
Biological control with Trichogramma pretiosum increases ...
Upload
independentCategory
view
0download
0
Journal of Chemical Ecology, Vol. 23, No. 11, 1997
VOLATILE PLANT INFOCHEMICALS MEDIATE PLANTPREFERENCE OF Trichogramma chilonis
JORG ROMEIS,1'2'* THOMAS G. SHANOWER,' andCLAUS P. W. ZEBITZ2
'Crop Protection DivisionInternational Crops Research Institute for the Semi-Arid Tropics (ICRISAT)
Patancheru, Andhra Pradesh 502 324, India
'Institute of PhytomedicineUniversity of Hohenheim
70593 Stuttgart, Germany
(Received January 28, 1997; accepted July 7, 1997)
Abstract—In India, eggs of the polyphagous noctuid moth Helicoverpa armi-gera on sorghum are parasitized to high levels by Trichogramma spp., butonly rarely are parasitized eggs found on pigeonpea. This study was conductedto test whether volatile plant infochemicals contribute to the different parasit-ism levels observed on these two crops. In a tour-armed airflow otfactometer,volatiles emitted by both sorghum and pigeonpea plants elicited a behavioralresponse form Trichogramma chilonis females. The parasitoids' response var-ied depending on the growth stage of the plant. Volatiles emitted by sorghumin the vegetative and reproductive stages arrested the parasitoids. T. chilonisfemales did not respond to volatiles from pigeonpea in the vegetative stage,but were repelled by volatiles from plants in the reproductive stage. Plants inthe reproductive stage are preferred for oviposition by H. armigera. Thus,sorghum is attractive and pigeonpea repellent to T. chilonis females at thetime when each plant is attractive to the host. This difference in the parasitoids'response may partly explain the different levels of egg parasitism reportedfrom these two crops. The infochemicals involved in these plant-parasitoidinteractions are discussed in the context of the current terminology.
Key Words— Trichogramma chilonis, Hymenoptera, Trichogrammatidae,pigeonpea, sorghum, Helicoverpa armigera, host location, airflow olfac-tometer, plant volatiles.
*To whom correspondence should be addressed at ICRISAT.
2455
0098-0331/97/II00- 2455/$I2 50/0 IS 1997 Plenum Publishing Corporation

2456 ROMEIS, SHANOWER, AND ZEBITZ
INTRODUCTION
Parasitoids of herbivores use chemical stimuli (infochemicals) from both plantsand hosts during the early steps in the host location process (Tumlinson et al.,1992; Vet and Dicke, 1992). These stimuli vary in their information contentand in their detectability. Stimuli derived from the host are more reliable butgenerally difficult to detect, while stimuli from the plant are easier to detect butgenerally carry less host-specific information (Vet et al., 1991). Some parasi-toids deal with this reliability-detectability problem by responding to herbivore-induced synomones, plant-produced infochemicals released after damage by theherbivore (Tumlinson et al., 1992; Vet and Dicke, 1992). Egg parasitoids suchas Trichogramma spp. (Hymenoptera: Trichogrammatidae) are not able to usestimuli derived from their hosts because insect eggs do not emit long-rangevolatiles (Kaiser et al., 1989; Noldus, 1989). Therefore, these parasitoids mustrely on other information indicating the presence of host eggs on a plant, suchas stimuli from another life stage of the host that may be easier to detect (info-chemical detour; Vet et al., 1991). Two types of potential information are hostsex pheromones and plant volatiles. Noldus (1989) listed several examples ofTrichogramma spp. responding to host sex pheromones, and a number of studieshave shown that Trichogramma spp. are attracted/arrested or repelled by plantvolatiles (Bar et al., 1979; Cabello and Vargas, 1985; Nordlund et al., 1985;Kaiser et al., 1989).
The following study was conducted to test if T. chilonis Ishii femalesrespond to volatiles emitted by pigeonpea [Cajanus cajan (L.) MillspaughJ andsorghum (Sorghum bicolor L.) plants. Pigeonpea is an important grain legumein the semiarid tropics (Nene and Sheila, 1990). Its most devastating pest in theOld World is the pod borer, Helicoverpa armigera (Hubner) (Lepidoptera: Noc-tuidae) (Lateef and Reed, 1990). One possible reason for the key pest status ofH. armigera on pigeonpea is the low incidence and small impact of naturalenemies, including Trichogramma spp., on this crop (Romeis and Shanower,1996). In India, for example, reported parasitism levels by Trichogramma spp.,mainly T. chilonis, can reach an average of 40% on eggs collected from sor-ghum, while less than 1 % of eggs on pigeonpea are parasitized (Romeis andShanower, 1996). Volatile plant infochemicals may partly explain the differentparasitism levels of H. armigera eggs on these two crops.
METHODS AND MATERIALS
Parasitoids. T. chilonis was reared at 26 ± 2°C, 50 ± 10% relativehumidity, and 16L:8D photoperiod on eggs of the factitious host Corcyra

Trichogramma PLANT PREFERENCE 2457
cephalonica Stainton (Lepidoptera: Phyralidae). C. cephalonica was reared onsorghum grain. The parasitoid strain was collected in 1980 on H. armigera eggsfrom cotton and tomato plants near Hyderabad, India, and has been rearedcontinuously on C. cephalonica. The female parasitoids used for the experimentswere 1-2 days old, mated, inexperienced, and fed with honey agar.
Olfactometer Bioassay, The four-armed airflow olfactometer used in thisstudy was slightly modified from the one used by Noldus (1988) and Noldus andvan Lenteren (1985) for Trichogramma spp. The exposure chamber consistedof a single piece of white acetal (Delrin) covered with a glass plate. The setupwas made airtight by a rubber ring fitted into a groove around the exposurechamber. Stainless-steel tubes were used for the connections. Each arm of theobservation chamber was connected to a treatment chamber made of polycar-bonate, a washing bottle filled with distilled water, and a flowmeter by whichthe airstream through each arm was separately regulated to 27 ml/min. Theboundaries of the four flow fields were drawn on the cover glass plate. Thirtyseconds after introducing one T. chilonis female in the entry tube, the tube wasdarkened and the parasitoid walked up into the exposure chamber (positivephototactic response). The airflow was established before the test insect enteredthe exposure chamber. Each wasp was observed for 10 min beginning when theparasitoid crossed a "first choice" line, 5 mm around the entry tube. This firstchoice, the total time spent by the parasitoid in each of the four flow fields, andwhether or not a parasitoid entered the arms connected to the flow fields wasrecorded using the computer software package The Observer® (Noldus Infor-mation Technology, 1993). The first flow field chosen by the parasitoid wasrecorded to see if the wasp differentiated between the treatments before exitingthe entry tube.
Plant material was offered in two opposite treatment chambers, leaving theother two as control. Air humidified with wet tissue paper (Kimwipes) was usedas the control, as a preliminary experiment, also described in this paper, showedthat T. chilonis females prefer humidified to unhumidified air. After every twoto three observations, the exposure chamber was cleaned with 70% ethanol toremove traces of previous parasitoids (Salt, 1958). After every five observations,the apparatus was rotated to prevent bias due to environmental factors. A min-imum of 40 wasps were observed for each test material. All experiments wereconducted at ambient conditions of 26 ± 2°C and 50 ± 10% relative humidity.
When the distribution of the parasitoids over the four flow fields was sig-nificant after analysis of variance on angular transformed percentage data, thetime spent in the two test fields was compared to the time spent in the twocontrol fields based on the single degree of freedom contrast using the F test(Steel and Torrie, 1980). The number of parasitoids that spent more than 50%

2458 ROMEIS, SHANOWER, AND ZKBITZ
of the observation time in the test fields as compared to the control fields wascompared by a \2 test (Dicke et al., 1984). The number of test females thatselected the flow fields containing the plant volatiles as their first choice wascompared to the number selecting the control fields with a X2 test. The nullhypothesis of equal distribution was used for both x2 tests.
Preliminary Experiment. As plant material offered in the treatment cham-bers increases the relative humidity of air, a preliminary experiment was con-ducted to test whether the parasitoids show any response to air humidified bywet tissue paper or plant material (20 pigeonpea pods, cv. ICPL 87; ~ 160 cm2
pod surface). Pigeonpea pods plus wet tissue paper were tested against wettissue paper alone to separate the effect of plant volatiles and relative humidity.
Sorghum Tests. Field collected sorghum (cv. CSH 9) leaves and panicleswere tested. Panicles were checked for H. armigera eggs and larvae to ensurethat they had no previous contact with the moth and were covered with paperbags one day prior to collection. The following quantities of each plant partwere offered in a single treatment chamber: leaves, —460 cm2 leaf area; newlyemerged panicle, one fresh exposed panicle of —15 cm length, no flowers;flowering panicle, one panicle of —15 cm length; soft dough panicle, onepanicle with developing seeds of - 15 cm length, no flowers.
Pigeonpea Tests. Plant material was collected from undamaged greenhousegrown plants (cv. ICPL 87), and the following quantities of each plant partwere offered in a single treatment chamber: vegetative tips, six tips of —20 cmlength, -460 cm2 leaf area; reproductive tips, three tips of -20 cm length,with leaves, buds, flowers and pods; buds, flowers, three tips of —20 cm length,with leaves, buds and flowers; and pods, 20 pods, - 160 cm2 pod surface.
With the exception of pigeonpea pods, the plant material was placed in thetreatment chamber with the cut end inserted in a glass vial filled with water andsealed with Parafilm.
RESULTS
The preliminary experiment showed that when offered a choice betweenair with a relatively high versus low humidity, the parasitoids spent a signifi-cantly longer time in the flow fields with the higher humidity (Table 1). Thiswas true whether the source of the humidity was wet tissue paper [Contrast (Q)± SE; Q = 5.0 ± 1.61; F, 244 = 9.6] or plant material (Q = 10.2 ± 2.30;F i , i56 = 19.7). The parasitoids were repelled by volatiles from pigeonpea podswhen offered together with wet tissue paper against humidified air (Q = -8.0± 1.59; FU96 = 25.3).
T. chilonis females were arrested by volatiles from flowering sorghum

Trichogramma PLANT PREFERENCE; 2459
TABLE 1 . RESPONSE OF FEMALE Trichogramma chilonis TO HUMIDITY OR PLANTMATERIAL VERSUS CLEAN, UNHUMIDIFIED OR HUMIDIFIED AIR IN A FOUR-ARMED
OLFACTOMETER
Test material
Wet tissue papervs.unhumidilied air
Pigeonpea podsvs.unhumidified air
Pigeonpea pods +wet tissue papervs.wet tissue paper
N
62
40
50
Time spent inflow fields (%)"
57.0**''
43.0
63.4***
36.6
38.3***
61.7
Longest time inflow fields N"
40*
22
27*
13
11***
39
Firstchoice N'
37
25
23
17
21
29
"Percentage of observation time spent in the test fields as compared to the control fields. Meanswere compared based on the single degree of freedoom contrast.
''Number of parasitoids spending more than 50% of the observation time in the test fields ascompared to the control fields. Numbers were analyzed using \2 test.
'Number of parasitoids crossing the first choice line in the test fields as compared to the controlfields. Numbers were analyzed using x~ test.
''* P < 0.05; ** P < 0.01; *** P < 0.001.
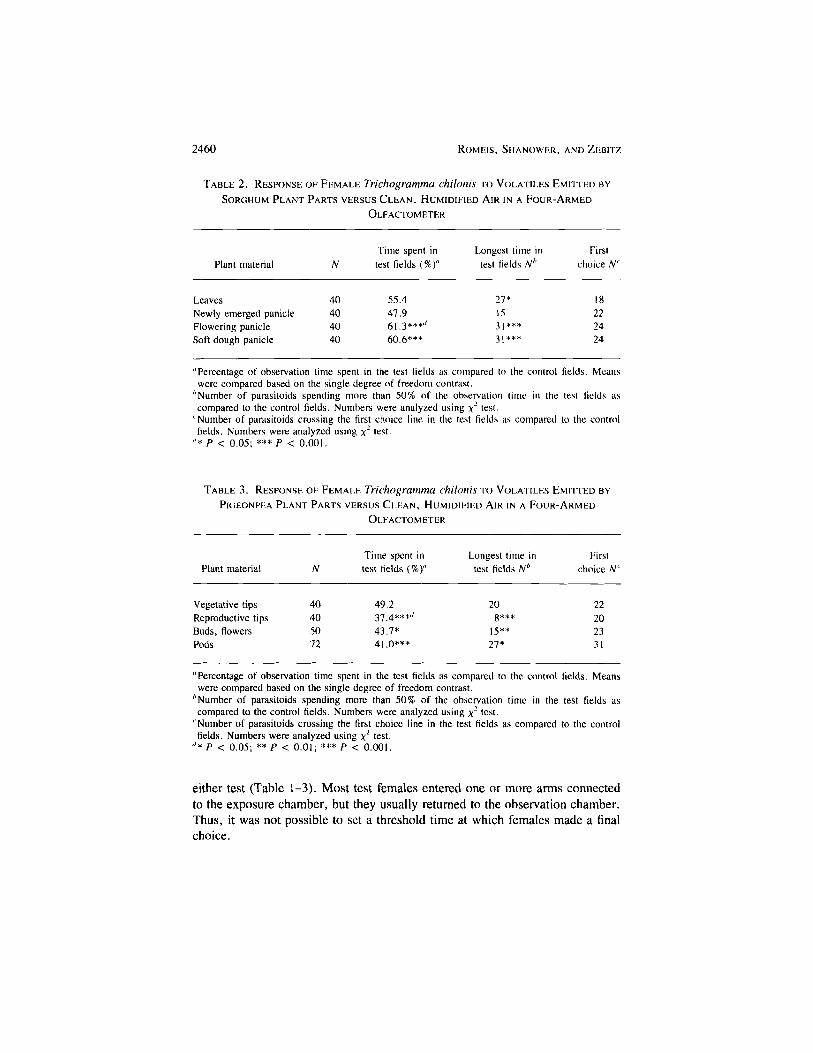
panicles (Q = 8.3 ± 1.95; FVise = 18.0) or panicles in the soft dough stage(2 = 7.6 ± 1.79; FU56 = 18.2) (Table 2). Volatiles from newly emergedpanicles without flowers, however, did not elicit a response from the parasitoids.
Volatiles emitted by pigeonpea plant material in the reproductive growthstage significantly repelled T. chilonis (Table 3). Repellency was observed byvolatiles from tips containing buds, flowers, and pods (Q = -8.4 ± 1.74;FU56 = 23.1), tips containing buds and flowers (Q = -3.9 ± 1.89; FU96 =4.3) and pods alone (Q = -6.5 ± 1.93;F, JM = 11.5). Prior to the reproductivegrowth stage, when only vegetative tissue was present, the wasps showed noresponse.
With the exception of the sorghum leaves (Table 2), the \2 test and thesingle degree of freedom comparisons produced similar results. For volatilesfrom sorghum leaves, a significant arrestment response was detected using theX2 test, while analysis based on the single degree of freedom comparison showedno significance.
No significant difference in the number of first choices was detected in
2460 ROMEIS, SHANOWER, AND ZEBITZ
TABLE 2. RESPONSE OF FEMALE Trichogramma chilonis TO VOLATILES EMITTED BY
SORGHUM PLANT PARTS VERSUS CLEAN, HUMIDIFIED AIR IN A FOUR-ARMEDOLFACTOMETER
Plant material
LeavesNewly emerged panicleFlowering panicleSoft dough panicle
N
40404040
Time spent intest fields (%)"
55.447.961.3***''60.6***
Longest time intest fields N1'
27*1531***31***
Firstchoice Nc
18222424
"Percentage of observation time spent in the test fields as compared to the control fields. Meanswere compared based on the single degree of freedom contrast.
''Number of parasitoids spending more than 50% of the observation time in the lest fields ascompared to the control fields. Numbers were analyzed using x2 test.
'Number of parasitoids crossing the first choice line in the test fields as compared to the controlfields. Numbers were analyzed using x2 test.
''* P < 0.05; *** P < 0.001.
TABLE 3. RESPONSE OF FEMALE Trichogramma chilonis TO VOLATILES EMITTED BY
PIGEONPEA PLANT PARTS VERSUS CLEAN, HUMIDIFIED AIR IN A FOUR-ARMEDOLFACTOMETER
Plant material
Vegetative tipsReproductive tipsBuds, flowersPods
N
40405072
Time spent intest fields (%)"
49.237.4***''43.7*41.0***
Longest time intest fields N1'
208***
15**27*
Firstchoice N'
22202331
"Percentage of observation time spent in the test fields as compared to the control fields. Meanswere compared based on the single degree of freedom contrast.
''Number of parasitoids spending more than 50% of the observation time in the test fields ascompared to the control fields. Numbers were analyzed using x2 test.
'Number of parasitoids crossing the first choice line in the test fields as compared to the controlfields. Numbers were analyzed using x2 test.
''* P < 0.05; ** P < 0.01; *** P < 0.001.
either test (Table 1-3). Most test females entered one or more arms connectedto the exposure chamber, but they usually returned to the observation chamber.Thus, it was not possible to set a threshold time at which females made a finalchoice.

Trichogramma PLANT PREFERENCE 2461
DISCUSSION
Sorghum plant material emitted volatiles that elicited an arrestment responsein T. chilonis. Volatiles from sorghum panicles, the oviposition site forH. armigera (Parsons, 1940), arrested the parasitoids only after the flowers hadopened. The infochemicals were not emitted by the flowers directly, however,as volatiles from panicles in the soft dough stage elicited the same response bythe parasitoids. T. chilonis is known to parasitize insects ovipositing on sorghumleaves such as Chilo partellus (Swinhoe) (Lepidoptera: Pyralidae) (Sharma,1985), and this may explain why the parasitoids were arrested by leaf volatiles.The sorghum volatiles that arrest T. chilonis are unknown. Whether info-chemicals emitted by panicles in the flowering and soft dough stages and thoseemitted by the leaves are the same is also unknown. Sorghum volatiles havebeen shown to be attractive to hymenopteran and dipteran larval parasitoids ofHeliothis/Helicoverpa spp. and stemborers (Schuster and Starks, 1974; Elzenet al., 1983; Martin et al., 1990; Ngi-Song et al., 1996).
The response of T. chilonis to volatiles from pigeonpea was strikinglydifferent. Pigeonpea became repellent to T. chilonis when it had reached thereproductive growth stage, the most attractive stage for H. armigera oviposition.Volatiles emitted by plants in the vegetative growth stage did not mediate thebehavior of searching parasitoids, and it is likely that the infochemicals respon-sible for the repellency of reproductive plants are emitted by the reproductivestructures. This is supported by the finding that volatiles from pods alone wererepellent. However, it is possible that leaves from plants in the reproductivegrowth stage may also emit the repellent infochemicals. Martin et al. (1990)reported that the response of Eucelatoria bryani Sabrosky (Diptera: Tachinidae),a tachinid parasitoid of Heliothis/Helicoverpa spp., to volatiles emitted bypigeonpea leaves varies with the growth stage of the plant. The pigeonpeainfochemicals responsible for the repellency of T. chilonis are unknown andonly a little information on pigeonpea volatiles exists. Six sesquiterpenes—a-bulnesene, a-humulene, a-guajene, /3-caryophyllene, y-muurolene anda-muurolene—are the major components in an aqueous steam distillate ofpigeonpea leaves collected from plants in the flowering growth stage. A mixtureof these compounds was attractive for female H. armigera in a flight tunnel andacted as an oviposition stimulant (Hartlieb and Rembold, 1996). These chemi-cals were emitted by pigeonpea leaves and are probably not responsible for therepellency of T. chilonis. Therefore it may be that different pigeonpea volatilesare involved in the attraction of H. armigera and the repellency of T. chilonis.
Our results show that T. chilonis is arrested by sorghum volatiles andrepelled by volatiles from pigeonpea in the reproductive growth stage, the stagepreferred for oviposition by the host H. armigera. Parasitoids were arrested byvolatiles from pigeonpea pods when offered against unhumidified air; thus, the

2462 ROMEIS, SHANOWER, AND ZEBITZ
attractiveness of humidity may overcome the repellency of pigeonpea. This mayindicate that these infochemicals do not play a major role in the searchingbehavior of T. chilonis in the field and that other plant characters also contributeto the low parasitism levels on pigeonpea. A study to evaluate the impact ofpigeonpea surface structures, i.e., trichomes and trichome exudates, on T. chi-lonis is currently in progress.
Using plant information is not expected to be adaptive for a generalistparasitoid such as Trichogramma spp. attacking a wide range of hosts (Vet andDicke, 1992). However, Trichogramma spp. have been reported to respond tovolatiles from tomato (Nordlund et al., 1985), maize (Kaiser et al., 1989), andcotton (Bar et al., 1979; Cabello and Vargas, 1985), in addition to sorghumand pigeonpea. Leaves from cabbage plants are the only example of plant mate-rial that did not elicit a response from Trichogramma spp. (Pak and van Len-teren, 1984). Therefore, the response of Trichogramma spp. to volatile plantinfochemicals seems to be a common phenomenon. Wind plays an importantrole in the distribution of Trichogramma spp. (Hendricks, 1967; Keller et al.,1985), and because the parasitoids are unable to fly against strong winds (Steen-burgh, 1934), it is likely that plant volatiles repel or arrest, rather than attract,trichogrammatids after entering a habitat. This may explain why in this studyT. chilonis did not walk up the olfactometer arms connected to an attractiveflow field as was reported for larval parasitoids (Dicke et al., 1984; Eller et al.,1988; Turlings et al., 1990). Arrestment rather than attraction of Trichogrammaspp. to host sex pheromones has been reported by Noldus (1988) and Noldusand van Lenteren (1985).
According to the current infochemical terminology (Dicke and Sabelis,1988), the sorghum volatiles responsible for the arrestment of T. chilonis areclassified as synomones as both the plant (emitter) and the parasitoid (receiver)benefit from the interaction. The infochemicals involved in the T. chi-/onw-pigeonpea interaction are most difficult to classify but may be an exampleof an antimone, as neither the plant nor the parasitoid seem to benefit from theinteraction. This term, first proposed by Whitman (1988), has been adapted tothe current infochemical terminology by Takabayashi and Takahashi (1990) (seealso Dicke and Sabelis, 1992).
Antimones have only been detected in a few systems. In two, the interactionwas nonadaptive for the emitter and maladaptive for the parasitoid (receiver)(Takabayashi and Takahashi, 1990; Takabayashi et al., 1991). In contrast tothese examples, T. chilonis-pigeonpea interactions seem to be maladaptive forboth the emitter and the receiver of the infochemicals. A similar interaction hasbeen reported for other Trichogramma species. Cotton is repellent to T. semi-fumatum Riley (Bar et al., 1979), and cotton leaves, but not flowers, are repel-lent to T. sp. p. buesi (Cabello and Vargas, 1985). T. cordubensis Vargas andCabello is not repelled by cotton (Cabello and Vargas, 1985), and it is therefore

Trichogramma PLANT PREFERENCE 2463
likely that the olfactory response varies between Trichogramma species. Dif-ferences in the olfactory response could be partly responsible for the reportedhabitat preference of Trichogramma spp. (Salt, 1935; Flanders, 1937; Lopez etal., 1982; Pinto and Oatman, 1988).
We believe that additional examples of volatile plant infochemicals that arerepellent to parasitoids will be detected in the future. Two reasons may explainwhy they have only rarely been reported: (1) researchers focus on the attractionas opposed to the repellency of parasitoids or predators to specific host plants,and (2) some of the currently used methods, such as wind tunnels, are notdesigned to detect a repellent response to volatiles.
It is surprising that antimones exist as one would expect that communicativeinteractions have evolved by positive selection for transmission as well as recep-tion (Otte, 1974). It remains unclear how such mutually disadvantageous inter-actions could evolve. One possible explanation is that production of or responseto compounds acting as antimones in one context may be beneficial in anothercontext. Most probably antimones will be found in systems where the interac-tants have not closely coevolved. Examples would be systems in which theinteractants are of different origin or generalist parasitoids (such as Tricho-gramma spp.) attacking polyphagous hosts (such as Heliothis/Helicoverpa spp.).Antimones might therefore be a limiting factor for the use of 'new associations'in biological control.
Acknowledgments—We thank S. Chandra and K. Rukmini Devi (ICRISAT, Statistics Unit)for statistical advice and M. Dicke, Department of Entomology, Wageningen Agricultural Univer-sity, for valuable comments on an earlier draft of the manuscript. We thank M. Gupta of the CentralIntegrated Pest Management Centre, Hyderabad, India, tor the T. chilonis strain and continuoussupply of C. cephalonica egg cards. Support to J.R. by Deutsche Gesellschaft fur TechnischeZusammenarbeit (GTZ) is gratefully acknowledged. This paper was approved as Journal ArticleNo. 1997 by the International Crops Research Institute tor the Semi-Arid Tropics (ICRISAT).
REFERENCES
BAR, D., GERLING, D., and ROSSLER, Y. 1979. Bionomics of the principal natural enemies attackingHeliothis armigera in cottom fields in Israel. Environ. Entomol. 8:468-474.
CABELLO, T., and VARGAS, P. 1985. Estudio con olfactometro de la influencia de la planta y delinsecto huesped en la actividad de busqueda de Trichogramma cordubensis Vargas y Cabelloy de T. sp. p. buesi (Hym., Trichogrammatidae). Bol. Serv. Plagas 11:237-241.
DICKE, M., and SABELIS, M. W. 1988. Infochemical terminology: Based on cost-benefit analysisrather than origin of compounds. Fund. Ecol. 2:131-139.
DICKE, M., and SABELIS, M. W. 1992. Costs and benefits of chemical information conveyance:proximate and ultimate factors, pp. 122-155, in B. D. Roitberg, and M. B. Isman (eds.).Insect Chemical Ecology. An Evolutionary Approach. Chapman & Hall, New York.
DICKE, M., VAN LENTEREN, J. C., BOSKAMP, G. J. F., and VAN DONGEN-VAN LEEUWRN, E. 1984.Chemical stimuli in host-habitat location by Leptopilina heierotoma (Thomson) (Hymenoptera:Eucoilidae), a parasite of Drosophila. J. Chem. Ecol. 10:695-712.

2464 ROMEIS, SHANOWER, AND ZEBITz
ELLER, F. J., TUMLINSON, J. H., and LEWIS, W. J. 1988. Beneficial arthropod behavior mediatedby airborne semiochemicals. II. Olfactometric studies of host location by the parasitoid Micro-plitis croceipes (Cresson) (Hymenoptera: Braconidae). J. Chem. Ecol. 14:425-434.
ELZEN, G. W., WILLIAMS, H. J., and VINSON, S. B. 1983. Response of Campoletis sonorensis(Hymenoptera: Ichneumonidae) to chemicals (synomones) in plants: Implications for host hab-itat location. Environ. Entomol. 12:1873-1877.
FLANDERS, S. E. 1937. Habitat selection by Trichogramma. Ann. Entomol. Soc: Am. 30:208-210.HARTLIEB, E., and REMBOLD, H. 1996. Behavioral response of female Helicoverpa (Heliothis)
armigera Hb. (Lepidoptera: Noctuidae) moths to synthetic pigeonpea (Cajanus cajan L.) kai-romone. J. Chem. Ecol. 22:821-837.
HeNDRICKS, D. E. 1967. Effect of wind on dispersal of Trichogramma semifumatum. J. Econ.Entomol. 60:1367-1373.
KAISER, L., PHAM-DELEGUE, M. H., BAKCHINE, E., and MASSON, C. 1989. Olfactory responses ofTrichogramma maidis Pint, et Voeg.: Effects of chemical cues and behavioral plasticity. J.Insect Behav. 2:701-712.
KELLER, M. A., LEWIS, W. J., and STINNER, R. E. 1985. Biological and practical significance ofmovement by Trichogramma species: A review. Southwest. Entomol. Suppl. 8:138-155.
LATEEF, S. S., and REED, W. 1990. Insect pests of pigeonpea, pp. 193-242, in S. R. Singh (ed.).Insect Pests of Tropical Food Legumes. John Wiley & Sons, New York.
LOPEZ, J. D., JONES, S. L., and HOUSE, V. S. 1982. Species of Trichogramma parasitizing eggsof Heliothis spp. and some associated lepidopterous pests in central texas. Southwest Entomol.7:87-93.
MARTIN, W. R., JR., NORDLUND, D. A., and NETTLES, W. C., JR. 1990. Response of parasitoidEucelatoria bryani to selected plant material in an olfactometer. J. Chem. Ecol. 16:499-508.
NENE, Y. L., and SHEILA, V. 1990. Geography and importance, pp. 1-14, in Y. L. Nene, S. D.Hall, and V.K. Sheila (eds.). The Pigeonpea. CAB International, Wallingford.
NGI-SONG, A. J., OVERHOLT, W. A., NJAGl, P. G. N., DICKE, M., AYeRTEY, J. N., and LWANDE,W. 1996. Volatile infochemicals used in host and host habitat location by Cotesia flavipesCameron and Cotesia sesamiae (Cameron) (Hymenoptera: Braconidae), larval parasitoids ofstemborers on graminae. J. Chem. Ecol. 22:307-323.
NOLDUS, L. P. J. J. 1988. Response of the egg parasitoid Trichogramma pretiosum to the sexpheromone of its host Heliothis zea. Entomol Exp. Appl. 48:293-300.
NOLDUS, L. P. J. J. 1989. Semiochemicals, foraging behaviour and quality of entomophagousinsects for biological control. J. Appl. Entomol. 108:425-451.
NOLDUS, L. P. J. J., and VAN L e N T E R E N , J. C. 1985. Kairomones for the egg parasite Trichogrammaevanescens Westwood. 1. Effect of volatile substances released by two of its hosts, Pierisbrassicae L. and Mamestra brassicae, L. J. Chem. Ecol. 11:781-791.
NOLDUS INFORMATION TECHNOLOGY. 1993. The Observer, Base Package for DOS. Reference Man-ual, Version 3.0. Wageningen, The Netherlands.
NORDLUND, D. A., CHALFANT, R. B., and LEWIS, W. J. 1985. Response of Trichogramma pre-tiosum females to volatile synomones from tomato plants. J. Entomol. Sci. 20:372-376.
OTTE, D. 1974. Effects and functions in the evolution of signalling systems. Annu. Rev. Ecol. Syst.5:385-417.
PAK, G. A., and VAN LENTEREN, J. C. 1984. Selection of a candidate Trichogramma sp. strain forinundative releases against lepidopterous pests of cabbage in the Netherlands. Med. Fac. Land-bouww. Rijksuniv. Gent 49(3a):827-837.
PARSONS, F. S. 1940. Investigations on the cotton bollworm, Heliothis armigera Hubn. Part III.Relationship between oviposition and the flowering curves of food-plants. Bull. Eitlomol. Res.31:147-177.
PINTO, J. D., and OATMAN, E. R. 1988. Trichogramma species in a chaparral community of southern

Trichogramma PLANT P R E f e r e N C E 2465
California, with a description of a new species (Hymenoptera: Trichogrammatidae). Pan-Pacific Entomol. 64:391-402.
ROMEIS, J., and SHANOWeR, T. G. 1996. Arthropod natural enemies of Helicoverpa armigera(Hubner) (Lepidoptera: Noctuidae) in India. Biocontrol Sci. Tcchnol. 6:481-508.
SALT, G. 1935. Experimental studies on insect parasitism. III. Host selection. Proc. R. Soc., London117:413-435.
SALT, G. 1958. Parasite behaviour and the control of insect pests. Endeavour 17:145-148.SCHUSTeR, D. J., and STARKS, K. J. 1974. Response of Lysiphlebus testaceipes in an olfactometer
to a host and a non-host insect and to plants. Environ. Entomol. 3:1034-1035.SHARMA, H. C. 1985. Strategies for pest control in sorghum in India. Trop. Pest Manage.
31:167-185.STEEL, R. G. D., and TORRIE, J. H. 1980. Principles and Procedures of Statistics, a Biometrical
Approach, 2nd ed. McGraw-Hill, Auckland.STEENBURGH, W. E. V A N . 1934. Trichogramma minutum Riley as a parasite of the Oriental fruit
moth (Laspeyresia molesta Busck) in Ontario. Can. J. Res. 10:287-314.TAKABAYASHI, J., NODA, T., and T A K A H A S H I , S. 1991. Plants produce attractants for Apantelex
kariyai, a parasitoid of Pseudaletia separata; cases of "communication" and "misunderstand-ing" in parasitoid-plant interactions. Appl. Entomol. Zool. 26:237-243.
TAKABAYASHI, J., and TAKAHASHI , S. 1990. An allelochemical elicits arrestment in Apanteles kariyaiin feces of nonhost larvae Acantholeucania loreyi. J. Chem. Ecol. 16:2009-2017.
TUMLINSON, J. H., TURLINgS, T. C. J., and LEWIS, W. J. 1992. The semiochemical complexesthat mediate insect parasitoid foraging. Agric. Zool. Rev. 5:221-252.
TURLINGS, T. C. J., SCHEEPMAKER, J. W. A., VET, L. E. M., TUMLINSON, J. H., and LEWIS,W. J. 1990. How contact foraging experiences affect preferences for host-related odors in thelarval parasitoid Cotesia marginiventris (Cresson) (Hymenoptera: Braconidae). J. Chem. Ecol.16:1577-1589.
VET, L. E. M., and DICKE, M. 1992. Ecology of infochemical use by natural enemies in a tritrophiccontext. Annu. Rev. Entomol. 37:141-172.
VET, L. E. M., WACKERS, F. L., and DICKE, M. 1991. How to hunt for hiding hosts: Thereliability-detectability problem in foraging parasitoids. Neth. J. Zool. 41:202-213.
WHITMAN, D. W. 1988. Allelochemical interaction among plants, herbivores and their predators,pp. 11-64, in P. Barbosa, and D. Letoumeau (eds.). Novel Aspects of Insect-Plant Interactions.John Wiley & Sons, New York.
Copyright © 2022 FDOKUMEN




















