
In vitro and in vivo interaction of Entamoeba histolytica Gal/GalNAc lectin with various target...
13
Parasitology International 53 (2004) 35–47 1383-5769/04/$ - see front matter 2003 Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.parint.2003.10.005 In vitro and in vivo interaction of Entamoeba histolytica Gal y GalNAc lectin with various target cells: an immunocytochemical analysis Judith Pacheco , Mineko Shibayama , Rafael Campos , David L. Beck , Erick Houpt , a,b a c d d William A. Petri, Jr. , Vıctor Tsutsumi * d a, ´ Department of Experimental Pathology, Center for Research and Advanced Studies, National Polytechnic Institute, a Cinvestav-IPN, Av. IPN 2508, Col. San Pedro Zacatenco, Mexico, D.F. 07360, Mexico Mexican Faculty of Medicine, Laboratory of Electron Microscopy, La Salle University, Fuentes 17, Tlalpan, Mexico, D.F. 14000, b Mexico Department of Biochemistry, School of Medicine, National Polytechnic Institute, Plan de San Luis y Dıaz Miron synum., Mexico, c ´ ´ ´ D.F. 11340, Mexico Division of Infectious Diseases and International Health, University of Virginia Health System, Charlottesville, VA 22908-1340, d USA Received 1 July 2003; accepted 1 October 2003 Abstract The GalyGalNAc lectin of Entamoeba histolytica trophozoites plays an important role in adhesion. The distribution and final destiny of the lectin during the interaction with host cells are poorly understood. Using monoclonal and polyclonal antibodies against the lectin we studied by immunocytochemistry the in vitro and in vivo interaction of E. histolytica trophozoites with human and hamster hepatocytes. We also analyzed the presence and distribution of the lectin in a mouse model of intestinal amoebiasis. In all cases, trophozoites were highly labeled by anti-lectin antibodies. Cultured human and hamster hepatocytes in contact with, or localized at the vicinity of parasites were also labeled by anti-lectin antibodies. Most of the labeled hepatocytes showed variable degrees of cell damage. Hepatocytes distantly localized from the parasites were also stained with the anti-lectin antibodies. Immunolabeling of tissue sections from different stages of the development of experimental amoebic liver abscess in hamsters showed inflammatory foci containing lectin-labeled trophozoites, hepatocytes, and sinusoidal and inflammatory cells. Lectin- containing hepatocytes had vacuolated cytoplasm with some nuclei with a condensed appearance. Damaged intestinal epithelium also was labeled with anti-lectin antibodies in a mouse model of intestinal amoebiasis. Electron microscopy of axenically cultured trophozoites using gold-labeled monoclonal and polyclonal anti-lectin antibody showed that plasma membrane, vacuole membranes and areas of cell cytosol were labeled. Higher deposits of gold particles in plasma membrane suggestive of cell secretion were observed. Our results demonstrated that GalyGalNAc lectin was *Corresponding author. Tel.:q52-5747-7138; fax: q52-5747-7107. E-mail address: [email protected] (V. Tsutsumi).
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of In vitro and in vivo interaction of Entamoeba histolytica Gal/GalNAc lectin with various target...

Parasitology International 53(2004) 35–47
1383-5769/04/$ - see front matter� 2003 Elsevier Ireland Ltd. All rights reserved.doi:10.1016/j.parint.2003.10.005
In vitro and in vivo interaction ofEntamoeba histolytica GalyGalNAc lectin with various target cells: an immunocytochemical
analysis
Judith Pacheco , Mineko Shibayama , Rafael Campos , David L. Beck , Erick Houpt ,a,b a c d d
William A. Petri, Jr. , Vıctor Tsutsumi *d a,´
Department of Experimental Pathology, Center for Research and Advanced Studies, National Polytechnic Institute,a
Cinvestav-IPN, Av. IPN 2508, Col. San Pedro Zacatenco, Mexico, D.F. 07360, MexicoMexican Faculty of Medicine, Laboratory of Electron Microscopy, La Salle University, Fuentes 17, Tlalpan, Mexico, D.F. 14000,b
MexicoDepartment of Biochemistry, School of Medicine, National Polytechnic Institute, Plan de San Luis y Dıaz Miron synum., Mexico,c ´ ´ ´
D.F. 11340, MexicoDivision of Infectious Diseases and International Health, University of Virginia Health System, Charlottesville, VA 22908-1340,d
USA
Received 1 July 2003; accepted 1 October 2003
Abstract
The GalyGalNAc lectin ofEntamoeba histolytica trophozoites plays an important role in adhesion. The distributionand final destiny of the lectin during the interaction with host cells are poorly understood. Using monoclonal andpolyclonal antibodies against the lectin we studied by immunocytochemistry the in vitro and in vivo interaction ofE.histolytica trophozoites with human and hamster hepatocytes. We also analyzed the presence and distribution of thelectin in a mouse model of intestinal amoebiasis. In all cases, trophozoites were highly labeled by anti-lectinantibodies. Cultured human and hamster hepatocytes in contact with, or localized at the vicinity of parasites werealso labeled by anti-lectin antibodies. Most of the labeled hepatocytes showed variable degrees of cell damage.Hepatocytes distantly localized from the parasites were also stained with the anti-lectin antibodies. Immunolabelingof tissue sections from different stages of the development of experimental amoebic liver abscess in hamsters showedinflammatory foci containing lectin-labeled trophozoites, hepatocytes, and sinusoidal and inflammatory cells. Lectin-containing hepatocytes had vacuolated cytoplasm with some nuclei with a condensed appearance. Damaged intestinalepithelium also was labeled with anti-lectin antibodies in a mouse model of intestinal amoebiasis. Electron microscopyof axenically cultured trophozoites using gold-labeled monoclonal and polyclonal anti-lectin antibody showed thatplasma membrane, vacuole membranes and areas of cell cytosol were labeled. Higher deposits of gold particles inplasma membrane suggestive of cell secretion were observed. Our results demonstrated that GalyGalNAc lectin was
*Corresponding author. Tel.:q52-5747-7138; fax:q52-5747-7107.E-mail address: [email protected](V. Tsutsumi).

36 J. Pacheco et al. / Parasitology International 53 (2004) 35–47
bound and captured by different target cells, and that host cells containing the lectin showed signs of cell damage.The contribution of lectin transfer to host cells in adherence and cell injury remains to be determined.� 2003 Elsevier Ireland Ltd. All rights reserved.
Keywords: Entamoeba histolytica; GalyGalNAc lectin; Cultured hepatocyte; Amoebic liver abscess; Intestinal ulcer;Immunohistochemistry
1. Introduction
Amoebiasis is an important worldwide diseasecaused by infection with the enteric protozoanEntamoeba histolytica. This parasite invades anddamages tissues through a sequence of events withthe participation of multiple factors related to boththe parasite and the host. Three basic steps havebeen described in the in vitro interaction of amoe-bae with different target cells: adherence, cytolysis,and phagocytosisw1–3x. Several molecules havebeen shown to be involved during the progressionof these events, and include lectins, pore formingpeptides, and proteases. It has been suggested thatcell adhesion is an initial requirement in theprocess of cell destruction by the amoebae.One of the best-characterized adherence mole-
cules on the surface ofE. histolytica trophozoitesis the GalyGalNAc inhibitable lectin. The lectin iscomposed of disulfide bonded 170 kDa heavy and35y31 kDa light subunits and a non-covalentlyassociated 150 kDa intermediate subunit. The ami-no acid sequence of the heavy subunit consists ofa putative extracellular domain divided into threeregions based on their composition, and a carboxy-terminal cytoplasmic domain formed by 41 aminoacids w4,5x. The heavy subunit is mainly respon-sible for mediating amoebic adherencew6x, and ithas been shown to participate in the in vitroadherence ofE. histolytica trophozoites to a varietyof target cellsw3,7–11x.It has previously been reported that amoebic
antigens are transferred to host cells. Using amonoclonal antibody(CD6) against an amoebasurface antigen, Leroy et al.w12x demonstrated acontact-dependent transfer of the CD6 antigen tothe lateral epithelial cell surface of cultured enter-ocytes after interaction withE. histolytica tropho-zoites. In other cell types such as fibroblasts testedin the same study, no immunostaining was
observed with the CD6 antibody. The nature ofthe CD6 antigen, initially reported to be the GalyGalNAc lectin, has yet to be conclusively identi-fied. No staining of epithelial cells by the amoebaewas observed by Leroy et al.w12x when anti-GalyGalNAc lectin antibodies were used. In contrast toprevious work, here we performed immunocyto-chemical studies using monoclonal or polyclonalantibodies specifically directed to the GalyGalNAclectin during the amoeba interaction with hepato-cytes, and in vivo during the formation of amoebicliver abscess(ALA ) and amoebic intestinal ulcers.Ultrastructural immunogold labeling of culturedE.histolytica trophozoites was also performed todetermine the subcellular localization of the lectinin the parasite.
2. Material and methods
2.1. Amoeba culture
Trophozoites ofE. histolytica strain HMI:IMSSwere grown axenically at 35.68C in TYI-S-33medium w13x supplemented with 15% bovineserum. The trophozoites were harvested at the endof the logarithmic growth phase(72 h) by chillingat 4 8C. They were concentrated by centrifugationfor 5 min at 300=g and used immediately.
2.2. Primary human hepatocyte culture
Normal human cryopreserved hepatocytes(hNheps) were obtained from Bio-Whittaker-Clonet-ics (Cambrex Co. Walkersville, MD). Primaryhuman hepatocytes were maintained in hepatocyteculture medium. The number of viable hepatocyteswas determined by Trypan blue exclusion, and theviability was 25%. The seeding density was150 000 viable cellsy3.24 cm . At light microsco-2
py, hepatic cells were seen randomly adhered on

37J. Pacheco et al. / Parasitology International 53 (2004) 35–47
the cover slips without forming uniform mono-layers.
2.3. Primary hamster hepatocytes culture
Livers were perfused through the portal veinand dissected from young adult male golden ham-sters(Mesocricetus auratus) weighing 90 g. Hepa-tocytes were isolated and cultured using a modifiedmethod of collagenase perfusion reported by Berryand Friendw14x. Cells were incubated at 36.88Cin 9% CO , and nonattached cells were discarded2
after 60 min of incubation. The number of viablehepatocytes as determined by Trypan blue exclu-sion was more than 90%. Cultured hamster hepa-tocytes formed uniform monolayers at 24 h ofincubation.
2.4. Interaction of E. histolytica trophozoites withcultured human hepatocytes
Human hepatocytes were cultured for 24 h oncover slips previously coated with a thin layer ofcollagen type I from rat tendon(19.4 mgy3.24cm cover slide area). Suspensions of freshly2
harvestedE. histolytica trophozoites were placedover the randomly distributed hepatocytes(notforming a confluent monolayer) at a ratio of 1y10(trophozoitesyhepatocytes), using a mixture ofTYI-S-33 and hepatocytes culture medium. Inter-actions were carried out for 15 and 30 min at 36.88C. To test the specificity of GalyGalNAc lectinmediated adhesion, interactions were also per-formed in the presence of 50 mMN-acetyl-D-galactosamine(GalNAc) to inhibit adhesion viathe lectin. Samples were fixed with 1.5% parafor-maldehyde before processing for immunocyto-chemistry.
2.5. Interaction of E. histolytica trophozoites withcultured hamster hepatocytes
The hamster hepatocytes were cultured for 24 hat 36.8 8C in 9% CO . Suspensions of freshly2
harvestedE. histolytica trophozoites were placedover the monolayer of cultured hepatocytes at aratio of 1y8 (trophozoitesyhepatocytes). Interac-tions were carried out for 15, 30 and 60 min at
36.8 8C. Inhibition of the GalyGalNAc lectinmediated adhesion was done as mentioned above.At the end of each period, samples were fixedwith 1.5% paraformaldehyde and processed forimmunocytochemistry. All interactions were donein duplicate and compared to interactions withsugars and controls not added with trophozoites.
2.6. Production of ALA
Two-month-old male adult golden hamsters(Mesocricetus auratus) weighing approximately100 g were used. All procedures and handling ofexperimental animals were performed according tothe institutional guidelines. Hamsters were intra-portally inoculated with 0.1 ml culture mediumcontaining 5=10 E. histolytica trophozoitesw15x.5
Groups of five animals were killed with an over-dose of anesthetic(10 mgy100 g body weight) at6, 12, 24 and 48 h post-inoculation. Lesionsassociated with normal hepatic tissue were fixed,embedded in paraffin, and sections were stainedwith hematoxilin and eosin. Representative areasof samples from each time point were selected andprocessed for immunohistochemistry.
2.7. Production of amoebic intestinal ulcer in mice
Five-week-old C3HyHej strain mice weighingapproximately 20 g were used. Mice were chal-lenged by inoculation of 2=10 xenically grown7
HMI:IMSS trophozoites that had been passagedthrough hamster liver. Mice were anaesthetizedwith ketamineyxylazine and a medial abdominalincision was performed. After identification of thececum, trophozoites were injected at the apex ofthe cecum using a tuberculin syringe. Mice werekilled 4 weeks post-inoculation. Approximately50% of the mice become infected, uninfected micewere used as controls. The cecum was removedand fixed with Bouin’s solution. Both infected anduninfected tissue was embedded in the same par-affin block.
2.8. Immunocytochemistry and immunohisto-chemistry
Nonspecific binding sites were blocked with10% fetal bovine serum in phosphate buffered

38 J. Pacheco et al. / Parasitology International 53 (2004) 35–47
Fig. 1. Immunocytochemistry of the in vitro interaction ofE. histolytica with hepatocytes using monoclonal anti-GalyGalNAc lectinantibody. Panels A–D human hepatocytes; E–H hepatocytes.(A) Fifteen minutes of interaction between human hepatocytes andE.histolytica trophozoites. Highly positive trophozoites(t) are seen contacting hepatocytes(h). Lectin positive hepatic cells are stainedas pink to brownish color and contain multiple perinuclear vesicles. An unlabeled hepatocyte(uh) is also observed, 75=. (B)Similar interaction time as before. A strongly labeled trophozoite(t) is seen closely contacting a hepatocyte(h). Peroxidase stainin the hepatocyte is more intense at perinuclear zone(arrows) and the hepatocyte contains multiple clear vacuoles, 220=. (C)Thirty minutes of interaction. Three severely damaged human hepatocytes(arrows) labeled with the GalyGalNAc lectin and con-taining irregular and condensed nuclei are seen closely contacting a trophozoite(t), 300=. (D) Effect of galactose on humanhepatocytes interacted withE. histolytica trophozoites. An amoeba is seen strongly labeled with the GalyGalNAc lectin; however,hepatocytes are only lightly marked, 75=. (E) Control. Non-infected human hepatocytes incubated with the anti-GalyGalNAc mAb.Non-confluent unlabeled normal hepatocytes are observed. Some cells show perinuclear vacuoles, 180=. (F) Fifteen minutes ofinteraction ofE. histolytica trophozoites with cultured hamster hepatocytes. A trophozoite(t) is strongly labeled with the lectin asare some of the neighbouring hepatic cells(h). These hepatocytes show signs of cell damage with loss of normal cell architecture,multiple vesicles, and damaged nuclei, 180=. (G) Similar interaction time and antibody as before. Some lectin labeled(h) andunlabeled hepatocytes(uh) are seen in contact with a trophozoite(t). Damaged hepatocytes are seen with the peroxidase label.Other undamaged hepatocytes are also shown, 180=. (H) Thirty minutes of interaction. Large lytic areas(ly) are seen at the rightside. Trophozoites(t) are strongly labeled and pleomorphic. Small amoeba at the upper part are closely contacting labeled anddamaged hepatocytes(arrow-heads) containing multiple vesicles. Trophozoites at the lower part of photo are associated with partiallydamaged non-labeled hepatocytes(arrows). Hepatic cells with dense nuclei are also observed, 180=.
saline (PBS) for 1 h at room temperature(RT).Detection of the amoebic GalyGalNAc lectin wascarried out using H85 mouse monoclonal antibody(IgG2b) against the 170 kDa heavy subunit of theGalyGalNAc lectin diluted 1:50. The backgroundstaining was eliminated after determining the opti-mal dilution of the primary and secondary antibod-ies, and also by using a non-immune serum fromthe same animal species in which the secondantibody was produced. Control samples wereincubated with an irrelevant primary mouse anti-CD8 antibody (IgG1) (Zymed, San Francisco,CA) or with the secondary antibody alone. Sam-ples were incubated overnight at 48C, washedwith PBS-Tween(PBS-T), and incubated with aperoxidase-conjugated goat anti-mouse antibody(Zymed) for 1 h and diluted 1:500 in PBS-Tcontaining 3% fetal bovine serum. The reactionwas developed by adding diaminobenzidine(DAB,Pierce, Rockford, IL) and H O as a substrate for2 2
3–10 min. Finally, the samples were counters-tained with hematoxilin. Immunostained interac-tions were examined by light microscopyw16x.For immunohistochemistry of hamster liver tis-
sues or mice intestinal ulcers, 6–7mm-thick par-affin sections were obtained and mounted on glassslides. After dewaxing with xylene and rehydra-tion, endogenous peroxidase activity was blockedby incubation with 1% H O in methanol for 1 h,2 2
and washing with PBS. Nonspecific reactions were
blocked similarly as in cultured hepatocytes. Inhepatic samples, slides were incubated overnightat 4 8C with a rabbit polyclonal antibody(Shiro)against the 170 kDa heavy subunit of the GalyGalNAc lectin, diluted 1:100 in PBS-T; a rabbitpreimmune serum or secondary antibody alonewere used as controls. For intestinal samples, slideswere incubated either with the anti-lectin polyclon-al antisera diluted 1:500 or with the anti-cytoplas-mic domain-specific antisera(gift of NancyGuillen) diluted 1:10 in blocking buffer. Slideswere immunostained using Elite Rabbit IgG Vec-tastain ABC kit from Vector Labs according tomanufactures directions. Hepatic samples wereincubated with a peroxidase-labeled goat anti-rabbit IgG antibody(Boehringer Mannheim, Indi-anapolis, IN) for 1 h and developed with DABw17x. Sections were finally stained with hematoxi-lin, dehydrated with alcohol, mounted with syn-thetic resin and examined with a light microscope.For intestinal samples, both infected and uninfect-ed ceca were processed on the same slide to ensureequivalent staining and development.
2.9. Immunoelectron microscopy of E. histolyticatrophozoites
Cultured trophozoites were centrifuged for 10min at 800=g. Cell pellets were fixed with a

39J. Pacheco et al. / Parasitology International 53 (2004) 35–47
Fig. 1.

40 J. Pacheco et al. / Parasitology International 53 (2004) 35–47
mixture of 1.5% paraformaldehydey0.5% glutar-aldehyde in 0.1 M cacodylate buffer, for 60 minat RT. Samples were dehydrated with increasingconcentrations of ethanol aty20 8C, and pre-embedded in 1:1 mixture of LR-WHITE resin(Polysciences, Warrington, PA) and absolute eth-anol(vyv). The next day, the mixture was replacedwith resinyethanol at 2:1 ratio, incubated 1 h aty20 8C, and then replaced twice with pure resinfor 1 h. Finally, the samples were embedded inresinqaccelerator(1 mly10 ml) and incubated for5 days at 508C. Sections 90 nm thick weremounted in 300 mesh nickel grids and blockedwith 10% fetal bovine serum in 0.005% PBS-T, 1h at RT. Samples were incubated overnight at 48C in humid chamber with monoclonal H85(mouse) and polyclonal(Shiro) rabbit anti-GalyGalNAc lectin antibody diluted 1:25 and 1:50,respectively, in PBS-T containing 3% fetal bovineserum. Removal of non-specific background andstaining of control sections were conducted aspreviously described above. A secondary goat anti-mouse and anti-rabbit antibody coupled to 20 nmgold particles diluted 1:40 in PBS-T with fetalbovine serum was added and samples were incu-bated for 1 h at RT. After washing, grids werecontrasted with uranyl acetate and lead citratew18x.Specimens were observed with a Zeiss EM-10transmission electron microscope.
3. Results
3.1. Immunodetection of the GalyGalNAc lectinduring the in vitro interactions of E. histolyticatrophozoites with cultured human hepatocytes
In 15 min of interaction, the monoclonal anti-body identified the GalyGalNAc lectin on theamoebae as well as on the contacting hepatocytes(Fig. 1A). Some non-contacting hepatocytes, butlocalized near the amoebae, were also labeled.Trophozoites presented a strong peroxidase label-ing shown as dark brownish color, whereas positivehepatocytes were observed with pink to lightbrown deposits in the cytoplasm with a higherdensity of label in the cytoplasmic perinuclearareas of hepatocytes(Fig. 1B). Large and clearcytoplasmic vacuoles suggestive of early degener-
ative hepatic cell changes were observed in manycells.In 30 min of interaction, abundant labeled hepa-
tocytes were randomly distributed on the slides.Most of the hepatic cells showed different degreesof damage, including the presence of multiple andlarge vacuoles in the cytoplasm, loss of normalcell architecture with small dense nuclei. Irregular-ly rounded or elongated small cells with densecytoplasmic label and condensed nuclei weresometimes seen attached to the surface ofE.histolytica trophozoites(Fig. 1C). Transfer of theGalyGalNAc lectin to host cells was blocked whenthe lectin was inhibited with excess GalNAc; therewas a decrease in both the number of trophozoitesattached to the human primary culture and in thelectin staining, which was mainly observed in thetrophozoites and significantly reduced in the hostcells(Fig. 1D). The control samples of hepatocytesincubated in the absence of parasites showed nolabeling with the anti-lectin mAb and the architec-ture of the human hepatocytes showed in generala normal appearance, except for the presence ofsome cells with perinuclear vacuoles(Fig. 1E).
3.2. Immunodetection of the GalyGalNAc lectinduring the in vitro interactions of E. histolyticatrophozoites with cultured hamster hepatocytes
In 15 min of interaction and during all subse-quent periods of interaction, trophozoites ofE.histolytica were strongly labeled with the mono-clonal anti-lectin antibody. Hepatocytes closelycontacting the amoebae were labeled and showedsigns of cell damage characterized by loss of celllimits, multiple cytoplasmic vacuolization andaltered nuclei(Fig. 1F and G). Other trophozoiteswere seen surrounded by partially labeled or unla-beled hepatocytes. Undamaged hepatocytes withonly scarce vacuolization were also observed atthe vicinity of the parasite(Fig. 1G). The lectinwas mainly distributed in the cytoplasmic region.In 30 and 60 min of interaction, lysed hepato-
cytes increased progressively leaving large blankareas on the cell monolayer(Fig. 1H). Stronglylabeled trophozoites were seen randomly distrib-uted throughout the monolayer, however, they weremore commonly localized at the edges of the

41J. Pacheco et al. / Parasitology International 53 (2004) 35–47
necrotic zones. Hepatocytes contacting amoebaewere either labeled or unlabeled with the lectin,and in both cases showed signs of cell damage.However, labeled hepatocytes showed more severesigns of damage characterized by loss of cell limitsand abundant microvesicles(Fig. 1H). In interac-tions performed in presence of excess GalNAc,the number of attached trophozoites was reducedapproximately 50%. Staining of the hepatocytesby anti-lectin antibodies was also diminished. Con-trol hepatocytes incubated in the absence of para-sites were not lectin-labeled or damaged(data notshown).
3.3. Presence of GalyGalNAc in the hepatic paren-chymal and inflammatory cells of the ALA
In 6 h post-intraportal inoculation and as report-ed earlierw15x, the parasites were localized in thehepatic parenchyma surrounded by acute inflam-matory infiltrates (Fig. 2A). Staining of theabscess with anti-lectin antibodies identified lectinon the surface of trophozoites and also in inflam-matory and sinusoidal cells and in the cytoplasmof neighbouring hepatocytes. No reactivity wasobserved in similarly infected liver samples incu-bated with a rabbit preimmune serum, or with theperoxidase-labeled secondary antibody alone(Fig.2A*).In 12 h the lesions increased in size, with a
larger and irregular inflammatory infiltrate associ-ated with strongly labeled trophozoites(Fig. 2B).Sinusoidal and parenchymal cells also revealed thepresence of GalyGalNAc lectin. At this stage,many hepatocytes labeled with peroxidase andlocalized close to the infiltrates showed differentdegrees of damage. Cells with condensed nucleiwere also observed. At 24 h the damage to theliver parenchyma was more evident with thenecrotic areas increased in size, hepatocytes show-ing signs of morphological damage, and abundantlysed leukocytes(Fig. 2C). The GalyGalNAc lec-tin was identified in the amoebae, inflammatorycells, and cytosol of hepatocytes bordering theinflammatory infiltrates. A fine deposit of peroxi-dase reaction was also present in the necroticcenter, suggesting degradation of some cells thatbound the lectin at earlier time-points. Trophozo-
ites, partially damaged and fused with each other,were seen in the inflammatory infiltrate.Finally in 48 h, the liver lesions showed typical
granulomatous lesions formed by a large chronicinflammatory infiltrate and necrosis outlined byepithelioid cells(Fig. 2D). The GalyGalNAc lectinwas still localized on the amoebae, as well as onacute and chronic inflammatory cells, epithelioidcells, and sinusoidal and parenchymal cells. Nolectin labeling was observed when infected liversamples were first incubated with a non-relevantantibody or when the secondary peroxidase-labeledantibody was used alone(Fig. 2A*). Normaluninfected liver samples incubated with anti-GalyGalNAc antibody were also unlabeled(data notshown).
3.4. Detection of the lectin and the cytoplasmicdomain in the mouse intestinal model ofamoebiasis
Infected and uninfected mouse ceca were stainedwith either polyclonal anti-whole lectin antisera,or polyclonal anti-lectin cytoplasmic domain anti-sera. As seen in Fig. 3 anti-whole lectin antiserastrongly stained the trophozoites, as well as thehost epithelium and necrotic tissue. A similarthough lighter staining was seen with anti-cyto-plasmic domain specific antisera. No staining wasseen in specimens of uninfected mouse cecum, orof infected cecum stained with secondary antibodyalone. Staining of infected mouse cecum onlyoccurred where the trophozoites were present, andin the absence of trophozoites no staining wasseen(data not shown).
3.5. Immunoelectronmicroscopic detection of theGalyGalNAc lectin in cultured E. histolyticatrophozoites
Membranes of large and medium sized cyto-plasmic vacuoles, as well as plasma membranesof E. histolytica trophozoites were highly labeledwith the polyclonal gold-labeled anti GalyGalNAclectin (Fig. 4A). Interestingly, some particularzones of the plasma membrane associated withmedium sized cytoplasmic vacuoles showed ahigher density of gold-labeled anti-lectin particles

42 J. Pacheco et al. / Parasitology International 53 (2004) 35–47
Fig. 2. Immunohistochemistry of the in vivo interaction ofE. histolytica with hamster liver using polyclonal anti-GalyGalNAc lectinantibodies. Hamster livers at(A) 6 h, (B) 12 h, (C) 24 h and(D) 48 h post-inoculation withE. histolytica trophozoites.(A) At 6h a focus of acute inflammatory infiltrate with a centrally located trophozoite is observed(t). Strong peroxidase labeling is observedin the trophozoite, inflammatory cells(i) and nearby hepatocytes(arrows), 20=. (A*) Control immunohistochemistry at 6 h post-inoculation demonstrating lack of reactivity with the peroxidase-labeled secondary antibody, 20=. (B) At 12 h an irregular andlarger inflammatory focus is seen with positive reaction in trophozoites(t), inflammatory cells(i), circumscribed hepatocytes(arrows) and some sinusoidal cells(s). Hepatocytes associated with the infiltrate are partially damaged, 20=. (C) At 24 h anexternal area of inflammatory reaction is seen. Necrotic tissue(n) associated with multiple trophozoites(t) is seen. Some trophozoitesare damaged and fused with each other. Inflammatory cells showed signs of damage. Surrounding hepatocytes(arrows) and sinu-soidal cells(s) are also labeled 40=. (D) At 48 h a typical granulomatous lesion is seen. Trophozoites(t), associated inflammatorycells (i), epithelioid cells(e) and nearby hepatocytes(arrows) are all labeled with the lectin. 40=.
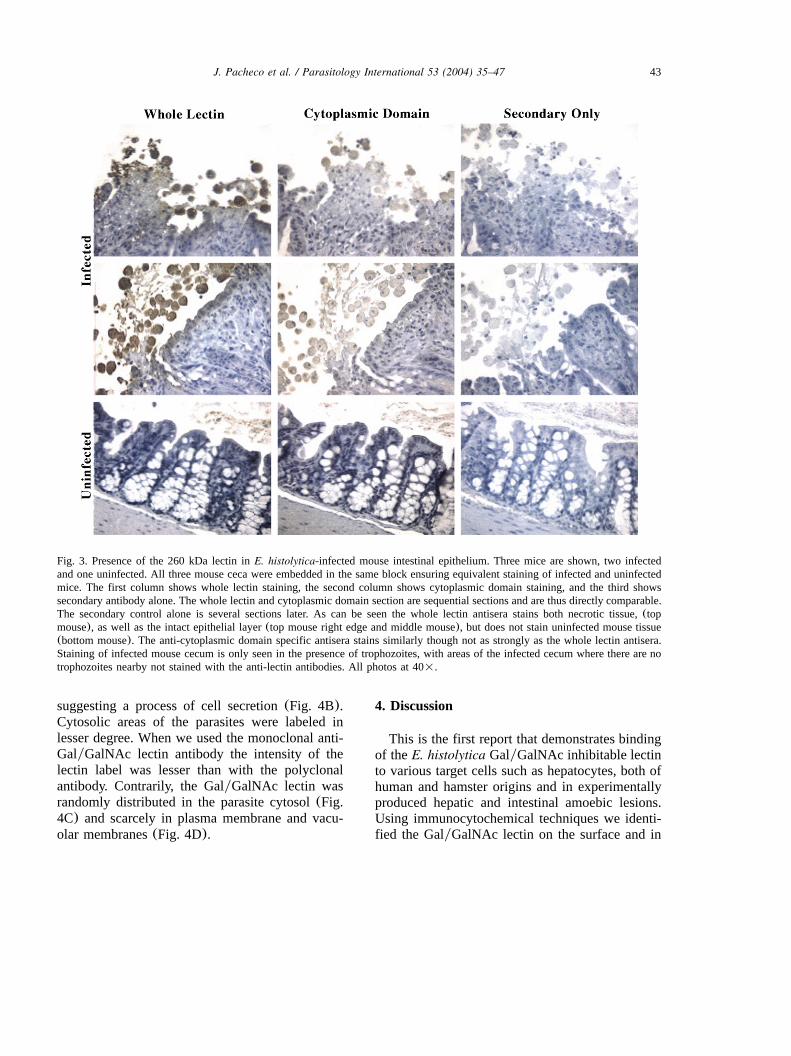
43J. Pacheco et al. / Parasitology International 53 (2004) 35–47
Fig. 3. Presence of the 260 kDa lectin inE. histolytica-infected mouse intestinal epithelium. Three mice are shown, two infectedand one uninfected. All three mouse ceca were embedded in the same block ensuring equivalent staining of infected and uninfectedmice. The first column shows whole lectin staining, the second column shows cytoplasmic domain staining, and the third showssecondary antibody alone. The whole lectin and cytoplasmic domain section are sequential sections and are thus directly comparable.The secondary control alone is several sections later. As can be seen the whole lectin antisera stains both necrotic tissue,(topmouse), as well as the intact epithelial layer(top mouse right edge and middle mouse), but does not stain uninfected mouse tissue(bottom mouse). The anti-cytoplasmic domain specific antisera stains similarly though not as strongly as the whole lectin antisera.Staining of infected mouse cecum is only seen in the presence of trophozoites, with areas of the infected cecum where there are notrophozoites nearby not stained with the anti-lectin antibodies. All photos at 40=.
suggesting a process of cell secretion(Fig. 4B).Cytosolic areas of the parasites were labeled inlesser degree. When we used the monoclonal anti-GalyGalNAc lectin antibody the intensity of thelectin label was lesser than with the polyclonalantibody. Contrarily, the GalyGalNAc lectin wasrandomly distributed in the parasite cytosol(Fig.4C) and scarcely in plasma membrane and vacu-olar membranes(Fig. 4D).
4. Discussion
This is the first report that demonstrates bindingof theE. histolytica GalyGalNAc inhibitable lectinto various target cells such as hepatocytes, both ofhuman and hamster origins and in experimentallyproduced hepatic and intestinal amoebic lesions.Using immunocytochemical techniques we identi-fied the GalyGalNAc lectin on the surface and in

44 J. Pacheco et al. / Parasitology International 53 (2004) 35–47
Fig. 4. Immunoelectronmicroscopy of culturedE. histolytica trophozoites using gold-labeled polyclonal(A and B) or monoclonal(C and D) anti-GalyGalNAc lectin antibodies.(A) Membranes of large and medium sized vacuoles and plasma membrane ofamoeba are labeled with the lectin(arrows). (B) Trophozoite plasma membrane associated to a cytoplasmic vacuole(v) shows agreater gathering of lectin labeling suggesting a process of cell secretion(arrow-heads). Some cytosolic areas are also labeled.(C)Trophozoite cytosol shows randomly distributed lectin labeling(arrows), (v) vacuoles.(D) The monoclonal antibody labeled onlyscarcely the amoeba plasma membrane, higher label is seen in the cytoplasm(arrows). Barss0.5mm.
the cytoplasm of cultured hepatocytes, suggestingthat these parenchymal cells express glycoproteinscapable of binding to the lectin and that thismolecule can be introduced inside the cellcytoplasm.Our results also showed that hepatocytes local-
ized at some distance from trophozoites werelabeled with the anti-lectin antibody, suggesting aprocess of secretion and diffusion of the lectinbeyond the vicinity of the parasites. These resultscontrast with data reported by Leroy et al.w12x,who showed that the antigen recognized by theCD6 antibody transfers to host cells only when
the amoebae are in direct contact with the epithelialcells. Based on fluorographs of SDS-PAGE, thesame authors showed that the CD6 monoclonalantibody recognizes bands approximately 170 and35 kDa. Although, this electrophoretic patternresembled to that obtained with the Mab 7F-4directed against GalyGalNAc lectin, immunocyto-chemistry using monoclonal and polyclonal anti-bodies against the same lectin did not label theepithelial cells. In our analysis, we used the samethree anti-GalyGalNAc antibodies(H85, 3F4 andShiro) tested by Leroy et al.w12x, and we observedthat intestinal cells, hepatocytes and inflammatory

45J. Pacheco et al. / Parasitology International 53 (2004) 35–47
cells were positively labeled, consistent with thetransfer of the lectin to these cells.Data obtained in the present work suggest that
capture of lectin by target cells could also berelated to the cytopathic effect of the parasite,since a large number of hepatocytes binding thisspecific lectin showed morphological changes sug-gestive of cell damage, including rounding, vacu-olization, loss of normal architecture andcondensed nuclei.Other lectins have been shown to mediate dam-
age to cellsw19–21x. Saffer and Petriw22x howeverdemonstrated that the purified GalyGalNAc lectinlacks any cytotoxic activity against Chinese ham-ster ovary cells. Recently, specific inhibitors wereused to assess the participation of three types ofmolecules in the cytotoxic effect ofE. histolyticaon MDCK monolayers: galactose-specific adhes-ins, phospholipase A, and cysteine proteinasesw23x. The only inhibitor that effectively blockedthe lost of transepithelial electrical resistancecaused by the parasite was galactose, suggestingthat the GalyGalNAc lectin constitutes an impor-tant factor for adherence-dependent amoebic cyto-toxicity. Of course, our results do not rule outtransfer, andyor a contribution, of the other amoe-bic molecules toE. histolytica cytotoxicity.Apoptosis has been shown to be induced by
micromolar concentrations of some lectinsw24,25x.In the case ofE. histolytica Ragland et al.w26xreported the damage of a murine myeloid cell line(FDC-P1) through an apoptotic mechanism thatshowed DNA fragmentation in a ladder pattern.This DNA fragmentation was inhibited by block-ing adherence with galactose, suggesting the par-ticipation of GalyGalNAc lectin. Although themorphological changes observed in some damagedcells in our samples suggest an apoptotic mecha-nism, this would have to be confirmed by bio-chemical and molecular techniques.This is the first study to determine the locali-
zation of the GalyGalNAc lectin during the evo-lution of ALA. The lectin was observed as earlyas 6 h post-inoculation on the sinusoidal cells,probably reflecting the presence of trophozoites inthe sinusoids during their arrival to the hepaticparenchyma. Capture of the lectin by inflammatorycells, including sinusoidal Kupffer cells had not
been clearly determined previously. Our in vivohistochemical studies performed at different timesof evolution of ALA showed that the lectin isrelatively easily captured by host acute(neutro-phils) as well as chronic inflammatory cells(mac-rophages, epithelioid and Kupffer cells). Moreover,the presence of large amount of lysed cells in theinflammatory foci suggests that this lectin mayalso be acting in their destruction. The presenceof the GalyGalNAc lectin during the ALA devel-opment could be associated with other amoebicproteins or molecules, since as mentioned before,similar qualitative immunolabeling was notedwhen whole amoebic lysates were used in thesame animal modelw27x.Immunodetection of anti-whole lectin as well as
the cytoplasmic domain of the molecule in theepithelium of mouse intestine interacting withtrophozoites indicates that this lectin are presentin the amoebic intestinal ulcers as well as hepaticabscesses.The apparent secretion of the GalyGalNAc lec-
tin during the host-parasite interaction observed inthese immunohistochemical studies led us to exam-ine the intracellular location of the lectin in tro-phozoites. Immunoelectron microscopy showedthat besides the plasma membrane of the parasites,intracellular vacuole membranes and cytosolic are-as of the cells were also labeled with the lectin.Interestingly, some particular zones ofE. histoly-tica plasma membrane associated to peripheralcytoplasmic vacuoles showed a high concentrationof gold-labeled anti-lectin, suggesting a process ofsecretion. These results support our previous lightmicroscopy observations in which diffusion ofGalyGalNAc lectin to the hepatocytes at the vicin-ity of the E. histolytica trophozoites was observed.In sum, our results demonstrate diffusion and
binding of the amoebic GalyGalNAc lectin tocultured human and hamster hepatocytes duringthe in vitro interaction, and to inflammatory andsinusoidal cells, and neighbouring hepatocytes dur-ing in vivo induction of ALA. Most cells bindingthe amoebic lectin showed different degrees ofmorphological damage, suggesting the participa-tion of the molecule in target cell injury. Detectionof the cytoplasmic tail of the heavy subunit oflectin in trophozoites and epithelium of intestinal

46 J. Pacheco et al. / Parasitology International 53 (2004) 35–47
mucosa of experimentally induced ulcers, alsosuggests that this specific fragment of the lectin ispresent during the host-parasite interaction andcould possibly be participating in some process ofcell damage. Absence of lectin labeling in someinteracting and damaged cells confirms that mech-anisms of pathogenesis of amoebae are multipleand other factors are also likely involved incytotoxicity.
Acknowledgments
The authors wish to thank Ascencion Hernan-´ ´dez, Lauren Lockhart, Adrian Rondan and Dago-´ ´berto Carrasco-Ceron for technical assistance. JP´received a scholarship from Conacyt(Mexico).´This work was partially supported by Conacyt(Mexico).´
References
w1x Martınez-Palomo A, Gonzalez-Robles A, Chavez B,´ ´ ´Orozco E, Fernandez-Castelo S, Cervantes A. Structural´bases of the cytolytic mechanisms ofEntamoeba histo-lytica. J Protozool 1985;32:166–75.
w2x Ravdin JI, Croft BY, Guerrant RL. Cytopathogenicmechanism of Entamoeba histolytica. J Exp Med1980;152:377–90.
w3x Ravdin JI, Guerrant RL. Role of adherence in cytopath-ogenic mechanism ofEntamoeba histolytica. Study withmammalian tissue culture cells and human erythrocytes.J Clin Invest 1981;68:1305–13.
w4x Mann BJ, Torian JBE, Vedvick TS, Petri WA Jr..Sequence of a cysteine-rich galactose-specific lectin ofEntamoeba histolytica. Proc Natl Acad Sci1991;88:3248–52.
w5x Mann BJ, Lockhart LA. Molecular analysis of the GalyGalNAc adhesin ofEntamoeba histolytica. J EukaryotMicrobiol 1998;45:13S–16S.
w6x Petri WA Jr., Chapman MD, Snodgrass T, Mann BJ,Broman J, Ravdin JI. Subunit structure of the galactoseandN-acetyl-D-galactosamine inhibitable adherence lec-tin of Entamoeba histolytica. J Biol Chem1989;264:3007–12.
w7x Ravdin JI, John JE, Johnston LI, Innes DJ, GuerrantRL. Adherence ofEntamoeba histolytica trophozoitesto rat and human colonic mucosa. Infect Immun1985;48:292–7.
w8x Ravdin JI, Murphy CF, Salata RA, Guerrant RL, HewlettEL. TheN-acetyl-D-galactosamine-inhibitable adherencelectin of Entamoeba histolytica. Partial purification andrelation to amoebic virulence in vitro. J Infect Dis1985;151:804–15.
w9x Bracha R, Kobiler D, Mirelman D. Attachment andingestion of bacteria by trophozoites ofEntamoebahistolytica. Infect Immun 1982;36:396–406.
w10x Salata RA, Pearson RD, Ravdin JI. The interaction ofhuman leukocytes withEntamoeba histolytica: killingof the virulent amoebae by the activated macrophage. JClin Invest 1985;76:491–9.
w11x Salata RA, Ravdin JI. The interaction of human neutro-phils andEntamoeba histolytica increases cytopathogen-icity for liver cell monolayers. J Infect Dis1986;154:19–26.
w12x Leroy A, De Bruyne G, Mareel M, Nokkaew C, BaileyG, Nelis H. Contact-dependent transfer of the galactose-specific lectin ofEntamoeba histolytica to the lateralsurface of enterocytes in culture. Infect Immun1995;63:4253–60.
w13x Diamond LS, Harlow DR, Cunnick CC. A new mediumfor the axenic cultivation ofEntamoeba histolytica andother Entamoeba. Trans R Soc Trop Med Hyg1978;72:431–2.
w14x Berry MN, Friend DS. High yield preparation of isolatedrat liver parenchymal cells. A biochemical and finestructural study. J Cell Biol 1969;43:506–20.
w15x Tsutsumi V, Mena-Lopez R, Anaya-Velazquez F,Martınez-Palomo A. Cellular bases of experimental´amoebic liver abscess formation. Am J Pathol1984;117:81–91.
w16x Cote SL, Ribeiro-Da-Silva A, Cuello AC. Currents´protocols for light microscopy immunocytochemistry.In: Cuello AC, editor. Immunohistochemistry II. NewYork: Wiley and Sons Press, 1993. p. 147–68.
w17x Shcerand JB, Thammapalerd N, Riganti M, TharavanijS, Punpoowong B. Monoclonal antibody based immu-nohistochemical demonstration ofEntamoeba histolyti-ca in liver tissue of experimentally infected hamster.Int J Parasitol 1994;24:909–16.
w18x Merighi A. Immunogold labelling procedures. In: PolakJM, Priestley JV, editors. Electron microscopic immu-nocytochemistry principles and practice. New York:Oxford University Press, 1992. p. 65–74.
w19x Schwartz RE, Wojciechowicz DC, Picon AI, SchwarzMA, Paty PB. Wheatgerm agglutinin-mediated toxicityin pancreatic cancer cells. Br J Cancer 1999;80:1754–62.
w20x Leist M, Wendel A. A novel mechanism of murinehepatocyte death inducible by concanavalin A. J Hepatol1996;25:948–59.
w21x Chen J, Lee KH, Steinhauer DA, Stevens DJ, SkehelJJ, Wiley DC. Structure of the hemagglutinin precursorclavage site, a determinant of influenza pathogenicityand the origin of the labile conformation. Cell1998;5:409–17.
w22x Saffer LD, Petri WA Jr.. Role of the galactose lectin ofEntamoeba histolytica in adherence-dependent killingof mammalian cells. Infect Immun 1991;9:4681–3.

47J. Pacheco et al. / Parasitology International 53 (2004) 35–47
w23x Lopez-Vancell R, Montfort I, Perez-Tamayo R. Galac-´ ´tose-specific adhesin and cytotoxicity ofEntamoebahistolytica. Parasitol Res 2000;6:226–31.
w24x Wada J, Ota K, Kumar A, Wallner EI, Kanwar YS.Developmental regulation, expression, and apoptoticpotential of galectin-9, a beta-galactoside binding lectin.J Clin Invest 1997;99:2452–61.
w25x Dong HD, Kimoto Y, Takai SI, Taguchi T. Apoptosisas a mechanism of lectin dependent monocyte-mediatedcytotoxicity. Immunol Invest 1996;25:65–78.
w26x Ragland BD, Ashley LS, Vaux DL, Petri WA Jr..Entamoeba histolytica: target cells killed by trophozoitesundergo DNA fragmentation which is not blocked byBcl-2. Exp Parasitol 1994;79:460–7.
w27x Ventura-Juarez J, Campos-Rodrıguez R, Tsutsumi V.´ ´
Early interactions ofEntamoeba histolytica trophozoiteswith parenchymal and inflammatory cells in the hamsterliver: an immunocytochemical study. Can J Microbiol2002;48:123–31.


















![Characterization of glycosphingolipids from Schistosoma mansoni eggs carrying Fuc(��1-3)GalNAc, GalNAc(��1-4)[Fuc(��1-3)]GlcNAc- and Gal(��1-4)[Fuc(��1-3)]GlcNAc-](https://static.fdokumen.com/doc/165x107/63176e09b6c3e3926d0ddfa3/characterization-of-glycosphingolipids-from-schistosoma-mansoni-eggs-carrying-fuc1-3galnac.jpg)

