
Leishmanolysin-like Molecules in Herpetomonas samuelpessoai Mediate Hydrolysis of Protein Substrates...
15
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
Transcript of Leishmanolysin-like Molecules in Herpetomonas samuelpessoai Mediate Hydrolysis of Protein Substrates...

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Protist, Vol. 161, 589–602, October 2010http://www.elsevier.de/protisPublished online date 2 April 2010
ORIGINAL PAPER
Leishmanolysin-like Molecules in Herpetomonassamuelpessoai Mediate Hydrolysis of ProteinSubstrates and Interaction with Insect
Fernanda M. Pereiraa, Felipe A. Diasa, Camila G.R. Eliasa, Claudia M. d’Avila-Levyb,Cristina S. Silvac, Jacenir R. Santos-Malletc, Marta H. Branquinhaa, and Andre L.S. Santosa,1
aLaboratorio de Estudos Integrados em Bioquımica Microbiana, Departamento de Microbiologia Geral,Instituto de Microbiologia Prof. Paulo de Goes (IMPPG), Bloco E-subsolo, Centro de Ciencias da Saude(CCS), Universidade Federal do Rio de Janeiro (UFRJ), Av. Carlos Chagas Filho, 373, Cidade Universitaria,Rio de Janeiro, RJ 21941-902, Brazil
bLaboratorio de Biologia Molecular e Doenc-as Endemicas, Instituto Oswaldo Cruz (IOC), Fundac- ao OswaldoCruz (FIOCRUZ), Rio de Janeiro, RJ, Brazil
cSetor de Entomologia Medica e Forense, Laboratorio de Transmissores de Leishmanioses, IOC-FIOCRUZ,Rio de Janeiro, RJ, Brazil
Submitted September 3, 2009; Accepted December 22, 2009Monitoring Editor: Larry Simpson
Herpetomonas samuelpessoai, an insect trypanosomatid, produces a 63-kDa metallopeptidase thathas similar biochemical/immunological properties to Leishmania leishmanolysin, a virulence factorthat participates in different stages of the parasite life cycle. Herein, we described some biochemicalcharacteristics of the major surface metallopeptidase of H. samuelpessoai that led us to infer someprobable functions for this peptidase during the parasite-invertebrate interaction. Gelatin-SDS-PAGE,flow cytometry and confocal fluorescence microscopy provided measurements for the relative levelsof surface leishmanolysin-like molecules in H. samuelpessoai. Immunocytochemical analysisdemonstrated the presence of leishmanolysin-like molecules on the surface and cytoplasm of theparasite. The surface metallopeptidase was active at a broad spectrum of pH and temperature,showing maximum activity at pH 6.0 at 37 1C, and an ability to degrade albumin, hemoglobin, IgG,mucin, casein and gut proteins obtained from Aedes aegypti. This wide substrate utilization mightsupport parasite growth and development. Curiously, H. samuelpessoai cells were able to colonizeA. aegypti guts. In an effort to implicate a possible role for the metallopeptidase fromH. samuelpessoai, living parasites were treated with different compounds before the interaction withgut cells. The pre-incubation with metallopeptidase inhibitors, phospholipase C or anti-leishmano-lysin antibodies promoted a significant reduction in the interaction with guts. Similarly, the pre-treatment of gut cells with purified leishmanolysin-like protein drastically diminished the adhesion
1Corresponding author; fax þ55 21 2560 8344e-mails [email protected], [email protected] (A.L.S. Santos).
& 2010 Elsevier Ltd. All rights reserved.doi:10.1016/j.protis.2010.02.001

Author's personal copy
process. Furthermore, the expression of surface leishmanolysin in H. samuelpessoai cells wasdrastically enhanced after passage in A. aegypti. These results suggest the participation ofhomologues of leishmanolysin in the interaction of H. samuelpessoai with the invertebrate vector.& 2010 Elsevier Ltd. All rights reserved.
Key words: Herpetomonas samuelpessoai; peptidases; leishmanolysin; cellular interaction; insect vector;Aedes aegypti.
Introduction
The monoxenous trypanosomatids, which encom-pass species belonging to the genera Leptomonas,Crithidia, Blastocrithidia and Herpetomonas, are lessstudied compared to their pathogenic relatives, suchas the human/animal (Trypanosoma and Leishmania)as well as the plant (Phytomonas) pathogens.Possessing all the molecular mechanisms uniqueto the trypanosomatid cells, such as trans-splicing(Murphy et al. 1986; Sutton and Boothroyd 1986),RNA editing (Shaw et al. 1988; Simpson and Shaw1989) and polycistronic transcription (Muhich andBoothroyd 1988; Tschudi and Ullu 1988), these‘‘non-mammalian, non-pathogenic’’ microorganismshave been used as experimental models of theTrypanosomatidae family for exploring their basicmechanisms at the genetic, biological and biochem-ical levels. In the same way, several research groupshave described common structures/molecules pro-duced by monoxenous and heteroxenous parasitesbelonging to the Trypanosomatidae family (Breganoet al. 2003; Elias et al. 2008; Lopes et al. 1981;Santos et al. 2006, 2007). For instance, our grouphas demonstrated that several insect and phyto-flagellate trypanosomatids are able to produce bothsurface and released leishmanolysin-like molecules(reviewed by Santos et al. 2006).
Leishmanolysin surface peptidase is a well-known virulence factor that participates in differ-ent stages of the Leishmania-mammalian hostrelationship, including (i) degradation of severalkey human proteinaceous structures (e.g., extra-cellular matrix proteins, complement components,albumin, immunoglobulins, hemoglobin, hostcytosolic MARCKS-related proteins that are majorprotein kinase C substrates expressed in diversecell types, surface CD4 antigen and intracellularpeptides presented by MHC class I molecules), (ii)adhesion and internalization of promastigotesthrough mammalian macrophage receptors (e.g.,CR3, Mac-1 (CD11b-Cd18), and fibronectinreceptors), (iii) survival inside the inhospitableconditions of the phagolysosome, and (iv) preven-tion of the antimicrobial peptide-mediatedapoptotic killing (Kulkarni et al. 2006, 2008;
reviewed by Yao et al. 2003). On the other hand,the real relevance of leishmanolysin molecules inthe Leishmania-insect vector is a controversialissue (Hajmova et al. 2004; Joshi et al. 2002;reviewed by Santos et al. 2006). Comparisonsbetween heteroxenous and monoxenous flagel-late trypanosomatids identify similar structuresthat have evolved in response to similar chal-lenges, such as colonization of the invertebratehosts. In this sense, our group has demonstratedthat leishmanolysin-like proteins from insect andphytoflagellate trypanosomatids can function asadhesive molecules, promoting the connection oflive parasites to the invertebrate host epithelialcells (reviewed by Santos et al. 2006).
Our prior publication reported that promastigotesof Herpetomonas samuelpessoai are able to releasea 63 kDa metallopeptidase that has similar biochem-ical and immunological properties to the leishmano-lysin of Leishmania (Elias et al. 2006; Santos et al.2003). H. samuelpessoai is an insect flagellateextensively used as a model for physiological,biochemical and ultrastructural aspects of trypano-somatids. In the present paper, we have aimed toinfer possible roles for the leishmanolysin-like mole-cule expressed on the surface of H. samuelpessoaicells. Experimentally, we have identified the metall-opeptidase activity as well as the leishmanolysi-n-related protein on the parasite surface and thenstudied the ability of this molecule to hydrolyzeseveral protein compounds as well as its influenceon the parasite-invertebrate host interaction.
Results and Discussion
H. samuelpessoai Expresses SurfaceLeishmanolysin-like Molecules
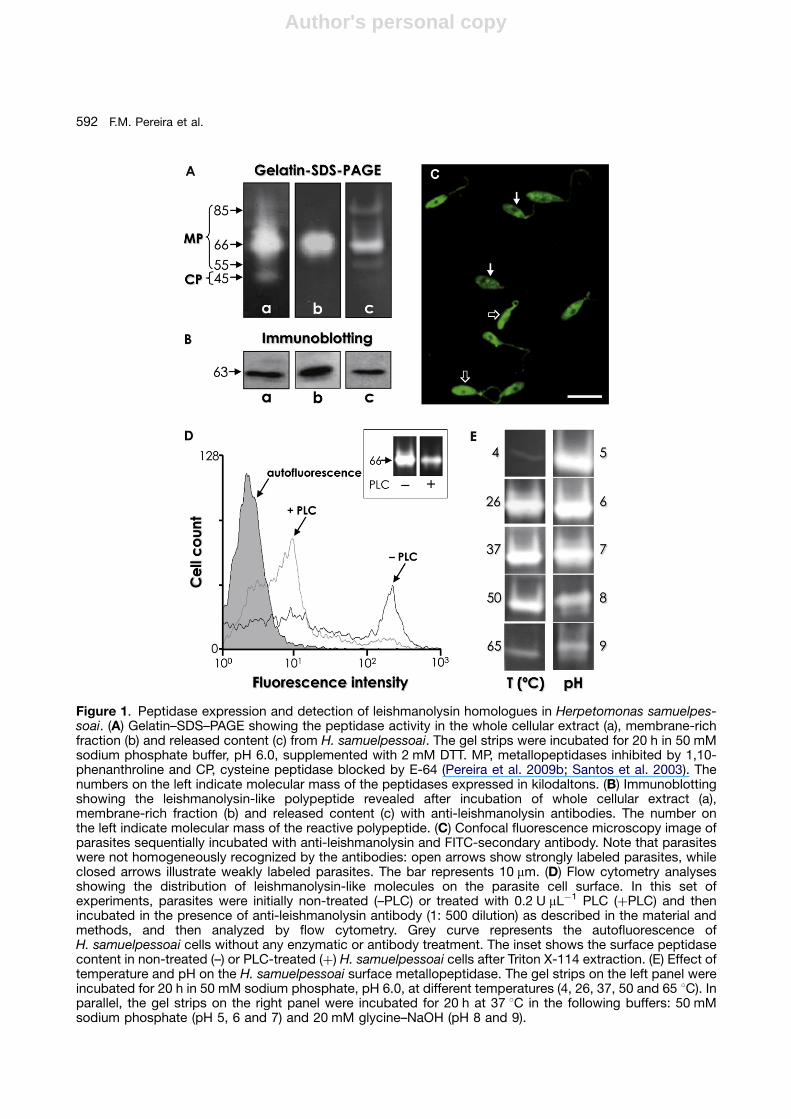
H. samuelpessoai promastigotes produced twomajor peptidase classes when analyzed by gelatin-sodium dodecyl sulfate-polyacrylamide gel electro-phoresis (SDS-PAGE): a prominent metallopeptidaseof 66 kDa (actually a broad hydrolytic activity ranging
F.M. Pereira et al.590

Author's personal copy
from 80 to 60 kDa), inhibited by 10 mM 1,10-phenanthroline, and a minor cysteine peptidaseactivity of 45 kDa, restrained by 1 mM trans-epox-ysuccinyl L-leucylamido-(4-guanidino) butane (E-64)(Fig. 1A, lane a). The 66 kDa metallopeptidase activitywas detected in the parasite membrane fraction afterTriton X-114 partition (Fig. 1A, lane b) as well as in theextracellular milieu as the major secreted peptidasecomponent (Fig. 1A, lane c). Leishmanolysin-likemolecules produced by H. samuelpessoai cellswere detected through immunoblotting using ananti-leishmanolysin raised against L. mexicana(Fig. 1B). Confocal fluorescence microscopyshowed that the leishmanolysin-like molecules weredisposed all over the parasite cellular body, except inthe nuclear region, and throughout the flagellum(Fig. 1C). Interestingly, the labeling was nothomogeneous, since some parasite cells weremore fluorescent than others (Fig. 1C). Flowcytometry analysis corroborated the heterogeneousexpression of surface leishmanolysin-like molecules(Fig. 1D). A first explanation for these observationswould be that the lack of equal expression iscorrelated to the H. samuelpessoai growth phase,since trypanosomatid cultures were notsynchronized. Furthermore, the occurrence ofdistinct subpopulations could alternatively denote adifferent expression of surface leishmanolysin or evena diminished accessibility to external ligands in thesecell subsets, as proposed for other glycoconjugatesin H. samuelpessoai (Santos et al. 2002). Theoccurrence of distinct subpopulations withdifferential expression of surface molecules is welldocumented in trypanosomatids (Matta et al. 1999;Santos et al. 2002; Silva et al. 2009), including surfaceleishmanolysin (d’Avila-Levy et al. 2006a; Eliaset al. 2006; Pereira et al. 2009a). In Leishmaniaspp., leishmanolysin gene products may be localizeddifferently in the cell, and these different isoformsare retained or secreted, suggesting that the minorsequence differences between leishmanolysinisoforms may have functional consequences.It is possible that sequence differences betweenmembers of these gene families could cause them tobe expressed in different parasite compart-ments and/or at different steps of the parasite lifecycle (Yao et al. 2005). In this context, leishmanolysinisoforms are differentially expressed during thepromastigote growth from logarithmic to stationaryphase, and between wild type and attenuatedpromastigotes (Lima et al. 2009; Sadlova et al.2006). In contrast, the expression of leishmanolysin-like molecules was not modulated during eithertemperature-or dimethylsulfoxide-elicited differentia-tion in H. samuelpessoai (Pereira et al. 2009b).
Phospholipase C (PLC)-treated H. samuelpes-soai cells showed a significant reduction in boththe surface binding of anti-leishmanolysin anti-bodies (Fig. 1D) and in the 66 kDa metallopepti-dase activity (Fig. 1D, inset) as compared to thenon-treated flagellates, substantiating that thesesurface molecules were glycosylphosphatidylino-sitol (GPI)-anchored, a feature shared with theleishmanolysin of Leishmania spp. and other lowertrypanosomatids like Herpetomonas megaseliae(Nogueira de Melo et al. 2006), Crithidia deanei(d’Avila-Levy et al. 2008), Crithidia fasciculata(Inverso et al. 1993), Blastocrithidia culicis(d’Avila-Levy et al. 2005) and Phytomonas serpens(d’Avila-Levy et al. 2006b). Interestingly, the zinc-metallopeptidase inhibitor 1,10-phenanthrolinewas able to restrain the secretion of the 66 kDametallopeptidase in a dose-dependent manner,while the PLC inhibitor p-chloromercuriphenylsul-fonic acid did not alter the secretion pattern inH. samuelpessoai (Elias et al. 2006). In addition,anti-cross-reacting determinant antibody failedto recognize any secreted polypeptide fromH. samuelpessoai cells (Elias et al. 2006). Collec-tively, these results suggest that the leishmanoly-sin-like molecule was released from theH. samuelpessoai surface by proteolysis insteadof phospholipolysis, in a similar mechanism to thatobserved in Leishmania cells (reviewed by Yaoet al. 2003).
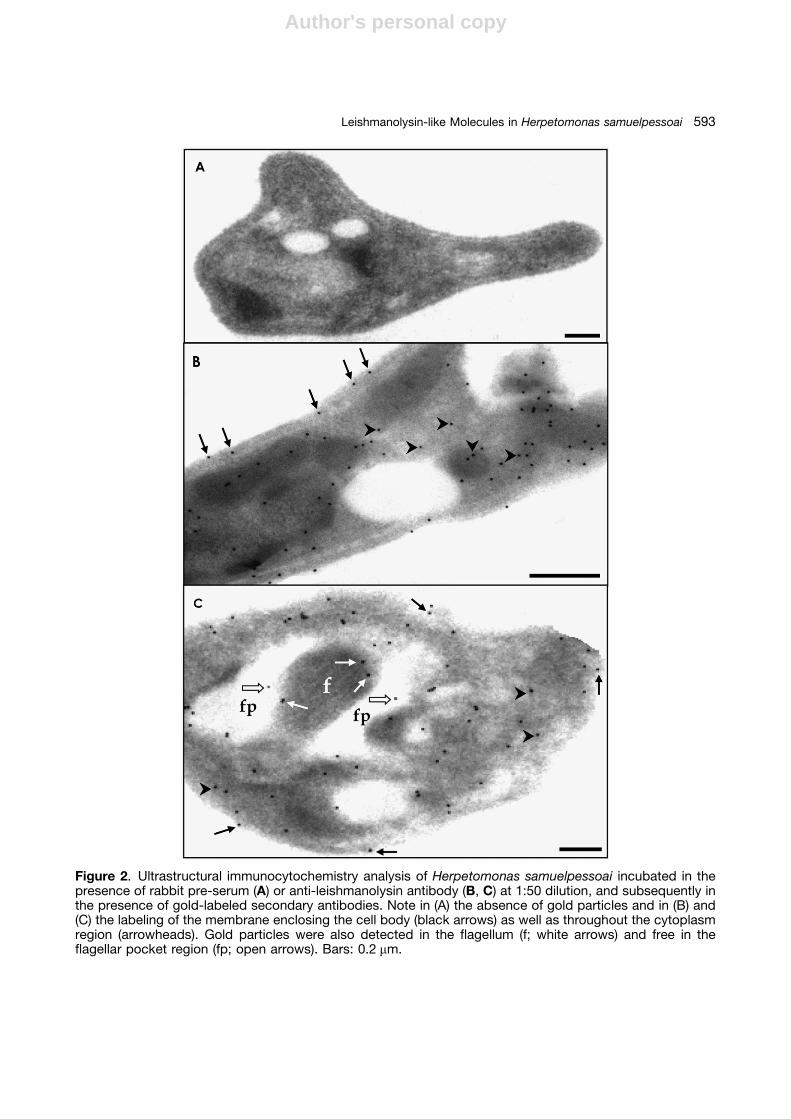
Taken together, our results suggest that leishma-nolysin-like molecules are distributed in differentcellular compartments of H. samuelpessoai, includ-ing cytoplasmic and membrane regions. In addi-tion, the cellular distribution of the leishmanolysin-like molecules on H. samuelpessoai cells could beinvestigated through immunocytochemistry analy-sis (Fig. 2). Gold particles could be clearly observedin all cellular compartments, including cytoplasm,membrane lining the cell body and flagellum,and flagellar pocket (Fig. 2B). Interestingly, somegold particles could be visualized free in theflagellar pocket region (Fig. 2C), suggesting therelease of the leishmanolysin-related molecules tothe extracellular milieu. It is important to note thatthe trypanosomatid flagellar pocket is a highlyspecialized area of the parasite membrane that isinvolved in several processes, including proteinsecretion to the external environment (reviewed byMcConville et al. 2002).
The pH dependence of the 66 kDa surfacemetallopeptidase of H. samuelpessoai was alsodetermined by overlay gels (Fig. 1E). On the gelatinsubstrate, the metallo-type enzyme was active ata broad spectrum of pH and temperature, showing
591Leishmanolysin-like Molecules in Herpetomonas samuelpessoai
Author's personal copy
Figure 1. Peptidase expression and detection of leishmanolysin homologues in Herpetomonas samuelpes-soai. (A) Gelatin–SDS–PAGE showing the peptidase activity in the whole cellular extract (a), membrane-richfraction (b) and released content (c) from H. samuelpessoai. The gel strips were incubated for 20 h in 50 mMsodium phosphate buffer, pH 6.0, supplemented with 2 mM DTT. MP, metallopeptidases inhibited by 1,10-phenanthroline and CP, cysteine peptidase blocked by E-64 (Pereira et al. 2009b; Santos et al. 2003). Thenumbers on the left indicate molecular mass of the peptidases expressed in kilodaltons. (B) Immunoblottingshowing the leishmanolysin-like polypeptide revealed after incubation of whole cellular extract (a),membrane-rich fraction (b) and released content (c) with anti-leishmanolysin antibodies. The number onthe left indicate molecular mass of the reactive polypeptide. (C) Confocal fluorescence microscopy image ofparasites sequentially incubated with anti-leishmanolysin and FITC-secondary antibody. Note that parasiteswere not homogeneously recognized by the antibodies: open arrows show strongly labeled parasites, whileclosed arrows illustrate weakly labeled parasites. The bar represents 10 mm. (D) Flow cytometry analysesshowing the distribution of leishmanolysin-like molecules on the parasite cell surface. In this set ofexperiments, parasites were initially non-treated (–PLC) or treated with 0.2 U mL�1 PLC (þPLC) and thenincubated in the presence of anti-leishmanolysin antibody (1: 500 dilution) as described in the material andmethods, and then analyzed by flow cytometry. Grey curve represents the autofluorescence ofH. samuelpessoai cells without any enzymatic or antibody treatment. The inset shows the surface peptidasecontent in non-treated (–) or PLC-treated (þ) H. samuelpessoai cells after Triton X-114 extraction. (E) Effect oftemperature and pH on the H. samuelpessoai surface metallopeptidase. The gel strips on the left panel wereincubated for 20 h in 50 mM sodium phosphate, pH 6.0, at different temperatures (4, 26, 37, 50 and 65 1C). Inparallel, the gel strips on the right panel were incubated for 20 h at 37 1C in the following buffers: 50 mMsodium phosphate (pH 5, 6 and 7) and 20 mM glycine–NaOH (pH 8 and 9).
F.M. Pereira et al.592
Author's personal copy
Figure 2. Ultrastructural immunocytochemistry analysis of Herpetomonas samuelpessoai incubated in thepresence of rabbit pre-serum (A) or anti-leishmanolysin antibody (B, C) at 1:50 dilution, and subsequently inthe presence of gold-labeled secondary antibodies. Note in (A) the absence of gold particles and in (B) and(C) the labeling of the membrane enclosing the cell body (black arrows) as well as throughout the cytoplasmregion (arrowheads). Gold particles were also detected in the flagellum (f; white arrows) and free in theflagellar pocket region (fp; open arrows). Bars: 0.2 mm.
593Leishmanolysin-like Molecules in Herpetomonas samuelpessoai
Author's personal copy
maximum hydrolytic activity at pH 6.0 at 37 1C(Fig. 1E). These large spectra of pH and tempera-ture in which the 66 kDa metallopeptidase is ableto be catalytically active could be used presum-ably to retain maximum flexibility for the trypano-somatid to survive under different environmentalconditions. Similar biochemical properties werealso observed for the extracellular leishmanolysin-like molecule from C. deanei (d’Avila-Levy et al.2003), C. guilhermei (Nogueira de Melo et al. 2001)and H. samuelpessoai (Elias et al. 2006); however,the corresponding molecule secreted by Phyto-monas franc-ai cells was more active at pH 5.0 at26 1C (Almeida et al. 2003).
Cleavage of Different Protein Substratesby the Surface Metallopeptidase Producedby H. samuelpessoai
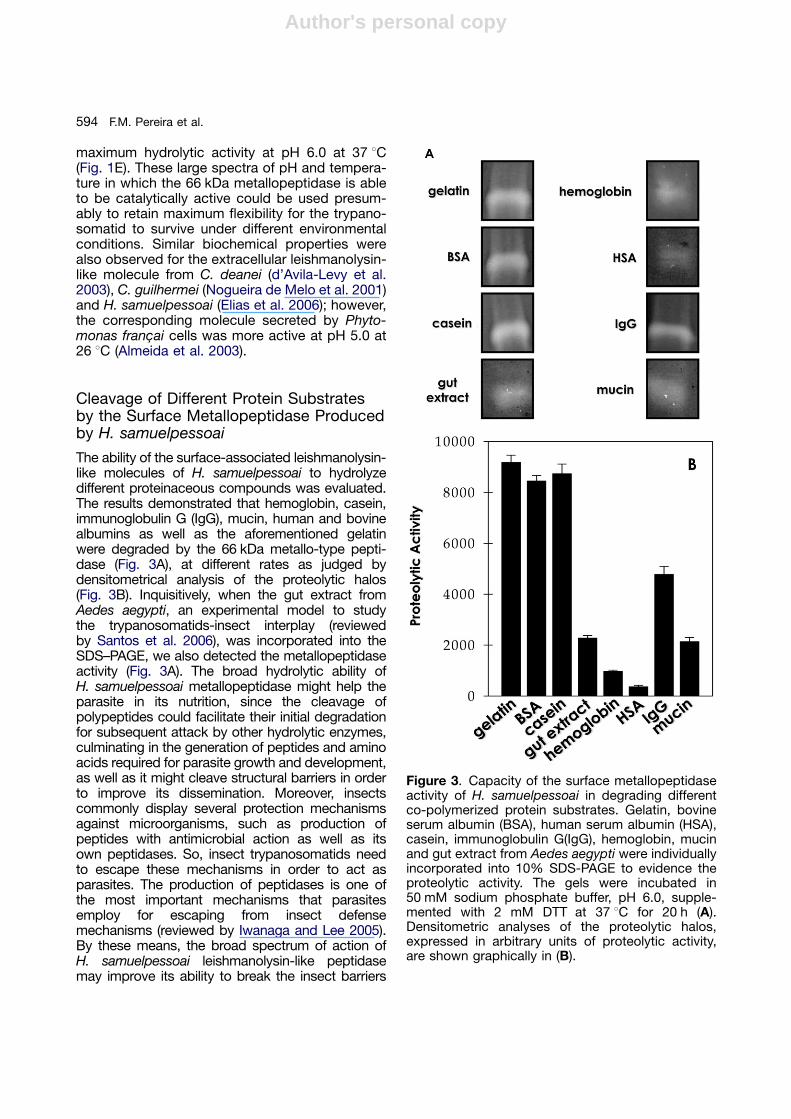
The ability of the surface-associated leishmanolysin-like molecules of H. samuelpessoai to hydrolyzedifferent proteinaceous compounds was evaluated.The results demonstrated that hemoglobin, casein,immunoglobulin G (IgG), mucin, human and bovinealbumins as well as the aforementioned gelatinwere degraded by the 66 kDa metallo-type pepti-dase (Fig. 3A), at different rates as judged bydensitometrical analysis of the proteolytic halos(Fig. 3B). Inquisitively, when the gut extract fromAedes aegypti, an experimental model to studythe trypanosomatids-insect interplay (reviewedby Santos et al. 2006), was incorporated into theSDS–PAGE, we also detected the metallopeptidaseactivity (Fig. 3A). The broad hydrolytic ability ofH. samuelpessoai metallopeptidase might help theparasite in its nutrition, since the cleavage ofpolypeptides could facilitate their initial degradationfor subsequent attack by other hydrolytic enzymes,culminating in the generation of peptides and aminoacids required for parasite growth and development,as well as it might cleave structural barriers in orderto improve its dissemination. Moreover, insectscommonly display several protection mechanismsagainst microorganisms, such as production ofpeptides with antimicrobial action as well as itsown peptidases. So, insect trypanosomatids needto escape these mechanisms in order to act asparasites. The production of peptidases is one ofthe most important mechanisms that parasitesemploy for escaping from insect defensemechanisms (reviewed by Iwanaga and Lee 2005).By these means, the broad spectrum of action ofH. samuelpessoai leishmanolysin-like peptidasemay improve its ability to break the insect barriers
Figure 3. Capacity of the surface metallopeptidaseactivity of H. samuelpessoai in degrading differentco-polymerized protein substrates. Gelatin, bovineserum albumin (BSA), human serum albumin (HSA),casein, immunoglobulin G(IgG), hemoglobin, mucinand gut extract from Aedes aegypti were individuallyincorporated into 10% SDS-PAGE to evidence theproteolytic activity. The gels were incubated in50 mM sodium phosphate buffer, pH 6.0, supple-mented with 2 mM DTT at 37 1C for 20 h (A).Densitometric analyses of the proteolytic halos,expressed in arbitrary units of proteolytic activity,are shown graphically in (B).
F.M. Pereira et al.594
Author's personal copy
of defense. Similarly, leishmanolysin moleculesproduced by Leishmania are able to hydrolyzeseveral relevant host protein structures andantimicrobial peptides (Kulkarni et al. 2008;McGwire et al. 2003).
Colonization of A. aegypti Gut byH. samuelpessoai Cells
The vectorial competence of an insect for differentspecies of trypanosomatids is an important ques-tion and it is well-known that this fitness iscontrolled by structural polymorphism of severalsurface molecules (Pimenta et al. 1994). In thissense, we have investigated whether A. aegypti isable to permit the colonization by livingH. samuelpessoai cells. Following feeding for24 h, sand flies were dissected and the numberof parasites associated to the guts was deter-mined as previously described (Correa-da-Silvaet al. 2006). One day after feeding, H. samuelpes-soai cells could be observed inside the gut ofA. aegypti (Fig. 4). From the second day onwards,the number of parasites diminished in relation tothe first day, but the parasites could be detectedup to 8 days after feeding (Fig. 4). This reductionprobably reflects immune mechanisms elicited inthe insect that partially controlled growth of theprotozoa. Microscopic examination of theexplanted and longitudinally opened gut showedthe parasite binding to the insect gut lumen (datanot shown). In vivo and in vitro binding assaysrevealed that H. samuelpessoai cells interactedwith the gut epithelial cells via flagellum and cell
body (data not shown). Similar results werepreviously reported for the adhesion of B. culicis,C. deanei, C. desouzai, C. oncopelti andH. roitmani with different insect cell lineages(C6/36 from Aedes albopictus, MOS-55 fromAnopheles gambiae and LL-5 from Lutzomyialongipalpis) or explanted insect guts (Fampaet al. 2003). These findings together withprevious similar data from Blastocrithidia culicis(Correa-da-Silva et al. 2006; Fampa et al. 2003)reinforce the idea that monoxenic trypanosomatids present low host specificity (Podlipaev2001; Podlipaev et al. 2004; Wallace 1966). Theinteraction of monoxenous trypanosomatidswith bloodsucking insects is not a novelty.These flagellates have been reported onTriatoma infestans, Aedes vexans, Anophelesquadrimaculatus and Culex pipiens (Cerisolaet al. 1971; Mezincescu 1908; Noguchi andTilden 1926; Novy et al. 1907; Wallace andJohnson 1961). These data are of particularrelevance especially in the light that monoxenoustrypanosomatids can be responsible forleishmaniosis-like symptoms (Chicharro andAlvar 2003). Indeed, Herpetomonas sp. is foundrepeatedly in A. aegypti (Weinman and Cheon1978) and a flagellated protozoan closely relatedto H. samuelpessoai was recently identified inthe blood of a HIV-infected patient (Morio et al.2008). Therefore, the occurrence of monoxenoustrypanosomatids in bloodsucking insects togetherwith repeated reports of their occurrence inhumans supports the view that monoxenoustrypanosomatids are phylogentically ancestralin relation to the heteroxenic ones (Simpsonet al. 2006).
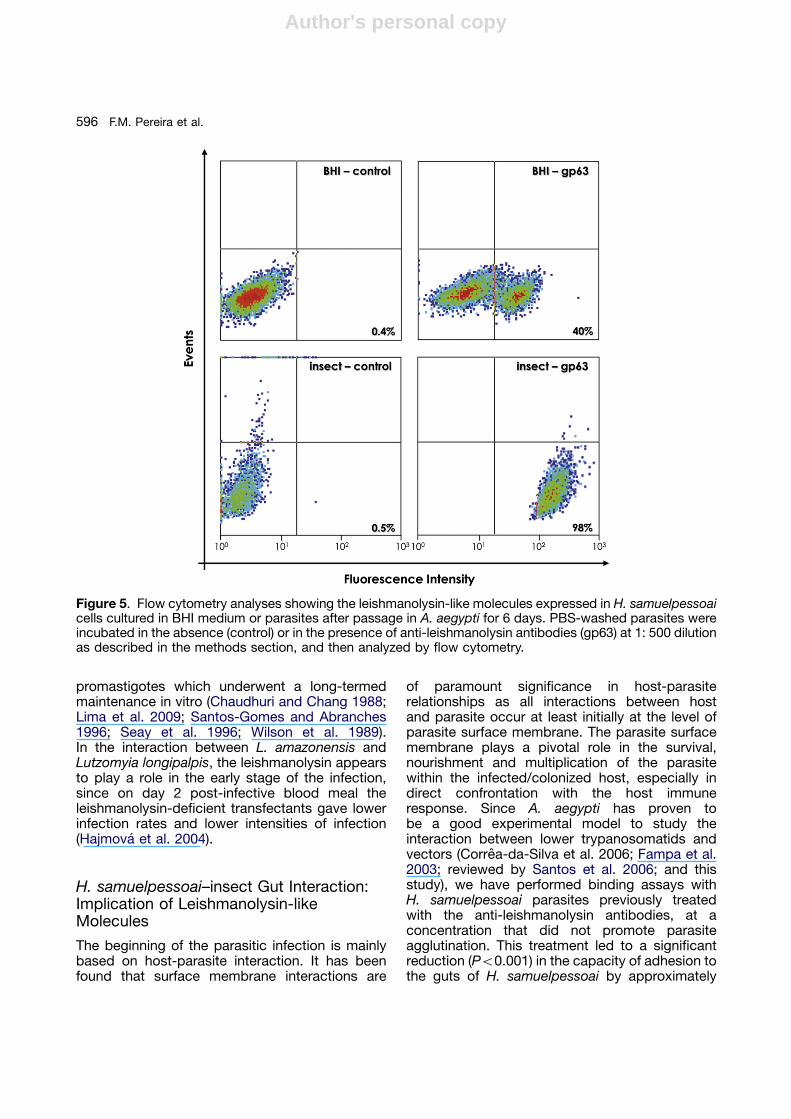
Modulation in the Leishmanolysin-likeMolecules of H. samuelpessoai afterPassage in vivo in A. aegypti
H. samuelpessoai passaged through A. aegyptisand flies expressed 10-fold as much leishmano-lysin-like molecules compared with parasitescultured in BHI medium (Fig. 5). Sadlova et al.(2006) demonstrated that an attenuated strainof L. major (LV561/AV line) produced low amountand low enzymatic activity of leishmanolysin;however, after serial passages of this avirulentstrain through either Phlebotomus duboscqi orthrough mice led to a recovery of leishmanolysinproteolytical activity to the level present in virulentL. major LV561/V line. Reduced leishmanolysinlevel is a frequent phenomenon in Leishmania
Figure 4. Outcome of infections in Aedes aegyptifed on H. samuelpessoai. Histograms represent thenumber of protozoa in mosquito guts (mean7SEM)of three experiments in which 10–20 insects wereexamined per time-point.
595Leishmanolysin-like Molecules in Herpetomonas samuelpessoai
Author's personal copy
promastigotes which underwent a long-termedmaintenance in vitro (Chaudhuri and Chang 1988;Lima et al. 2009; Santos-Gomes and Abranches1996; Seay et al. 1996; Wilson et al. 1989).In the interaction between L. amazonensis andLutzomyia longipalpis, the leishmanolysin appearsto play a role in the early stage of the infection,since on day 2 post-infective blood meal theleishmanolysin-deficient transfectants gave lowerinfection rates and lower intensities of infection(Hajmova et al. 2004).
H. samuelpessoai–insect Gut Interaction:Implication of Leishmanolysin-likeMolecules
The beginning of the parasitic infection is mainlybased on host-parasite interaction. It has beenfound that surface membrane interactions are
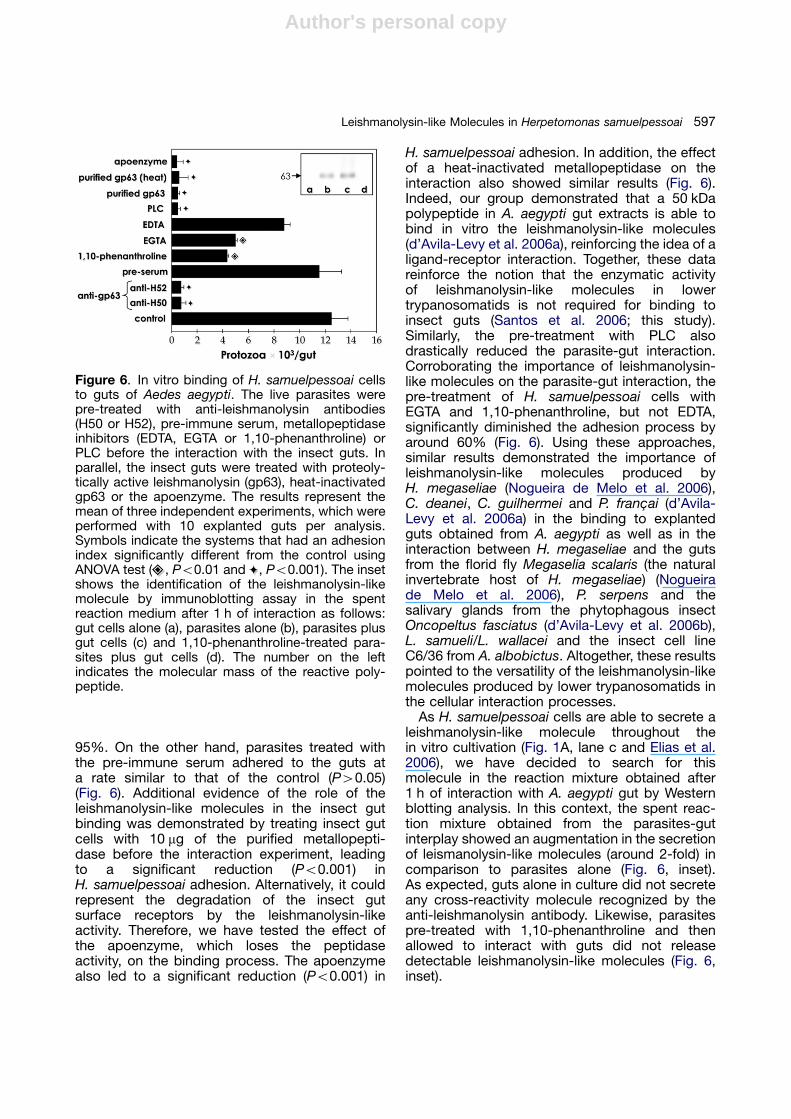
of paramount significance in host-parasiterelationships as all interactions between hostand parasite occur at least initially at the level ofparasite surface membrane. The parasite surfacemembrane plays a pivotal role in the survival,nourishment and multiplication of the parasitewithin the infected/colonized host, especially indirect confrontation with the host immuneresponse. Since A. aegypti has proven tobe a good experimental model to study theinteraction between lower trypanosomatids andvectors (Correa-da-Silva et al. 2006; Fampa et al.2003; reviewed by Santos et al. 2006; and thisstudy), we have performed binding assays withH. samuelpessoai parasites previously treatedwith the anti-leishmanolysin antibodies, at aconcentration that did not promote parasiteagglutination. This treatment led to a significantreduction (Po0.001) in the capacity of adhesion tothe guts of H. samuelpessoai by approximately
Figure 5. Flow cytometry analyses showing the leishmanolysin-like molecules expressed in H. samuelpessoaicells cultured in BHI medium or parasites after passage in A. aegypti for 6 days. PBS-washed parasites wereincubated in the absence (control) or in the presence of anti-leishmanolysin antibodies (gp63) at 1: 500 dilutionas described in the methods section, and then analyzed by flow cytometry.
F.M. Pereira et al.596
Author's personal copy
95%. On the other hand, parasites treated withthe pre-immune serum adhered to the guts ata rate similar to that of the control (P40.05)(Fig. 6). Additional evidence of the role of theleishmanolysin-like molecules in the insect gutbinding was demonstrated by treating insect gutcells with 10 mg of the purified metallopepti-dase before the interaction experiment, leadingto a significant reduction (Po0.001) inH. samuelpessoai adhesion. Alternatively, it couldrepresent the degradation of the insect gutsurface receptors by the leishmanolysin-likeactivity. Therefore, we have tested the effect ofthe apoenzyme, which loses the peptidaseactivity, on the binding process. The apoenzymealso led to a significant reduction (Po0.001) in
H. samuelpessoai adhesion. In addition, the effectof a heat-inactivated metallopeptidase on theinteraction also showed similar results (Fig. 6).Indeed, our group demonstrated that a 50 kDapolypeptide in A. aegypti gut extracts is able tobind in vitro the leishmanolysin-like molecules(d’Avila-Levy et al. 2006a), reinforcing the idea of aligand-receptor interaction. Together, these datareinforce the notion that the enzymatic activityof leishmanolysin-like molecules in lowertrypanosomatids is not required for binding toinsect guts (Santos et al. 2006; this study).Similarly, the pre-treatment with PLC alsodrastically reduced the parasite-gut interaction.Corroborating the importance of leishmanolysin-like molecules on the parasite-gut interaction, thepre-treatment of H. samuelpessoai cells withEGTA and 1,10-phenanthroline, but not EDTA,significantly diminished the adhesion process byaround 60% (Fig. 6). Using these approaches,similar results demonstrated the importance ofleishmanolysin-like molecules produced byH. megaseliae (Nogueira de Melo et al. 2006),C. deanei, C. guilhermei and P. franc-ai (d’Avila-Levy et al. 2006a) in the binding to explantedguts obtained from A. aegypti as well as in theinteraction between H. megaseliae and the gutsfrom the florid fly Megaselia scalaris (the naturalinvertebrate host of H. megaseliae) (Nogueirade Melo et al. 2006), P. serpens and thesalivary glands from the phytophagous insectOncopeltus fasciatus (d’Avila-Levy et al. 2006b),L. samueli/L. wallacei and the insect cell lineC6/36 from A. albobictus. Altogether, these resultspointed to the versatility of the leishmanolysin-likemolecules produced by lower trypanosomatids inthe cellular interaction processes.
As H. samuelpessoai cells are able to secrete aleishmanolysin-like molecule throughout thein vitro cultivation (Fig. 1A, lane c and Elias et al.2006), we have decided to search for thismolecule in the reaction mixture obtained after1 h of interaction with A. aegypti gut by Westernblotting analysis. In this context, the spent reac-tion mixture obtained from the parasites-gutinterplay showed an augmentation in the secretionof leismanolysin-like molecules (around 2-fold) incomparison to parasites alone (Fig. 6, inset).As expected, guts alone in culture did not secreteany cross-reactivity molecule recognized by theanti-leishmanolysin antibody. Likewise, parasitespre-treated with 1,10-phenanthroline and thenallowed to interact with guts did not releasedetectable leishmanolysin-like molecules (Fig. 6,inset).
Figure 6. In vitro binding of H. samuelpessoai cellsto guts of Aedes aegypti. The live parasites werepre-treated with anti-leishmanolysin antibodies(H50 or H52), pre-immune serum, metallopeptidaseinhibitors (EDTA, EGTA or 1,10-phenanthroline) orPLC before the interaction with the insect guts. Inparallel, the insect guts were treated with proteoly-tically active leishmanolysin (gp63), heat-inactivatedgp63 or the apoenzyme. The results represent themean of three independent experiments, which wereperformed with 10 explanted guts per analysis.Symbols indicate the systems that had an adhesionindex significantly different from the control usingANOVA test ( , Po0.01 and , Po0.001). The insetshows the identification of the leishmanolysin-likemolecule by immunoblotting assay in the spentreaction medium after 1 h of interaction as follows:gut cells alone (a), parasites alone (b), parasites plusgut cells (c) and 1,10-phenanthroline-treated para-sites plus gut cells (d). The number on the leftindicates the molecular mass of the reactive poly-peptide.
597Leishmanolysin-like Molecules in Herpetomonas samuelpessoai

Author's personal copy
Conclusion
H. samuelpessoai cells provide an additional toolfor the investigation of ancestral functions ofleishmanolysin-like molecules during the insectcolonization. This ubiquitous surface metallo-typepeptidase is able to cleave numerous proteins,being active at a wide range of temperatures(including both insect and mammalian ones) andpH, and also contributes to the parasite adhesionto explanted guts of A. aegypti, an experimentalinsect model used in studies with trypanosoma-tids. The role of leishmanolysin in insect interac-tion is reinforced by its enhanced secretion duringthe interaction processes with explanted guts aswell as its increased exposition at the parasitesurface after passage in A. aegypti.
Methods
Parasite and growth conditions: The protozoan Herpetomonassamuelpessoai (CT–IOC–067) was provided by the Colec- ao deTripanossomatıdeos, Instituto Oswaldo Cruz, FIOCRUZ, Rio deJaneiro, Brazil. The trypanosomatid was maintained by weeklytransfers in brain heart infusion (BHI) medium. For the experi-ments, cells were grown in 100-mL Erlenmeyer flasks containing20 mL of autoclaved BHI medium. The inoculum consisted of1 mL of a 48-h culture, containing about 2� 106 cells m L�1.Cells were cultivated at 26 1C for 48 h (exponential growthphase) (Santos et al. 2001). Parasite growth was estimated bydetermining the cell concentration in a Neubauer chamber.Cellular viability was assessed by motility and exclusion oftrypan blue dye from cells.
Insect: Aedes aegypti adult female mosquitoes werereared and maintained at the Laboratorio de Fisiologia eControle de Vetores, Departamento de Entomologia (InstitutoOswaldo Cruz, Rio de Janeiro, Brazil) and kindly provided byDr. Denise Valle. Insects were allowed to feed with a 2%sucrose water solution. Guts were carefully dissected, rinsedthree times in cold phosphate-buffered saline (PBS; 150 mMNaCl, 20 mM phosphate buffer, pH 7.2) at 4 1C and immedi-ately used for the experiments.
Parasite extracts: Parasites were collected by centrifuga-tion (500 g/5 min/4 1C), washed three times with PBS,resuspended in 200 mL (1� 108 cells) of PBS and lysed bythe addition of 1% SDS. The cells were broken in a vortex byalternating 1 min shaking and 2 min cooling intervals, followedby a centrifugation at 5000 g for 15 min at 4 1C. The super-natants obtained after centrifugation corresponded to thewhole parasite cellular extracts (Santos et al. 2006). To obtainthe membrane-rich fraction, parasites (1�109 cells) wereextracted with 500 mL of 2% pre-condensed Triton X-114 inTBS (150 mM NaCl; 10 mM Tris, pH 7.4) as described byBordier (1981). In parallel, culture supernatant fluids werefiltered through a 0.22-mm pore-size membrane (Millipore) andthen concentrated 10-fold by ultrafiltration in a 10000molecular mass cut-off Centricon microconcentrator(AMICON, Beverly, MA, USA) at 4 1C (Santos et al. 2005).The same volume of fresh BHI medium was also concentratedand used as a control to test for possible proteolytic
activity, yielding negative results. Protein concentration wasdetermined by the method of Lowry et al. (1951), using bovineserum albumin (BSA) as standard. The parasite extracts werealiquoted and the fractions stored at �80 1C for furtheranalyses.
Insect gut extract: To obtain soluble gut extract, 20 insectguts were resuspended in 500 mL of PBS, supplementedwith a proteolytic inhibitor cocktail (1 mM E-64, 1 mM1,10-phenanthroline, 1 mM phenylmethylsulfonyl fluoride[PMSF], and 1 mM pepstatin A), homogenized with a Teflon-coated microtissue grinder and vigorously vortexed (threetimes for 30 sec) in the presence of 1% SDS, followed bycentrifugation at 5000 g for 10 min at 4 1C. The supernatantobtained after the centrifugation corresponded to the insectgut extract and contained approximately 1 mg protein per mL.Then, the gut extract was heated for 5 min at 100 1C,alternating 10 min cooling intervals (three times), to abolishthe endogenous proteolytic activity (Elias et al. 2008).
Peptidase activity assay: The proteolytic activity wasassayed by 10% SDS–PAGE with 0.1% gelatin incorporatedinto the gel as protein substrate. The gels were loaded with50 mg of parasite extracts that were added to SDS–PAGEsample buffer (125 mM Tris, pH 6.8, 4% SDS, 20% glyceroland 0.002% bromophenol blue). Electrophoresis wasperformed at a constant current of 60 mA at 4 1C for 2 h.After electrophoresis, SDS was removed by incubation with10 volumes of 2.5% Triton X-100 for 1 h under constantagitation. Subsequently, the gels were incubated in 50 mMsodium phosphate buffer, pH 6.0, supplemented with 2 mMdithiothreitol (DTT) at 37 1C, in the absence or in thepresence of the following proteolytic inhibitors: 1 mM E-64or 10 mM 1,10-phenanthroline. After incubation for 20 h, thegels were washed twice with distilled water, stained for 2 hwith 0.2% Coomassie brilliant blue R-250 in methanol-acetic acid-water (50:10:40), and destained overnight in asolution containing methanol-acetic acid-water (5:10:85), tointensify the proteolytic halos. The molecular mass of thepeptidases was estimated by comparison with the mobilityof low molecular mass protein standards. The gels werephotographed and the density profiles digitally processed(Elias et al. 2008).
Effect of pH and temperature on the surface peptidaseactivity: In this set of experiments, the proteolytic activity wasevaluated using the standard assay conditions described above.The effect of the pH was determined by incubating the gel stripsat 37 1C for 20 h in the following buffers: 50 mM sodiumphosphate (pH 5.0–7.0) and 20 mM glycine-NaOH (pH 8.0–9.0). The proteolytic enzyme was also analyzed in gelatin-SDS-PAGE at different temperatures (4, 26, 37, 50 and 65 1C) for 20 hin 50 mM sodium phosphate buffer, pH 6.0 (Elias et al. 2006).
Effect of different protein substrates on the surfacepeptidase activity: The ability of the surface peptidase todegrade different proteinaceous substrates was also evalu-ated by 10% SDS–PAGE co-polymerized with 0.1% gelatin,casein, hemoglobin, BSA, human serum albumin (HSA),mucin, IgG or 100 mg of the gut extract from A. aegypti (Eliaset al. 2006, 2008). Electrophoresis was performed asdescribed above and the gel strips were incubated for 20 hin 50 mM sodium phosphate buffer, pH 6.0.
Immunoblotting: Samples containing both cellular and super-natant extracts, equivalent to 50 mg of protein, were added toSDS–PAGE sample buffer and mixed with 10% b-mercaptoetha-nol, followed by heating at 100 1C for 5 min. Thereafter, proteinextracts were separated in 12% SDS–PAGE and the polypeptideselectrophoretically transferred at 4 1C at 100 V/300 mA for 2 h toa nitrocellulose membrane. The membrane was blocked in 5%
F.M. Pereira et al.598

Author's personal copy
low-fat dried milk in PBS containing 0.5% Tween 20 (PBS/Tween)for 1 h at room temperature. Then, membranes were washedthree times (10 min each) with the blocking solution and incubatedfor 1 h with the polyclonal anti-leishmanolysin antibodies at 1:500(H50, serum against recombinant leishmanolysin, and H52, serumagainst the native protein without the diacylglycerol-anchor fromLeishmania mexicana). These antibodies were kindly provided byDr Peter Overath, Max-Planck-Institut fur Biologie, AbteilungMembranbiochemie, Germany. The secondary antibody usedwas peroxidase-conjugated goat anti-rabbit IgG at 1:25000followed by chemiluminescence immunodetection (Santos et al.2006). The relative molecular masses of the reactive polypeptideswere calculated by comparison with the mobility of low molecularmass protein standards.
Fluorescence microscopy and flow cytometry analysesfor cell surface leishmanolysin-like molecule: Parasites(1� 107 cells) were fixed in 0.4% paraformaldehyde diluted inPBS for 30 min at room temperature, followed by extensivewashing in the same buffer. These fixed cells maintained theirmorphological integrity, as verified by microscopic observa-tion. Cells (5�106 cells) were incubated with 0.2 U mL�1 ofBacillus thuringiensis PLC (BtPLC) in a final reaction mixture(400 mL) containing 100 mM Tris-acetate, pH 7.4, supplemen-ted with 0.1% Triton X-100 for 1 h at 37 1C (Santos et al. 2002).Control cells were subjected to the same experimentalconditions, except for the presence of BtPLC. Untreatedand BtPLC-treated H. samuelpessoai cells were then incu-bated for 1 h at room temperature with a 1:500 dilution ofrabbit anti-leishmanolysin antibody, and then incubated for anadditional hour with a 1:250 dilution of fluorescein isothiocya-nate (FITC)–labeled goat anti-rabbit IgG. The cells were thenwashed three times in PBS and observed in an OlympusFluoview confocal microscope equipped with a �100 oilobjective. The images were digitally recorded using a cooledCCD camera (Color View XS, Analysis GmBH, Germany) andanalyzed with AnalySIS system software. Alternatively, theparasite-associated fluorescence was excited at 488 nm andquantified on a fluorescence-activated cell sorter (FACS)(FACSCalibur,BD Bioscience, USA) equipped with a 15-mWargon laser emitting at 488 nm. Non-treated cells and thosetreated with the secondary antibody alone were run in parallelas controls. Each experimental population was then mappedby using a two-parameter histogram of forward-angle lightscatter versus side scatter. The mapped population(n=10,000) was then analyzed for log green fluorescence byusing a single-parameter histogram.
Immunocytochemistry analysis: The parasites werecollected by centrifugation, washed twice in PBS and fixedin a solution containing 0.1% glutaraldehyde, 2% formalde-hyde in 0.1% cacodylate buffer, pH 7.2. After 1 h fixation, cellswere washed in PBS, dehydrated in methanol and embeddedin Lowicryl K4M resin at �20 1C. Ultrathin sections werecollected in nickel grids and incubated in 0.1 M TBS followedby a 15 min incubation in 50 mM ammonium chloride. Afterthree washings in TBS, the grids were incubated with thepre-serum or with the anti-leishmanolysin antibody diluted at1:50 in TBS, 1% BSA, 1% Tween 20 (TBSBT). Grids werewashed three times in TBSBT and subsequently incubatedwith gold-labeled goat anti-rabbit IgG (15 nm, dilution 1:10) for60 min (Souto-Padron et al. 1990). Ultrathin sections werestained with uranyl acetate and lead citrate and observed in aZeiss ETM 10 C transmission electron microscope.
Feeding of A. aegypti with H. samuelpessoai: Femalemosquitoes were starved 15 h before feeding on a cottonswab dipped in the trypanosomatid suspension (5� 107 cellsm L�1 in Hanks’ balanced salt solution, supplemented with
2% BSA and 2% sucrose). Fed females were separated andtheir guts were dissected after different times post-feeding upto 8 days, transferred to microcentrifuge tubes containing50 mL of PBS and homogenized. The released trypanosoma-tids were counted in a Neubauer chamber. The results areshown as means7standard error (SE) of three independentexperiments in which 10–20 mosquitoes were examined perday (Correa-da-Silva et al. 2006; Fampa et al. 2003). Toanalyze the possible modulation on the expression of surfaceleishmanolysin-like molecules in H. samuelpessoai afterin vivo passage in A. aegypti, insect guts were dissectedand parasite cells were recovered, washed in PBS andprocessed for flow cytometry as described above using theanti-leishmanolysin antibody.
Protozoa–insect gut interaction: In this set of experi-ments, live parasites (1� 107 cells) were incubated for 1 h atroom temperature in the absence (control) or in the presenceof different modulators: (1) metallopeptidase inhibitors (EDTA,EGTA, and 1,10-phenanthroline) at a final concentration of1 mM; (2) anti-leishmanolysin antibodies (H52, and H50) or anirrelevant IgG (rabbit pre-immune serum), both at 1:100dilution; or (3) BtPLC at 0.2 U mL�1. The viability of theorganisms throughout the experiment was assessed bymobility, trypan blue dye exclusion and by monitoring lactatedehydrogenase, an intracellular enzyme, in the supernatantfrom the incubation systems (Elias et al. 2006). The cells werethen washed three times in cold PBS, resuspended in PBS(5� 106 cells in 100 mL) and added to dissected guts (10 pergroup) that were sliced open longitudinally. Alternatively, thedissected guts were incubated for 30 min in PBS (100 mL)containing 10 mg of either the purified and proteolyticallyactive purified leishmanolysin-like molecule, heat-inactivatedprotein or an inactive apoenzyme obtained after dialysisagainst a solution containing 1 mM of the following metallo-peptidase inhibitors: 1,10-phenanthroline, EDTA and EGTA(d’Avila-Levy et al. 2003, 2006a, 2006b); after that, theinteraction process was performed. The parasites wereallowed to interact with insect gut for 1 h at room temperaturein PBS. After the interaction period, the guts were extensivelywashed with PBS, individually transferred to microcentrifugetubes containing 50 mL of PBS and homogenized (Fampaet al. 2003). The released trypanosomatids were counted ina Neubauer chamber. The results are shown as means7standard error (SE) of three independent experiments.
Statistical analysis: All experiments were performed intriplicate, in three independent experimental sets. The meanand standard error of at least three distinct experiments weredetermined. Statistical analysis was calculated by using EPI-INFO 6.04 (Database and Statistics Program for Public Health)computer software. The analysis of variance between groupswas performed by means of ANOVA test. P values of 0.01 orless were considered statistically significant.
Acknowledgements
We thank Dr. Peter Overath (Max-Planck-Institutfur Biologie, Abteilung Membranbiochemie,Germany) for donating the anti-leishmanolysinantibody and the rabbit pre-immune antiserum.This work was supported by grants from Funda-c- aoCarlos Chagas Filho de Amparo �a Pesquisa(FAPERJ), Conselho Nacional de Desenvolvimento
599Leishmanolysin-like Molecules in Herpetomonas samuelpessoai

Author's personal copy
Cientıfico e Tecnologico (MCT/CNPq), Fundac- aoUniversitaria Jose Bonifacio (FUJB), Coordenac- aode Aperfeic-oamento Pessoal de Nıvel Superior(CAPES), Conselho de Ensino para Graduados ePesquisa (CEPG/UFRJ), and Funda-c- ao OswaldoCruz (FIOCRUZ).
References
Almeida FVS, Giovanni-de-Simone S, Branquinha MH,Vermelho AB (2003) Extracellular metalloproteinase activityin Phytomonas franc-ai. Parasitol Res 89:320–322
Bordier C (1981) Phase separation of integral membraneproteins in Triton X-114 solution. J Biol Chem 256:1604–1607
Bregano JW, Picao RC, Grac-a VK, Menolli RA, ItowJankevicius S, Pinge Filho P, Jankevicius JV (2003)Phytomonas serpens, a tomato parasite, shares antigens withTrypanosoma cruzi that are recognized by human sera andinduce protective immunity in mice. FEMS Immunol MedMicrobiol 39:257–264
Cerisola JA, Rohwedder R, Bozzini JP, Del Prado CE (1971)Blastocrithidia triatomae n. sp. found in Triatoma infestansfrom Argentina. J Protozool 18:503–506
Chaudhuri G, Chang KP (1988) Acid protease activity of amajor surface membrane glycoprotein (gp63) from Leishmaniamexicana promastigotes. Mol Biochem Parasitol 27:43–52
Chicharro C, Alvar J (2003) Lower trypanosomatids in HIV/AIDS patients. Ann Trop Med Parasitol 97:75–78
Correa-da-Silva MS, Fampa P, Lessa LP, Silva ER,Santos-Mallet JR, Saraiva EM, Motta MCM (2006) Coloni-zation of Aedes aegypti midgut by the endosymbiont-bearingtrypanosomatid Blastocrithidia culicis. Parasitol Res 99:384–391
d’Avila-Levy CM, Souza RF, Gomes RC, Vermelho AB,Branquinha MH (2003) A metalloproteinase extracellularlyreleased by Crithidia deanei. Can J Microbiol 49:625–632
d’Avila-Levy CM, Santos LO, Marinho FA, Dias FA, LopesACHS, Santos ALS, Branquinha MH (2006b) Gp63-likemolecules in Phytomonas serpens: possible role on the insectinteraction. Curr Microbiol 36:415–422
d’Avila-Levy CM, Dias FA, Nogueira-de-Melo AC, MartinsJL, Lopes ACHS, Santos ALS, Vermelho AB, BranquinhaMH (2006a) Insights into the role of gp63-like proteins in lowertrypanosomatids. FEMS Microbiol Lett 254:149–156
d’Avila-Levy CM, Santos LO, Marinho FA, Matteoli FP,Lopes ACHS, Motta MCM, Santos ALS, Branquinha MH(2008) Crithidia deanei: influence of parasite gp63 homologueon the interaction of endosymbiont-harboring and aposym-biotic strains with Aedes aegypti midgut. Exp Parasitol118:345–353
d’Avila-Levy CM, Silva BA, Dias FA, Hayasi EA, VermelhoAB, Alviano CS, Saraiva EMB, Branquinha MH, Santos ALS(2005) Influence of the endosymbiont of Blastocrithidia culicisand Crithidia deanei on the glycoconjugate expression and onAedes aegypti interaction. FEMS Microbiol Lett 252:279–286
Elias CGR, Pereira FM, Silva BA, Alviano CS, Soares RMA,Santos ALS (2006) Leishmanolysin (gp63 metallopeptidase)-
like activity extracellularly released by Herpetomonas samuel-pessoai. Parasitology 132:37–47
Elias CGR, Pereira FM, Dias FA, Silva TL, Lopes AH,d’Avila-Levy CM, Branquinha MH, Santos ALS (2008)Cysteine peptidases in the tomato trypanosomatid Phytomo-nas serpens: influence of growth conditions, similarities withcruzipain and secretion to the extracellular environment. ExpParasitol 120:343–352
Fampa P, Correa-da-Silva MS, Lima DC, Oliveira SM,Motta MCM, Saraiva EM (2003) Interaction of insecttrypanosomatids with mosquitoes, sand fly and the respectiveinsect cell lines. Int J Parasitol 33:1019–1026
Hajmova M, Chang KP, Kolli B, Volf P (2004) Down-regulation of gp63 in Leishmania amazonensis reduces itsearly development in Lutzomyia longipalpis. Microbes Infect6:646–649
Inverso JA, Medina-Acosta E, O’Connor J, Russell DG,Cross GA (1993) Crithida fasciculata contains a transcribedleishmanial surface peptidase (gp63) gene homologue. MolBiochem Parasitol 57:47–54
Iwanaga S, Lee BL (2005) Recent advances in the innateimmunity of invertebrate animals. J Biochem Mol Biol 38:128–150
Joshi PB, Kelly BL, Kamhawi S, Sacks DL, McMaster WR(2002) Targeted gene deletion in Leishmania major identifiesleishmanolysin (GP63) as a virulence factor. Mol BiochemParasitol 120:33–40
Kulkarni MM, Jones EA, McMaster WR, McGwire BS (2008)Fibronectin binding and proteolytic degradation by Leishma-nia and effects on macrophage activation. Infect Immun76:1738–1747
Kulkarni MM, McMaster WR, Kamysz E, Kamysz W,Engman DM, McGwire BS (2006) The major surface-metalloprotease of the parasitic protozoan, Leishmania,protects against antimicrobial peptide-induced apoptotickilling. Mol Microbiol 62:1484–1497
Lima AKC, Elias CGR, Souza JE, Santos ALS, Dutra PML(2009) Dissimilar peptidase production by avirulent andvirulent promastigotes of Leishmania braziliensis: inferenceon the parasite proliferation and interaction with macro-phages. Parasitology 136:1179–1191
Lopes JD, Caulada Z, Barbieri CL, Camargo EP (1981)Cross-reactivity between Trypanosoma cruzi and insecttrypanosomatids as a basis for the diagnosis of Chagas’disease. Am J Trop Med Hyg 30:1183–1188
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951)Protein measurement with the Folin phenol reagent. J BiolChem 193:265–275
Matta MAV, Alviano DS, Couceiro JNSS, Meirelles MNL,Alviano CS, Angluster J (1999) Cell-surface sialoglyconju-gate strucutures in wild-type and mutant Crithidia fasciculata.Parasitol Res 85:293–299
McConville MJ, Mullin KA, Ilgoutz SC, Teasdale RD (2002)Secretory pathway of trypanosomatid parasites. MicrobiolMol Biol Rev 66:122–154
McGwire BS, Chang KP, Engman DM (2003) Migrationthrough the extracellular matrix by the parasitic protozoan
F.M. Pereira et al.600

Author's personal copy
Leishmania is enhanced by surface metalloprotease gp63.Infect Immun 71:1008–1010
Mezincescu D (1908) Les trypanosomes des moustiques etleur relation avec les Haemoproteus des oiseaux. C RSeances Soc Biol Fil 64:975–976
Morio F, Reynes J, Dollet M, Pratlong F, Dedet JP, Ravel C(2008) Isolation of a protozoan parasite genetically related tothe insect trypanosomatid Herpetomonas samuelpessoai froma human immunodeficiency virus-positive patient. J ClinMicrobiol 46:3845–3847
Muhich ML, Boothroyd JC (1988) Polycistronic transcripts intrypanosomes and their accumulation during heat shock:evidence for a precursor role in mRNA synthesis. Mol Cell Biol8:3837–3846
Murphy WJ, Watkins KP, Agabian N (1986) Identification of anovel Y branch structure as an intermediate in trypanosomemRNA processing: evidence for trans-splicing. Cell 47:517–525
Noguchi H, Tilden EB (1926) Comparative studies ofHerpetomonads and Leishmanias. 1. Cultivation ofHerpetomonads from insects and plants. J Exp Med44:307–325
Nogueira de Melo AC, Giovanni-de-Simmone S, Bran-quinha MH, Vermelho AB (2001) Crithidia guilhermei pur-ification and partial characterization of a 62-kDa extracellularmetalloproteinase. Exp Parasitol 97:1–8
Nogueira de Melo AC, d’Avila-Levy CM, Dias FA, ArmadaJLA, Silva HD, Lopes ACHS, Santos ALS, Branquinha MH,Vermelho AB (2006) Peptidases and gp63-like proteinsin Herpetomonas megaseliae: possible involvementin the adhesion to the invertebrate host. Int J Parasitol36:415–422
Novy FG, McNeal WJ, Torrey HN (1907) The trypanosomesof mosquitoes and other insects. J Infect Dis 4:223–276
Pereira FM, Elias CGR, d’Avila-Levy CM, Branquinha MH,Santos ALS (2009b) Cysteine peptidases in Herpetomonassamuelpessoai are modulated by temperature anddimethylsulfoxide-triggered differentiation. Parasitology136:45–54
Pereira FM, Bernardo PS, Dias Junior PFF, Silva BA,Romanos MTV, d’ Avila-Levy CM, Branquinha MH, SantosALS (2009a) Differential influence of gp63-like molecules inthree distinct Leptomonas species on the adhesion to insectcells. Parasitol Res 104:347–353
Pimenta PF, Saraiva EM, Rowton E, Modi GB, GarrawayLA, Beverley SM, Turco SJ, Sacks DL (1994) Evidence thatthe vectorial competence of phlebotomine sand flies fordifferent species of Leishmania is controlled by structuralpolymorphisms in the surface lipophosphoglycan. Proc NatlAcad Sci USA 91:9155–9159
Podlipaev SA (2001) The more insect trypanosomatids understudy—the more diverse Trypanosomatidae appears. Int JParasitol 31:648–652
Podlipaev SA, Sturm NR, Fiala I, Fernandes O, WestenbergSJ, Dollet M, Campbell DA, Lukes J (2004) Diversity ofinsect trypanosomatids assessed from the splice leader RNAand 5S rRNA genes and intergenic region. J EukaryotMicrobiol 51:283–290
Sadlova J, Volf P, Victoir K, Dujardin JC, Votypka J (2006)Virulent and attenuated lines of Leishmania major: DNAkaryotypes and differences in metalloproteinase GP63. FoliaParasitol 53:81–90
Santos ALS, Branquinha MH, d’Avila-Levy CM (2006) Theubiquitous gp63-like metalloprotease from lower trypanoso-matids: in the search for a function. An Acad Bras Cienc78:687–714
Santos ALS, Abreu CM, Alviano CS, Soares RMA (2002)Activation of the glycosylphosphatidylinositol membraneproteinase upon released from Herpetomonas samuelpessoaiby phospholipase C. Curr Microbiol 45:293–298
Santos ALS, Abreu CM, Alviano CS, Soares RMA (2005)Use of proteolytic enzymes as an additional tool fortrypanosomatid identification. Parasitology 130:79–88
Santos ALS, d’Avila-Levy CM, Elias CG, Vermelho AB,Branquinha MH (2007) Phytomonas serpens immunologicalsimilarities with the human trypanosomatid pathogens. MicrobInfect 9:915–921
Santos ALS, Rodrigues ML, Alviano CS, Angluster J,Soares RMA (2003) Herpetomonas samuelpessoai: dimethyl-sulfoxide-induced differentiation is influenced by proteinaseexpression. Curr Microbiol 46:11–17
Santos ALS, Batista LM, Abreu CM, Alviano CS, AnglusterJ, Soares RMA (2001) Developmentally regulated proteinexpression mediated by dimethylsulfoxide in Herpetomonassamuelpessoai. Curr Microbiol 42:111–116
Santos-Gomes GM, Abranches P (1996) Comparative studyof infectivity caused by promastigotes of Leishmania infantumMON-1, L. infantum MON-24 and L. donovani MON-18. FoliaParasitol 43:7–12
Seay MB, Heard PL, Chaudhuri G (1996) SurfaceZn-proteinase as a molecule for defense of Leishmaniamexicana amazonensis promastigotes against cytolysisinside macrophage phagolysosomes. Infect Immun 64:5129–5137
Shaw JM, Feagin JE, Stuart K, Simpson L (1988) Editing ofkinetoplastid mitochondrial mRNAs by uridine addition anddeletion generates conserved amino acid sequences andAUG initiation codons. Cell 53:401–411
Silva BA, Pereira FM, de Jesus JB, Branquinha MH, SantosALS, d’Avila-Levy CM (2009) Sialoglycoconjugates in Herpe-tomonas megaseliae: role in the adhesion to insect hostepithelial cells. FEMS Microbiol Lett 295:274–280
Simpson AGB, Stevens JR, Lukes J (2006) The evolutionand diversity of kinetoplastid flagellates. Trends Parasitol22:168–174
Souto-Padron T, Campetella OE, Cazzulo JJ, de Souza W(1990) Cysteine proteinase in Trypanosoma cruzi: immunocy-tochemical localization and involvement in parasite-host cellinteraction. J Cell Sci 96:485–490
Sutton RE, Boothroyd JC (1986) Evidence for trans-splicingin trypanosomes. Cell 47:527–535
Tschudi C, Ullu E (1988) Polygene transcripts are precursorsto calmodulin mRNAs in trypanosomes. EMBO J 7:455–463
Wallace FG (1966) The trypanosomatids parasites of insectsand arachnids. Exp Parasitol 18:124–193
601Leishmanolysin-like Molecules in Herpetomonas samuelpessoai

Author's personal copy
Wallace FG, Johnson A (1961) The infectivity of old culturedstrains of mosquito flagellates. J Insect Pathol 3:75–80
Weinman D, Cheon WH (1978) Herpetomonas, with bacter-ium-like inclusions, in Malaysian Aedes aegypti and Aedesalbopictus. J Protozool 25:167–169
Wilson ME, Hardin KK, Donelson JE (1989) Expression of themajor surface glycoprotein of Leishmania donovani chagasi invirulent and attenuated promastigotes. J Immunol 143:678–684
Yao C, Donelson JE, Wilson ME (2003) The major surfaceprotease (MSP or GP63) of Leishmania sp. Biosynthesis,regulation of expression, and function. Mol Biochem Parasitol132:1–16
Yao C, Luo J, Hsiao C, Donelson JE, Wilson ME (2005)Internal and surface subpopulations of the major surfaceprotease (MSP) of Leishmania chagasi. Mol Biochem Parasitol139:173–183
F.M. Pereira et al.602




















