
Mutant nuclear lamin A leads to progressive alterations of epigenetic control in premature aging

Lamin A Ser404 Is a Nuclear Target of Akt Phosphorylation in C2C12
Cells
Vittoria Cenni,†,‡,# Jessika Bertacchini,†,# Francesca Beretti,† Giovanna Lattanzi,‡
Alberto Bavelloni,§ Massimo Riccio,† Maria Ruzzene,| Oriano Marin,| Giorgio Arrigoni,|
Veena Parnaik,∇ Manfred Wehnert,O Nadir M. Maraldi,‡,§ Anto de Pol,† Lucio Cocco,⊥ andSandra Marmiroli*,†,‡
Department of Anatomy and Histology and CIPro Proteomics Centre, University of Modena and Reggio Emilia,Via Del Pozzo 71, I-41100 Modena, Italy, I.G.M.-CNR, Unit of Bologna, c/o IOR, via di Barbiano,
I-40125 Bologna, Italy, Laboratory of Cellular Biology and Electron Microscopy, IOR, I-40125 Bologna, Italy,Department of Biological Chemistry, Viale Colombo, I-35121 Padova, Italy, Department of Anatomical
Sciences, Cellular Signalling Laboratory, Via Irnerio 48, I-40126 Bologna, Italy, Centre for Cellular & MolecularBiology, Hyderabad, IN-500 007, India, and Institute of Human Genetics, Greifswald, D-17484, Germany
Received April 8, 2008
Akt/PKB is a central activator of multiple signaling pathways coupled with a large number of stimuli.Although both localization and activity of Akt in the nuclear compartment are well-documented, mostAkt substrates identified so far are located in the cytoplasm, while nuclear substrates have remainedelusive. A proteomic-based search for nuclear substrates of Akt was undertaken, exploiting 2D-electrophoresis/MS in combination with an anti-Akt phosphosubstrate antibody. This analysis indicatedlamin A/C as a putative substrate of Akt in C2C12 cells. In vitro phosphorylation of endogenous laminA/C by recombinant Akt further validated this result. Moreover, by phosphopeptide analysis and pointmutation, we established that lamin A/C is phosphorylated by Akt at Ser404, in an evolutionaryconserved Akt motif. To delve deeper into this, we raised an antibody against the lamin A Ser404phosphopeptide which allowed us to determine that phosphorylation of lamin A Ser404 is triggeredby the well-known Akt activator insulin, and is therefore to be regarded as a physiological response.Remarkably, expression of S404A lamin A in primary cells from healthy tissue caused the nuclearabnormalities that are a hallmark of Emery-Dreifuss muscular dystrophy (EDMD) cells. Indeed, it isknown that mutations at several sites in lamin A/C cause autosomal dominant EDMD. Very importantly,we show here that Akt failed to phosphorylate lamin A/C in primary cells from an EDMD-2 patient withlamin A/C mutated in the Akt consensus motif. Together, our data demonstrate that lamin A/C is anovel signaling target of Akt, and implicate Akt phosphorylation of lamin A/C in the correct function ofthe nuclear lamina.
Keywords: Akt/PKB • nucleus • Lamin A/C • proteomics • 2D-electrophoresis • phosphorylation
IntroductionSince its discovery as an oncogene in the mouse leukemia
virus AKT81,2 and as a homologue of protein kinase C,3 the 63kDa serine/threonine kinase Akt/PKB has been involved inpromotion of cell survival, proliferation and metabolic re-sponses downstream the phosphoinositide-3-kinase (PI3 ki-nase) signaling pathway.4-6 In resting cells, Akt is a predomi-nantly cytosolic enzyme; however, generation of PI3 kinase lipid
products recruits Akt to the plasma membrane,4,5,7 resultingin a conformational change which confers full enzymaticactivity through the phosphorylation of the membrane-boundprotein at two residues, Thr308 and Ser473. Thr308 lies withinthe T loop of the catalytic domain, and is phosphorylated bythe 3-phosphoinositide-dependent-protein kinase 1, PDK-1.8
Conversely, the identity of the Ser473, in the C-terminalhydrophobic motif, originally termed PDK-2, has remained amystery for several years, the results from various laboratorybeing very controversial.9-12 Nevertheless, the regulation ofSer473 has been unravelled recently by the identification ofboth the kinase and the phosphatase that specifically act onthis residue.13-15 Indeed, a rapamycin-insensitive form of themTOR protein kinase complexed to Rictor:G�L (mTORC2) hasbeen shown to phosphorylate Ser473 directly in vivo, inresponse to serum.13
* To whom correspondence should be addressed. E-mail: [email protected].
† University of Modena and Reggio Emilia.‡ I.G.M.-CNR, Unit of Bologna.# These authors contributed equally.§ Laboratory of Cellular Biology and Electron Microscopy.| Department of Biological Chemistry.∇ Centre for Cellular & Molecular Biology.O Institute of Human Genetics.⊥ Cellular Signalling Laboratory.
10.1021/pr800262g CCC: $40.75 2008 American Chemical Society Journal of Proteome Research 2008, 7, 4727–4735 4727Published on Web 09/23/2008

The mechanism by which Akt is inactivated was initiallysuggested to depend on protein phosphatase 2A, PP2A. How-ever, recent work has provided strong evidence that the PHdomain leucine-rich repeat protein phosphatases PHLPP1/2dephosphorylate Ser473 directly and terminate Akt signaling,thus, triggering apoptosis and suppressing tumor growth.14,15
Activated Akt redistributes to cytoplasm and nucleus, wherephosphorylation of specific substrates, such as GSK3, Bad,Mdm2 and TSC2, occurs.6,16,17
Both the presence and the activity of Akt in the nucleus havebeen described thoroughly.18,19 In particular, a recent studyemployed a genetically encoded fluorescent reporter for Aktactivity that allowed real-time imaging of phosphorylationcatalyzed by Akt.18 By this technology, Akt signaling in thenucleus was found to be more stable compared with that inthe cytosol, suggesting that Akt activity in these two compart-ments is regulated by different mechanisms.18 However, thesignal in the nucleus was also less rapid, probably reflectingthe time it takes for activated Akt to translocate into the nuclearcompartment, and thus raising the question of what mecha-nism mediates Akt nuclear translocation. Our recent workdemonstrated that, in response to precise signals, Akt istargeted to the cytoskeleton by direct interaction of its N-terminal PH domain with actin, and that this process isregulated by the small GTP-binding proteins cdc42 and Rac.20
We suggested that this mechanism might contribute to Aktredistribution to diverse subcellular locations, such as thenucleus. Another interesting mechanism that mediates nucleartranslocation of Akt has been described in human mature T-cellleukemia: the product of TCL1 gene, Tcl1, interacts with thePH domain of phosphorylated Akt, thus, driving Akt to thenucleus.21 In this context, Tcl1 may act as a direct transporterof Akt or may contribute to the formation of a complex thatpromotes the transport of active Akt to the nucleus, where itcan phosphorylate nuclear substrates. A well described nuclearsubstrate is Foxo. IGF-1 triggers phosphorylation of Foxo byAkt inside the nucleus, where phosphorylated Foxo associateswith 14.3.3 proteins that, in turn, promote its export to thecytoplasm.22 Remarkably, Foxo phosphorylation by Akt hasbeen shown to be a crucial event in Akt-dependent myogen-esis.23 In spite of these striking observations, most Akt nuclearsubstrates, as well as nuclear Akt functions, have so farremained elusive. This lack of information prompted us toundertake a search of substrates of Akt in the nucleus. Reckon-ing that functional proteomics is a very powerful approach tothe discovery of kinase targets, we decided to combine 2D-separation/mass spectrometry with Western blotting by anti-Akt-phosphosubstrate antibody and bioinformatic screening forthe Akt phosphorylation motif.24-26
Lamin A and lamin C, both products of the LMNA gene, aretype V intermediate filaments protein that participate in theformation of the nuclear lamina, a protein meshwork liningthe nucleoplasmic surface of the inner nuclear membranethought to provide a framework for organizing nuclear envelopestructures and anchoring the interphase chromatin.27-29 LaminA/C have essential roles in maintaining lamina stability, andin regulating transcription factors. Our data establish lamin A/Cas new genuine signaling targets of Akt, and suggest that theirphosphorylation is involved in the correct function of thelamina scaffold.
Experimental Procedures
Cell Culture and Transfections. Human embryonic kidneyepithelial (HEK) 293T cells and C2C12 mouse myoblasts weregrown in DMEM-HG (Dulbecco’s Modified Eagle’s MediumHigh Glucose, Sigma-Aldrich) supplemented with 10% FetalCalf Serum (FCS, EuroClone Ltd., U.K.).
Transient transfections of 293T cells were performed by thecalcium-phosphate method.20 After transfection, cells wereserum-starved for 24 h and used for experiments.
Plasmids. Wild-type, flag-tagged rat lamin A, subcloned inpCI expression vector as described by Kumaran et al.,30 wasused as template to generate S301A, S404A and S301A/S404Amutants by means of the QuikChange strategy (Stratagene, LaJolla, CA). The following mutation primers were used: S301Amutant, forward primer CGAATCCGCATTGACGCCCTCTCAGC-CCAG; S301A mutant, reverse primer CTGGGCTGAGAGGGCGT-CAATGCGGATTCG; S404A mutant, forward primer GCCGCGC-CTCCGCCCACTCCTCCC; S404A mutant, reverse primer GGGAG-GAGTGGGCGGAGGCGCGGC. All constructs were verified byDNA sequencing service (MWG Biotech Germany).
To transiently silence the expression of Akt1, C2C12 cellswere transfected with vectors that express Akt1 (or scramble,where indicated) hairpin siRNAs under the control of the mouseU6 promoter, as described previously,31 with the Cell LineNucleofector kit V and Amaxa Nucleoporator (InstrumentationLaboratory, Milano, Italy), following manufacturer’s instruc-tions. To monitor transfection efficiency, pEGFPN1 was cotrans-fected at a 1:10 ration with Akt vectors and 15 mm glasscoverslips were added to each plate before seeding. Count ofGFP-positive cells was 70% ( 5% in all samples.
All other constructs were as described previously.32,33
Preparation of Cell Extracts. Subconfluent cells were ex-tracted by addition of RIPA buffer (20 mM Tris-Cl, pH 7.0, 1%NP-40, 150 mM NaCl, 10% Glycerol, 10 mM EDTA, 20 mM NaF,5 mM Sodium Pyrophosphate, 1 mM Na3VO4, and freshlyadded Sigma-Aldrich Protease Inhibitor Cocktail) at 4 °C for10 min. Lysates were cleared by centrifugation and used forimmunoprecipitation experiments, as described below. A totalof 50 µL of the total lysate was immediately boiled in SDSsample buffer, resolved, and immunoblotted with the indicatedantibodies. Separation of nuclei was obtained by hypotonicshock and shearing; nuclei were obtained as pellet by a 300gcentrifugation at 4 °C. The purity of nuclear fraction waschecked by evaluating the presence of cytoplasmic contami-nants (tubulin, not shown).
Immunoprecipitation and Electrophoresis. Equal amountsof precleared lysates, whose protein concentration was deter-mined by the Bradford method, were incubated for 3 h withanti-flag (M2 clone, Sigma-Aldrich), anti-lamin A/C (SantaCruz) (3 µg all) or mouse IgG to provide a negative control,plus 30 µL of 50% (v/v) of protein A/G agarose slurry (SantaCruz Biotech.) at 4 °C with gentle rocking. Pellets were washedtwice with PBS plus 1% NP-40, twice in TNE (10 mM Tris-Cl,pH 7.5, 100 mM NaCl, 1 mM EDTA), once with 10 mM Tris-Cl,pH 7.4, boiled in Laemmli sample buffer, and centrifuged.Supernatants were loaded onto SDS-polyacrylamide gel, blottedon Immobilon-P membranes (Millipore, MA), and processedby Western blot with the indicated antibody, detected bySupersignal substrate chemiluminescence detection kit (Pierce,U.K.). Quantitation of the signal was obtained by chemilumi-nescence detection on a Kodak Image Station 440CF andanalysis with the Kodak 1D Image software.
research articles Cenni et al.
4728 Journal of Proteome Research • Vol. 7, No. 11, 2008

For 2D-electrophoresis, a total volume of 120 µL of TSU(Urea 7M, Thiourea 2M, CHAPS 4%, DTT 1% Tris-Cl, pH 7.4,20 mM, Ampholines 0.2% plus fresh protease inhibitor cocktail,DNase and RNase) containing nuclear lysates (120 µg) or anti-M2 immunoprecipitates (eluted with the 3× FLAG peptide,Sigma-Aldrich, following manufacturer’s instructions) wasloaded onto 3-10 or 5-8 IEF strips (Bio-Rad) and left focusedin a Protean IEF cell (from Bio-Rad) until a final VoltHour of12000 was reached. Strips were then reduced for 15 min with125 mM DTT in EB (equilibration buffer: Tris-Cl, pH 8.6, 5 mM,Urea 6M, Glycerol 30%, SDS 2%) and alkylated with 250 mMiodacetamide in EB for 8 min. Strips and samples were resolved
on SDS-polyacrylamide gel, blotted onto nitrocellulose, probedwith specific primary antibodies and HRP-conjugated second-ary antibodies (Pierce, U.K.), and developed with Pierce Su-persignal substrate chemiluminescence detection kit. Thefollowing antibodies were used: anti-flag, clone M2, fromSigma-Aldrich, anti-lamin A/C (N-terminal), anti-lamin A /C(C-terminal), anti-actin, anti-tubulin, anti-parp, anti-Akt, fromSanta Cruz Biotechnology; anti-Akt phosphoserine 473 (pAktS473) and anti-Akt phosphosubstrate, from Cell SignallingTechnology (MA). Anti-phospholamin A (S404) and its unphos-phorylated counterpart were raised in New Zealand rabbitsagainst two synthetic peptides corresponding to amino acids[400-413] of the Akt protein, and phosphorylated in Ser404 ornot, respectively; the antisera were purified using an im-mobilized peptide affinity resin (Sulfo Link Coupling Gel,Pierce, U.K.) according to the manufacturer’s instructions.
In-Gel Digestion and LC-MS/MS Analysis. The spots wereexcised from the gel with the help of an Ettan spot picker (GEHealthcare), and the protein digestion was performed in-gelusing sequencing grade modified trypsin (Promega, Madison,WI). Briefly, gel spots were washed with acetonitrile, driedunder vacuum and treated with 12.5 ng/µL of trypsin at 37 °Covernight. After digestion, the peptides were extracted by 3changes of 50% acetonitrile/0.1% formic acid (20 min betweenchanges), dried under vacuum, resuspended with 10 µL of 0.1%formic acid and analyzed by LC-MS/MS.
The samples were analyzed using a LCQ XP (ThermoElectron, San Jose, CA) interfaced with a nano-LC system 1100series (Agilent Technologies, Santa Clara, CA) and a capillarycolumn Zorbax 300SB C18, 3.5 µm, 150 mm × 75 µm (Agilent),
Figure 1. Lamin A/C is a nuclear substrate of Akt. Nuclear lysatesfrom C2C12 myoblasts were purified and subjected to 2D-electrophoresis on a pH 3-10 isoelectric focusing (IEF) gradientand a 10% polyacrilammide gel. (a) Nuclear proteins werevisualized by Coomassie blue staining; asterisks indicate proteinsidentified as A-type lamins; (b) a replicate gel was blotted andprobed with anti-Akt phosphosubstrate (Akt psubstrate) antibody(1:1000, Cell Signaling Technology); (c) the filter was strippedand reprobed with an anti-lamin A/C antibody (1:500, Santa CruzBiotech). Arrows indicate lamin A (72 kDa) and lamin C (68 kDa).
Table 1. Tryptic Peptides Derived from Lamin A/C andIdentified by LC-MS/MS Analysisa
sequence Mascot score exp. value
R.LQLELSK.V 50 0.019R.LVEIDNGK.Q 47 0.042K.LEAALGEAK.K 53 0.0097R.DLEDSLAR.E 36 0.37R.SSFSQHAR.T 58 0.002K.DLEALLNSK.E 63 0.001R.LADALQELR.A 82 8.7e-006K.LEAALGEAKK.Q 49 0.023K.QLQDEMLR.R 44 0.064R.KLLEGEEER.L 63 0.00087R.LVEIDNGKQR.E 56 0.0039R.TLEGELHDLR.G 53 0.0064K.LRDLEDSLAR.E 57 0.0021R.VAVEEVDEEGK.F 71 0.0001R.LKDLEALLNSK.E 70 0.00011K.AAYEAELGDARK.T 72 6.7e-005R.SGAQASSTPLSPTR.I 62 0.00075R.AQHEDQVEQYK.K 56 0.0034K.NIYSEELRETK.R 58 0.0014R.TVLCGTCGQPADK.A 73 5.4e-005R.IDSLSAQLSQLQK.Q 70 0.00011R.TAPINSTGEEVAMR.K 49 0.015R.AQHEDQVEQYKK.E 63 0.00057R.SVGGSGGGSFGDNLVTR.S 118 1.3e-009R.VAVEEVDEEGKFVR.L 77 1.8e-005R.TLEGELHDLRGQVAK.L 55 0.003R.IRIDSLSAQLSQLQK.Q 100 7.4e-008R.NSNLVGAAHEELQQSR.I 83 4.1e-006R.ITESEEVVSR.E 58 0.0028
a The MS/MS spectra were searched with Mascot search engine.Peptide sequence, Mascot score, and expectation value are reported.
Lamin A Ser404 Is a Target of Akt Phosphorylation research articles
Journal of Proteome Research • Vol. 7, No. 11, 2008 4729

using a linear gradient of acetonitrile/0.1% formic acid from5% to 40% in 30 min. Data were acquired in a data dependentmode: a full MS scan was followed by a Zoom scan and a MS/MS scan on the 3 most intense peaks. MS/MS data weresearched using Mascot (Matrix Science, London, U.K.) againstthe human section of the Swiss-Prot database (version 51.5,149 052 entries). Enzyme specificity was set to trypsin with 1missed cleavage using carbamidomethylcysteine as fixed modi-fication. The tolerance of the precursor ion was set to 1 massunit for both peptide and fragment ion matches.
Akt Kinase Assay and Phosphopeptide Analysis. Lamin A/Cwas immunoprecipiatetd as described above, then phospho-rylated in vitro with 3 µg of recombinant, active Akt1 (∆PH,S473D, kind gift of Dario Alessi, University of Dundee, U.K.) ina buffer comprising 20 mM Tris-Cl, pH 7.5, 25 mM MgCl2, 80mM ATP and 5 µCi/assay (γ-32P)ATP (3000 Ci/mmol), (Amer-
sham-GE Healthcare), and carried out for 20 min at 25 °C.Pellets were then washed and treated as described above.Radioactive bands were localized by autoradiography (Cyclonestorage phosphor system, Packard). For phosphopeptide analy-sis, nitrocellulose membranes from a duplicate experimentwere cut and digested by trypsin. The tryptic peptides weresubjected to acidic hydrolysis (6 M HCl for 4 h at 110 °C) andthe phospho-amino acids were detected by high voltage paperelectrophoresis at pH 1.9. As migration reference, nonradioac-tive phospho-amino acids were added to each sample anddetected by ninhydrin reaction.
Results
Lamin A/C Is a Nuclear Substrate of Akt. Nuclear lysatesfrom C2C12 cells were resolved by 2D-electrophoresis indupicates, using a 3-10 IEF strip for the first dimension. Then,one gel was stained with MS-compatible Coomassie blue(Figure 1a), while the replicate gel was blotted and revealedwith an anti-Akt phosphosubstrate antibody24 to detect putativesubstrates of Akt (Figure 1, panel b). This approach has beenvalidated by previous studies.26,34,35 The overlapping spots wereexcised from the Coomassie-blue stained gel and peptides wereextracted by in-gel digestion for MS analysis. Our attention wasdrawn by two trains of spots (Figure 1b, arrows and asterisks)which, according to MS analysis, corresponded to A-type lamins(Table 1). Consequently, the filter was probed with an anti-lamin A/C antibody which recognizes an N-terminal epitope,common to both lamin A and its splicing product lamin C, thatdiffers from lamin A by a short stretch of amino acids at theC-terminus. The antibody reacted with a doublet at theexpected isoelectric point (6.5-6) and molecular weight oflamin A (72 kDa) and lamin C (68 kDa), confirming the identityof the protein (Figure 1c). Interestingly, a recent study alsoreported lamin A/C as a candidate Akt substrate identified bymass spectrometry upon in vitro phosphorylation of mesangialcell lysate with active recombinant Akt.34
This prompted us to investigate whether Akt and lamin Ainteract in vivo. Therefore, lysates from C2C12 cells wereimmunoprecipitated with anti-lamin A/C, and the pelletsprobed with anti-Akt. The presence of endogenous Akt in theIP suggested that the two proteins interact in vivo (Figure 2a).Lamin A/C is also clearly detectable in anti-Akt immunopre-cipitates. Besides this, the filter was probed with the anti-Aktphosphosubstrate antibody showing that lamin A/C co-immu-noprecipitated with Akt is phosphorylated at a putative Aktmotif (Figure 2a).
293T cells were therefore transiently transfected with a pCI-flagLMNA plasmid, which we have already described to expressboth lamin A (72 kDa) and its precursor prelamin A (74 kDa).30
Then lysates were immunoprecipitated with anti-flag antibodyand the pellets were phosphorylated in vitro in the presenceof rAkt or buffer alone. Pellets were run on SDS-polyacrylamidegel and stained with Coomassie blue. In the sample in whichlamin A has been phosphorylated by rAkt, an additional bandwith a retarded mobility is clearly visible, compatible with theappearance of a phosphorylated form (Figure 2b). Therefore,replicate pellets were run on 2DE, using 5-8 IEF strips,transferred to nitrocellulose and probed with anti-flag. Inagreement with the previous result, the spots indicated as 1 ofprelamin A and 2, 3 of lamin A, upon phosphorylation by Aktare shifted to the left side of the gel, indicating a more acidicpI compatible with a phosphorylation (Figure 2c, asteriks).
Figure 2. Lamin A/C co-immunoprecipitates with Akt. (a) Endog-enous lamin A/C immunoprecipitates (R-lamin A/C IP), andendogenous Akt immunoprecipitates (R-Akt IP), obtained fromC2C12 nuclear lysates, were probed with anti-Akt phosphosub-strate (R-Akt psubstrate WB), then checked for the presence ofAkt (R-Akt WB) and of lamin A/C (R-lamin A/C WB). The result isrepresentative of three independent experiments. (b) 293T cellswere transfected with pCI flag-lamin A, expressing prelamin A(74 kDa) and lamin A (72 kDa), then pellets from anti-flagimmunoprecipitation were phosphorylated in vitro in the pres-ence (+) or not (-) of recombinant Akt (rAkt). Proteins wereresolved by SDS-polyacrylamide gel and stained with colloidalCoomassie blue. An asterisk indicates the band with retardedmobility in the sample phosphorylated by rAkt. (c) Duplicatepellets from the above experiment were resolved by 2D-electro-phoresis on pH 5-8 IEF gradient and an 8% polyacrilammide gel,blotted onto nitrocellulose and incubated with an anti-flagantibody. In the sample phosphorylated by rAkt, spots 1, 2, and3 are clearly shifted toward the acidic side of the gel (marked byasterisks).
research articles Cenni et al.
4730 Journal of Proteome Research • Vol. 7, No. 11, 2008

Akt Phosphorylates Lamin A at Ser404. On the basis of theabove findings, we then asked whether Akt phosphorylatesA-type lamins directly. Therefore, cells were transfected withflag-lamin A, and the anti-flag immunoprecipitate was com-bined with recombinant, active Akt in the presence of γ32P-ATP. The result shows that lamin A can indeed be phospho-rylated by Akt in vitro (Figure 3a). A weak phosphorylation oflamin A is detected in the control sample, most probably dueto endogenous Akt which, as shown above, co-immunopre-cipitates with lamin A. A replicate sample was digested withtrypsin for phosphopeptide analysis. The result plainly indicatesthat Akt phosphorylates lamin A at a serine, while threoninephosphorylation is undetectable (Figure 3b).
In good agreement with the above result, a lamin A sequenceanalysis at high stringency with the scansite algorithm (http://scansite.mit.edu) produced two putative Akt phosphorylationsites, serine 301 and serine 404. Therefore, the correspondingpeptides were synthesized and phosphorylated in vitro withrecombinant Akt. It is clear that both peptides can be phos-phorylated in vitro, though the peptide containing serine 404
to a greater extent (Figure 3c). On this basis, both serines weremutated to alanine, then wild-type, S301A or S404A flag-laminA were expressed into cells, and anti-flag pellets were phos-phorylated in vitro with active, recombinant Akt. The resultclearly shows that lamin A S301A is phosphorylated by Akt tothe same extent as wild-type lamin A. Conversely, Akt failed tophosphorylate lamin A S404A, indicating Ser404 as a genuineAkt phosphorylation site (Figure 3d). It is worth noting thatthe S404-containing Akt motif is evolutionary conserved invertebrate A-type lamins.
Phosphorylation of Lamin A by Akt Is Elicited by Insulin.To delve deeper into lamin A phosphorylation by Akt, weinvestigated whether this event can be evoked in vivo inresponse to physiological stimuli known to activate Akt. To thisgoal, we raised a phosphoantibody against the lamin A GRASp-SHSSQTQGGC peptide phosphorylated at Ser 404. Lamin AS404 phosphorylation was then probed in cells in which Akthad been activated by insulin treatment. As shown in Figure4a, insulin-dependent Akt activation correlated with phospho-rylation of lamin A/C. That phosphorylation of lamin A/C is
Figure 3. Akt phosphorylates lamin A at Ser404. (a) Flag-lamin A overexpressed in 293T cells was immunoprecipitated with an anti-flag antibody, then phosphorylated in vitro in the presence (+) or not (-) of recombinant, active Akt (rAkt) and γ32P-ATP. One samplewas incubated with heat inactivated rAkt. Pellets were resolved on SDS-polyacrylamide gel and blotted, and the radioactive bandswere detected and quantified by autoradiography (upper panel) (Cyclone storage phosphor system, Packard). Next, the filter was probedwith anti-Akt and anti-flag antibodies (lower panel). (b) A duplicate filter was digested with trypsin for phosphopeptide analysis andresolved by high voltage paper electrophoresis at pH 1.9. A phospho-amino acid mixture was run as control. (c) The 0.1 or 0.5 mMpeptides containing either the Ser301 [QSRIRIDSLSAQLSQ(NH2)] peptide, the Ser404 [RSRGRASSHSSQTQG(NH2)] peptide, or theSer404 peptide modified with the mutation R401C [RSRGCASSHSSQTQG(NH2)] were phosphorylated in vitro with rAkt. Crosstidesubstrate peptide (Sigma-Aldrich) was used as positive control. Values are expressed as cpm. (d) Flag-tagged lamin A wild-type (wt),or the lamin mutants S301A and S404A were expressed in 293T cells, immunoprecipitated and subjected to an in vitro kinase assaywith rAkt as described for panel a. As control, filters were probed with anti-flag and anti-Akt antibodies. The data shown are representativeof three independent experiments.
Lamin A Ser404 Is a Target of Akt Phosphorylation research articles
Journal of Proteome Research • Vol. 7, No. 11, 2008 4731

due to Akt was revealed using siRNA against Akt1, the Aktisoform expressed in growing myoblast.36 Reduction of endog-enous Akt1 abrogated the increase of lamin phosphorylationobserved in the presence of insulin alone. The filter was thenprobed with antipoly(ADP-ribose)polymerase (parp), anti-caspase 9 and anti-caspase 3. The absence in all samples ofthe 80 kDa form cleaved parp and 17/19 kDa cleaved caspase3, as well as equal amounts of full-length caspase 9 demonstratecell viability. Moreover, Figure 4b shows that the anti-phos-
pholamin A S404 antibody recognizes both endogenous laminA and flag-lamin A, but not lamin A S404A, immunoprecipitatedfrom cells overexpressing Akt, thus, confirming the antibodyspecificity.
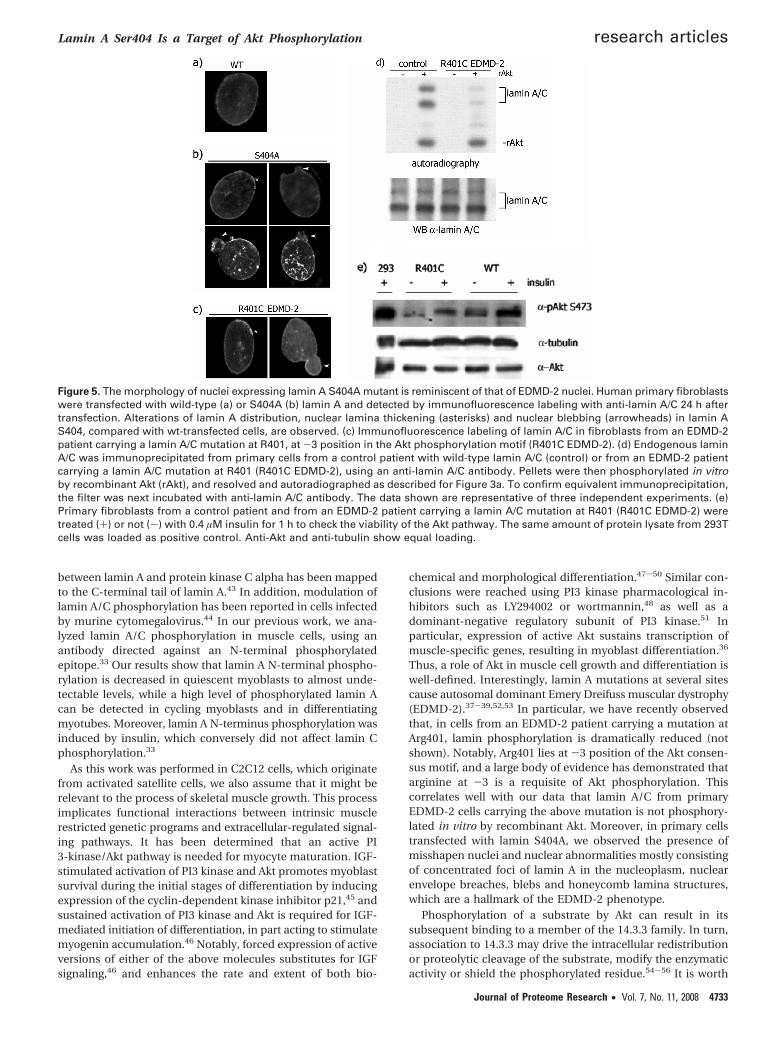
Expression of Lamin A S404A Affects the Nuclear Morphol-ogy. To gain insight into the physiological function of lamin Aphosphorylation by Akt, human primary cells were transfectedwith wild-type or S404A-mutated lamin A (Figure 5a,b) and thenuclear lamina was analyzed by immunofluorescence. Com-pared to primary cells transfected with wild-type lamin A(Figure 5, panel a), cells transfected with lamin A S404A (Figure5, panel b) are featured by the presence of blebs and honey-comb lamina structures reminiscent of the phenotype of thenuclear lamina observed in Emery-Dreifuss muscular dystrophy(EDMD).37-39 Indeed, it is well-known that mutations at severalsites in lamin A cause autosomal dominant EDMD (EDMD-2).33 To investigate the extent of lamin A S404 phosphorylationin EDMD-2 cells, we obtained primary fibroblasts from anEDMD-2 patient carrying a mutation at R401 (R401C laminA/C), in the Akt phosphorylation motif. It is worth noting thatarginine at -3 position is regarded as a minimal requirementfor Akt phosphorylation.16 Observed by immunofluorescence,these cells exhibit disorganization of the nuclear lamina typicalof the EDMD-2 cells reported above in 10% of nuclei (Figure5c), as well as focal loss of peripheral heterochromatin (ourunpublished results).
Therefore, endogenous lamin A/C was immunoprecipitatedfrom either EDMD-2 cells or from control primary cells, thenphosphorylated in vitro by recombinant Akt. Clearly, the extentof Akt phosphorylation of lamin A/C R401C was greatly reduced(Figure 5d). To rule out that the low level of phosphorylationof lamin A/C is due to down-regulation of Akt itself in thesecells, EDMD-2 cells were treated with insulin and Akt activitywas detected by anti-Akt phosphoserine-473 antibody (Figure5e). Moreover, the R401C peptide was phosphorylated in vitrowith recombinant Akt (Figure 3c). As expected, in conditionsin which both S301 and S404 peptides were phosphorylatedby Akt, phosphorylation of the R401C peptide was undetectable.
Discussion
The identification of genuine substrates of kinases is re-garded as an important issue to the complete dissection of anysignaling pathway. Given the important roles of Akt in bothcell metabolism, proliferation and survival, and its potentialas therapeutic target, we have undertaken a search for newsubstrates of Akt by a functional proteomic approach, com-bining high-throughput 2DE/MS with anti-phosphomotif an-tibodies and bioinformatic search with algorithms for thedetection of phosphomotifs associated with specific kinases.Here we present evidence of lamin A/C as a novel nuclearsubstrate of Akt. By coimmunoprecipitation, we demonstratedthat endogenous lamin A/C and Akt proteins interact, and thatlamin A/C is phosphorylated by Akt both in vitro and in vivo.Moreover, by phospho-amino acid analysis and mutagenesis,we further demonstrated that Akt phosphorylates lamin A atSer404, in the evolutionary conserved RSRGRASSH Akt motif.Lamin phosphorylation is a key event in the mitotic breakdownof the nuclear lamina.40 In cells undergoing mitosis, cyclin-dependent kinases mediate phosphorylation of lamins A, B andC.41 Moreover, interphase phosphorylation of lamins A and Cwas reported following insulin treatment of quiescent fibro-blasts.42 Phosphorylation of lamins by PKC in interphase cellshas also been described, and the site of molecular interaction
Figure 4. Lamin A S404 phosphorylation is triggered by insulin.(a) C2C12 cells were transfected for 48 h with either pU6-Akt1siRNA, pU6-scramble siRNA or pU6 empty vector (ev), togetherwith pEGFP to monitor tansfection, using the Cell Line Nucleo-fector kit V and Amaxa Nucleoporator, serum starved for 16 hthen treated for 1 h with 0.4 µM insulin. Endogenous lamin A/Cwas immunoprecipitated from whole cell lysates and the filterrevealed with anti-phospholamin S404 (pS404), then with anti-lamin A/C. Whole lysates were probed with anti-phosphoAktSer473 (pAkt) and anti-Akt1 to confirm siRNA efficacy. Equivalentamounts of uncleaved poly(ADP-ribose)polymerase (parp, 113kDa, Santa Cruz Biotech) and caspase 9 (49 kDa, Cell Signaling#9504) demonstrate cell viability. Since anti-caspase 3 (CellSignalling #9661) detects only the large fragment (17/19 kDa) ofcleaved caspse-3, Jurkat cells treated with 5 µM sodium arsenite(As2O3) for 1 day were loaded as positive control (As2O3, lane5); (b) Cells were co-transfected with caax-Akt and either emptyvector (ev), lamin A wild-type (wt) or lamin A S404A, serumstarved for 16 h then treated for 1 h with 0.4 µM insulin and equalamounts of proteins were probed with anti-phospholamin S404(pS404). The absence of labeling at the expected molecularweight in the lamin A S404A sample confirms the antibodyspecificity.
research articles Cenni et al.
4732 Journal of Proteome Research • Vol. 7, No. 11, 2008
between lamin A and protein kinase C alpha has been mappedto the C-terminal tail of lamin A.43 In addition, modulation oflamin A/C phosphorylation has been reported in cells infectedby murine cytomegalovirus.44 In our previous work, we ana-lyzed lamin A/C phosphorylation in muscle cells, using anantibody directed against an N-terminal phosphorylatedepitope.33 Our results show that lamin A N-terminal phospho-rylation is decreased in quiescent myoblasts to almost unde-tectable levels, while a high level of phosphorylated lamin Acan be detected in cycling myoblasts and in differentiatingmyotubes. Moreover, lamin A N-terminus phosphorylation wasinduced by insulin, which conversely did not affect lamin Cphosphorylation.33
As this work was performed in C2C12 cells, which originatefrom activated satellite cells, we also assume that it might berelevant to the process of skeletal muscle growth. This processimplicates functional interactions between intrinsic musclerestricted genetic programs and extracellular-regulated signal-ing pathways. It has been determined that an active PI3-kinase/Akt pathway is needed for myocyte maturation. IGF-stimulated activation of PI3 kinase and Akt promotes myoblastsurvival during the initial stages of differentiation by inducingexpression of the cyclin-dependent kinase inhibitor p21,45 andsustained activation of PI3 kinase and Akt is required for IGF-mediated initiation of differentiation, in part acting to stimulatemyogenin accumulation.46 Notably, forced expression of activeversions of either of the above molecules substitutes for IGFsignaling,46 and enhances the rate and extent of both bio-
chemical and morphological differentiation.47-50 Similar con-clusions were reached using PI3 kinase pharmacological in-hibitors such as LY294002 or wortmannin,48 as well as adominant-negative regulatory subunit of PI3 kinase.51 Inparticular, expression of active Akt sustains transcription ofmuscle-specific genes, resulting in myoblast differentiation.36
Thus, a role of Akt in muscle cell growth and differentiation iswell-defined. Interestingly, lamin A mutations at several sitescause autosomal dominant Emery Dreifuss muscular dystrophy(EDMD-2).37-39,52,53 In particular, we have recently observedthat, in cells from an EDMD-2 patient carrying a mutation atArg401, lamin phosphorylation is dramatically reduced (notshown). Notably, Arg401 lies at -3 position of the Akt consen-sus motif, and a large body of evidence has demonstrated thatarginine at -3 is a requisite of Akt phosphorylation. Thiscorrelates well with our data that lamin A/C from primaryEDMD-2 cells carrying the above mutation is not phosphory-lated in vitro by recombinant Akt. Moreover, in primary cellstransfected with lamin S404A, we observed the presence ofmisshapen nuclei and nuclear abnormalities mostly consistingof concentrated foci of lamin A in the nucleoplasm, nuclearenvelope breaches, blebs and honeycomb lamina structures,which are a hallmark of the EDMD-2 phenotype.
Phosphorylation of a substrate by Akt can result in itssubsequent binding to a member of the 14.3.3 family. In turn,association to 14.3.3 may drive the intracellular redistributionor proteolytic cleavage of the substrate, modify the enzymaticactivity or shield the phosphorylated residue.54-56 It is worth
Figure 5. The morphology of nuclei expressing lamin A S404A mutant is reminiscent of that of EDMD-2 nuclei. Human primary fibroblastswere transfected with wild-type (a) or S404A (b) lamin A and detected by immunofluorescence labeling with anti-lamin A/C 24 h aftertransfection. Alterations of lamin A distribution, nuclear lamina thickening (asterisks) and nuclear blebbing (arrowheads) in lamin AS404, compared with wt-transfected cells, are observed. (c) Immunofluorescence labeling of lamin A/C in fibroblasts from an EDMD-2patient carrying a lamin A/C mutation at R401, at -3 position in the Akt phosphorylation motif (R401C EDMD-2). (d) Endogenous laminA/C was immunoprecipitated from primary cells from a control patient with wild-type lamin A/C (control) or from an EDMD-2 patientcarrying a lamin A/C mutation at R401 (R401C EDMD-2), using an anti-lamin A/C antibody. Pellets were then phosphorylated in vitroby recombinant Akt (rAkt), and resolved and autoradiographed as described for Figure 3a. To confirm equivalent immunoprecipitation,the filter was next incubated with anti-lamin A/C antibody. The data shown are representative of three independent experiments. (e)Primary fibroblasts from a control patient and from an EDMD-2 patient carrying a lamin A/C mutation at R401 (R401C EDMD-2) weretreated (+) or not (-) with 0.4 µM insulin for 1 h to check the viability of the Akt pathway. The same amount of protein lysate from 293Tcells was loaded as positive control. Anti-Akt and anti-tubulin show equal loading.
Lamin A Ser404 Is a Target of Akt Phosphorylation research articles
Journal of Proteome Research • Vol. 7, No. 11, 2008 4733

noting that, according to the scansite algorithm, phospho-Ser404 represents a good 14.3.3 phosphomotif, suggesting thatAkt phosphorylation of lamin A at Ser404 creates putativebinding sites for 14.3.3 proteins. Remarkably, a recently pub-lished proteomic analysis of 14.3.3 binding partner demon-strates that lamin A associates directly with 14.3.3 proteins inmitotic cells.57 Thus, it is conceivable that failure to phospho-rylate Ser404 might prevent binding to 14.3.3, thus, affectingeither stability or localization of lamin A. Though we have noevidence at present that 14.3.3 binding can affect the correctintranuclear location of lamin A, it has been previously shownthat phosphorylation at Ser403 and Ser404, which lie inproximity to the nuclear localization signal, can regulate thecorrect positioning of human lamin A/C into the nucleus.58 Onthe other hand, Ser404 phosphorylation might control laminA stability, hence affecting its ability to form a functionalscaffold or a correct compartmentalization of chromatin. In thisregard, it is worth noting that a disorganization of the nuclearlamina as well as focal loss of peripheral heterochromatin inthe cells carrying the Arg401 mutant lamin has been describedrecently. Moreover, we have shown previously that in musclebiopsies from EDMD-2 patients the extent of lamin phospho-rylation is dramatically reduced compared to the health tis-sue.33
In summary, the results presented here establish lamin A asa genuine nuclear substrate of Akt, phosphorylated in responseto physiological stimuli at a precise residue, and pave the wayfor a functional link between a central nuclear structuralcomponent, lamin A, and an essential regulator of multiplesignaling pathways, Akt, for the first time implicating a phos-phorylation defect of the nuclear lamina in muscle diseasepathologies.
Acknowledgment. This work was supported by thefollowing grants: FAR-University of Modena and ReggioEmilia; MIUR (PRIN 2005 grant 2005055737, FIRB inter-nazionalizzazione grant RBIN04NR2S_001; FIRB ItalianHuman Proteome Net.); EU FP6“Euro-laminopathies” grantn 018690; Italian ISS “Rare diseases Italy-USA program”grant n 526/D30; Fondazione Cassa Risparmio Modena. Wethank Dario Alessi for providing reagents.
References(1) Bellacosa, A.; Testa, J. R.; Staal, S. P.; Tsichlis, P. N. A retroviral
oncogene, akt, encoding a serine-threonine kinase containing anSH2-like region. Science 1991, 254, 274–277.
(2) Staal, G. E.; Kalff, A.; Heesbeen, E. C.; van Veelen, C. W.; Rijksen,G. Subunit composition, regulatory properties, and phosphoryla-tion of phosphofructokinase from human gliomas. Cancer Res.1987, 47, 5047–5051.
(3) Jones, P. F.; Jakubowicz, T.; Pitossi, F. J.; Maurer, F.; Hemmings,B. A. ) Molecular cloning and identification of a serine/threonineprotein kinase of the second-messenger subfamily. Proc. Natl.Acad. Sci. U.S.A. 1991, 88, 4171–4175.
(4) Franke, T.; Kaplan, R. D.; Cantley, L. C.; Toker, A. Direct regulationof the Akt proto-oncogene product by phosphatidylinositol-3,4-bisphosphate. Science 1997, 275, 665–668.
(5) Stokoe, D.; Stephens, L. R.; Copeland, T.; Gaffney, P. R. J.; Painter,G. F.; Holmes, A. B.; McCormick, F.; Hawkins, P. T. Dual role ofphosphatidylinositol-3,4,5-trisphosphate in the activation of pro-tein kinase B. Science 1997, 277, 567–570.
(6) Yang, Z. Z.; Tschopp, O.; Baudry, A.; Dummler, B.; Hynx, D.;Hemmings, B. A. Physiological functions of protein kinase B/Akt.Biochem. Soc. Trans. 2004, 32, 350–354.
(7) Andjelkovic, M.; Alessi, D. R.; Meier, R.; Fernandez, A.; Lamb, N. J.;Frech, M.; Cron, P.; Cohen, P.; Lucocq, J. M.; Hemmings, B. A.Role of translocation in the activation and function of proteinkinase B. J. Biol. Chem. 1997, 272, 31515–31524.
(8) Alessi, D. R.; James, S. R.; Downes, C. P.; Holmes, A. B.; Gaffney,P. R.; Reese, C. B.; Cohen, P. Characterization of a 3-phosphoi-nositide-dependent protein kinase which phosphorylates andactivates protein kinase Balpha. Curr. Biol. 1997, 7 (4), 261–269.
(9) Toker, A.; Newton, A. C. Akt/protein kinase B is regulated byautophosphorylation at the hypothetical PDK-2 site. J. Biol. Chem.2000, 275, 8271–8274.
(10) Persad, S.; Attwell, S.; Gray, V.; Mawji, N.; Deng, J. T.; Leung, D.;Yan, J.; Sanghera, J.; Walsh, M. P.; Dedhar, S. Regulation of proteinkinase B/Akt-serine 473 phosphorylation by integrin-linked kinase:critical roles for kinase activity and amino acids arginine 211 andserine 343. J. Biol. Chem. 2001, 276, 27462–27469.
(11) Feng, J.; Park, J.; Cron, P.; Hess, D.; Hemmings, B. A. Identificationof a PKB/Akt hydrophobic motif Ser-473 kinase as DNA-dependentprotein kinase. J. Biol. Chem. 2004, 279, 41189–41196.
(12) Scheid, M. P.; Marignani, P. A.; Woodgett, J. R. Multiple phos-phoinositide 3-kinase-dependent steps in activation of proteinkinase B. Mol. Cell. Biol. 2002, 22, 6247–6260.
(13) Sarbassov, D. D.; Guertin, D. A.; Siraj, M. A.; Sabatini, D. M.Phosphorylation and regulation of Akt/PKB by the rictor-mTORcomplex. Science 2005, 307, 1098–1101.
(14) Gao, T.; Furnari, F.; Newton, A. C. PHLPP: a phosphatase thatdirectly dephosphorylates Akt, promotes apoptosis, and suppressestumor growth. Mol. Cell 2005, 18, 13–24.
(15) Brognard, J.; Sierecki, E.; Gao, T.; Newton, A. C. PHLPP and asecond isoform, PHLPP2, differentially attenuate the amplitudeof Akt signaling by regulating distinct Akt isoforms. Mol. Cell 2007,25, 917–931.
(16) Manning, B. D.; Cantley, L. C. AKT/PKB signaling: navigatingdownstream. Cell 2007, 129, 1261–1274.
(17) Martelli, A. M.; Nyåkern, M.; Tabellini, G.; Bortul, R.; Tazzari, P. L.;Evangelisti, C.; Cocco, L. Phosphoinositide 3-kinase/Akt signalingpathway and its therapeutical implications for human acutemyeloid leukemia. Leukemia 2006, 20, 911–928.
(18) Kunkel, M. T.; Tsien, R. Y.; Zhang, J.; Newton, A. C. Spatio-temporaldynamics of protein kinase B/Akt signaling revealed by a geneti-cally encoded fluorescent reporter. J. Biol. Chem. 2005, 280, 5581–5587.
(19) Zhu, L.; Hu, C.; Li, J.; Xue, P.; He, X.; Ge, C.; Qin, W.; Yao, G.; Gu,J. Real-time imaging nuclear translocation of Akt1 in HCC cells.Biochem. Biophys. Res. Commun. 2007, 356, 1038–1043.
(20) Cenni, V.; Sirri, A.; Riccio, M.; Lattanzi, G.; Santi, S.; de Pol, A.;Maraldi, N. M.; Marmiroli, S. Targeting of the Akt/PKB kinase tothe actin skeleton. Cell. Mol. Life Sci. 2003, 60, 2710–2720.
(21) Laine, J.; Kunstle, G.; Obata, T.; Sha, M.; Noguchi, M. Differentialregulation of Akt kinase isoforms by the members of the TCL1oncogene family. Mol. Cell 2000, 6, 395–407.
(22) Brunet, A.; Bonni, A.; Zigmond, M. J.; Lin, M. Z.; Juo, P.; Hu, L. S.;Anderson, M. J.; Arden, K. C.; Blenis, J.; Greenberg, M. E. Aktpromotes cell survival by phosphorylating and inhibiting a Fork-head transcription factor. Cell 1999, 96, 857–868.
(23) Hribal, M. L.; Nakae, J.; Kitamura, T.; Shutter, J. R.; Accili, D.Regulation of insulin-like growth factor-dependent myoblast dif-ferentiation by Foxo forkhead transcription factors. J. Cell. Biol.2003, 162, 535–541.
(24) Zhang, H.; Zha, X.; Tan, Y.; Hornbeck, P. V.; Mastrangelo, A. J.;Alessi, D. R.; Polakiewicz, R. D.; Comb, M. J. Phosphoproteinanalysis using antibodies broadly reactive against phosphorylatedmotifs. J. Biol. Chem. 2002, 277, 39379–39387.
(25) Manning, B. D.; Tee, A. R.; Logsdon, M. N.; Blenis, J.; Cantley, L. C.Identification of the tuberous sclerosis complex-2 tumor suppres-sor gene product tuberin as a target of the phosphoinositide3-kinase/akt pathway. Mol. Cell 2002, 10, 151–162.
(26) Konishi, H.; Namikawa, K.; Shikata, K.; Kobatake, Y.; Tachibana,T.; Kiyama, H. Identification of peripherin as a Akt substrate inneurons. J. Biol. Chem. 2007, 282, 23491–23499.
(27) Hutchison, C. J.; Worman, H. J. A-type lamins: guardians of thesoma. Nat. Cell Biol. 2004, 6, 1062–1067.
(28) Shumaker, D. K.; Kuczmarski, E. R.; Goldman, R. D. The nucleosk-eleton: lamins and Actin are major players in essential nuclear.Curr. Opin. Cell. Biol. 2003, 15, 358–366.
(29) Bridger, J. M.; Foeger, N.; Kill, I. R.; Herrmann, H. The nuclearlamina. Both a structural framework and a platform for genomeorganization. FEBS J. 2007, 274, 1354–1361.
(30) Kumaran, R. I.; Muralikrishna, B.; Parnaik, V. K. Lamin A/C specklesmediate spatial organization of splicing factor compartments andRNA polymerase II transcription. J. Cell Biol. 2002, 159, 783–793.
(31) Vojtek, A. B.; Taylor, J.; DeRuiter, S. L.; Yu, J. Y.; Figueroa, C.; Kwok,R. P.; Turner, D. L. Akt regulates basic helix-loop-helix transcriptionfactor-coactivator complex formation and activity during neuronaldifferentiation. Mol. Cell. Biol. 2003, 23, 4417–4427.
research articles Cenni et al.
4734 Journal of Proteome Research • Vol. 7, No. 11, 2008

(32) Cenni, V.; Sirri, A.; De Pol, A.; Maraldi, N. M.; Marmiroli, S.Interleukin-1-receptor-associated kinase 2 (IRAK2)-mediated in-terleukin-1-dependent nuclear factor kappaB transactivation inSaos2 cells requires the Akt/protein kinase B kinase. Biochem. J.2003, 376, 303–311.
(33) Cenni, V.; Sabatelli, P.; Mattioli, E.; Marmiroli, S.; Capanni, C.;Ognibene, A.; Squarzoni, S.; Maraldi, N. M.; Bonne, G.; Columbaro,M.; Merlini, L.; Lattanzi, G. J. Lamin A N-terminal phosphorylationis associated with myoblast activation: impairment in Emery-Dreifuss muscular dystrophy. J. Med. Genet. 2005, 42, 214–220.
(34) Barati, M. T.; Rane, M. J.; Klein, J. B.; McLeish, K. R. A proteomicscreen identified stress-induced chaperone proteins as targets ofAkt phosphorylation in mesangial cells. J. Proteome Res. 2006, 5,1636–1646.
(35) Vandermoere, F.; El Yazidi-Belkoura, I.; Demont, Y.; Slomianny,C.; Antol, J.; Lemoine, J.; Hondermarck, H. Proteomics explorationreveals that Actin is a signaling target of the kinase Akt. Mol. Cell.Proteomics 2007, 6, 114–124.
(36) Wilson, E. M.; Rotwein, P. Selective control of skeletal muscledifferentiation by Akt1. J. Biol. Chem. 2007, 282, 5106–5110.
(37) Muchir, A.; Worman, H. J. Emery-Dreifuss muscular dystrophy.Curr. Neurol. Neurosci. Rep. 2007, 7, 78–83.
(38) Gruenbaum, Y.; Margalit, A.; Goldman, R. D.; Shumaker, D. K.;Wilson, K. L. The nuclear lamina comes of age. Nat. Rev. Mol. Cell.Biol. 2005, 6, 21–31.
(39) Bonne, G.; Di Barletta, M. R.; Varnous, S.; Becane, H. M.; Han-nouda, E. H.; Merlini, L.; Muntoni, F.; Greenberg, C. R.; Gary, F.;Urtizberea, J. A. Mutations in the gene encoding lamin A/C causeautosomal dominant Emery-Dreifuss muscular dystrophy. Nat.Genet. 1999, 21, 285–288.
(40) Lee, K. K.; Gruenbaum, Y.; Spann, P.; Liu, J.; Wilson, K. L. C. elegansnuclear envelope proteins emerin, MAN1, lamin, and nucleoporinsreveal unique timing of nuclear envelope breakdown duringmitosis. Mol. Biol. Cell 2000, 11, 3089–3099.
(41) Buendia, B.; Courvalin, J. C.; Collas, P. Dynamics of the nuclearenvelope at mitosis and during apoptosis. Cell. Mol. Life Sci. 2001,58, 1781–1789.
(42) Friedman, D,L.; Ken, R. Insulin stimulates incorporation of 32Piinto nuclear lamins A and C in quiescent BHK-21 cells. J. Biol.Chem. 1988, 263, 1103–1106.
(43) Martelli, A. M.; Bortul, R.; Tabellini, G.; Faenza, I.; Cappellini, A.;Bareggi, R.; Manzoli, L.; Cocco, L. Molecular characterization ofprotein kinase C-alpha binding to lamin A. J. Cell Biochem. 2002,86, 320–330.
(44) Muranyi, W.; Haas, J.; Wagner, M.; Krohne, G.; Koszinowski, U. H.Cytomegalovirus recruitment of cellular kinases to dissolve thenuclear lamina. Science 2002, 297, 854–857.
(45) Lawlor, M. A.; Rotwein, P. Insulin-like growth factor-mediatedmuscle cell survival: central roles for Akt and cyclin-dependentkinase inhibitor p21. Mol. Cell. Biol. 2000, 20, 8983–8995.
(46) Wilson, E. M.; Tureckova, J.; Rotwein, P. Permissive roles ofphosphatidyl inositol 3-kinase and Akt in skeletal myocyte matura-tion. Mol. Biol. Cell 2004, 15, 497–505.
(47) Xu, Q.; Wu, Z. The insulin-like growth factor-phosphatidylinositol3-kinase-Akt signaling pathway regulates myogenin expression innormal myogenic cells but not in rhabdomyosarcoma-derived RDcells. J. Biol. Chem. 2000, 275, 36750–36757.
(48) Jiang, B. H.; Aoki, M.; Zheng, J. Z.; Li, J.; Vogt, P. K. Myogenicsignaling of phosphatidylinositol 3-kinase requires the serine-threonine kinase Akt/protein kinase B. Proc. Natl. Acad. Sci. U.S.A.1999, 96, 2077–2081.
(49) Rommel, C.; Bodine, S. C.; Clarke, B. A.; Rossman, R.; Nunez, L.;Stitt, T. N.; Yancopoulos, G. D.; Glass, D. J. Mediation of IGF-1-induced skeletal myotube hypertrophy by PI(3)K/Akt/mTOR andPI(3)K/Akt/GSK3 pathways. Nat. Cell Biol. 2001, 3, 1009–1013.
(50) Takahashi, A.; Kureishi, Y.; Yang, J.; Luo, Z.; Guo, K.; Mukho-padhyay, D.; Ivashchenko, Y.; Branellec, D.; Walsh, K. MyogenicAkt signaling regulates blood vessel recruitment during myofibergrowth. Mol. Cell. Biol. 2002, 22, 4803–4814.
(51) Tamir, Y.; Bengal, E. Phosphoinositide 3-kinase induces thetranscriptional activity of MEF2 proteins during muscle dif-ferentiation. J. Biol. Chem. 2000, 275, 34424–34432.
(52) Maraldi, N. M.; Lattanzi, G. Involvement of prelamin A in lami-nopathies. Crit. Rev. Eukaryotic Gene Expression 2007, 317, 17–34.
(53) Maraldi, N. M.; Squarzoni, S.; Sabatelli, P.; Capanni, C.; Mattioli,E.; Ognibene, A.; Lattanzi, G. Laminopathies: involvement ofstructural nuclear proteins in the pathogenesis of an increasingnumber of human diseases. J. Cell Physiol. 2005, 203, 319–327.
(54) Yaffe, M. B. How do 14-3-3 proteins work? Gatekeeper phospho-rylation and the molecular anvil hypothesis. FEBS Lett. 2002, 513,53–57.
(55) Aitken, A. 14-3-3 proteins: a historic overview. Semin. Cancer Biol.2006, 16, 162–172.
(56) Bridges, D.; Moorhead, G. B. 14-3-3 Proteins: a number of functionsfor a numbered protein. Sci. STKE 2005, 296, re10.
(57) Meek, S. E.; Lane, W. S.; Piwnica-Worms, H. M. Comprehensiveproteomic analysis of interphase and mitotic 14-3-3-bindingproteins. J. Biol. Chem. 2004, 279, 32046–32054.
(58) Leukel, M.; Jost, E. Two conserved serines in the nuclear localiza-tion signal flanking region are involved in the nuclear targeting ofhuman lamin A. Eur. J. Cell. Biol. 1995, 68, 133–142.
(59) Capanni, C.; Mattioli, E.; Columbaro, M.; Lucarelli, E.; Parnaik,V. K.; Novelli, G.; Wehnert, M.; Cenni, V.; Maraldi, N. M.; Squarzoni,S.; Lattanzi, G. Altered pre-lamin A processing is a commonmechanism leading to lipodystrophy. Hum. Mol. Genet. 2005, 14,1489–1502.
PR800262G
Lamin A Ser404 Is a Target of Akt Phosphorylation research articles
Journal of Proteome Research • Vol. 7, No. 11, 2008 4735
Copyright © 2022 FDOKUMEN




















