
The Legionella pneumophila effector protein DrrA is a Rab1 guanine nucleotide-exchange factor
19
LETTERS NATURE CELL BIOLOGY VOLUME 8 | NUMBER 9 | SEPTEMBER 2006 971 The Legionella pneumophila effector protein DrrA is a Rab1 guanine nucleotide-exchange factor Takahiro Murata 1,4 , Anna Delprato 2 , Alyssa Ingmundson 1 , Derek K. Toomre 3 , David G. Lambright 2 and Craig R. Roy 1,5 The intracellular pathogen Legionella pneumophila avoids fusion with lysosomes and subverts membrane transport from the endoplasmic reticulum to create an organelle that supports bacterial replication 1,2 . Transport of endoplasmic reticulum- derived vesicles to the Legionella-containing vacuole (LCV) requires bacterial proteins that are translocated into host cells by a type IV secretion apparatus called Dot/Icm 3–7 . Recent observations have revealed recruitment of the host GTPase Rab1 to the LCV by a process requiring the Dot/Icm system 8,9 . Here, a visual screen was used to identify L. pneumophila mutants with defects in Rab1 recruitment. One of the factors identified in this screen was DrrA, a new Dot/Icm substrate protein translocated into host cells. We show that DrrA is a potent and highly specific Rab1 guanine nucleotide-exchange factor (GEF). DrrA can disrupt Rab1-mediated secretory transport to the Golgi apparatus by competing with endogenous exchange factors to recruit and activate Rab1 on plasma membrane-derived organelles. These data establish that intracellular pathogens have the capacity to directly modulate the activation state of a specific member of the Rab family of GTPases and thus further our understanding of the mechanisms used by bacterial pathogens to manipulate host vesicular transport. The collective activities of proteins injected into host cells by the Dot/Icm system are essential for successful transport of L. pneumophila to the endoplasmic reticulum; however, the identity and biochemical activities of the individual Dot/Icm substrate proteins controlling LCV transport remain largely unknown. In addition to these bacterial proteins, cell biological studies have revealed that several host GTPases regulate bio- genesis of the organelle that supports L. pneumophila replication 2,8–11 . The coordinated activities of the GTPases Sar1 and Arf are important for transport and fusion of endoplasmic reticulum-derived vesicles to the LCV 2,10,11 . To date, the only L. pneumophila protein delivered into host cells by the Dot/Icm system for which a biochemical function has been demonstrated is RalF, which is a bacterial member of the Sec7 family of Arf GEFs 5,12 . Recent studies have shown that the host GTPase Rab1 is recruited to the LCV by a Dot/Icm-dependent mechanism 8,9 . Inhibition of Rab1 function delays the kinetics of endoplasmic reticulum-vesicle recruitment to the LCV and impairs intracellular replication of L. pneu- mophila 8 . Although there are at least 38 functionally distinct Rab protein family members in the human genome that regulate events important for the movement and fusion of vesicles through the endocytic and secre- tory pathways 13–16 , Rab1 is the only Rab protein that has been identi- fied on the LCV membrane. Although it is attractive to speculate that intracellular pathogens may subvert host vesicular transport pathways by directly manipulating the function of host Rab proteins, bacterial proteins that specifically alter the activities of host Rab GTPases have not been identified. A visual screen was conducted to identify L. pneumophila mutants with defects in Rab1 recruitment. A total of 3,948 independently-derived L. pneumophila transposon mutants were individually examined following infection of macrophages and 113 mutants were isolated that showed impaired Rab1 recruitment to the LCV (see Supplementary Information, Table S1). This screen isolated a total of 45 transposon insertions in genes encoding different Dot and Icm proteins necessary for type IV secretion system function, 45 mutants that had a partial defect in their ability to recruit Rab1 to the LCV and 19 mutants that had a severe Rab1 recruit- ment defect. It was determined that 10 of the 19 mutants with a severe Rab1 recruitment defect also had impaired Dot/Icm transporter func- tion, suggesting that elimination of these gene products was not directly responsible for the Rab1 recruitment defect. The nine remaining trans- poson insertions were in eight different genes that were not essential for Dot/Icm function (Table 1), making these genes potential candidates for encoding Dot/Icm substrate proteins important for Rab1 recruitment. To determine whether any of the genes listed in Table 1 encode pro- teins that are translocated into host cells by the Dot/Icm system, and thus have the potential to be directly involved in Rab1 recruitment to the LCV, the calmodulin-dependent adenylate cyclase domain from the 1 Section of Microbial Pathogenesis, Yale University School of Medicine, Boyer Center for Molecular Medicine, 295 Congress Avenue, New Haven, CT 06536, USA. 2 Department of Molecular Medicine, UMASS Medical School Two Biotech, 373 Plantation Street, Worcester, MA 01605, USA. 3 Department of Cell Biology, Yale University School of Medicine, Boyer Center for Molecular Medicine, 295 Congress Avenue, New Haven, CT 06536, USA. 4 Current address: Department of Pediatrics, Mizonokuchi Hospital, Teikyo University School of Medicine, 3-8-3 Mizonokuchi, Takatsuku, Kawasaki 213-8507, Japan. 5 Correspondence should be addressed to C.R.R. (e-mail: [email protected]) Received 5 June 2006; accepted 10 July 2006; published online 13 August 2006; DOI: 10.1038/ncb1463 Nature Publishing Group ©2006
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The Legionella pneumophila effector protein DrrA is a Rab1 guanine nucleotide-exchange factor

L E T T E R S
NATURE CELL BIOLOGY VOLUME 8 | NUMBER 9 | SEPTEMBER 2006 971
The Legionella pneumophila effector protein DrrA is a Rab1 guanine nucleotide-exchange factorTakahiro Murata1,4, Anna Delprato2, Alyssa Ingmundson1, Derek K. Toomre3, David G. Lambright2 and Craig R. Roy1,5
The intracellular pathogen Legionella pneumophila avoids fusion with lysosomes and subverts membrane transport from the endoplasmic reticulum to create an organelle that supports bacterial replication1,2. Transport of endoplasmic reticulum-derived vesicles to the Legionella-containing vacuole (LCV) requires bacterial proteins that are translocated into host cells by a type IV secretion apparatus called Dot/Icm3–7. Recent observations have revealed recruitment of the host GTPase Rab1 to the LCV by a process requiring the Dot/Icm system8,9. Here, a visual screen was used to identify L. pneumophila mutants with defects in Rab1 recruitment. One of the factors identified in this screen was DrrA, a new Dot/Icm substrate protein translocated into host cells. We show that DrrA is a potent and highly specific Rab1 guanine nucleotide-exchange factor (GEF). DrrA can disrupt Rab1-mediated secretory transport to the Golgi apparatus by competing with endogenous exchange factors to recruit and activate Rab1 on plasma membrane-derived organelles. These data establish that intracellular pathogens have the capacity to directly modulate the activation state of a specific member of the Rab family of GTPases and thus further our understanding of the mechanisms used by bacterial pathogens to manipulate host vesicular transport.
The collective activities of proteins injected into host cells by the Dot/Icm system are essential for successful transport of L. pneumophila to the endoplasmic reticulum; however, the identity and biochemical activities of the individual Dot/Icm substrate proteins controlling LCV transport remain largely unknown. In addition to these bacterial proteins, cell biological studies have revealed that several host GTPases regulate bio-genesis of the organelle that supports L. pneumophila replication2,8–11. The coordinated activities of the GTPases Sar1 and Arf are important for transport and fusion of endoplasmic reticulum-derived vesicles to the LCV2,10,11. To date, the only L. pneumophila protein delivered into host cells by the Dot/Icm system for which a biochemical function has been
demonstrated is RalF, which is a bacterial member of the Sec7 family of Arf GEFs5,12. Recent studies have shown that the host GTPase Rab1 is recruited to the LCV by a Dot/Icm-dependent mechanism8,9. Inhibition of Rab1 function delays the kinetics of endoplasmic reticulum-vesicle recruitment to the LCV and impairs intracellular replication of L. pneu-mophila8. Although there are at least 38 functionally distinct Rab protein family members in the human genome that regulate events important for the movement and fusion of vesicles through the endocytic and secre-tory pathways13–16, Rab1 is the only Rab protein that has been identi-fied on the LCV membrane. Although it is attractive to speculate that intracellular pathogens may subvert host vesicular transport pathways by directly manipulating the function of host Rab proteins, bacterial proteins that specifically alter the activities of host Rab GTPases have not been identified.
A visual screen was conducted to identify L. pneumophila mutants with defects in Rab1 recruitment. A total of 3,948 independently-derived L. pneumophila transposon mutants were individually examined following infection of macrophages and 113 mutants were isolated that showed impaired Rab1 recruitment to the LCV (see Supplementary Information, Table S1). This screen isolated a total of 45 transposon insertions in genes encoding different Dot and Icm proteins necessary for type IV secretion system function, 45 mutants that had a partial defect in their ability to recruit Rab1 to the LCV and 19 mutants that had a severe Rab1 recruit-ment defect. It was determined that 10 of the 19 mutants with a severe Rab1 recruitment defect also had impaired Dot/Icm transporter func-tion, suggesting that elimination of these gene products was not directly responsible for the Rab1 recruitment defect. The nine remaining trans-poson insertions were in eight different genes that were not essential for Dot/Icm function (Table 1), making these genes potential candidates for encoding Dot/Icm substrate proteins important for Rab1 recruitment.
To determine whether any of the genes listed in Table 1 encode pro-teins that are translocated into host cells by the Dot/Icm system, and thus have the potential to be directly involved in Rab1 recruitment to the LCV, the calmodulin-dependent adenylate cyclase domain from the
1Section of Microbial Pathogenesis, Yale University School of Medicine, Boyer Center for Molecular Medicine, 295 Congress Avenue, New Haven, CT 06536, USA. 2Department of Molecular Medicine, UMASS Medical School Two Biotech, 373 Plantation Street, Worcester, MA 01605, USA. 3Department of Cell Biology, Yale University School of Medicine, Boyer Center for Molecular Medicine, 295 Congress Avenue, New Haven, CT 06536, USA. 4Current address: Department of Pediatrics, Mizonokuchi Hospital, Teikyo University School of Medicine, 3-8-3 Mizonokuchi, Takatsuku, Kawasaki 213-8507, Japan.5Correspondence should be addressed to C.R.R. (e-mail: [email protected])
Received 5 June 2006; accepted 10 July 2006; published online 13 August 2006; DOI: 10.1038/ncb1463
print_ncb1463.indd 971print_ncb1463.indd 971 10/8/06 12:05:46 pm10/8/06 12:05:46 pm
Nature Publishing Group ©2006
972 NATURE CELL BIOLOGY VOLUME 8 | NUMBER 9 | SEPTEMBER 2006
L E T T E R S
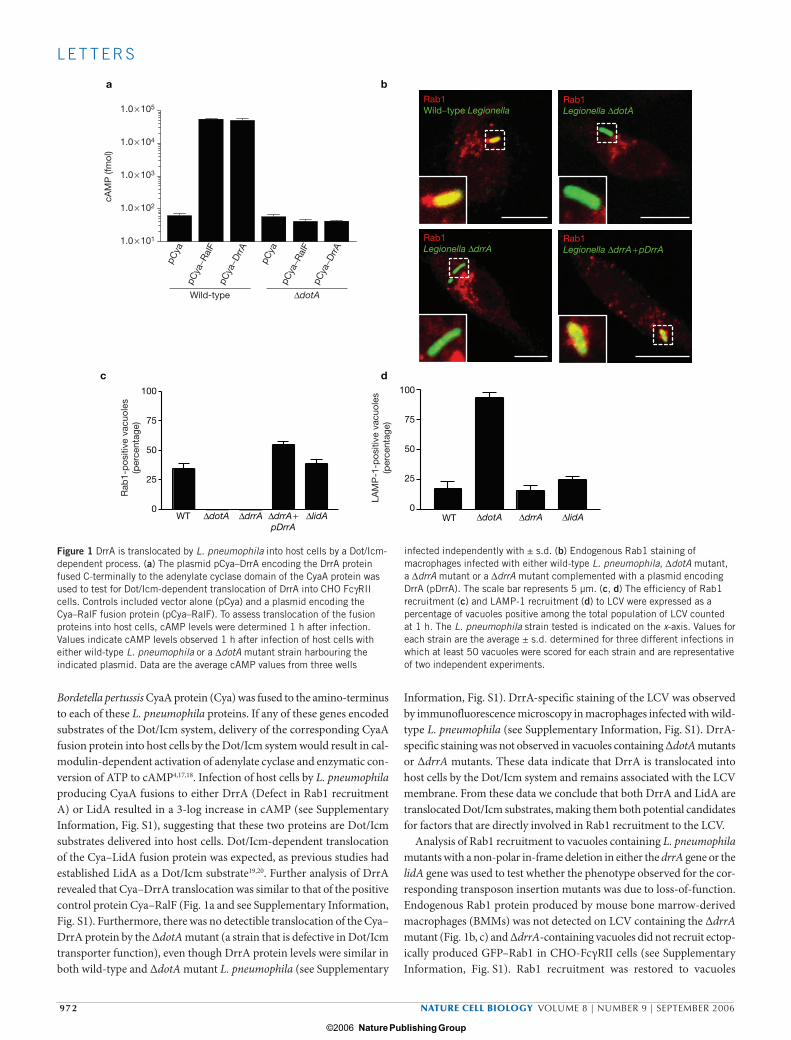
Bordetella pertussis CyaA protein (Cya) was fused to the amino-terminus to each of these L. pneumophila proteins. If any of these genes encoded substrates of the Dot/Icm system, delivery of the corresponding CyaA fusion protein into host cells by the Dot/Icm system would result in cal-modulin-dependent activation of adenylate cyclase and enzymatic con-version of ATP to cAMP4,17,18. Infection of host cells by L. pneumophila producing CyaA fusions to either DrrA (Defect in Rab1 recruitment A) or LidA resulted in a 3-log increase in cAMP (see Supplementary Information, Fig. S1), suggesting that these two proteins are Dot/Icm substrates delivered into host cells. Dot/Icm-dependent translocation of the Cya–LidA fusion protein was expected, as previous studies had established LidA as a Dot/Icm substrate19,20. Further analysis of DrrA revealed that Cya–DrrA translocation was similar to that of the positive control protein Cya–RalF (Fig. 1a and see Supplementary Information, Fig. S1). Furthermore, there was no detectible translocation of the Cya–DrrA protein by the ∆dotA mutant (a strain that is defective in Dot/Icm transporter function), even though DrrA protein levels were similar in both wild-type and ∆dotA mutant L. pneumophila (see Supplementary
Information, Fig. S1). DrrA-specific staining of the LCV was observed by immunofluorescence microscopy in macrophages infected with wild-type L. pneumophila (see Supplementary Information, Fig. S1). DrrA-specific staining was not observed in vacuoles containing ∆dotA mutants or ∆drrA mutants. These data indicate that DrrA is translocated into host cells by the Dot/Icm system and remains associated with the LCV membrane. From these data we conclude that both DrrA and LidA are translocated Dot/Icm substrates, making them both potential candidates for factors that are directly involved in Rab1 recruitment to the LCV.
Analysis of Rab1 recruitment to vacuoles containing L. pneumophila mutants with a non-polar in-frame deletion in either the drrA gene or the lidA gene was used to test whether the phenotype observed for the cor-responding transposon insertion mutants was due to loss-of-function. Endogenous Rab1 protein produced by mouse bone marrow-derived macrophages (BMMs) was not detected on LCV containing the ∆drrA mutant (Fig. 1b, c) and ∆drrA-containing vacuoles did not recruit ectop-ically produced GFP–Rab1 in CHO-FcγRII cells (see Supplementary Information, Fig. S1). Rab1 recruitment was restored to vacuoles
aRab1Wild−type Legionella
Rab1Legionella ∆dotA
Rab1Legionella ∆drrA
Rab1Legionella ∆drrA + pDrrA
c100
75
50
25
0
100
75
50
25
0WT ∆dotA ∆drrA ∆drrA +
pDrrA∆lidA WT ∆dotA ∆drrA ∆lidA
d
b
1.0 × 105
1.0 × 104
1.0 × 103
1.0 × 102
1.0 × 101
cAM
P (f
mol
)
pCya
pCya
−Ral
FpC
ya−D
rrA
pCya
pCya
−Ral
FpC
ya−D
rrA
Wild-type ∆dotA
Rab
1-p
ositi
ve v
acuo
les
(per
cent
age)
LAM
P-1
-pos
itive
vac
uole
s(p
erce
ntag
e)
Figure 1 DrrA is translocated by L. pneumophila into host cells by a Dot/Icm-dependent process. (a) The plasmid pCya–DrrA encoding the DrrA protein fused C-terminally to the adenylate cyclase domain of the CyaA protein was used to test for Dot/Icm-dependent translocation of DrrA into CHO FcγRII cells. Controls included vector alone (pCya) and a plasmid encoding the Cya–RalF fusion protein (pCya–RalF). To assess translocation of the fusion proteins into host cells, cAMP levels were determined 1 h after infection. Values indicate cAMP levels observed 1 h after infection of host cells with either wild-type L. pneumophila or a ∆dotA mutant strain harbouring the indicated plasmid. Data are the average cAMP values from three wells
infected independently with ± s.d. (b) Endogenous Rab1 staining of macrophages infected with either wild-type L. pneumophila, ∆dotA mutant, a ∆drrA mutant or a ∆drrA mutant complemented with a plasmid encoding DrrA (pDrrA). The scale bar represents 5 µm. (c, d) The efficiency of Rab1 recruitment (c) and LAMP-1 recruitment (d) to LCV were expressed as a percentage of vacuoles positive among the total population of LCV counted at 1 h. The L. pneumophila strain tested is indicated on the x-axis. Values for each strain are the average ± s.d. determined for three different infections in which at least 50 vacuoles were scored for each strain and are representative of two independent experiments.
print_ncb1463.indd 972print_ncb1463.indd 972 10/8/06 12:05:50 pm10/8/06 12:05:50 pm
Nature Publishing Group ©2006
NATURE CELL BIOLOGY VOLUME 8 | NUMBER 9 | SEPTEMBER 2006 973
L E T T E R S
containing the ∆drrA mutant complemented with a plasmid encoding drrA (Fig. 1b, c), confirming the importance of DrrA for recruitment of Rab1. No defect in Rab1 recruitment was observed for LCV containing the ∆lidA mutants (Fig. 1c), suggesting that the Rab1 recruitment defect observed for the five different lidA transposon insertion mutants identi-fied in this study resulted from the activities of truncated lidA products produced by these mutants. These data establish that drrA is important for Rab1 recruitment to the LCV.
Whether drrA and lidA were required for L. pneumophila evasion of endocytic maturation was determined by measuring the proportion of LCV containing ∆drrA and ∆lidA mutants that acquired the late endo-somal protein LAMP-1 after infection of BMMs. There was no appar-ent defect in avoidance of endocytic maturation detected for either the ∆drrA or ∆lidA mutants (Fig. 1d). Additionally, no defect was observed when intracellular growth of the ∆drrA mutant was measured in BMMs and the protozoan host Acanthamoeba castellanii (see Supplementary Information, Fig. S1). Thus, the defect in Rab1 recruitment observed for the ∆drrA mutant is not due to an indirect effect caused by an altera-tion in trafficking of the LCV before Rab1 acquisition or an effect that might decrease the efficiency of translocation of other Dot/Icm substrates required for intracellular growth.
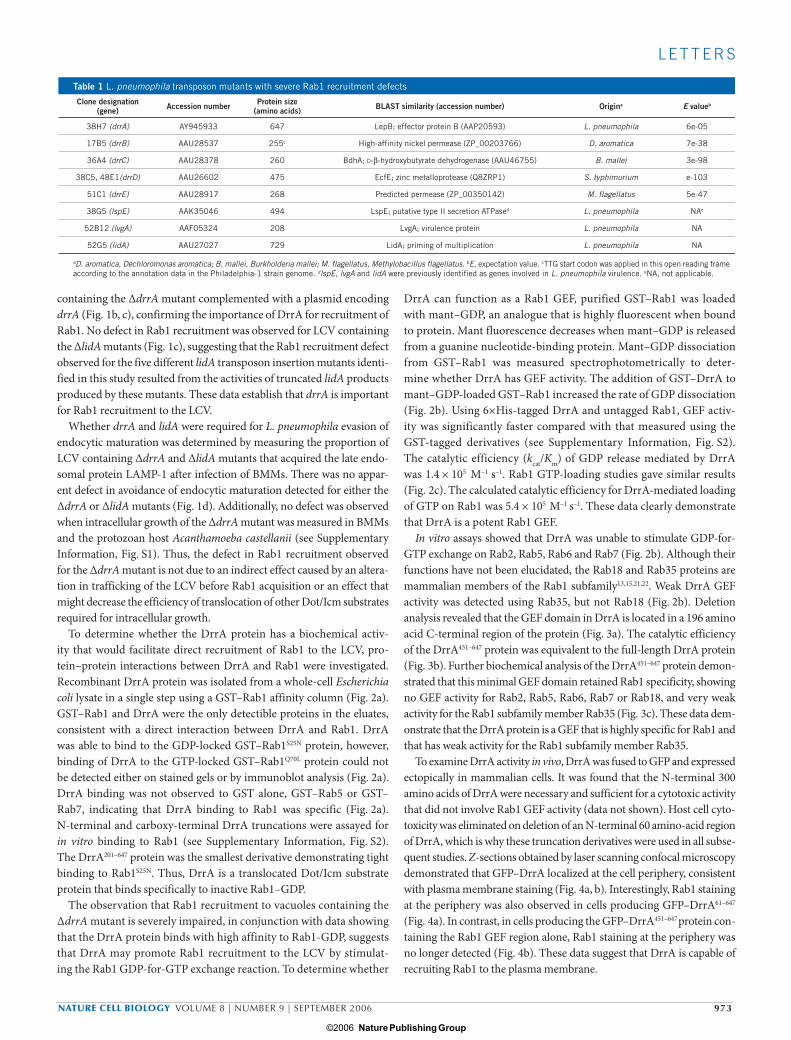
To determine whether the DrrA protein has a biochemical activ-ity that would facilitate direct recruitment of Rab1 to the LCV, pro-tein–protein interactions between DrrA and Rab1 were investigated. Recombinant DrrA protein was isolated from a whole-cell Escherichia coli lysate in a single step using a GST–Rab1 affinity column (Fig. 2a). GST–Rab1 and DrrA were the only detectible proteins in the eluates, consistent with a direct interaction between DrrA and Rab1. DrrA was able to bind to the GDP-locked GST–Rab1S25N protein, however, binding of DrrA to the GTP-locked GST–Rab1Q70L protein could not be detected either on stained gels or by immunoblot analysis (Fig. 2a). DrrA binding was not observed to GST alone, GST–Rab5 or GST–Rab7, indicating that DrrA binding to Rab1 was specific (Fig. 2a). N-terminal and carboxy-terminal DrrA truncations were assayed for in vitro binding to Rab1 (see Supplementary Information, Fig. S2). The DrrA201–647 protein was the smallest derivative demonstrating tight binding to Rab1S25N. Thus, DrrA is a translocated Dot/Icm substrate protein that binds specifically to inactive Rab1–GDP.
The observation that Rab1 recruitment to vacuoles containing the ∆drrA mutant is severely impaired, in conjunction with data showing that the DrrA protein binds with high affinity to Rab1-GDP, suggests that DrrA may promote Rab1 recruitment to the LCV by stimulat-ing the Rab1 GDP-for-GTP exchange reaction. To determine whether
DrrA can function as a Rab1 GEF, purified GST–Rab1 was loaded with mant–GDP, an analogue that is highly fluorescent when bound to protein. Mant fluorescence decreases when mant–GDP is released from a guanine nucleotide-binding protein. Mant–GDP dissociation from GST–Rab1 was measured spectrophotometrically to deter-mine whether DrrA has GEF activity. The addition of GST–DrrA to mant–GDP-loaded GST–Rab1 increased the rate of GDP dissociation (Fig. 2b). Using 6×His-tagged DrrA and untagged Rab1, GEF activ-ity was significantly faster compared with that measured using the GST-tagged derivatives (see Supplementary Information, Fig. S2). The catalytic efficiency (kcat/Km) of GDP release mediated by DrrA was 1.4 × 105 M–1 s–1. Rab1 GTP-loading studies gave similar results (Fig. 2c). The calculated catalytic efficiency for DrrA-mediated loading of GTP on Rab1 was 5.4 × 105 M–1 s–1. These data clearly demonstrate that DrrA is a potent Rab1 GEF.
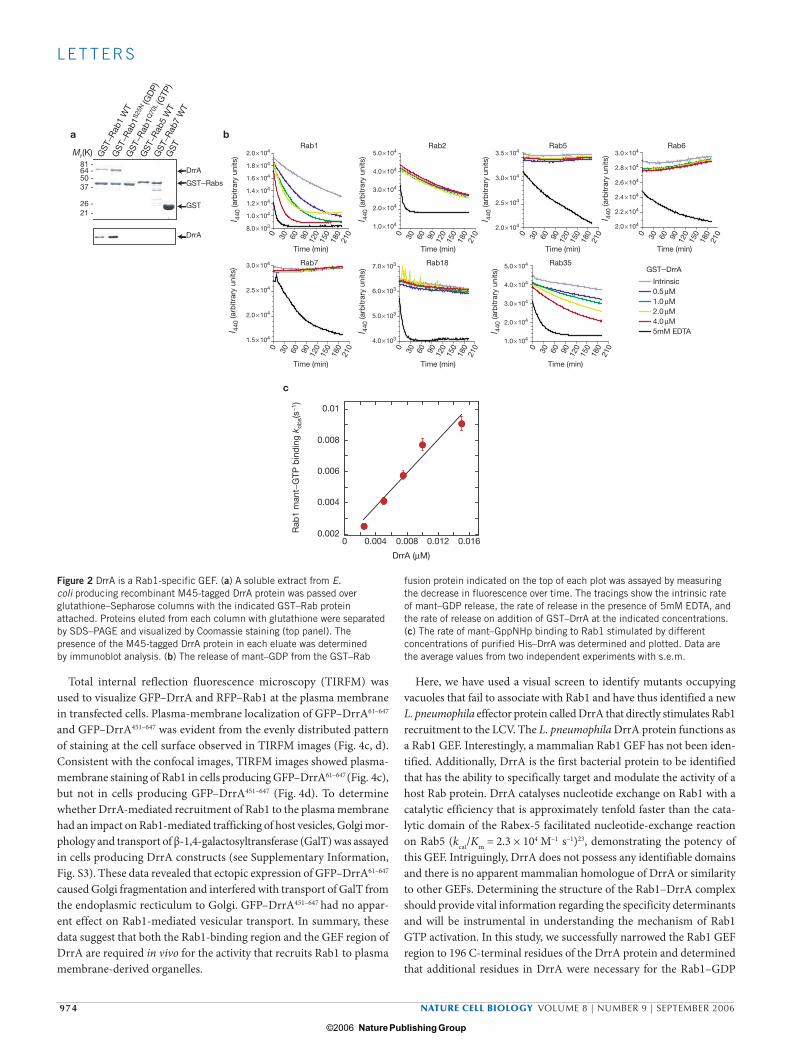
In vitro assays showed that DrrA was unable to stimulate GDP-for-GTP exchange on Rab2, Rab5, Rab6 and Rab7 (Fig. 2b). Although their functions have not been elucidated, the Rab18 and Rab35 proteins are mammalian members of the Rab1 subfamily13,15,21,22. Weak DrrA GEF activity was detected using Rab35, but not Rab18 (Fig. 2b). Deletion analysis revealed that the GEF domain in DrrA is located in a 196 amino acid C-terminal region of the protein (Fig. 3a). The catalytic efficiency of the DrrA451–647 protein was equivalent to the full-length DrrA protein (Fig. 3b). Further biochemical analysis of the DrrA451–647 protein demon-strated that this minimal GEF domain retained Rab1 specificity, showing no GEF activity for Rab2, Rab5, Rab6, Rab7 or Rab18, and very weak activity for the Rab1 subfamily member Rab35 (Fig. 3c). These data dem-onstrate that the DrrA protein is a GEF that is highly specific for Rab1 and that has weak activity for the Rab1 subfamily member Rab35.
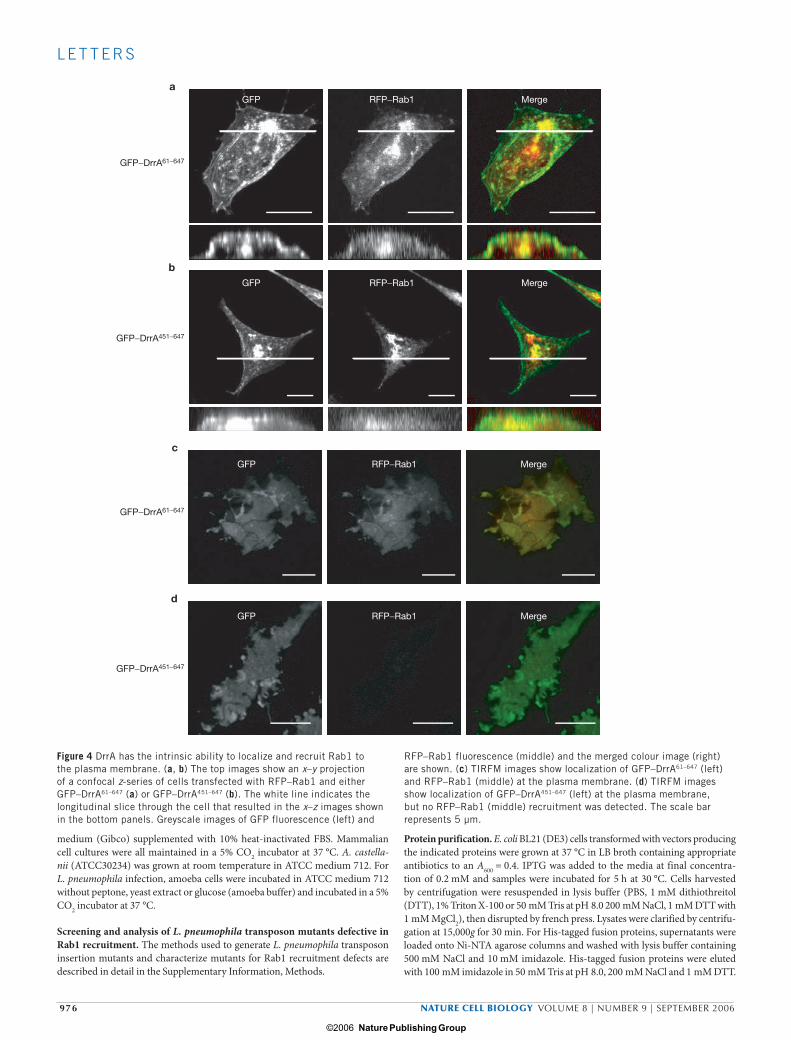
To examine DrrA activity in vivo, DrrA was fused to GFP and expressed ectopically in mammalian cells. It was found that the N-terminal 300 amino acids of DrrA were necessary and sufficient for a cytotoxic activity that did not involve Rab1 GEF activity (data not shown). Host cell cyto-toxicity was eliminated on deletion of an N-terminal 60 amino-acid region of DrrA, which is why these truncation derivatives were used in all subse-quent studies. Z-sections obtained by laser scanning confocal microscopy demonstrated that GFP–DrrA localized at the cell periphery, consistent with plasma membrane staining (Fig. 4a, b). Interestingly, Rab1 staining at the periphery was also observed in cells producing GFP–DrrA61–647 (Fig. 4a). In contrast, in cells producing the GFP–DrrA451–647 protein con-taining the Rab1 GEF region alone, Rab1 staining at the periphery was no longer detected (Fig. 4b). These data suggest that DrrA is capable of recruiting Rab1 to the plasma membrane.
Table 1 L. pneumophila transposon mutants with severe Rab1 recruitment defects
Clone designation (gene) Accession number Protein size
(amino acids) BLAST similarity (accession number) Origina E valueb
38H7 (drrA) AY945933 647 LepB; effector protein B (AAP20593) L. pneumophila 6e-05
17B5 (drrB) AAU28537 255c High-affinity nickel permease (ZP_00203766) D. aromatica 7e-38
36A4 (drrC) AAU28378 260 BdhA; D-β-hydroxybutyrate dehydrogenase (AAU46755) B. mallei 3e-98
38C5, 48E1(drrD) AAU26602 475 EcfE; zinc metalloprotease (Q8ZRP1) S. typhimurium e-103
51C1 (drrE) AAU28917 268 Predicted permease (ZP_00350142) M. flagellatus 5e-47
38G5 (lspE) AAK35046 494 LspE; putative type II secretion ATPased L. pneumophila NAe
52B12 (lvgA) AAF05324 208 LvgA; virulence protein L. pneumophila NA
52G5 (lidA) AAU27027 729 LidA; priming of multiplication L. pneumophila NA
aD. aromatica, Dechloromonas aromatica; B. mallei, Burkholderia mallei; M. flagellatus, Methylobacillus flagellatus. bE, expectation value. cTTG start codon was applied in this open reading frame according to the annotation data in the Philadelphia-1 strain genome. dlspE, lvgA and lidA were previously identified as genes involved in L. pneumophila virulence. eNA, not applicable.
print_ncb1463.indd 973print_ncb1463.indd 973 10/8/06 12:05:52 pm10/8/06 12:05:52 pm
Nature Publishing Group ©2006
974 NATURE CELL BIOLOGY VOLUME 8 | NUMBER 9 | SEPTEMBER 2006
L E T T E R S
Total internal reflection fluorescence microscopy (TIRFM) was used to visualize GFP–DrrA and RFP–Rab1 at the plasma membrane in transfected cells. Plasma-membrane localization of GFP–DrrA61–647
and GFP–DrrA451–647 was evident from the evenly distributed pattern of staining at the cell surface observed in TIRFM images (Fig. 4c, d). Consistent with the confocal images, TIRFM images showed plasma-membrane staining of Rab1 in cells producing GFP–DrrA61–647 (Fig. 4c), but not in cells producing GFP–DrrA451–647 (Fig. 4d). To determine whether DrrA-mediated recruitment of Rab1 to the plasma membrane had an impact on Rab1-mediated trafficking of host vesicles, Golgi mor-phology and transport of β-1,4-galactosyltransferase (GalT) was assayed in cells producing DrrA constructs (see Supplementary Information, Fig. S3). These data revealed that ectopic expression of GFP–DrrA61–647 caused Golgi fragmentation and interfered with transport of GalT from the endoplasmic recticulum to Golgi. GFP–DrrA451–647 had no appar-ent effect on Rab1-mediated vesicular transport. In summary, these data suggest that both the Rab1-binding region and the GEF region of DrrA are required in vivo for the activity that recruits Rab1 to plasma membrane-derived organelles.
Here, we have used a visual screen to identify mutants occupying vacuoles that fail to associate with Rab1 and have thus identified a new L. pneumophila effector protein called DrrA that directly stimulates Rab1 recruitment to the LCV. The L. pneumophila DrrA protein functions as a Rab1 GEF. Interestingly, a mammalian Rab1 GEF has not been iden-tified. Additionally, DrrA is the first bacterial protein to be identified that has the ability to specifically target and modulate the activity of a host Rab protein. DrrA catalyses nucleotide exchange on Rab1 with a catalytic efficiency that is approximately tenfold faster than the cata-lytic domain of the Rabex-5 facilitated nucleotide-exchange reaction on Rab5 (kcat/Km = 2.3 × 104 M–1 s–1)23, demonstrating the potency of this GEF. Intriguingly, DrrA does not possess any identifiable domains and there is no apparent mammalian homologue of DrrA or similarity to other GEFs. Determining the structure of the Rab1–DrrA complex should provide vital information regarding the specificity determinants and will be instrumental in understanding the mechanism of Rab1 GTP activation. In this study, we successfully narrowed the Rab1 GEF region to 196 C-terminal residues of the DrrA protein and determined that additional residues in DrrA were necessary for the Rab1–GDP
DrrA
81 -
21 -26 -
37 -50 -64 -
GST−Rabs
GST
DrrA
a b
c
0.002
0.004
0.006
0.008
0.01
0 0.004 0.008 0.012 0.016
Mr(K) 2.0 × 104 5.0 × 104
1.0 × 104
2.0 × 104
3.0 × 104
4.0 × 104
5.0 × 104
1.0 × 104
2.0 × 104
3.0 × 104
4.0 × 104
3.0 × 104
2.0 × 104
2.2 × 104
2.4 × 104
2.6 × 104
2.8 × 1041.8 × 104
1.6 × 104
1.4 × 104
1.2 × 104
1.0 × 104
8.0 × 103
l 440
(arb
itrar
y un
its)
l 440
(arb
itrar
y un
its)
l 440
(arb
itrar
y un
its) 3.5 × 104
2.0 × 104
2.5 × 104
3.0 × 104
3.0 × 104
1.5 × 104
2.0 × 104
2.5 × 104
l 440
(arb
itrar
y un
its)
l 440
(arb
itrar
y un
its)7.0 × 103
4.0 × 103
5.0 × 103
6.0 × 103
l 440
(arb
itrar
y un
its)
l 440
(arb
itrar
y un
its)
0 30 60 90 120
150
180
210
0 30 60 90 120
150
180
210 0 30 60 90 120
150
180
210 0 30 60 90 120
150
180
210
0 30 60 90 120
150
180
210 0 30 60 90 120
150
180
210 0 30 60 90 120
150
180
210
Time (min) Time (min) Time (min) Time (min)
Time (min) Time (min) Time (min)
Rab1 Rab2 Rab5
Rab7 Rab18 Rab35
Rab6
GST−DrrA
Intrinsic0.5 µM1.0 µM2.0 µM4.0 µM5mM EDTA
Rab
1 m
ant−
GTP
bin
din
g k o
bs(
s−1 )
DrrA (µM)
GST
−Rab
1 W
TG
ST−R
ab1
S25N
(GD
P)
GST
−Rab
1Q
70L (
GTP
)
GST
−Rab
5 W
T
GST
−Rab
7 W
T
GST
Figure 2 DrrA is a Rab1-specific GEF. (a) A soluble extract from E. coli producing recombinant M45-tagged DrrA protein was passed over glutathione–Sepharose columns with the indicated GST–Rab protein attached. Proteins eluted from each column with glutathione were separated by SDS–PAGE and visualized by Coomassie staining (top panel). The presence of the M45-tagged DrrA protein in each eluate was determined by immunoblot analysis. (b) The release of mant–GDP from the GST–Rab
fusion protein indicated on the top of each plot was assayed by measuring the decrease in fluorescence over time. The tracings show the intrinsic rate of mant–GDP release, the rate of release in the presence of 5mM EDTA, and the rate of release on addition of GST–DrrA at the indicated concentrations. (c) The rate of mant–GppNHp binding to Rab1 stimulated by different concentrations of purified His–DrrA was determined and plotted. Data are the average values from two independent experiments with s.e.m.
print_ncb1463.indd 974print_ncb1463.indd 974 10/8/06 12:05:56 pm10/8/06 12:05:56 pm
Nature Publishing Group ©2006

NATURE CELL BIOLOGY VOLUME 8 | NUMBER 9 | SEPTEMBER 2006 975
L E T T E R S
binding activity detected in vitro. In vivo data indicate that the Rab1–GDP binding activity may facilitate DrrA-mediated recruitment of Rab1 to membranes, enabling DrrA to compete with endogenous Rab1 exchange factors for cytosolic pools of Rab1–GDP and activate Rab1 on plasma membrane-derived organelles.
During L. pneumophila infection, we hypothesize that active Rab1 on the LCV facilitates the recruitment and fusion of endoplasmic reticulum-derived vesicles with the LCV. Our data predict that con-version of the LCV from a plasma membrane-derived vacuole to an endoplasmic reticulum-derived organelle could account for the release of DrrA and Rab1 from the mature replicative vacuole, where DrrA and Rab1 may no longer provide beneficial functions, suggesting a role for membrane localization in temporal regulation of DrrA func-tion. In summary, our investigations on DrrA-mediated recruitment of Rab1 to the LCV provide a unique insight on a mechanism by which an intracellular pathogen can control the localization and activity of a mammalian Rab protein.
Note added in proof: While this manuscript was in press, a paper was published that showed that DrrA (SidM) has Rab1 GEF activity.27
METHODSBacterial strains, plasmids and media. All strains, plasmids and primers used in this study are listed the Supplementary Information, Table S1. L. pneumophila strains were grown on charcoal-yeast extract plates or in aces-buffered yeast extract broth as previously described24. Antibiotics were added to the media as necessary at the following concentrations: streptomycin, 100 µg ml–1; kanamycin, 10 µg ml—1; chloramphenicol, 10 µg ml–1. E. coli strains were cultivated in LB or 2YT media with antibiotics at the following concentrations: ampicillin, 100 µg ml–1; kanamycin, 30 µg ml–1; chloramphenicol, 20 µg ml–1.
Cell culture. Murine BMMs were prepared from female A/J mice as previously described25. Cells were replated in RPMI-1640 medium (Gibco, Carlsbad, CA) supplemented with 10% heat-inactivated fetal bovine serum (FBS; Gibco) and 5% conditioned medium from L929 cells. CHO cells producing the FcγRII protein (CHO FcγRII) were grown in minimal essential medium α (Gibco) containing 10% heat-inactivated FBS26. COS7 cells were grown in Dulbecco’s modified Eagle’s
GEF domainDrrA 647451
6471
1 647
5001
4481
2001
546 647
201 647
647451
201 647Rab1-binding domain
a
0.125 µM
0.016 µM
0.032 µM
0.064 µM
Time (min)
0 906030
Rab1
His−DrrA451-647
b
Rab6Rab5
1.0 × 1041.2 × 104
0.4 × 104
1.2 × 104
1.0 × 104
0.8 × 104
0.6 × 104
0.4 × 104
0.2 × 104
0.6 × 1040.8 × 104
1.8 × 104
1.6 × 104
1.4 × 104
1.0 × 1041.2 × 104
0.4 × 1040.6 × 1040.8 × 104
1.8 × 1041.0 × 104
0.8 × 104
0.6 × 104
0.4 × 104
0.2 × 104
1.6 × 104
1.4 × 104
Time (min)
0 906030
Time (min)
0 906030
Time (min)
0 906030
Time (min)
0 906030
Time (min)
0 906030
Time (min)
0 906030
Rab2
4.0 × 104
1.0 × 104
2.0 × 104
3.0 × 104
5.0 × 104Rab35
0.2 × 104
0.4 × 104
0.6 × 104
0.8 × 104Rab18Rab7
8.0 µm
1.0 µm
2.0 µm
4.0 µm
His−DrrA451-647
c
1.40 × 105
1.0 × 104
0.8 × 104
0.6 × 104
0.4 × 104
0.2 × 104
1.6 × 103
35
No activity
1.2 × 104
1.0 × 105
2.0 × 105
100
1.1
0.0003
0
8.6
71
143
+
−
−
−
−
+
−
Rab
1 G
EF a
ctiv
ity (k
cat/
K m)
Rab
1 G
EF a
ctiv
ity (p
erce
ntag
e of
WT)
Rab
1S2
5N b
indi
ng
l 440
(arb
itrar
y un
its)
l 440
(arb
itrar
y un
its)
l 440
(arb
itrar
y un
its)
l 440
(arb
itrar
y un
its)
l 440
(arb
itrar
y un
its)
l 440
(arb
itrar
y un
its)
l 440
(arb
itrar
y un
its)
Figure 3 Deletion analysis reveals regions in DrrA necessary for Rab1 binding and GEF activity. (a) Rab1 GEF and Rab1-binding activities determined for the indicated His-tagged DrrA truncation derivatives are shown. The schematic shows regions in DrrA that were determined to be required for wild-type levels of Rab1 binding and GEF activity. (b) His-tagged DrrA451–647 was assayed for guanine nucleotide-exchange
activity on Rab1 protein. The traces show the rate of mant–GDP release on addition of His–DrrA451–647 at the indicated concentrations. (c) The release of mant–GDP from the indicated Rab was assayed by measuring the decrease in fluorescence over time. The traces show the rate of mant–GDP release on addition of His–DrrA451–647 at the indicated concentrations.
print_ncb1463.indd 975print_ncb1463.indd 975 10/8/06 12:05:57 pm10/8/06 12:05:57 pm
Nature Publishing Group ©2006
976 NATURE CELL BIOLOGY VOLUME 8 | NUMBER 9 | SEPTEMBER 2006
L E T T E R S
medium (Gibco) supplemented with 10% heat-inactivated FBS. Mammalian cell cultures were all maintained in a 5% CO2 incubator at 37 °C. A. castella-nii (ATCC30234) was grown at room temperature in ATCC medium 712. For L. pneumophila infection, amoeba cells were incubated in ATCC medium 712 without peptone, yeast extract or glucose (amoeba buffer) and incubated in a 5% CO2 incubator at 37 °C.
Screening and analysis of L. pneumophila transposon mutants defective in Rab1 recruitment. The methods used to generate L. pneumophila transposon insertion mutants and characterize mutants for Rab1 recruitment defects are described in detail in the Supplementary Information, Methods.
Protein purification. E. coli BL21 (DE3) cells transformed with vectors producing the indicated proteins were grown at 37 °C in LB broth containing appropriate antibiotics to an A600 = 0.4. IPTG was added to the media at final concentra-tion of 0.2 mM and samples were incubated for 5 h at 30 °C. Cells harvested by centrifugation were resuspended in lysis buffer (PBS, 1 mM dithiothreitol (DTT), 1% Triton X-100 or 50 mM Tris at pH 8.0 200 mM NaCl, 1 mM DTT with 1 mM MgCl2), then disrupted by french press. Lysates were clarified by centrifu-gation at 15,000g for 30 min. For His-tagged fusion proteins, supernatants were loaded onto Ni-NTA agarose columns and washed with lysis buffer containing 500 mM NaCl and 10 mM imidazole. His-tagged fusion proteins were eluted with 100 mM imidazole in 50 mM Tris at pH 8.0, 200 mM NaCl and 1 mM DTT.
GFP RFP−Rab1 Mergea
bGFP RFP−Rab1 Merge
GFP RFP−Rab1 Merge
c
d
GFP RFP−Rab1 Merge
GFP−DrrA61−647
GFP−DrrA451−647
GFP−DrrA61−647
GFP−DrrA451−647
Figure 4 DrrA has the intrinsic ability to localize and recruit Rab1 to the plasma membrane. (a, b) The top images show an x–y projection of a confocal z-series of cells transfected with RFP–Rab1 and either GFP–DrrA61–647 (a) or GFP–DrrA451–647 (b). The white line indicates the longitudinal slice through the cell that resulted in the x–z images shown in the bottom panels. Greyscale images of GFP fluorescence (left) and
RFP–Rab1 fluorescence (middle) and the merged colour image (right) are shown. (c) TIRFM images show localization of GFP–DrrA61–647 (left) and RFP–Rab1 (middle) at the plasma membrane. (d) TIRFM images show localization of GFP–DrrA451–647 (left) at the plasma membrane, but no RFP–Rab1 (middle) recruitment was detected. The scale bar represents 5 µm.
print_ncb1463.indd 976print_ncb1463.indd 976 10/8/06 12:05:59 pm10/8/06 12:05:59 pm
Nature Publishing Group ©2006

NATURE CELL BIOLOGY VOLUME 8 | NUMBER 9 | SEPTEMBER 2006 977
L E T T E R S
For GST-fusion proteins, supernatants were loaded onto glutathione–Sepharose columns (GE Healthcare, Piscataway, NJ) and washed with lysis buffer contain-ing 1 mM MgCl2. Bound proteins were eluted in lysis buffer containing 10 mM glutathione. For GST–Rab protein-binding assays, lysates from E. coli produc-ing the indicated proteins were incubated with the indicated GST–Rab-charged glutathione resin for 30 min at 4 °C, columns were washed extensively with lysis buffer containing 1 mM MgCl2, then proteins were eluted with glutathione. To assess binding of His-tagged DrrA proteins to GST–Rab1S25N, purified proteins were dialysed in binding buffer (20 mM HEPES at pH 7.4, 0.1 M KCl, 5 mM MgCl2 and 1 mM DTT). Proteins (1 nM) were incubated together in 0.2 ml of binding buffer with 25 µl of glutathione resin for 30 min. Glutathione resins were extensively washed with binding buffer and eluted with 10 mM glutathione.
GFP–DrrA localization studies. CHO FcγRII cells were plate at a density of 1 × 105 cells per well in 24-well tissue culture plates with glass coverslips of 12-mm diameter. Plasmids expressing GFP–DrrA and truncation derivatives were cotransfected into CHO FcγRII cells with plasmids encoding monomeric RFP fused to Rab1 (pRFP–rab1) using FuGENE6 (Roche, Indianapolis, IN). After transfection, cells were incu-bated for 20 h, fixed with 3% PFA and permeabilized with ice-cold methanol before visualization. Images were acquired with a Zeiss LSM510 microscope using a 100× 1.4 N.A. objective. For multi-channel TIRFM, CHO FcγRII cells grown in 35-mm glass bottom dishes (MatTek, Ashland, MA) were excited at 488 nm and 568 nm with an Olympus objective-type TIRFM setup using a triple dichroic filter (DAPI–FITC–TexasRed; Semrock, Rochester, NY) and a 60× 1.45 N.A. objective. The penetration depth of this setup at 570 nm is approximately 100 nm for TIR excitation at the outer rim of the back focal plane of the objective and assuming a cytosolic refractive index of approximately 1.36. Fluorescence was detected with an iXon 987 back-thinned EMCCD camera (5112 ×x 512; Andor, Belfast, UK) using Andor’s IQM software.
Nucleotide exchange assays. Rab proteins were loaded with the fluorescent GDP analogue 1′(3)-bis-O-(N-methylanthraniloyl) GDP (mant–GDP; Molecular Probes, Carlsbad, CA) by incubating for 30 min at 37 °C in loading buffer (20 mM Tris at pH 8.0, 150 mM NaCl, 5 mM EDTA and 1 mM DTT) containing a 25 mM excess of mant–GDP. The exchange reaction mixture was transferred to ice, the reaction was terminated by the addition of 10 mM MgCl2 and free mant–GDP removed by gel-filtration on a prepacked d-salt column (Pierce, Rockford, IL). For nucleotide-release assays, Rab proteins bound to mant–GDP were diluted to a final concentration of 1 µM in reaction buffer (20 mM Tris at pH 8.0, 150 mM NaCl and 0.5 mM MgCl2.
The dissociation of mant–GDP at various concentrations of DrrA was deter-mined by measuring the decrease in fluorescence that accompanies release of mant–GDP in the presence of a large excess of GppNHp (200 µM final concentra-tion), with or without varying concentrations of DrrA. Samples were excited at 360 nm and the emission monitored at 440 nm using a Saphire multimode micro-plate spectrophotometer (Tecan, Zurich, Switzerland). Observed pseudo-first-order rate constants (kobs) at each concentration of DrrA were derived from a non-linear least squares fit to the exponential function I(t) = (I
0–I
∞) exp(–kobs t) + I∞ where
I(t) represents the emission intensity as a function of time and I0 and I
∞ are the emis-
sion intensities at t = 0 and t = ∞, respectively. The catalytic efficiency (kcat/Km) was obtained from the slope of a linear least squares fit to kobs = ( kcat/Km) [DrrA] + kintr, where kintr is the intrinsic rate constant for GDP release in the absence of DrrA. For GTP-loading studies, Rab1 protein was loaded with GDP as described above and diluted to 1 µM in 50 mM Tris at pH 8.0, 150 mM NaCl, 0.5 mM MgCl2. To initi-ate the reaction mant–GppNHp was added to a final concentration of 10 µM and His–DrrA was added at the concentrations indicated. Data were collected on an ISS spectrofluorometer with the excitation wavelength set to 290 nm and the emission monitored at 460 nm. Catalytic efficiency was calculated as described above.
Accession numbers. The accession number for drrA is AY945933.
Note: Supplementary Information is available on the Nature Cell Biology website.
ACKNOWLEDGEMENTSWe thank G. Warren, A. Wandinger-Ness, W. Moss and E. Rubin for providing plasmids used in this study. This work was supported by National Institutes of Health (NIH) grants to C.R.R. (AI41699) and D.G.L. (GM56324), the Ludwig Institute for Cancer Research (D.T.), a Charles A. King Postdoctoral Fellowship (A.D.) and an NSF Predoctoral Award (A.I.).
COMPETING FINANCIAL INTERESTSThe authors declare that they have no competing financial interests.
Published online at http://www.nature.com/naturecellbiology/Reprints and permissions information is available online at http://npg.nature.com/reprintsandpermissions/
1. Horwitz, M. A. The Legionnaires’ disease bacterium (Legionella pneumophila) inhibits phagosome-lysosome fusion in human monocytes. J. Exp. Med. 158, 2108–2126 (1983).
2. Kagan, J. C. & Roy, C. R. Legionella phagosomes intercept vesicular traffic from endo-plasmic reticulum exit sites. Nature Cell Biol. 4, 945–954 (2002).
3. Luo, Z. Q. & Isberg, R. R. Multiple substrates of the Legionella pneumophila Dot/Icm system identified by interbacterial protein transfer. Proc. Natl Acad. Sci. USA 101, 841–846 (2004).
4. Chen, J. et al. Legionella effectors that promote nonlytic release from protozoa. Science 303, 1358–1361 (2004).
5. Nagai, H., Kagan, J. C., Zhu, X., Kahn, R. A. & Roy, C. R. A bacterial guanine nucleotide exchange factor activates ARF on Legionella phagosomes. Science 295, 679–682 (2002).
6. Vogel, J. P., Andrews, H. L., Wong, S. K. & Isberg, R. R. Conjugative transfer by the virulence system of Legionella pneumophila. Science 279, 873–876 (1998).
7. Segal, G., Purcell, M. & Shuman, H. A. Host cell killing and bacterial conjugation require overlapping sets of genes within a 22-kb region of the Legionella pneumophila genome. Proc. Natl Acad. Sci. USA 95, 1669–1674 (1998).
8. Kagan, J. C., Stein, M. P., Pypaert, M. & Roy, C. R. Legionella subvert the functions of Rab1 and Sec22b to create a replicative organelle. J. Exp. Med. 199, 1201–1211 (2004).
9. Derre, I. & Isberg, R. R. Legionella pneumophila replication vacuole formation involves rapid recruitment of proteins of the early secretory system. Infect. Immun. 72, 3048–3053 (2004).
10. Robinson, C. G. & Roy, C. R. Attachment and fusion of endoplasmic reticulum with vacuoles containing Legionella pneumophila. Cell Microbiol. 8, 793–805 (2006).
11. Dorer, M. S., Kirton, D., Bader, J. S. & Isberg, R. R. RNA interference analysis of Legionella in Drosophila cells: exploitation of early secretory apparatus dynamics. PLoS Pathog. 2, e34 (2006).
12. Amor, J. C. et al. The structure of RalF, an ADP-ribosylation factor guanine nucleotide exchange factor from Legionella pneumophila, reveals the presence of a cap over the active site. J. Biol. Chem. 280, 1392–1400 (2005).
13. Pereira-Leal, J. B. & Seabra, M. C. Evolution of the Rab family of small GTP-binding proteins. J. Mol. Biol. 313, 889–901 (2001).
14. Pfeffer, S. R. Rab GTPases: specifying and deciphering organelle identity and function. Trends Cell Biol. 11, 487–491 (2001).
15. Zerial, M. & McBride, H. Rab proteins as membrane organizers. Nature Rev. Mol. Cell Biol. 2, 107–117 (2001).
16. Bock, J. B., Matern, H. T., Peden, A. A. & Scheller, R. H. A genomic perspective on membrane compartment organization. Nature 409, 839–841 (2001).
17. Nagai, H. et al. A C-terminal translocation signal required for Dot/Icm-dependent delivery of the Legionella RalF protein to host cells. Proc. Natl Acad. Sci. USA 102, 826–831 (2005).
18. Sory, M. P. & Cornelis, G. R. Translocation of a hybrid YopE-adenylate cyclase from Yersinia enterocolitica into HeLa cells. Mol. Microbiol. 14, 583–594 (1994).
19. Conover, G. M., Derre, I., Vogel, J. P. & Isberg, R. R. The Legionella pneumophila LidA protein: a translocated substrate of the Dot/Icm system associated with maintenance of bacterial integrity. Mol. Microbiol. 48, 305–321 (2003).
20. Derre, I. & Isberg, R. R. LidA, a translocated substrate of the Legionella pneumophila type IV secretion system, interferes with the early secretory pathway. Infect. Immun. 73, 4370–4380 (2005).
21. Zhu, A. X., Zhao, Y. & Flier, J. S. Molecular cloning of two small GTP-binding proteins from human skeletal muscle. Biochem. Biophys. Res. Commun. 205, 1875–1882 (1994).
22. Yu, H., Leaf, D. S. & Moore, H. P. Gene cloning and characterization of a GTP-binding Rab protein from mouse pituitary AtT-20 cells. Gene 132, 273–278 (1993).
23. Delprato, A., Merithew, E. & Lambright, D. G. Structure, exchange determinants, and family-wide rab specificity of the tandem helical bundle and Vps9 domains of Rabex-5. Cell 118, 607−617 (2004).
24. Roy, C. R. & Isberg, R. R. Topology of Legionella pneumophila DotA: an inner mem-brane protein required for replication in macrophages. Infect. Immun. 65, 571–578 (1997).
25. Celada, A., Gray, P. W., Rinderknecht, E. & Schreiber, R. D. Evidence for a γ-interferon receptor that regulates macrophage tumoricidal activity. J. Exp. Med. 160, 55–74 (1984).
26. Joiner, K. A., Fuhrman, S. A., Miettinen, H. M., Kasper, L. H. & Mellman, I. Toxoplasma gondii: fusion competence of parasitophorous vacuoles in Fc receptor-transfected fibroblasts. Science 249, 641–646 (1990).
27. Machner, M. P., Isberg, R. R. Targeting of host rab GTPase function by the intravacuolar pathogen Legionella pneumophila. Dev Cell. 11, 47−56 (2006).
print_ncb1463.indd 977print_ncb1463.indd 977 10/8/06 12:06:01 pm10/8/06 12:06:01 pm
Nature Publishing Group ©2006
S U P P L E M E N TA RY I N F O R M AT I O N
WWW.NATURE.COM/NATURECELLBIOLOGY 1
1 .0 x 1 0 1
1 .0 x 1 0 2
1 .0 x 1 0 3
1 .0 x 1 0 4
1 .0 x 1 0 5
pcya
-drr
A1-
647
pcya
-drr
B92
-255
pcya
pcya
-ral
F1-3
74
pcya
-lidA
378-
729
pcya
-lvgA
98-2
08
pcya
-lspE
1-49
4
pcya
-drr
E66
-268
pcya
-drr
D1-
475
pcya
-drr
C1-
260
Murata Supplementary Information Figure 1
Wild
-typ
e
Δdo
tA
Δdr
rA/p
Drr
A
Δdr
rA
81
50
kDa
DrrA
anti-DrrA
anti-DrrALegionellaDNA
L. pneumophila (wt) L. pneumophila (∆dotA)
anti-DrrA
anti-DrrALegionellaDNA
anti-DrrA
anti-DrrALegionellaDNA
L. pneumophila (∆drrA)
GFP-Rab1Legionella wild type
GFP-Rab1Legionella ΔdotA
GFP-Rab1Legionella ΔdrrA
GFP-Rab1Legionella ΔdrrA/pDrrA
a b
f
c d e
g
Rel
ativ
e C
FU
Time (h) Time (h)
Rel
ativ
e C
FU
Macrophage A. castellanii
0 24 481.0x10-1
1.0x100
1.0x101
1.0x102
1.0x103
1.0x104
wt
ΔdotA
ΔdrrA
0 24 481.0x10-1
1.0x100
1.0x101
1.0x102
wt
ΔdotA
ΔdrrA
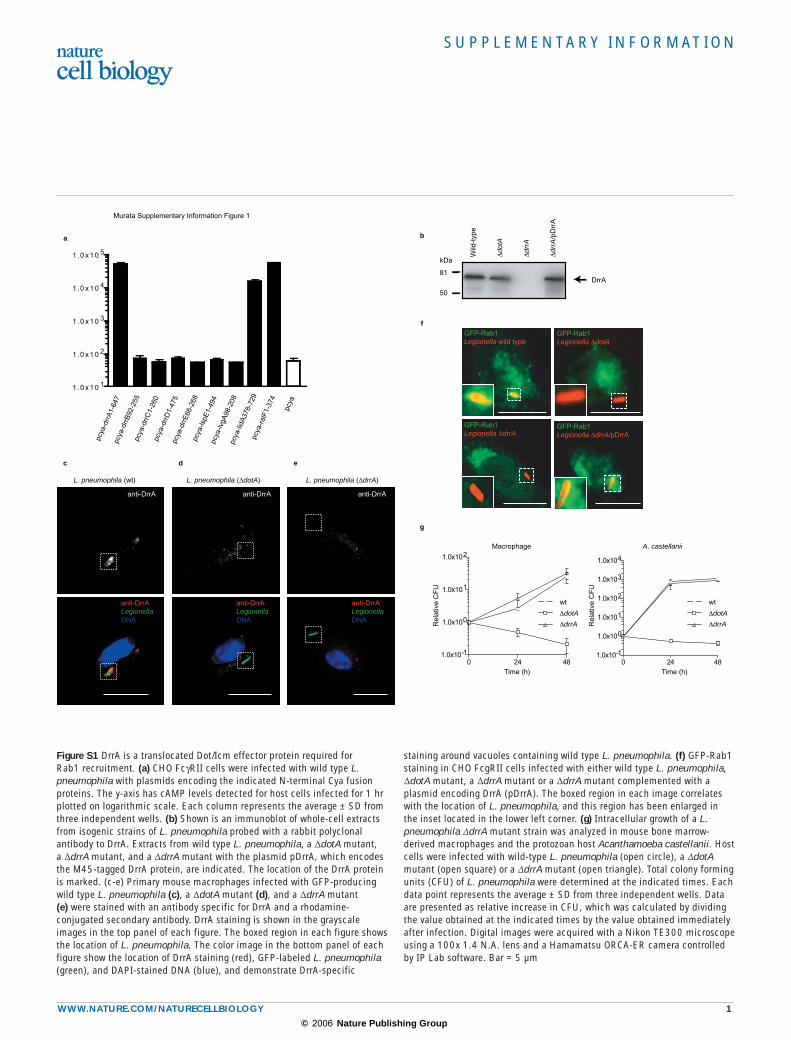
Figure S1 DrrA is a translocated Dot/Icm effector protein required for Rab1 recruitment. (a) CHO FcγRII cells were infected with wild type L. pneumophila with plasmids encoding the indicated N-terminal Cya fusion proteins. The y-axis has cAMP levels detected for host cells infected for 1 hr plotted on logarithmic scale. Each column represents the average ± SD from three independent wells. (b) Shown is an immunoblot of whole-cell extracts from isogenic strains of L. pneumophila probed with a rabbit polyclonal antibody to DrrA. Extracts from wild type L. pneumophila, a ∆dotA mutant, a ∆drrA mutant, and a ∆drrA mutant with the plasmid pDrrA, which encodes the M45-tagged DrrA protein, are indicated. The location of the DrrA protein is marked. (c-e) Primary mouse macrophages infected with GFP-producing wild type L. pneumophila (c), a ∆dotA mutant (d), and a ∆drrA mutant (e) were stained with an antibody specific for DrrA and a rhodamine-conjugated secondary antibody. DrrA staining is shown in the grayscale images in the top panel of each figure. The boxed region in each figure shows the location of L. pneumophila. The color image in the bottom panel of each figure show the location of DrrA staining (red), GFP-labeled L. pneumophila (green), and DAPI-stained DNA (blue), and demonstrate DrrA-specific
staining around vacuoles containing wild type L. pneumophila. (f) GFP-Rab1 staining in CHO FcgRII cells infected with either wild type L. pneumophila, ∆dotA mutant, a ∆drrA mutant or a ∆drrA mutant complemented with a plasmid encoding DrrA (pDrrA). The boxed region in each image correlates with the location of L. pneumophila, and this region has been enlarged in the inset located in the lower left corner. (g) Intracellular growth of a L. pneumophila ∆drrA mutant strain was analyzed in mouse bone marrow-derived macrophages and the protozoan host Acanthamoeba castellanii. Host cells were infected with wild-type L. pneumophila (open circle), a ∆dotA mutant (open square) or a ∆drrA mutant (open triangle). Total colony forming units (CFU) of L. pneumophila were determined at the indicated times. Each data point represents the average ± SD from three independent wells. Data are presented as relative increase in CFU, which was calculated by dividing the value obtained at the indicated times by the value obtained immediately after infection. Digital images were acquired with a Nikon TE300 microscope using a 100x 1.4 N.A. lens and a Hamamatsu ORCA-ER camera controlled by IP Lab software. Bar = 5 µm
© 2006 Nature Publishing Group
S U P P L E M E N TA RY I N F O R M AT I O N
2 WWW.NATURE.COM/NATURECELLBIOLOGY
0.5 μM5 mM EDTA
Intrinsic0.008 μM0.032 μM0.125 μM
Rab1-DrrA
time (min)
I 440
(ar
bitr
ary
uits
)
0 30 60 901.0x104
2.0x104
3.0x104
4.0x104
a
81
114
64
50
3726
21
15
GST-Rab1 S25N
kDa
His
-Drr
A (
1-64
7)
His
-Drr
A (
201-
647)
His
-Drr
A (
1-50
0)
His
-Drr
A (
546-
647)
His
-Drr
A (
451-
647)
His
-Drr
A (
1-44
8)
(1%) (1%)
Input
His-DrrA (anti-His)
b
Murata Supplementary Information, Figure 2
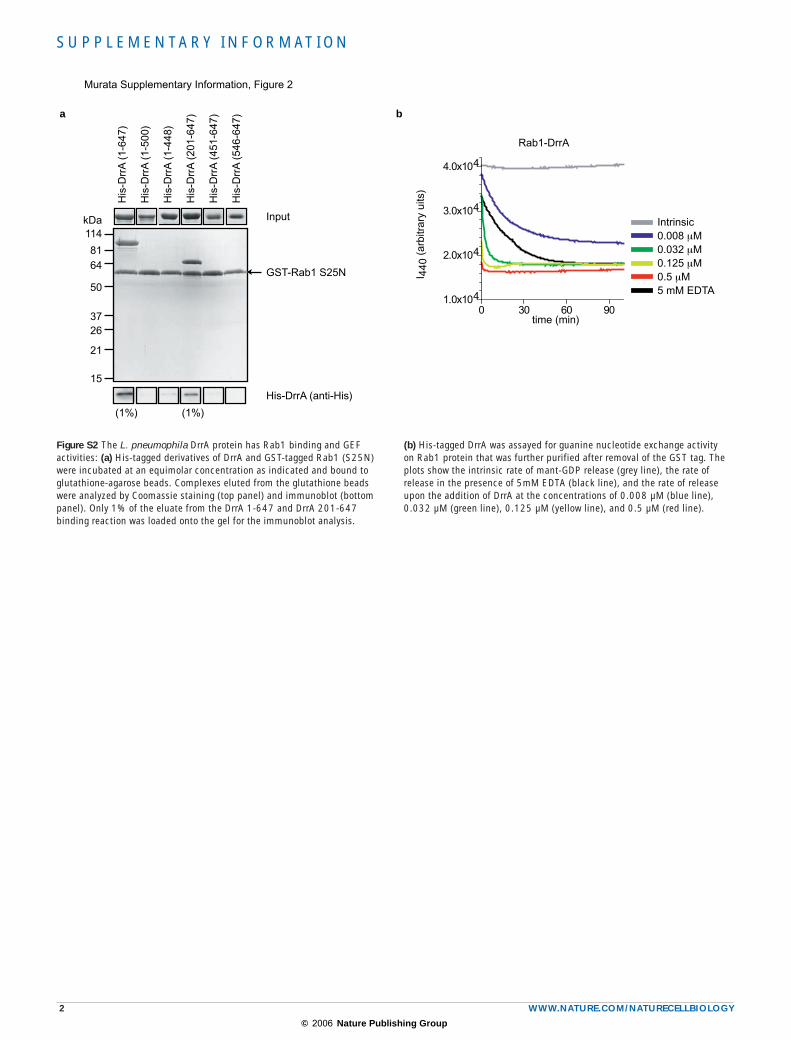
Figure S2 The L. pneumophila DrrA protein has Rab1 binding and GEF activities: (a) His-tagged derivatives of DrrA and GST-tagged Rab1 (S25N) were incubated at an equimolar concentration as indicated and bound to glutathione-agarose beads. Complexes eluted from the glutathione beads were analyzed by Coomassie staining (top panel) and immunoblot (bottom panel). Only 1% of the eluate from the DrrA 1-647 and DrrA 201-647 binding reaction was loaded onto the gel for the immunoblot analysis.
(b) His-tagged DrrA was assayed for guanine nucleotide exchange activity on Rab1 protein that was further purified after removal of the GST tag. The plots show the intrinsic rate of mant-GDP release (grey line), the rate of release in the presence of 5mM EDTA (black line), and the rate of release upon the addition of DrrA at the concentrations of 0.008 µM (blue line), 0.032 µM (green line), 0.125 µM (yellow line), and 0.5 µM (red line).
© 2006 Nature Publishing Group
S U P P L E M E N TA RY I N F O R M AT I O N
WWW.NATURE.COM/NATURECELLBIOLOGY 3
GFP-DrrA GM130 Merge
GF
P-D
rrA
61-
647
GFP RFP-GalT Merge
GF
P-D
rrA
451
-647
GF
P-D
rrA
61-
647
GF
P-D
rrA
451
-647
Murata Supplementary Infromation Figure 3
aa
b
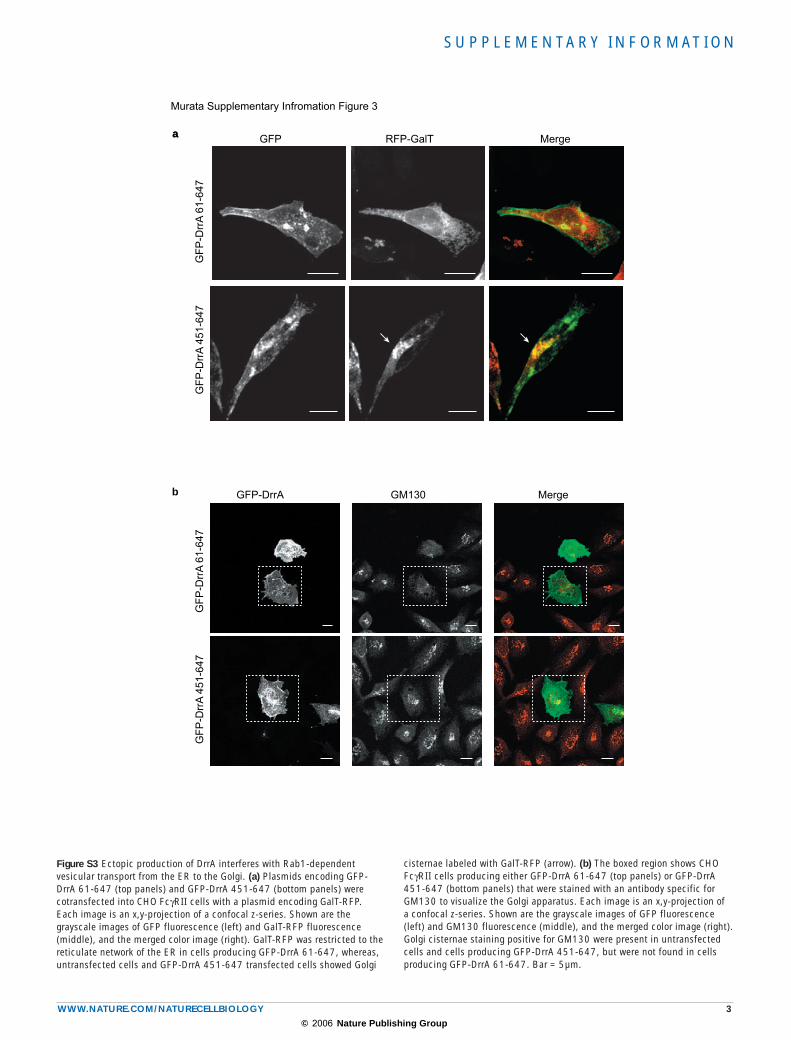
Figure S3 Ectopic production of DrrA interferes with Rab1-dependent vesicular transport from the ER to the Golgi. (a) Plasmids encoding GFP-DrrA 61-647 (top panels) and GFP-DrrA 451-647 (bottom panels) were cotransfected into CHO FcγRII cells with a plasmid encoding GalT-RFP. Each image is an x,y-projection of a confocal z-series. Shown are the grayscale images of GFP fluorescence (left) and GalT-RFP fluorescence (middle), and the merged color image (right). GalT-RFP was restricted to the reticulate network of the ER in cells producing GFP-DrrA 61-647, whereas, untransfected cells and GFP-DrrA 451-647 transfected cells showed Golgi
cisternae labeled with GalT-RFP (arrow). (b) The boxed region shows CHO FcγRII cells producing either GFP-DrrA 61-647 (top panels) or GFP-DrrA 451-647 (bottom panels) that were stained with an antibody specific for GM130 to visualize the Golgi apparatus. Each image is an x,y-projection of a confocal z-series. Shown are the grayscale images of GFP fluorescence (left) and GM130 fluorescence (middle), and the merged color image (right). Golgi cisternae staining positive for GM130 were present in untransfected cells and cells producing GFP-DrrA 451-647, but were not found in cells producing GFP-DrrA 61-647. Bar = 5µm.
© 2006 Nature Publishing Group
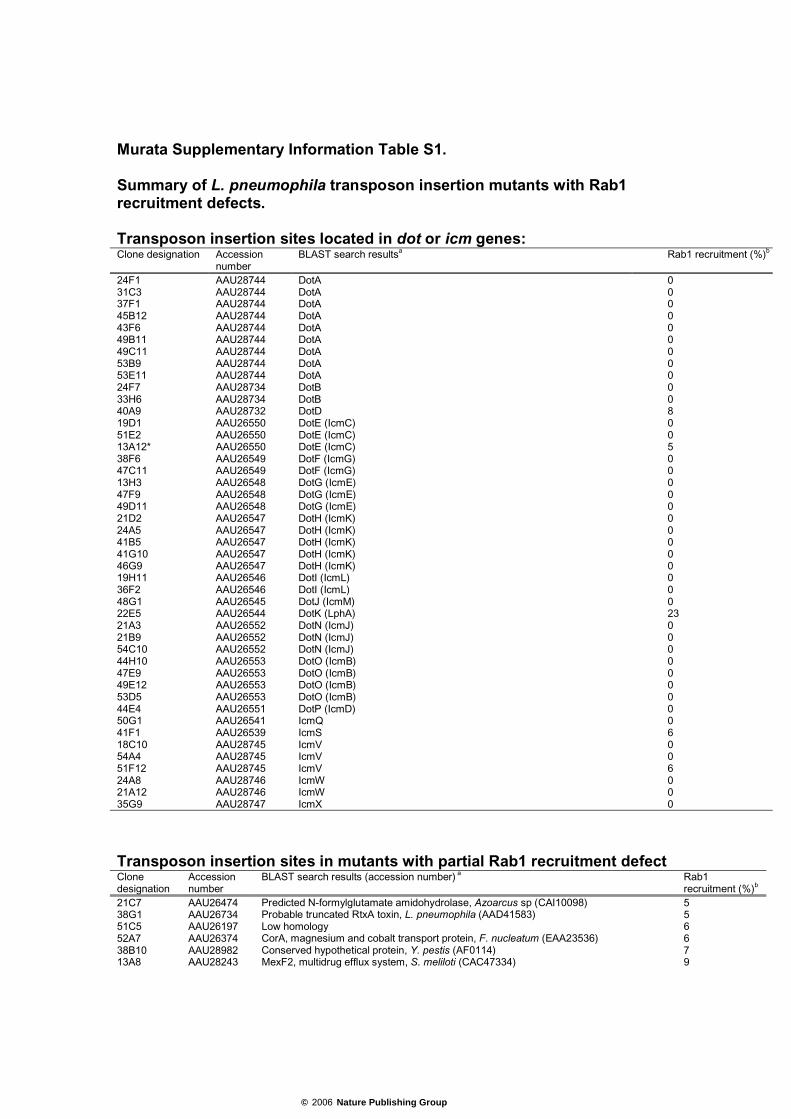
Murata Supplementary Information Table S1. Summary of L. pneumophila transposon insertion mutants with Rab1 recruitment defects. Transposon insertion sites located in dot or icm genes: Clone designation Accession
number BLAST search resultsa Rab1 recruitment (%)b
24F1 AAU28744 DotA 0 31C3 AAU28744 DotA 0 37F1 AAU28744 DotA 0 45B12 AAU28744 DotA 0 43F6 AAU28744 DotA 0 49B11 AAU28744 DotA 0 49C11 AAU28744 DotA 0 53B9 AAU28744 DotA 0 53E11 AAU28744 DotA 0 24F7 AAU28734 DotB 0 33H6 AAU28734 DotB 0 40A9 AAU28732 DotD 8 19D1 AAU26550 DotE (IcmC) 0 51E2 AAU26550 DotE (IcmC) 0 13A12* AAU26550 DotE (IcmC) 5 38F6 AAU26549 DotF (IcmG) 0 47C11 AAU26549 DotF (IcmG) 0 13H3 AAU26548 DotG (IcmE) 0 47F9 AAU26548 DotG (IcmE) 0 49D11 AAU26548 DotG (IcmE) 0 21D2 AAU26547 DotH (IcmK) 0 24A5 AAU26547 DotH (IcmK) 0 41B5 AAU26547 DotH (IcmK) 0 41G10 AAU26547 DotH (IcmK) 0 46G9 AAU26547 DotH (IcmK) 0 19H11 AAU26546 DotI (IcmL) 0 36F2 AAU26546 DotI (IcmL) 0 48G1 AAU26545 DotJ (IcmM) 0 22E5 AAU26544 DotK (LphA) 23 21A3 AAU26552 DotN (IcmJ) 0 21B9 AAU26552 DotN (IcmJ) 0 54C10 AAU26552 DotN (IcmJ) 0 44H10 AAU26553 DotO (IcmB) 0 47E9 AAU26553 DotO (IcmB) 0 49E12 AAU26553 DotO (IcmB) 0 53D5 AAU26553 DotO (IcmB) 0 44E4 AAU26551 DotP (IcmD) 0 50G1 AAU26541 IcmQ 0 41F1 AAU26539 IcmS 6 18C10 AAU28745 IcmV 0 54A4 AAU28745 IcmV 0 51F12 AAU28745 IcmV 6 24A8 AAU28746 IcmW 0 21A12 AAU28746 IcmW 0 35G9 AAU28747 IcmX 0
Transposon insertion sites in mutants with partial Rab1 recruitment defect Clone designation
Accession number
BLAST search results (accession number) a Rab1 recruitment (%)b
21C7 AAU26474 Predicted N-formylglutamate amidohydrolase, Azoarcus sp (CAI10098) 5 38G1 AAU26734 Probable truncated RtxA toxin, L. pneumophila (AAD41583) 5 51C5 AAU26197 Low homology 6 52A7 AAU26374 CorA, magnesium and cobalt transport protein, F. nucleatum (EAA23536) 6 38B10 AAU28982 Conserved hypothetical protein, Y. pestis (AF0114) 7 13A8 AAU28243 MexF2, multidrug efflux system, S. meliloti (CAC47334) 9
© 2006 Nature Publishing Group

32E9 AAU27663 NAD-glutamate dehydrogenase, P. profundum (CAG20173) 12 50F12* AAU28400 DksA, DnaK suppressor protein, E. coli (BAB96722) 13 37A9 AAU26765 Conserved hypothetical protein, P. luminescens (CAE12376) 14 38G3 No ORF 14 41A3* AAU27013 Restin, G. gallus (O42184) 18 52B1 AAU27185 Hypothetical protein, C. albicans (EAK93571) 19 42G5 AAU27817 Cell surface antigen, R. conorii (AAL02557) 20 38G6 AAU28658 Acyltransferase family protein, P. syringae (AAO54531) 22 42C9* AAU26359 Putative membrane protein, E. carotovora (CAG77251) 24 52A9 AAU27808 sn-glycerol-3-phosphate ABC type transport system, Y. pseudotuberculosis (CAH19481) 26 43B1 AAU27807 Hypothetical protein, R. conorii (AAL03274) 29 24G5 AAU26954 3-methyladenine DNA glycosylase, C. acetobutylicum (AAK80911) 32 49C7 AAU28703 UvrC, excinuclease ABC subunit, P. fluorescence (P32966) 32 31E7 AAU27838 FliG, flagellar motor switch protein, P. syringae (AAO55477) 32 31G8 AAU27402 LspE, type II secretory pathway, L. pneumophila 32 51C6 No ORF 32 37B8 AAU26319 PhrB, deoxyribodipyrimidine photolyase, C. burnetii (NP_820171) 33 39A11* AAU28320 Hypothetical protein, R. gelatinosus (ZP_00244406) 33 42G4 AAU27745 Putative membrane protein, Y. pseudotuberculosis (CAH20309) 33 13G8 AAU27677 Conserved hypothetical protein. P. aeruginosa (AAG06242) 35 13C3 AAU28168 Sterol desaturase, V. vulnificus (AAO10919) 36 42D4 AAU28067 Histidinol phosphate aminotransferase. P. aeruginosa (AAG06553) 36 22D12 AAU27027 LidA; priming of multiplication, L. pneumophila 36 14F3 AAU27027 LidA; priming of multiplication, L. pneumophila 37 48B7 AAU28810 Hypothetical protein 40 48G5 AAU26248 Oligopeptidase A, V. parahaemolyticus (BAC58333) 40 42H5 No ORF 40 42C6 AAU26188 Low homology 42 33A6 AAU27027 LidA; priming of multiplication, L. pneumophila 43 43C2 AAU28255 DotI/IcmL like protein, L. pneumophila 44 44C5 AAU27717 Polysaccharide deacetylase, P. profundum (CAG20649) 44 42H1 No ORF 47 41A10 AAU27047 Peptide ABC transporter, permease protein, C. burnetii (NP_820836) 48 41A12 AAU28566 Sensor histidine kinase/response regulator, B. fungorum (ZP_00281724) 48 48A6 AAU26878 AlgI, alginate O-acetyltransferase, R. baltica (CAD73424) 48 31A7 AAU27947 PulL, type II secretory pathway, I. loihiensis (AAV82850) 48 35B12 No ORF 48 15A5 AAU26290 Predicted NAD/FAD-binding protein, B. cepacia (ZP_00224402) 50 45A3 AAU27027 LidA; priming of multiplication, L. pneumophila 50
Transposon insertion sites in mutants with a complete Rab1 recruitment defects Clone designation
Accession number
BLAST search results (accession number) a DotA secretion phenotypec
17B5 AAU28537 High-affinity nickel permease, D. aromatica (ZP_00203766) + 36A4 AAU28378 BdhA; D-beta-hydroxybutyrate dehydrogenase, B. mallei (AAU46755) + 38C5 AAU26602 EcfE; zinc metalloprotease, S. typhimurium (Q8ZRP1) + 38G5 AAK35046 LspE; putative type II secretion ATPase, L. pneumophila + 38H7 AY945933 LepB; effector protein B, L. pneumophila (AAP20593) + 48E1 AAU26602 EcfE; zinc metalloprotease, S. typhimurium (Q8ZRP1) + 51C1 AAU28917 Predicted permease, M. flagellatus (ZP_00350142) + 52B12 AAF05324 LvgA; virulence protein, L. pneumophila + 52G5 AAU27027 LidA; priming of multiplication, L. pneumophila + 13A4 AAU26733 Conserved hypothetical protein, B. japonicum (BAC50736) - 15C3 AAU27520 CpxR, transcriptional regulatory protein, L. pneumophila - 36D8 AAU27543 RecJ, single stranded DNA specific exonuclease, P. aeruginosa (CAH12568) - 38A5 No ORF - 38E3 AAU27450 Hypothetical protein - 41D4 AAU28556 RNA methyltransferase, TrmH family, group 2, C. burnetii (AAO91014) - 41E7 AAU28220 Similar to SdeD, L. pneumophila (AAU28569) - 42A4 AAU27950 Deoxyguanosinetriphosphate triphosphohydrolase, V. cholerae (AAF95127) - 46G4 AAU26987 Conserved hypothetical protein, C. burnetii (AAO90465) - 53C2 AAU28799 EngC GTPase, H. influenzae (P45339) -
© 2006 Nature Publishing Group

* indicated that the transposon insertion loci were not in ORFs but upstream region of indicated genes. a. Proteins that showing highest similarity based on E value (at least less than 0.05) were listed in this column. b. Efficiency of Rab1 recruitment was shown as the ratio comparing to the Rab1 recruitment of wild type L. pneumophila in each experiments. c. Bacterial culture supernatants were precipitated with 10% TCA and analyzed by SDS-PAGE as described in Materials and methods.
© 2006 Nature Publishing Group

Table S2. Bacterial strains, plasmids and primers used in this study. Strains Genotype Reference CR39 L. pneumopnila serogroup 1, strain LP01 rpsL 1 CR58 LP01 ∆dotA 2 CR1614 LP01 ∆lidA781 In this study CR1616 LP01 ∆drrA24 In this study CR1630 LP01 ∆drrA24 (pdrrA) In this study Plasmids Properties Reference pMMB207 Cloning vector derived from RSF1010 3 pMMB207-M45 Amino-terminal M45 epitope-tag vector 4 pdrrA pMMB207-M45NT encoding M45-drrA In this study plidA pMMB207-M45NT encoding M45-lidA In this study pM45-ralF pMMB207-M45NT encoding M45-ralF 5 pcya pMMB207-M45NT encoding M45-cya 5 pcya-drrA pcya encoding M45-cya-drrA In this study pcya-drrB92-255 pcya encoding M45-cya-drrB92-255 In this study pcya-drrC pcya encoding M45-cya-drrC In this study pcya-drrD pcya encoding M45-cya-drrD In this study pcya-drrE66-268 pcya encoding M45-cya-drrE66-268 In this study pcya-lidA378-729 pcya encoding M45-cya-lidA378-729 In this study pcya-lspE pcya encoding M45-cya-lspE In this study pcya-lvgA98-208 pcya encoding M45-cya-lvgA98-208 In this study pcya-ralF pcya encoding M45-cya-ralF 5 pET15b Expression vector for Histidine-tagged proteins Novagen pH6-drrA 1-647 pET15b encoding his-tagged drrA 1-647 In this study pH6-drrA 1-500 pET15b encoding his-tagged drrA 2-615 In this study pH6-drrA 1-488 pET15b encoding his-tagged drrA 2-545 In this study pH6-drrA 1-366 pET15b encoding his-tagged drrA 2-450 In this study pH6-drrA 1-200 pET15b encoding his-tagged drrA 2-366 In this study pH6-drrA 201-647 pET15b encoding his-tagged drrA 90-366 In this study pH6-drrA 451-647 pET15b encoding his-tagged drrA 117-366 In this study pH6-drrA 546-647 pET15b encoding his-tagged drrA 150-366 In this study pGEX-2TK Expression vector for GST-tagged proteins Amersham pGST-drrA pGEX encoding gst-drrA In this study pGSTrab1A pGEX encoding gst-rab1 6 pGSTrab1Adn pGEX encoding gst-rab1 S25N 6 pGSTrab1Aca pGEX encoding gst-rab1 Q70L 6 pGSTrab2 pGEX encoding gst-rab2 7 pGSTrab5 pGEX encoding gst-rab5 from A. Wandinger-
Ness
pGSTrab6 pGEX encoding gst-rab6 7 pGSTrab7 pGEX encoding gst-rab7 from A. Wandinger-
Ness
pGSTrab18 pGEX encoding gst-rab18 7 pGSTrab35 pGEX encoding gst-rab35 7 pGFP-drrA61-647 pEGFP encoding gfp-drrA61-647 In this study pGFP-drrA451-64 pEGFP encoding gfp-drrA451-647 In this study pEGFP GFP expression vector Clontech pmRFP-C1 monomeric RFP expression vector From W. Moss pmRFP-N1 monomeric RFP expression vector From W. Moss pRFP-rab1 pmRFP encoding rfp-rab1 In this study pgalT-RFP pmRFP encoding galT-rfp In this study pSR47 Suicide vector (Kanr, SacB) 8 pSR47S Gene replacement vector 8 pSRdrrA Gene replacement vector of drrA In this study pSRlidA Gene replacement vector of lidA In this study pMEnt Mariner plasmid 9 pKH5002 Suicide vector 10 pNH3503 pKH5002 derivative encoding mariner transposon In this study Primers Sequence (5’-3’) Restriction enzyme Resulting
plasmids CyaDrrA1 CGCGGATCCTTATGAGCATAATGGGGAGAATTAAAATG BamHI pcya-drrA, pdCyaDrrA2 AAACTGCAGTATCTTCATAATTGAAATGCGCTGG PstI 17B5N CGCGGATCCTGAGTTTGATCATTGGCGGGGG BamHI pcya-drrB92-25
© 2006 Nature Publishing Group

17B5C AAACTGCAGACGCTTACATAACCATAATTCATGC PstI 36A4N CGCGGATCCAGATGAAACTGAAGAATAAAGTCGC BamHI pcya-drrC 36A4C AAACTGCAGCAATATGGCGATACTCAATCTGTC PstI 48E1N CGCGGATCCTTATGGTTTTACTTTGGATTAACCTACA BamHI pcya-drrD 48E1C AAACTGCAGTACAGAAATTAGAATTACTCTCGCC PstI 51C1N CGCGGATCCTTATGATTTTTACTTCTTATGCATCCTTA BamHI pcya-drrE66-26
51C1C AAACTGCAGGCAATTGGATTAATATTTGGATAGGT PstI 38G5N CGCGGATCCTTATGGAAAATGAAGAAAAATCACTTAAAA BamHI pcya-lspE 38G5C AAACTGCAGTAATATCTTGAAGATGTGGTGTGAC PstI 52B12N CGCGGATCCTTATGTTTAGTGCGGGTATGTCCATG BamHI pcya-lvgA98-20
52B12C AAACTGCAGCTGCGTTACGTTGATAACTGA PstI CyaLidA1 CGCGGATCCAAGACTATAGCCAATCTAAACATGAT BamHI pcya-lidA378-7
CyaLidA2 AAACTGCAGGTAGGGAGATGAAAGATAATTCTATC PstI Bam1b CGCGGATCCCATGAGCATAATGGGGAGAATTAAAATG BamHI Forward primBam201b CGCGGATCCGTTACGAGATAAAATCAAACAAGAACC BamHI for drrA
truncation Bam451b CGCGGATCCAAATGAGCGAGCAACCCCTGATC BamHI Bam546b CGCGGATCCTACGGCAAGCACGGAAAACTTTAAA BamHI Bam200b CGCGGATCCTTACTCTTCTCCTAATTCATAGCGAGG BamHI Reverse primBam365b CGCGGATCCTTAGATGTTACCCTTTTCAAGCATGCT BamHI for drrA
truncation Bam448b CGCGGATCCTTATAATTTACAATCCATAAAGAATGCTGA BamHI Bam500b CGCGGATCCTTAAGCAGCAATTTTATGATTCATCAAAG BamHI Bam647b CGCGGATCCTATCTTCATAATTGAAATGCGCTGG BamHI GSTDrrA1 CGCGGATCCATGAGCATAATGGGGAGAATTAAAATG BamHI pGST-drrA GSTDrrA2 CGGAATTCCTGCAGTATCTTCATAATTGAAATGCGCTGG EcoRI SalSap GCGGCGTCGACCGGAAGAGCGCCCAATACGC SapI pNH3503 SalXbaNcoTR GGCCCGTCGACTCTAGACCATGGTTGTGTCAGACCGGGGACTTATC SalI Mari5a GACTCTAGACCATGGTTGTGTC - Inverse PCRMari7a ATGCCTCTTCCGACCATCAAGC - sequencing LidA1 GCGAGCTCCACATAGATAGCAAAGCGGGTGA SacI pSRlidA LidA2 TAATTCTATCCATAAGTAGGATCCTTAATCAATCG - LidA3 CCTACTTATGGATAGAATTATCTTTCATCTCCCTA - LidA4 GCTCTAGAGCACTTGTTTAAATCCAATATTCGC XbaI DrrA1 GCGAGCTCTAGTATGAATGAGTGATTCGGTAAC SacI pSRdrrA DrrA2 AATGCGCTGGCATGTACTCGAGGATTCATAAGTC - DrrA3 CGAGTACATGCCAGCGCATTTCAATTATGAAGATA - DrrA4 GCTCTAGAAGGAATACCCCATCTTATACAAGC XbaI GalT1 GGAATTCGCATGAGGCTTCGGGAGCCGCTC EcoRI pgalt-RFP GalT2 GCTCCTCGCCCTTGCTCACC - pgalt-RFP Forward primers were listed first followed by reverse primers. a. These two primers were used for inverse PCR and sequencing to determine the loci of transposon insertion. b. A series of drrA truncation were constructed by PCR with combination of these primers
References: 1. Berger, K.H. & Isberg, R.R. Two distinct defects in intracellular growth
complemented by a single genetic locus in Legionella pneumophila. Mol Microbiol 7, 7-19 (1993).
2. Zuckman, D.M., Hung, J.B. & Roy, C.R. Pore-forming activity is not sufficient for Legionella pneumophila phagosome trafficking and intracellular growth. Mol Microbiol 32, 990-1001 (1999).
3. Morales, V.M., Backman, A. & Bagdasarian, M. A series of wide-host-range low-copy-number vectors that allow direct screening for recombinants. Gene 97, 39-47 (1991).
4. Coers, J. et al. Identification of Icm protein complexes that play distinct roles in the biogenesis of an organelle permissive for Legionella pneumophila intracellular growth. Mol Microbiol 38, 719-36 (2000).
© 2006 Nature Publishing Group

5. Nagai, H. et al. A C-terminal translocation signal required for Dot/Icm-dependent delivery of the Legionella RalF protein to host cells. Proc Natl Acad Sci U S A 102, 826-31 (2005).
6. Satoh, A., Wang, Y., Malsam, J., Beard, M.B. & Warren, G. Golgin-84 is a rab1 binding partner involved in Golgi structure. Traffic 4, 153-61 (2003).
7. Delprato, A., Merithew, E. & Lambright, D.G. Structure, exchange determinants, and family-wide rab specificity of the tandem helical bundle and Vps9 domains of Rabex-5. Cell 118, 607-17 (2004).
8. Merriam, J.J., Mathur, R., Maxfield-Boumil, R. & Isberg, R.R. Analysis of the Legionella pneumophila fliI gene: intracellular growth of a defined mutant defective for flagellum biosynthesis. Infect Immun 65, 2497-501 (1997).
9. Rubin, E.J. et al. In vivo transposition of mariner-based elements in enteric bacteria and mycobacteria. Proc Natl Acad Sci U S A 96, 1645-50 (1999).
10. Ohmori, H. et al. The pcsA gene is identical to dinD in Escherichia coli. J Bacteriol 177, 156-65 (1995).
© 2006 Nature Publishing Group

Supplementary Information Methods: Plasmid construction. The plasmid pNH3503 used for the delivery of the mariner minitransposon was constructed in two sequential steps. First the DNA fragment containing the mariner transposase and a minimal terminal repeat of mariner minitransposon was PCR amplified from plasmid pMEnt 1 as a template using the combination of primers SalSap and SalXbaNcoTR. The generated PCR fragment was digested with SapI (blunted) and SalI, and subcloned into SacI (blunted)/SalI sites of a suicide vector pKH5002 2. Next, the mariner mini-transposon was reconstructed by inserting the SalI/BsaA1 fragment of pMEnt into SalI/HindIII (blunted) sites of pKH5002. The resulting plasmid contains the mariner transposase and a modified mariner mini-transposon with a minimal terminal repeat followed by the unique restriction enzyme sites, NcoI/XbaI/SalI. This plasmid was designated as pNH3503. L. pneumophila genes were amplified and fused C-terminally to the Cya protein in the vector pCya-RalF, where the DNA fragment encoding RalF was removed and replaced with the DNA fragment encoding the indicated L. pneumophila protein. M45-tagged derivatives of DrrA, LidA and RalF were constructed in the plasmid pMMB207-M45NT. GST fusion proteins were constructed in plasmid pGEX-2TK. His-tagged fusion proteins were constructed in the plasmid pET15b. Mammalian expression vectors encoding N-terminal GFP fusions to DrrA were constructed using pEGFP-C3 (Clontech), in which the fragments were inserted into the BglII restriction site. Screening L. pneumophila transposon mutants for Rab1 recruitment defects. Plasmid pNH3503, was transformed into wild-type L. pneumophila LP01 (CR39) by electroporation and independently-derived transposon mutants were isolated by plating bacteria on CYE agar plates containing kanamycin. Initial screenings were performed using a 96-well plate format. Using a sterile toothpick, individual transposon mutants were selected, transferred to a single well, and cultivated at 37 C. for two days in 96-well plates containing CYE agar and kanamycin. Roughly 12 hours before infection, murine BMMs were added to glass-bottom 96-well plates (96 MicroWellTM Optical Bottom Plates, Nalgenunc) at a density of 2x104 cells per well. Each well of the glass-bottom plate was infected with a different transposon mutant. This was done by inserting a 96-pin replicator into the agar of a plate of arrayed L. pneumophila mutants and resuspending the bacteria attached to each pin in the medium of the 96-well glass-bottom plate. The L. pneumophila transposon mutants were co-incubated with BMMs in the glass-bottom plates for 30 min at 37 C., and then the wells were washed with PBS to remove uninternalized bacteria.
After infection, cells in each well were fixed with 4% paraformaldehyde in PBS for 20 min and permeabilized with ice-cold methanol for 1 min. PBS containing 2% goat serum (GIBCO) was added to each well and the plates were incubated for 1 hr at room temperature. Rab1 protein was stained by adding to each well 40 µl of rabbit anti-Rab1b antibody (Santa Cruz Biotech. Inc.) diluted 1:100 into PBS containing 2% goat serum for 1 hr at room temperature. After the wells were washed with PBS, rabbit anti-Rab1b was detected by adding to each well 40 µl of rhodamine-conjugated anti-rabbit IgG secondary antibody (Molecular Probes) diluted 1:100 into PBS containing 2% goat serum for 1 hr at room temperature. The cells were stained with 0.1 µg/ml 4,6-diamidino-2-phenylindole (DAPI, Sigma) in PBS for 10 min. PBS was added to cover the cells in each well and the plate was mounted onto a Nikon Eclipse TE300 inverted microscope. Random fields from each well were visualized by fluorescence microscopy using a CFI-
© 2006 Nature Publishing Group

Plan Apo 100X/1.4na oil immersion lens and mutants with a suspected Rab1 recruitment defect were identified based on wells that showed an empirical difference in Rab1 staining of LCV. A total of 3,948 independently-derived mutants were analyzed in the initial screen. A total of 441 mutants were selected for additional screening based on an apparent decrease in the number of Rab1-positive LCV that could be identified in random fields.
Secondary screening and quantitative analysis of L. pneumophila mutants was conducted using BMMs incubated overnight on glass coverslips in 24 well plates at a density of 2x105 cells per well. For these assays, individual transposon mutants were grown on CYE agar plates for 48 hours and resuspended in sterile distilled water. L. pneumophila was added to the well at a MOI of 50. Plates were centrifuged for 5 min at 150g and placed in a 37 C waterbath for 5 min to promote bacterial internalization. The cells were washed with PBS to remove extracellular bacteria and fresh medium was added to each well. The plates were incubated for 25 min in a 5% CO2 incubator at 37 C. Cells were fixed and stained as in the initial screening. For each mutant, 50 vacuoles were scored for Rab1 colocalization and these numbers were compared to the Rab1 recruitment numbers obtained for an internal control well on the same plate that was infected with wild type L. pneumophila. Confocal images were acquired with a Zeiss LSM 510 laser-scanning confocal microscope, exported as TIFF files and labeled in Adobe Illustrator (Adobe). LAMP-1 staining was performed as described previously 3. Construction of L. pneumophila ∆drrA and ∆lidA mutant strains. In-frame deletions in the drrA and lidA genes were constructed in Lp01 using allelic exchange, as described previously 4. Briefly, PCR was used to generate DNA fragments consisting of 5' and 3' regions of homology for each gene. The corresponding 5' and 3' DNA fragments were ligated together in the gene replacement vector pSR47S, resulting in a deletion allele having the start codon of the gene of interest fused in-frame to the stop codon from that same gene. Plasmids encoding the deletion alleles were mated into Lp01 and bacteria in which the plasmid had co-integrated onto the chromosome were selected on CYE agar plates containing Kan. After the strains were passaged once non-selectively, sucrose selection was used to identify clones in which the co-integrated plasmid had been lost. PCR analysis was used to identify clones in which the deletion allele had replaced the wild type allele on the chromosome and confirmed by immunoblot analysis using a rabbit polyclonal antibody generated against the full-length DrrA protein. DotA secretion assay L. pneumophila transposon mutants were grown for 2 days on CYE media with Kan. Bacterial cells were collected from the CYE plates and resuspended in AYE broth at OD600 0.3. After cultivation for 20 hours at 37° C, bacterial cells were collected by centrifugation. Cell-free culture supernatants were prepared by centrifugation followed by filtration through 0.22 µm filter (Ultrafree-MC, MILLIPORE). Protein in the culture supernatant were precipitated in 10% trichloroacetic acid. Cell pellets and precipitated supernatant proteins were dissolved in SDS-PAGE sample buffer by vortexing and separated by SDS-PAGE. The presence of DotA protein in the cellular lysates and culture supernatant was assessed by immunoblot analysis using a polyclonal rabbit anti-DotA antibody, as described previously 4. Determining the site of insertion of the mariner transposon. Genomic DNA from individual L. pneumophila transposon mutants purified and digested to completion using either the restriction enzymes Sau3AI or HindIII. The linear DNA
© 2006 Nature Publishing Group

fragments were circularized by incubation with T4 DNA ligase (New England Biolabs). The circularized DNA was amplified by inverse PCR using the primers mari5 and mari7. PCR products were purified and served as template for sequencing using the mari5 primer. The drrA sequence was deposited into GenBank under the accession number AY945933. Translocation assay Translocation of Cya fusion protein from L. pneumophila into CHO FcγRII cells was assayed as described previously 5,6. Briefly, CHO FcγRII cells were added to 48-well tissue culture plates at a density of 4x104 cells per well. L. pneumophila producing the indicated Cya fusion proteins were added to the CHO FcγRII cells at an MOI of 30 for 1 hr. Cells were washed with PBS then lysed in 50 mM HCl, 0.1 % Triton X-100 solution. The pH of the samples was neutralized with NaOH prior to boiling and extraction with ethanol. Cyclic AMP (cAMP) levels were measured using cAMP Biotrak enzyme immunoassay system (Amersham Bioscience RPN-225). DrrA staining of infected macrophages by immunofluorescence microscopy was conducted using rabbit polyclonal antiserum generated against the full-length DrrA protein, as described previously 7. Intracellular growth assay BMMs or A. castellanii were plated at a density of 2x105 cells per well in 24-well plates. The host cells were infected with the indicated strain of L. pneumophila at a MOI of 10. After 1 hr of infection, wells were washed with PBS to remove extracellular L. pneumophila and fresh medium was added to each well. At the indicated times post-infection, bacterial counts were determined for individual wells by combining the supernatant from each well with a hypotonic lysate containing intracellular bacteria, and plating dilutions on CYE agar plates to measure colony forming units (CFUs). Each data point represents the average number of bacteria from 3 independent wells. References: 1. Rubin, E.J. et al. In vivo transposition of mariner-based elements in enteric
bacteria and mycobacteria. Proc Natl Acad Sci U S A 96, 1645-50 (1999). 2. Ohmori, H. et al. The pcsA gene is identical to dinD in Escherichia coli. J
Bacteriol 177, 156-65 (1995). 3. Roy, C.R., Berger, K.H. & Isberg, R.R. Legionella pneumophila DotA protein is
required for early phagosome trafficking decisions that occur within minutes of bacterial uptake. Mol Microbiol 28, 663-74 (1998).
4. Nagai, H. & Roy, C.R. The DotA protein from Legionella pneumophila is secreted by a novel process that requires the Dot/Icm transporter. Embo J 20, 5962-70 (2001).
5. Sory, M.P. & Cornelis, G.R. Translocation of a hybrid YopE-adenylate cyclase from Yersinia enterocolitica into HeLa cells. Mol Microbiol 14, 583-94 (1994).
6. Nagai, H. et al. A C-terminal translocation signal required for Dot/Icm-dependent delivery of the Legionella RalF protein to host cells. Proc Natl Acad Sci U S A 102, 826-31 (2005).
7. Nagai, H., Kagan, J.C., Zhu, X., Kahn, R.A. & Roy, C.R. A bacterial guanine nucleotide exchange factor activates ARF on Legionella phagosomes. Science 295, 679-82 (2002).
© 2006 Nature Publishing Group














![Solution Structure of the Na+ form of the Dimeric Guanine Quadruplex [d(G3T4G3)]2](https://static.fdokumen.com/doc/165x107/6318f44265e4a6af370f95cf/solution-structure-of-the-na-form-of-the-dimeric-guanine-quadruplex-dg3t4g32.jpg)





