
ATP Hydrolysis by the Proteasome Regulatory Complex PAN ...
10
Molecular Cell, Vol. 11, 69–78, January, 2003, Copyright 2003 by Cell Press ATP Hydrolysis by the Proteasome Regulatory Complex PAN Serves Multiple Functions in Protein Degradation et al., 2000; Kim et al., 2000; Navon and Goldberg, 2001; Singh et al., 2000; Weber-Ban et al., 1999). The amount of ATP consumed and the specific functions of ATP hydrolysis are particularly difficult to analyze with 26S proteasomes due to the requirement for ubiquitin conju- Nadia Benaroudj, 1,3 Peter Zwickl, 2 Erika Seemu ¨ ller, 2 Wolfgang Baumeister, 2 and Alfred L. Goldberg 1,* 1 Harvard Medical School Department of Cell Biology 240 Longwood Avenue gation to most substrates, the low intrinsic stability of Boston, Massachusetts 02115 the 19S and its complexity (it contains at least 17 distinct 2 Max-Planck-Institute for Biochemistry subunits), and the presence of six different ATPases Department of Molecular Structural Biology that probably serve distinct functions (Glickman et al., Am Klopferspitz 18a 1998; Holzl et al., 2000; Rubin et al., 1998). D-82152 Martinsried Although prokaryotes lack ubiquitin and 26S protea- Germany somes, archaebacteria and certain eubacteria contain 20S proteasomes whose structural organization and catalytic mechanism are very similar to those of the Summary eukaryotic 20S particles (Baumeister et al., 1998; De Mot et al., 1999; Zwickl et al., 2000). Consequently, studies of To clarify the role of ATP in proteolysis, we studied archaeal proteasomes have provided important insights archaeal 20S proteasomes and the PAN (proteasome- into the structure and enzymatic mechanisms of their activating nucleotidase) regulatory complex, a homo- eukaryotic homologs. 20S particles have a barrel-shaped log of the eukaryotic 19S ATPases. PAN’s ATPase ac- structure composed of two outer rings and two central tivity was stimulated similarly by globular (GFPssrA) rings, which contain the proteolytic active sites se- and unfolded (casein) substrates, and by the ssrA rec- questered within a central chamber (Lowe et al., 1995; ognition peptide. Denaturation of GFPssrA did not ac- Wenzel and Baumeister, 1995). Because of the tight celerate its degradation or eliminate the requirement packing of subunits, substrates enter through a narrow for PAN and ATP. During degradation of one molecule channel in the ring which only permits entrance of of globular or unfolded substrates, 300–400 ATP mole- unfolded polypeptide chains or short peptides (Lowe et cules were hydrolyzed. An N-terminal deletion in the al., 1995). In the native form of yeast 20S particles, this 20S subunits caused opening of the substrate-entry channel is blocked by the amino-terminal extremities of channel and rapid degradation of unfolded proteins the subunits (Groll et al., 1997). As a consequence, without PAN; however, degradation of globular GFPssrA eukaryotic 20S particles exhibit low peptidase activity still required PAN’s ATPase activity, even after because substrates cannot readily enter the internal PAN-catalyzed unfolding. Thus, substrate binding ac- chamber. Deletion of the N termini of the 3 subunit tivates ATP hydrolysis, which promotes three pro- (residues 2–10) in yeast 20S prevents closing of the cesses: substrate unfolding, gate opening in the 20S, channel and greatly enhances peptide hydrolysis (Groll and protein translocation. et al., 2000). Recent studies have suggested that the 19S ATPase Rpt2 is particularly important in opening Introduction the axial pore (Kohler et al., 2001). In contrast to the eukaryotic particles, the X-ray diffraction analysis of the In all cells, degradation of intracellular proteins requires 20S particles from Thermoplasma acidophilum did not ATP hydrolysis (Goldberg, 1992). In eukaryotes, ATP reveal a closed channel, presumably because the amino- hydrolysis is necessary for ubiquitination of protein sub- terminal extremities of the subunits are disordered strates, as well as for the proteolytic activity of 26S and therefore not visible (Lowe et al., 1995). Moreover, proteasomes, the principal site of proteolysis in the cyto- these particles by themselves appear fully active in deg- sol and nucleus (Coux et al., 1996). 26S proteasomes radation of fluorogenic tetrapeptides, but not of pro- contain six homologous ATPase subunits in their 19S teins. Therefore, it has been generally assumed that regulatory particles. The precise roles of those ATPases the channel formed by the rings of the archaeal 20S in proteasome function, as well as the amount of ATP particles is in an open state (Groll et al., 2000; Kohler et consumed during proteolysis, are uncertain. It has been al., 2001; Lowe et al., 1995). proposed that the ATPases catalyze unfolding of protein We have identified a 650 kDa ATPase complex from the substrates and promote their translocation into the cen- archaebacterium Methanococcus jannaschii named PAN, tral chamber of 20S proteasomes where proteolysis oc- which is probably the evolutionary precursor of the eukary- curs (Lupas et al., 1993). More recently, it has been otic 19S (Zwickl et al., 1999). It is composed of identical shown that bacterial ATP-dependent proteases (ClpAP subunits closely homologous to the six ATPases of the and ClpXP) and the PAN (proteasome-activating nucleo- 19S and stimulates markedly the degradation of pro- tidase) complex (the archaeal 19S homolog) unfold glob- teins, but not of tetrapeptides, by archaeal 20S protea- ular proteins (Benaroudj and Goldberg, 2000; Hoskins somes. This process requires hydrolysis of ATP (Zwickl et al., 1999), although the mechanisms linking ATP hy- drolysis and protein degradation are unknown. Because *Correspondence: [email protected] of its simple structural organization, lack of requirement 3 Present address: Institut Pasteur, Unite ´ de Repliement et Mode ´ lisa- tion des Prote ´ ines, 25-28 rue du Dr Roux, 75015 Paris, France. for substrate ubiquitination, and ease of expression in
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of ATP Hydrolysis by the Proteasome Regulatory Complex PAN ...

Molecular Cell, Vol. 11, 69–78, January, 2003, Copyright 2003 by Cell Press
ATP Hydrolysis by the ProteasomeRegulatory Complex PAN Serves MultipleFunctions in Protein Degradation
et al., 2000; Kim et al., 2000; Navon and Goldberg, 2001;Singh et al., 2000; Weber-Ban et al., 1999). The amountof ATP consumed and the specific functions of ATPhydrolysis are particularly difficult to analyze with 26Sproteasomes due to the requirement for ubiquitin conju-
Nadia Benaroudj,1,3 Peter Zwickl,2 Erika Seemuller,2
Wolfgang Baumeister,2 and Alfred L. Goldberg1,*
1Harvard Medical SchoolDepartment of Cell Biology240 Longwood Avenue
gation to most substrates, the low intrinsic stability ofBoston, Massachusetts 02115the 19S and its complexity (it contains at least 17 distinct2 Max-Planck-Institute for Biochemistrysubunits), and the presence of six different ATPasesDepartment of Molecular Structural Biologythat probably serve distinct functions (Glickman et al.,Am Klopferspitz 18a1998; Holzl et al., 2000; Rubin et al., 1998).D-82152 Martinsried
Although prokaryotes lack ubiquitin and 26S protea-Germanysomes, archaebacteria and certain eubacteria contain20S proteasomes whose structural organization andcatalytic mechanism are very similar to those of theSummaryeukaryotic 20S particles (Baumeister et al., 1998; De Motet al., 1999; Zwickl et al., 2000). Consequently, studies ofTo clarify the role of ATP in proteolysis, we studiedarchaeal proteasomes have provided important insightsarchaeal 20S proteasomes and the PAN (proteasome-into the structure and enzymatic mechanisms of theiractivating nucleotidase) regulatory complex, a homo-eukaryotic homologs. 20S particles have a barrel-shapedlog of the eukaryotic 19S ATPases. PAN’s ATPase ac-structure composed of two outer � rings and two centraltivity was stimulated similarly by globular (GFPssrA)� rings, which contain the proteolytic active sites se-and unfolded (casein) substrates, and by the ssrA rec-questered within a central chamber (Lowe et al., 1995;ognition peptide. Denaturation of GFPssrA did not ac-Wenzel and Baumeister, 1995). Because of the tightcelerate its degradation or eliminate the requirementpacking of subunits, substrates enter through a narrowfor PAN and ATP. During degradation of one moleculechannel in the � ring which only permits entrance ofof globular or unfolded substrates, 300–400 ATP mole-unfolded polypeptide chains or short peptides (Lowe etcules were hydrolyzed. An N-terminal deletion in theal., 1995). In the native form of yeast 20S particles, this20S � subunits caused opening of the substrate-entrychannel is blocked by the amino-terminal extremities ofchannel and rapid degradation of unfolded proteinsthe � subunits (Groll et al., 1997). As a consequence,without PAN; however, degradation of globular GFPssrAeukaryotic 20S particles exhibit low peptidase activitystill required PAN’s ATPase activity, even afterbecause substrates cannot readily enter the internalPAN-catalyzed unfolding. Thus, substrate binding ac-chamber. Deletion of the N termini of the �3 subunittivates ATP hydrolysis, which promotes three pro-(residues 2–10) in yeast 20S prevents closing of thecesses: substrate unfolding, gate opening in the 20S,channel and greatly enhances peptide hydrolysis (Grolland protein translocation.et al., 2000). Recent studies have suggested that the19S ATPase Rpt2 is particularly important in opening
Introduction the axial pore (Kohler et al., 2001). In contrast to theeukaryotic particles, the X-ray diffraction analysis of the
In all cells, degradation of intracellular proteins requires 20S particles from Thermoplasma acidophilum did notATP hydrolysis (Goldberg, 1992). In eukaryotes, ATP reveal a closed channel, presumably because the amino-hydrolysis is necessary for ubiquitination of protein sub- terminal extremities of the � subunits are disorderedstrates, as well as for the proteolytic activity of 26S and therefore not visible (Lowe et al., 1995). Moreover,proteasomes, the principal site of proteolysis in the cyto- these particles by themselves appear fully active in deg-sol and nucleus (Coux et al., 1996). 26S proteasomes radation of fluorogenic tetrapeptides, but not of pro-contain six homologous ATPase subunits in their 19S teins. Therefore, it has been generally assumed thatregulatory particles. The precise roles of those ATPases the channel formed by the � rings of the archaeal 20Sin proteasome function, as well as the amount of ATP particles is in an open state (Groll et al., 2000; Kohler etconsumed during proteolysis, are uncertain. It has been al., 2001; Lowe et al., 1995).proposed that the ATPases catalyze unfolding of protein We have identified a 650 kDa ATPase complex from thesubstrates and promote their translocation into the cen- archaebacterium Methanococcus jannaschii named PAN,tral chamber of 20S proteasomes where proteolysis oc- which is probably the evolutionary precursor of the eukary-curs (Lupas et al., 1993). More recently, it has been otic 19S (Zwickl et al., 1999). It is composed of identicalshown that bacterial ATP-dependent proteases (ClpAP subunits closely homologous to the six ATPases of theand ClpXP) and the PAN (proteasome-activating nucleo- 19S and stimulates markedly the degradation of pro-tidase) complex (the archaeal 19S homolog) unfold glob- teins, but not of tetrapeptides, by archaeal 20S protea-ular proteins (Benaroudj and Goldberg, 2000; Hoskins somes. This process requires hydrolysis of ATP (Zwickl
et al., 1999), although the mechanisms linking ATP hy-drolysis and protein degradation are unknown. Because*Correspondence: [email protected] its simple structural organization, lack of requirement3 Present address: Institut Pasteur, Unite de Repliement et Modelisa-
tion des Proteines, 25-28 rue du Dr Roux, 75015 Paris, France. for substrate ubiquitination, and ease of expression in

Molecular Cell70
E. coli, PAN offers many advantages for mechanistic molar excess of 20S proteasomes was added so thatall PAN should be bound to proteasomes, the basal andstudies. PAN exhibits certain activities characteristic of
molecular chaperones, e.g., it prevents aggregation and the substrate-stimulated ATPase activity of PAN werenot significantly affected (Figure 1B). Thus, the protein-promotes the refolding of denatured proteins (Benaroudj
and Goldberg, 2000). In addition, in the presence of activated PAN’s ATPase activity is not increased furtherwhen PAN binds to 20S and when the substrate is trans-ATP, PAN catalyzes the unfolding of the globular green
fluorescent protein when its C terminus is fused to the located into the proteolytic chamber and degraded.11 residue ssrA peptide (GFPssrA) (Benaroudj and Gold-berg, 2000). This ATP-dependent “unfoldase” activity is Peptide Binding to PAN Stimulates Its ATPase Activityessential for the degradation of GFPssrA by archaeal Native GFP lacking the ssrA peptide at its C terminus is20S proteasomes, which otherwise do not degrade not unfolded or degraded by PAN and 20S proteasomesGFPssrA. Like ATP-dependent proteases in E. coli and (Benaroudj and Goldberg, 2000), and did not stimulateother molecular chaperones, PAN’s ATPase activity is ATP hydrolysis by PAN, presumably because it does notstimulated by protein substrates (our unpublished data), interact with PAN. Thus, the binding of PAN to GFPssrAbut the mechanism of this substrate-activated ATP hy- seems to depend on the 11 residue ssrA extension. Wedrolysis and its role in translocation and degradation of first tested whether this peptide could interact with PANproteins remain unclear. and inhibit its ability to unfold GFPssrA. Upon transfer
In the present studies, we have used the PAN complex of GFPssrA to 45�C, there was an immediate small dropto analyze further the coupling between ATP hydrolysis, in its fluorescence signal (Figure 1C, closed circle),substrate unfolding, and translocation. We have investi- which by 5 min reached a new steady state correspond-gated how different types of protein substrates influence ing to GFPssrA fluorescence at 45�C. This decreaseATP hydrolysis by PAN and the actual amount of ATP reflected the effects of high temperature on the quantumconsumed during hydrolysis of globular and denatured efficiency of fluorescence and does not indicate anyproteins. In order to clarify the process of protein entry unfolding of GFP, which is a very thermostable proteininto 20S particles, we have investigated whether arch- (Tm � 65�C) (Bokman and Ward, 1981). However, uponaeal 20S proteasomes, like eukaryotic 20S, have a func- addition of PAN (Figure 1C, closed triangle), there wastional gate that precludes protein entry. We have ana- a large further decrease in fluorescence, indicating anlyzed the degradation of different proteins by a 20S ATP-dependent unfolding of GFPssrA catalyzed by PANproteasome deletion mutant ��(2-12)20S that lacks the (Benaroudj and Goldberg, 2000). When the ssrA peptideresidues of � subunits which in eukaryotic 20S function was added to the reaction containing GFPssrA-PAN andas a gate for the channel. This study shows that one ATP, the fall was prevented. Thus, the ssrA peptidefunction of PAN and ATP is to open the gate in the � inhibited the unfolding of GFPssrA by PAN, presumablyring, and that ATP hydrolysis also serves an additional because this peptide binds to the same site on PAN asrole in facilitating the entry of substrates after their un- GFPssrA. Interestingly, the ssrA peptide also inhibitedfolding. the PAN-stimulated degradation of casein by 20S parti-
cles (Figure 1D), suggesting that these two very differentprotein substrates bind to the same site on PAN.Results
We then tested whether the ssrA peptide could stimu-late PAN’s ATPase activity. By itself, the ssrA peptideGlobular and Unfolded Substrates Stimulate PAN’sstimulated PAN’s ATPase activity to a comparable ex-ATPase Activity to Similar Extentstent as GFPssrA or casein when used at the same con-Like the ATPase component of bacterial ATP-dependentcentration (Figures 1E). Together, these findings indicateproteases (lon, ClpA, and HslU) (Hwang et al., 1988;that the binding of a peptide to PAN is sufficient toMenon et al., 1987; Seol et al., 1997), PAN’s ATPasestimulate its ATPase activity. It is very likely that sub-activity is stimulated by protein substrates (our unpub-strate binding induces a change in the conformationlished data). In order to understand the mechanism ofof PAN to a form that has a higher maximal ability tothis activation, we compared the rates of ATP hydrolysishydrolyze ATP.by PAN in the presence of casein, which contains little
secondary or tertiary structures (Creamer et al., 1981),and GFPssrA, a globular protein that is unfolded by PAN Substrate Denaturation Does Not Eliminate the
Requirement for PAN and ATP in Degradation(Benaroudj and Goldberg, 2000). These experimentswere performed with a 1000-fold molar excess of these by 20S Proteasomes
Archaeal 20S proteasomes by themselves can degradesubstrates, which causes maximal stimulation of PAN’sATPase activity (data not shown). Casein stimulated the certain unfolded proteins, e.g., the fully reduced �-lact-
albumin (Wenzel and Baumeister, 1995). However, rapidATPase activity of PAN 5-fold. Surprisingly, PAN’sATPase activity was stimulated by GFPssrA to a similar degradation of even the loosely folded casein, like the
breakdown of the globular GFPssrA, required PAN’sextent as casein (Figure 1A). Therefore, the maximalstimulation of ATP hydrolysis is the same whether or not ATPase activity (Benaroudj and Goldberg, 2000; Zwickl
et al., 1999). To further investigate the influence of thethe substrate is globular. Prior observations suggestedthat only proteolytic substrates of the PAN-20S complex substrate’s folding status, we tested whether denatur-
ation of GFPssrA would make it competent for degrada-enhance ATP hydrolysis (our unpublished data). Accord-ingly, native GFP (i.e., lacking the ssrA peptide), which tion by 20S independently of PAN and ATP. GFPssrA
was denatured by treatment with acid, which causesis not unfolded by PAN (Benaroudj and Goldberg, 2000),did not stimulate PAN’s ATPase activity. When a 5-fold a complete loss of fluorescence, modification of the
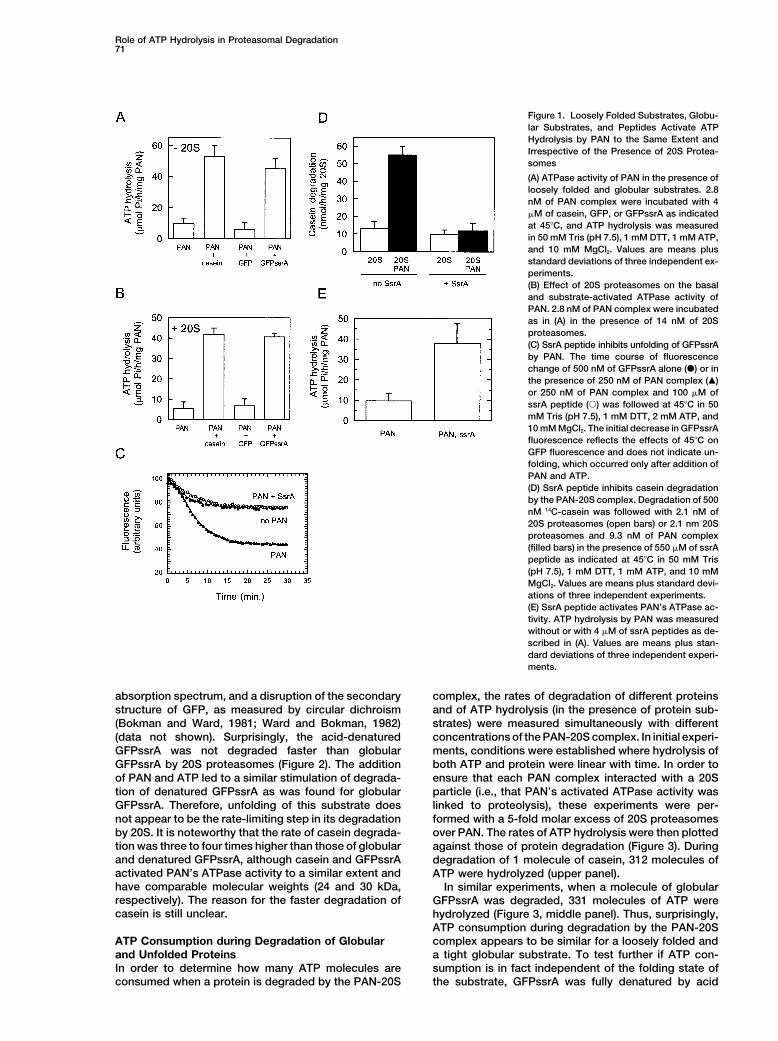
Role of ATP Hydrolysis in Proteasomal Degradation71
Figure 1. Loosely Folded Substrates, Globu-lar Substrates, and Peptides Activate ATPHydrolysis by PAN to the Same Extent andIrrespective of the Presence of 20S Protea-somes
(A) ATPase activity of PAN in the presence ofloosely folded and globular substrates. 2.8nM of PAN complex were incubated with 4�M of casein, GFP, or GFPssrA as indicatedat 45�C, and ATP hydrolysis was measuredin 50 mM Tris (pH 7.5), 1 mM DTT, 1 mM ATP,and 10 mM MgCl2. Values are means plusstandard deviations of three independent ex-periments.(B) Effect of 20S proteasomes on the basaland substrate-activated ATPase activity ofPAN. 2.8 nM of PAN complex were incubatedas in (A) in the presence of 14 nM of 20Sproteasomes.(C) SsrA peptide inhibits unfolding of GFPssrAby PAN. The time course of fluorescencechange of 500 nM of GFPssrA alone (�) or inthe presence of 250 nM of PAN complex (�)or 250 nM of PAN complex and 100 �M ofssrA peptide (�) was followed at 45�C in 50mM Tris (pH 7.5), 1 mM DTT, 2 mM ATP, and10 mM MgCl2. The initial decrease in GFPssrAfluorescence reflects the effects of 45�C onGFP fluorescence and does not indicate un-folding, which occurred only after addition ofPAN and ATP.(D) SsrA peptide inhibits casein degradationby the PAN-20S complex. Degradation of 500nM 14C-casein was followed with 2.1 nM of20S proteasomes (open bars) or 2.1 nm 20Sproteasomes and 9.3 nM of PAN complex(filled bars) in the presence of 550 �M of ssrApeptide as indicated at 45�C in 50 mM Tris(pH 7.5), 1 mM DTT, 1 mM ATP, and 10 mMMgCl2. Values are means plus standard devi-ations of three independent experiments.(E) SsrA peptide activates PAN’s ATPase ac-tivity. ATP hydrolysis by PAN was measuredwithout or with 4 �M of ssrA peptides as de-scribed in (A). Values are means plus stan-dard deviations of three independent experi-ments.
absorption spectrum, and a disruption of the secondary complex, the rates of degradation of different proteinsand of ATP hydrolysis (in the presence of protein sub-structure of GFP, as measured by circular dichroism
(Bokman and Ward, 1981; Ward and Bokman, 1982) strates) were measured simultaneously with differentconcentrations of the PAN-20S complex. In initial experi-(data not shown). Surprisingly, the acid-denatured
GFPssrA was not degraded faster than globular ments, conditions were established where hydrolysis ofboth ATP and protein were linear with time. In order toGFPssrA by 20S proteasomes (Figure 2). The addition
of PAN and ATP led to a similar stimulation of degrada- ensure that each PAN complex interacted with a 20Sparticle (i.e., that PAN’s activated ATPase activity wastion of denatured GFPssrA as was found for globular
GFPssrA. Therefore, unfolding of this substrate does linked to proteolysis), these experiments were per-formed with a 5-fold molar excess of 20S proteasomesnot appear to be the rate-limiting step in its degradation
by 20S. It is noteworthy that the rate of casein degrada- over PAN. The rates of ATP hydrolysis were then plottedtion was three to four times higher than those of globular against those of protein degradation (Figure 3). Duringand denatured GFPssrA, although casein and GFPssrA degradation of 1 molecule of casein, 312 molecules ofactivated PAN’s ATPase activity to a similar extent and ATP were hydrolyzed (upper panel).have comparable molecular weights (24 and 30 kDa, In similar experiments, when a molecule of globularrespectively). The reason for the faster degradation of GFPssrA was degraded, 331 molecules of ATP werecasein is still unclear. hydrolyzed (Figure 3, middle panel). Thus, surprisingly,
ATP consumption during degradation by the PAN-20Scomplex appears to be similar for a loosely folded andATP Consumption during Degradation of Globular
and Unfolded Proteins a tight globular substrate. To test further if ATP con-sumption is in fact independent of the folding state ofIn order to determine how many ATP molecules are
consumed when a protein is degraded by the PAN-20S the substrate, GFPssrA was fully denatured by acid
Molecular Cell72
Figure 2. PAN Stimulates the Degradation of Globular and UnfoldedProteins by 20S Proteasomes
The degradation of 1 �M of 14C-GFPssrA, acid-denatured 14C-GFPssrA,or 14C-casein was followed with 2.1 nM of 20S proteasomes alone(open bars), with 9.3 nM of PAN complex added (striped bars), orwith 9.3 nM of PAN complex and 1 mM ATP added (filled bars) at45�C in 50 mM Tris (pH 7.5), 1 mM DTT, and 10 mM MgCl2. Values aremeans plus standard deviations of three independent experiments.
treatment just before incubation with PAN and 20S pro-teasomes. Three hundred and seventy three moleculesof ATP were hydrolyzed during the degradation of amolecule of acid-denatured GFPssrA (Figure 3, lowerpanel). Together, these findings indicate that a largenumber of ATP molecules (at least 300) are hydrolyzedduring degradation of a protein substrate, regardless ofits folding status. These findings are consistent with ourobservation that folded and unfolded protein substrates
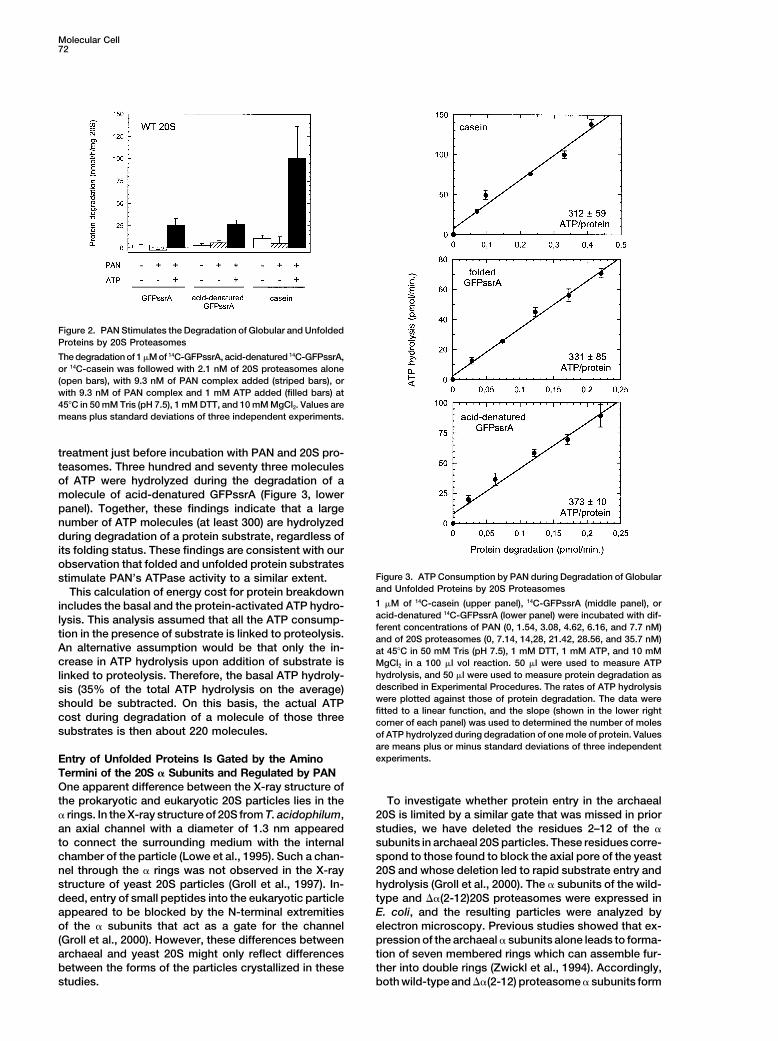
Figure 3. ATP Consumption by PAN during Degradation of Globularstimulate PAN’s ATPase activity to a similar extent.and Unfolded Proteins by 20S ProteasomesThis calculation of energy cost for protein breakdown1 �M of 14C-casein (upper panel), 14C-GFPssrA (middle panel), orincludes the basal and the protein-activated ATP hydro-acid-denatured 14C-GFPssrA (lower panel) were incubated with dif-lysis. This analysis assumed that all the ATP consump-ferent concentrations of PAN (0, 1.54, 3.08, 4.62, 6.16, and 7.7 nM)
tion in the presence of substrate is linked to proteolysis. and of 20S proteasomes (0, 7.14, 14,28, 21.42, 28.56, and 35.7 nM)An alternative assumption would be that only the in- at 45�C in 50 mM Tris (pH 7.5), 1 mM DTT, 1 mM ATP, and 10 mMcrease in ATP hydrolysis upon addition of substrate is MgCl2 in a 100 �l vol reaction. 50 �l were used to measure ATP
hydrolysis, and 50 �l were used to measure protein degradation aslinked to proteolysis. Therefore, the basal ATP hydroly-described in Experimental Procedures. The rates of ATP hydrolysissis (35% of the total ATP hydrolysis on the average)were plotted against those of protein degradation. The data wereshould be subtracted. On this basis, the actual ATPfitted to a linear function, and the slope (shown in the lower right
cost during degradation of a molecule of those three corner of each panel) was used to determined the number of molessubstrates is then about 220 molecules. of ATP hydrolyzed during degradation of one mole of protein. Values
are means plus or minus standard deviations of three independentexperiments.Entry of Unfolded Proteins Is Gated by the Amino
Termini of the 20S � Subunits and Regulated by PANOne apparent difference between the X-ray structure ofthe prokaryotic and eukaryotic 20S particles lies in the To investigate whether protein entry in the archaeal
20S is limited by a similar gate that was missed in prior� rings. In the X-ray structure of 20S from T. acidophilum,an axial channel with a diameter of 1.3 nm appeared studies, we have deleted the residues 2–12 of the �
subunits in archaeal 20S particles. These residues corre-to connect the surrounding medium with the internalchamber of the particle (Lowe et al., 1995). Such a chan- spond to those found to block the axial pore of the yeast
20S and whose deletion led to rapid substrate entry andnel through the � rings was not observed in the X-raystructure of yeast 20S particles (Groll et al., 1997). In- hydrolysis (Groll et al., 2000). The � subunits of the wild-
type and ��(2-12)20S proteasomes were expressed indeed, entry of small peptides into the eukaryotic particleappeared to be blocked by the N-terminal extremities E. coli, and the resulting particles were analyzed by
electron microscopy. Previous studies showed that ex-of the � subunits that act as a gate for the channel(Groll et al., 2000). However, these differences between pression of the archaeal � subunits alone leads to forma-
tion of seven membered rings which can assemble fur-archaeal and yeast 20S might only reflect differencesbetween the forms of the particles crystallized in these ther into double rings (Zwickl et al., 1994). Accordingly,
both wild-type and ��(2-12) proteasome � subunits formstudies.

Role of ATP Hydrolysis in Proteasomal Degradation73
Figure 4. Deletion of Amino-Terminal Residues of 20S � SubunitsOpens the Axial Channel of � Rings
Electron microscopy top views of the � subunits of wild-type (A)and ��(2-12) 20S proteasomes (B) were performed as described inExperimental Procedures. (C) represents the difference mapping ofthe averaged pictures. The diameter of both complexes is approxi-mately 11 nm.
particles with 7-fold symmetry (Figures 4A and 4B). Dif-ference maps of averaged images of the wild-type andthe �(2-12) � ring showed the presence of mass in thecenter of the � rings formed by the wild-type 20S �subunits, which was lacking in the center of the �(2-12)� ring (Figure 4C). Therefore, the wild-type archaeal pro-teasomes normally have their axial channel in a closedform, and deletion of residues 2–12 in the � subunitsled to the opening of this channel.
The ��(2-12) 20S variant enabled us to examine theeffect of gate opening as an isolated process, and thusthe ability of the ��(2-12) 20S variant to degrade pep-tides, unfolded proteins, or globular proteins was tested.In contrast to eukaryotic 20S particles, which degradesmall peptides at a slow rate when their axial channelis closed, the wild-type and the ��(2-12) archaeal 20Sparticles did not differ in degradating tetrapeptides (Fig- Figure 5. Deletion of Amino-Terminal Residues of 20S � Subunits
Increases Rate of Degradation of Unfolded Proteins and Eliminatesure 5A). However, the ��(2-12) 20S degraded caseinStimulation by PAN29 times and acid-denatured GFPssrA 46 times faster(A) Wild-type and ��(2-12) 20S proteasomes degrade a tetrapeptidethan wild-type 20S proteasomes (Figure 5B). In fact, theat similar rates. The degradation of 100 �M of Suc-LLVY-amc was��(2-12) 20S degraded protein substrates even morecarried out as described in Experimental Procedures with 420 nMefficiently than the wild-type 20S in the presence ofof wild-type (solid line) or ��(2-12) 20S (dashed line) at 45�C in 50
PAN and ATP (three and five times more for casein and mM Tris (pH 7.5), 1 mM DTT, and 10 mM MgCl2.denatured GFPssrA, respectively). Moreover, degrada- (B) Degradation of GFPssrA, acid-denatured GFPssrA, and caseintion of casein and acid-denatured GFPssrA by the by ��(2-12) 20S. Degradation of 1 �M of 14C-radiolabeled protein
was followed as in Figure 2. Values are means plus standard devia-��(2-12) 20S variant was not stimulated further by thetions of three independent experiments.addition of PAN and ATP. However, the ��(2-12) 20S,(C) PAN promotes degradation of GFPssrA by the ��(2-12) 20S. Thelike the wild-type 20S, was not able to degrade thetime course of fluorescence change of 250 nM of GFPssrA was
globular GFPssrA, in the absence of PAN or ATP (Figures followed at 45�C in 50 mM Tris (pH 7.5), 1 mM DTT, and 10 mM5B and 5C). Presumably, the folded GFPssrA molecule, MgCl2 in the presence of 20 nM of ��(2-12)20S and 2 mM of ATPunlike the unfolded one, is incapable of entering the (�); or of 20 nM of ��(2-12)20S, 10 nM of PAN complex, and 2 mM
of AMPPNP (�); or of 20 nM of ��(2-12)20S, 10 nM of PAN complex,20S particles even when the channel is open. Therefore,and 2 mM of ATP (�).although the amino-terminal extremities of the � sub-
units are not visible in the X-ray structure of archaealproteasomes, they function in gating, as they do in
After Unfolding by PAN, GFPssrA Degradationeukaryotic 20S. The axial channel of archaeal 20S pro-by the ��(2-12)20S Proteasome Still Requiresteasomes in its native state is still permeable to tetra-ATP Hydrolysis by PANpeptides, but is not large enough to accommodate poly-It is noteworthy that the ��(2-12)20S by itself couldpeptides, which require the presence of PAN and ATPdegrade the acid-denatured GFPssrA, but needed PANfor efficient degradation. Once the channel of the 20Sand ATP to hydrolyze the globular GFPssrA. We there-proteasome is opened by deleting the N-terminal ex-fore tested whether this 20S variant could by itself de-tremities of the � subunits, the dependence on PAN andgrade GFPssrA after this protein had first been unfoldedATP is abrogated for unfolded proteins. Therefore, weby PAN. We first incubated GFPssrA with PAN and ATP,conclude that PAN, in the presence of ATP, facilitatesand when the loss of fluorescence had reached its maxi-the opening of the gate in the 20S proteasome, enabling
the translocation of unfolded polypeptides. mal extent, further ATP hydrolysis was prevented by

Molecular Cell74
Figure 6. After Unfolding of GFPssrA by PAN,Hydrolysis of ATP by PAN Is Still Essentialfor Its Degradation by the ��(2-12)20S Pro-teasome Variant
(A) The time course of fluorescence changeof 250 nM of GFPssrA was followed in 50 mMTris (pH 7.5), 1 mM DTT, 1 mM ATP, and 10mM MgCl2 at 45�C in the presence of 50 nMof PAN (�, �, �). After 15 min of incubationto allow unfolding of GFPssrA, 0 nM (�) or 20nM of ��(2-12)20S was added to the samplestogether with 2 mM of ATP�S (�) or the equiv-alent volume of buffer (�, �). If ��(2-12)20Swere present together with PAN and ATPfrom the onset (�), only buffer was added.(B) GFPssrA was incubated as in (A), and atthe indicated times, aliquots were withdrawn,loaded on a 12% SDS-PAGE, and analyzedby Western blot using an anti-histidine anti-body. The antibody also recognizes the �
subunit of 20S proteasomes which carries a(His)6 tag.(C) GFPssrA amount on (B) was quantifiedand relative amounts were determined usingthe 0 time point as 100%. Relative amountsof GFPssrA in lanes 1–3 (�), 4–6 (�), 7–9 (�),and 10–12 (�) are plotted as indicated.
addition of an excess of ATP�S that totally inhibited cuvette just after addition of 20S and the loading on theelectrophoretic gel due to the very high ability of thisPAN’s ATPase activity, as well as its ability to unfold
GFPssrA (data not shown). Then, the ��(2-12)20S was variant to degrade unfolded substrates as noted in Fig-ure 5B. Such degradation was minimized in experimentsadded, and degradation of GFPssrA was assayed by
monitoring its fluorescence (Figure 6A) and the amount using the wild-type 20S and when aliquots were with-drawn before addition of proteasomes at 15 min (dataof the protein on a SDS-PAGE gel (Figures 6B and 6C).
After incubating GFPssrA with PAN and ATP, when not shown). Thus, although GFPssrA is unfolded by PAN,its degradation by 20S still requires ATP hydrolysis, eventhe 20S variant was added to the preincubated mixture
of GFPssrA, PAN, and ATP (Figure, 6, closed triangles, when the axial channel in the proteasome � ring is inan open state.lanes 7–9) degradation of GFPssrA occurred to a similar
extent as when the ��(2-12)20S, PAN, and ATP were all This finding strongly suggests that ATP hydrolysisby PAN serves an additional function after substratepresent from the onset (open circles, lanes 1–3). How-
ever, when the ��(2-12)20S was added in the presence unfolding and gate opening. This additional functionvery likely is translocation of unfolded substrates. Oneof ATP�S to prevent further ATP hydrolysis, GFPssrA
was not degraded (closed circles, lanes 10–12), even alternative possible explanation of this lack of degrada-tion of unfolded GFPssrA by the open channel 20S vari-though most of it had been unfolded by PAN during
the preincubation (open triangles). Similar results were ant in the presence of ATP�S would be that, in theabsence of ATP hydrolysis, the unfolded substrateobtained with wild-type 20S proteasomes (data not
shown). Some disappearance of GFPssrA was observed forms a stable association with PAN, which precludesentry of GFPssrA into the proteolytic chamber. However,on the electrophoretic gel at 15 min just after the addition
the ��(2-12)20S (lanes 7–9). This decrease corresponds prior work has established that the complex betweenGFPssrA and PAN is quite short lived, since PAN andprobably to inevitable degradation that occurs between
the withdrawing of the sample from the fluorescence GFPssrA, even when it is acid denatured, could not be

Role of ATP Hydrolysis in Proteasomal Degradation75
coimmunoprecipitated, even in the presence of ATP�S proteasome function, the energy costs for protein break-down by two E. coli ATP-dependent proteases have(Navon and Goldberg, 2001) (our unpublished data). Ac-
cordingly, PAN could not be retained by a GFPssrA- been investigated. It has been shown that during degra-dation of Arc-ssrA, 150 mol of ATP were hydrolyzed bycontaining resin (our unpublished data). In addition,
when PAN was added without ATP, it did not affect the ClpXP, and this number was independent of the sub-strate’s thermodynamic stability (Burton et al., 2001).degradation of acid-denatured GFPssrA or of casein
(Figure 5B), as would be expected if these substrates Thus, in degrading this small globular protein (about 75residues), ClpXP consumed almost as much ATP asremained bound to PAN in the absence of ATP hydroly-
sis. This observation also allows us to rule out a model found here for PAN-20S proteasome degrading themuch larger polypeptides casein and GFPssrA (220 andwhereby PAN hinders the entry of unfolded substrates
into the open channel 20S particle in the presence of 268 residues). Menon et al. (1987) found that ATP hydro-lysis by the lon protease is proportional to the numberATP�S.
Thus, once a globular protein has been unfolded by of cleavages made in the substrate, but with this type ofenzyme (which contains ATPase and proteolytic activePAN, it is not simply released into the medium and does
not reach the 20S by passive diffusion. Instead, the sites in the same polypeptide chain and where theseprocesses may be more tighly coupled), significantlyunfolded polypeptide has to be actively translocated by
PAN into 20S proteasomes, by a process that requires less ATP is consumed than by PAN-20S complex.ATP hydrolysis, even if 20S proteasomes have their axialchannel sufficiently open to digest the denatured protein. The Rate-Limiting Step in Protein Degradation
by ProteasomesBecause acid treatment of GFPssrA destroyed its globu-Discussionlar conformation and its ability to fluoresce, but did notaccelerate its rate of degradation, gross unfolding ofATP Consumption during Protein Degradation
Although it has long been appreciated that intracellular this protein can not be the rate-limiting step in its degra-dation. One alternative explanation of this result mightproteolysis requires ATP hydrolysis, the actual amount
of ATP required for the breakdown of a protein by pro- be that once the acid-denatured GFPssrA was dilutedinto the reaction buffer (pH 7.5) at 45�C, it refolded rap-teasomes is not known. The present studies show that
when the protein-activated ATP hydrolysis and proteoly- idly and therefore showed similar rates of proteolysisand ATP dependence as globular GFPssrA. However,sis proceed at linear rates, there is an apparent stoichi-
ometry between the amount of ATP molecules hy- several findings make this trivial explanation very un-likely. First, after removing the acid, the denatureddrolyzed and protein molecules degraded that appears
independent of the nature of the protein substrate. At GFPssrA did not regain its fluorescence, and after 2 hr,its absorption spectrum was still indistinguishable fromleast 300 ATP molecules are hydrolyzed by PAN when
proteasomes degrade 1 molecule of the protein sub- that of denatured GFPssrA maintained in acid (data notshown). Second, the yield of GFP refolding at 50�C isstrate. This surprisingly high number indicates that pro-
tein unfolding and translocation into the 20S particle only 19% of that at 37�C (Makino, 1997). Perhaps thestrongest evidence against refolding was that the acid-requires many cycles of ATP hydrolysis. Even more sur-
prising was the finding that the amount of ATP hydrolysis denatured GFPssrA, unlike globular GFPssrA, was rap-idly degraded by the ��(2-12)20S proteasome. There-per protein degraded is similar with the globular and
unfolded substrates. Thus, the rates of ATP hydrolysis fore, its structure must differ from that of the globularGFPssrA at 45�C. Although gross unfolding of globularby PAN are independent of whether or not PAN has to
catalyze substrate unfolding. This conclusion is also substrates does not appear to be the rate-limiting step indegradation, translocation of the unfolded polypeptidesupported by our finding that globular and unfolded
substrates stimulate ATP hydrolysis to similar extents. through the ATPase ring into the 20S particle could berate limiting. Consistent with this conclusion was theATP-driven changes in the conformation of PAN must
underlie both the unfolding and translocation through finding that deletion of the N-terminal residues of the �subunit that obstruct entry into the 20S led to muchthe ATPase ring. With PAN alone, the protein is probably
released into the solution. With 20S particles present, faster degradation of unfolded proteins, even faster thanwith PAN and ATP.these events are linked to substrate injection into the
proteolytic chamber. ATP consumption was identical It has been generally assumed that protein unfoldingis the rate-limiting step in the functioning of ATP-depen-whether or not proteasomes were present. In contrast,
with the ATP-dependent proteases from E. coli, the dent proteases. For example, it has been shown thatwhen isolated ClpX ATPase complex catalyzes the un-ATPase activity is altered by the proteolytic component;
e.g., ATP consumption by ClpA and ClpX decreases folding of GFPssrA, fluorescence of GFPssrA decreasesat a similar rate as GFPssrA is degraded by the holoen-upon addition of the ClpP protease (Burton et al., 2001;
Hwang et al., 1988) while ATP hydrolysis by HslU in- zyme ClpXP (Kim et al., 2000). The authors concludedthat unfolding is the rate-limiting step during ClpXP-creases in the presence of HslV (Seol et al., 1997).
Because the 11 residue recognition sequence in mediated proteolysis. On the other hand, Singh et al.(2000) observed a difference in the rates of GFPssrAGFPssrA (ssrA peptide) by itself stimulated ATP hydroly-
sis to a similar extent as GFPssrA or casein, the activa- unfolding and degradation. Nevertheless, both studiessuggested that the rate-limiting step precedes peptidetion of ATP consumption must be triggered by the inter-
action between the ATPase and the substrate. Although bond cleavage, in agreement with our conclusion thatthe rate-limiting step occurs after protein unfolding, andno prior studies have addressed the energy costs for

Molecular Cell76
Figure 7. Model for the Energy-Dependent Steps of PAN and Proteasomes-Mediated Protein Degradation
Cut side views of 20S particles are schematized with the active sites represented as blue dots in the internal chamber. PAN is schematizedas a hexameric ring. The circle and pentagonal shape of the subunits represent the substrate-free and substrate-bound forms of PAN,respectively. The gate which precludes entry of protein into 20S particles is represented in red.
thus probably is the translocation into the 20S particle. X-ray structure of archaeal proteasomes showed anopen axial channel (Lowe et al., 1995). By interactingFor the unfolded proteins, translocation into the 20S iswith 20S particles, PAN probably induces an ATP-clearly rate limiting, since their degradation was acceler-dependent movement of the N termini of the � subunitsated dramatically by deletion that enlarges the gate andthat displaces them from the surface of the axial pore.facilitates entry into the 20S (see below).A similar mechanism has been suggested for the activa-tion of 20S proteasomes by PA28 (Whitby et al., 2000).Gating of the Archaeal 20S ProteasomesWhen the channel into the 20S was opened by theThese studies have demonstrated that archaeal 20S pro-�(2-12) deletion, degradation of casein and acid-dena-teasomes contain a functional gate in their outer ringtured GFPssrA were rapid and independent of ATP andwhose opening appears to be regulated by PAN. Indeed,PAN, while in the wild-type, where the channel was in itssubstrate entry into the archaeal 20S is normally blockedclosed state, degradation of these unfolded substratesby the N termini of the � subunits, since deletion ofrequired PAN’s ATPase activity. Therefore, PAN appearsresidues 2–12 of those subunits dramatically increasedto open the gate in the � ring. Interestingly, the rates ofthe degradation of unfolded proteins and led to an open-degradation of unfolded proteins by the PAN-wild-typeing in the � ring visible by electron microscopy analysis.20S complex was lower than those by the ��(2-12) 20S.Those residues are highly conserved among prokaryoticTherefore, protein binding to PAN and ATP-dependentand eukaryotic proteasomes, and the present findingstranslocation through the ATPase ring into the 20S parti-are similar to those obtained with analogous deletionscle appear to be slower than diffusion directly throughin yeast 20S particles (Groll et al., 2000). With eukaryoticthe open axial channel, perhaps because the PAN-20S proteasomes in their native state, entry and exit ofinduced opening in the � ring may not be as large as insmall peptides requires gate opening by the 19S regulatorythe deletion mutant.particle (Kohler et al., 2001) or by PA28-related activa-
In eukaryotes, a homologous ATPase in the 19S parti-tors (Whitby et al., 2000). By contrast, entry of tetrapep-cle activates proteolysis apparently by promoting thetide into archaeal 20S particles occurs readily since theopening of the gate into the 20S particles. Mutation in
��(2-12) 20S variant and the wild-type 20S-PAN com-one of the six ATPases (rpt2RF) strongly reduced the
plex did not degrade these small peptides faster thanability of 20S particles to hydrolyze peptides (Rubin et
the wild-type 20S proteasome (this work; Zwickl et al.,al., 1998). This defect could be suppressed by a mutation
1999). Thus, although archaeal proteasomes show ATP-in the 20S particle that leads to the opening of the chan-
regulated gating that prevents entry of polypeptides, in nel in the � ring (Kohler et al., 2001). Therefore, Rpt2the basal state these particles do appear open to small appears to be particularly important in regulating open-peptides, unlike the eukaryotic 20S. ing of the axial channel and peptide entry, although
analogous mutations in the other five ATPase subunitsATP-Dependent Gate Opening were not analyzed.and Substrate Translocation Even after its unfolding by PAN, and even when theThe most fundamental function of the PAN ATPase is axial channel of the 20S particles was open, hydrolysisto facilitate entry of protein substrates into proteasomes of ATP by PAN was still required for degradation offor degradation. Since the axial channel in the � ring globular GFPssrA. In contrast, when the GFPssrA washas a gate that normally prevents protein entry, it is denatured with acid, PAN was not required for its trans-not surprising that PAN and ATP trigger gate opening. location into the ��(2-12)20S. This difference in the re-
quirement for PAN between GFPssrA unfolded by PANHowever, such a function was unexpected because the

Role of ATP Hydrolysis in Proteasomal Degradation77
Expression and Purification of 20S � Ringsand GFPssrA denatured by acid is intriguing. Perhaps,The � gene was elongated at the 3� end by an affinity tag that codeswhen PAN unfolds this substrate, it does not adopt afor 6 histidine residues and cloned into pRSET5a expression vector.stable extended conformation that can diffuse readilyThen, the tagged � gene was deleted for 33 nucleotides at the 5�
into the 20S particles. The gross unfolding of GFPssrA end, which codes for residues 2–12, thus generating the �(2-12)�occurs on the surface of PAN’s ring, and probably pre- gene. Wild-type and mutant � proteins were expressed in BL21(DE3)
and purified as 20S proteasomes.cedes its PAN-mediated substrate threading throughPAN’s central opening into the 20S (Navon and Gold-
Hydrolysis of ATPberg, 2001). Perhaps, this ATP-dependent threadingHydrolysis of ATP was assayed by following production of inorganicthrough PAN might also eliminate residual secondaryphosphate at 45�C as described elsewhere (Ames, 1966; Zwickl et
structural elements of GFPssrA that prevent its diffusion al., 1999).into ��(2-12)20S particles.
In summary, these findings allow us to dissect several Degradation of 14C-Radiolabeled Proteins14C-radiolabeled casein and GFPssrA were prepared by reductivedistinct steps where PAN is required during protein deg-
alkylation, a gentle procedure that does not produce dramaticradation (Figure 7). The substrate binds to PAN, whichchanges in protein structure (Rice and Means, 1971) and stored intriggers a change in its conformation leading to an in-50 mM Tris-HCl (pH 7.5). The radiolabeling of GFPssrA did not altercreased capacity to hydrolyze ATP. Substrate bindingits fluorescence properties. Breakdown of these 14C-radiolabeled
to PAN can occur on the isolated PAN or on the PAN-20S proteins was conducted in the presence of 20S proteasomes, PAN,complex. This substrate-activated ATPase (i) facilitates and ATP as indicated in 50 mM Tris (pH 7.5), 1 mM DTT, and 10
mM MgCl2 at 45�C. After 40–60 min incubation, trichloroacetic acidunfolding of the substrate, probably at the surface ofat a final concentration of 12% was added together with 100 �g ofthe ATPase ring, (ii) favors an open form of the 20S axialBSA as a carrier. Acid-soluble radioactive peptides products werechannel, and (iii) promotes substrate translocation intoquantify by scintillation counting.the internal chamber of the 20S particles where peptide
bond cleavage occurs. (Because ATP hydrolysis by PAN Peptides Hydrolysiscan also occur in the absence of a protein substrate, 100 �M of fluorogenic Suc-LLVY-amc peptide diluted in 100%although at a lower rate, it seems likely that gate opening DMSO was incubated with 420 nM 20S proteasomes at 45�C in 50
mM Tris (pH 7.5), 1 mM DTT, and 10 mM MgCl2. Hydrolysis of thein the 20S by PAN occurs also in the absence of apeptide was followed by measuring the relase of amc (7-amido-4-substrate.) These distinct functions of PAN’s ATPasemethylcoumarin) in a spectrofluorometer (ex 380 nm; em 440 nm).activity are very likely to apply to the closely homologous
19S ATPases, although in addition one ATPase subunit GFPssrA Fluorescencein the 19S is involved in interacting with ubiquitinated Measurements of GFPssrA fluorescence were obtained at 45�C withproteins (Lam et al., 2002). excitation at 400 nm and emission at 510 nm. GFPssrA was diluted
in 50 mM Tris (pH 7.5), 1 mM DTT, and 10 mM MgCl2, in the presencePAN belongs to the large family of AAA ATPasesor the absence of PAN, 20S proteasomes, and the indicated nucle-(Ogura and Wilkinson, 2001), many of which have beenotide.shown to catalyze protein remodeling. The present find-
ings on substrate-activated ATPase activity, the energyDenaturation of GFPssrA
costs of unfolding and translocation, and the multiple GFPssrA or [14C]-GFPssrA were incubated with 50 mM HCl (pH 2.0)roles of ATP hydrolysis are likely to apply also to these for 10 min at room temperature. Denatured GFPssrA was diluted
directly into reaction mixture at 45�C and used immediately.related multimeric ATPases. Analogous studies of theseAAA family members may also help clarify their molecu-
Electron Microscopy and Image Analysislar mechanisms.Purified proteins were negatively stained with 2% uranyl acetate.600 end-on views of wild-type � rings and of mutant � rings wereExperimental Proceduresextracted and aligned by conventional cross-correlation methods.After performing correspondence analysis, the projections were in-Proteins and Peptidesterpreted according to their coordinates along the dominating firstPAN and archaeal 20S proteasomes were purified as previouslyfactor.described (Benaroudj and Goldberg, 2000; Zwickl et al., 1999). Molar
concentrations of PAN were expressed on the basis of a 650 kDaAcknowledgmentscomplex (Zwickl et al., 1999). (His)6-GFP and (His)10-GFPssrA were
purified as described earlier (Benaroudj and Goldberg, 2000) withThis work was supported by research grants from NIH to A.L.G.the following modifications. After purification on an Ni-NTA agarose(GM46147 and GM 51923), postdoctoral fellowships from Associa-column, the fractions containing GFP proteins were pooled andtion pour la Recherche contre le Cancer and Human Frontier Sciencesubjected to a MonoQ HR5/5 (Amersham Pharmacia Biotech) anionProgram Organization to N.B., and by a postdoctoral fellowshipexchange column equilibrated with 50 mM Tris-HCl (pH 7.5) andfrom the Deutsche Forschungsgemeinschaft to P.Z. Authors are1 mM DTT. A linear gradient of NaCl from 0 to 500 mM was applied,very grateful to Gunter Pfeifer for her excellent technical assistanceand the GFP proteins eluted at approximately 150–200 mM. Frac-in electron microscopy and image analysis. We thank Prasannations containing (His)6-GFP or (His10)-GFPssrA were pooled andVariath, Marion Schmidt, and Tom Rapoport for critically readingdialyzed against 50 mM Tris-HCl (pH 7.5). The ssrA peptide (NH2-this manuscript, and Sarah Trombley for assistance in preparationCAANDENYALAA-CO2H) was a generous gift from Y.I. Kim (Kim etof the manuscript.al., 2000). Casein was purchased from Sigma.
Received: June 23, 2002Construction of ��(2-12)20S Proteasome VariantRevised: October 31, 2002Deletion of residues 2–12 in the � subunits of archaeal 20S protea-
somes was performed by reverse PCR mutagenesis using theReferencespRSET5a-�(His)6-� described earlier (Seemuller et al., 1995; Zwickl
et al., 1992). The resulting pRSET5a-�(His)6-��(2-12) plasmid wasthen introduced in E. coli BL21(DE3) cells, and the ��(2-12)20S Ames, B.N. (1966). Assay of inorganic phosphate, total phosphate
and phosphatases. Methods Enzymol. 8, 115–118.proteasome variant was purified as the wild-type 20S proteasomes.

Molecular Cell78
Baumeister, W., Walz, J., Zuhl, F., and Seemuller, E. (1998). The surface of the ATPase ring before transport into the proteasome.Mol. Cell 8, 1339–1349.proteasome: paradigm of a self-compartmentalizing protease. Cell
92, 367–380. Ogura, T., and Wilkinson, A.J. (2001). AAA superfamily ATPase:common structure-diverse function. Genes Cells 6, 575–597.Benaroudj, N., and Goldberg, A.L. (2000). PAN, the proteasome-
activating nucleotidase from archaebacteria, is a protein-unfolding Rice, R.H., and Means, G.E. (1971). Radioactive labeling of proteinsmolecular chaperone. Nat. Cell Biol. 2, 833–839. in vitro. The J. Biol. Chem. 246, 831–832.Bokman, S.H., and Ward, W.W. (1981). Renaturation of Aequorea Rubin, D.M., Glickman, M.H., Larsen, C.N., Dhruvakumar, S., andgreen-fluorescent protein. Biochem. Biophys. Res. Commun. 101, Finley, D. (1998). Active site mutants in the six regulatory particle1372–1380. ATPases reveal multiple roles for ATP in the proteasome. EMBO J.
17, 4909–4919.Burton, R.E., Siddiqui, S.M., Kim, Y.-I., Baker, T.A., and Sauer, R.T.(2001). Effects of protein stability and structure on substrate pro- Seemuller, E., Lupas, A., Stock, D., Lowe, J., Huber, R., and Baumeis-cessing by the ClpXP unfolding and degradation machine. EMBO ter, W. (1995). Proteasome from Thermoplasma acidophilum: a thre-J. 20, 3092–3100. onine protease. Science 268, 579–582.
Coux, O., Tanaka, K., and Goldberg, A.L. (1996). Structure and func- Seol, J.H., Yoo, S.J., Shin, D.H., Shim, Y.K., Kang, M.S., Goldberg,tions of the 20S and 26S proteasomes. Annu. Rev. Biochem. 65, A.L., and Chung, C.H. (1997). The heat-shock protein HslVU from801–847. Escherichia coli is a protein-activated ATPase as well as an ATP-
dependent proteinase. Eur. J. Biochem. 247, 1143–1150.Creamer, L.K., Richardson, T., and Parry, D.A.D. (1981). Secondarystructure of bovine �s1- and �-casein in solution. Arch. Biochem. Singh, S.K., Grimaud, R., Hoskins, J.R., Wickner, S., and Maurizi,
M.R. (2000). Unfolding and internalization of proteins by the ATP-Biophys. 211, 689–696.dependent proteases ClpXP and ClpAP. Proc. Natl. Acad. Sci. USADe Mot, R., Nagy, I., Walz, J., and Baumeister, W. (1999). Protea-97, 8898–8903.somes and other self-compartmentalizing proteases in prokaryotes.Ward, W.W., and Bokman, S.H. (1982). Reversible denaturation ofTrends Microbiol. 7, 88–92.Aequorea green-fluorescent protein: physical separation and char-Glickman, M.H., Rubin, D.M., Fried, V.A., and Finley, D. (1998). Theacterization of the renatured protein. Biochemistry 21, 4535–4540.regulatory particle of the Saccharomyces cerevisiae proteasome.Weber-Ban, E.U., Reid, B.G., Miranker, A.D., and Horwich, A.L.Mol. Cell. Biol. 18, 3149–3162.(1999). Global unfolding of a substrate protein by the Hsp100 chap-Goldberg, A.L. (1992). The mechanism and functions of ATP-depen-erone ClpA. Nature 401, 90–93.dent proteases in bacterial and animal cells. Eur. J. Biochem. 203,Wenzel, T., and Baumeister, W. (1995). Conformational constraints9–23.in protein degradation by the 20S proteasome. Nat. Struct. Biol. 2,Groll, M., Ditzel, L., Lowe, J., Stock, D., Bochtler, M., Bartunik, H.D.,199–204.and Huber, R. (1997). Structure of 20S proteasome from yeast atWhitby, F.G., Masters, E.I., Kramer, L., Knowlton, J.R., Yao, Y., Wang,2.4 A resolution. Nature 386, 463–471.C.C., and Hill, C.P. (2000). Structural basis for the activation of 20SGroll, M., Bajorek, M., Kohler, A., Moroder, L., Rubin, D.M., Huber,proteasomes by 11S regulators. Nature 408, 115–120.R., Glickman, M.H., and Finley, D. (2000). A gated channel into theZwickl, P., Lottspeich, F., and Baumeister, W. (1992). Expression ofproteasome core particle. Nat. Struct. Biol. 7, 1062–1067.functional Thermoplasma acidophilum proteasomes in Escherichia
Holzl, H., Kapelari, B., Kellermann, J., Seemuller, E., Sumegi, M.,coli. FEBS Lett. 312, 157–160.
Udvardy, A., Medalia, O., Sperling, J., Muller, S.A., Engel, A., et al.Zwickl, P., Kleinz, J., and Baumeister, W. (1994). Critical elements(2000). The regulatory complex of Drosophila melanogaster 26Sin proteasome assembly. Nat. Struct. Biol. 1, 765–770.proteasomes: subunit composition and localization of a deubiquityl-Zwickl, P., Ng, D., Woo, K.M., Klenk, H.P., and Goldberg, A.L. (1999).ating enzyme. J. Cell Biol. 150, 119–129.An archaebacterial ATPase, homologous to ATPases in the eukary-Hoskins, J.R., Singh, S.K., Maurizi, M.R., and Wickner, S. (2000).otic 26S proteasome, activates protein breakdown by 20 S protea-Protein binding and unfolding by the chaperone ClpA and degrada-somes. J. Biol. Chem. 274, 26008–26014.tion by the protease ClpAP. Proc. Natl. Acad. Sci. USA 97, 8892–Zwickl, P., Goldberg, A.L., and Baumeister, W. (2000). Proteasomes8897.in prokaryotes. In Proteasomes: The World of Regulatory Proteoly-Hwang, B.J., Woo, K.M., Goldberg, A.L., and Chung, C.H. (1988).sis, W. Hilt and D. Wolf, eds. (Georgetown, TX: Landes Bioscience),Protease Ti, a new ATP-dependent protease in Escherichia coli,pp. 8–20.contains protein-activated ATPase and proteolytic functions in dis-
tinct subunits. J. Biol. Chem. 263, 8727–8734.
Kim, Y.I., Burton, R.E., Burton, B.M., Sauer, R.T., and Baker, T.A.(2000). Dynamics of substrate denaturation and translocation by theClpXP degradation machine. Mol. Cell 5, 639–648.
Kohler, A., Cascio, P., Leggett, D.S., Woo, K.M., Goldberg, A.L., andFinley, D. (2001). The axial channel of the proteasome core particleis gated by the Rpt2 ATPase and controls both substrate entry andproduct release. Mol. Cell 7, 1143–1152.
Lam, Y.A., Lawson, T.G., Velayutham, M., Zweier, J.L., and Pickart,C.M. (2002). A proteasomal ATPase subunit recognizes the polyubi-quitin degradation signal. Nature 416, 763–767.
Lowe, J., Stock, D., Jap, B., Zwickl, P., Baumeister, W., and Huber,R. (1995). Crystal structure of the 20S proteasome from the archaeonT. acidophilum at 3.4 A resolution. Science 268, 533–539.
Lupas, A., Koster, A.J., and Baumeister, W. (1993). Structural fea-tures of 26S and 20S proteasomes. Enzyme Protein 47, 252–273.
Makino, Y. (1997). Chaperonin-mediated folding of green fluorescentprotein. J. Biol. Chem. 272, 5326–5334.
Menon, A.S., Waxman, L., and Goldberg, A.L. (1987). The energyutilized in protein breakdown by the ATP-dependent protease (La)from Escherichia coli. J. Biol. Chem. 262, 722–726.
Navon, A., and Goldberg, A.L. (2001). Proteins are unfolded on the




















