
ARF-GEP100, a guanine nucleotide-exchange protein for ADP-ribosylation factor 6
6
ARF-GEP 100 , a guanine nucleotide-exchange protein for ADP-ribosylation factor 6 Akimasa Someya* , Makoto Sata* ‡ , Kazuyo Takeda § , Gustavo Pacheco-Rodriguez*, Victor J. Ferrans § , Joel Moss*, and Martha Vaughan* *Pulmonary–Critical Care Medicine Branch and § Pathology Section, National Heart, Lung, and Blood Institute, National Institutes of Health, Bethesda, MD 20892 Contributed by Martha Vaughan, December 29, 2000 A human cDNA encoding an 841-aa guanine nucleotide-exchange protein (GEP) for ADP-ribosylation factors (ARFs), named ARF-GEP 100 , which contains a Sec7 domain, a pleckstrin homology (PH)-like do- main, and an incomplete IQ-motif, was identified. On Northern blot analysis of human tissues, a ’8-kb mRNA that hybridized with an ARF-GEP 100 cDNA was abundant in peripheral blood leukocytes, brain, and spleen. ARF-GEP100 accelerated [ 35 S]GTPgS binding to ARF1 (class I) and ARF5 (class II) 2- to 3-fold, and to ARF6 (class III) ca. 12-fold. The ARF-GEP 100 Sec7 domain contains Asp 543 and Met 555 , correspond- ing to residues associated with sensitivity to the inhibitory effect of the fungal metabolite brefeldin A (BFA) in yeast Sec7, but also Phe 535 and Ala 536 , associated with BFA-insensitivity. The PH-like domain differs greatly from those of other ARF GEPs in regions involved in phospholipid binding. Consistent with its structure, ARF-GEP 100 ac- tivity was not affected by BFA or phospholipids. After subcellular fractionation of cultured T98G human glioblastoma cells, ARF6 was almost entirely in the crude membrane fraction, whereas ARF-GEP 100 , a 100-kDa protein detected with antipeptide antibodies, was cytoso- lic. On immunofluorescence microscopy, both proteins had a punctate pattern of distribution throughout the cells, with apparent colocal- ization only in peripheral areas. The coarse punctate distribution of EEA-1 in regions nearer the nucleus appeared to coincide with that of ARF-GEP 100 in those areas. No similar coincidence of ARF-GEP 100 with AP-1, AP-2, catenin, LAMP-1, or 58K was observed. The new human BFA-insensitive GEP may function with ARF6 in specific endocytic processes. F ormation of vesicles during intracellular protein trafficking requires activation and membrane association of GTPases, including ADP-ribosylation factors (ARFs) and Sar1 (1– 4). ARFs, ’20-kDa guanine nucleotide-binding proteins, participate in the initiation of COPI- and clathrin-coated vesicles at the Golgi, trans-Golgi network, and plasma membrane (1). The role of ARFs as modulators of vesicle formation may be both structural, e.g., coatomer binding, and catalytic, e.g., activation of phospholipase D (5–8) andyor phosphoinositide kinases (9). Activation of ARF- GDP is favored by the enzymatic action of guanine nucleotide- exchange proteins (GEPs); the cycle of ARF action is closed by its inactivation with the help of GTPase-activating proteins (2, 10). Among the several ARF-GEPs are the cytohesins, cytohesin-1, -2, -3, and -4 (11–14), and EFA6 (15); both families are insensitive to the inhibitory effects of the fungal metabolite, brefeldin A (BFA). The initial observation of BFA-sensitive ARF GEP activity lead to the cloning and characterization of BFA-inhibitable mam- malian BIG1 and BIG2 (16, 17), and yeast Gea1y2 (18), as well as recognition of the activity of the yeast Sec7, which had been identified earlier (19). Biochemically, ARF GEPs accelerate gua- nine nucleotide binding to ARFs with a phospholipid requirement for activity (13), leading to the proposal that membrane association of GEPs and ARFs is required for guanine nucleotide exchange (20, 21). Additional regulation of ARF GEP function is poorly under- stood, but local turnover of phosphoinositides may be important in their activation and intracellular localization (22). The ARF GEPs are modular proteins that contain a Sec7 domain, first recognized in the yeast Sec7 protein, which was identified by its role in protein secretion (19, 23). Proteins contain- ing Sec7 domains are found in multi- and unicellular organisms, suggesting a pivotal role in intracellular transport and homeostasis (22). The Sec7 domains of BFA-insensitive cytohesin-2 (ARNO) (24) and cytohesin-1 (25), as well as the BFA-sensitive Gea2 (26), show very similar arrangements of 10 a-helices grouped in two sets of five, which form a hydrophobic groove that is conserved in all ARF GEPs. The sequences of motifs 1 and 2 in the Sec7 domain, which are highly conserved among ARF GEPs, contain all of the major residues involved in ARF interactions, as demonstrated by cocrystallization of nucleotide-free ARF and a Sec7 domain (26). Of note, Motif 1 contains a conserved critical glutamic acid involved in catalysis of guanine nucleotide exchange (27). The Sec7 domain also includes regions that confer BFA sensitivity (28, 29). Among the functional structures found in ARF GEPs is the pleckstrin homology (PH) domain, required for phosphoinositide binding and membrane association by the cytohesin family and EFA6 (22). The PH domain is a protein module of approximately 120 amino acids, which generates an orthogonal fold of several b-sheets and a C-terminal a-helix (30). Amino acid sequences of the PH domains of EFA6 and cytohesin-1 are 26% identical, but, among the cytohesin family, they are ’80% identical. Binding of specific phosphoinositides by cytohesins differs, however, as revealed by biochemical assays (13) and crystallization of the PH domain of GRP1 (cytohesin-3 or ARNO3) (31). ARFs are members of the ras superfamily that can be classified into class I (ARF1, -2, -3), class II (ARF4, -5), and class III (ARF6) based on size, sequence identity, and gene structure (2, 32). The activation of each ARF depends on a specific interaction and concomitant localization with a GEP. In vitro studies have focused on the identification of ARF GEPs, based on rates of guanine nucleotide exchange, intracellular localization, and yeast two- hybrid screening. Almost all ARF GEPs are able to increase guanine nucleotide binding to class 1 ARFs (ARF1). GBF1 was shown to increase preferentially guanine nucleotide binding to ARF5 (33), and ARF6 appears to be the best substrate for EFA6 (15) and cytohesin-2 (34). Here, we describe a modular protein with ARF GEP activity that contains an IQ-like motif, and Sec7 and PH-like domains. The widely distributed ARF-GEP 100 preferentially increased guanine nucleotide binding to ARF6 in vitro. In cells, endoge- nous ARF-GEP 100 appeared to colocalize in part with ARF6 and in part with the endosomal protein EEA1, consistent with involvement in the activation of ARF6 during endocytosis. Abbreviations: ARF, ADP-ribosylation factor; GEP, guanine nucleotide-exchange protein; BFA, brefeldin A; GTPgS, guanosine 59-g-(thio) triphosphate; PS, phosphatidyl serine; PIP2, phosphatidylinositol bis-4,5-phosphate; PIP3, phosphatidylinositol tris-3,4,5-phosphate; PH, pleckstrin homology; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; CaM, calmodulin. Data deposition: The sequence reported in this paper has been deposited in the GenBank database (accession no. AB018306). To whom reprint requests should be addressed. E-mail: [email protected]. ‡ Present address: Yamagata University School of Medicine, 2-2-2 Iida-Nishi, Yamagata, Japan 990-9585. The publication costs of this article were defrayed in part by page charge payment. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. §1734 solely to indicate this fact. www.pnas.orgycgiydoiy10.1073ypnas.051634798 PNAS u February 27, 2001 u vol. 98 u no. 5 u 2413–2418 CELL BIOLOGY
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of ARF-GEP100, a guanine nucleotide-exchange protein for ADP-ribosylation factor 6

ARF-GEP100, a guanine nucleotide-exchange proteinfor ADP-ribosylation factor 6Akimasa Someya*†, Makoto Sata*‡, Kazuyo Takeda§, Gustavo Pacheco-Rodriguez*, Victor J. Ferrans§, Joel Moss*,and Martha Vaughan*
*Pulmonary–Critical Care Medicine Branch and §Pathology Section, National Heart, Lung, and Blood Institute, National Institutes of Health, Bethesda, MD 20892
Contributed by Martha Vaughan, December 29, 2000
A human cDNA encoding an 841-aa guanine nucleotide-exchangeprotein (GEP) for ADP-ribosylation factors (ARFs), named ARF-GEP100,which contains a Sec7 domain, a pleckstrin homology (PH)-like do-main, and an incomplete IQ-motif, was identified. On Northern blotanalysis of human tissues, a '8-kb mRNA that hybridized with anARF-GEP100 cDNA was abundant in peripheral blood leukocytes,brain, and spleen. ARF-GEP100 accelerated [35S]GTPgS binding to ARF1(class I) and ARF5 (class II) 2- to 3-fold, and to ARF6 (class III) ca. 12-fold.The ARF-GEP100 Sec7 domain contains Asp543 and Met555, correspond-ing to residues associated with sensitivity to the inhibitory effect ofthe fungal metabolite brefeldin A (BFA) in yeast Sec7, but also Phe535
and Ala536, associated with BFA-insensitivity. The PH-like domaindiffers greatly from those of other ARF GEPs in regions involved inphospholipid binding. Consistent with its structure, ARF-GEP100 ac-tivity was not affected by BFA or phospholipids. After subcellularfractionation of cultured T98G human glioblastoma cells, ARF6 wasalmost entirely in the crude membrane fraction, whereas ARF-GEP100,a 100-kDa protein detected with antipeptide antibodies, was cytoso-lic. On immunofluorescence microscopy, both proteins had a punctatepattern of distribution throughout the cells, with apparent colocal-ization only in peripheral areas. The coarse punctate distribution ofEEA-1 in regions nearer the nucleus appeared to coincide with that ofARF-GEP100 in those areas. No similar coincidence of ARF-GEP100 withAP-1, AP-2, catenin, LAMP-1, or 58K was observed. The new humanBFA-insensitive GEP may function with ARF6 in specific endocyticprocesses.
Formation of vesicles during intracellular protein traffickingrequires activation and membrane association of GTPases,
including ADP-ribosylation factors (ARFs) and Sar1 (1–4). ARFs,'20-kDa guanine nucleotide-binding proteins, participate in theinitiation of COPI- and clathrin-coated vesicles at the Golgi,trans-Golgi network, and plasma membrane (1). The role of ARFsas modulators of vesicle formation may be both structural, e.g.,coatomer binding, and catalytic, e.g., activation of phospholipase D(5–8) andyor phosphoinositide kinases (9). Activation of ARF-GDP is favored by the enzymatic action of guanine nucleotide-exchange proteins (GEPs); the cycle of ARF action is closed by itsinactivation with the help of GTPase-activating proteins (2, 10).
Among the several ARF-GEPs are the cytohesins, cytohesin-1,-2, -3, and -4 (11–14), and EFA6 (15); both families are insensitiveto the inhibitory effects of the fungal metabolite, brefeldin A(BFA). The initial observation of BFA-sensitive ARF GEP activitylead to the cloning and characterization of BFA-inhibitable mam-malian BIG1 and BIG2 (16, 17), and yeast Gea1y2 (18), as well asrecognition of the activity of the yeast Sec7, which had beenidentified earlier (19). Biochemically, ARF GEPs accelerate gua-nine nucleotide binding to ARFs with a phospholipid requirementfor activity (13), leading to the proposal that membrane associationof GEPs and ARFs is required for guanine nucleotide exchange (20,21). Additional regulation of ARF GEP function is poorly under-stood, but local turnover of phosphoinositides may be important intheir activation and intracellular localization (22).
The ARF GEPs are modular proteins that contain a Sec7domain, first recognized in the yeast Sec7 protein, which wasidentified by its role in protein secretion (19, 23). Proteins contain-
ing Sec7 domains are found in multi- and unicellular organisms,suggesting a pivotal role in intracellular transport and homeostasis(22). The Sec7 domains of BFA-insensitive cytohesin-2 (ARNO)(24) and cytohesin-1 (25), as well as the BFA-sensitive Gea2 (26),show very similar arrangements of 10 a-helices grouped in two setsof five, which form a hydrophobic groove that is conserved in allARF GEPs. The sequences of motifs 1 and 2 in the Sec7 domain,which are highly conserved among ARF GEPs, contain all of themajor residues involved in ARF interactions, as demonstrated bycocrystallization of nucleotide-free ARF and a Sec7 domain (26).Of note, Motif 1 contains a conserved critical glutamic acid involvedin catalysis of guanine nucleotide exchange (27). The Sec7 domainalso includes regions that confer BFA sensitivity (28, 29). Amongthe functional structures found in ARF GEPs is the pleckstrinhomology (PH) domain, required for phosphoinositide binding andmembrane association by the cytohesin family and EFA6 (22). ThePH domain is a protein module of approximately 120 amino acids,which generates an orthogonal fold of several b-sheets and aC-terminal a-helix (30). Amino acid sequences of the PH domainsof EFA6 and cytohesin-1 are 26% identical, but, among thecytohesin family, they are '80% identical. Binding of specificphosphoinositides by cytohesins differs, however, as revealed bybiochemical assays (13) and crystallization of the PH domain ofGRP1 (cytohesin-3 or ARNO3) (31).
ARFs are members of the ras superfamily that can be classifiedinto class I (ARF1, -2, -3), class II (ARF4, -5), and class III (ARF6)based on size, sequence identity, and gene structure (2, 32). Theactivation of each ARF depends on a specific interaction andconcomitant localization with a GEP. In vitro studies have focusedon the identification of ARF GEPs, based on rates of guaninenucleotide exchange, intracellular localization, and yeast two-hybrid screening. Almost all ARF GEPs are able to increaseguanine nucleotide binding to class 1 ARFs (ARF1). GBF1 wasshown to increase preferentially guanine nucleotide binding toARF5 (33), and ARF6 appears to be the best substrate for EFA6(15) and cytohesin-2 (34).
Here, we describe a modular protein with ARF GEP activitythat contains an IQ-like motif, and Sec7 and PH-like domains.The widely distributed ARF-GEP100 preferentially increasedguanine nucleotide binding to ARF6 in vitro. In cells, endoge-nous ARF-GEP100 appeared to colocalize in part with ARF6 andin part with the endosomal protein EEA1, consistent withinvolvement in the activation of ARF6 during endocytosis.
Abbreviations: ARF, ADP-ribosylation factor; GEP, guanine nucleotide-exchange protein;BFA, brefeldin A; GTPgS, guanosine 59-g-(thio) triphosphate; PS, phosphatidyl serine; PIP2,phosphatidylinositol bis-4,5-phosphate; PIP3, phosphatidylinositol tris-3,4,5-phosphate;PH, pleckstrin homology; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; CaM,calmodulin.
Data deposition: The sequence reported in this paper has been deposited in the GenBankdatabase (accession no. AB018306).
†To whom reprint requests should be addressed. E-mail: [email protected].
‡Present address: Yamagata University School of Medicine, 2-2-2 Iida-Nishi, Yamagata,Japan 990-9585.
The publication costs of this article were defrayed in part by page charge payment. Thisarticle must therefore be hereby marked “advertisement” in accordance with 18 U.S.C.§1734 solely to indicate this fact.
www.pnas.orgycgiydoiy10.1073ypnas.051634798 PNAS u February 27, 2001 u vol. 98 u no. 5 u 2413–2418
CELL
BIO
LOG
Y

Materials and MethodsMaterials. cDNA encoding KIAA0763 (GenBank accession no.AB018306) was kindly provided by T. Nagase (Kazusa DNAResearch Institute, Chiba, Japan). BL21 (DE3) Singles competentcells and pET-30 LIC vector were purchased from Novagen. Sf9insect cells were from Invitrogen. Baculovirus transfer vectorpAcHLT-C and linearized BaculoGold were from Becton Dickin-son. Plasmid purification kits and Ni21-nitrilotriacetic acid agarosewere from Qiagen (Chatsworth, CA). CONCERT Rapid GelExtraction System, SF-900 II SFM, MEM, and FBS were from LifeTechnologies (Grand Island, NY). Pfu DNA polymerase was fromStratagene. Rapid DNA Ligation kit, PCR Nucleotide mix, andrestriction enzymes were from Roche Molecular Biochemicals.[35S]-guanosine 59-g-(thio) triphosphate (GTPgS) (1250 Ciymmol;1 Ci 5 37 GBq) and [a-32P]dATP (3,000 Ciymmol) were fromNEN. GTPgS and phosphatidylserine (PS) were from Sigma.Phosphatidylinositol tris-3,4,5-phosphate (PIP3) and phosphatidyl-inositol bis-4,5-phosphate (PIP2) were from Matreya (PleasantGap, PA). Human brain calmodulin was from Calbiochem.
Cell Culture. Sf9 cells (Invitrogen) were grown in Sf-900 IImedium supplemented with 10% FBS and gentamycin (50mgyml) at 27°C. Human glioblastoma T98G cells, purchasedfrom American Type Culture Collection, were grown in MEMsupplemented with 10% FBS, nonessential amino acids (100 mMeach), penicillin G (100 unitsyml), and streptomycin (100 mgyml)at 37°C with an atmosphere of 5% CO2.
Preparation of Recombinant ARFs. DNA constructs for hARF1,hARF5, hARF6, and yeast ARF2 were prepared in the pET7vector and purified as described (35, 36). Recombinant ARFproteins were obtained as follows. A single colony expressingrecombinant protein was incubated overnight at 37°C in 25 ml ofLB medium containing ampicillin (100 mgyml), which was addedto 500 ml of the same medium. Incubation with 1 mM isopro-pyl-1- thio-b-D-galactopyranoside was started at an A600 of 0.6.Cells were harvested 2 h later and incubated with lysozyme (0.5mM) in TE buffer, followed by sonication. After centrifugation(100,000 3 g, 35 min), the supernatant was applied to a column(2.5 3 100 cm) of Ultrogel AcA 54, which was eluted with bufferA (20 mM TriszHCl, pH 8.0, 1 mM EDTA, 1 mM NaN3, 200 mMsucrose, 100 mM NaCl, 5 mM MgCl2). Fractions containingARF activity were pooled and stored in small portions at 280°C.
Preparation of ARF-GEP100 by BaculovirusyInsect Cell System. Se-quence encoding ARF-GEP100 was amplified from KIAA0763cDNA in pBluescript II SK(1) vector by PCR using the forwardprimer 59-ACGTACTCCATATGCATGCTAGAACGAAA-GTATGGGGGGCGCCT-39 (underlined sequence is a NdeIrestriction site) and reverse primer 59-ACGTACGTGGGTACC-GTCCTCGGGTCCCATGGCTTAGGAGCACAGCACTGA-39(underlined sequence is a KpnI restriction site). The PCR productwas gel-purified and subcloned into pCR-Blunt vector. The result-ing plasmid was digested with NdeI and KpnI. After purification byagarose gel electrophoresis, the fragment was ligated into baculo-virus transfer vector pAcHLT-C, which had been treated with NdeIand KpnI (named pAcHLT-Cyencoding ARF-GEP100).
Sf9 cells (70% confluent) cotransfected with transfer vectorpAcHLT-CyARF-GEP100 (3 mg) and linearized BaculoGold Bac-ulovirus DNA (0.5 mg) were incubated for 5 days at 27°C accordingto the manufacturer’s instructions. Viral amplification was per-formed twice to obtain a high viral titer, and the recombinant virusstock solution was stored at 4°C. Recombinant virus was tested forits ability to induce expression of His6-tagged recombinant proteinin infected Sf9 cells by Western blots of cell lysate with anti-Hismonoclonal antibody (data not shown). Monolayers (2.8 3 107 cellsin a 225-cm2 flask) were infected with recombinant baculovirus and
incubated at 27°C for 3–4 days before collection in lysis buffer (50mM sodium phosphate, pH 8.0, 0.5 mM NaCl, 5 mM b2mercap-toethanol, benzamidine, 16 mgyml, with o-phenanthroline, aproti-nine, leupeptin, and pepstatin A, each 10 mgyml, plus 1 mM PMSFand 10% glycerol). After incubation on ice for 1 h followed byfreezing and thawing, cells were sonified. The lysate was centrifuged(100,000 3 g, 50 min) to pellet the cellular debris, and thesupernatant plus 20 mM imidazole was incubated with Ni21-nitrilotriacetic acid agarose at 4°C overnight with constant agita-tion. After washing the matrix extensively with 50 mM sodiumphosphate, pH 8.0, 50 mM imidazole, 1 M NaCl, 5 mM b2mer-captoethanol, 0.5 mM 4-(2-aminoethyl) bensenesulfonyl fluoride,and 10% glycerol, bound protein was eluted with 150 mM imidazolein 50 mM sodium phosphate, pH 8.0, 0.5 M NaCl, and 10% glycerol,and dialyzed overnight against 20 mM TriszHCl, pH 8.0, containing1 mM EDTA,1 mM DTT, 3 mM MgCl2, 30 mM NaCl, 1 mM NaN3,and 10% glycerol. Fractions containing recombinant protein wereidentified by SDSyPAGE, pooled, and stored in small portions at280°C.
Northern Blot Analysis. The DNA probe, a 526-bp PCR productcorresponding to bases 321–847 in the ARF-GEP100 coding region,was labeled with [a-32P]dATP (3000 mCiyml) using RandomPrimer labeling kit (Roche Molecular Biochemicals), and hybrid-ized in ExpressHyb solution (CLONTECH) with a human multipletissue Northern blot (CLONTECH) at 68°C for 16 h. The mem-brane was washed with 23 SSCy0.05% SDS at room temperaturefor 40 min and 0.13 SSCy0.1% SDS at 50°C for 40 min, followedby autoradiography at 280°C. After stripping to remove the labeledprobe, the same blot was hybridized with a glyceraldehyde-3-phosphate dehydrogenase (GAPDH) cDNA.
GTPgS-Binding Assay. [35S]GTPgS binding to purified recombinantARFs was assayed in a total volume of 50 ml, using a rapid filtrationprocedure. Briefly, 25 pmol (0.5 mg) of recombinant human ARFand 4 mM [35S]GTPgS (2.5 3 106 cpm) without or with GEP (2.5pmol unless otherwise indicated), were incubated in assay buffer(20 mM TriszHCl, pH 8.0y1 mM DTTy3 mM MgCl2y1 mM EDTA)with 15 mg of BSA and 5 mg of PS for 20 min at 30°C. Incubationwas terminated by addition of 300 ml of ice-cold washing buffer(25 mM TriszHCl, pH 8.0y5 mM MgCl2y100 mM NaCly1 mMEDTAy1 mM DTT). The mixture was transferred to a nitrocellu-lose filter in a manifold (Millipore) for rapid filtration, followed bywashing six times, each with 2 ml of ice-cold washing buffer.Scintillation fluid was added to dried filters before radioassay ofprotein-bound [35S]GTPgS. Data are reported as means 6 SEM ofvalues from triplicate assays in a representative experiment.
Preparation of Antibody. Rabbits were immunized with a peptide(RARDTEPQTALHGMDHRKLDEMTAC), correspondingto amino acids 153–176 of ARF-GEP100, with the underlinedcysteine added to facilitate coupling to keyhole limpet hemocy-anin. Antibodies were affinity-purified with the peptide used forimmunization coupled to epoxy-activated Sepharose 6B (Am-ersham Pharmacia) as described by the manufacturer. Anti-serum was incubated with bead-bound peptide in TBS for 1 h at4°C, followed by washing of beads with TBS containing 0.5%Triton X-100 and 0.15 M NaCl, and elution of bound IgG with0.1 M glycinezHCl (pH 2.5). The solution was neutralized with1y20 volume of 1 M Tris, and stored at 280°C until use.
Cell Fractionation and Western Blotting. T98G cells (5 3 107) werewashed with ice-cold PBS, collected by scraping, and homogenized(Dounce tissue grinder) in sucrose buffer (20 mM Tris, pH 7.4, 1mM CaCl2, and 250 mM sucrose, containing 1 mM PMSF andbenzamidine, 16 mgyml, plus o-phenanthroline, aprotinin, leupep-tin, and pepstatin A (each 10 mgyml) with 1 mM MgCl2 or 1 mMEDTA). The homogenate was centrifuged (400 3 g, 10 min) to
2414 u www.pnas.orgycgiydoiy10.1073ypnas.051634798 Someya et al.

remove unbroken cells, nuclei, and cell debris; the post nuclearsupernatant was further centrifuged (100,000 3 g, 1.5 h) to separatecytosol and crude membrane fractions. Equivalent samples offractions were subjected to SDSyPAGE in 8% gel and transferredto nitrocellulose membranes. Blots were incubated in TBS con-taining 3% nonfat dry milk and 0.05% thimerosal, and thenantibodies against ARF-GEP100 (0.1 mgyml) or ARF6, followed byhorseradish peroxidase-conjugated goat anti-rabbit (Promega), be-fore development using Super Signal Chemiluminescent substrate(Pierce).
Confocal Immunofluorescence Microscopy. T98G cells were grown on12-mm round glass coverslips for 24 h and at 37°C in 5%CO2, thenfixed with 4% paraformaldehyde in PBS for 20 min at roomtemperature. After washing with PBS, cells were permeabilized inPBS containing 0.1% Triton X-100 for 4 min, washed with PBS, andincubated with blocking buffer (PBS containing 10% goat serumand 3% BSA). After washing with PBS, cells were incubatedovernight at 4°C with primary antibodies diluted in blocking buffer.Anti-ARF-GEP100 was used at final concentration of 0.2 mgyml.Mouse monoclonal antibodies against ARF6 (1y50 dilution; cloneno. 3A-6; Santa Cruz Biotechnology), b-catenin (1y500 dilution;clone no. 7D8; Upstate Biotechnology, Lake Placid, NY), LAMP-1(1y50 dilution; clone no. 25; Transduction Laboratories, Lexington,KY), Golgi 58K protein (1y200 dilution; clone no. 58K-9; Sigma),EEA-1 (1y200 dilution; clone no. 15; Transduction Laboratories),AP-1 (1y200 dilution; clone no.100y3; Sigma), and AP-2 (1y25dilution; clone no. 100y2; Sigma) were used in colocalizationstudies. After washing with PBS, cells were incubated with TexasRed-conjugated goat anti-rabbit IgG (1y200 dilution; Vector Lab-oratories) and FITC-conjugated goat anti-mouse IgG (1y200 dilu-tion; Sigma) for 1 h at room temperature, and washed with PBS.Cover slips were mounted in Vectashield (Vector Laboratories)and inspected with a confocal microscope (Leica TCS-SP).
ResultsIdentification of ARF-GEP100 cDNA. Using DNA sequence of thecytohesin-1 Sec7 domain, a BLAST search of the databaseidentified a Sec7 domain-containing sequence, referred to asARF-GEP100 (Fig. 1A). This cDNA (originally called KIAA0763and derived from human brain) encodes an ORF of 841 aminoacids, with an IQ-like motif near the N terminus, a central Sec7domain (residues 399–590), and a 112-residue PH-like domain(residues 631–742). By structural comparison, it is clearly not amember of the cytohesin family of smaller ('40 kDa) proteinsthat contain Sec7 and PH domains. The predicted protein alsocontains a nuclear localization signal (residues 403–420) in theSec7 domain and a serine-rich region (residues 337–372). Theamino acid sequences of Sec7 domains of ARF-GEP100, humancytohesin-1, and BIG1, which are aligned in Fig. 1B, exhibit onlylimited identity; overall, the ARF-GEP100 Sec7 domain is 27–44% identical to those of other ARF GEPs.
Tissue Distribution of ARF-GEP100 mRNA. On a blot of poly(A)1
RNA from several human tissues, a cDNA probe correspondingin sequence to bases 107–282 of the coding region of theARF-GEP100 cDNA hybridized with a band of '8 kb (Fig. 2). Inbrain, a '7-kb band was also detected. After quantification bydensitometry, when expressed as a fraction of the density of theGAPDH band, the '8-kb mRNA was most abundant in leuko-cytes, brain, and spleen. Lesser amounts were detected in lung,placenta, small intestine, liver, and kidney.
Effect of ARF-GEP100 on GTPgS Binding to ARF1, ARF5, and ARF6.Recombinant His-tagged ARF-GEP100 enhanced GTPgS bind-ing to ARF1, ARF5, and ARF6 in a concentration-dependentmanner (Fig. 3A). Activity with ARF6 was more than five timesthat with ARF1 and about twice that with ARF5. The rate of
GTPgS binding to ARF6 was maximal with about 5 pmol (100nM) of ARF-GEP100 (Fig. 3B). The GEP activities of 2.5 pmol(50 nM) of ARF-GEP100 were similar to those of the sameconcentration of the cytohesin-1 Sec7 domain with ARF1 andARF5 as substrates (Fig. 3B). With ARF6, however, ARF-GEP100 was clearly more effective, accelerating binding .10-fold, with a rate that was essentially constant for 20 min at 30°C.For reasons that are not clear, ARF-GEP100 activity appeared todecline more rapidly in the assays with ARF1 and ARF5 thanARF6 (Fig. 3B). As noted earlier, activity of the cytohesin-1 Sec7domain is unstable in assays at 37°C.
As shown in Fig. 4A, at concentrations that almost completelyinhibited GEP activity of the Sec7 domain of yeast Sec7 acting on
Fig. 1. Predicted structure of ARF-GEP100. (A) Schematic representation ofputative functional domains and location of cDNA probe sequence in ARF-GEP100. (B) Amino acid sequence alignment of Sec7 domain of ARF-GEP100
(p100) with those of cytohesin 1 (C-1) and BIG1. (C) Alignment of Sec7 domainmotif 2 and adjacent sequence from p100, EFA6, ARNO, C-1, yeast Sec7 domain(Sec7), Gea2, and BIG1.
Fig. 2. NorthernblotanalysisofARF-GEP100 mRNAinhumantissues.Ablotwithpoly(A)1 RNA from the indicated tissues was hybridized with the 586-bp ARF-GEP100 (p100) cDNA. After stripping, the blot was hybridized with GAPDH cDNA.
Someya et al. PNAS u February 27, 2001 u vol. 98 u no. 5 u 2415
CELL
BIO
LOG
Y

yeast ARF2, BFA had no effect on the activity of ARF-GEP100 withARF6 as substrate. This was surprising because two amino acids inthe Sec7 domain that had been shown to confer BFA sensitivitywhen introduced into the BFA-insensitive cytohesin-1 (28) arepresent in the predicted ARF-GEP100 sequence (Fig. 1C). It isnotable, however, that two other amino acids (phenylalanine andalanine) nearby in the sequence are those found in BFA-sensitiveGEPs and are replaced by tyrosine and serine in BFA-sensitivemolecules, as reported by Peyroche et al. (29).
Effect of Phospholipids on GEP Activity of ARF-GEP100. GTP bindingto ARFs, with and without GEP, is influenced by phospholipids anddetergents (13, 27, 37). Enhancement of GEP activity by phospho-lipids is perhaps most dramatic with the cytohesin family, in whichthe PH domain is responsible for functional interaction with specificphosphatidylinositol phosphates (12). The GEP activity of ARF-GEP100, however, was unaffected by the addition of PIP2, PIP3, orPS, whether ARF6 or ARF1 was the substrate (Fig. 4B). TheARF-GEP100 PH domain differs significantly from those of thecytohesins, notably in regions believed to be responsible for phos-phatidylinositol phosphate binding. Whether ARF-GEP100 inter-acts with other phospholipids remains to be determined.
Effect of Calcium and Calmodulin on GEP Activity of ARF-GEP100. Thereis an IQ-like sequence (residues 14–24) near the N terminus ofARF-GEP100. The IQ motif is reported to bind to calmodulin(CaM) and serve as a regulatory domain in CaM-binding proteins
(38, 39). In assays of GTPgS binding to ARF6, calcium at concen-trations below 500 mM had little effect without or with 100 nM CaM(Fig. 5), but 1 and 2 mM CaCl2 inhibited binding (without or withCaM), by ca. 35%. Similarly, with 100 mM CaCl2, CaM at concen-trations of up to 400 nM was without effect on GTPgS binding toARF6, without or with ARF-GEP100 (Fig. 5).
Intracellular Localization of ARF-GEP100. T98G cells were homoge-nized in buffer containing 1 mM MgCl2 or 1 mM EDTA. Aftercentrifugal fractionation in either buffer, ARF6 was recoveredalmost entirely in the crude membrane fraction, whereas ARF-GEP100 was detected only in cytosol (Fig. 6).
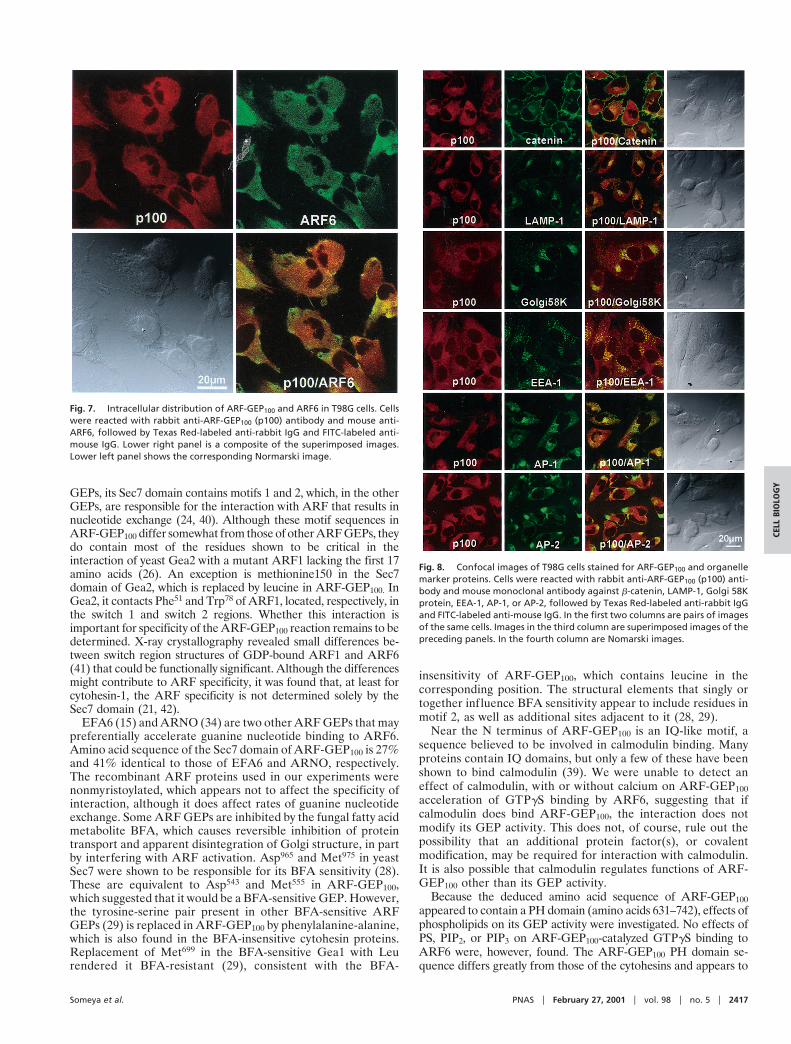
On confocal laser-scanning immunofluorescence microscopy ofT98G cells, both endogenous ARF-GEP100 and ARF6 were dis-tributed in punctate patterns throughout the cytoplasm, with ap-parent colocalization only in restricted areas, for the most part in thecell periphery (Fig. 7). On comparison of the distribution ofARF-GEP100 with that of selective markers to identify subcellularcompartments, only the coarse punctate concentrations of EEA-1immunoreactivity in regions nearer the nucleus appeared to coin-cide with ARF-GEP100. There was no similar coincidence ofARF-GEP100 reactivity with that of AP-1, AP-2, catenin, LAMP-1,or Golgi 58K (Fig. 8). No ARF-GEP100 was detected in nuclei,although its amino acid sequence contains a predicted nuclearlocalization signal.
DiscussionAlthough ARF-GEP100 accelerates GTPgS binding by ARFs of allthree classes, it appears to function preferentially as a GEP forARF6. The deduced amino acid sequence of ARF-GEP100 includesseveral putative functional domains (Fig. 1). Like all known ARF
Fig. 3. Binding of [35S]GTPgS to ARFs 1, 5, and 6. (A) Samples (25 pmol) ofARF1 (F), ARF5 (Œ), or ARF6 (■) were incubated for 20 min at 30°C with 4 mM[35S]GTPgS and the indicated amount of ARF-GEP100. (B) Samples (25 pmol) ofARF1, ARF5, or ARF6 were incubated with 4 mM [35S]GTPgS for indicated timeat 30°C without (E), or with 2.5 pmol of ARF-GEP100 (■) or cytohesin-1 Sec7 (Œ).Data are means 6 SEM of values from triplicate assays. Findings were repli-cated at least three times with different protein preparations.
Fig. 4. Effect of BFA and phospholipids on [35S]GTPgS binding to ARF 6. (A)Samples (25 pmol) of ARF6 plus 2.5 pmol of ARF-GEP100 (■) or yeast ARF2 (25pmol) plus 2.5 pmol of yeast Sec7 (F) were incubated for 4 h at 4°C with 4 mM[35S]GTPgS and the indicated amounts of BFA. (B) Samples (25 pmol) of ARF6or ARF1 were incubated without or with 2.5 pmol of ARF-GEP100 (p100) for 20min at 30°C with 4 mM [35S]GTPgS and the indicated amount of phospholipid(PS, PIP2, PIP3). Data are means 6 SEM of values from triplicate assays. Exper-iments were repeated twice with similar results.
Fig. 5. Effect of calcium and calmodulin on [35S]GTPgS. Samples (25 pmol) ofARF6 were incubated for 20 min at 30°C with 2.5 pmol of ARF-GEP100 and 4 mM[35S]GTPgS without (h) or with (■) 100 nM calmodulin and the indicatedconcentration of CaCl2(Ca21) (Left), or without (E) or without (F) 100 mM CaCl2and the indicated concentration of CaM (Right). Data are means 6 SEM ofvalues from triplicate assays. IQ motif sequence above is compared withsequence of ARF-GEP100 (p100).
Fig. 6. Subcellular distribution of ARF-GEP100 and ARF6. T98G cells werehomogenized in buffer with 1 mM MgCl2 (Mg21) or 1 mM EDTA, and post-nuclear supernatant, crude membrane (M), and cytosol (C) fractions wereprepared. Samples were subjected to SDSyPAGE and immunoblotting withanti-ARF-GEP100 (p100) and anti-ARF6 antibodies. Experiment was replicatedthree times.
2416 u www.pnas.orgycgiydoiy10.1073ypnas.051634798 Someya et al.
GEPs, its Sec7 domain contains motifs 1 and 2, which, in the otherGEPs, are responsible for the interaction with ARF that results innucleotide exchange (24, 40). Although these motif sequences inARF-GEP100 differ somewhat from those of other ARF GEPs, theydo contain most of the residues shown to be critical in theinteraction of yeast Gea2 with a mutant ARF1 lacking the first 17amino acids (26). An exception is methionine150 in the Sec7domain of Gea2, which is replaced by leucine in ARF-GEP100. InGea2, it contacts Phe51 and Trp78 of ARF1, located, respectively, inthe switch 1 and switch 2 regions. Whether this interaction isimportant for specificity of the ARF-GEP100 reaction remains to bedetermined. X-ray crystallography revealed small differences be-tween switch region structures of GDP-bound ARF1 and ARF6(41) that could be functionally significant. Although the differencesmight contribute to ARF specificity, it was found that, at least forcytohesin-1, the ARF specificity is not determined solely by theSec7 domain (21, 42).
EFA6 (15) and ARNO (34) are two other ARF GEPs that maypreferentially accelerate guanine nucleotide binding to ARF6.Amino acid sequence of the Sec7 domain of ARF-GEP100 is 27%and 41% identical to those of EFA6 and ARNO, respectively.The recombinant ARF proteins used in our experiments werenonmyristoylated, which appears not to affect the specificity ofinteraction, although it does affect rates of guanine nucleotideexchange. Some ARF GEPs are inhibited by the fungal fatty acidmetabolite BFA, which causes reversible inhibition of proteintransport and apparent disintegration of Golgi structure, in partby interfering with ARF activation. Asp965 and Met975 in yeastSec7 were shown to be responsible for its BFA sensitivity (28).These are equivalent to Asp543 and Met555 in ARF-GEP100,which suggested that it would be a BFA-sensitive GEP. However,the tyrosine-serine pair present in other BFA-sensitive ARFGEPs (29) is replaced in ARF-GEP100 by phenylalanine-alanine,which is also found in the BFA-insensitive cytohesin proteins.Replacement of Met699 in the BFA-sensitive Gea1 with Leurendered it BFA-resistant (29), consistent with the BFA-
insensitivity of ARF-GEP100, which contains leucine in thecorresponding position. The structural elements that singly ortogether influence BFA sensitivity appear to include residues inmotif 2, as well as additional sites adjacent to it (28, 29).
Near the N terminus of ARF-GEP100 is an IQ-like motif, asequence believed to be involved in calmodulin binding. Manyproteins contain IQ domains, but only a few of these have beenshown to bind calmodulin (39). We were unable to detect aneffect of calmodulin, with or without calcium on ARF-GEP100acceleration of GTPgS binding by ARF6, suggesting that ifcalmodulin does bind ARF-GEP100, the interaction does notmodify its GEP activity. This does not, of course, rule out thepossibility that an additional protein factor(s), or covalentmodification, may be required for interaction with calmodulin.It is also possible that calmodulin regulates functions of ARF-GEP100 other than its GEP activity.
Because the deduced amino acid sequence of ARF-GEP100appeared to contain a PH domain (amino acids 631–742), effects ofphospholipids on its GEP activity were investigated. No effects ofPS, PIP2, or PIP3 on ARF-GEP100-catalyzed GTPgS binding toARF6 were, however, found. The ARF-GEP100 PH domain se-quence differs greatly from those of the cytohesins and appears to
Fig. 7. Intracellular distribution of ARF-GEP100 and ARF6 in T98G cells. Cellswere reacted with rabbit anti-ARF-GEP100 (p100) antibody and mouse anti-ARF6, followed by Texas Red-labeled anti-rabbit IgG and FITC-labeled anti-mouse IgG. Lower right panel is a composite of the superimposed images.Lower left panel shows the corresponding Normarski image.
Fig. 8. Confocal images of T98G cells stained for ARF-GEP100 and organellemarker proteins. Cells were reacted with rabbit anti-ARF-GEP100 (p100) anti-body and mouse monoclonal antibody against b-catenin, LAMP-1, Golgi 58Kprotein, EEA-1, AP-1, or AP-2, followed by Texas Red-labeled anti-rabbit IgGand FITC-labeled anti-mouse IgG. In the first two columns are pairs of imagesof the same cells. Images in the third column are superimposed images of thepreceding panels. In the fourth column are Nomarski images.
Someya et al. PNAS u February 27, 2001 u vol. 98 u no. 5 u 2417
CELL
BIO
LOG
Y

lack residues critical for phosphoinositide binding in that family ofARF GEPs (22), consistent with the lack of effect of PIP2 and PIP3.Other regions of the cytohesin proteins involved in phospholipidbinding include the polybasic region that follows the PH domain(20) and is not found in ARF-GEP100. Despite their similarmolecular structures, PH domains differ widely in amino acidsequence and specific motifs predictive of binding partners orfunction have not been identified. The PH domain in ARF-GEP100may participate in specific protein interactions or serve to bindphospholipids other than PS or the phosphatidylinositol phosphatesthat were tested. Alternatively, as suggested by the inconsistentresults of repeated computer sequence analyses, the region initiallyidentified may not, in fact, represent a PH domain structure.
Our evidence is consistent with a physiological function forARF-GEP100 in the regulation of ARF6 activity and perhaps inother aspects of endocytosis. Immunoreactive endogenous ARF6and ARF-GEP100 were partially colocalized in vesicular structuresthat remain to be identified, and ARF-GEP100 immunofluores-cence also coincided with that of EEA-1, a marker for earlyendosomes. We were unable to demonstrate colocalization ofARF-GEP100 and ARF6 at the plasma membrane, which is not
inconsistent with a role for ARF-GEP100, in the regulation of ARF6activity at endosomal structures. It is possible that ARF-GEP100associates with other membranes in response to external or endog-enous signals. After cell fractionation, ARF-GEP100 was detectedonly in the cytosol, suggesting that it may be rather easily dissociatedfrom endosomal or other membranes. In the same cell fractions,only very little ARF6 was cytosolic. The presence of MgCl2 in thehomogenization buffer, which with Chinese hamster ovary cellsdramatically increased cytosolic ARF6 (43), had little effect on thedistribution of either protein in T98G cells. Considerable differ-ences were found among several cell lines in the fraction of ARF6that was cytosolic in homogenates containing MgCl2. It was sug-gested that ARF6 release from membranes results from Mg21
enhancement of GTPase activity (43). The colocalization of ARF-GEP100 with EEA-1 (44) suggests that it might function in ARFactivation at specific sites involved in vesicle formation, fusion, ortethering in an endocytic pathway.
We thank Dr. Julie G. Donaldson for providing helpful advice and rabbitanti-ARF6 antibody, Dr. T. Nagase for providing cDNA encodingKIAA0763, and Carol Kosh for expert secretarial assistance.
1. Robinson, M. S. (1997) Trends Cell Biol. 7, 99–102.2. Moss, J. & Vaughan, M. (1995) J. Biol. Chem. 270, 12327–12330.3. Rothman, J. E. & Wieland, F. T. (1996) Science 272, 227–234.4. Springer, S., Spang, A. & Schekman, R. (1999) Cell 97, 145–148.5. Brown, H. A., Gutowski, S., Moomaw, C. R., Slaughter, C. & Sternweis, P. C.
(1993) Cell 75, 1137–1144.6. Cockcroft, S., Thomas, G. M. H., Fensome, A., Geny, B., Cunningham, E.,
Gout, I., Hiles, I., Totty, N. F., Troung, O. & Hsuan, J. J. (1994) Science 263,523–526.
7. Massenburg, D., Han, J.-S., Liyanage, M., Patton, W. A., Rhee, S. G., Moss, J.& Vaughan, M. (1994) Proc. Natl. Acad. Sci. USA 91, 11718–11722.
8. Roth, M. G. & Sternweis, P. C. (1997) Curr. Opin. Cell. Biol. 9, 519–526.9. Honda, A., Nogami, M., Yokozeki, T., Yamazaki, M., Nakamura, H., Wa-
tanabe, H., Kawamoto, K., Nakayama, K., Morris, A. J., Frohman, M. A. &Kanaho, Y. (1999) Cell 99, 521–532.
10. Moss, J. & Vaughan, M. (1998) J. Biol. Chem. 273, 21431–21434.11. Meacci, E., Tsai, S.-C., Adamik, R., Moss, J. & Vaughan, M. (1997) Proc. Natl.
Acad. Sci. USA 94, 1745–1748.12. Chardin, P., Paris, S., Antonny, B., Robineau, S., Beraud-Dufour, S., Jackson,
C. L. & Chabre, M. (1996) Nature (London) 384, 481–484.13. Klarlund, J. K., Rameh, L. E., Cantley, L. C., Buxton, J. M., Holik, J. J., Sakelis,
C., Patki, V., Corvera, S. & Czech, M. P. (1998) J. Biol. Chem. 273, 1859–1862.14. Ogasawara, M., Kim, S.-C., Adamik, R., Togawa, A., Ferrans, V. J., Takeda, K.,
Kirby, H., Moss, J. & Vaughan, M. (2000) J. Biol. Chem. 275, 3221–3230.15. Franco, M., Peters, P. J., Boretto, J., van Donselaar, E., Neri, A., D’Souza-
Schorey, C. & Chavrier, P. (1999) EMBO J. 18, 1480–1491.16. Morinaga, N., Moss, J. & Vaughan, M. (1997) Proc. Natl. Acad. Sci. USA 94,
12926–12931.17. Togawa, A., Morinaga, N., Ogasawara, M., Moss, J. & Vaughan, M. (1999)
J. Biol. Chem. 274, 12308–12315.18. Peyroche, A., Paris, S. & Jackson, C. L. (1996) Nature (London) 384, 479–481.19. Achstetter, T., Franzusoff, A., Field, C. & Schekman, R. (1988) J. Biol. Chem.
263, 11711–11717.20. Macia, E., Paris, S. & Chabre, M. (2000) Biochemistry 39, 5893–5901.21. Pacheco-Rodriguez, G., Meacci, E., Vitale, N., Moss, J. & Vaughan, M. (1998)
J. Biol. Chem. 273, 26543–26548.22. Jackson, T. R., Kearns B. G. & Theibert, A. B. (2000) Trends Biochem. Sci. 25,
489–495.23. Franzusoff, A. & Schekman, R. (1989) EMBO J. 8, 2695–2702.
24. Mossessova, E., Gulbis, J. M. & Goldberg, J. (1998) Cell 92, 415–423.25. Betz, S. F., Schnuchel, A., Wang, H., Olejniczak, E. T., Meadows, R. P., Lipsky,
B. P., Harris, E. A. S., Staunton, D. E. & Fesik, S. W. (1998) Proc. Natl. Acad.Sci. USA 95, 7909–7914.
26. Goldberg, J. (1998) Cell 95, 237–248.27. Beraud-Dufour, S., Robineau, S., Chardin, P., Paris, S., Chabre, M., Cherfils,
J. & Antonny, B. (1998) EMBO J. 17, 3651–3659.28. Sata, M., Moss, J. & Vaughan, M. (1999) Proc. Natl. Acad. Sci. USA 96,
2752–2757.29. Peyroche, A., Antonny, B., Robineau, S., Acker, J., Cherfils, J. & Jackson, C. L.
(1999) Mol. Cell. 3, 275–285.30. Rebecchi, M. J. & Scarlata, S. (1998) Annu. Rev. Biophys. Biomol. Struct. 27,
503–528.31. Ferguson, K. M., Kavran, J. M., Sankaran, V. G., Fournier, E., Isakoff, S. J.,
Skolnik, E. Y. & Lemmon, M. A. (2000) Mol. Cell. 6, 373–384.32. Tsuchiya, M., Price, S. R., Tsai, S.-C., Moss, J. & Vaughan, M. (1991) J. Biol.
Chem. 266, 2772–2777.33. Claude, A., Zhao, B.-P., Kuziemsky, C. E., Dahan, S., Berger, S. J., Yan, J.-P.,
Arnold, A. D., Sullivan, E. M. & Melancon, P. (1999) J. Cell Biol. 146, 71–84.34. Frank, S., Upender, S., Hansen, S. H. & Casanova, J. E. (1998) J. Biol. Chem.
273, 23–27.35. Hong, J.-X., Haun, R. S., Tsai, S.-C., Moss, J. & Vaughan, M. (1994) J. Biol.
Chem. 269, 9743–9745.36. Haun, R. S., Tsai, S.-C., Adamik, R., Moss, J. & Vaughan M. (1993) J. Biol.
Chem. 269, 7064–7068.37. Tsai, S.-C., Adamik, R., Moss, J. & Vaughan, M. (1996) Proc. Natl. Acad. Sci.
USA 93, 305–309.38. Houdusse, A. & Cohen, C. (1995) Proc. Natl. Acad. Sci. USA 92, 10644–10647.39. Rhoads, A. & Friedberg, F. (1997) FASEB J. 11, 331–340.40. Cherfils, J., Menetrey, J., Mathieu, M., Le Bras, G., Robineau, S., Beraud-
Dufour, S., Antonny, B. & Chardin, P. (1998) Nature (London) 392, 101–105.41. Menetrey, J., Macia, E., Pasqualato, S., Franco, M. & Cherfils, J. (2000) Nat.
Struct. Biol. 7, 466–469.42. Vitale, N., Pacheco-Rodriguez G., Ferrans, V. J., Riemenschneider, W., Moss,
J. & Vaughan, M. (2000) J. Biol. Chem. 275, 21331–21339.43. Gaschet, J. & Hsu, V. W. (1999) J. Biol. Chem. 274, 20040–20045.44. Rubino, M., Miaczynska, M., Lippe, R. & Zerial, M. (2000) J. Biol. Chem. 275,
3745–3748.
2418 u www.pnas.orgycgiydoiy10.1073ypnas.051634798 Someya et al.















![Solution Structure of the Na+ form of the Dimeric Guanine Quadruplex [d(G3T4G3)]2](https://static.fdokumen.com/doc/165x107/6318f44265e4a6af370f95cf/solution-structure-of-the-na-form-of-the-dimeric-guanine-quadruplex-dg3t4g32.jpg)




