
เชื้ออะมีบา Entamoeba histolytica ซึ่งเป็นสาเหตุของโรคบิด ...
Upload
independentCategory
view
3download
0
Innate immunity prevents tissue invasion byEntamoeba histolytica
Mineko Shibayama, Vıctor Rivera-Aguilar, Elizabeth Barbosa-Cabrera,Saul Rojas-Hernandez, Adriana Jarillo-Luna, Vıctor Tsutsumi,Judith Pacheco-Yepez, and Rafael Campos-Rodrıguez
Abstract: Although innate and adaptive immunity both play a role in amoebiasis, the mechanisms involved in the elimina-tion of Entamoeba histolytica are poorly understood. To provide more information about the innate immune mechanismsthat may confer protection against invasive amoebiasis, we administered inflammatory substances (bacillus Calmette-Gue-rin, lipopolysaccharide, complete Freund’s adjuvant, or mineral oil) into the peritoneum of hamsters. The animals werethen challenged with pathogenic trophozoites of E. histolytica and, after 7 days, the protective host response was analysed.We found that the nonspecific inflammatory response induced in the peritoneum was sufficient to prevent liver invasionby E. histolytica. In vitro experiments showed that the killing of trophozoites was mediated by peritoneal macrophagesand a protein of 68 kDa with peroxidase activity.
Key words: Entamoeba histolytica, innate immunity, macrophages, nitric oxide, peroxidase.
Resume : Meme si l’immunite innee et acquise jouent toutes deux un role lors de l’amoebiase, les mecanismes impliquesdans l’elimination d’Entameoba histolytica sont mal compris. Afin d’obtenir plus d’informations sur les mecanismes del’immunite innee qui pourraient conferer une protection contre l’amoebiase invasive, nous avons administre des agents in-flammatoires (bacillus Calmette-Guerin, lipopolysaccharide, adjuvant complet de Freund ou de l’huile minerale) dans leperitoine du hamster. Les animaux ont ete soumis a une provocation avec les trophozoıtes pathogenes d’E. histolytica et laprotection de l’hote a ete analysee apres 7 jours. Nous avons trouve que la reponse inflammatoire non-specifique induitedans le peritoine etait suffisante pour prevenir l’invasion hepatique par E. histolytica. Des experiences in vitro ont demon-tre que la destruction des trophozoıtes etait realisee par l’intermediaire des macrophages peritoneaux et d’une proteine de68 kDa possedant une activite peroxydase.
Mots-cles : Entomoeba histolytica, immunite innee, macrophages, oxyde nitrique, peroxydase.
[Traduit par la Redaction]
IntroductionEntamoeba histolytica causes 2 major clinical syndromes,
amoebic colitis and amoebic liver abscess (ALA), which re-sult in 40 000 – 100 000 deaths worldwide each year (Anony-
mous 1997; Petri and Singh 1999). In Mexico, whereamoebiasis is endemic, >8% of the population has had anepisode of amoebiasis (Caballero-Salcedo et al. 1994). Thedevelopment of an effective vaccine would require a pro-found knowledge of the mechanisms of protective immunityagainst E. histolytica (Stanley 2006).
Little is known about protective immunity to amoebiasis,but apparently both innate and acquired immune responseslimit amoebic infection (Huston and Petri 1998; Seydel etal. 2000; Haque et al. 2002; Houpt et al. 2002; Asgharpouret al. 2005; Ivory et al. 2007). Although the mechanisms ofthe innate immunity involved in amoebiasis are poorlyunderstood, research has shown that innate immunity is es-sential for the elimination of the parasite.
The efficient innate immune response of genetically re-sistant individuals or species can prevent the invasion of theintestinal epithelium by E. histolytica (Seydel et al. 1997;Houpt et al. 2002). Their ability to resist tissue invasionmay be related to the following: (i) the innate capacity ofneutrophils and macrophages to generate reactive oxygen in-termediates (ROI) and reactive nitrogen intermediates(RNI); (ii) the expression of effector molecules, such asantimicrobial peptides, and components of the alternativeand mannan-binding lectin pathways of the complementsystem; (iii) the production of proinflammatory or anti-in-
Received 26 May 2008. Revision received 15 August 2008.Accepted 21 August 2008. Published on the NRC ResearchPress Web site at cjm.nrc.ca on 18 December 2008.
M. Shibayama and V. Tsutsumi. Department of ExperimentalPathology, CINVESTAV-IPN, Av. IPN No. 2508 Zacatenco,CP 07360, Mexico, DF.V. Rivera-Aguilar. Department of Microbiology, UBIPRO,FES-Iztacala, UNAM, Avenida de los Barrios s/n, TlalnepantlaEdo. de Mexico, CP 54090, Mexico, DF.E. Barbosa-Cabrera, S. Rojas-Hernandez, andR. Campos-Rodrıguez.1 Seccion de Estudios de Posgrado eInvestigacion, Escuela Superior de Medicina, InstitutoPolitecnico Nacional, Plan de San Luis y Diaz Miron, CP 11340,Mexico, DF.A. Jarillo-Luna. Department of Morphology, School ofMedicine, IPN, Plan de San Luis y Diaz Miron, CP 11340,Mexico, DF.J. Pacheco-Yepez. Laboratory of Electron Microscopy, MexicanFaculty of Medicine, La Salle University, Fuentes 17, Tlalpan,CP 14000, Mexico, DF.
1Corresponding author (e-mail: [email protected]).
1032
Can. J. Microbiol. 54: 1032–1042 (2008) doi:10.1139/W08-106 # 2008 NRC Canada

flammatory cytokines; (iv) the activity of nonhaematopoieticcells; and (v) the generation of an effective inflammatory re-sponse that contributes to the elimination of the parasite(Seydel et al. 2000; Shibayama et al. 2000; Haque et al.2001, 2002; Duggal et al. 2004; Asgharpour et al. 2005;Campos-Rodriguez and Jarillo-Luna 2005; Hamano et al.2006).
As further evidence that innate immunity is a key factorin the elimination of the parasite, individuals who are in-fected with human immunodeficiency virus (HIV) frequentlyshow asymptomatic colonization of the intestinal mucosawith E. histolytica (Reed et al. 1991; Jessurun et al. 1992).As acquired immunity is severely impaired in HIV patients,their innate intestinal immunity is probably responsible forthe success of these individuals in defending themselvesagainst systemic invasion by E. histolytica.
Finally, humoral and cellular components of the innateimmune system, such as the complement system, nitric ox-ide (NO), neutrophils, and macrophages, have been shownto kill trophozoites in vitro (Denis and Chadee 1989a,1989b; Lin and Chadee 1992). The data obtained frommouse models of amoebic colitis and hepatic amoebiasissupport the notion that innate immunity plays a major rolein protecting against tissue invasion by E. histolytica (Velaz-quez et al. 1997; Seydel et al. 2000; Houpt et al. 2002; Ja-rillo-Luna et al. 2002).
In this paper, the hypothesis that innate immunity is suffi-cient for the elimination of E. histolytica was analyzed withthe model of ALA produced by the intraperitoneal (i.p.) in-oculation of pathogenic trophozoites. Although the model ofi.p. injection of trophozoites to induce ALA is less commonthan the conventional intrahepatic (i.h.) or intraportal ALAmodels, we chose it because it also permits the analysis ofthe immunological and physiological responses of the host(Gold and Keisari 1990; Shibayama et al. 1998; Tsutsumiand Shibayama 2006). The i.p. model constitutes an easy toperform, highly reproducible procedure for inducing ALAsin hamsters (Jarumilinta 1966; Gold and Keisari 1990; Shi-bayama et al. 1998), and allows a rapid and efficient isola-tion of peritoneal inflammatory cells and molecules so thatthey can be analyzed in ex vivo and in vitro experiments. Itis important to mention that there are still no animal modelsin which cysts administered orally produce intestinal andspontaneous hepatic disease (Ackers and Mirelman 2006;Tsutsumi and Shibayama 2006).
In this study, we analyzed the effect of nonspecificallystimulated peritoneal exudate on the production of ALA,and examined some of the possible components of the peri-toneal exudate responsible for the destruction of amoebas.We found that the induced innate response was sufficient toprevent tissue invasion by E. histolytica. More specifically,a protein of 68 kDa was identified as having peroxidase andmarked amoebicidal activity.
Materials and methods
Amoebic culturesTrophozoites of the E. histolytica strain HMI:IMSS were
maintained axenically in BI-S33 medium, as described else-where (Diamond et al. 1978). The strain was subsequentlycloned and passaged 3 times in hamster liver to increase its
virulence. The inoculum was prepared from amoebic cul-tures at 72 h, when they were in the logarithmic growthphase.
Animals and production of ALAsMale adult golden hamsters (Mesocricetus auratus
(Waterhouse, 1839)), 2 months of age and weighing approx-imately 100 g, were used in all the experiments. The use andexperimentation to which they were subjected were carriedout in accordance with the institutional animal care commit-tee (IACUC, ID No. 244/05, Mexico). Groups of hamsters(n = 6) were challenged by the i.h. or i.p. route with 1.25 �105 and 2 � 106 trophozoites, respectively, as describedelsewhere (Tsutsumi et al. 1984; Shibayama et al. 1998).Seven days after inoculation, the animals were euthanizedwith an i.p. injection of sodium pentobarbital (10 mg / 100 gbody weight). To determine the percentage of liver tissue in-volved in the lesion, the abscess was dissected from theliver, and the mass of the abscess was divided by the massof the whole liver and multiplied by 100.
Determination of antibodies against E. histolytica inserum
Antibodies against E. histolytica IgG that were present inthe serum of the experimental and control groups werequantified with the ELISA method, as described elsewhere(Jarillo-Luna et al. 2002).
Administration of bacillus Calmette-Guerin andcomplete Freund’s adjuvant
One group of hamsters was injected weekly with 0.5 mgof bacillus Calmette-Guerin (BCG; Sigma-Aldrich, St.Louis, Missouri), and another group was injected with 1 mLof complete Freund’s adjuvant (CFA; 1 mg/mL of heat-killed Mycobacterium tuberculosis H37Ra). Three weekslater, the animals were challenged with 2 � 106 i.p. tropho-zoites of E. histolytica, were euthanized 7 days later, and theALAs were assessed.
Administration of incomplete Freund’s adjuvant andsterile mineral oil
Hamsters received a single dose of either 1 mL of incom-plete Freund’s adjuvant (IFA) or 3 mL of mineral oil(Sigma-Aldrich). Seven days later, animals were challengedwith i.p. trophozoites of E. histolytica, were euthanized7 days later, and the ALAs were assessed.
Peritoneal exudate cell samplesCells were harvested by peritoneal lavage with ice-cold
Hanks balanced saline solution (HBSS), containing 5 U/mLheparin. Cells were collected by centrifugation and washedwith ice-cold HBSS. The number of cells was counted in ahemocytometer, and viability was determined by trypan blueexclusion.
Purification of peritoneal macrophagesFor the cytotoxic assay, macrophages from the peritoneal
exudates of hamster treated with mineral oil were isolatedby density gradient centrifugation on Percoll (Sigma-Al-drich). The osmolarity of the Percoll was adjusted to matchnormal physiological conditions by adding 1 mL of 10-fold
Shibayama et al. 1033
# 2008 NRC Canada

concentrated HBSS to 9 mL of Percoll. This was diluted fur-ther with HBSS to give 30%, 60%, and 80% Percoll solu-tions, which were layered (2.5 mL each) into a 15 mL tube.The cell suspension (2 � 107 to 4 � 107 cells) was placedon top of the layers and centrifuged for 20 min (400g) atroom temperature. The cells at each interface were col-lected, then washed by pelleting (5 min at 400g) 3 times(Katayama et al. 1982; Hirata et al. 1989). The cells werediluted to 1 � 106 cells/mL HBSS, containing 2% heat-in-activated bovine serum and 5.7 mmol/L cysteine. The cellsrich in macrophages were recovered from the 60%–80% in-terface, and their viability was 80%–90%. These cells wereused immediately in the cytotoxic assay.
Cytochemical detection of trophozoites in peritonealexudate cells
Samples of peritoneal exudate cell (PEC) suspensions(100 mL) were incubated for 1 h at room temperature withrabbit polyclonal serum, prepared in our laboratory, againstE. histolytica diluted 1:200 in 0.025% PBS–Tween (PBST).Control samples were incubated with PBS alone. After incu-bation, the samples were washed 3 times for 5 min in PBST,and then incubated for 1 h at room temperature with perox-idase-labelled goat IgG directed against rabbit antigens(Sigma-Aldrich) diluted 1:250 in PBST. The reaction wasdeveloped using immunopure metal-enhanced diaminobenzi-dine (Pierce; Rockford, Illinois) for 10 min; the plates werethen washed with PBS and examined using a light micro-scope.
Passive immunization studiesGroups of 10–12 animals were inoculated by the i.p. route
with 8 mL of mineral oil (Sigma-Aldrich) or saline.Seven days after inoculation, the animals were euthanizedand the peritoneal cavities of normal and mineral-oil-treatedhamsters were washed with 10 mL of cold HBSS, contain-ing 5 U/mL heparin. Cells were collected by centrifugation(400g for 10 min at room temperature), washed twice withcold HBSS, and adjusted to 4 � 106 cells/mL. The superna-tants were also collected and stored at –70 8C. Five milli-litres of the supernatant was inoculated by the i.p. route intoa group of recipient animals (n = 6). Another 2 groups ofrecipient animals (n = 6–8) were inoculated by the i.p. routewith 1 or 3 mL of the peritoneal cell suspension, containing4 � 106 or 12 � 106 cells, respectively. Next, noninoculated(control group) and recipient groups were inoculated with2 � 106 trophozoites; 7 days later the hamsters were euthan-ized, and the percentage of liver affected by the lesion wasdetermined.
Cytotoxicity assayMeasurement of the in vitro cytotoxic activity of the peri-
toneal macrophages against E. histolytica trophozoites wascarried out, as described by Ravdin et al. (1985), with minorchanges. Amoebas and peritoneal macrophages were mixedin a ratio of 1:10 in HBSS, containing 2% heat-inactivatedbovine serum and 5.7 mmol/L cysteine, then centrifuged at150g for 5 min at room temperature to promote cell to cellcontact. After incubation for 1–3 h at 37 8C, the pellet wasdisrupted by vortexing. The number of amoebas and macro-phages was counted in a hemocytometer, and their viability
was determined by the trypan blue exclusion assay. Datawere expressed as percentages of viable cells at each inter-val studied (Ravdin et al. 1985).
The in vitro cytotoxic activity of the peritoneal superna-tants against E. histolytica trophozoites was assayed by incu-bating 100 or 500 mL aliquots of the supernatant with 5 �104 trophozoites for 3 h at 37 8C; upon completion, the via-bility of the amoebae was assessed by the trypan blue exclu-sion assay.
Amoebicidal effect of low-molecular-mass moleculesTo analyse the possible amoebicidal effect of low-molec-
ular-mass molecules in vivo, we introduced 1 � 106 troph-ozoites in dialysis bags (10 mm; molecular mass cutoff, 12–14 kDa) that were placed in the peritoneal cavity of anaes-thetized hamsters to which mineral oil had previously beenadministered and in control animals. After 4 h, the tropho-zoites inside the bags were harvested by centrifugation, andtheir viability was determined by the trypan blue exclusionassay. Nitrate and nitrite concentrations in the supernatantwere determined using the Griess method (Green et al.1982).
Gel-filtration chromatographyThe supernatant of the peritoneal lavage was concen-
trated 10 times by lyophilization, dialyzed in loading buffer(0.01 mol/L phosphate (pH 8.5) and applied to a DEAE-Sepharose column (2.5 cm � 48 cm). The column waseluted with a 50–500 mmol/L NaCl gradient in 0.01 mol/Lphosphate (pH 8.5). Flow rate was maintained at 40 mL/hand 2 mL fractions were collected. The presence of pro-teins in the column effluent was monitored by UV absorb-ance at 280 nm. The amoebicidal activity in the differentfractions (I–VII) was assayed using a trypan blue exclusionassay, and SDS–PAGE was carried out to examine theircomposition.
Fraction II was concentrated by lyophilization, dissolvedin 2 mL of 0.01 mol/L phosphate (pH 8.5), and then appliedto a Sepharose 4B-CL column (2.5 cm � 48 cm). Flow ratewas maintained at 1 mL/min and 1 mL fractions were col-lected. The presence of proteins in the column effluent wasmonitored by UV absorbance at 280 nm. The amoebicidalactivity in the different fractions was assayed as before, andSDS–PAGE was carried out to examine their composition.
SDS–PAGEThe peritoneal fractions were mixed with buffer (2%
SDS, 10% 0.5 mol/L Tris (pH 6.8), and glycerol) and boiledfor 3 min. Twenty microlitres of the sample was electro-phoresed using 10% SDS–PAGE. Gels were washed andstained with Coomassie Brilliant Blue R-250 (Bio-Rad,Mexico City, Mexico) for detection of proteins.
Spectrophotometric assay of peroxidaseThe enzyme activity for peroxidase was determined by
measuring the H2O2-dependent oxidation of o-dianisidinespectrophotometrically at 490 nm. In brief, 1769 mL of50 mmol/L sodium phosphate buffer and 30 mL of0.04 mmol/L o-dianisidine were mixed with 200 mL of peri-toneal fluid, and the background value was set at 0. Afterthe addition of 1 mL of 30% H2O2 to the mixture to start
1034 Can. J. Microbiol. Vol. 54, 2008
# 2008 NRC Canada

the reaction, the absorbance at 490 nm was recorded every30 s for 9 min, using the kinetic spectrophotometer. Opticaldensity units were converted into units of concentration, us-ing the molar absorption coefficient for oxidized o-dianisi-dine (3 = 10 062 (mol/L � cm)–1) (Xia and Zweier 1997).
Data analysisThe data obtained from each experiment were expressed
as mean ± SD, and the statistical difference between experi-mental groups was calculated using a one-way analysis ofvariance (ANOVA). If a significant difference in the maineffect was identified (p < 0.05), the means of the respectivegroups were compared using the Bonferroni test. Differencesbetween 2 means were analysed by the nonpaired Student’s ttest. All analyses were performed using the statistical pro-gram SigmaStat for Windows, version 2.03 (SPSS Inc.).
Results
Immunization with BCG or CFA prevented hepaticamoebiasis
Hamsters that were challenged intraperitoneally with E.histolytica trophozoites 7 days after i.p. administration ofBCG or CFA showed no significant liver damage. In con-trast, control hamsters treated with saline solution developedALAs that were approximately 40% of the total hepatic
mass (Fig. 1). However, in hamsters challenged by the i.h.route, the size of the abscesses was similar to those of theuntreated controls. The results show that bacterial compo-nents induced protective immunity against E. histolytica,which operated exclusively in the microenvironment of theperitoneum.
Amoebae were destroyed before the production ofspecific serum antibodies
When CFA was administered to hamsters by the i.p.route, the amoebae inoculated by the same route disappeared48 h postinoculation (Fig. 2), before the production of spe-cific antibodies. This effect was probably mediated by fastand nonspecific innate mechanisms that killed the amoebaeand, consequently, prevented tissue damage.
Administration of nonbiologic and nonspecific stimulants(IFA or mineral oil) prevented hepatic amoebiasis
Seven days before challenge with E. histolytica tropho-zoites, hamsters were treated with nonbiological and nonspe-cific peritoneal stimulants, such as IFA or mineral oil, whichelicit inflammatory exudates with numerous mononuclearphagocytes and neutrophils. Seven days after this challenge,animals were euthanized and no liver damage was found(100% protection). In contrast, untreated control hamsterspresented ALAs that were approximately 40% of the totalhepatic mass (Fig. 3). Thus, inflammatory cells or their
Fig. 1. Protection of golden hamsters (Mesocricetus auratus) fromamoebic liver abscess (ALA) by intraperitoneal (i.p.) treatment withbacillus Calmette-Guerin (BCG) and complete Freund’s adjuvant(CFA). Hamsters inoculated with BCG or CFA 7 days after treat-ment were challenged with intrahepatic (i.h.) or i.p. trophozoites;7 days later they were euthanized, and the size of the ALA was de-termined. Data are expressed as the mean ± SD of the percentageof the liver affected by abscesses. In hamsters inoculated with CFAor BCG and challenged by the i.p. route, ALAs were significantlysmaller than in the control group (*, p < 0.001). In contrast, inhamsters challenged by the i.h. route, ALAs were similar in size tothose of the control group.
Fig. 2. Amoebae were destroyed before the development of specificserum immunoglobulin (Ig)G antibodies. Golden hamsters (Meso-cricetus auratus; n = 6) were administered 1 mL of completeFreund’s adjuvant (CFA), and 7 days later were challenged withi.p. trophozoites. The peritoneal exudate cells (PECs) were obtainedfrom hamsters euthanized at 6 h, 24 h, 48 h, 5 days, and 7 days.The number of trophozoites in PECs was counted using immuno-cytochemical analysis, and serum IgG antibodies were quantifiedby ELISA. Data are expressed as the mean ± SD of the number oftrophozoites per 100 PECs and as the absorbance at 490 nm.Although trophozoites disappeared at 48 h postinoculation, the le-vels of IgG antibodies against Entamoeba histolytica increased sig-nificantly up to day 7.
Shibayama et al. 1035
# 2008 NRC Canada
products that lack specificity for amoebic antigens were ableto prevent tissue damage inflicted by E. histolytica. Whenthe i.p. administration of CFA, IFA, or mineral oil wasdone simultaneously with the administration of E. histolytca,ALAs were similar to those of the controls (data not shown),indicating that these compounds are not directly responsiblefor damage to trophozoites.
Transfer of immunity by cells and peritoneal supernatantTo characterize the mechanisms involved in the destruc-
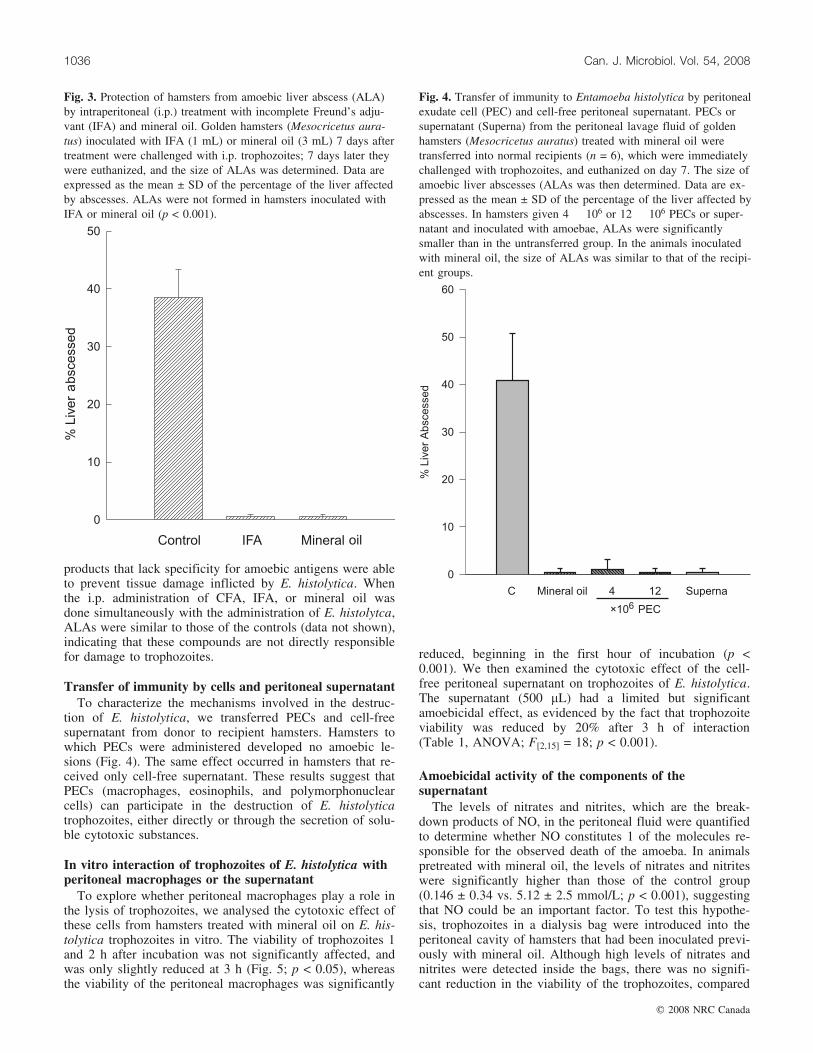
tion of E. histolytica, we transferred PECs and cell-freesupernatant from donor to recipient hamsters. Hamsters towhich PECs were administered developed no amoebic le-sions (Fig. 4). The same effect occurred in hamsters that re-ceived only cell-free supernatant. These results suggest thatPECs (macrophages, eosinophils, and polymorphonuclearcells) can participate in the destruction of E. histolyticatrophozoites, either directly or through the secretion of solu-ble cytotoxic substances.
In vitro interaction of trophozoites of E. histolytica withperitoneal macrophages or the supernatant
To explore whether peritoneal macrophages play a role inthe lysis of trophozoites, we analysed the cytotoxic effect ofthese cells from hamsters treated with mineral oil on E. his-tolytica trophozoites in vitro. The viability of trophozoites 1and 2 h after incubation was not significantly affected, andwas only slightly reduced at 3 h (Fig. 5; p < 0.05), whereasthe viability of the peritoneal macrophages was significantly
reduced, beginning in the first hour of incubation (p <0.001). We then examined the cytotoxic effect of the cell-free peritoneal supernatant on trophozoites of E. histolytica.The supernatant (500 mL) had a limited but significantamoebicidal effect, as evidenced by the fact that trophozoiteviability was reduced by 20% after 3 h of interaction(Table 1, ANOVA; F[2,15] = 18; p < 0.001).
Amoebicidal activity of the components of thesupernatant
The levels of nitrates and nitrites, which are the break-down products of NO, in the peritoneal fluid were quantifiedto determine whether NO constitutes 1 of the molecules re-sponsible for the observed death of the amoeba. In animalspretreated with mineral oil, the levels of nitrates and nitriteswere significantly higher than those of the control group(0.146 ± 0.34 vs. 5.12 ± 2.5 mmol/L; p < 0.001), suggestingthat NO could be an important factor. To test this hypothe-sis, trophozoites in a dialysis bag were introduced into theperitoneal cavity of hamsters that had been inoculated previ-ously with mineral oil. Although high levels of nitrates andnitrites were detected inside the bags, there was no signifi-cant reduction in the viability of the trophozoites, compared
Fig. 3. Protection of hamsters from amoebic liver abscess (ALA)by intraperitoneal (i.p.) treatment with incomplete Freund’s adju-vant (IFA) and mineral oil. Golden hamsters (Mesocricetus aura-tus) inoculated with IFA (1 mL) or mineral oil (3 mL) 7 days aftertreatment were challenged with i.p. trophozoites; 7 days later theywere euthanized, and the size of ALAs was determined. Data areexpressed as the mean ± SD of the percentage of the liver affectedby abscesses. ALAs were not formed in hamsters inoculated withIFA or mineral oil (p < 0.001).
Fig. 4. Transfer of immunity to Entamoeba histolytica by peritonealexudate cell (PEC) and cell-free peritoneal supernatant. PECs orsupernatant (Superna) from the peritoneal lavage fluid of goldenhamsters (Mesocricetus auratus) treated with mineral oil weretransferred into normal recipients (n = 6), which were immediatelychallenged with trophozoites, and euthanized on day 7. The size ofamoebic liver abscesses (ALAs was then determined. Data are ex-pressed as the mean ± SD of the percentage of the liver affected byabscesses. In hamsters given 4 � 106 or 12 � 106 PECs or super-natant and inoculated with amoebae, ALAs were significantlysmaller than in the untransferred group. In the animals inoculatedwith mineral oil, the size of ALAs was similar to that of the recipi-ent groups.
1036 Can. J. Microbiol. Vol. 54, 2008
# 2008 NRC Canada

with the control group, ruling out the possibility that NOwas responsible for the observed amoebicidal effect (Fig. 6).
Purification of molecules with amoebicidal activityTo purify the molecules with possible amoebicidal activ-
ity, the peritoneal supernatant was concentrated 10 times bylyophilization, and then protein fractions were separated ona DEAE-Sepharose column eluted with a discontinuous gra-dient of phosphates (Fig. 7). The amoebicidal activity ineach fraction was evaluated by the trypan blue exclusion as-say. The greatest antiamoebic activity was found in fractionsII and III, which reduced the amoebic viability by 78% and68%, respectively, in comparison with the control (Table 2).The fractions eluted were subjected to SDS–PAGE underdenaturing conditions. The fractions with the greatest amoe-bicidal activity contain proteins of 68 and 160 kDa (Fig. 7,insert). However, it is possible that other proteins of lowerabundance were present with the 68 and 160 kDa proteins.
To further purify the molecules with amoebicidal activity,fraction II was concentrated by lyophilization, and protein
fractions with different molecular masses were separated ona Sepharose CL-4B column eluted with PBS (Fig. 8). Thegreatest antiamoebic activity was found in fraction II-2,which reduced the amoebic viability by 60% in comparisonwith the control (Table 3). The fractions eluted from the Se-pharose CL-4B column were subjected to SDS–PAGE underdenaturing conditions. Fraction II-2, with the greatest amoe-bicidal activity, contains a protein of 68 kDa (Fig. 8, insert).The amoebicidal activity of fraction II-2 was heat labile(data not shown).
The protein of 68 kDa has the activity of peroxidaseSince the molecule with the greater antiamoebic activity
has a molecular mass of 68 kDa, similar to that of the heavychains of myeloperoxidase (60 kDa) and eosinophil peroxi-dase (55 kDa), we determined the peroxidase activity in thefractions obtained by chromatography. The activity of per-oxidase was mainly found in fraction II-2, which containsthe 68 kDa protein (Fig. 9).
DiscussionThe most important finding of this work is that nonspe-
cific immune mechanisms were able to eliminate tropho-zoites of E. histolytica without the participation ofantibodies or T cells specific for amoebic antigens.
In our initial experiment, we found a significant decreasein the size of ALAs in hamsters immunized with CFA orBCG and inoculated with trophozoites by the i.p. route, butnot when trophozoites were inoculated directly into the liverparenchyma. According to the results of another study, theapplication of BCG does not induce resistance to E. histoly-tica when the trophozoites are inoculated by the i.p. route(Gold and Keisari 1990). The differences between the
Fig. 5. Interaction of amoebae and peritoneal macrophages. Tro-phozoites (Eh) and an enriched population of macrophages (Mac)were mixed, and cell viability was determined by trypan blue ex-clusion. Whereas the viability of Eh was slightly reduced only at3 h (p < 0.05), the viability of Mac was significantly reduced fromthe first hour of interaction (p < 0.001). Eh alone and Mac alonerepresent the viability of Ehs and Macs incubated in Hanks ba-lanced saline solution.
Table 1. Amoebicidal effect of peritoneal fluid.
Medium Amoebic viability (%) pNaCl 78±6Supernatant
100 mL 73±5 0.312500 mL 61±3 <0.001*
Note: Trophozoites were incubated for 3 h with differentvolumes of supernatant from the peritoneal lavage fluid, and theviability of trophozoites was determined by trypan blueexclusion assay. The data are expressed as the mean ± SD oftriplicate experiments.
*Only 500 mL of supernatant reduced the number of amoebae,compared with the saline control (p < 0.001, Bonferroni test).
Fig. 6. Nitric oxide did not show amoebicidal activity. Tropho-zoites in a dialysis bag were introduced into the peritoneal cavity ofgolden hamsters (Mesocricetus auratus) previously injected withmineral oil. There was no reduction in the viability of the amoebaein relation to the control group injected with saline (p > 0.5),although the levels of nitrates and nitrates were significantly greaterin the animals given mineral oil (*, p < 0.01).
Shibayama et al. 1037
# 2008 NRC Canada

2 studies could have resulted from the different treatment re-gimes or doses of BCG used. However, in the same study,resistance to E. histolytica was induced by treatment withglucan, an agent that stimulates a strong inflammatory re-sponse (Gold and Keisari 1990). Although i.h. inoculationof trophozoites is a commonly used technique to causeALA, we found only 1 previous study that employed it andused BCG as the immunostimulant (Ghadirian and Meero-vitch 1982). Those results show a significant reduction inthe size of ALAs after challenge, which is contrary to theresults of this study. These differences may have resultedfrom the higher doses of BCG used in the earlier study(2 mg vs. 0.5 mg).
The fact that BCG inoculated by the i.p. route only in-duced protection when amoebae were also inoculated by thesame route suggests the participation of cells or molecules
located in the inflamed peritoneum (e.g., macrophages orNO). Even though BCG inoculated by the i.p. route can ac-tivate Kupffer cells (McCuskey et al. 1983; Matsuo et al.1985), the cells have been shown to be incapable of elimi-nating amoebae that have invaded the liver, possibly be-cause the cells are few in number or show enhancedsusceptibility to the inhibitory effects of amoebic proteins(Wang and Chadee 1995).
Although several studies have shown that activated mac-rophages are the main effector cell in the host defenceagainst E. histolytica (Ghadirian and Meerovitch 1982; Or-tiz-Ortiz et al. 1990; Denis and Ghadirian 1992; Lin andChadee 1992), the mechanism by which i.p. infection withBCG induces nonspecific resistance to E. histolytica has notbeen clearly defined. Following inoculation of BCG, perito-neal macrophages are probably activated by cytokines, espe-cially interferon-g (IFN-g), produced by CD4+ T cellsstimulated by mycobacterial antigens (Kaufmann 2001; vanCrevel et al. 2002). The absence of amoebic antigens pre-cludes the possibility that macrophages were directly acti-vated by T cells with specificity for amoebic antigens(Perez-Tamayo et al. 1986).
To analyze whether induced nonspecific inflammatory re-sponse is sufficient to prevent liver invasion by E. histoly-tica, we injected agents such as IFA or mineral oil. ALAswere not formed in hamsters inoculated with those agentsand challenged with E. histolytica by the i.p. route, indicat-ing 100% protection. It is probable that nonspecific stimula-tion of PECs, among which neutrophils and macrophagesare the most abundant, prevented amoebic invasion of theliver. In animals injected with mineral oil, the inoculatedtrophozoites had almost completely disappeared 48 h postin-oculation, at which time IgG antibodies specific to E. histo-
Fig. 7. Chromatography of peritoneal proteins on a DEAE-Sepharose column. Two millilitres of concentrated supernatant of the peritonealexudate was applied to a column of DEAE-Sepharose (2.5 cm � 48 cm), the flow rate was adjusted to 40 mL/h, and 2 mL fractions werecollected and assayed for absorbance at 280 nm. Seven fractions were selected, and their amoebicidal effect was assayed by trypan blueexclusion assay. Insert: Fractions II, III, and IV, with the highest amoebicidal activity, were analysed by SDS–PAGE. Lane 1: molecularstandards. Lanes II, III, and IV: fractions II, III, and IV, respectively.
Table 2. Amoebicidal activity in fractions obtained by chroma-tography on a DEAE-Sepharose column.
Fraction Amoebic viability (%) pNaCl 78±3I 71±15 0.34II 17±2 <0.001*III 25±2 <0.001*IV 33±6 <0.001*V 50±14 0.15VI 70±10 0.13VII 73±6 0.19
Note: Trophozoites were incubated with the fractions for 3 h, andtheir viability was quantified by the trypan blue exclusion assay. Dataare mean ± SD.
*The amoebic viability was significantly lower in trophozoitesincubated with fractions II, III, and IV (p < 0.01, Bonferroni test).
1038 Can. J. Microbiol. Vol. 54, 2008
# 2008 NRC Canada

lytica had not yet been produced. This suggests that theelimination of amoebae depends on rapid nonspecific effec-tor mechanisms. IFA and mineral oil may induce resistanceto amoebic infection by a mechanism that involves a T cell-independent pathway of macrophage activation, similar tothe model of infection by Listeria monocytogenes in athymicmice, lethally irradiated mice, and severe combined immu-nodeficiency mice (Cole 1975; Bancroft et al. 1991; Rogerset al. 1992).
Because our previous results did not allow for a differen-tiation between the effects mediated by inflammatory cellsand those mediated by soluble factors, we did passive trans-fer experiments with PECs and peritoneal supernatant. Theresults showed that the destruction of E. histolytica tropho-zoites resulted from the activity of the PECs and soluble cy-totoxic substances. Therefore, we decided to test the in vitroeffect of each of these agents separately.
The viability of trophozoites was lightly but significantlyaffected by peritoneal macrophages only at 3h of incubation.These results confirm previous reports that showed that acti-vated neutrophils and macrophages are directly involved inthe killing of E. histolytica trophozoites, through a contact-
dependent process (Denis and Chadee 1989b; Lin and Cha-dee 1992; Ghadirian and Salimi 1993; Salata et al. 1985).However, the viability of peritoneal macrophages was sig-nificantly affected, starting at the first hour of incubation.These results are in agreement with previous studies showingthat E. histolytica kills host cells through contact-dependentmechanisms (Guerrant et al. 1981; Ravdin et al. 1985; Salataet al. 1985). In summary, the principal cytotoxic activity isrelated to the trophozoites, although the macrophages had acertain cytotoxic effect.
Fig. 8. Chromatography of fraction II on a Sepharose 4B-CL column. Two millilitres of fraction II obtained from the DEAE-Sepharosecolumn was applied to a Sepharose 4B-CL column, the flow rate was adjusted to 1 mL/min, and 1 mL fractions were collected and assayedfor absorbance at 280 nm. Two fractions were selected, and their amoebicidal effect was assayed by trypan blue exclusion assay. Insert:Fraction II-2, with the highest amoebicidal activity, was analysed by SDS–PAGE. Lane 1, molecular standards; lane 2, fraction II-2.
Fig. 9. Peroxidase activity. The peroxidase activity in fractions II-1and II-2 was assayed by spectrophotometric measurements at460 nm, using o-dianisidine as a substrate. The reaction mixturecontained 0.53 mmol/L o-dianisidine and 0.15 mmol/L H2O2 atroom temperature. Activity was only found in fraction II-2. Valuesare means of triplicate wells from 1 representative experiment of 3.
Table 3. Amoebicidal activity in fractions obtained by chromato-graphy on a Sepharose CL-4B column.
Fraction Amoebic viability (%) pNaCl 83±4II-1 76±7 0.27II-2 33±8 <0.001*
Note: Trophozoites were incubated with the fractions for 3 h, and theirviability was quantified by the trypan blue exclusion assay. Data aremean ± SD.
*The amoebic viability was significantly lower in trophozoites incubatedwith fraction II-2 (p < 0.001, Bonferroni test).
Shibayama et al. 1039
# 2008 NRC Canada

However, cell-free supernatant reduced amoebic viabilitysignificantly, indicating that soluble molecules have amoebi-cidal properties in vitro. The specific cytotoxic moleculesresponsible for the amoebicidal effect are unclear. Previousin vitro studies have shown that the amoebicidal activity ofactivated macrophages depends mainly on the synthesis ofNO (Lin and Chadee 1992; Lin et al. 1994), and that induci-ble NO synthase activity is important for the control of hep-atic amoebiasis in immunocompetent mice (Seydel et al.2000). This suggests that the amoebicidal mechanisms maybe dependent on ROI and RNI. In support of this hypothesis,we found that the level of NO production in the peritonealcavity of animals inoculated with mineral oil, measured asthe levels of nitrates and nitrites, was significantly higherthan that of untreated controls. However, trophozoites main-tained inside dialysis bags that were permeable to NO andother low-molecular-mass molecules (<10 kDa) were not de-stroyed when placed in the peritoneum of hamsters treatedwith mineral oil. These results suggest that NO, other RNIand ROI, and microbicidal peptides of low molecular mass(Oppenheim et al. 2003; Brogden 2005) probably did notparticipate in the destruction of amoebae, which has beensuggested by other studies (Pacheco-Yepez et al. 2001;Ramirez-Emiliano et al. 2005). However, since it took about36–48 h to see substantial killing of trophozoites (Fig. 2),4 h may not be long enough for NO, or other small mole-cules, to induce measurable cell death. Consequently, therole of NO in the innate immune response to E. histolyticacannot be excluded on the basis of this set of experiments.
To continue our search for the molecules responsible for thekilling of amoebae, the proteins in the peritoneal supernatantwere purified by chromatography on DEAE-Sepharose and inSepharose Cl-4B, and analysed by SDS–PAGE. A proteinwith a molecular mass of 68 kDa showed significant amoebi-cidal and peroxidase activity in vitro. Such activity was shownto be heat labile, indicating that it is indeed a protein that isresponsible for this activity. This protein likely correspondsto myeloperoxidase produced by monocytes and neutrophils,or to peroxidase produced by eosinophils. It is known thatmonocytes, neutrophils, and eosinophils utilize a peroxidaseenzyme, in combination with H202 derived from the respira-tory burst, and a halide to produce cytocidal hypohalous acidoxidants (Thomas and Fishman 1986; Klebanoff 2005; Hans-son et al. 2006; Wang et al. 2006). Hydrogen peroxide and hy-pochlorite can oxidize and halogenate essential structural andfunctional components in E. histolytica trophozoites, causinginhibition of their growth and loss of amoebic viability(Murray et al. 1981; Ghadirian et al. 1986; Denis and Chadee1989a).
In summary, we found that the induced innate responsewas sufficient to prevent tissue invasion by E. histolyticaand, consequently, adaptive immunity was not a relevantmechanism of protection in our animal model of amoebiasis.The amoebicidal activity was mediated by peritoneal macro-phages and a protein of 68 kDa. This protein had peroxidaseactivity and, therefore, probably kills the trophozoites byproducing hypohalous acid oxidants.
AcknowledgementsThis work was supported in part by grants from SIP-IPN
(Mexico). The authors thank A. Larsen for reading the
manuscript and for providing helpful comments. This studyand the experiments comply with the current laws on animalexperimentation in Mexico.
ReferencesAckers, J.P., and Mirelman, D. 2006. Progress in research on Enta-
moeba histolytica pathogenesis. Curr. Opin. Microbiol. 9: 367–373. doi:10.1016/j.mib.2006.06.014. PMID:16824782.
Anonymous. 1997. Amoebiasis. Wkly. Epidemiol. Rec. 72: 97–99.PMID:9100475.
Asgharpour,A.,Gilchrist,C.,Baba,D.,Hamano,S.,andHoupt,E.2005.Resistance to intestinal Entamoeba histolytica infection is conferredby innate immunity and Gr-1+ cells. Infect. Immun. 73: 4522–4529.doi:10.1128/IAI.73.8.4522-4529.2005.PMID:16040963.
Bancroft, G.J., Schreiber, R.D., and Unanue, E.R. 1991. Naturalimmunity: a T-cell-independent pathway of macrophage activa-tion, defined in the scid mouse. Immunol. Rev. 124: 5–24.doi:10.1111/j.1600-065X.1991.tb00613.x. PMID:1804781.
Brogden, K.A. 2005. Antimicrobial peptides: pore formers or meta-bolic inhibitors in bacteria? Nat. Rev. Microbiol. 3: 238–250.doi:10.1038/nrmicro1098. PMID:15703760.
Caballero-Salcedo, A., Viveros-Rogel, M., Salvatierra, B., Tapia-Conyer, R., Sepulveda-Amor, J., Gutierrez, G., and Ortiz-Ortiz,L. 1994. Seroepidemiology of amebiasis in Mexico. Am. J.Trop. Med. Hyg. 50: 412–419. PMID:8166347.
Campos-Rodriguez, R., and Jarillo-Luna, A. 2005. The pathogeni-city of Entamoeba histolytica is related to the capacity of evad-ing innate immunity. Parasite Immunol. 27: 1–8. doi:10.1111/j.1365-3024.2005.00743.x. PMID:15813717.
Cole, P. 1975. Activation of mouse peritoneal cells to kill Listeriamonocytogenes by T-lymphocyte products. Infect. Immun. 12:36–41. PMID:806534.
Denis, M., and Chadee, K. 1989a. Human neutrophils activated byinterferon-g and tumour necrosis factor-a kill Entamoeba histoly-tica trophozoites in vitro. J. Leukoc. Biol. 46: 270–274.PMID:2547889.
Denis, M., and Chadee, K. 1989b. Cytokine activation of murinemacrophages for in vitro killing of Entamoeba histolytica tro-phozoites. Infect. Immun. 57: 1750–1756. PMID:2542164.
Denis, M., and Ghadirian, E. 1992. Activated mouse macrophageskill Entamoeba histolytica trophozoites by releasing reactive ni-trogen intermediates. Microb. Pathog. 12: 193–198. doi:10.1016/0882-4010(92)90053-Q. PMID:1614330.
Diamond, L.S., Harlow, D.R., and Cunnick, C.C. 1978. A newmedium for the axenic cultivation of Entamoeba histolytica andother Entamoeba. Trans. R. Soc. Trop. Med. Hyg. 72: 431–432.doi:10.1016/0035-9203(78)90144-X. PMID:212851.
Duggal, P., Haque, R., Roy, S., Mondal, D., Sack, R.B., Farr, B.M.,et al. 2004. Influence of human leukocyte antigen class II alleleson susceptibility to Entamoeba histolytica infection in Banglade-shi children. J. Infect. Dis. 189: 520–526. doi:10.1086/381272.PMID:14745711.
Ghadirian, E., and Meerovitch, E. 1982. Macrophage requirementfor host defence against experimental hepatic amoebiasis in thehamster. Parasite Immunol. 4: 219–225. doi:10.1111/j.1365-3024.1982.tb00433.x. PMID:6289225.
Ghadirian, E., and Salimi, A. 1993. In vitro effect of recombinantinterferon gamma in combination with LPS on amoebicidal ac-tivity of murine Kupffer cells. Immunobiology, 188: 203–219.PMID:8406558.
Ghadirian, E., Somerfield, S.D., and Kongshavn, P.A. 1986. Sus-ceptibility of Entamoeba histolytica to oxidants. Infect. Immun.51: 263–267. PMID:2867045.
1040 Can. J. Microbiol. Vol. 54, 2008
# 2008 NRC Canada

Gold, D., and Keisari, Y. 1990. The effect of immunosuppressiveand immunostimulatory treatment on experimental amoebiasis.Ann. Trop. Med. Parasitol. 84: 573–580. PMID:2076035.
Green, L.C., Wagner, D.A., Glogowski, J., Skipper, P.L., Wishnok,J.S., and Tannenbaum, S.R. 1982. Analysis of nitrate, nitrite,and [15N]nitrate in biological fluids. Anal. Biochem. 126: 131–138. doi:10.1016/0003-2697(82)90118-X. PMID:7181105.
Guerrant, R.L., Brush, J., Ravdin, J.I., Sullivan, J.A., and Mandell,G.L. 1981. Interaction between Entamoeba histolytica and hu-man polymorphonuclear neutrophils. J. Infect. Dis. 143: 83–93.PMID:6260869.
Hamano, S., Asgharpour, A., Stroup, S.E., Wynn, T.A., Leiter,E.H., and Houpt, E. 2006. Resistance of C57BL/6 mice to amoe-biasis is mediated by nonhemopoietic cells but requires hemo-poietic IL-10 production. J. Immunol. 177: 1208–1213.PMID:16818779.
Hansson, M., Olsson, I., and Nauseef, W.M. 2006. Biosynthesis,processing, and sorting of human myeloperoxidase. Arch. Bio-chem. Biophys. 445: 214–224. doi:10.1016/j.abb.2005.08.009.PMID:16183032.
Haque, R., Ali, I.M., Sack, R.B., Farr, B.M., Ramakrishnan, G.,and Petri, W.A., Jr. 2001. Amebiasis and mucosal IgA antibodyagainst the Entamoeba histolytica adherence lectin in Banglade-shi children. J. Infect. Dis. 183: 1787–1793. doi:10.1086/320740. PMID:11372032.
Haque, R., Duggal, P., Ali, I.M., Hossain, M.B., Mondal, D., Sack,R.B., et al. 2002. Innate and acquired resistance to amebiasis inBangladeshi children. J. Infect. Dis. 186: 547–552. doi:10.1086/341566. PMID:12195383.
Hirata, K., Pele, J.P., Robidoux, C., and Sirois, P. 1989. Guinea piglung eosinophil: purification and prostaglandin production. J.Leukoc. Biol. 45: 523–528. PMID:2723536.
Houpt, E.R., Glembocki, D.J., Obrig, T.G., Moskaluk, C.A.,Lockhart, L.A., Wright, R.L., et al. 2002. The mouse model ofamebic colitis reveals mouse strain susceptibility to infectionand exacerbation of disease by CD4+ T cells. J. Immunol. 169:4496–4503. PMID:12370386.
Huston, C.D., and Petri, W.A., Jr. 1998. Host–pathogen interactionin amebiasis and progress in vaccine development. Eur. J. Clin.Microbiol. Infect. Dis. 17: 601–614. doi:10.1007/s100960050143. PMID:9832261.
Ivory, C., Kammanadiminti, S., and Chadee, K. 2007. Innate resis-tance to Entamoeba histolytica in murine models. Trends Parasi-tol. 23: 46–48. doi:10.1016/j.pt.2006.12.006. PMID:17185037.
Jarillo-Luna, R.A., Campos-Rodriguez, R., and Tsutsumi, V. 2002.Entamoeba histolytica: immunohistochemical study of hepaticamoebiasis in mouse. Neutrophils and nitric oxide as possiblefactors of resistance. Exp. Parasitol. 101: 40–56. doi:10.1016/S0014-4894(02)00021-8. PMID:12243737.
Jarumilinta, R. 1966. A simple method of inducing amoebic liverabscess in hamsters. Ann. Trop. Med. Parasitol. 60: 139–145.PMID:4289920.
Jessurun, J., Barron-Rodriguez, L.P., Fernandez-Tinoco, G., andHernandez-Avila, M. 1992. The prevalence of invasive amebia-sis is not increased in patients with AIDS. AIDS, 6: 307–309.doi:10.1097/00002030-199203000-00007. PMID:1567575.
Katayama, I., Parker, D., and Turk, J.L. 1982. In vivo macrophagesuppression of delayed hypersensitivity in the guinea pig. Immu-nology, 47: 709–716. PMID:6216203.
Kaufmann, S.H. 2001. How can immunology contribute to the con-trol of tuberculosis? Nat. Rev. Immunol. 1: 20–30. doi:10.1038/35095558. PMID:11905811.
Klebanoff, S.J. 2005. Myeloperoxidase: friend and foe. J. Leukoc.Biol. 77: 598–625. doi:10.1189/jlb.1204697. PMID:15689384.
Lin, J.Y., and Chadee, K. 1992. Macrophage cytotoxicity againstEntamoeba histolytica trophozoites is mediated by nitric oxidefrom L-arginine. J. Immunol. 148: 3999–4005.PMID:1318338.
Lin, J.Y., Seguin, R., Keller, K., and Chadee, K. 1994. Tumor ne-crosis factor a augments nitric oxide-dependent macrophage cy-totoxicity against Entamoeba histolytica by enhanced expressionof the nitric oxide synthase gene. Infect. Immun. 62: 1534–1541.PMID:7513301.
Matsuo, S., Nakagawara, A., Ikeda, K., Mitsuyama, M., andNomoto, K. 1985. Enhanced release of reactive oxygen inter-mediates by immunologically activated rat Kupffer cells. Clin.Exp. Immunol. 59: 203–209. PMID:2982526.
McCuskey, R.S., Urbaschek, R., McCuskey, P.A., and Urbaschek,B. 1983. In vivo microscopic observations of the responses ofKupffer cells and the hepatic microcirculation to Mycobacteriumbovis BCG alone and in combination with endotoxin. Infect. Im-mun. 42: 362–367. PMID:6352499.
Murray, H.W., Aley, S.B., and Scott, W.A. 1981. Susceptibility ofEntamoeba histolytica to oxygen intermediates. Mol. Biochem.Parasitol. 3: 381–391. doi:10.1016/0166-6851(81)90038-4.PMID:6272108.
Oppenheim, J.J., Biragyn, A., Kwak, L.W., and Yang, D. 2003.Roles of antimicrobial peptides such as defensins in innate andadaptive immunity. Ann. Rheum. Dis. 62(Suppl 2): ii17–ii21.doi:10.1136/ard.62.suppl_2.ii17. PMID:14532141.
Ortiz-Ortiz, L., Ruiz, B., and Gonzalez, A. 1990. Cellular immuneresponses. In Amebiasis: infection and disease by Entamoebahistolytica. Edited by R.R. Kretschmer. CRC Press, Boca Raton,Florida. pp. 77–103.
Pacheco-Yepez, J., Campos-Rodriguez, R., Shibayama, M.,Ventura-Juarez, J., Serrano-Luna, J., and Tsutsumi, V. 2001. En-tamoeba histolytica: production of nitric oxide and in situ activ-ity of NADPH diaphorase in amebic liver abscess of hamsters.Parasitol. Res. 87: 49–56. doi:10.1007/s004360000287. PMID:11199849.
Perez-Tamayo, R., Martinez-Villegas, J.E., and Perez-Montfort, R.1986. Effect of cellular immunity on the interaction between theperitoneal cell and the ameba in vitro. Arch. Invest. Med.(Mex.), 17(Suppl 1): 259–267. PMID:2884966.
Petri, W.A., Jr., and Singh, U. 1999. Diagnosis and management ofamebiasis. Clin. Infect. Dis. 29: 1117–1125. doi:10.1086/313493. PMID:10524950.
Ramirez-Emiliano, J., Gonzalez-Hernandez, A., and Arias-Negrete,S. 2005. Expression of inducible nitric oxide synthase mRNAand nitric oxide production during the development of liverabscess in hamster inoculated with Entamoeba histolytica. Curr.Microbiol. 50: 299–308. doi:10.1007/s00284-005-4503-z.PMID:15968502.
Ravdin, J.I., Murphy, C.F., Salata, R.A., Guerrant, R.L., andHewlett, E.L. 1985. N-Acetyl-D-galactosamine-inhibitableadherence lectin of Entamoeba histolytica. I. Partial purificationand relation to amoebic virulence in vitro. J. Infect. Dis. 151:804–815. PMID:2859338.
Reed, S.L., Wessel, D.W., and Davis, C.E. 1991. Entamoeba histo-lytica infection and AIDS. Am. J. Med. 90: 269–271.PMID:1996598.
Rogers, H.W., Sheehan, K.C., Brunt, L.M., Dower, S.K., Unanue,E.R., and Schreiber, R.D. 1992. Interleukin 1 participates in thedevelopment of anti-Listeria responses in normal and SCIDmice. Proc. Natl. Acad. Sci. U.S.A. 89: 1011–1015. doi:10.1073/pnas.89.3.1011. PMID:1531270.
Salata, R.A., Pearson, R.R., and Ravdin, J.I. 1985. Interaction ofhuman leukocytes and Entamoeba histolytica. Killing of virulent
Shibayama et al. 1041
# 2008 NRC Canada

amebae by the activated macrophage. J. Clin. Invest. 76(2):491–499. doi:10.1172/JCI111998. PMID:2863284.
Seydel, K.B., Zhang, T., and Stanley, S.L., Jr. 1997. Neutrophilsplay a critical role in early resistance to amebic liver abscessesin severe combined immunodeficient mice. Infect. Immun. 65:3951–3953. PMID:9284178.
Seydel, K.B., Smith, S.J., and Stanley, S.L., Jr. 2000. Innate immu-nity to amebic liver abscess is dependent on g interferon andnitric oxide in a murine model of disease. Infect. Immun. 68:400–402. PMID:10603416.
Shibayama, M., Campos-Rodriguez, R., Ramirez-Rosales, A.,Flores-Romo, L., Espinosa-Cantellano, M., Martinez-Palomo,A., and Tsutsumi, V. 1998. Entamoeba histolytica: liver invasionand abscess production by intraperitoneal inoculation oftrophozoites in hamsters, Mesocricetus auratus. Exp. Parasitol.88: 20–27. doi:10.1006/expr.1998.4218. PMID:9501845.
Shibayama, M., Campos-Rodriguez, R., Ramirez, C., Pacheco-Yepez, J., and Tsutsumi, V. 2000. Studies on the natural immu-nity in hamsters using the intraperitoneal model of amebic liverabscess. Arch. Med. Res. 31: S78–S80. doi:10.1016/S0188-4409(00)00187-9. PMID:11070232.
Stanley, S.L., Jr. 2006. Vaccines for amoebiasis: barriers and op-portunities. Parasitology, 133(Suppl): S81–S86. doi:10.1017/S003118200600182X. PMID:17274850.
Thomas, E.L., and Fishman, M. 1986. Oxidation of chloride and thio-cyanate by isolated leukocytes. J. Biol. Chem. 261: 9694–9702.PMID:3015901.
Tsutsumi, V., and Shibayama, M. 2006. Experimental amebiasis: a
selected review of some in vivo models. Arch. Med. Res. 37:210–220. doi:10.1016/j.arcmed.2005.09.011. PMID:16380321.
Tsutsumi, V., Mena-Lopez, R., Anaya-Velazquez, F., and Martinez-Palomo, A. 1984. Cellular bases of experimental amebic liverabscess formation. Am. J. Pathol. 117: 81–91. PMID:6385728.
van Crevel, R., Ottenhoff, T.H., and van der Meer, J.W. 2002. In-nate immunity to Mycobacterium tuberculosis. Clin. Microbiol.Rev. 15: 294–309. doi:10.1128/CMR.15.2.294-309.2002. PMID:11932234.
Velazquez, C., Tsutsumi, V., Shibayama-Salas, M., Aguirre-Garcia,J., and Calderon, J. 1997. Role of the neutrophil in the patho-genesis of the amebic liver lesion in mice. Arch. Med. Res.28(Spec No): 230–232. PMID:9033082.
Wang, W., and Chadee, K. 1995. Entamoeba histolytica suppressesg interferon-induced macrophage class II major histocompatibil-ity complex Ia molecule and I-A b mRNA expression by aprostaglandin E2-dependent mechanism. Infect. Immun. 63:1089–1094. PMID:7868232.
Wang, J.-G., Mahmud, S.A., Thompson, J.A., Geng, J.-G., Key,N.S., and Slungaard, A. 2006. The principal eosinophil peroxi-dase product, HOSCN, is a uniquely potent phagocyte oxidantinducer of endothelial cell tissue factor activity: a potential me-chanism for thrombosis in eosinophilic inflammatory states.Blood, 107: 558–565. doi:10.1182/blood-2005-05-2152. PMID:16166591.
Xia, Y., and Zweier, J.L. 1997. Measurement of myeloperoxidasein leukocyte-containing tissues. Anal. Biochem. 245: 93–96.doi:10.1006/abio.1996.9940. PMID:9025973.
1042 Can. J. Microbiol. Vol. 54, 2008
# 2008 NRC Canada
Copyright © 2022 FDOKUMEN




















