
Cell Nuclei Spin in the Absence of Lamin B1
12
Cell Nuclei Spin in the Absence of Lamin B1 * □ S Received for publication, December 4, 2006, and in revised form, May 1, 2007 Published, JBC Papers in Press, May 8, 2007, DOI 10.1074/jbc.M611094200 Julie Y. Ji ‡ , Richard T. Lee ‡ , Laurent Vergnes § , Loren G. Fong ¶ , Colin L. Stewart , Karen Reue § , Stephen G. Young §¶ , Qiuping Zhang**, Catherine M. Shanahan**, and Jan Lammerding ‡1 From the ‡ Cardiovascular Division, Brigham and Women’s Hospital, Cambridge, Massachusetts 02139, the Laboratory of Cancer and Developmental Biology, NCI, National Institutes of Health, Frederick, Maryland 21702, the ¶ Department of Medicine, University of California, Los Angeles, California 90095, the § Department of Human Genetics, University of California, Los Angeles, California 90073, and the **Department of Medicine, Division of Cardiovascular Medicine, University of Cambridge , Box 110, Addenbrooke’s Hospital, Hills Road, Cambridge CB2 2QQ, United Kingdom Mutations of the nuclear lamins cause a wide range of human diseases, including Emery-Dreifuss muscular dystrophy and Hutchinson-Gilford progeria syndrome. Defects in A-type lamins reduce nuclear structural integrity and affect transcrip- tional regulation, but few data exist on the biological role of B-type lamins. To assess the functional importance of lamin B1, we examined nuclear dynamics in fibroblasts from Lmnb1 / and wild-type littermate embryos by time-lapse videomicros- copy. Here, we report that Lmnb1 / cells displayed striking nuclear rotation, with 90% of Lmnb1 / nuclei rotating at least 90° during an 8-h period. The rotation involved the nuclear interior as well as the nuclear envelope. The rotation of nuclei required an intact cytoskeletal network and was eliminated by expressing lamin B1 in cells. Nuclear rotation could also be abol- ished by expressing larger nesprin isoforms with long spectrin repeats. These findings demonstrate that lamin B1 serves a fun- damental role within the nuclear envelope: anchoring the nucleus to the cytoskeleton. Mutations in nuclear lamins and lamin-associated proteins cause a panoply of human diseases (laminopathies) including Emery-Dreifuss muscular dystrophy, dilated cardiomyopathy with conduction system disease, Dunnigan-type familial partial lipodystrophy, limb-girdle muscular dystrophy, Charcot-Marie tooth disorder type 2, and Hutchinson-Gilford progeria syn- drome (1–7). The mechanism by which these ubiquitously expressed proteins cause such diverse phenotypes is unclear. This is not particularly surprising, though, because the func- tions of the nuclear lamins themselves are currently incom- pletely understood. Lamins are the principal components of the nuclear lamina, an intermediate filament meshwork that lines the inner nuclear membrane (8, 9). Lamins are associated with chromatin, other integral proteins of the inner nuclear membrane, inner portions of the nuclear pore complexes (NPCs), 2 and several transcrip- tion factors such as SREBP1, retinoblastoma protein, and MOK (10). Thus, lamins are critical for the structural integrity of the nucleus and also play a role in DNA replication, chromatin organization, and transcriptional regulation (11). In mammalian cells, the major A-type lamins, lamins A and C, are alternatively spliced products of LMNA, whereas the major B-type lamins, lamin B1 and lamin B2, are encoded by two distinct genes, LMNB1 and LMNB2, respectively (12). Although the A- and B-type lamins share a similar structure, they differ in their behavior during cell division and their pat- terns of expression. B-type lamins are found in all cell types and are expressed throughout development, whereas A-type lamins are not present in early embryos (13). Within the nucleus, lamin B1 binds directly to chromatin and histones (14) and interacts with several chromatin-binding inner nuclear membrane pro- teins (e.g. lamina-associated proteins, lamin B receptor (LBR), and the nuclear pore protein nucleoporin 153) (15). Following mitosis, B-type lamins assemble first into the nuclear lamina, followed by lamin A, and subsequently by lamin C (16). Few data exist on the biological role of the B-type lamins. B-type lamins may have a direct role in DNA synthesis (16), and silenc- ing of lamin B by siRNA causes cell death in human cells and in Caenorhabditis elegans (17, 18). For these reasons, B-type lamins are generally assumed to be essential. Mice deficient in lamin B1 (Lmnb1 / ), which were created with a gene-trap embryonic stem cell line, die in the perinatal period with defects in lung and bone (19). Lmnb1 / embryonic fibroblasts display nuclear shape abnormalities, chromosomal abnormalities, and impaired differentiation into adipocytes (19). In humans, dupli- cation of LMNB1 causes autosomal dominant leukodystrophy (20). To further elucidate the function of lamin B1, we decided to examine nuclear shape and dynamics in fibroblasts from Lmnb1 / and wild-type embryos by quantitative time-lapse videomicroscopy. We made a stunning observation: nuclei spin in the absence of lamin B1, and we show that this nuclear rota- tion could be rescued by transfection with a GFP-lamin B1 * This work was supported by National Institutes of Health Grants HL073809, HL64858, AR050200, and HL082792, the American Heart Association Grant 0635359N, the Progeria Research Foundation, March of Dimes, and a NRSA postdoctoral fellowship HL079862 (to J. Y. J.). The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. □ S The on-line version of this article (available at http://www.jbc.org) contains supplemental videos S1–S15, Figs. S1–S3, and additional text. 1 To whom correspondence should be addressed: Partners Research Facility- Rm. 283, 65 Landsdowne St., Cambridge, MA 02139. Tel.: 617-768-8273; Fax: 617-768-8280; E-mail: [email protected]. 2 The abbreviations used are: NPC, nuclear pore complex; MEF, mouse embryo fibroblast; LBR, lamin B receptor; KASH, Klarsicht/ANC-1/Syne-1 homology; GFP, green fluorescent protein; EGFP, enhanced GFP; RFP, red fluorescent protein; ER, endoplasmic reticulum; Az, sodium azide; DOG, 2-deoxyglucose; Bis-Tris, 2-[bis(2-hydroxyethyl)amino]-2-(hydroxymethyl)- propane-1,3-diol. THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 282, NO. 27, pp. 20015–20026, July 6, 2007 Printed in the U.S.A. JULY 6, 2007 • VOLUME 282 • NUMBER 27 JOURNAL OF BIOLOGICAL CHEMISTRY 20015 at Massachusetts General Hospital Treadwell Library on July 1, 2007 www.jbc.org Downloaded from http://www.jbc.org/cgi/content/full/M611094200/DC1 Supplemental Material can be found at:
Transcript of Cell Nuclei Spin in the Absence of Lamin B1

Cell Nuclei Spin in the Absence of Lamin B1*□S
Received for publication, December 4, 2006, and in revised form, May 1, 2007 Published, JBC Papers in Press, May 8, 2007, DOI 10.1074/jbc.M611094200
Julie Y. Ji‡, Richard T. Lee‡, Laurent Vergnes§, Loren G. Fong¶, Colin L. Stewart�, Karen Reue§, Stephen G. Young§¶,Qiuping Zhang**, Catherine M. Shanahan**, and Jan Lammerding‡1
From the ‡Cardiovascular Division, Brigham and Women’s Hospital, Cambridge, Massachusetts 02139, the �Laboratory of Cancerand Developmental Biology, NCI, National Institutes of Health, Frederick, Maryland 21702, the ¶Department of Medicine,University of California, Los Angeles, California 90095, the §Department of Human Genetics, University of California,Los Angeles, California 90073, and the **Department of Medicine, Division of Cardiovascular Medicine,University of Cambridge , Box 110, Addenbrooke’s Hospital, Hills Road, Cambridge CB2 2QQ, United Kingdom
Mutations of the nuclear lamins cause a wide range of humandiseases, including Emery-Dreifuss muscular dystrophy andHutchinson-Gilford progeria syndrome. Defects in A-typelamins reduce nuclear structural integrity and affect transcrip-tional regulation, but few data exist on the biological role ofB-type lamins. To assess the functional importance of lamin B1,we examined nuclear dynamics in fibroblasts from Lmnb1�/�
and wild-type littermate embryos by time-lapse videomicros-copy. Here, we report that Lmnb1�/� cells displayed strikingnuclear rotation, with �90% of Lmnb1�/� nuclei rotating atleast 90° during an 8-h period. The rotation involved the nuclearinterior as well as the nuclear envelope. The rotation of nucleirequired an intact cytoskeletal network and was eliminated byexpressing laminB1 in cells.Nuclear rotation could also be abol-ished by expressing larger nesprin isoforms with long spectrinrepeats. These findings demonstrate that lamin B1 serves a fun-damental role within the nuclear envelope: anchoring thenucleus to the cytoskeleton.
Mutations in nuclear lamins and lamin-associated proteinscause a panoply of human diseases (laminopathies) includingEmery-Dreifuss muscular dystrophy, dilated cardiomyopathywith conduction system disease, Dunnigan-type familial partiallipodystrophy, limb-girdlemuscular dystrophy, Charcot-Marietooth disorder type 2, and Hutchinson-Gilford progeria syn-drome (1–7). The mechanism by which these ubiquitouslyexpressed proteins cause such diverse phenotypes is unclear.This is not particularly surprising, though, because the func-tions of the nuclear lamins themselves are currently incom-pletely understood.Lamins are the principal components of the nuclear lamina,
an intermediate filamentmeshwork that lines the inner nuclearmembrane (8, 9). Lamins are associated with chromatin, other
integral proteins of the inner nuclearmembrane, inner portionsof the nuclear pore complexes (NPCs),2 and several transcrip-tion factors such as SREBP1, retinoblastoma protein, andMOK(10). Thus, lamins are critical for the structural integrity of thenucleus and also play a role in DNA replication, chromatinorganization, and transcriptional regulation (11).In mammalian cells, the major A-type lamins, lamins A and
C, are alternatively spliced products of LMNA, whereas themajor B-type lamins, lamin B1 and lamin B2, are encoded bytwo distinct genes, LMNB1 and LMNB2, respectively (12).Although the A- and B-type lamins share a similar structure,they differ in their behavior during cell division and their pat-terns of expression. B-type lamins are found in all cell types andare expressed throughout development, whereas A-type laminsare not present in early embryos (13).Within the nucleus, laminB1 binds directly to chromatin and histones (14) and interactswith several chromatin-binding inner nuclear membrane pro-teins (e.g. lamina-associated proteins, lamin B receptor (LBR),and the nuclear pore protein nucleoporin 153) (15). Followingmitosis, B-type lamins assemble first into the nuclear lamina,followed by lamin A, and subsequently by lamin C (16). Fewdata exist on the biological role of the B-type lamins. B-typelaminsmay have a direct role inDNA synthesis (16), and silenc-ing of lamin B by siRNA causes cell death in human cells and inCaenorhabditis elegans (17, 18). For these reasons, B-typelamins are generally assumed to be essential. Mice deficient inlamin B1 (Lmnb1�/�), which were created with a gene-trapembryonic stemcell line, die in the perinatal periodwith defectsin lung and bone (19). Lmnb1�/� embryonic fibroblasts displaynuclear shape abnormalities, chromosomal abnormalities, andimpaired differentiation into adipocytes (19). In humans, dupli-cation of LMNB1 causes autosomal dominant leukodystrophy(20).To further elucidate the function of lamin B1, we decided to
examine nuclear shape and dynamics in fibroblasts fromLmnb1�/� and wild-type embryos by quantitative time-lapsevideomicroscopy.Wemade a stunning observation: nuclei spinin the absence of lamin B1, and we show that this nuclear rota-tion could be rescued by transfection with a GFP-lamin B1
* This work was supported by National Institutes of Health Grants HL073809,HL64858, AR050200, and HL082792, the American Heart Association Grant0635359N, the Progeria Research Foundation, March of Dimes, and a NRSApostdoctoral fellowship HL079862 (to J. Y. J.). The costs of publication ofthis article were defrayed in part by the payment of page charges. Thisarticle must therefore be hereby marked “advertisement” in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
□S The on-line version of this article (available at http://www.jbc.org) containssupplemental videos S1–S15, Figs. S1–S3, and additional text.
1 To whom correspondence should be addressed: Partners Research Facility-Rm. 283, 65 Landsdowne St., Cambridge, MA 02139. Tel.: 617-768-8273;Fax: 617-768-8280; E-mail: [email protected].
2 The abbreviations used are: NPC, nuclear pore complex; MEF, mouseembryo fibroblast; LBR, lamin B receptor; KASH, Klarsicht/ANC-1/Syne-1homology; GFP, green fluorescent protein; EGFP, enhanced GFP; RFP, redfluorescent protein; ER, endoplasmic reticulum; Az, sodium azide; DOG,2-deoxyglucose; Bis-Tris, 2-[bis(2-hydroxyethyl)amino]-2-(hydroxymethyl)-propane-1,3-diol.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 282, NO. 27, pp. 20015–20026, July 6, 2007Printed in the U.S.A.
JULY 6, 2007 • VOLUME 282 • NUMBER 27 JOURNAL OF BIOLOGICAL CHEMISTRY 20015
at Massachusetts G
eneral Hospital T
readwell Library on July 1, 2007
ww
w.jbc.org
Dow
nloaded from
http://www.jbc.org/cgi/content/full/M611094200/DC1Supplemental Material can be found at:

fusion protein. Using fluorescence labeling of discrete nuclearenvelope components, we found that the nuclear rotationincludes rotational movement of chromatin, the nuclear lam-ina, the inner nuclear membrane, NPC proteins, as well as theendoplasmic reticulum (ER) immediately adjacent to thenucleus, but not the extended ER or the surrounding cytoskel-eton. Furthermore, the nuclear rotationwas energy-dependent,required an intact cytoskeleton, and the rotation could bereduced by transfection of Lmnb1�/� cells with larger nesprinisoforms. These data suggest a critical role for lamin B1 as amolecular anchor at the nuclear-cytoplasm interface, specifi-cally in facilitating physical coupling between the outer nuclearmembrane and the surrounding cytoskeleton.
EXPERIMENTAL PROCEDURES
Cell Culture and Reagents—Wild-type and Lmnb1-deficient(Lmnb1�/� and Lmnb1�/�, respectively) mouse embryo fibro-blasts (MEFs) were derived from Lmnb1�/� mouse embryosand wild-type littermates (19). Lmna�/� and Lmna�/� MEFswere obtained fromDr. Colin Stewart (21). All cells weremain-tained in Dulbecco’s modified Eagle’s medium containing 10%fetal bovine serum (HyClone, Logan, UT) and penicillin/strep-tomycin (Invitrogen) at 37 °C. To study the role of the cytoskel-eton in nuclear dynamics, cells were treated with media con-taining either 200 nM taxol, 300 nM nocodazole, or 200 nMcytochalasin D (all from Sigma) before starting time-lapseimaging experiments. For recovery studies, drugs wereremoved and replacedwith freshmedia before the second time-lapse experiment. In all cases, fluorescence immunohistochem-istry confirmed that each drug was disrupting its intended tar-get cytoskeletal component, while leaving other structuresintact. For ATP inhibitor studies, subconfluent Lmnb1�/�
MEFs were treated with media containing 5 mM sodium azide(Az) and 1mM2-deoxyglucose (DOG), both fromSigma, beforestarting the time-lapse imaging experiment. Cell nuclei werelabeled with Hoechst 33342; the endoplasmic reticulum wasstained with ER-Tracker Blue-White DPX; and mitochondriawere stained with MitoTracker Green FM (all from MolecularProbes).Plasmid Construct and Transfection—The pGFP-Lmnb1
construct was generated by inserting the human lamin B1sequence into the multiple cloning site of the pEGFPC1 vector(Clontech), generating lamin B1 fused to the carboxyl terminusof EGFP. The nesprin constructs were generated by cloninghuman cDNAs for nespirn-1�, -2�, and -2� into pEGFPC1 vec-tors. Nesprin-1�KASH (amino acids: nespirn-1�-(918–982))or 2�KASH (amino acids: nespirn-2�-(483–542)) contain onlythe KASH domain sequences for each isoform. All nesprins arefused to the carboxyl terminus of EGFP. The pGFP-emerinplasmid was a gift from Dr. Chris Hutchison of University ofDurham; the pRFP-Lmna and pLBR-GFP plasmids were pro-vided by Dr. HowardWorman of Columbia University; and thepPOM121-GFP plasmidwas provided byDr. Brian Burke of theUniversity of Florida. GFP-actin and GFP-tubulin were fromDr. FrankGertler of theMassachusetts Institute of Technology.All constructs contain full-length human proteins, which,except for LBR and POM121, are placed at the carboxyl termi-nus of fluorescence tags. Transfection was conducted with
GeneJammer Transfection Reagent (Stratagene) at a ratio of 3�l to 1 �g of DNA.Time-lapse Imaging and Fluorescent Confocal Microscopy—
Lmnb1�/�, Lmnb1�/�, and Lmna�/�MEFswere grown to sub-confluent density in 35-mm polystyrene cell culture dishes(Corning), sealed with parafilm, and equilibrated to room tem-perature. Images of cells were acquired automatically at either20� or 60� magnification on an Olympus IX-70 microscopewith a digital charge-coupled device camera (CoolSNAP HQ,Roper Scientific) for 100 frames at 5-min intervals (correspond-ing to 8 h and 20 min). Phase contrast images alone wereacquired with ImagePro image acquisition software (MediaCybernetics). Consecutive phase contrast and fluorescentimages were acquired with an automated, motorized shuttersystem (Prior ProScan II) controlled by IPLab version 3.7(Scanalytics) software. Fluorescence images were processed withDeconvolution 7.0 (Vaytech Image). For confocal microscopy,cells were plated on 35-mmglass-bottomculture dishes (MatTek)and imaged with a Plan-Apochromat 63�/1.4 oil differentialinterference contrast oil-immersion objective on a Zeiss Axiovert100M microscope. Laser scanning microscopy was done with anargon laser module. Photobleaching and image acquisition werecontrolled with the Zeiss LSM510 software.Image Acquisition and Manipulation—Phase contrast and
fluorescence time-lapse images and confocal microscopy werecarried out as described above. Time-lapse images were takenwith an Olympus LCPlanF 20� phase contrast objective(numerical aperture, 0.40). Confocal imageswere acquiredwitha Plan-Apochromat 63� oil immersion objective (numericalaperture, 1.4). All imaging experiments were carried out atroom temperature, and cells were maintained in complete cul-ture media. Videos of time-lapse images were generated withMATLAB, and fluorescence grayscale images from IPLab werecolorized in MATLAB for the appropriate fluorochromes(green for GFP, and red for RFP). Photographic films fromWestern blot studies were digitized on an Epson Perfection2450 scanner with linear intensity settings. Digital images wereprocessed in Adobe Photoshop (version 6.0) by adjusting thelinear image intensity display range.Nuclear Movement Analysis—Customized MATLAB algo-
rithms were used to track the position of three to six distinctnucleoli within each nucleus as described previously (22). Foreach frame, the centroid of selected nucleoli was calculated, andthe linear conformal image transformation was computed thatbestmapped the current centroid positions to the original posi-tions while minimizing the least-square error. The linear con-formal transformations can account for a combination of trans-lation, rotation, and scaling, and also preserve the relativeposition of objects to each other. The deviation from the best fit(i.e. the error between the least-square fit transformation andthe actual nucleoli positions) was used as a measure of nucleardeformation, as it describes the extent of nuclear deformationfrom its initial shape independent of absolute nuclear move-ment or uniform changes in size. Nuclei of each cell type wereanalyzed for the time-averaged rotational movement (indegree), translational movement (in microns), and nuclearshape deformation (in microns) as well as normalized nuclearsize, or the scaling factor. Time-lapse videos were also qualita-
Cell Nuclei Spin in the Absence of Lamin B1
20016 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 27 • JULY 6, 2007
at Massachusetts G
eneral Hospital T
readwell Library on July 1, 2007
ww
w.jbc.org
Dow
nloaded from

tively scored by an observer blinded to genotype, as either rotat-ing or not rotating. A rotating nucleuswas defined as rotating atleast 90° in either direction within the 8-h, 20-min time frame.Western Analysis—Cells were lysed in radioimmune precip-
itation assay buffer with 1 mM dithiothreitol, 0.5 mM phenyl-methylsulfonyl fluoride, and protease inhibitor mixture(Sigma) at 1:1000 dilution. Equal amounts of protein in 12 �l ofsample buffer were electrophoresed on 10% Bis-Tris polyacryl-amide gels (Invitrogen), and then transferred onto polyvinyli-dene difluoride membrane (PerkinElmer Life Sciences). Blotswere probed with either primary rabbit polyclonal antibodyagainst human lamin B1 (sc20682, Santa Cruz Biotechnology,1:500 dilution), goat polyclonal antibody against human laminA/C (sc6215, Santa Cruz Biotechnology, 1:500 dilution), rabbitpolyclonal antibody against human emerin (Abcam, 1:3000dilution), goat polyclonal horseradish peroxidase-conjugateantibody against GFP (Abcam, 1:500 dilution), or rabbit anti-actin antibody (Sigma, 1:5000 dilution), followed by chemilu-minescence detection (PerkinElmer Life Sciences).Immunofluorescence Microscopy—Cells were grown on glass
slides, transfectedwithGFP constructs of nesprin isoforms, andfixed in 4% paraformaldehyde for 10 min, 48 h after the trans-fection. Cells were then permeabilized for 10 min with either0.2% Triton X-100 in phosphate-buffered saline, or 0.003% dig-itonin (Sigma) in water (in experiments involving selective per-meabilization of the plasma membrane only). The cells werethen blocked with 1% bovine serum albumin and labeled withappropriate primary and secondary antibodies. Fluorescenceimages were acquired at 20� magnification using an OlympusIX-70 microscope with a digital charge-coupled device camera(CoolSNAP HQ, Roper Scientific) driven by IPLab version 3.7(Scanalytics) software. Cells were probed with either goat poly-clonal antibody against human lamin A/C (sc6215, Santa CruzBiotechnology) followed by Alexa Fluor 568 (red) rabbit anti-goat IgG (A11079, Invitrogen) or rabbit polyclonal antibodyagainst GFP (ab6556, Abcam) followed by Alexa Fluor 350(blue) goat anti-rabbit IgG (A11046, Invitrogen). All antibodieswere used at 1:200 dilution in 1% bovine serum albumin.Statistical Analysis—All experiments were performed at
least three independent times. Statistical analyses were per-formed with the PRISM 3.0 and INSTAT (GraphPad, SanDiego, CA). The unpaired Student’s t test (allowing for differentvariance) was used to analyze if the means of two data groupswere statistically different. For all experiments, a two-tailed pvalue of �0.05 was considered significant. All data areexpressed as mean � S.E.
RESULTS
Lmnb1�/� Nuclei Spin—To assess the functional importanceof lamin B1, we examined nuclear dynamics in fibroblastsderived from littermate Lmnb1�/� and wild-type mouseembryos with time-lapse videomicroscopy. Representative vid-eos of wild-type, Lmnb1�/�, and lamin A/C-null (Lmna�/�)cells are included in Video 1 (supplemental material). Remark-ably, a high percentage of Lmnb1�/� nuclei displayed rotationalmovement around an axis perpendicular to the image plane (i.e.the nuclei rotated parallel to the plane of the cell substrate).Rotation in an orthogonal axis was never observed. The rota-
tional motion was intermittent, and the degree and speed ofrotation varied; the direction of spinning was random. Occa-sionally, nuclei underwent rapid rotation and turned severaltimes before stopping. Fig. 1A shows a series of 14 images(corresponding to 65 min) capturing one nucleus with rapidcounterclockwise rotation of up to 145°. As demonstrated inVideo 1, the positions of nucleoli relative to each other didnot change in the rotating nuclei, despite the striking rota-tion. Also, despite the spinning, Lmnb1�/� nuclei maintained acircular shape and appeared normally positioned within thecell, suggesting that the entire nucleus rotates as a solid body. Incontrast, Lmna�/� nuclei displayed dynamic nuclear deforma-tion but little rotational movement; wild-type cells displayedstable nuclear shape and only minimal nuclear rotation.To quantify the frequency of nuclear rotation, we counted
the fraction of nuclei that rotated at least 90° during an 8-hobservation period. Themajority (90� 2.7%) of Lmnb1�/� cellsmet this criterion, whereas wild-type and Lmna�/� cells rarelydisplayed any nuclear rotation (p � 0.005) (Fig. 1B). Subse-quently, we analyzed nuclear dynamics in more detail (e.g.nuclear rotation, translation, and deformation) based on thetrajectories of selected nucleoli within each nucleus. We calcu-lated the incremental rotation at each time point and deter-mined the absolute angles of nuclear rotation at the end of eachtime-lapse period for all of the nuclei observed. On average,Lmnb1�/� nuclei rotated 90.0 � 20.8° in either direction, withone nucleus turning as much as 593° (i.e. more than 1.5 fullturns) (Fig. 1C). Despite the intermittent nature of nuclear rota-tion, the time-averaged angle of rotation for Lmnb1�/� nucleiwas larger than in wild-type and Lmna�/� nuclei. BothLmnb1�/� and Lmna�/� cells had slightly increased nucleartranslational motion (Fig. 1D), predominantly caused byincreased cell movement. However, because Lmna�/� andLmnb1�/� cells did not differ statistically in their translationalmovement, this effect appears to be unrelated to nuclear rota-tional movement, which was unique to Lmnb1�/� cells. UnlikeLmna�/� cells, Lmnb1�/� cells did not show increased time-averaged nuclear deformation compared with wild-type cells(Fig. 1E), and nuclear fragility was normal (Fig. S1, supplemen-tal material).Rotation of Lmnb1�/� Nuclei Is Attenuated by the Expression
of a GFP-lamin B1 Fusion Protein—To confirm that nuclearspinning was attributable to the loss of wild-type lamin B1, weanalyzed nuclearmovement in Lmnb1�/� cells transfected witha GFP-lamin B1 fusion construct. Experiments were donewithin 48 h post-transfection, enough time for mitosis andnuclear envelope reformation to occur in most cells. Nucleiexpressing GFP-lamin B1 exhibited reduced nuclear rotationcompared with nontransfected Lmnb1�/� controls (45% reduc-tion, p � 0.02), but expression of a GFP-emerin fusion con-struct did not alter nuclear rotation (Fig. 1F) and failed torestore wild-type behavior. Similarly, transfection with a redfluorescent protein-lamin A fusion protein (RFP-lamin A) didnot prevent nuclear rotation (not shown). Average translationalmovement was not affected by any of the constructs. Westernanalysis confirmed the presence of both GFP-lamin B1 andGFP-emerin 1 day after transfection (Fig. 1G). Expression ofGFP-lamin B1, however, did not completely restore the wild-
Cell Nuclei Spin in the Absence of Lamin B1
JULY 6, 2007 • VOLUME 282 • NUMBER 27 JOURNAL OF BIOLOGICAL CHEMISTRY 20017
at Massachusetts G
eneral Hospital T
readwell Library on July 1, 2007
ww
w.jbc.org
Dow
nloaded from

type phenotype (45% reduction in nuclear rotation), possiblydue to variations in transfection efficiency or expression ofGFP-lamin B1. These data indicate that rotation in Lmnb1�/�
nuclei is due to the loss of functional lamin B1.Nuclear Rotation in Lmnb1�/� Cells Includes Chromatin and
the Nuclear Envelope—Although time-lapse analysis of phase-contrast images is well suited to quantify the extent of nuclearrotation based on nucleoli movement, it is insufficient to deter-mine if the rotational movement encompasses the entirenuclear interior and the nuclear envelope. Therefore, we selec-tively labeled chromatin, the nuclear lamina, the inner nuclearmembrane, and NPCs with fluorescent probes and then ana-lyzed, by time-lapse videomicroscopy, which nuclear structuresparticipated in the nuclear rotation in Lmnb1�/� fibroblasts.Staining cells with Hoechst 33342, a DNA minor groove-bind-
ing fluorescent dye, demonstratedthat the entire chromatin contentswere involved in the nuclear rota-tion (Fig. 2A andVideo 2). This find-ingwas in keepingwith our observa-tion that nucleoli retained theirrelative position to each other.Because lamins can directly bind
to DNA and thus attach the nuclearlamina to chromatin (14), we inves-tigated whether the nuclear laminaalso rotated in the Lmnb1�/� fibro-blasts. Time-lapse microscopy ofcells expressing RFP-lamin Arevealed distinct rotation of thenuclear lamina (Video 3). Theseresults were further confirmed bytime lapse-analysis of partially pho-tobleached nuclei in RFP-laminA-labeled Lmnb1�/� cells. In oneexample, images taken immediatelyafter photobleaching showed adarkened band through the fluores-cent nuclear lamina (Fig. 2F), whichturned �50° clockwise during thesubsequent 78-min period of time-lapse confocal laser scanningmicroscopy (see also Video 7, sup-plemental material).Subsequently, we tracked rota-
tion of the inner nuclear membraneby fluorescent time-lapse videomi-croscopy in cells transfected withGFP-tagged lamin B receptor (LBR-GFP) or emerin (GFP-emerin). LBRand emerin reside at the innernuclear membrane (23) and there-fore can serve as an indicator forinner nuclear membrane move-ment. We found that both LBR,which interacts with lamin B1, andemerin, which binds to lamin A,participated in the nuclear rotation
in Lmnb1�/� cells (Fig. 2C and 3A). Rotation of the innernuclear membrane was clearly demonstrated by the movementof the fluorescent nuclear periphery in the supplementalVideos4 and 8. Importantly, the inner nuclear membrane (marked byGFP-emerin) rotation occurred in unison with the rotation ofthe nuclear interior, as indicated by the numbered nucleoli inthe phase-contrast images (Fig. 3A). We also attempted photo-bleaching studies on GFP-emerin- and LBR-GFP-labelednuclei, but the high diffusional mobility of these inner nuclearmembrane proteins and the resulting rapid fluorescence recov-ery (�10min) did not allow long term observations of the pho-tobleached sections.Lamin B1 can bind to NPC proteins such as nucleoporin 153
(15), so that loss of lamin B1 could potentially allow the innernucleus to rotate relative to the nuclear pores and the outer
FIGURE 1. Time-lapse microscopy of Lmnb1�/� nuclei revealed striking rotational movement of thenuclear content and the lamina. A, series of 14 phase-contrast images of one Lmnb1�/� nucleus was extractedfrom 100 time-lapse images taken every 5 min over an 8-h, 20-min period at 20� magnification with anOlympus IX-70 microscope. The nucleus showed significant rotational movement in a counterclockwise direc-tion (�145°), as indicated by the numbered nucleoli. Representative videos of Lmnb1�/�, wild-type (Lmnb1�/�),and Lmna�/� nuclei are included in Video 1. Quantitative analysis revealed significant rotational movement ofLmnb1�/� nuclei compared with wild-type (Lmnb1�/�) and Lmna�/� cells, and GFP-lamin B1 expression spe-cifically attenuated nuclear rotation in Lmnb1�/� cells. B, Lmnb1�/� cells had a significantly larger fraction (90 �2.7%; n � 121) of nuclei that rotated at least 90° compared with wild-type (7.0 � 2.4%; n � 113) and Lmna�/�
(4.9 � 2.4%; n � 81) cells. C, the cumulative degree of nuclear rotation was also significantly higher forLmnb1�/� cells (n � 33) compared with wild-type (n � 40) and Lmna�/� cells (n � 35). *, p � 0.005 versuswild-type and Lmna�/�. D, average nuclear translational movement of Lmnb1�/� cells was higher than inwild-type cells (8.8 � 0.94 versus 6.5 � 0.53 �m) but not significantly different from Lmna�/� nuclei. *, p � 0.05versus wild-type. E, Lmna�/� cells displayed significant time-averaged nuclear shape deformations, whereasLmnb1�/� and wild-type nuclei only exhibited minor variations in shape over time. *, p � 0.005 versus wild-typeand Lmnb1�/�. F, time-lapse microscopy experiments were conducted in Lmnb1�/� cells within 48 h aftertransfection with GFP-Lmnb1 or GFP-emerin. Lmnb1�/� nuclei expressing GFP-lamin B1 rotated significantlyless than nuclei of nontransfected Lmnb1�/� control cells. The average angle of rotation at the end of thetime-lapse period for transfected nuclei was 55.4 � 9.3% of controls (n � 32), averaging 34.7° versus 62.7° fornontransfected nuclei (p � 0.02). In contrast, expression of GFP-emerin in Lmnb1�/� cells did not reducenuclear rotation; the average angle of rotation was 94.3 � 21.6% of control (n � 15). Error bars indicate mean �S.E. *, p � 0.05 versus nontransfected controls. G, expression of GFP-emerin (61 kDa) or GFP-lamin B1 (94 kDa)was confirmed by Western analysis of whole cell lysates taken 1 day post-transfection probed with antibodiesagainst GFP. Control bands were probed with antibodies against emerin or actin.
Cell Nuclei Spin in the Absence of Lamin B1
20018 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 27 • JULY 6, 2007
at Massachusetts G
eneral Hospital T
readwell Library on July 1, 2007
ww
w.jbc.org
Dow
nloaded from
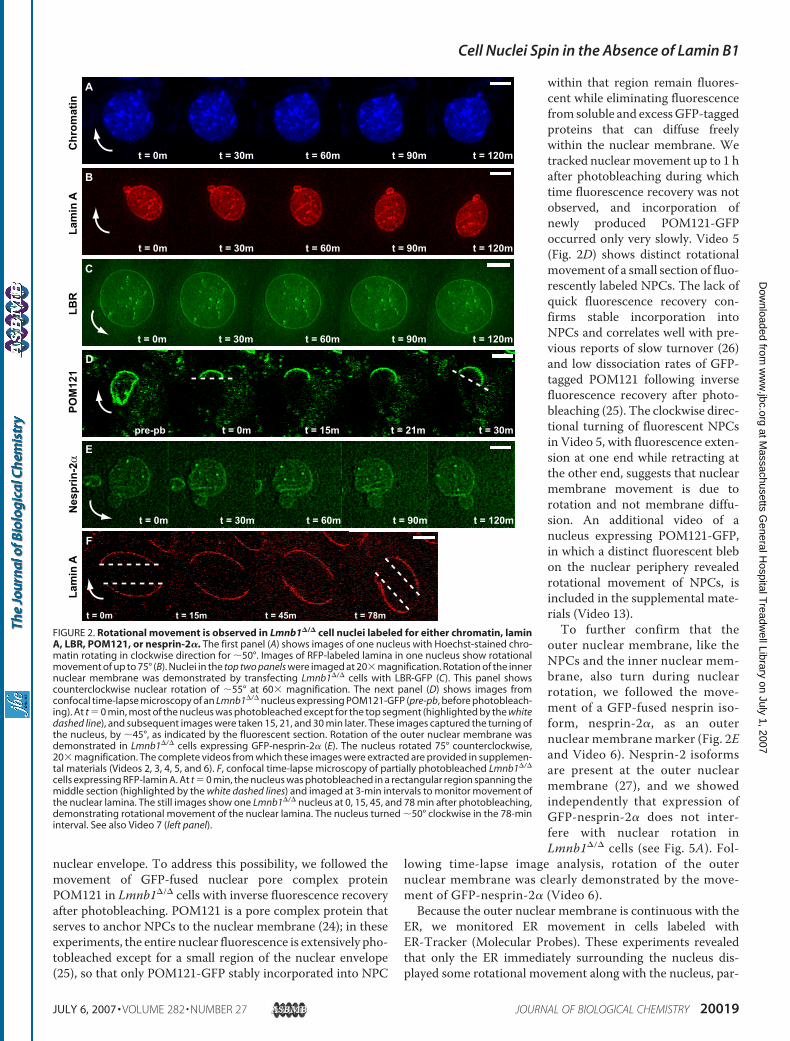
nuclear envelope. To address this possibility, we followed themovement of GFP-fused nuclear pore complex proteinPOM121 in Lmnb1�/� cells with inverse fluorescence recoveryafter photobleaching. POM121 is a pore complex protein thatserves to anchor NPCs to the nuclear membrane (24); in theseexperiments, the entire nuclear fluorescence is extensively pho-tobleached except for a small region of the nuclear envelope(25), so that only POM121-GFP stably incorporated into NPC
within that region remain fluores-cent while eliminating fluorescencefrom soluble and excessGFP-taggedproteins that can diffuse freelywithin the nuclear membrane. Wetracked nuclearmovement up to 1 hafter photobleaching during whichtime fluorescence recovery was notobserved, and incorporation ofnewly produced POM121-GFPoccurred only very slowly. Video 5(Fig. 2D) shows distinct rotationalmovement of a small section of fluo-rescently labeled NPCs. The lack ofquick fluorescence recovery con-firms stable incorporation intoNPCs and correlates well with pre-vious reports of slow turnover (26)and low dissociation rates of GFP-tagged POM121 following inversefluorescence recovery after photo-bleaching (25). The clockwise direc-tional turning of fluorescent NPCsin Video 5, with fluorescence exten-sion at one end while retracting atthe other end, suggests that nuclearmembrane movement is due torotation and not membrane diffu-sion. An additional video of anucleus expressing POM121-GFP,in which a distinct fluorescent blebon the nuclear periphery revealedrotational movement of NPCs, isincluded in the supplemental mate-rials (Video 13).To further confirm that the
outer nuclear membrane, like theNPCs and the inner nuclear mem-brane, also turn during nuclearrotation, we followed the move-ment of a GFP-fused nesprin iso-form, nesprin-2�, as an outernuclear membranemarker (Fig. 2Eand Video 6). Nesprin-2 isoformsare present at the outer nuclearmembrane (27), and we showedindependently that expression ofGFP-nesprin-2� does not inter-fere with nuclear rotation inLmnb1�/� cells (see Fig. 5A). Fol-
lowing time-lapse image analysis, rotation of the outernuclear membrane was clearly demonstrated by the move-ment of GFP-nesprin-2� (Video 6).
Because the outer nuclear membrane is continuous with theER, we monitored ER movement in cells labeled withER-Tracker (Molecular Probes). These experiments revealedthat only the ER immediately surrounding the nucleus dis-played some rotational movement along with the nucleus, par-
FIGURE 2. Rotational movement is observed in Lmnb1�/� cell nuclei labeled for either chromatin, laminA, LBR, POM121, or nesprin-2�. The first panel (A) shows images of one nucleus with Hoechst-stained chro-matin rotating in clockwise direction for �50°. Images of RFP-labeled lamina in one nucleus show rotationalmovement of up to 75° (B). Nuclei in the top two panels were imaged at 20� magnification. Rotation of the innernuclear membrane was demonstrated by transfecting Lmnb1�/� cells with LBR-GFP (C). This panel showscounterclockwise nuclear rotation of �55° at 60� magnification. The next panel (D) shows images fromconfocal time-lapse microscopy of an Lmnb1�/� nucleus expressing POM121-GFP (pre-pb, before photobleach-ing). At t � 0 min, most of the nucleus was photobleached except for the top segment (highlighted by the whitedashed line), and subsequent images were taken 15, 21, and 30 min later. These images captured the turning ofthe nucleus, by �45°, as indicated by the fluorescent section. Rotation of the outer nuclear membrane wasdemonstrated in Lmnb1�/� cells expressing GFP-nesprin-2� (E). The nucleus rotated 75° counterclockwise,20� magnification. The complete videos from which these images were extracted are provided in supplemen-tal materials (Videos 2, 3, 4, 5, and 6). F, confocal time-lapse microscopy of partially photobleached Lmnb1�/�
cells expressing RFP-lamin A. At t � 0 min, the nucleus was photobleached in a rectangular region spanning themiddle section (highlighted by the white dashed lines) and imaged at 3-min intervals to monitor movement ofthe nuclear lamina. The still images show one Lmnb1�/� nucleus at 0, 15, 45, and 78 min after photobleaching,demonstrating rotational movement of the nuclear lamina. The nucleus turned �50° clockwise in the 78-mininterval. See also Video 7 (left panel).
Cell Nuclei Spin in the Absence of Lamin B1
JULY 6, 2007 • VOLUME 282 • NUMBER 27 JOURNAL OF BIOLOGICAL CHEMISTRY 20019
at Massachusetts G
eneral Hospital T
readwell Library on July 1, 2007
ww
w.jbc.org
Dow
nloaded from

ticularly at the start of rotation, but that the majority of the ERremained stationary throughout the rotation, suggesting thatthe lipid membranes can flow sufficiently on the slow experi-mental time scales (Fig. 3B and Video 9).The Nucleus Rotates Relative to the Stationary Cytoskeleton—
To address the possibility that nuclear rotation occurred as partof overall cellular and cytoskeletal movement, we performedtime-lapse videomicroscopy in Lmnb1�/� cells expressingeither GFP-actin or GFP-tubulin. These experiments showedthat the actin cytoskeleton remained stationary, whereas thenucleus rotated within a filament network cage (Fig. 4A andVideo 10). The majority of the cytoplasmic microtubule net-work, away from the nucleus, also appeared tomove independ-ently of the rotating nucleus (Fig. 4B and Video 11). Microtu-bules immediately surrounding the nucleus, however, appearedmore closely associated with the nucleus and its movement,with some rotating with the nucleus. It is at this perinuclearspace that interactions of microtubules with the nuclear mem-brane are potentially driving rotational movement of thenucleus, but the resolution of videomicroscopy is insufficient toobserve the dynamics of individual microtubules at the nuclearperiphery. Additional videos of rotating nuclei with eitherGFP-actin or GFP-tubulin are included in supplemental materials(Videos 14 and 15). Furthermore, time-lapse video analysis ofcells with fluorescent mitochondrial stain also revealed thatthe mitochondria, which anchor to the cytoskeleton, do notrotate with the nucleus (Fig. 4C and Video 12). Consistent
with the normal cytoskeletaldynamics seen in the videomicros-copy experiments, we found thatcytoskeletal structure in fixedLmnb1�/� was indistinguishablefrom wild-type cells when viewedby immunofluorescence labelingagainst F-actin, microtubules, andvimentin (Fig. S2).Taken together, these data dem-
onstrate that nuclear rotation inLmnb1�/� cells involves movementof the entire nucleus (i.e. chromatin,the nuclear lamina, the nuclearmembranes, and the nuclear pores)relative to most of the ER and otherstable cytoplasm contents. Thus, itappears that lamin B1 is notrequired for attaching the intranu-clear contents to the nuclear laminaor the nuclear pores but instead isnecessary to anchor the nucleus tothe surrounding cytoskeleton.Lmnb1�/� Nuclear Rotation Can
Be Abolished by Overexpression ofLarge Nesprin Isoforms—In wild-type cells, large nesprin isoformslocated on the outer nuclear mem-brane can bind to cytoskeletal struc-tures with their amino-terminalcytoplasmic domain. At the same
time, these nesprins can interact with SUN proteins on theinner nuclear membrane via their carboxyl-terminal perinu-clear domain. SUN proteins in turn bind to lamins, chromatin,and other, yet unknown nuclear envelope proteins, thus com-pleting the link from the cytoskeleton to the nucleus (27–30).We hypothesized that loss of lamin B1 could interfere with thisnuclear-cytoskeletal linker and thus allow nuclear rotation rel-ative to the cytoskeleton. To test if overexpression of nesprincould overcome this deficit and reduce nuclear rotation, weanalyzed Lmnb1�/� cells expressing GFP constructs of variousnesprin isoforms (28) by time-lapse videomicroscopy. In par-ticular, we examined the effect of (i) nesprin-l�, a 112-kDa iso-form with five spectrin repeat domains; (ii) nesprin-2�, a61-kDa isoform with two spectrin repeat domains; and (iii)nesprin-2�, a 87-kDa isoform with four spectrin repeatdomains (Fig. 5A, left panel). In addition, we tested two trun-cated mutant forms (nesprin-1�KASH and nesprin-2�KASH)consisting of only the 60-residue carboxyl-terminal KASH(Klarsicht/ANC-1/Syne-1 homology) domain that includes thetransmembrane domain and the luminal domain essential fornuclear localization of nesprin-1 and nesprin-2. Our data showthat expression of the GFP-tagged versions of the largest iso-forms, GFP-nesprin-1� or GFP-nesprin-2�, almost completelyabolished nuclear rotation in Lmnb1�/� cells (Fig. 5A, rightpanel). In contrast, expression of the shorter isoform GFP-ne-sprin-2� or the truncated KASH domain mutants (GFP-ne-sprin-1�KASH or GFP-nesprin-2�KASH) did not significantly
FIGURE 3. Nuclear rotation includes the nuclear interior and emerin proteins of the nuclear membrane,but not the cytoplasmic endoplasmic reticulum or mitochondria. Phase-contrast (top) and epifluorescence(bottom) time-lapse images of Lmnb1�/� nuclei expressing GFP-emerin, or labeled for either mitochondria orER. A, this series of 5 images, taken at 20� magnification and spanning a 3-h period, demonstrates distinctclockwise nuclear rotation (�150°) of the inner nuclear membrane (emerin) and the nuclear interior (phasecontrast), as indicated by the numbered nucleoli. See also Video 8. Except for the layer directly associated withthe nuclear envelope, the majority of the cytoplasmic ER remained stationary during nuclear rotation (B). (Thisis more apparent in the accompanying Video 9.) In B, the nucleus rotated a complete turn of 360°. Fluorescenceimages were deconvoluted for clarity. The arrows indicated the direction of rotation; scale bars � 10 �m.
Cell Nuclei Spin in the Absence of Lamin B1
20020 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 27 • JULY 6, 2007
at Massachusetts G
eneral Hospital T
readwell Library on July 1, 2007
ww
w.jbc.org
Dow
nloaded from

reduce nuclear rotational movement (Fig. 5A, right panel). Seealso Fig. 2D (GFP-nesprin-2� and Video 5). The reason for theselective effect of the larger isoforms is unclear, but we specu-late that the larger cytoplasmic, amino-terminal domains in thenesprin-1� and nesprin-2� isoforms could interact with thecytoskeleton in a size-dependent manner and thus restoreanchoring of the nuclear envelope to the cytoskeleton. To con-firm that the amino-terminal nesprin domains face the cyto-plasm, we treated fixed cells with low concentrations of digito-nin, which selectively permeabilizes the plasma membranewhile leaving the nuclear membranes and ER intact (29). Posi-
tive immunofluorescent staining ofGFP showed that the amino-termi-nal domains of the GFP-nesprinconstructs were accessible to thecytoplasm, suggesting that thesenesprin constructs are located onthe outer nuclear membrane (Fig.5B). Similar results were observedwith all nesprin constructs tested(data not shown). However, it isimportant to note that theseresults do not exclude the possibil-ity of nesprin isoforms inside thenucleus and do not necessarilyreflect the localization of endoge-nous nesprins.Cytoskeleton Disruption Attenu-
ates Lmnb1�/� Nuclear Rotation—We hypothesized that nuclear rota-tion is driven by the surroundingcytoskeleton. Consistent with thishypothesis, treatment of Lmnb1�/�
cells with either taxol (a microtu-bule stabilizer), nocodazole (a tubu-lin polymerization inhibitor), orcytochalasin D (an F-actin filamentdisruptor) significantly decreasedthe fraction of rotating nuclei (Fig.6A) and reduced the time-averagedangle of rotation in Lmnb1-defi-cient nuclei, by 54%, 66%, and 38%,respectively (Fig. 6B). In thoseexperiments, the reagents targetingmicrotubules (i.e. taxol and nocoda-zole) had a significantly strongereffect on reducing nuclear rotationin Lmnb1�/� cells than the actindepolymerization drug cytochalasinD. These data suggest that nuclearrotation is largely driven by micro-tubule dynamics, consistent withprevious observations of microtu-bule-dependent nuclear movementin response to fluid shear stress (31).In our cells, stabilizingmicrotubulesdid not affect nuclear translationalmotion, whereas depolymerization
of tubulin and F-actin restored nuclear translocation inLmnb1�/� cells to levels observed in wild-type cells (Fig. 6C).In all cases, drug target specificity and efficiency were con-firmed by immunofluorescence imaging of F-actin andmicrotubules (data not shown). We confirmed that theattenuating effect of cytoskeleton disruptors on nuclearmovement was not due to cell death or other permanentfunctional effects by continued observation of cells afterreplacing drugs with fresh media. Whereas cytoskeleton dis-ruption significantly decreased nuclear rotation in responseto drug treatment, removing the cytoskeleton-disrupting
FIGURE 4. Cytoplasmic actin and microtubule networks are not disrupted during nuclear rotation. Epif-luorescence (top) and phase-contrast (bottom) time-lapse images of one Lmnb1�/� nucleus expressing GFP-actin showed no rotational movements of the acting filaments during nuclear rotation (A). The series of fiveimages, taken at 20� magnification and spanning a 3-h period, demonstrates distinct clockwise nuclear rota-tion (�105°) in phase contrast, without disrupting cytoplasmic actin filaments (fluorescent). Time-lapse imagesof a Lmnb1�/� nucleus expressing GFP-tubulin (also at 20�) showed significant rotation of the nucleus (over180°) in phase contrast, while most of the fluorescent microtubule structures in the cytoplasm were unaffectedand remained stationary (B). See also Videos 10 and 11. Fluorescently labeled mitochondria in the cytoplasmremained stationary as one nucleus rotated for �100°, as indicated by the numbered nucleoli in phase-contrast(C and Video 12).
Cell Nuclei Spin in the Absence of Lamin B1
JULY 6, 2007 • VOLUME 282 • NUMBER 27 JOURNAL OF BIOLOGICAL CHEMISTRY 20021
at Massachusetts G
eneral Hospital T
readwell Library on July 1, 2007
ww
w.jbc.org
Dow
nloaded from

drugs restored nuclear rotational and translational move-ments in post-treatment cells to levels comparable to pre-treatment Lmnb1�/� cells (Fig. 6, D and E).Nuclear Rotation Is Energy-dependent—To determine if
nuclear rotation was energy-dependent, we depleted ATP lev-els in Lmnb1�/� cells with sodium azide and 2-deoxyglucose(Az/DOG). ATP depletion significantly reduced nuclear rota-tion in Lmnb1�/� cells, resulting in a 63% reduction after treat-ment with Az/DOG compared with untreated cells (Fig. 7A). Incontrast, translational movement was not altered by Az/DOGtreatment (Fig. 7B). Thus, nuclear rotation is ATP-dependent,whereas translational movement is not.
DISCUSSION
We investigated the nuclear dynamics of Lmnb1�/� MEFs tobetter understand the role of lamin B1 in nuclear function andarchitecture. Using quantitative analysis of time-lapse videomi-croscopy sequences, we monitored nuclear rotation, transla-tion, and deformation in wild-type, Lmna�/�, and Lmnb1�/�
cells. We found that Lmnb1�/� fibroblasts displayed distinctand prominent nuclear rotation that could be abolished byexpression of wild-type lamin B1. The rotational movementencompassed the nuclear interior (chromatin), nuclear lamina,inner and outer nuclear membranes, NPCs, and regions of theER adjacent to the nucleus. In contrast to Lmna�/� cells, laminB1 deficiency did not alter nuclear shape or rigidity, and mostLmnb1�/� nuclei retained their spherical shape. It appears thatin mammalian cells, A-type and B-type lamin proteins havedifferent roles in maintaining nuclear structure and stability,and that lamin B1 has a previously unrecognized function ofanchoring the nucleus to the cytoskeleton. These observationsare also supported by previous experiments, which showed thatlamins A and C, but not lamin B1, are the main contributors tonuclear stiffness (32). In contrast, C. elegans express only a sin-gle lamin isoform (a B-type lamin), and silencing of this laminby siRNA results in rapid changes in nuclear shape (17).The coordinated movement of chromatin and nuclear enve-
lope suggests that the entire nucleus is rotating relative to thecytoplasm and the surrounding cytoskeleton. Thus, nuclearrotation occurs presumably via disruption of coupling betweenthe nuclear envelope and the cytoskeleton. The molecularmechanism by which the nucleus is connected to the cytoskel-eton has puzzled researchers for years, as it was unclear howforces required for nuclear positioning and anchoring could betransmitted across the 50 nm wide perinuclear space. Recentwork in C. elegans, Drosophila melanogaster, and mammaliancells has led to the discovery of two new families of nuclearenvelope proteins (nesprins and SUN proteins) that are ideallysuited to transmit forces from the cytoskeleton across thenuclear envelope to the nuclear interior (27, 29, 30, 33–39).These findings have led to the current model of nuclear-cy-toskeletal coupling, in which larger nesprin isoforms located onthe outer nuclear membrane can bind to cytoskeletal F-actin,intermediate filaments, and microtubules. At the same time,nesprins physically interact across the perinuclear space withSUNproteins (SUN1 and SUN2), which are located at the innernuclear membrane. There, SUN proteins can bind to lamins,chromatin, and other, as yet unknown nuclear envelope pro-teins, thus creating a physical link between the cytoskeleton andthe nucleus (36, 40).Based on these observations, we speculate that loss of lamin
B1 can disrupt nuclear-cytoskeletal coupling by disturbinglocalization of SUN proteins or other inner nuclear membraneproteins that are required for localization of nesprin-1 andnesprin-2 to the nuclear envelope (29, 30, 36). Libotte et al. (34)recently demonstrated that expression of a dominant-negativelamin B1 mutant can lead to redistribution of lamin A/C fromthe nuclear lamina and results in mislocalization of nesprin-2away from the nuclear envelope. Although localization of laminA/C and transfected nesprin isoforms appeared normal in our
FIGURE 5. Lmnb1�/� nuclear rotation is significantly reduced by theectopic expression of nesprin-1� and nesprin-2� but not by smallernesprin isoforms. The left panel in A shows the predicted protein structuresbased on SMART program, including the number of spectrin repeats fornesprin-1�, -2�, and -2�, and their KASH domains. The right panel in A showscorresponding results for time-lapse nuclear rotational studies of wild-type,Lmnb1�/� control cells, and Lmnb1�/� cells expressing nesprin constructs.Expression of a GFP-nesprin-1� construct decreased average angle of nuclearrotation from 29.54 � 4.5° (n � 32) for control cells to 11.76 � 2.6° (n � 17, p �0.001). Expression of nesprin-2�-GFP construct also decreased the averageangle of rotation to 7.6 � 1.6° (n � 15), from 24.22 � 7.4° (n � 19) for controlcells, p � 0.04. Average angle of rotation for wild-type nuclei was 7.77 � 2.1°(n � 8). GFP constructs of nesprin-l�KASH, nesprin-2�, or nesprin-2�KASHdid not significantly reduce nuclear rotation of Lmnb1�/� cells. Error bars indi-cate mean � S.E.; *, p � 0.05 for comparison between control and transfectednuclei. B, immunofluorescence staining of cells expressing GFP-nesprin-1�and GFP-2�KASH (green) confirmed localization of these isoforms at the outernuclear membrane. Fixed cells were treated with digitonin to selectively per-meabilize the plasma membrane but not the nuclear membranes. Cells weresubsequently probed with antibodies against GFP (blue) and lamin A/C (red).Only the cytoplasmic GFP domains were accessible to antibodies; the lackof lamin A/C staining confirmed that the nuclear membranes remainedintact. Similar results were observed with all nesprin isoforms tested. Scalebar � 10 �m.
Cell Nuclei Spin in the Absence of Lamin B1
20022 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 27 • JULY 6, 2007
at Massachusetts G
eneral Hospital T
readwell Library on July 1, 2007
ww
w.jbc.org
Dow
nloaded from

Lmnb1�/� cells, we speculate that loss of functional lamin B1could still affect the function of nesprins, resulting in a lack ofnuclear anchoring and nuclear rotation. In C. elegans, weakforces of interaction between nesprin and SUN proteins can be
FIGURE 6. Nuclear rotation in Lmnb1�/� cells requires an intact cytoskel-eton, and the attenuating effect of cytoskeleton disruptors on nuclearrotation is reversible. A, Lmnb1�/� cells treated with the cytoskeletal dis-rupting agents, 200 nM taxol (n � 58), 300 nM nocodazole (n � 79), or 200 nM
cytochalasin D (n � 61), had significantly reduced fractions of rotating nucleicompared with untreated Lmnb1�/� cells (n � 95). For wild-type cells(Lmnb1�/�), n � 119. B, the time-averaged angles of rotation for Lmnb1�/�
cells treated with taxol (n � 36), nocodazole (n � 20), or cytochalasin D (n �20) were reduced by 54%, 66%, and 38%, respectively, compared withuntreated Lmnb1�/� cells (n � 35), although all were still significantly higherthan in wild-type cells (n � 22). C, except for taxol, cytoskeleton-disruptingdrug also reduced nuclear translation in Lmnb1�/� cells, restoring levels closeto those in wild-type cells. *, p � 0.05; **, p � 0.005, both versus untreatedLmnb1�/� controls. #, p � 0.005 versus wild-type cells. Error bars indicatemean � S.E. However, Lmnb1�/� cells treated with either cytoskeleton disrup-tor remained alive and healthy, and when drugs were removed from themedia, time-lapse image analysis revealed that nuclear rotation is re-estab-lished in the same cells (after taxol, n � 24; after nocodazole, n � 25; and aftercytochalasin D. n � 25). #, p � 0.005 versus untreated Lmnb1�/� cells, and*, p � 0.05, versus wild-type control (D). Again, either nocodazole or cytocha-lasin D also reduced nuclear translational movement in Lmnb1�/� cells, and,upon removing drugs from cells, increased overall nuclear movement wasobserved (E).
FIGURE 7. Nuclear rotation in Lmnb1�/� cells is energy-dependent. A, thefinal angle of nuclear rotation for Lmnb1�/� cells treated with Az/DOG (n � 81)was significantly lower than in untreated cells (n � 32): 26.58 � 2.7° versus60.58 � 17.02°; although still higher than in wild-type cells (16.78 � 4.73°; n �16; p � 0.08). #, p � 0.005; *, p � 0.05, all in comparison to Lmnb1�/� cells.Translational movement for Lmnb1�/� nuclei was not significantly altered byATP inhibition (B). However, Az/DOG-treated and control Lmnb1�/� cellsshowed significantly higher nuclear translation compared with wild-type (#,p � 0.005). Error bars indicate mean � S.E.
Cell Nuclei Spin in the Absence of Lamin B1
JULY 6, 2007 • VOLUME 282 • NUMBER 27 JOURNAL OF BIOLOGICAL CHEMISTRY 20023
at Massachusetts G
eneral Hospital T
readwell Library on July 1, 2007
ww
w.jbc.org
Dow
nloaded from

sufficient for correct localization of the proteins to the nuclearenvelope, but higher forces are required during nuclear migra-tion and anchoring of the nucleus to the cytoskeleton (41).Thus,mutations that onlyweaken the interactionwith nesprin/SUN proteins might not be detected in the localization studiesbut could nonetheless result in impaired nuclear-cytoskeletalcoupling that allows for nuclear rotation. Overexpression oflarger nesprin isoforms on the outer nuclear membrane couldthen compensate for a partial loss of function. Consistent withthis model, we found that expression of GFP constructs of thelarger nesprin isoforms (nesprin-1� or -2�) significantlyreduced nuclear rotation in Lmnb1�/� cells. Expression ofsmaller isoforms (nesprin-2�), or of the KASH domain alone,which can displace endogenous nesprins from the outernuclear envelope (34, 42), had no effect on nuclear rotation.Although the exact nuclear envelope localization of many
nesprin isoforms is not clear, larger-sized nesprins are generallythought to be located in the outer nuclear membrane. Smallerisoforms (��60 kDa) can diffuse to the inner nuclear mem-brane where they are in direct contact with lamins and thenucleoplasm, although recent studies of nuclear pore transportsuggest that inner nuclear membrane proteins that containnuclear localization sequences can be actively transportedacross nuclear pores (43). Here, we confirmed that all nesprinconstructs used in our experiments could localize to the outernuclearmembrane and the ER (Fig. 5B), although our results donot exclude the possibility that these isoforms can also be foundon the inner nuclear membrane, and some of the ER localiza-tion could result from overexpression. Interestingly, bothnesprin-1� and -2� contain the entire lamin- and emerin-bind-ing regions (34, 44), which might further stabilize nuclear-cy-toplasmic interactions at the inner nuclear membrane. On theother hand, nesprin-2� has just one emerin-binding region,KASH domains have no such binding regions; these proteinshad no effect on nuclear rotation. It is also possible that thecytoskeletal interaction of excess exogenous nesprins in the ERmay compete against and thus disrupt normal nesprin-depend-ent interactions between outer nuclear membrane and the sur-rounding cytoskeleton. Alternatively, lamin B1 could interactwith yet unknown nuclear envelope proteins important fornuclear-cytoskeleton linkage (independent of nesprins), andoverexpression of nesprins attenuates nuclear rotation nonspe-cifically by providing increased entanglement with thecytoskeleton.At this point, the driving force for the nuclear rotation
remains unclear. Actin filaments,microtubules, and intermedi-ate filaments not only act as a rigid scaffold towhich the nucleuscan be anchored, but their dynamic remodeling at the nuclearinterface could also provide the forces driving the intermittentnuclear rotational movement in Lmnb1�/� cells (35, 45, 46).Lmnb1�/� cells have intact networks of actin, microtubule, aswell as intermediate filaments in the cytoskeleton, and cyto-plasm actin and microtubule organizations are not disruptedduring nuclear rotation, a finding that agrees well with the oth-erwise normal cellular structure andphysiology of laminB1-de-ficient cells. However, in cytoskeleton disruption studies, wefound that the nuclear rotation was more dependent onmicro-tubules than on the actin network (Fig. 6, A and B). This is
consistent with previous findings that flow-induced nuclearrotation in Swiss 3T3 fibroblasts is eliminated by microtubuledisruption, but not F-actin depolymerization, and is accompa-nied by repositioning of the microtubule organization center(31). Similarly, nuclearmigration and positioning in cells platedon micropatterned substrates also requires an intact microtu-bule network (47). Specific nesprin-1 isoforms can interactwiththe kinesin II subunit kinesin family member 3B (KIF3B), sug-gesting a potential link between the nuclear envelope and themicrotubule network (46). The observation that nuclear rota-tion is reduced by energy depletion also implicatesmicrotubulemotors such as dynein, which has been shown to aid nuclearenvelope breakdown during mitosis (48), in facilitating nuclearrotational movement in the absence of lamin B1. One has toconsider that actin is present in both the cytoplasm andnucleus, so that cytochalasin D treatment might also affectnuclear actin and thus affect nuclear dynamics. However, wedid not observe increased nuclear deformations in the cytocha-lasin D-treated cells, and it is not clear if nuclear actin formspolymers or short filaments. It is important to note that therecently discovered nesprin-3 isoforms might anchor thenuclear lamina to the intermediate filament cage surroundingthe nucleus, and disruption of this function could also result innuclear rotation (39). Furthermore, there is evidence for dock-ing of lamin B containing mitotic vesicles with vimentin fila-ments (49). Although we did not investigate disruption of theintermediate filament network in the present study, nuclearrotation has been reported in neuronal interphase nuclei fol-lowing acrylamide treatment, which results in neurofilamentdisruption and a decrease in nuclear lamina thickness (50).Spinning nuclei have been observed previously in a variety of
cells and tissues (51, 52), but here we present the first study toidentify molecular details underlying nuclear rotation.Although nuclear rotation has previously been reported inL-929 fibroblasts (57), our image analysis of L-929 cells showedqualitatively and quantitatively different behaviors from thenuclear rotation observed in Lmnb1�/� fibroblasts, and theoverall rotation of L-929 nuclei was not different from wild-type cells (Fig. S3A). Western analysis revealed normal expres-sion of laminB1 and laminsA/C in these cells (Fig. S3B), furthersuggesting that extensive nuclear rotation is specific to laminB1 deficiency. Most recently, nuclear rotation has beenreported in the early stages of brain development in zebrafishembryos (53), and factors that alter gene expression can alsoaffect nuclear rotation (54). Because lower levels of lamin Bexpression are found in brain and spleen of rat tissues (55) andlevels of specific lamin subtypes can vary dramatically betweendifferent tissues and during development (13), localized andtransient low expression of lamin B1may contribute to nuclearrotation in the developing zebrafish brains. Given that nesprinsare highly expressed inmuscle cells, share similarities with dys-trophin associated with Duchenne and Becker muscular dys-trophy, and could play a role in muscle differentiation (28), wespeculate that, through nesprins, lamin B1 may play a tissue-specific role in balancing cytoskeletal and intranuclear forces atthe nuclear-cytoplasm interface, particularly in muscles cellsunder high force. Defects in nuclear-cytoskeletal coupling canaffect nuclear positioning in the neuromuscular junctions (42)
Cell Nuclei Spin in the Absence of Lamin B1
20024 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 27 • JULY 6, 2007
at Massachusetts G
eneral Hospital T
readwell Library on July 1, 2007
ww
w.jbc.org
Dow
nloaded from

and could also play an important role in muscular dystrophiescaused by lamin mutations (29, 35).In summary, we show that lamin B1 is important for nuclear-
cytoskeletal anchoring and that loss of lamin B1 results in thestriking nuclear rotation in Lmnb1�/� cells that can be reducedby overexpression of larger nesprin isoforms. We propose amodel in which cytoskeletal forces at the interface of nucleus/cytoskeleton can, in the absence of lamin B1, result in intermit-tent rotational movement of the entire nucleus. Consistentwith our phase-contrast and confocal microscopy time-lapsedata, the nuclear envelope along with the nuclear contentscould then rotate relative to the cytoplasm and the surroundingcytoskeleton. The ER, which is continuous with the outernuclear membrane but not connected through a stable proteinnetwork, displays only minimal movement at the immediatenuclear periphery, because lipidmembranes can remodel easilywithin the time scales of our experiments. Thus, lamin B1serves as a molecular anchor at the nuclear envelope to preventnuclear rotation. We suspect that the nuclear rotation associ-ated with the loss of lamin B1 could affect cellular structure andtranscriptional regulation (56). Further investigation into thecellular roles of lamins may help us to better understand howdisorders of the nuclear envelope lead to remarkably diversehuman diseases.
Acknowledgments—Confocal imaging was performed at the W. M.Keck Microscopy Facility at the Whitehead Institute. The pGFP-emerin plasmid was a gift from Dr. Chris Hutchison of University ofDurham, and the pRFP-Lmna plasmid was provided by Dr. HowardWorman of Columbia University. The pPOM121-GFP construct wasgenerated by Dr. Einar Hallberg of University College in Sweden. Wealso thank Dr. Frank Gertler for providing us with the GFP-actin andGFP-tubulin plasmids.
REFERENCES1. Bonne, G., Di Barletta, M. R., Varnous, S., Becane, H. M., Hammouda,
E. H., Merlini, L., Muntoni, F., Greenberg, C. R., Gary, F., Urtizberea, J. A.,Duboc, D., Fardeau, M., Toniolo, D., and Schwartz, K. (1999) Nat. Genet.21, 285–288
2. Bione, S., Maestrini, E., Rivella, S., Mancini, M., Regis, S., Romeo, G., andToniolo, D. (1994) Nat. Genet. 8, 323–327
3. Fatkin, D., MacRae, C., Sasaki, T., Wolff, M. R., Porcu, M., Frenneaux, M.,Atherton, J., Vidaillet, H. J., Jr., Spudich, S., DeGirolami, U., Seidman, J. G.,Seidman, C., Muntoni, F., Muehle, G., Johnson, W., and McDonough, B.(1999) N. Engl. J. Med. 341, 1715–1724
4. Cao, H., and Hegele, R. A. (2000) Hum. Mol. Genet. 9, 109–1125. Muchir, A., Bonne, G., van der Kooi, A. J., van Meegen, M., Baas, F.,
Bolhuis, P. A., de Visser, M., and Schwartz, K. (2000)Hum. Mol. Genet. 9,1453–1459
6. De Sandre-Giovannoli, A., Chaouch,M., Kozlov, S., Vallat, J.M., Tazir,M.,Kassouri, N., Szepetowski, P., Hammadouche, T., Vandenberghe, A.,Stewart, C. L., Grid, D., and Levy, N. (2002) Am. J. Hum. Genet. 70,726–736
7. Eriksson, M., Brown, W. T., Gordon, L. B., Glynn, M.W., Singer, J., Scott,L., Erdos, M. R., Robbins, C. M., Moses, T. Y., Berglund, P., Dutra, A., Pak,E., Durkin, S., Csoka, A. B., Boehnke, M., Glover, T. W., and Collins, F. S.(2003) Nature 423, 293–298
8. Aebi, U., Cohn, J., Buhle, L., and Gerace, L. (1986) Nature 323, 560–5649. Burke, B., and Stewart, C. L. (2002) Nat. Rev. Mol. Cell. Biol. 3, 575–58510. Goldman, R. D., Gruenbaum, Y., Moir, R. D., Shumaker, D. K., and Spann,
T. P. (2002) Genes Dev. 16, 533–547
11. Hutchison, C. J. (2002) Nat. Rev. Mol. Cell. Biol. 3, 848–85812. Stuurman, N., Heins, S., and Aebi, U. (1998) J. Struct. Biol. 122, 42–6613. Broers, J. L., Machiels, B. M., Kuijpers, H. J., Smedts, F., van den Kieboom,
R., Raymond, Y., and Ramaekers, F. C. (1997) Histochem. Cell Biol. 107,505–517
14. Taniura, H., Glass, C., and Gerace, L. (1995) J. Cell Biol. 131, 33–4415. Smythe, C., Jenkins, H. E., and Hutchison, C. J. (2000) EMBO J. 19,
3918–393116. Hutchison, C. J., Alvarez-Reyes, M., and Vaughan, O. A. (2001) J. Cell Sci.
114, 9–1917. Liu, J., Ben-Shahar, T. R., Riemer, D., Treinin, M., Spann, P., Weber, K.,
Fire, A., and Gruenbaum, Y. (2000)Mol. Biol. Cell 11, 3937–394718. Harborth, J., Elbashir, S. M., Bechert, K., Tuschl, T., andWeber, K. (2001)
J. Cell Sci. 114, 4557–456519. Vergnes, L., Peterfy, M., Bergo, M. O., Young, S. G., and Reue, K. (2004)
Proc. Natl. Acad. Sci. U. S. A. 101, 10428–1043320. Padiath, Q. S., Saigoh, K., Schiffmann, R., Asahara, H., Yamada, T., Koep-
pen, A., Hogan, K., Ptacek, L. J., and Fu, Y. H. (2006) Nat. Genet. 38,1114–1123
21. Sullivan, T., Escalante-Alcalde, D., Bhatt, H., Anver, M., Bhat, N., Na-gashima, K., Stewart, C. L., and Burke, B. (1999) J. Cell Biol. 147, 913–920
22. Lammerding, J., Hsiao, J., Schulze, P. C., Kozlov, S., Stewart, C. L., and Lee,R. T. (2005) J. Cell Biol. 170, 781–791
23. Georgatos, S. D. (2001) EMBO J. 20, 2989–299424. Soderqvist, H., Imreh, G., Kihlmark, M., Linnman, C., Ringertz, N., and
Hallberg, E. (1997) Eur. J. Biochem. 250, 808–81325. Rabut, G., Doye, V., and Ellenberg, J. (2004) Nat. Cell Biol. 6, 1114–112126. Daigle, N., Beaudouin, J., Hartnell, L., Imreh, G., Hallberg, E., Lippincott-
Schwartz, J., and Ellenberg, J. (2001) J. Cell Biol. 154, 71–8427. Zhang, Q., Ragnauth, C. D., Skepper, J. N., Worth, N. F., Warren, D. T.,
Roberts, R. G., Weissberg, P. L., Ellis, J. A., and Shanahan, C. M. (2005)J. Cell Sci. 118, 673–687
28. Zhang, Q., Skepper, J. N., Yang, F., Davies, J. D., Hegyi, L., Roberts, R. G.,Weissberg, P. L., Ellis, J. A., and Shanahan, C. M. (2001) J. Cell Sci. 114,4485–4498
29. Crisp, M., Liu, Q., Roux, K., Rattner, J. B., Shanahan, C., Burke, B., Stahl,P. D., and Hodzic, D. (2006) J. Cell Biol. 172, 41–53
30. Padmakumar, V. C., Libotte, T., Lu, W., Zaim, H., Abraham, S., Noegel,A. A., Gotzmann, J., Foisner, R., and Karakesisoglou, I. (2005) J. Cell Sci.118, 3419–3430
31. Lee, J. S., Chang, M. I., Tseng, Y., and Wirtz, D. (2005)Mol. Biol. Cell 16,871–880
32. Lammerding, J., Fong, L. G., Ji, J. Y., Reue, K., Stewart, C. L., Young, S. G.,and Lee, R. T. (2006) J. Biol. Chem. 281, 25768–25780
33. Padmakumar, V. C., Abraham, S., Braune, S., Noegel, A. A., Tunggal, B.,Karakesisoglou, I., and Korenbaum, E. (2004) Exp. Cell Res. 295, 330–339
34. Libotte, T., Zaim, H., Abraham, S., Padmakumar, V. C., Schneider, M., Lu,W., Munck, M., Hutchison, C., Wehnert, M., Fahrenkrog, B., Sauder, U.,Aebi, U., Noegel, A. A., and Karakesisoglou, I. (2005) Mol. Biol. Cell 16,3411–3424
35. Worman, H. J., and Gundersen, G. G. (2006) Trends Cell Biol. 16, 67–6936. Haque, F., Lloyd, D. J., Smallwood, D. T., Dent, C. L., Shanahan, C.M., Fry,
A. M., Trembath, R. C., and Shackleton, S. (2006) Mol. Cell. Biol. 26,3738–3751
37. Pare, G. C., Easlick, J. L., Mislow, J. M., McNally, E. M., and Kapiloff, M. S.(2005) Exp. Cell Res. 303, 388–399
38. Munter, S., Enninga, J., Vazquez-Martinez, R., Delbarre, E., David-Watine,B., Nehrbass, U., and Shorte, S. L. (2006) BMC Cell Biol. 7, 23
39. Wilhelmsen, K., Litjens, S. H., Kuikman, I., Tshimbalanga, N., Janssen, H.,van den Bout, I., Raymond, K., and Sonnenberg, A. (2005) J. Cell Biol. 171,799–810
40. Hasan, S., Guttinger, S., Muhlhausser, P., Anderegg, F., Burgler, S., andKutay, U. (2006) FEBS Lett. 580, 1263–1268
41. McGee, M. D., Rillo, R., Anderson, A. S., and Starr, D. A. (2006)Mol. Biol.Cell 17, 1790–1801
42. Grady, R.M., Starr, D. A., Ackerman, G. L., Sanes, J. R., andHan,M. (2005)Proc. Natl. Acad. Sci. U. S. A. 102, 4359–4364
43. King, M. C., Lusk, C. P., and Blobel, G. (2006) Nature 442, 1003–1007
Cell Nuclei Spin in the Absence of Lamin B1
JULY 6, 2007 • VOLUME 282 • NUMBER 27 JOURNAL OF BIOLOGICAL CHEMISTRY 20025
at Massachusetts G
eneral Hospital T
readwell Library on July 1, 2007
ww
w.jbc.org
Dow
nloaded from

44. Mislow, J. M., Holaska, J. M., Kim, M. S., Lee, K. K., Segura-Totten, M.,Wilson, K. L., and McNally, E. M. (2002) FEBS Lett. 525, 135–140
45. Malone, C. J.,Misner, L., Le Bot, N., Tsai,M. C., Campbell, J.M., Ahringer,J., and White, J. G. (2003) Cell 115, 825–836
46. Fan, J., and Beck, K. A. (2004) J. Cell Sci. 117, 619–62947. Szabo, B., Kornyei, Z., Zach, J., Selmeczi, D., Csucs, G., Czirok, A., and
Vicsek, T. (2004) Cell. Motil. Cytoskeleton 59, 38–4948. Salina, D., Bodoor, K., Eckley, D. M., Schroer, T. A., Rattner, J. B., and
Burke, B. (2002) Cell 108, 97–10749. Maison, C., Pyrpasopoulou, A., and Georgatos, S. D. (1995) EMBO J. 14,
3311–3324
50. Hay, M., and De Boni, U. (1991) Cell Motil. Cytoskeleton 18, 63–7551. Pomerat, C. M. (1953) Exp. Cell Res. 5, 191–19652. Englander, L. L., and Rubin, L. L. (1987) J. Cell Biol. 104, 87–9553. Herbomel, P. (1999) Curr. Biol. 9, R627–R62854. De Boni, U. (1988) Anticancer Res 8, 885–89855. Worman, H. J., Yuan, J., Blobel, G., and Georgatos, S. D. (1988) Proc. Natl.
Acad. Sci. U. S. A. 85, 8531–853456. Malhas, A., Lee, C. F., Sanders, R., Saunders, N. J., and Vaux, D. J. (2007)
J. Cell Biol. 176, 593–60357. Bard, F., Bourgeois, C. A., Costagliola, D., and Bouteille, M. (1985) Biol.
Cell 54, 135–142
Cell Nuclei Spin in the Absence of Lamin B1
20026 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 282 • NUMBER 27 • JULY 6, 2007
at Massachusetts G
eneral Hospital T
readwell Library on July 1, 2007
ww
w.jbc.org
Dow
nloaded from




















