
The Jacob2 Lectin of the Entamoeba histolytica Cyst Wall Binds Chitin and Is Polymorphic
Upload
independentCategory
view
5download
0
Experimental Parasitology 109 (2005) 150–162
www.elsevier.com/locate/yexpr
Entamoeba histolytica: inhibition of cellular functions by overexpression of EhGEF1, a novel Rho/Rac guanine
nucleotide exchange factor
Arturo Aguilar-Rojasa, Ma. de Jesús Almaraz-Barreraa, Mickaël Krzeminskid, Martha Robles-Floresb, Rosaura Hernández-Rivasa, Nancy Guillénc,
Rachid C. Maround, Miguel Vargasa,¤
a Departamento de Biomedicina Molecular, Centro de Investigación y de Estudios, Avanzados del IPN, Mexicob Facultad de Medicina, Universidad Nacional Autónoma de México, México, D.F., Mexico
c Unité de Biologie Cellulaire du Parasitisme, Institut National de la Santé et de la Recherche Médicale U389, Franced Unité de Bioinformatique Structurale, Institut Pasteur, Paris, France
Received 18 June 2004; received in revised form 8 December 2004; accepted 13 December 2004Available online 26 January 2005
Abstract
The molecular, biochemical, and cellular characterization of EhGEF1 protein is described. Complete cDNA sequence of 1890 bprevealed an open reading frame that encodes a protein of 69 kDa. EhGEF1 is constituted of Dbl homology domain, pleckstrinhomology domain, and several putative regulation sites. Studies of guanine nucleotide exchange activity of EhGEF1 on severalGTPases from Entamoeba histolytica and Homo sapiens showed preferential activation on EhRacG, suggesting that EhGEF1 pro-tein could be involved in mechanisms related to actin cytoskeleton activation, cytokinesis, capping, and uroid formation in trophozo-ite. Confocal microscopy studies of pExEhNeo/HSV-tagged-EhGEF1-transfected cells showed that trophozoites stimulated withConA, EhGEF1, and EhRacG were localized at plasma membrane. Cellular studies showed that F-actin content of pExEhNeo/HSV-tagged-EhGEF1-transfected trophozoites as well as cellular migration and cell damage capacity were signiWcantly altered. Theobservations suggest that EhRacG was the principal target of EhGEF1 and that EhGEF1 may provide a link between F-actindynamics and EhRacG signaling. 2004 Elsevier Inc. All rights reserved.
Index Descriptors and Abbreviations: Entamoeba histolytica; GEF, guanine nucleotide exchange factor; Dbl, diVuse B-cell lymphoma; mant-GDP,N-methylanthraniloyl-GDP; DH, Dbl homology; PH, pleckstrin homology; HSV, Herpes simplex virus glycoprotein D; GST, glutathione S-transferase
1. Introduction
The enteric protozoan parasite Entamoeba histolyticais the causative agent of amebiasis, a common healthproblem in many developing countries of the world.E. histolytica, like other enteric ameba of the genus Ent-amoeba, resides in the colon as a motile, feeding form
* Corresponding author. Fax: +52 55 57477134.E-mail address: [email protected] (M. Vargas).
0014-4894/$ - see front matter 2004 Elsevier Inc. All rights reserved.doi:10.1016/j.exppara.2004.12.013
(trophozoite) and travels from host to host in anencysted form. Hallmarks of E. histolytica virulence areintimately related to its motile capacities to penetratehuman tissues and its powerful cytotoxic activity onhuman cells. Close contact between the parasite and tar-get eukaryotic cells is mediated by speciWc surface mole-cules, suggesting that parasite–cell membraneinteractions have an important role in virulence.Cytolytic as well as phagocytic and motility activity ofE. histolytica are based on the highly dynamic actin

A. Aguilar-Rojas et al. / Experimental Parasitology 109 (2005) 150–162 151
polymerization. These cellular functions are essential fortissue invasion in host.
Many features of the cell behavior of eukaryotic cellsare regulated by Rho family GTPases, and their promi-nent eVects targetted on the actin cytoskeleton. Severalstudies have shown that there exist diVerent types ofsmall GTPases in E. histolytica; moreover, it has beensuggested that the function and regulation of these mole-cules are intimately related to pathogenic mechanisms ofE. histolytica such as phagocytosis for EhRacA (Ghoshand Samuelson, 1997), cytokinesis, morphogenesis, andmigration for EhRacG (Guillén et al., 1998; Labruyèreet al., 2002). However, knowledge of the functional regu-lation and molecular target of small GTPases in E. his-tolytica is scarce. Members of the Ras superfamily ofGTPases and their regulators have been studied in higheukaryotic cells, and the results of these studies suggestthat small GTPases have a critical role in regulating awide range of cellular processes such as cellular growth,diVerentiation, vesicle transport, nuclear transport, andactin cytoskeleton regulation, among others (Burridgeand Wennerberg, 2004). The small GTPases act asbinary switches between active GTP-bound and inactiveGDP-bound states due to their intrinsic GTPase activity(Boguski and McCormick, 1993). The GTP-bound stateforms complexes that activate downstream eVector tar-get molecules to stimulate cytoplasmic signaling cas-cades. Subsequently, intrinsic GTP hydrolysis activityregenerates the GDP-bound form to terminate the activestate. Spontaneous GTP hydrolysis and the GDP/GTPnucleotide exchange reactions of Rho and other Rassuperfamily GTPases are both very slow. Three classesof regulatory proteins inXuence the active and inactiveconformations: the GTPase activating proteins (GAPs);the guanine nucleotide exchange factors (GEFs); and theguanine nucleotide dissociation inhibitors (GDIs). GAPsenhance intrinsic GTP hydrolysis rate and attenuatethereby small GTPase protein activity by promoting theGDP-bound state (Bishop and Hall, 2000; Boguski andMcCormick, 1993). In contrast, GEFs serve as positiveregulators of Rho family proteins by catalyzing theexchange of GDP for GTP, which results in increasedlevels of active GTP-bound Rho GTPases in cell. More-over, GDIs bind GTPases and stabilize their conforma-tion either in the GDP- or GTP-bound state. Most RhoGEF proteins belong to the Dbl family. However, someunconventional, non-Dbl RhoGEFs also exist (Ericksonand Cerione, 2004). The Wrst member of Dbl familyidentiWed was Dbl (for the transforming protein ofdiVuse B-cell lymphoma) (Eva et al., 1998). Characteris-tic members of this family show potent transforming andinvasion properties, which are largely dependent on theirability to activate Rho GTPases (Collard, 1996; White-head et al., 1997; Zheng et al., 1995). Rho GEFs shareapproximately 300 homologous amino acids consistingof two tandem and functionally inter-related segments
known as Dbl homology (DH) and pleckstrin homology(PH) domains (Cerione and Zheng, 1996; CherWls andChardin, 1999). The DH domain (»200 amino acid resi-dues) is involved in the catalytic reaction that stimulatesGDP/GTP exchange through binding to the GTPaseand induces a conformational change in the latter (Cher-Wls and Chardin, 1999). The PH domain (»100 aminoacid residues) is located immediately C-terminal to theDH domain. It has been reported that the primary roleof the PH domain is to anchor the protein to the mem-brane by interacting with phosphoinositides (Whiteheadet al., 1999); however, it was recently reported that thisdomain seems to have, in addition, other functionsrelated to regulation of GTPase activity in vivo and cel-lular transformation (Rossman et al., 2003).
In the present study, the molecular, biochemical, andcellular characterization of the Wrst guanine nucleotideexchange factor from E. histolytica, named EhGEF1,shares homologies with the Dbl oncogene family.EhGEF1 was able to properly activate in vitro EhRacGand EhRho1 from E. histolytica, but EhRacA, EhRacCor EhRas1 was not well activated. EhGEF1 andEhRacG activation was intimately related to F-actinpolymerization and some cellular processes such asmotility and cell damage capacity of E. histolytica.
2. Materials and methods
2.1. Entamoeba histolytica and bacterial strain growth conditions
Trophozoites from pathogenic HM1:IMSS strainwere grown at 37 °C in TY1-S-33 medium and harvestedas previously described (Diamond et al., 1978). Bacterialstrains Escherichia coli DH5� and BL21(�E3) were usedfor recombinant DNA techniques and grown at 37 °C inLuria–Bertani (LB) medium (Sambrook and Russell,2001).
2.2. Polymerase chain reaction ampliWcation of the EhGEF1 protein-encoding gene, computer analysis, and nucleotide sequencing
A fragment of 258 bp of the EhGEF1 gene was ampli-Wed by PCR. Sequences of direct and reverse oligonucle-otides were 5�-GAWTTMACWTCRTTMTTSATWACVCCVATWCAM-3� and 5�-WATSTAAGAWGGACCACCAACWTTWAAWTGWAG-3�, respectively. Thesewere designed according to conserved amino acidsequences of DH domain and using the codon usage ofE. histolytica. PCR was carried out in a 100 �l reactionvolume using buVer provided by the manufacturer(USB), 20 pmol of each primer, 100 ng E. histolyticagenomic DNA as template, 1.5 mM deoxynucleotide tri-phosphates (Pharmacia, La Jolla, CA, USA), 1.5 mM

152 A. Aguilar-Rojas et al. / Experimental Parasitology 109 (2005) 150–162
MgCl2, and 2.5 U Taq DNA polymerase (USB). Condi-tions for PCR were 35 cycles of denaturation at 92 °C for1 min, primer annealing at 50 °C for 1 min, and extension72 °C for 1 min; at the end of ampliWcation reaction, sup-plementary incubation was performed for 7 min at 72 °C.PCR products were separated electrophoretically in 1%agarose gels and cloned into plasmid pCR 2.1 (Invitro-gen). DNA sequencing was carried out for both DNAsemploying overlapping oligonucleotide primers and ABIPRISM 310 DNA sequencer using Ampli Taq DNApolymerase and FS dye terminator cycle sequencingchemistry (Qiagen, Germany). To determine the pres-ence of potential open reading frame (ORF) in thesequence, the BlastX program was used (Altschul et al.,1997). Selection of related sequences and comparison ofamino acid sequences were, respectively, carried outusing the BlastP (Altschul et al., 1997) and ClustalWcomputer programs (Thompson et al., 1994).
2.3. Isolation of cDNA clones encoding the EhGEF1 protein
A �ZAP II library (kindly provided by Dr. E.Tannich, B.N. Institute, Hamburg, Germany) wasscreened with 258 bp PCR-ampliWed fragment thatwas described previously and used as probe. Theprobe was labeled using the Megaprime DNA labelingsystem kit (USB). One positive clone was isolated,zapped into pBluescript SK (+), and puriWed accordingto manufacturer’s recommendations (Stratagene). DNAsequencing was performed on both DNA strands, usingoverlapping oligonucleotide primers, and sequenced aspreviously described. This clone had a cDNA insert of996 bp. The corresponding EhGEF1 gene was truncatedat 5� and 3� ends. The missing 3� region of EhGEF1 wasampliWed by PCR using speciWc oligonucleotides andprimers derived from vector and cDNA sequence ofEhGEF1, and using a cDNA �ZAP II library from E.histolytica as DNA template. With this PCR fragment itwas possible to amplify an 800 pb fragment with a stopsite at the end. The missing 5� region was ampliWed byPCR using speciWc oligonucleotides and overlapping oli-gonucleotide primers derived from vector and cDNAsequence of EhGEF1, and using our genomic �gt11library of E. histolytica as DNA template.
2.4. DNA, RNA isolation, Northern and Southern blot analyses, and RNA dot blots
Amoebic DNA was isolated in a discontinuouscesium chloride gradient. Genomic DNA (approxi-mately 3�g) isolated from E. histolytica was digestedwith EcoRI and HindIII restriction endonucleases, ana-lyzed on 0.8% agarose gels in 1£ TBE buVer, and trans-ferred to nylon membranes. Following transfer, DNAwas cross-linked by exposing the membrane for 3 min to
UV (312 nm wavelength). Hybridization was carried outovernight at 42 °C in a buVer containing 50% formam-ide, 5% Denhardt’s, 5£ SSC, 0.5% SDS, and 100 �g ml¡1
salmon sperm DNA, and was hybridized with anEhGEF1 probe. The complete ORF of the EhGEF1gene was labeled with [32P]ATP using the multiprimerDNA labeling kit (Amersham, UK). The membraneswere washed to Wnal stringency of 0.1£ SCS, 0.1% SDSat 65 °C for 3 h and subsequently exposed to an X-rayWlm at ¡80 °C for 18 h. Total RNA was isolated fromE. histolytica by using Trizol reagent (Invitrogene, USA)and treated with DNAase RNAase free; then samples(20 �g total RNA per lane) were electrophoresed andtransferred to nylon membranes. An [�32P]ATP labeleddouble strand DNA of complete EhGEF1 gene was usedas probe in Northern and Southern blot assays, as wellas in RNA dot blot experiments. Whereas a fragment ofribosomal RNA 16S (876 bp) from E. histolytica (Clarkand Diamond, 1991) was ampliWed using Psp5(ggccaattcattcaatgaattgag) and Psp3 primers (ctca-gatctagaaacaatgcttctc) and used as control in RNA dotblot experiments.
2.5. Production of polyclonal antibodies and immunodetection against the EhGEF1 protein in total protein from E. histolytica
The 98 kDa protein corresponding to the GST-EhGEF1 fusion protein was puriWed according to stan-dard procedure of manufacturers. Eight mice wereintraperitoneally inoculated with 100 �g GST-EhGEF1fusion protein four times at 15-day intervals. For theWrst dose, this protein was emulsiWed with incompleteFreund’s adjuvant. Following the inoculation series,animals were sacriWced and serum was collected.Immunoglobulins were puriWed from serum by proteinA–Sepharose chromatography following standard pro-cedures (Pharmacia). Trophozoites were washed withphosphate-buVered saline (PBS) and lysed in the pres-ence of 100 mM p-hydroxymercurybenzoate (PHMB)(Sigma) in 100 mM Tris–HCl, pH 8.0. The mixture wasvortexed at 4 °C, diluted with Laemmli buVer (Laemmli,1970), and boiled for 5 min in a water bath. Protein con-centration was determined by the Bradford technique(Bradford, 1976). Aliquots containing 100 �g of proteinwere subjected to 10% sodium dodecyl sulfate (SDS)–polyacrylamide gel electrophoresis (PAGE). Proteinswere then transferred to nitrocellulose membrane (NC)(Towbin et al., 1979). The membrane was blocked over-night with 0.05% PBS-Tween (PBS-T) and 5% milkpowder. Subsequently, it was incubated for 15 h at 4 °Cwith polyclonal antibody against EhGEF1, washed withPBS-T, and incubated with secondary anti-mouse IgGantibody conjugated to horseradish peroxidase (SantaCruz Biotechnology). Finally, the blot was developedwith enhanced chemiluminescence kit ECL (Western

A. Aguilar-Rojas et al. / Experimental Parasitology 109 (2005) 150–162 153
blot kit, Amersham) or with 4-chloro-1-naphthol(Sigma).
2.6. Guanine nucleotide exchange assays
The analysis of N-methylanthraniloyl-GDP is similarto that described previously (Abe et al., 2000). To evalu-ate guanine nucleotide exchange activity of GST-EhGEF1from E. histolytica, full-length ORF of tagged EhGEF1protein was expressed and puriWed as mentioned, as wellas several reported GTPases, GST-EhRho1 (Lohia andSamuelson, 1993), GST-EhRacA and GST-EhRacC(Lohia and Samuelson, 1996), GST-EhRacG (Guillénet al., 1998), EhRas1 (Shen et al., 1994) from E. histolyticaand GST-HsRac1 and GST-HsCdc42 from Homo sapi-ens, were employed. To simplify experimental procedures,EhRacB and EhRacD were omitted in these experiments.EhRacB showed a high deduced amino acid identity tothe already described GTPase EhRacA (Ghosh and Sam-uelson, 1997) while EhRacD protein apparently is nottranscribed in cultured E. histolytica parasite (Lohia andSamuelson, 1996). Each recombinant GTPase was loaded(20�g Wnal concentration) with 400 nM N-methylanthra-niloyl-GDP (mant-GDP) and equilibrated undercontinuous stirring in exchange buVer containing 20 mMTris–HCl, pH 7.5, 10 mM MgCl2, 1 mM DTT, 50�g ml¡1
BSA, and 10% glycerol at 25 °C. After 20 min, GST-EhGEF1 (2 �g Wnal concentration; 1:10 enzyme to sub-strate ratio) was added in the presence of 10�M GTP.Relative decrease of mant-GDP Xuorescence(excitation D 360 nm, emission D 440 nm) was monitoredby Xuorescence spectroscopy. The Xuorescence was mea-sured using an ISS PC1 Photon Counting spectrometer.Experiments were performed in triplicate. GST proteinwas used instead of GST-EhGEF1 as control in guaninenucleotide exchange assays. Positive activation criteriawere established based on the amount of Xuorescenceremains between control and treatment (t test p < 0.01).The rate (K) of guanine nucleotide exchange was deter-mined by Wtting data as single exponential decays usingthe GraphPad Prism program.
2.7. Construction, expression, and puriWcation of HSV-tagged-EhGEF1, GST-EhGEF1, and GST-GTPase hybrid proteins
Full-length ORF of EhGEF1 as well as GTPasesEhRho1, EhRacA, EhRacC, EhRacG, and EhRas1 werecloned in a pGEX6P expression vector (Gibco, LifeTechnologies). BrieXy, all the sequences were ampliWedby PCR using genomic DNA as a template. EcoRI andXhoI restriction sites were introduced in the sense andantisense speciWc primers, respectively. 5�-GAAATTCCATGCTTGCATTTTCTGATATG-3�, and 5�-CCGCTCGAGCGGTCATACTAATAAAAGAAAAAT-3� forEhRho1; 5�-GGAATTCCATGCAAGCTGTCAAAT
GTGTC-3� and 5�-CCGCTCGAGCGGTTAGAATAATAAACATCCTCT-3� for EhRacA; 5�-GAATTCCATGAGTGAAAAACCCACATCA-3� and 5�-CCGCTCGAGCGGTTATAAAAGAGCACACTTTGA-3� forEhRacC; 5�-GGAATTCCATGAGACCAGTGAAACTTGTC-3� and 5�-CCGCTCGAGCGGTTAGAATAATGAGCATCCTTT-3� for EhRacG; and 5�-GGAATTCCATGACTGCCAATACATATAAA-3� and 5�-CCGCTCGAGCGGTTAGAACATTATGCATTTCTT-3�for EhRas1. PCR fragments were digested, puriWed, andcloned into pGEX6P expression vector (Gibco, LifeTechnologies) to obtain the GST-GTPase fusion pro-teins. GST-HsCdc42 and GST-HsRac1 vectors werekindly provided by A. Hall (University College London,UK), The DH domain (177 aa–391 aa) from EhGEF1was ampliWed by PCR using the following primers 5�-CGCGGATCCGCGATGAATAAATTATATGCATATAGAACTAAT-3� and 5�-TCCCCCGGGGGACCATCAGACGGTAAGTCAGTTATTCTTTCACTC-3� senseand antisense primers, respectively, and cloned atBamHI and SmaI sites of the pGEX2TK expression vec-tor (Gibco, Life Technologies). On the other hand, full-length ORF of EhGEF1 Xanked by SacI and NotI siteswas ampliWed by PCR using the following primers: 5�-CGAGCTCGTTATGATGGATTTATTCCCAGACCAAAAAGAATGT-3� and 5�-TTTTCCTTTTGCGGCCGCTTTTTTCCTTTTACTATCTGGT TTAGGAGATGAAGGAATTGAAACTGA-3�, and cloned usingthe pET43b expression vector (Novagen). To obtain theHVS-tagged-EhGEF1 gene, it was ampliWed by PCRusing the pET43b vector (Novagene) containing theEhGEF1 cloned gene and using the following primers5�-CGGGGTACCCCGATGATGGCTGCCCGTAATATTTGCTGGTTCGGTGACGAAGCG-3� and 5�-CGGGATCCCGTCTGTTTTATCAGCCTAGGAACGCCCAACTTAATTAACA-3�. Each primer was Xankedby KpnI-5� and BamHI-3� sites, respectively. PCR prod-ucts were puriWed and cloned at KpnI and BamHI sitesof the pExEhNeo episomal vector. All constructs weresequence veriWed.
PuriWcation of these GST fusion proteins with gluta-thione–Sepharose (Amersham) was carried out accord-ing to the manufacturer’s suggested protocol andanalyzed by SDS–PAGE (Laemmli, 1970). Brief,GST-fusion bacterial expression vector was transformedinto E. coli BL21 (�E3) strain. An overnight bacterialculture was grown at 37 °C in LB medium containing50�g ml¡1 ampicillin. In 250 ml of fresh medium con-taining antibiotic, 2.5 ml of overnight culture was added.Bacteria were grown for 2 h at 37 °C and 1 mM of ITPGwas added. The bacterial culture was grown for 4 h more.The cells were harvested by centrifugation at 4000g for15 min and disrupted by cell sonication in 50 mM Tris,150 mM NaCl, 5 mM MgCl2, and 100 mM dithiothreitol.The recombinant proteins were puriWed by aYnitychromatography using glutathione–Sepharose 4B

154 A. Aguilar-Rojas et al. / Experimental Parasitology 109 (2005) 150–162
(Amersham Biosciences). Proteins were washed andeluted with free and reduced glutathione in Tris-buVeredsaline medium (50 mM Tris, 150 mM NaCl, 5 mMMgCl2, and 1 mM dithiothreitol) and stored in 30% glyc-erol. The puriWed recombinant proteins were dialyzedagainst exchange buVer before the nucleotide exchangeassay.
2.8. Transfection of E. histolytica trophozoites with the pExEhNeo/HSV-tagged-EhGEF1 vector
Construction of pExEhNeo/HSV-tagged-EhGEF1involved the replacement of the Cat Xanking region ofpExEhNeo/Cat vector (kindly provided by Dr. E. Tan-nich, Hamburg, Germany) with full-length ORF ofHSV-tagged-EhGEF1 gene. The pExEhNeo episomalvector without Cat (Hamann et al., 1995) and recombi-nant plasmid pExEhNeo/HSV-tagged-EhGEF1 weretransformed into E. coli and puriWed using a maxi kitfrom Qiagen (Germany). E. histolytica was transfectedby electroporation with 200�g DNA/107 cells. Transfec-ted cells were grown for 48 h after electroporation andselected for resistance from 10 to 50�g ml¡1 G418.Expression of the EhGEF1 gene in pExEhNeo/HSV-tagged-EhGEF1 vector was under the control of anE. histolytica lectin gene promoter. Overexpression ofEhGEF1 in transfected cells was measured with anEagle Eye densitometer. To determine the subcellularlocalization of EhGEF1 protein by Xuorescence micros-copy in wild type trophozoites as well as in pExEhNeo/HSV-tagged-EhGEF1-transfected cells, we used a poly-clonal antibody against EhGEF1 protein and a mono-clonal antibody against HSV-Tag (HSV monoclonal,Novagen), respectively. Secondary FITC-labeling anti-bodies were used (Molecular Probes). Samples were pre-pared as previously described (Guillén et al., 1998).Fluorescent samples were examined using an immuno-Xuorescence microscope (BX-60 Olympus).
2.9. Confocal laser-scanning microscopy of EhGEF1 and EhRacG
Transfected E. histolytica trophozoites withpExEhNeo/HSV-tagged-EhGEF1 vector were trans-ferred to pre-warmed acetone-washed coverslips con-taining amoeba culture medium and allowed to adherefor 10 min at 37 °C. Culture medium was removed andtrophozoites were Wxed with 3.7% paraformaldehydepre-warmed at 37 °C and permeabilized with 0.1% Tri-ton/PBS for 1 min. To localize EhGEF1, the cells werewashed with PBS, quenched for 30 min in PBS contain-ing 50 mM NH4Cl, then blocked with 1% BSA/PBS for30 min, and incubated with monoclonal antibody againstHVS-Tag (HSV monoclonal antibody, Novagen). Apolyclonal antibody against EhRacG (Guillén et al.,1998) was diluted in blocked buVer, then added to cells,
and incubated for 1 h at 37 °C. Coverslips were washedwith 1% BSA/PBS. Subsequently, trophozoites wereincubated for 3 min with 1% BSA/PBS and incubated for2 h at 37 °C with secondary TRIC-labeling goat anti-rab-bit of EhRacG (Molecular Probes) and Cy5-labelinganti-mouse of EhGEF1 (Caltag Laboratories) pre-adsorbed on trophozoites. Preparations were furtherwashed with PBS and mounted on 10 �l CitiXuor (TedPella, CA, USA). Fluorescent samples were examinedwith an immunoXuorescence microscope (BMX-60Olympus) and then with a Zeiss confocal laser-scanningmicroscope. Observations were performed in 10 planesfrom bottom to top of each cell. Distance between scan-ning planes was 0.5 �m. For uroid formation and cap-ping of surface receptors, trophozoites were incubatedwith FITC-conjugated Concanavalin A (ConA) (Molec-ular Probes), as previously described (Guillén et al.,1998).
3. Evaluation of F-actin content in pExEhNeo/HSV-tagged-EhGEF1-transfected cells
Transfected E. histolytica trophozoites withpExEhNeo/EhGEF1 at 10 and 50 �g ml¡1 of G418 andcells transfected with pExEhNeo at 50 �g ml¡1 of G418were used to measure F-actin polymerization for 5 and10 min at 37 and 4 °C. Then, cells were Wxed for 30 minwith 3.5% paraformaldehyde in PBS 1£, washed withPBS, quenched in 0.1 M glycine in PBS for 30 min,washed twice, and permeabilized (0.2% Triton X-100 inPBS, 1% BSA) for 10 min. Trophozoites were stainedwith 0.5 �M Rhodamine phalloidin (Molecular Probes)for 30 min, washed two times, and analyzed on a FAC-SCalibur (Becton–Dickinson) at 554 nm of excitationand 573 nm of emission (Menu et al., 2002). The experi-ment was carried out three times for each strain using1 £ 106 cells every time and mean Xuorescence intensitywas used.
3.1. Evaluation of cellular migration and cytopathic capacity in pExEhNeo/HSV-tagged-EhGEF1-transfected cells
Cell migration was assayed as previously described(Franco et al., 1997). In brief, transfected E. histolyticatrophozoites with pExEhNeo/HSV-tagged-EhGEF1and cells transfected with pExEhNeo, both grown at50 �g ml¡1 G418, were incubated in TY1-S-33 mediumfor 3 h (serum starvation) at 37 °C. Then trophozoiteswere collected and washed three times in TBS–CaCl2buVer (50 mM Tris, 150 mM NaCl, and 1 mM CaCl2)and pelleted. After the washes, cells were resuspended inTBS–CaCl2 containing 100�g ml¡1 BSA. This suspen-sion (1 £ 104 cells resuspended in 125 �l TBS–CaCl2) wasadded to the upper chamber of Transwells (Costar). The

A. Aguilar-Rojas et al. / Experimental Parasitology 109 (2005) 150–162 155
lower chamber was Wlled with either 600 �l TY1-S-33 orTBS–CaCl2 as control. After 2 h, chambers were chilledin ice for 10 min and tapped several times on top with ametal spatula to dislodge all trophozoites that hadmigrated through Wlter pores but remained attached toit. Cells were Wxed with 600 �l of 7% PFA that was addedto lower wells incubated at 37 °C for 60 min and countedusing an inverted microscope. Chemotactic activity wasexpressed as the chemotactic index (CI). In pExEhNeoand pExEhNeo/HSV-EhGEF1 strains, %migration(random and chemotactic) was then quantiWed by thefollowing formula: %migration D number of cells in thedown chamber/total cells £ 100. The chemotactic activ-ity was then expressed as CI: CI D %chemotactic migra-tion/%random migration; CI D 1 indicates the absence ofchemotactic response (Capsoni et al., 1989). CytopathiceVect on Madin Darby Canine Kidney Epithelial cells(MDCK) was carried out as described (Garcia-Riveraet al., 1999) with pExEhNeo/HSV-tagged-EhGEF1-transfected trophozoites grown at 50 �g ml¡1 G418, aswell as with cells with pExEhNeo-transfected cellsgrown at 50 �g ml¡1 G418.
4. Results
4.1. Isolation and sequence analysis of the cDNA encoding the complete open reading frame of the guanine nucleotide exchange factor (EhGEF1) from E. histolytica
To identify the EhGEF1 gene, PCR on E. histolyticagenomic DNA was performed using degenerate primers.Among the PCR products, a DNA fragment of 280 bpencoding an ORF was found. The 280 bp product pre-sented homology to human faciogenital dysplasia 1 gene(FGD1). FDG1 encodes a guanine nucleotide exchangefactor for GTPase Cdc42, a member of the Rho familyproteins, which is involved in regulating cellular polarityand the assembly of Wlopodia (Zheng et al., 1996). Theidentity in sequence presented between FGD1 and280 bp fragment showed that it was part of the GEFgene from E. histolytica. The product was used as aprobe to screen a �ZAPII cDNA library from E. histoly-tica. One positive clone with an insert of 996 bp wasfound and sequenced. The cDNA corresponding toEhGEF1 was truncated at 5� and 3� ends. AnchoredPCR was used to obtain the missing segments. Oligonu-cleotides corresponding to the 5� or 3� ends of the Wrstpositive clone which contained the EhGEF1 were usedas well as primers from the vector. A genomic library�gt11 from E. histolytica was used as the DNA template;the ampliWed DNA fragment of 861 bp carrying the 5�end of gene overlapped 151 bp of the known sequence.To identify the 3� end, a �ZAPII cDNA library from E.histolytica was used as template. The ampliWed DNAfragment of 800 bp carrying the 3� end of the gene over-
lapped 109 bp of the known sequence. To conWrm thecontiguous nature of ampliWed DNA fragments, full-length EhGEF1 gene was ampliWed by PCR and theentire DNA sequence of EhGEF1 was performed asdescribed in Section 2.
The complete nucleotide sequence of the EhGEF1gene, consisting of 1893 bp, was submitted to GenBankwith Accession No. AY281052. The predicted protein of630 amino acids (Fig. 1) has a theoretical molecular massof 72.547 kDa and a calculated isoelectric point of 9.4,both calculated using the ExPASy server. A BlastPsearch revealed that the DH domain of EhGEF1 has26% identity with FGD1 from H. sapiens, an X-linkedskeletal dysplasia (FGDY; Arskog syndrome) thatadversely aVects skeletal development (Pasteris et al.,1994). The homology is restricted only to the DHdomain since EhGEF1 is smaller than FGD1. DH andPH are the only two domains present in EhGEF1,closely related to all the RhoGEFs. The DH domain ofEhGEF1 (Tyr183-Glu367, according to SMART) has ashort 26 amino acid fragment (Leu316-Pro342), whichwas identiWed as the signature domain of Dbl familyproteins (Fig. 1). Moreover, a search of sequence motifswith the ProScan server (Sigrist et al., 2002) resulted innine potential phosphorylation sites by protein kinase C(PKC), 11 potential casein kinase II (CK2) sites, twopotential cAMP and cGMP-dependent protein kinase(cA-PrK) sites, one potential tyrosine kinase (TYRK)site, two potential glycosylation (Gly) sites, one potentialtyrosine sulfation site, and two potential myristoylation(Myr) sites (Fig. 1).
The DH domain of EhGEF1 showed high evolution-ary conservation at the designated conserved regions(CR1 Tyr183-Phe209, CR2 Ile227-Gln246, and CR3Phe314-Thr341). In addition, essential amino acidswhose mutations have resulted in inactivation of thefunction in other GEFs were detected in these sequences(Aghazadeh et al., 1998). Aside from the DH domain,EhGEF1 has a PH domain (Asn393-Phe501 accordingto SMART). This contains some conserved residues aswell as the highly conserved C-terminal Trp500, whichhas been reported to interfere with global structure;interference that results in loss of cellular transformationin NIH3T3 murine Wbroblasts (Whitehead et al., 1995).A short conserved fragment (Lys433-Val453) was foundat the N-terminal domain of PH. This fragment has beenreported to be required for high-aYnity binding to phos-phatidylinositol-4,5-bisphosphate (PIP2) (Snyder et al.,2002). The organization of EhGEF1 in the genome of E.histolytica was analyzed by Southern blotting of geno-mic DNA digested with EcoRI and HindIII. In bothcases, two fragments hybridized with the EhGEF1-labeled probe, indicating that only one copy of EhGEF1was present per E. histolytica genome (Fig. 2A). To Wndout whether EhGEF1 is transcribed and translated,Northern blot and Western blot assays were performed,

156 A. Aguilar-Rojas et al. / Experimental Parasitology 109 (2005) 150–162
respectively. A unique band of about 2000 bases in size (64.3 § 1.3). These results are in good agreement with the
Fig. 1. Deduced amino acid sequence of EhGEF1. Amino acid numbers are shown on the left and right. The Dbl homology domain (DH) is shadedblue and its domain signature is underlined with a continuous line, whereas the pleckstrin homology (PH) domain is shaded pink. Sequence motifsspeciWc to PKC are shaded green; speciWc to CK2, yellow; speciWc to Myr, red; speciWc to TYRK, brown; speciWc to Gly are indicated with fourpoints; and speciWc to cA-PrK are indicated with double underlining. The CK2 sites 364SVNE367 and 506SAYE509 belong to the DH and PHdomains, respectively. Conserved regions CR1, CR2, and CR3 are shown in boxes. High-aYnity binding PIP2 sequence in PH domain (Snyder et al.,2002) is shown (black dotted line). Essential amino acids involved in inactivation (Aghazadeh et al., 1998; Whitehead et al., 1995) are labeled (¤).
corresponding to EhGEF1 was detected by Northernblotting (Fig. 2B), while Western blot analysis revealed aprotein of about 69 kDa in total extracts from E. histoly-tica (Fig. 2C). These results indicated that the EhGEF1gene was expressed in this parasite.
4.2. EhGEF1 displayed in vitro exchange activity toward EhRacG and EhRho1 from E. histolytica
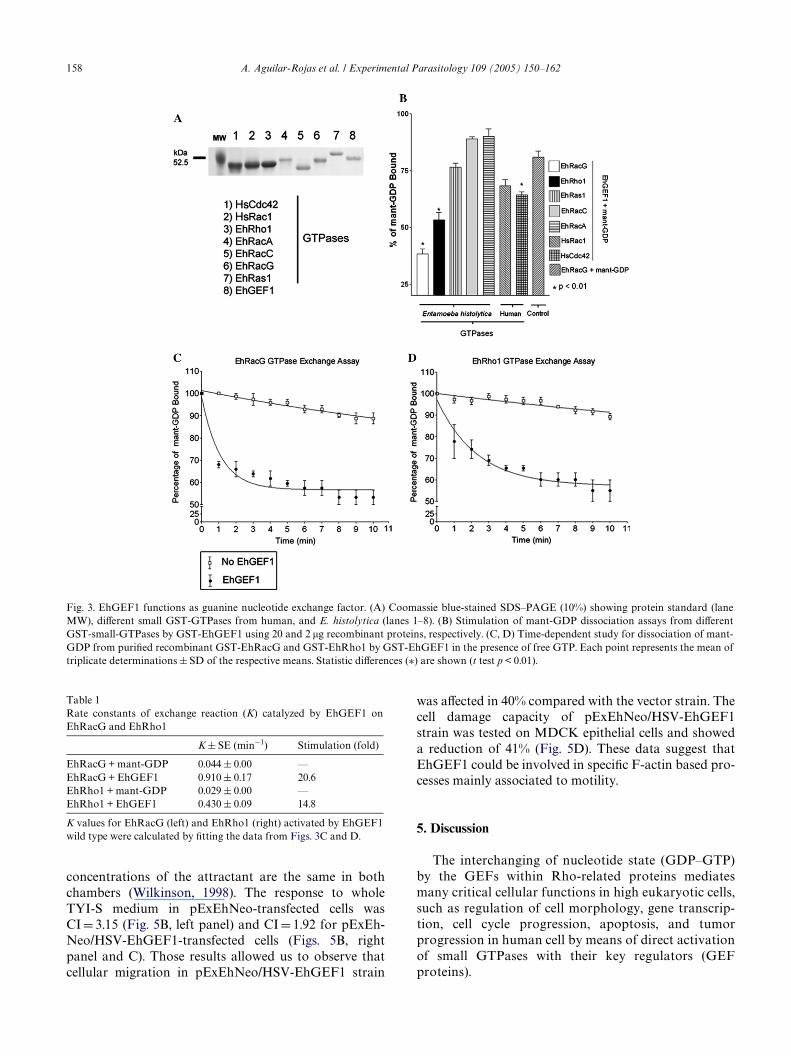
To examine the potential guanine nucleotideexchange activity of EhGEF1 as well as its speciWcity fordistinct Rho subfamily proteins, it was necessary to con-struct, express, and purify the following recombinantproteins: GST-EhGEF1; GST-EhRho1; GST-EhRacA;GST-EhRacC; and GST-EhRacG and GST-EhRas1from E. histolytica. Already existent GST-HsCdc42 andGST-HsRac1 from human were also used (Fig. 3A). Allof these proteins were expressed and puriWed from E.coli to evaluate the guanine nucleotide exchange activityof EhGEF1 on these diVerent small GTPases by Xuores-cence spectroscopy. It was found that EhGEF1 was notable to properly activate EhRacC (89.0 § 00), EhRacA(90.0 § 3.3) or EhRas1 (76.6 § 1.6) but displayed someactivity on HsRac1 (68.3 § 2.6) and HsCdc42
identity in sequence presented between EhGEF1 andFGD1. However, only two of the Wve proteins fromE. histolytica tested were eYciently activated byEhGEF1: EhRacG (40.1 § 3.8) and EhRho1 (53.3 § 3.3)(Fig. 3B); being EhRacG the main target in vitro. Toobtain the rate constants of guanine nucleotide exchangereaction, the data were best Wtted as a sum of threeexponential decays in Xuorescence (Figs. 3C and D).Results indicated that EhGEF1 eYciently catalyzed theexchange of bound GDP for GTP on EhRacG up to20.6-fold, whereas EhRho1 was of 14.8 (Table 1). Thisanalysis corroborated the preferential activation ofthe small GTPase EhRacG by the EhGEF1 factor ofE. histolytica.
4.3. EVect of EhGEF1 gene overexpression and its pathogenic-related functions
For a functional analysis of EhGEF1, the gene overex-pression strategy leading to a down-regulated phenotypewas used; therefore, the EhGEF1 gene was cloned inpExEhNeo episomal vector and the recombinant plasmidwas transfected into E. histolytica. Transfected parasiteswere selected at 10�g ml¡1 and grown at 50�g ml¡1 of

A. Aguilar-Rojas et al. / Experimental Parasitology 109 (2005) 150–162 157
G418. Dot blot using total RNA from transfected cellsshowed that there was 3-fold more transcription of theEhGEF1 gene in pExEhNeo/HSV-tagged-EhGEF1-transfected trophozoites (referred to as GEF1+ strain)grown at 50�g ml¡1 than in cells transfected with pExEh-Neo vector (vector strain) used as a control (Fig. 4A).These results conWrmed that the expression occurs intransfected cells. To verify if the high levels of EhGEF1protein in transfected cells did not aVect normal localiza-tion of this protein, microscopy assays were performed(Fig. 4B). Fluorescence microscopy analysis showed thatthe endogen EhGEF1 protein in wild type cells and alsoin pExEhNeo/HSV-EhGEF1-transfected cells has thesame immunodetection pattern, since in both cases the
Fig. 2. (A) Southern blot analysis of EhGEF1 gene in E. histolytica.DNA from E. histolytica was EcoRI (lane 1) or HindIII (lane 2)digested, separated in 0.8% agarose gel, transferred to nylon mem-branes, and hybridized with EhGEF1 gene used as probe as describedin Materials and methods. Kodak Wlms were exposed for 5 days tomembranes. (B) Detection of EhGEF1 in E. histolytica RNA. TotalRNA from E. histolytica was separated in 12% denatured agarose geland hybridized with EhGEF1 gene used as probe. (C) Detection ofEhGEF1 protein in E. histolytica. Protein extracts (200 �g) fromE. histolytica were electrophoresed in 10% SDS–PAGE, transferred tonitrocellulose, and analyzed by immunoblot for reactivity with poly-clonal anti-EhGEF1.
EhGEF1 was located at the plasma membrane regionand mainly at the pseudopod region.
To analyze the eVect of overexpression of EhGEF1on the pathogenic mechanism of E. histolytica, biologi-cal functions such as cellular localization during cappingof surface receptors, F-actin content, cellular migration,as well as the impact of the EhGEF1 overexpression oncell damage were studied.
Previous works have suggested a role for EhRacG inthe signaling triggered during capping of surface recep-tors in E. histolytica and uroid formation (Guillén et al.,1998). Therefore, it was analyzed whether these morpho-logical and signaling changes correlated to modiWcationsin EhGEF1 cell localization, as well as its co-localizationwith EhRacG in pExEhNeo/HSV-tagged-EhGEF1-transfected cells (HSV-GEF1+). Trophozoites werestimulated for capping of surface receptors and for uroidformation with Concanavalin A (ConA). Confocalmicroscopy showed that ConA did not produce anychange in EhGEF1 immunolocalization since cellsshowed a similar pattern to control cells (Fig. 4C)restricting its localization at the plasma membrane, placewhere it would activate its GTPase target. In addition,and as expected, EhRacG was mobilized towards theuroid region, as previously shown (Guillén et al., 1998),and co-localized with EhGEF1, which suggests thatEhRacG may be activated in vivo by this GEF (Fig. 4D).Therefore, EhGEF1 as well as EhRacG could haveeither a direct or indirect role in uroid formation.
On the other hand, using antibodies directed againstthe PVPKASVTTQAKSQE peptide from the carboxy-terminal domain of the small GTPase EhRho1 showedthat it was localized at the uroid region in pExEhNeo/HSV-EhGEF1-transfected cells stimulated with ConA(data not shown), place where both, the small GTPaseEhRacG and EhGEF1, were co-localized, these resultssuggesting the double GEF function of EhGEF1 onEhRacG and EhRho1 proteins.
The F-actin rearrangement is aVected whenEhRacG-V12 mutant is over-expressed (Guillén et al.,1998). In GEF1+ strain, using Xow cytometry analysis onF-actin amount clearly indicated a decrease in F-actincontent to 42% (Fig. 5A). To verify if there are changesin cell motility by overexpression of EhGEF1, a cellularmigration assay was carried out using the pExEhNeo/HSV-EhGEF1- and pExEhNeo-transfected cells. Acheckerboard analysis and also a chemotactic index wereobtained of the cellular migration assays (Fig. 5B).Those results allowed us to discriminate between directmigratory response (chemotactic) and the randommovement (chemokinetic) using diVerent dilutions ofTYI-S33 medium above and below the Wlters. The valuesbelow the diagonal (Fig. 5B) indicate responses to apositive gradient, whereas those up the diagonal indicateresponse to a negative gradient. The values along thediagonal indicate random motility because the
158 A. Aguilar-Rojas et al. / Experimental Parasitology 109 (2005) 150–162
was aVected in 40% compared with the vector strain. The
Fig. 3. EhGEF1 functions as guanine nucleotide exchange factor. (A) Coomassie blue-stained SDS–PAGE (10%) showing protein standard (laneMW), diVerent small GST-GTPases from human, and E. histolytica (lanes 1–8). (B) Stimulation of mant-GDP dissociation assays from diVerentGST-small-GTPases by GST-EhGEF1 using 20 and 2 �g recombinant proteins, respectively. (C, D) Time-dependent study for dissociation of mant-GDP from puriWed recombinant GST-EhRacG and GST-EhRho1 by GST-EhGEF1 in the presence of free GTP. Each point represents the mean oftriplicate determinations § SD of the respective means. Statistic diVerences (¤) are shown (t test p < 0.01).
Table 1
concentrations of the attractant are the same in bothchambers (Wilkinson, 1998). The response to wholeTYI-S medium in pExEhNeo-transfected cells wasCI D 3.15 (Fig. 5B, left panel) and CI D 1.92 for pExEh-Neo/HSV-EhGEF1-transfected cells (Figs. 5B, rightpanel and C). Those results allowed us to observe thatcellular migration in pExEhNeo/HSV-EhGEF1 strain
Rate constants of exchange reaction (K) catalyzed by EhGEF1 onEhRacG and EhRho1
K values for EhRacG (left) and EhRho1 (right) activated by EhGEF1wild type were calculated by Wtting the data from Figs. 3C and D.
K § SE (min¡1) Stimulation (fold)
EhRacG + mant-GDP 0.044 § 0.00 —EhRacG + EhGEF1 0.910 § 0.17 20.6EhRho1 + mant-GDP 0.029 § 0.00 —EhRho1 + EhGEF1 0.430 § 0.09 14.8
cell damage capacity of pExEhNeo/HSV-EhGEF1strain was tested on MDCK epithelial cells and showeda reduction of 41% (Fig. 5D). These data suggest thatEhGEF1 could be involved in speciWc F-actin based pro-cesses mainly associated to motility.
5. Discussion
The interchanging of nucleotide state (GDP–GTP)by the GEFs within Rho-related proteins mediatesmany critical cellular functions in high eukaryotic cells,such as regulation of cell morphology, gene transcrip-tion, cell cycle progression, apoptosis, and tumorprogression in human cell by means of direct activationof small GTPases with their key regulators (GEFproteins).

A. Aguilar-Rojas et al. / Experimental Parasitology 109 (2005) 150–162 159
Invasive amebiasis by human protozoan parasite is Rho/Rac GEF family at amino acid level indicates that
Fig. 4. Overexpression and immunolocalization of EhGEF1 and EhRacG during uroid formation in pExEhNeo/HSV-EhGEF1-transfected cells. (A)Expression of EhGEF1 in total RNA from transfected E. histolytica. RNA dot blots of pExEhNeo and pExEhNeo/HSV-EhGEF1-transfected cellswere hybridized on nylon membrane with EhGEF1 gene used as probe and 16S RNA ribosomal from E. histolytica used as control. Kodak Wlmswere exposed for 16 h to membranes and overexpression of EhGEF1 in transfected cells was measured with Eagle Eye densitometer. (B) Immunolo-calization of native EhGEF1 protein in wild type trophozoites using an anti-EhGEF1 serum (upper panels) and HSV-tagged-EhGEF1 protein usinga monoclonal antibody against HSV-tagged-EhGEF1 fusion protein in pExEhNeo/HSV-EhGEF1-transfected cells (lower panels), arrows indicatethe EhGEF1 membrane association, binding was detected with FITC-conjugated goat anti-mouse IgG. (C) Distribution of EhGEF1 and EhRacG innon-stimulated pExEhNeo/HSV-EhGEF1-transfected cells. (D) Confocal microscopy of EhGEF1 and EhRacG in stimulated pExEhNeo/HSV-EhGEF1-transfected cells, co-localization of both proteins is indicated by an arrow. Transfected cells were incubated with anti-HSV-tag monoclonalantibody for EhGEF1 and anti-EhRacG serum, and incubated with FITC-ConA to induce capping and uroid. Binding was detected with IgGs-Cy5-labeled or IgGs-TRIC-labeled antibodies, respectively. Bar D 10 �M.
one of the most common diseases present in lessdeveloped countries. The motile form of the causativeagent of amebiasis, the trophozoite, expresses its patho-genetic ability by invading and destroying human epi-thelial tissues. This phenotype has been characterized byproduction of proteolytic activity, adherence phagocyto-sis, metastatic and invasive behavior, and also by travel-ing out of the colon due to hematogenous disseminationand high mobility. These characteristics are similar tothose of certain transformed cell phenotypes (Schmitzet al., 2000). During these invasive and metastasis pro-cesses, trophozoites interact with diVerent host cells andtissues where the small GTPases as well as their masterregulators (GEFs) could be involved in the metastaticand invasive behavior of E. histolytica, however in thisparasite these last molecules remain poorly studied.
In this work, the Wrst Dbl family member to be identi-Wed in the human parasite E. histolytica is described. It isa new guanine nucleotide exchange factor that wenamed EhGEF1 on the basis of its sequence similarity toother Rho/Rac GEF family members. EhGEF1 has thetwo structural motifs shared by all these family mem-bers, i.e., the DBL homology (DH) domain responsiblefor the GEF activity, and the pleckstrin homologydomain (PH). Over the DH domain, EhGEF1 showedthat high sequence conservation with members of the
this is a new Rho-GEF in E. histolytica. Although thePH domain from EhGEF1 showed less similarity toother domains of the family, studies concerning its possi-ble target suggest that it is a Ptdlns(3,4)P2-recognitiondomain (data not shown). The N- and C-termini nearthe DH and PH domains in EhGEF1 do not show sig-niWcant stretches of homology with other Dbl proteins,and it remains to be established if they are involved inregulation and activation of EhGEF1, like in other GEFproteins (Bustelo, 2000).
Biochemical studies conWrmed that EhGEF1 pro-motes in vitro nucleotide exchange from GDP to GTP,mostly in the small GTPases EhRacG and EhRho1. Rel-atively low activity was observed in HsCdc42 andHsRac1 promoted by EhGEF1, while EhRacA,EhRacC, and EhRas1 were not well activated byEhGEF1. These data indicated that EhGEF1 is relatedto diVerent cell-signaling pathways in E. histolytica. Oneof them is eVected through EhRacG activation, which isinvolved in uroid formation, F-actin polymerization(Guillén et al., 1998), and cellular migration (Tavareset al., 2000). Another may be related to EhRho1 activa-tion by EhGEF1. Although to date there is no informa-tion available regarding the biological activity ofEhRho1 in E. histolytica, its 47% identity with thehuman small GTPase RhoA that controls the assembly

160 A. Aguilar-Rojas et al. / Experimental Parasitology 109 (2005) 150–162
of actin stress Wbers and focal adhesion (Mackay and observed previously in other DBL family members.
Fig. 5. QuantiWcation of F-actin measured by Xow cytometry of pExEhNeo and pExEhNeo/HSV-EhGEF1-transfected cells. (A) Transfectant cellswere incubated for 5 and 10 min at 37 °C and then they were Wxed and stained with rhodamine-phalloidine. After staining, F-actin was measured byFACS as indicated in Materials and methods. Transfected cell incubated at 4 °C served as control as it disrupts F-actin cytoskeleton. (B) Checker-board analysis of chemotactic and chemokinetic activity responses of pExEhNeo (left) and pExEhNeo/HSV-EhGEF1-transfected cells (right) toTY1-S-33. (C) Motility response of trophozoites is expressed as chemotactic index, graphic representation of chemotactic response was more intenseby pExEhNeo (lane 1) and pExEhNeo/HSV-EhGEF1-transfected cells (lane 2) to TY1-S-33. (D) Cytopathic eVect (CP) of E. histolytica on MDCKcell monolayers by pExEhNeo and pExEhNeo/HSV-EhGEF1-transfected cells were measured as described in Materials and methods. Error barsindicate standard deviation (SD) of three experiments.
Hall, 1998; Van and Souza, 1997) suggests that EhGEF1and EhRho1 could be involved in similar functions in E.histolytica. The speciWc constant for the exchange activ-ity of EhGEF1 over EhRacG and EhRho1 was deter-mined by the kinetic analysis of nucleotide exchangefrom GDP to GTP in these small GTPases. These resultsindicate that the rates of exchange of EhRacG andEhRho1 promoted by EhGEF1 are very similar. Themultifunctional activity detected in EhGEF1 was
Thus, it was reported that human guanine nucleotideexchange factor T (GEFT) is able to activate Rac andalso Cdc42, which suggests that GEFT is involved inseveral cellular functions such as morphology, prolifera-tion, and cell transformation (Guo et al., 2003).
Confocal laser microscopy studies with trophozoitesstimulated with ConA showed that EhGEF1 and its pri-mary target EhRacG were both located at a well-deWnedrear zone of membrane accumulation known as the

A. Aguilar-Rojas et al. / Experimental Parasitology 109 (2005) 150–162 161
uroid (Guillén et al., 1998). The results of this study giveevidence that both EhGEF1 and EhRacG proteins wereco-located during uroid formation, thus suggesting thatthe two proteins participate jointly in this cellular pro-cess. This correlates with the guanine nucleotideexchange assays in which it was demonstrated that,EhRacG was the principal target for EhGEF1 protein;however, the necessary tools to demonstrate that theseinteractions exist in vivo are being developed at themoment.
Previously was shown that 1.3-fold EhRacG-V12overexpression produced defects in F-actin polymeriza-tion (Guillén et al., 1998). The 3.0-fold overexpression ofEhGEF1 also had a negative eVect at diVerent levels,such as F-actin content, cellular migration, and decreaseof cell damage ability of E. histolytica. Therefore, over-expression of EhGEF1 generates down-regulated phe-notypes in E. histolytica; this suggests that EhGEF1 aswell as its target GTPase play an important role in theorchestration of actin cytoskeleton organization, aVect-ing cellular functions such as motility and consequentlythe pathogenic mechanisms that are closely related tocytoskeletal integrity in trophozoites. Taken together,these data show that EhGEF1 and EhRacG play impor-tant roles in the pathogenic activity of E. histolytica.
The mechanism by EhGEF1 overproduction aVectcytoskeleton is unknown. Therefore, it has been shownthat F-actin organization can be regulated by severalproteins interactions. Recently, it was shown that homo-logues in E. histolytica to PAK proteins EhPAK are ableto bind HsRac1 protein which share 99% of homologyto EhRacG and also overproduction of its proteinaVects cytoskeleton integrity and down-regulated motil-ity (Labruyère et al., 2002). Moreover, it has been shownthat F-actin organization can be regulated by proteinrelations such as GEF-PAK, as it has been revealed forother GEFs in high eukaryotic cells, such as PDZ-RhoGEF protein, which is down-regulated by an over-expression of PAK4 kinase. This protein can bind toPDZRhoGEF favoring its phosphorylation and pro-moting an inadequate formation of actin stress Wbers inresponse to serum or LPA stimulation (Barac et al.,2004). Therefore, it is being proposed that in E. histoly-tica there may exist a similar mechanism to that of higheukaryotic cells, through which EhPAK or another PAKkinase from E. histolytica may regulate the guaninenucleotide exchange activity of EhGEF1 on EhRacGand turn oV its cellular function. This process requiresthe combined participation of EhRacG and high levelsof the overexpressed EhGEF1 protein, which aVects theactivity of diVerent eVector molecules, and therefore reg-ulates diVerent cellular processes. In this case, bothEhGEF1 and EhRacG proteins were located at theuroid region, corroborating the relationship betweenthese two proteins and perhaps forming the necessarycomplex to be regulated and therefore promoting a
change in F-actin polymerization which, in turn, maypromote a decrease in the cellular events previouslymentioned.
Further studies will be directed to dissecting the regu-latory eVects outside and inside the DH and PH domainsso as to modulate the rate of nucleotide exchange ofEhGEF1 and to understand the complex role of GEFs,GTPases, and their eVector molecules such as EhPAK,their relation with regulation, and in fundamental cellu-lar activities of E. histolytica, such as in the metastaticand invasive behavior of this parasite.
Acknowledgments
This work was supported by grants from CONACyT(Mexico) (No. 39511-A1) and European CommissionResearch Programs (ICA4-CT-2001-10073). A. Aguilar-Rojas is grateful for a doctoral fellowship from CONA-CyT (Exp. No. 129538) and also with Bruce and BettyAlberts, Frank R. Lillie, PWzer Inc. foundations, for anInternational Fellowship and Marine Biological Labora-tory. R.C. Maroun is a research scientist of the InstitutNational de la Santé et de la Recherche Médicale(INSERM). Authors are grateful to Dr. Alejandro Sosafor laboratory facilities.
References
Abe, K., Rossman, K.L., Liu, B., Ritola, K.D., Chiang, D., Campbel,S.L., et al., 2000. VAV2 is an activator of Cdc42, Rac1, and RhoA.The Journal of Biological Chemistry 275, 10141–10149.
Aghazadeh, B., Zhu, K., Kubiseski, T.J., Liu, G.A., Pawson, T., Zheng,Y., Rosen, M.K, 1998. Structure and mutagenesis of the Dblhomology domain. Nature Structural Biology 5, 1007–1098.
Altschul, S.F., Madden, T.L., SchaVer, A.A., Zhang, J., Zhang, Z.,Miller, W., Lipman, D.J., 1997. Gapped BLAST and PSI-BLAST: anew generation of protein database search programs. Nucleic AcidsResearch 25, 2402–3389.
Barac, A., Basile, J., Vazquez-Prado, J., Gao, Y., Zheng, Y., Gutkind, S.,2004. Direct interaction of PAK4 with PDZ-RhoGEF, a G protein-linked Rho guanine exchange factor. The Journal of BiologicalChemistry 279, 6182–6189.
Bishop, A.L., Hall, A., 2000. RhoGTPases and their eVector proteins.The Biochemical Journal 348, 241–255.
Boguski, M.S., McCormick, F., 1993. Proteins regulating Ras and itsrelatives. Nature 366, 643–654.
Bradford, M., 1976. A rapid and sensitive method for the quantitationof microgram quantities of protein dye binding. Analytical Bio-chemistry 72, 248–254.
Burridge, K., Wennerberg, K., 2004. Rho and Rac take center stage.Cell 166, 167–179.
Bustelo, X., 2000. Regulatory and signalling properties of the Vav fam-ily. Molecular and Cellular Biology 20, 1461–1477.
Capsoni, F., Minonzio, F., Ongari, A.M., Zanussi, C., 1989. A new sim-pliWed single-Wlter assay for “in vitro” evaluation of chemotaxis of51Cr-labeled polymorphonuclear leukocytes. Journal of Immuno-logical Methods 120, 125–131.
Cerione, R.A., Zheng, Y., 1996. The Dbl family of oncogenes. CurrentOpinion in Cell Biology 8, 216–222.

162 A. Aguilar-Rojas et al. / Experimental Parasitology 109 (2005) 150–162
CherWls, J., Chardin, P., 1999. GEFs: structural basis for their activa-tion of small GTP-binding proteins. Trends in Biochemical Sci-ences 24, 306–311.
Clark, C.G., Diamond, L.S., 1991. Ribosomal RNA genes of patho-genic and non-pathogenic Entamoeba histolytica are distinct.Molecular and Biochemical Parasitology 49, 297–302.
Collard, J., 1996. Signalling pathways regulated by rho proteins: possi-ble role in tumor formation and metastasis. International Journalof Oncology 8, 131–138.
Diamond, L.S., Harlow, D.R., Cunnick, C.C., 1978. A new medium forthe axenic cultivation of Entamoeba histolytica. Transactions of theRoyal Society of Tropical Medicine and Hygiene 72, 431–432.
Erickson, J.W., Cerione, R.A., 2004. Structural elements, mechanism,and evolutionary convergence of Rho protein-guanine nucleotideexchange factor complexes. Biochemistry 43, 837–842.
Eva, A., Vecchio, G., Rao, C.D., Tronick, S.R., Aaronson, S.A., 1998.The predicted DBL oncogenes product deWnes a distinct class oftransforming proteins. Proceedings of the National Academy ofSciences of the United States of America 85, 2061–2065.
Franco, E., Vazquez-Prado, J., Meza, I., 1997. Fibronectin-derivedfragments as inducers of adhesion and chemotaxis of Entamoebahistolytica trophozoites. The Journal of Infectious Diseases 176,1597–1602.
Garcia-Rivera, G., Rodriguez, M.A., Arroyo, R., Gonzáles-Robles, A.,Orozco, E., 1999. Entamoeba histolytica: a novel cysteine proteaseand an adhesin from the 112 kDa surface protein. MolecularMicrobiology 33, 556–568.
Ghosh, S.K., Samuelson, J., 1997. Involvement of p21racA, phosphoino-sitide 3-kinase, and vacuolar ATPase in phagocytosis of bacteriaand erythrocytes by Entamoeba histolytica: suggestive evidence forcoincidental evolution of amebic invasiveness. Infection and Immu-nity 65, 4243–4249.
Guillén, N., Boquet, P., Sansonetti, P., 1998. The small GTP-bindingprotein RacG regulates uroid formation in the protozoan parasiteEntamoeba histolytica. Journal of Cell Science 111, 1729–1739.
Guo, X., StaVord, L.J., Bryan, B., Xia, C., Ma, W., Wu, X., et al., 2003.A rac/Cdc42-speciWc exchange factor, GEFT, induces cell prolifera-tion, transformation and migration. The Journal of BiologicalChemistry 278, 13207–13215.
Hamann, L., Nickel, R., Tannich, E., 1995. Transfection and continu-ous expression of heterologous genes in the protozoan parasite Ent-amoeba histolytica. Proceedings of the National Academy ofSciences of the United States of America 92, 8975–8979.
Labruyère, E., Zimmer, C., Galy, V., Olivo-Marin, J.C., Guillén, N.,2002. EhPAK, a member of the p21-activated kinase family isinvolved in the control of Entamoeba histolytica migration andphagocytosis. Journal of Cell Science 116, 61–71.
Laemmli, U.K., 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 227, 680–685.
Lohia, A., Samuelson, J., 1993. Molecular cloning of a rho family geneof Entamoeba histolytica. Molecular and Biochemical Parasitology58, 177–180.
Lohia, A., Samuelson, J., 1996. Heterogeneity of Entamoeba histolyticarac genes encoding p21rac homologues. Gene 173, 205–208.
Mackay, D.J., Hall, A., 1998. RhoGTPases. The Journal of BiologicalChemistry 273, 20685–20688.
Menu, E., Braet, F., Timmers, M., Van, R.I., Van, C.B., Vanderkerken,K., 2002. The F-actin content of multiple myeloma cells as a mea-surement of their migration. Annals of the New York Academy ofSciences 973, 124–134.
Pasteris, N.G., Cadle, A., Logie, L.J., Porteous, M.E.M., Schwartz, C.E.,Stevenson, R.E., et al., 1994. Isolation and characterization of thefaciogenital dysplasia (Aarskog-Scott syndrome) gene: a putativerho/rac guanine nucleotide exchange factor. Cell 79, 669–678.
Rossman, K.L., Cheng, L., Mahon, G.M., Rojas, R.J., Snyder, J.T.,Whitehead, I.P., Sondek, J., 2003. Multifunctional roles for the PHdomain of Dbs in regulating Rho GTPase activation. The Journalof Biological Chemistry 278, 18393–18400.
Sambrook, J., Russell, D.W., 2001. Molecular Cloning. A LaboratoryManual. Cold Spring Harbor Laboratory Press, Cold Spring Har-bor, USA.
Schmitz, A.A.P., Govek, E.E., Bottner, B., Aelst, L.V., 2000. Rho GTP-ases: signaling, migration, and invasion. Experimental CellResearch 261, 1–12.
Shen, P.S., Lohia, A., Samuelson, J., 1994. Molecular cloning of ras andrap genes from Entamoeba histolytica. Molecular and BiochemicalParasitology 64, 111–120.
Sigrist, C.J.A., Cerutti, L., Hulo, N., Gattiker, A., Falquet, L., Pagni,M., et al., 2002. PROSITE: a documented database using patternsand proWles as motif descriptors. BrieWngs in Bioinformatics 3,265–274.
Snyder, J.T., Worthylake, D.K., Rossman, K.L., Betts, L., Pruitt, W.M.,Siderovski, D.P., Der, C.J., Sondek, J., 2002. Structural basis for theselective activation of Rho GTPases by Dbl exchange factors.Nature Structural Biology 9, 468–475.
Tavares, P., Sansonetti, P., Guillén, N., 2000. Cell polarization andadhesion in a motile pathogenic protozoan: role and fate of theEntamoeba histolytica Gal/GalNAc lectin. Microbes and Infection/Institut Pasteur 2, 643–649.
Thompson, J.D., Higgins, D.G., Gibson, T.J., 1994. CLUSTAL W:improving the sensitivity of progressive multiple sequencealignment through sequence weighing, position speciWc gap penal-ties and weight matrix choice. Nucleic Acids Research 22, 4673–4680.
Towbin, H., Stachelin, T., Gordon, J., 1979. Electrophoretic transfer ofproteins from polyacrylamide gels to nitrocellulose sheets:procedures and some applications. Proceedings of the NationalAcademy of Sciences of the United States of America 76, 4350–4354.
Van, A.L., Souza, S.C., 1997. RhoGTPases and signalling networks.Genes and Development 11, 2295–2322.
Whitehead, I.P., Campbell, S., Rossman, K.L., Der, C.J., 1997. Dbl fam-ily proteins. Biochimica et Biophysica Acta 1332, F1–F23.
Whitehead, I.P., Kirk, H., Kay, R., 1995. Retroviral transductionand oncogenic selection of a cDNA encoding Dbs, a homolog ofthe Dbl guanine nucleotide exchange factor. Oncogene 10, 713–721.
Whitehead, I.P., Lambert, Q.T., Glaven, J.A., Abe, K., Rossman, K.L.,Mahon, G.M., et al., 1999. Dependence of Dbl and Dbs transforma-tion on MEK and NF-kappaB activation. Molecular and CellularBiology 19, 7759–7770.
Wilkinson, P.C., 1998. Micropore Wlter methods for leukocyte chemo-taxis. Methods in Enzymology 162, 38–50.
Zheng, Y., Hart, M.J., Cerione, R.A., 1995. Guanine nucleotideexchange catalyzed by dbl oncogene product. Methods in Enzymol-ogy 256, 77–84.
Zheng, Y., Fischer, D.J., Tigyi, G., Pasteris, N.G., Gorski, J.L., Xu, Y.,1996. The faciogenital dysplasia gene product FGD1 functions as aCdc42Hs-speciWc guanine-nucleotide exchange factor. The Journalof Biological Chemistry 271, 33169–33172.
Copyright © 2022 FDOKUMEN




















