of microemulsion kinetics. J.Chem. Soc., Faraday Trans. 1 1987, 83, 2407-2419.
Armstrong, D. W.; Li, W. Highly selective protein separationswith reverse micellar liquid membranes. Anal. Chem. 1988,60, 86-88.
Ayala, G. A.; Kamat, S.; Beckmen, E. J.; Russel, A. J. Proteinextraction and activity in reverse micelles of a non ionicdetergent. Biotechnol. Bioeng. 1986, 39, 806-814.
Barlow, D. J.; Thornton, J. M. The distribution of charged groupsin proteins. Biopolymers 1986, 25, 1717-1733.
Bausch, T. E.; Plucinski, P. K.; and Nitsch, W. Kinetics of thereextraction of hydrophilic solutes out of AOT-reversed mi-celles. J. Colloid Interface Sci. 1992, 150, 226-234.
Cabral, J. M. S.; Aires-Barros, M. R. In Recovery Processes forBiological Materials; Kennedy, J. F., Cabral, J. M. S., Eds.;John Wiley & Sons Ltd: Chichester, U.K., 1993; pp 247-271.
Camarinha-Vicente, M. L. C.; Aires-Barros, M. R.; Cabral, J.M. S. Purification of Chromobacterium viscosum Lipasesusing reverse micelles. Biotechnol. Tech. 1990, 4, 137-142.
Carneiro-da-Cunha, M. G.; Cabral, J. M. S.; Aires-Barros, M.R. Studies on the extraction and back-extraction of a recom-binant cutinase in a reversed micellar extraction process.Bioprocess Eng. 1994a, 11, 203-208.
Carneiro-da-Cunha, M. G.; Aires-Barros, M. R.; Tambourgi, E.B.; Cabral, J. M. S. Recovery of a recombinant cutinase withreversed micelles in a continuous perforated rotating disccontactor. Biotechnol. Tech. 1994b, 8, 413-418.
Castro, M. J. M.; Moura, J. J. G.; Cabral, J. M. S. In Advancesin Gene Technology: Protein Engineering and Protein Produc-tion; Brew, K., et al., Eds.; Oxford IRL Press: Oxford, U.K.,1988; pp 136.
Coughlin, R. W.; Baclaski, J. B. Biotechnol. Prog. 1990, 6, 307-309.
Dekker, M. Doctoral Thesis, University of Wageningen, TheNetherlands, 1990.
Dekker, M.; van’t Riet, K.; Weijers, S. R.; Baltussen, J. W. A.;Laane, C.; Bijsterbosh, B. H. Enzyme recovery by liquid-liquidextraction using reversed micelles. Chem. Eng. J. 1986, 33,B27-B33.
Dekker, M.; van’t Riet, K.; Baltussen, J. W. A.; Bijsterbosh, B.H.; Hilhorst, R.; Laane, C. In Proceedings of the 4th EuropeanCongress on Biotechnology; Neijssed, O. M., van der Meer,R. R., Luyben, K.Ch. A. M., Eds.; Amsterdam, 1987; Vol. 2,pp 507-510.
Dekker, M.; Hilhorst, R.; Laane, C. Isolating enzymes byreversed micelles. Anal. Biochem. 1989, 178, 217-226.
Dungan, S. R.; Bausch, T.; Hatton, T. A.; Plucinski, P.; Nitsch,W. Interfacial transport processes in the reversed micellarextraction of proteins. J. Colloid Interface Sci. 1991, 145, 33-50.
Eicke, H.-F.; Rehak, J. On the formation of water/oil-micro-emulsions. Helv. Chim. Acta 1976, 59, 2883-2891.
Fletcher, P. D. I.; Howe, A. M.; Robinson, B. H. The kinetics ofsolubilisate exchange between water droplets of water-in-oilmicroemulsion. J. Chem. Soc., Faraday Trans. 1987, 83,985-1006.
Giovenco, S.; Laane, C.; Hilhorst, R. In Proceedings of the 4thEuropean Congress on Biotechnology; Neijssed, O. M., vander Meer, R. R., Luyben, K.Ch. A. M., Eds.; Amsterdam, 1987;Vol. 2, pp 503-506.
Goklen, K. E. Doctoral Thesis, Massachusetts Institute ofTechnology, MA, 1986.
Goklen, K. E.; Hatton, T. A. Protein extraction using reversemicelles. Biotechnol. Prog. 1985, 1, 69-74.
Goklen, K. E.; Hatton, T. A. Proceedings, ISEC86 Munich, 1986,Vol. III, pp 587-595.
Goklen, K. E.; Hatton, T. A. Liquid liquid extraction of lowmolecular proteins by selective solubilization in reversedmicelles. Sep. Sci. Technol. 1987, 22, 831-841.
Hatton, T. A. In Surfactant-based processes; Scamehorn, J. F.,Horwell, J. H., Eds.; Marcel-Dekker: New York, 1989; Vol.33, pp 55-90.
Hentsch, M.; Menoud, P.; Steiner, L.; Flaschel, E.; Renken, A.Optimization of the surfactant (AOT) concentration in areverse micellar extraction process. Biotechnol. Tech. 1992,6, 359-364.
Huddleston, J.; Veide, A.; Kohler, K.; Flanagan, J.; Enfors, S.-O.; Lyddiatt, A. The molecular basis of partitioning inaqueous two phase systems. TibTech 1991, 9, 381-388.
Ishikawa, H.; Noda, K.; Oka, T. Kinetic Properties of Enzymesin AOT-Isooctane Reversed Micelles. J. Ferment. Bioeng.1990, 6, 381-385.
Israelachvili, J. In Surfactants in solution; Mittal, K. L.,Lindman, B., Eds. Plenum Press: New York, 1984; Vol. 1,pp 3-33.
Jada, A.; Lang, J.; Zana, R. Ternary water in oil microemulsionsmade of cationic surfactants, water and aromatic solvents.1. Water solubility studies. J. Phys. Chem. 1990, 94, 381-387.
Jolivalt, C.; Minier, M.; Reinon, R. Extraction of R-chymotrypsinusing reverse micelles. J. Colloid Interface Sci. 1990, 135,86-96.
Kahlweit, M.; Strey, R.; and Busse, G. Effect of alcohols on thephase behaviour of microemulsions J. Phys. Chem. 1991, 95,5344-5352.
300 Biotechnol. Prog., 1996, Vol. 12, No. 3

Kelley, B. D.; Wang, D. I. C.; Hatton, T. A. Affinity-basedreversed micellar protein extraction: I. Principles andprotein-ligand systems. Biotechnol. Bioeng. 1993, 42, 1199-1208.
Krei, G. A.; Hustedt, H. Extraction of enzymes by reversemicelles. Chem. Eng. Sci. 1992, 47, 99-111.
Kula, M. R.; Kroner, K. H. In Biochemical Engineering; Fietcher,A., Ed.; Springer-Verlag: New York, 1982; Vol. 24, pp 73-118.
Lauwereys, M.; De Geus, P.; De Meutter, J.; Stanssens, P.;Matthyssens, G. In Lipases: Structure,mechanism and geneticengineering; Arberghina, L., Schmid, R. D., Verger, R., Eds.;GBF Monographs: Braunschweig, Germany, 1990; Vol. 16,pp 243-251.
Leodidis, E. B.; Hatton, T. A. Amino Acids in AOT ReversedMicelles. 1. Determination of Interfacial Partition Coef-ficients Using the Phase-Transfer Method. J. Phys. Chem.1990a, 94, 6400-6411.
Leodidis, E. B.; Hatton, T. A. Amino Acids in AOT ReversedMicelles. 2. The Hydrophobic Effect and Hydrogen Bondingas Driving Forces for Interfacial Solubilization. J. Phys.Chem. 1990b, 94, 6411-6420.
Leser, E. M.; Wei, G.; Luisi, P. L.; Maestro, M. Application ofreversed micelles for the extraction of proteins. Biochem.Biophys. Res. Commun. 1986, 135, 629.
Luisi, P. L.; Magid, L. J. Solubilization of Enzymes and NucleicAcids in Hydrocarbon Micellar Solutions. Crit. Rev. Biochem.1986, 20, 409-474.
Luisi, P. L.; Henninger, F.; Joppich, M.; Dossena, A.; Casnati,G. Solubilization and spectroscopic properties of R-chymot-rypsin in cyclohexane. Biochem. Biophys. Res. Commun.1977, 74, 1384-1389.
Luisi, P. L.; Bonner, F. J.; Pellegrini, A.; Wiget, P. E.; Wolf, R.Micellar solubilization of proteins in aprotic solvents and theirspectroscopic characterization. Helv. Chim. Acta 1979, 62,3, 77, 740-752.
Luisi, P. L.; Imre, V. E.; Kaeckle, H.; Pande, H. In Topics inPharmaceutical Sciences; Breiner, D. D., Speiser, P., Eds.;Elsevier: Amsterdam, 1983; pp 243-254.
Marcozzi, G.; Correa, N.; Luisi, P. L.; Caselli, M. Proteinextraction by reverse micelles: a study of the factors affectingthe forward and backward transfer of R-chymotrypsin andits activity. Biotechnol. Bioeng. 1991, 38, 1239-1246.
Martinez, C.; DeGeus, P.; Lauwereys, M.; Matthyssens, G.;Cambillau, C. Fusarium solani cutinase is a lipolytic enzymewith a catalytic serine accessible to solvent. Nature 1992,356, 615-618.
Meier, P.; Imre, E.; Flesher, M.; Luisi, P. L. In Surfactants inSolution; Mital, K. L., Lindam, B., Eds.; Plenum Press: NewYork, 1984; pp 999-1012.
Melo, E. P.; Costa, S. M. B.; Cabral, J. M. S. In Water andreversed micelles of AOT; CEC International Workshop onLipases: Structure, Mechanism and Genetic Engineering;Capri, Italy, October 1992.
Paradkar, V. M.; Dordick, J. S. Purification of glycoproteins byselective transport using concanavalin-mediated reverse mi-cellar extraction. Biotechnol. Prog. 1991, 7, 330-334.
Pires, M. J.; Cabral, J. M. S. Liquid-liquid extraction of arecombinant protein with a reverse micelle phase. Biotechnol.Prog. 1993, 9, 647-650.
Pires, M. J.; Martel, P.; Baptista, A.; Petersen, S. B.; Willson,R.; Cabral, J. M. S. Improving protein extraction in reversedmicellar systems through surface charge engineering. Bio-technol. Bioeng. 1994, 44, 773-780.
Rahaman, R. S.; Chee, J. Y.; Cabral, J. M. S.; Hatton, T. A.Recovery of an extracellular alkaline protease from wholefermentation broth using reverse micelles. Biotechnol. Prog.1988, 4, 217-224.
Sebastiao, M. J.; Cabral, J. M. S.; Aires-Barros, M. R. Synthesisof fatty acid esters by a recombinant cutinase in reversedmicelles. Biotechnol. Bioeng. 1993, 42, 326-332.
Shire, S. J.; Hanania, G. I. H.; Gurd, F. R. N. Electrostatic effectsin Myoglobin. Hydrogen ion equilibria in sperm whaleferrimyoglobin. Biochemistry 1974, 13, 2967-2974.
Tanford, C.; Roxby, R. Interpretation of protein titration curves.Application to lysozyme. Biochemistry 1972, 11, 2192-2198.
Toprakeioglu, C.; Dore, J. C.; Robinson, B. H.; Howe, A. Small-angle neutron scattering study of microemulsions stabilizedby aerosol OT. Part 2sCritical scattering and phase stability.J. Chem. Soc., Faraday Trans. 1984, 80, 413-422.
Van Brakel, J.; Kleizen, H. H. In Chemical engineering problemsin biotechnology; Critical Report on Applied Chemistry;Winkler, A., ed.; Elsevier Science Publishers: New York,1990; Vol. 29, pp 95-166.
van’t Riet, K.; Dekker, M. In Proceedings of the 3rd EuropeanCongress on Biotechnology, Munich, 1984; Vol. 3, p 341.
von Bodman, S. B.; Schuller, M. A.; Jollie, D. R.; Sligar, S. G.Synthesis, bacterial expression, and mutagenesis of the genecoding for mammalian cytochrome b5. Proc. Natl. Acad. Sci.U.S.A. 1986, 83, 9443-9447.
Winsor, P. A. Hydrotopy solubilization and related emulsifica-tion processes. Trans. Faraday Soc. 1984, 54, 376-398.
Wolbert, R. B. G.; Hilhorst, R.; Voskuilen, G.; Nachtegaal, H.;Dekker, M.; van’t Riet, K.; Bijsterbosch, B. H. Protein transferfrom an aqueous phase into reversed micellessthe effect ofprotein size and charge distribution. Eur. J. Biochem. 1989,184, 627-633.
Woll, J. Doctoral Thesis, Massachusetts Institute of Technology,MA, 1987.
Woll, J. M.; Dillon, A. S.; Rahaman, R. S.; Hatton, T. A. InProtein purification: micro to macro; Burgess, R., Ed.; Alan,R. Bliss: New York, 1987; pp 117-130.
Woll, J. M.; Hatton, T. A.; Yarmush, M. L. Bioaffinity separa-tions using reversed micellar extraction. Biotechnol. Prog.1989, 5, 57-62.
Zulauf, M.; Eicke, H. F. Inverted micelles and microemulsionsin the ternary system H2O/Aerosol OT/isooctane as studiedby photon correlation spectroscopy. J. Phys. Chem. 1979, 83,480-486.
Accepted August 4, 1995.X
BP950050L
X Abstract published in Advance ACS Abstracts, February 1,1996.
Biotechnol. Prog., 1996, Vol. 12, No. 3 301
Copyright © 2022 FDOKUMEN




















