
Phosphatidylinositol 3Kinase Is a Negative Regulator of Cellular Differentiation

International Journal for Parasitology 39 (2009) 645–653
Contents lists available at ScienceDirect
International Journal for Parasitology
journal homepage: www.elsevier .com/locate / i jpara
A unique phosphatidylinositol 4-phosphate 5-kinase is activatedby ADP-ribosylation factor in Plasmodium falciparum
Werner Leber a,*,1, Alison Skippen b, Quinton L. Fivelman a, Paul W. Bowyer c, Shamshad Cockcroft b,David A. Baker a
a Department of Infectious and Tropical Diseases, London School of Hygiene & Tropical Medicine, Keppel Street, London WC1E 7HT, United Kingdomb Department of Cell and Developmental Biology, University College London, London WC1E 6JJ, United Kingdomc Division of Cell and Molecular Biology, Centre for Molecular Microbiology and Infection, Imperial College London, London SW7 2AZ, United Kingdom
a r t i c l e i n f o a b s t r a c t
Article history:Received 8 September 2008Received in revised form 9 November 2008Accepted 13 November 2008
Keywords:PlasmodiumPhosphatidylinositol 4-phosphate 5-kinaseNeuronal calcium sensor proteinADP-ribosylation factorProtein traffickingCytoskeleton
0020-7519/$36.00 � 2009 Australian Society for Paradoi:10.1016/j.ijpara.2008.11.015
* Corresponding author. Tel.: +44 (0) 20 7882 2509E-mail address: [email protected] (W. L
1 Present address: Centre for Health Sciences, BartMedicine and Dentistry, Queen Mary, University of Lon2AT, UK.
In eukaryotes, calcium signalling has been linked to hydrolysis of the phosphatidylinositol 4,5-bisphos-phate (PtdIns(4,5)P2). The final enzyme in the synthesis of this phosphoinositide, a Type I phosphatidyl-inositol 4-phosphate 5-kinase (PIP5K), is activated by the small G protein ADP-ribosylation factor 1(ARF1). In mammals, the ARF-PIP5K pathway is a key regulator of cell motility, secretion and cell signal-ling. We report the characterisation of a unique, putative bifunctional PIP5K in the human malaria par-asite Plasmodium falciparum. The protein comprises a C-terminal, functional PIP5K domain withcatalytic specificity for phosphatidylinositol 4-phosphate. The recombinant enzyme is activated byARF1 but not phosphatidic acid. The protein also incorporates an unusual N-terminal domain with poten-tial helix-loop-helix EF-hand-like motifs that is a member of the neuronal calcium sensor family (NCS).Intriguingly, NCS-1 has been shown to stimulate phosphatidylinositol 4-phosphate synthesis by activat-ing mammalian and yeast phosphatidylinositol 4-kinase b in vitro in a calcium-dependent manner. Theunexpected physical attachment of an NCS-like domain to the plasmodial PIP5K might reflect a uniquefunctional link between the calcium and PtdIns(4,5)P2 pathways allowing modulation of PtdIns(4,5)P2
production in response to changes in intracellular calcium concentrations within the parasite.� 2009 Australian Society for Parasitology Inc. Published by Elsevier Ltd. All rights reserved.
1. Introduction
Malaria is a systemic infectious disease caused by protozoa ofthe genus Plasmodium. By far the most harmful species is Plasmo-dium falciparum and it is estimated that at least one million peopledie from malaria each year; many of these deaths occur amongstyoung children in Africa. Development of drug resistance in P. fal-ciparum has been noted with great concern, and measures to erad-icate malaria in endemic areas have failed to date (White et al.,1999). New drugs and effective vaccines are urgently needed.The life cycle of P. falciparum is complex and comprises both asex-ual and sexual stages. Asexual blood stages cause pathologywhereas sexual stages mediate transmission of the parasite be-tween human hosts via the Anopheles mosquito vector. Sexual par-asites (gametocytes) form within the human blood and are takenup by a mosquito together with a blood meal. On entering the mid-gut, gametocytes emerge from their host erythrocytes as mature
sitology Inc. Published by Elsevier
; fax: +44 (0) 20 7882 2552.eber).s and The London School ofdon, 2 Newark St., London E1
gametes. Male gametogenesis involves three endomitotic divi-sions, giving rise to eight highly motile, flagellated gametes (a pro-cess known as exflagellation).
In vitro studies of exflagellation have shown that a drop in tem-perature coupled with either an increase in pH or a mosquito-de-rived factor, xanthurenic acid, can trigger this process (Sindenet al., 1976; Nijhout and Carter, 1978; Billker et al., 1998). Previouswork has also implicated calcium, the cyclic nucleotide cGMP,diacylglycerol and inositol 1,4,5-trisphosphate in exflagellation(Kawamoto et al., 1990; Ogwan’g et al., 1993; Martin et al.,1994). Hydrolysis of phosphatidylinositol 4,5-bisphosphate(PtdIns(4,5)P2) by phospholipase C produces the second messen-gers diacylglycerol and inositol 1,4,5-trisphosphate that are associ-ated with calcium signalling. In mammalian cells, PtdIns(4,5)P2 ismainly generated by sequential phosphorylation of phosphatidyl-inositol via phosphatidylinositol 4-phosphate (PtdIns4P) throughthe activities of phosphatidylinositol 4-kinases (PI4Ks) and Type Iphosphatidylinositol 4-phosphate 5-kinases (PIP5Ks). By regulat-ing the levels of PtdIns(4,5)P2, PIP5Ks play a key role in membranetrafficking, cytoskeletal reorganisation and cell signalling in a widerange of eukaryotic cells (Cockcroft, 2000; Martin, 2001).
As part of a study to investigate the potential role of cGMP inexflagellation, we identified a putative guanylyl cyclase-activating
Ltd. All rights reserved.

646 W. Leber et al. / International Journal for Parasitology 39 (2009) 645–653
protein (GCAP) homologue, a member of neuronal calcium sensorproteins (NCS), in the P. falciparum genome database. NCS are N-terminally acylated, EF-hand calcium-trigger proteins that undergoconformational changes in response to calcium (Burgoyne andWeiss, 2001). Remarkably, the predicted NCS-like protein sequenceis potentially a bifunctional protein that incorporates a C-terminalPIP5K domain. We show that the two domains are potentially tran-scribed and translated as a single unit, called PfPIP5K/NCS, and thatthe phosphoinositide kinase domain is a functional Type I PIP5Kthat is activated by ADP-ribosylation factor 1 (ARF1).
2. Materials and methods
2.1. Biochemical reagents
All reagents were purchased from Sigma–Aldrich (UK) unlessotherwise stated. [c-32P] ATP was obtained from Amersham Biosci-ences (UK). Bovine PtdIns4P was obtained from both Sigma–Aldrich (UK) and Avanti Phospholipids (USA). The syntheticD-myo-phospholipids PtdIns3P, PtdIns4P and PtdIns5P wereobtained from Echelon Biosciences Incorporated (USA), and wereof >99% isomeric purity according to the manufacturer’s analysis.Oligonucleotides were custom-synthesised and purchased fromSigma-Genosys (UK).
2.2. Cloning and expression of the recombinant P. falciparum PIP5Kdomain
For the generation of the P. falciparum PIP5K domain, a 1380-bpcDNA sequence was amplified from mixed-parasite stage 3D7mRNA by random priming using the ProSTARTM Ultra HF RT-PCRSystem according to the manufacturer’s instructions (Stratagene,UK). The sequence encoded the C-terminal open reading frame(ORF) and comprised the PIP5K dimerisation region, the kinase cat-alytic core, the activation loop including the natural stop codon ofthe PfPIP5K/NCS gene. For the amplification reaction, 40 ll of RNasefree water, 5 ll of Ultra HF PCR buffer, 1 ll of 40 mM dNTP mix(40 mM), 1 ll of each primer (25 lM), 1 ll of first-strand cDNAand 1 ll of Pfu Turbo DNA polymerase were mixed. The primersused were GCAP2EF (50-CGCGGATCCTCATATGATTCATTAGATGATAGTAGTAATATG, sense primer containing a BamHI site) andGCAP2HR (50-CGCAAGCTTTCATTTCATATGATTTCTATAAATGTAAC,antisense primer containing the natural stop codon and a HindIIIsite). The cycling reactions were as follows: 1 cycle of 95 �C for1 min, 60 �C for 30 s and 65 �C for 5 min, 33 cycles of 95 �C for30 s, 60 �C for 30 s, and 65 �C for 5 min, and finally 1 cycle of95 �C for 30 s, 60 �C for 30 s and 65 �C for 10 min. To confirm thatthe amplified cDNA encoded the recombinant P. falciparum PIP5Kdomain, cDNA cloned into pTrcHis A (Invitrogen, UK) was ex-pressed in Escherichia coli JM107, and detected from bacterial ly-sate expressing the PIP5K-cDNA but not vector alone using SDS–PAGE and Western blotting. An overnight subculture from a single,freshly transformed bacterial colony was diluted 1:10 in Luria–Ber-tani medium (LB)/ampicillin (100 lg ml�1) and grown at 37 �Cwith shaking at 220 rpm for 1.5 h before induction with 0.1 lMisopropyl b-D-thiogalactosidase (IPTG) at 26 �C for 3 h. Cells werepelleted and re-suspended in lysis buffer (50 mM NaH2PO4,300 mM NaCl, pH 8.0). The lysate was supplemented with DNAse(10 U/ll final concentration), lysozyme (1 mg/ml final concentra-tion) and protease inhibitors (Complete Mini EDTA-free, Roche),sonicated on ice and finally centrifuged at 15,000g for 30 min at4 �C. Using elution buffer (50 mM NaH2PO4, 300 mM NaCl,20 mM imidazole, pH 8.0) and Econopac 10DG ChromatographyColumns (Bio-Rad, UK), the recombinant N-terminally His6-taggedPIP5K domain was batch affinity-purified on Ni2+-nitrilotriacetic
acid agarose (Qiagen) and buffer exchanged using 1,4-piperazine-diethanesulfonic acid (PIPES)/salts buffer (20 mM PIPES, 137 mMNaCl, 2.7 mM KCl, pH 6.8).
2.3. Generation of recombinant myristoylated PfARF1
Myristoylated P. falciparum ARF1 (PfARF1) was generated simi-larly to previously published protocols for mammalian ARF (Duro-nio et al., 1990; Jones et al., 1999). The PfARF1 gene was modifiedfrom Stafford et al. (1996) to contain the wild-type N-terminusand encode a His6-tag at the C-terminus in pET-28a (Bowyer,P.W., 2007. Studies on the N-myristoyl transferase of P. falciparum.PhD Thesis, University of London, UK; Stafford et al., 1996). Myri-stoylated PfARF1 was obtained by co-expression of the modifiedPfARF1 and P. falciparum N-myristoyl transferase in E. coliBL21(DE3)pLysS in the presence of myristic acid (Duronio et al.,1990; Jones et al., 1999). Expression of protein was induced with1 mM IPTG at OD600 0.5 for 4 h. His6-tagged proteins were purifiedin a single step from bacterial lysate on Talon (TM) metal affinityresin (Clontech, USA) according to the manufacturer’s protocol.
2.4. In vitro assay for measuring phosphatidylinositol 4-phosphate 5-kinase activity
An adapted phospholipid kinase assay was used to determinethe incorporation of [c-32P] ATP into PtdIns(4,5)P2 in the presenceof the recombinant P. falciparum PIP5K domain (Jones et al., 2000).Phospholipids were separated by thin layer chromatography. Thereaction product of the recombinant P. falciparum PIP5K domainwas identified by co-migration of a radio-labelled PtdIns(4,5)P2
generated by recombinant mammalian Type I PIP5Ka. Thin layerchromatography plates were exposed to a phosphor screen (GEHealthcare/Amersham Biosciences, UK), radioactive signals werevisualised by a Storm 840 phosphor imager and quantified usingImageQuant software (both Molecular Dynamics, USA). MouseType I PIP5Ka, rat ARF nucleotide-binding site opener (ARNO),and myristoylated rat ARF1 were generated as described previ-ously (Cockcroft et al., 1994; Ishihara et al., 1996; Jones et al.,2000).
2.5. Cultivation and purification of P. falciparum blood stage parasites
Synchronous asexual malaria parasites were continuously cul-tured according to the method of Trager and Jensen (1976), withslight modifications (Fairlamb et al., 1985). Gametocytogenesiswas induced from 3D7A asexual parasites and gametocytes werecultured in the presence of 50 mM N-acetylglucosamine (Sigma–Aldrich) as previously described (Fivelman et al., 2007). Trophozo-ite stage parasites were enriched using MACS CS separation col-umns (Miltenyi Biotech, Germany) and stage V parasites wereenriched using Nycodenz (Nycoprep 1.077TM, Axis-Shield) as de-scribed (Fivelman et al., 2007). The number of gametocytes wascounted on a haemocytometer and split into aliquots of 5 � 106
cells. Trophozoites were split into aliquots of 1 � 109 cells. All par-asite aliquots were snap-frozen in liquid nitrogen and stored at�80 �C until required.
2.6. Northern blotting
Parasite RNA was obtained from parasitised red blood cells atthe required lifecycle stage using TriZol (Invitrogen, UK) accordingto manufacturer’s instructions. Northern blots were prepared asdescribed previously (Kyes et al., 2000). [32P] dCTP-labelled P. falci-parum PIP5K domain probes were produced by the random primermethod using Rediprime II kit (Amersham Biosciences, UK).

W. Leber et al. / International Journal for Parasitology 39 (2009) 645–653 647
2.7. Western blotting
Purified parasites and red blood cells were lysed in ice-coldradioimmunoprecipitation assay (RIPA) buffer (10 mM Tris, pH7.2, 150 mM NaCl, SDS 0.1%, Triton X-100 1%, sodium deoxycholate1%, EDTA 5 mM) followed by six cycles of freeze thawing and pass-ing through a 21 gauge needle. Lysates were spun down at 15,000gfor 5 min. An equal volume of purified recombinant protein or par-asite supernatant was mixed with 2� loading buffer and subjectedto SDS–PAGE followed by blotting onto nitrocellulose membrane.In order to visualise recombinant protein, blots were probed withanti-Xpress antibody (Invitrogen, UK) and goat anti-mouse second-ary antibody (Bio-Rad, UK; both diluted 1:37,500). Purified poly-clonal antibodies against the Keyhole Limpet Hemocyanin (KLH)-conjugated peptide ‘FYGIKSKEYLTSVGP’ from the P. falciparumPIP5K domain, raised in rabbits, were commercially obtained fromEurogentech, Belgium. Blots were probed with these polyclonalantibodies (1:400) and horseradish peroxidase (HRP)-conjugatedgoat anti-rabbit secondary antibody (Bio-Rad, UK; 1:25,000). Sig-nals were detected using the ECL Advance chemiluminescence kitand ECL Hyperfilm (both Amersham Biosciences, UK).
3. Results
3.1. Predicted domain structure of a putative bifunctional PIP5K/NCSin P. falciparum
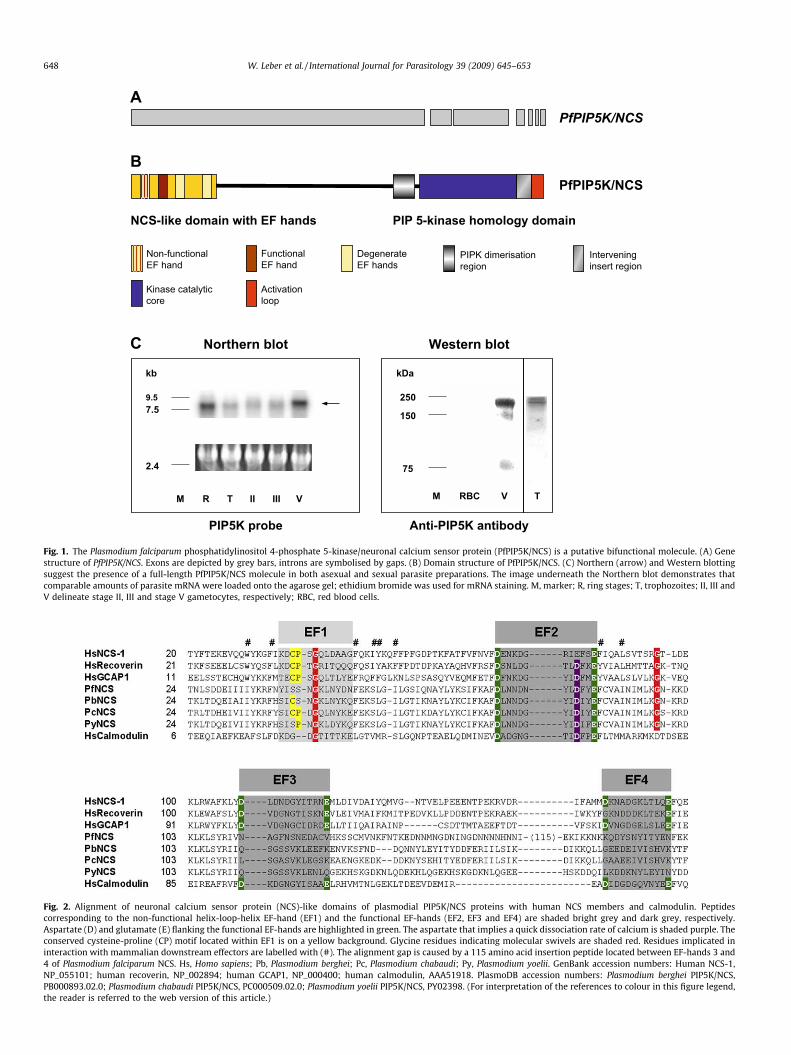
Searches of the P. falciparum genome project database withmammalian GCAP sequence revealed a single high probabilitymatch to chromosome 1 in the Sanger Centre database – here des-ignated PfPIP5K/NCS. The annotated genome data (PFA0515w;http://www.plasmodb.org) suggested that the full-length PfPIP5K/NCS gene forms an ORF, which is interrupted by six introns atthe 30 end (Fig. 1A). The gene identified is predicted to encode a1710 amino acid, putatively bifunctional, protein comprising anN-terminal NCS-like domain and also a PIP5K domain at the C-ter-minus (Fig. 1B).
3.2. PfPIP5K/NCS is expressed as a potentially bifunctional molecule
To investigate whether the PfPIP5K/NCS gene is transcribed as asingle mRNA molecule, total RNA isolated from both asexual andsexual blood stage parasites was separated and analysed usingNorthern blotting with a radio-labelled P. falciparum PIP5K domainprobe. This revealed large transcripts (�8000 nucleotides) in allblood stage parasites, with strongest signals being present in ringstages and stage V gametocytes (Fig. 1C). The transcript size is con-sistent with both functional domains being encoded by a singletranscript. To investigate whether the PfPIP5K/NCS protein is ex-pressed as a contiguous polypeptide in stage V gametocytes, para-site proteins were separated by SDS–PAGE and probed withpurified anti-P. falciparum PIP5K domain peptide antibody. This re-sulted in a major band of approximately 200 kDa, which again cor-responds to the predicted combined size of PfPIP5K/NCS suggestingthat it may also be translated as a single bifunctional protein. Aband of equal size was detected in asexual trophozoite stage prep-arations on a Western blot that was performed under similar con-ditions (Fig. 1C).
3.3. The PfPIP5K/NCS has sequence domains corresponding to bothmammalian PIP5K and NCS
Members of the NCS family of proteins typically comprise onenon-functional EF-hand and either two (e.g. recoverin) or three(e.g. mammalian GCAP) additional functional EF-hands for cal-
cium-binding (Burgoyne et al., 2004). The plasmodial NCS-like do-main is unusual and is predicted to have one non-functional (EF-hand 1) and a single functional EF-hand (EF-hand 2). The two C-terminal EF-hands (EF-hands 3 and 4) are partially conserved(see Fig. 2). EF-hand 2 contains an aspartate at position nine sug-gesting that it is potentially a calcium-trigger rather than a cal-cium-buffer (Renner et al., 1993; Ames et al., 1997). The two N-terminal glycines highlighted in Fig. 2 (Gly-45 and Gly-98) mayfacilitate a calcium-mediated conformational switch, and the cys-teine at position three (1-Met-Lys-Cys-3) is a potential target forposttranslational palmitoylation (Takimoto et al., 2002). Both theN-terminal patch of hydrophobic residues and a cysteine-proline(CP) motif within EF-hand 1 may mediate interaction with down-stream effectors such as GCs (Ermilov et al., 2001; Hwang et al.,2004; Ames et al., 2006; Strahl et al., 2007). The CP has been re-placed by two serine residues in the P. falciparum NCS-like domain.In the mouse orthologues Plasmodium berghei PIP5K/NCS (Ser-43-Pro) and Plasmodium yoelii PIP5K/NCS (Ser-42-Cys), only single res-idue replacements have occurred. Intriguingly, this CP motif is en-tirely conserved in the NCS-like domain of the mouse parasitePlasmodium chabaudi.
The P. falciparum PIP5K domain is 31% identical and 48% similarto human Type I PIP5Ka over the C-terminal 269 amino acids. Thedomain structure of the P. falciparum PIP5K is conserved and com-prises the following putative functional units (see Fig. 3A–C): (i) aPIP5K dimerisation region that is rich in polar, positively chargedamino acids possibly used for homodimerisation and attachmentto cellular membranes (Rao et al., 1998); (ii) a kinase catalytic corecontaining two conserved amino acids for ATP-binding (Ishiharaet al., 1998; Powner et al., 2005); (iii) an intervening insert region(Rao et al., 1998); and finally (iv) an activation loop that includesresidues for substrate binding and a dilysine motif for membraneattachment and actin remodelling (Kunz et al., 2000, 2002).
In mammalian cells, PtdIns(4,5)P2 is synthesised from phospha-tidylinositol via two different routes: either by phosphorylation ofPtdIns4P on the D-5 position by a Type I PIP5K (major route), or byphosphorylation of PtdIns5P on the D-4 position by a Type II phos-phatidylinositol 5-phosphate 4-kinase (PIP4K) (minor route)(Cockcroft, 2000). Both enzymes are structurally very similar butdiffer in their substrate preference (Fig. 3A and C), which is deter-mined by a single diagnostic residue located at position 16 withinthe activation loop (Rao et al., 1998; Kunz et al., 2002). The pres-ence of a conserved alanine determines substrate specificity ofType II PIP4Ks for PtdIns5P. The presence of a glutamate residueat the corresponding position of Type I PIP5Ks, however, mediatesbinding of PtdIns4P (Kunz et al., 2002). This specific glutamate res-idue is present in the P. falciparum PIP5K domain suggesting that itis a Type I PIP5K but not a PIP4K (Fig. 3C).
3.4. The plasmodial recombinant PIP5K domain has PIP5K enzymeactivity that is insensitive to phosphatidic acid
The P. falciparum PIP5K domain was amplified from mixed tro-phozoite and gametocyte cDNA and expressed as a His6-tag fusionprotein in E. coli. Production of the recombinant protein was con-firmed by Western blotting (Fig. 4B). In a typical reaction, the re-combinant P. falciparum PIP5K domain was assayed for its abilityto phosphorylate bovine PtdIns4P to generate PtdIns(4,5)P2. Re-combinant mammalian full-length Type I PIP5Ka is a well charac-terised enzyme and was assayed in parallel to confirm the identityof the PtdIns(4,5)P2 reaction products generated by the recombi-nant P. falciparum PIP5K domain (data not shown). Commercial bo-vine PtdIns4P preparations are known to be contaminated withtraces of other phospholipids. PtdIns(4,5)P2 derived from the affin-ity-purified recombinant P. falciparum PIP5K domain could have,therefore, resulted from the phosphorylation of a monophosphate
Kinase catalytic core
Activation loop
PIPK dimerisation region
Intervening insert region
Functional EF hand
Non-functional EF hand
Degenerate EF hands
C
kb
9.5 7.5
2.4
M R T II III V M RBC V
kDa
250
150
75
T
Northern blot Western blot
PIP5K probe Anti-PIP5K antibody
PIP 5-kinase homology domain NCS-like domain with EF hands
PfPIP5K/NCS
PfPIP5K/NCS B
A
Fig. 1. The Plasmodium falciparum phosphatidylinositol 4-phosphate 5-kinase/neuronal calcium sensor protein (PfPIP5K/NCS) is a putative bifunctional molecule. (A) Genestructure of PfPIP5K/NCS. Exons are depicted by grey bars, introns are symbolised by gaps. (B) Domain structure of PfPIP5K/NCS. (C) Northern (arrow) and Western blottingsuggest the presence of a full-length PfPIP5K/NCS molecule in both asexual and sexual parasite preparations. The image underneath the Northern blot demonstrates thatcomparable amounts of parasite mRNA were loaded onto the agarose gel; ethidium bromide was used for mRNA staining. M, marker; R, ring stages; T, trophozoites; II, III andV delineate stage II, III and stage V gametocytes, respectively; RBC, red blood cells.
Fig. 2. Alignment of neuronal calcium sensor protein (NCS)-like domains of plasmodial PIP5K/NCS proteins with human NCS members and calmodulin. Peptidescorresponding to the non-functional helix-loop-helix EF-hand (EF1) and the functional EF-hands (EF2, EF3 and EF4) are shaded bright grey and dark grey, respectively.Aspartate (D) and glutamate (E) flanking the functional EF-hands are highlighted in green. The aspartate that implies a quick dissociation rate of calcium is shaded purple. Theconserved cysteine-proline (CP) motif located within EF1 is on a yellow background. Glycine residues indicating molecular swivels are shaded red. Residues implicated ininteraction with mammalian downstream effectors are labelled with (#). The alignment gap is caused by a 115 amino acid insertion peptide located between EF-hands 3 and4 of Plasmodium falciparum NCS. Hs, Homo sapiens; Pb, Plasmodium berghei; Pc, Plasmodium chabaudi; Py, Plasmodium yoelii. GenBank accession numbers: Human NCS-1,NP_055101; human recoverin, NP_002894; human GCAP1, NP_000400; human calmodulin, AAA51918. PlasmoDB accession numbers: Plasmodium berghei PIP5K/NCS,PB000893.02.0; Plasmodium chabaudi PIP5K/NCS, PC000509.02.0; Plasmodium yoelii PIP5K/NCS, PY02398. (For interpretation of the references to colour in this figure legend,the reader is referred to the web version of this article.)
648 W. Leber et al. / International Journal for Parasitology 39 (2009) 645–653
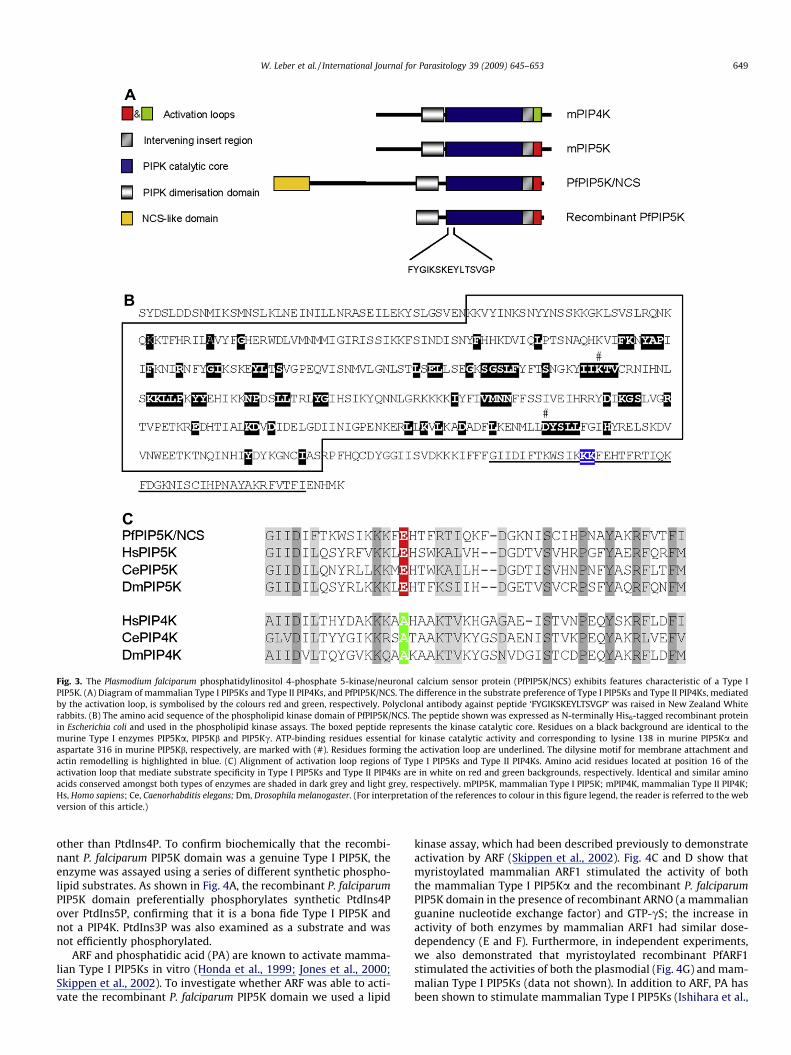
Fig. 3. The Plasmodium falciparum phosphatidylinositol 4-phosphate 5-kinase/neuronal calcium sensor protein (PfPIP5K/NCS) exhibits features characteristic of a Type IPIP5K. (A) Diagram of mammalian Type I PIP5Ks and Type II PIP4Ks, and PfPIP5K/NCS. The difference in the substrate preference of Type I PIP5Ks and Type II PIP4Ks, mediatedby the activation loop, is symbolised by the colours red and green, respectively. Polyclonal antibody against peptide ‘FYGIKSKEYLTSVGP’ was raised in New Zealand Whiterabbits. (B) The amino acid sequence of the phospholipid kinase domain of PfPIP5K/NCS. The peptide shown was expressed as N-terminally His6-tagged recombinant proteinin Escherichia coli and used in the phospholipid kinase assays. The boxed peptide represents the kinase catalytic core. Residues on a black background are identical to themurine Type I enzymes PIP5Ka, PIP5Kb and PIP5Kc. ATP-binding residues essential for kinase catalytic activity and corresponding to lysine 138 in murine PIP5Ka andaspartate 316 in murine PIP5Kb, respectively, are marked with (#). Residues forming the activation loop are underlined. The dilysine motif for membrane attachment andactin remodelling is highlighted in blue. (C) Alignment of activation loop regions of Type I PIP5Ks and Type II PIP4Ks. Amino acid residues located at position 16 of theactivation loop that mediate substrate specificity in Type I PIP5Ks and Type II PIP4Ks are in white on red and green backgrounds, respectively. Identical and similar aminoacids conserved amongst both types of enzymes are shaded in dark grey and light grey, respectively. mPIP5K, mammalian Type I PIP5K; mPIP4K, mammalian Type II PIP4K;Hs, Homo sapiens; Ce, Caenorhabditis elegans; Dm, Drosophila melanogaster. (For interpretation of the references to colour in this figure legend, the reader is referred to the webversion of this article.)
W. Leber et al. / International Journal for Parasitology 39 (2009) 645–653 649
other than PtdIns4P. To confirm biochemically that the recombi-nant P. falciparum PIP5K domain was a genuine Type I PIP5K, theenzyme was assayed using a series of different synthetic phospho-lipid substrates. As shown in Fig. 4A, the recombinant P. falciparumPIP5K domain preferentially phosphorylates synthetic PtdIns4Pover PtdIns5P, confirming that it is a bona fide Type I PIP5K andnot a PIP4K. PtdIns3P was also examined as a substrate and wasnot efficiently phosphorylated.
ARF and phosphatidic acid (PA) are known to activate mamma-lian Type I PIP5Ks in vitro (Honda et al., 1999; Jones et al., 2000;Skippen et al., 2002). To investigate whether ARF was able to acti-vate the recombinant P. falciparum PIP5K domain we used a lipid
kinase assay, which had been described previously to demonstrateactivation by ARF (Skippen et al., 2002). Fig. 4C and D show thatmyristoylated mammalian ARF1 stimulated the activity of boththe mammalian Type I PIP5Ka and the recombinant P. falciparumPIP5K domain in the presence of recombinant ARNO (a mammalianguanine nucleotide exchange factor) and GTP-cS; the increase inactivity of both enzymes by mammalian ARF1 had similar dose-dependency (E and F). Furthermore, in independent experiments,we also demonstrated that myristoylated recombinant PfARF1stimulated the activities of both the plasmodial (Fig. 4G) and mam-malian Type I PIP5Ks (data not shown). In addition to ARF, PA hasbeen shown to stimulate mammalian Type I PIP5Ks (Ishihara et al.,

650 W. Leber et al. / International Journal for Parasitology 39 (2009) 645–653
1998). To examine whether the same was true for the plasmodialPIP5K, the recombinant P. falciparum PIP5K domain was assayedin the absence or presence of PA. In contrast to recombinant mam-malian Type I PIP5Ka, which shows robust increase in lipid kinaseactivity, the recombinant P. falciparum PIP5K domain was not acti-vated by PA (Fig. 4H).
4. Discussion
Here we describe and biochemically characterise a novel Type IPIP5K protein, designated PfPIP5K/NCS, isolated from the humanmalaria parasite P. falciparum. We show that this molecule ispotentially a bifunctional protein that has an additional N-terminalEF-hand-containing domain that is related to the NCS protein sub-family of calcium-binding proteins with similarities to both GCAPand NCS-1. We provide evidence that both functional domainsare transcribed as a single mRNA and translated as a contiguouspolypeptide. A recombinant protein corresponding to the P. falcipa-rum PIP5K domain is biochemically functional and activated byARF1 but, unusually, not by PA.
In mammalian cells, PtdIns(4,5)P2 is mainly synthesised fromPtdIns by sequential phosphorylation by PI4Ks and Type I PIP5Ks.In the P. falciparum genome a single copy gene of each of these en-zymes is predicted to be present, and here we show that the PIP5Kdomain of PfPIP5K/NCS is a functional enzyme suggesting that themajor pathway for PtdIns(4,5)P2 synthesis from PtdIns is intact inthe malaria parasite. Mammalian Type I PIP5Ks are highly regu-lated and the small G protein ARF, for example, is a well knownactivator of these enzymes (Honda et al., 1999; Jones et al., 2000;Skippen et al., 2002). At least two different ARFs are predicted tobe encoded in the genome of P. falciparum but only one of these(designated PfARF1) has been partially characterised (Staffordet al., 1996; Lee et al., 1997). The PfARF1 gene was shown to betranscriptionally up-regulated in asexual parasites undergoing nu-clear fission during early schizogony (Stafford et al., 1996). In a sec-ond study, recombinant PfARF1 was demonstrated to stimulateboth mammalian phospholipase D (PLD) and cholera toxin ADP-ribosyltransferase activity in vitro (Lee et al., 1997). Here we showthat plasmodial and mammalian ARF1 activate both the plasmodialand the mammalian Type I PIP5Ka. ARFs are highly conserved GTP-ases. PfARF1 for instance is 75% identical and 95% similar to mam-malian ARF1. This probably explains the observed cross-speciesactivation of Type I PIP5Ks by both the mammalian and plasmodialARF1 reported here as well as the plasmodial ARF1-mediated acti-vation of mammalian PLD and cholera toxin (Lee et al., 1997).
PA, a product of PLD-mediated hydrolysis of phosphatidylcho-line, is believed to specifically activate mammalian Type I PIP5K(Ishihara et al., 1998). PLD is a ubiquitous enzyme found in bacte-ria, fungi, plants and mammals and comprises two copies of an
Fig. 4. The recombinant Plasmodium falciparum phosphatidylinositol 4-phosphate 5-kinPtdIns(4,5)P2 and is sensitive to ADP-ribosylation factor (ARF) but not phosphatidic acid (assayed for PtdIns(4,5)P2 synthesis in the presence of bovine PtdIns4P or a synthetic phoshown above. The reaction products originating from the synthetic D-myo-phospholipduplicate of a single representative experiment, which was reproduced on three differentthe affinity-purified recombinant P. falciparum PIP5K domain in Escherichia coli eluate.mammalian Type I PIP5Ka using liposomes consisting of PtdIns4P and phosphatidylchoduplicates of a single representative experiment, which was reproduced on three differemammalian PIP5K were assayed with 250 lM mammalian ARF1 in the absence or presenwere also assayed in the presence of GTP-cS with increasing concentrations of recombinin the absence or presence of recombinant P. falciparum ARF1 in vitro (250 lM final condomain was assayed in the absence (solid bars) or presence (grey bars) of 40 lM PA. Thesingle representative experiment, which was reproduced on four different occasions. ErroPIP5K; mARF1, mammalian ARF1; pARF1, P. falciparum ARF1; sPI3P, synthetic phosphosphatidylinositol 4-phosphate; GN, guanine nucleotide.
amino acid signature motif (HXK(X)4D), essential for its catalyticactivity (Exton, 1998; Cockcroft, 2001). Here we show that the re-combinant P. falciparum PIP5K domain is not activated by PA. Thisobservation is intriguing, as searches of the P. falciparum databaseusing the BLAST algorithm and using peptides containing theHXK(X)4D motif were negative, suggesting that a PLD orthologuemay be absent from the genome of the parasite. Evidence to sup-port plasmodial PLD activity is lacking in the literature. In mam-mals, ARF is also a direct activator of PLD, and both ARF and PLDhave been shown to co-localise to the same subcellular compart-ments as Type I PIP5K to participate in PtdIns(4,5)P2-mediatedevents (Honda et al., 1999; Divecha et al., 2000). It has been arguedthat PLD-derived PA is required for ARF-mediated activation ofType I PIP5K (Arneson et al., 1999; Siddhanta et al., 2000). How-ever, a recent study suggested that ARF is able to stimulate TypeI PIP5K directly in the absence of PLD-derived PA (Skippen et al.,2002). Our own experiments demonstrate that plasmodial PIP5Kis ARF-sensitive but non-responsive to PA, providing further evi-dence that PA and ARF regulate Type I PIP5K activityindependently.
A growing list of bifunctional proteins in which the componentsfeed into the same metabolic or signalling pathways have been re-ported in the malaria parasite (Bzik et al., 1987; Carucci et al.,2000; Clarke et al., 2001; Salcedo et al., 2001; Wrenger et al.,2001; Weber et al., 2004; Promeneur et al., 2007). In mammaliancells, a functional cross-talk that links the calcium-dependentNCS-1 pathway and the ARF-dependent pathway has been pro-posed recently (Haynes et al., 2005). Mammalian ARF co-stimu-lates phosphatidylinositol 4-kinase b (PI4Kb) to provide sufficientPtdIns4P substrate for ARF-mediated PtdIns(4,5)P2 synthesis (Godiet al., 1999), and NCS-1 has been shown to stimulate mammalianand yeast PI4Kb in vitro in a calcium-dependent manner (Hen-dricks et al., 1999; Zhao et al., 2001; Rajebhosale et al., 2003). Inthe human genome multiple PI4K genes are found, but only two se-quences in the plasmodial genome have features characteristic oftypical PI4Ks. Here, sequence analysis indicates that one of themis a PI4Kb orthologue, which might serve as a target enzyme forboth ARF and an NCS-like upstream activator. More recently,NCS-1 has also been shown to interact directly with both ARFand (hippocampal) membrane GC indicating conservation of theNCS-1-mediated cross-talk between the calcium, cGMP, andPtdIns(4,5)P2 signalling pathways (Fik-Rymarkiewicz et al., 2006;Haynes et al., 2007). The physical attachment of the NCS-like do-main to the plasmodial PIP5K might therefore reflect the functionallink between these pathways within the parasite.
Mammalian ARF1 regulates protein transport from the endo-plasmic reticulum (ER) to Golgi where it is activated by a brefeldinA (BFA)-sensitive guanine nucleotide exchange factor (ARF-GEF). InP. falciparum asexual blood stage parasites, a classical BFA-sensi-tive protein secretion pathway has been described (Crary and Hal-
ase (PIP5K) domain preferably converts the synthetic phospholipid PtdIns4P intoPA). (A) Nickel column affinity-purified recombinant P. falciparum PIP5K domain wasspholipid substrate (all 40 lM final concentration). The respective lipid images are
id substrates migrate slower due to different fatty acid chains. Data are means inoccasions. Error bars show the SEM. (B) Western blot demonstrating the presence of
(C–G) Lipid kinase assays of both the recombinant P. falciparum PIP5K domain andline at final concentrations of 8 and 100 lM, respectively. Data represent means ofnt occasions. (C) and (D), both the recombinant P. falciparum PIP5K domain and thece of either GDP or GTP-cS (40 lM final concentration). E and F, both types of PIP5Ksant mammalian ARF1. (G) The recombinant P. falciparum PIP5K domain was assayedcentration). (H) Generation of PtdIns(4,5)P2 by the recombinant P. falciparum PIP5Kfinal concentration of PtdIns4P was 40 lM. Data represent means of duplicates of a
r bars represent the SEM. mPIP5Ka, mammalian Type I PIP5Ka; pPIP5K, P. falciparumphatidylinositol 3-phosphate; PIP2, phosphatidylinositol 4,5-bisphosphate; PI4P,
"

W. Leber et al. / International Journal for Parasitology 39 (2009) 645–653 651
dar, 1992). Incubation of cultures of asexual parasites with BFAprevents parasite maturation and results in the reshaping of theGolgi and in the formation of an ER-like compartment in the
periphery of the parasite cell (Crary and Haldar, 1992; Elmendorfand Haldar, 1993; Wickham et al., 2001). In addition, traffickingof parasite proteins such as the P. falciparum erythrocyte mem-

652 W. Leber et al. / International Journal for Parasitology 39 (2009) 645–653
brane proteins PfEMP1 and PfEMP3, and the knob-associated histi-dine-rich protein (KHARP) to the surface of the infected red bloodcell (Wickham et al., 2001), and transport of invasion proteins suchas the rhoptry-associated membrane protein (RAMA), the rhoptry-associated protein 1 (RAP-1), and the merozoite surface proteinsMSP1 and MSP7 through the Golgi cisterna have been shown tobe BFA-sensitive (Howard and Schmidt, 1995; Noe et al., 2000;Topolska et al., 2004; Pachebat et al., 2007). BFA prevents thenucleotide exchange on ARF from its inactive (ARF-GDP) into its ac-tive form (ARF-GTP) by interfering with the Sec7 domain of a largeARF-GEF (Chardin and McCormick, 1999). In contrast to mamma-lian cells, where a vast number of different GEFs have been de-scribed, only a single large BFA-sensitive ARF-GEF (PfARF-GEF) ispredicted to be present in the plasmodial genome (Jackson and Casa-nova, 2000; Baumgartner et al., 2001). Most likely the activation ofARF proteins in Plasmodium is regulated by the single PfARF-GEF(Baumgartner et al., 2001). Recently, a BFA-resistant parasite linewas generated by exchanging methionine for isoleucine at position1473 located within the Sec7 domain of wild-type PfARF-GEFthrough allelic replacement (Wiek et al., 2004). In these transfec-tants, viability, protein trafficking and the Golgi structure of asexualparasites were intact in the presence of BFA (Wiek et al., 2004).
The presence of a BFA-sensitive secretory pathway and the evi-dence for ARF-mediated activation of plasmodial PIP5K presentedin this paper strongly suggest that the entire (ARF-GEF)-ARF-PIP5Ksignalling pathway is intact in the malaria parasite. Parasite hostcell invasion, trafficking of proteins to the surface of infected redblood cells and exflagellation are sophisticated cellular events thatrequire the interaction of several signalling pathways to regulatethe cytoskeletal rearrangements and membrane dynamics re-quired for these processes. Through its individual domains, plas-modial PIP5K/NCS might form a focal point in unifying thesedifferent pathways.
Acknowledgements
W. Leber and Q.L. Fivelman were supported by Wellcome Trustgrants to D.A. Baker (Ref. 066742 and 069972). W. Leber was addi-tionally funded by the University of Innsbruck, Austria. S. Cockcroftthanks the Wellcome Trust for their support, and A. Skippen was inreceipt of a Wellcome Trust Prize studentship (Ref. 043166). P.W.Bowyer was supported by a Wellcome Trust studentship (Ref.065514). We also thank Deborah Smith from Imperial Collegeand Tony Holder from the National Institute for Medical Research(both UK) for providing recombinant myristoylated PfARF1. Wethank Sarah Sharp from Colin Sutherland’s group at the LondonSchool of Hygiene & Tropical Medicine for providing the Northernblot. We are also grateful to Wensheng Deng for providing techni-cal advice. We acknowledge the contribution of the sequencingcentres (The Institute of Genomic Research, the Wellcome TrustSanger Institute, and the Stanford DNA Sequencing and TechnologyCenter) that provided genomic sequences used in this study. Weare also grateful to GenBank and PlasmoDB for making genomicdata freely available (Bahl et al., 2003; Benson et al., 2008).
References
Ames, J.B., Ishima, R., Tanaka, T., Gordon, J.I., Stryer, L., Ikura, M., 1997. Molecularmechanics of calcium-myristoyl switches. Nature 389, 198–202.
Ames, J.B., Levay, K., Wingard, J.N., Lusin, J.D., Slepak, V.Z., 2006. Structural basis forcalcium-induced inhibition of rhodopsin kinase by recoverin. J. Biol. Chem. 281,37237–37245.
Arneson, L.S., Kunz, J., Anderson, R.A., Traub, L.M., 1999. Coupled inositidephosphorylation and phospholipase D activation initiates clathrin-coatassembly on lysosomes. J. Biol. Chem. 274, 17794–17805.
Bahl, A., Brunk, B., Crabtree, J., Fraunholz, M.J., Gajria, B., Grant, G.R., Ginsburg, H.,Gupta, D., Kissinger, J.C., Labo, P., Li, L., Mailman, M.D., Milgram, A.J., Pearson,D.S., Roos, D.S., Schug, J., Stoeckert Jr., C.J., Whetzel, P., 2003. PlasmoDB: the
Plasmodium genome resource. A database integrating experimental andcomputational data. Nucleic Acids Res. 31, 212–215.
Baumgartner, F., Wiek, S., Paprotka, K., Zauner, S., Lingelbach, K., 2001. A pointmutation in an unusual Sec7 domain is linked to brefeldin A resistance in aPlasmodium falciparum line generated by drug selection. Mol. Microbiol. 41,1151–1158.
Benson, D.A., Karsch-Mizrachi, I., Lipman, D.J., Ostell, J., Wheeler, D.L., 2008.GenBank. Nucleic Acids Res. 36, D25–D30.
Billker, O., Lindo, V., Panico, M., Etienne, A.E., Paxton, T., Dell, A., Rogers, M., Sinden,R.E., Morris, H.R., 1998. Identification of xanthurenic acid as the putativeinducer of malaria development in the mosquito. Nature 392, 289–292.
Burgoyne, R.D., Weiss, J.L., 2001. The neuronal calcium sensor family of Ca2+-binding proteins. Biochem. J. 353, 1–12.
Burgoyne, R.D., O’Callaghan, D.W., Hasdemir, B., Haynes, L.P., Tepikin, A.V., 2004.Neuronal Ca2+-sensor proteins: multitalented regulators of neuronal function.Trends Neurosci. 27, 203–209.
Bzik, D.J., Li, W.B., Horii, T., Inselburg, J., 1987. Molecular cloning and sequenceanalysis of the Plasmodium falciparum dihydrofolate reductase-thymidylatesynthase gene. Proc. Natl. Acad. Sci. USA 84, 8360–8364.
Carucci, D.J., Witney, A.A., Muhia, D.K., Warhurst, D.C., Schaap, P., Meima, M., Li, J.L.,Taylor, M.C., Kelly, J.M., Baker, D.A., 2000. Guanylyl cyclase activity associatedwith putative bifunctional integral membrane proteins in Plasmodiumfalciparum. J. Biol. Chem. 275, 22147–22156.
Chardin, P., McCormick, F., 1999. Brefeldin A: the advantage of being uncompetitive.Cell 97, 153–155.
Clarke, J.L., Scopes, D.A., Sodeinde, O., Mason, P.J., 2001. Glucose-6-phosphatedehydrogenase-6-phosphogluconolactonase. A novel bifunctional enzyme inmalaria parasites. Eur. J. Biochem. 268, 2013–2019.
Cockcroft, S., Thomas, G.M., Fensome, A., Geny, B., Cunningham, E., Gout, I., Hiles, I.,Totty, N.F., Truong, O., Hsuan, J.J., 1994. Phospholipase D: a downstream effectorof ARF in granulocytes. Science 263, 523–526.
Cockcroft, S., 2000. Biology of Phosphoinositides. Oxford University Press.Cockcroft, S., 2001. Signalling roles of mammalian phospholipase D1 and D2. Cell.
Mol. Life Sci. 58, 1674–1687.Crary, J.L., Haldar, K., 1992. Brefeldin A inhibits protein secretion and parasite
maturation in the ring stage of Plasmodium falciparum. Mol. Biochem. Parasitol.53, 185–192.
Divecha, N., Roefs, M., Halstead, J.R., D’Andrea, S., Fernandez-Borga, M., Oomen, L.,Saqib, K.M., Wakelam, M.J., D’Santos, C., 2000. Interaction of the type IalphaPIPkinase with phospholipase D: a role for the local generation ofphosphatidylinositol 4,5-bisphosphate in the regulation of PLD2 activity.EMBO J. 19, 5440–5449.
Duronio, R.J., Jackson-Machelski, E., Heuckeroth, R.O., Olins, P.O., Devine, C.S.,Yonemoto, W., Slice, L.W., Taylor, S.S., Gordon, J.I., 1990. Protein N-myristoylation in Escherichia coli: reconstitution of a eukaryotic proteinmodification in bacteria. Proc. Natl. Acad. Sci. USA 87, 1506–1510.
Elmendorf, H.G., Haldar, K., 1993. Identification and localization of ERD2 in themalaria parasite Plasmodium falciparum: separation from sites of sphingomyelinsynthesis and implications for organization of the Golgi. EMBO J. 12, 4763–4773.
Ermilov, A.N., Olshevskaya, E.V., Dizhoor, A.M., 2001. Instead of binding calcium,one of the EF-hand structures in guanylyl cyclase activating protein-2 isrequired for targeting photoreceptor guanylyl cyclase. J. Biol. Chem. 276,48143–48148.
Exton, J.H., 1998. Phospholipase D. Biochim. Biophys. Acta 1436, 105–115.Fairlamb, A.H., Warhurst, D.C., Peters, W., 1985. An improved technique for the
cultivation of Plasmodium falciparum in vitro without daily medium change.Ann. Trop. Med. Parasitol. 79, 379–384.
Fik-Rymarkiewicz, E., Duda, T., Sharma, R.K., 2006. Novel frequenin-modulatedCa2+-signaling membrane guanylate cyclase (ROS-GC) transduction pathway inbovine hippocampus. Mol. Cell. Biochem. 291, 187–204.
Fivelman, Q.L., McRobert, L., Sharp, S., Taylor, C.J., Saeed, M., Swales, C.A.,Sutherland, C.J., Baker, D.A., 2007. Improved synchronous production ofPlasmodium falciparum gametocytes in vitro. Mol. Biochem. Parasitol. 154,119–123.
Godi, A., Pertile, P., Meyers, R., Marra, P., Di Tullio, G., Iurisci, C., Luini, A., Corda, D.,De Matteis, M.A., 1999. ARF mediates recruitment of PtdIns-4-OH kinase-betaand stimulates synthesis of PtdIns(4,5)P2 on the Golgi complex. Nat. Cell Biol. 1,280–287.
Haynes, L.P., Thomas, G.M., Burgoyne, R.D., 2005. Interaction of neuronal calciumsensor-1 and ADP-ribosylation factor 1 allows bidirectional control ofphosphatidylinositol 4-kinase beta and trans-Golgi network-plasmamembrane traffic. J. Biol. Chem. 280, 6047–6054.
Haynes, L.P., Sherwood, M.W., Dolman, N.J., Burgoyne, R.D., 2007. Specificity,promiscuity and localization of ARF protein interactions with NCS-1 andphosphatidylinositol-4 kinase-III beta. Traffic 8, 1080–1092.
Hendricks, K.B., Wang, B.Q., Schnieders, E.A., Thorner, J., 1999. Yeast homologue ofneuronal frequenin is a regulator of phosphatidylinositol-4-OH kinase. Nat. CellBiol. 1, 234–241.
Honda, A., Nogami, M., Yokozeki, T., Yamazaki, M., Nakamura, H., Watanabe, H.,Kawamoto, K., Nakayama, K., Morris, A.J., Frohman, M.A., Kanaho, Y., 1999.Phosphatidylinositol 4-phosphate 5-kinase alpha is a downstream effector ofthe small G protein ARF6 in membrane ruffle formation. Cell 99, 521–532.
Howard, R.F., Schmidt, C.M., 1995. The secretary pathway of Plasmodium falciparumregulates transport of p82/RAP1 to the rhoptries. Mol. Biochem. Parasitol. 74,43–54.

W. Leber et al. / International Journal for Parasitology 39 (2009) 645–653 653
Hwang, J.Y., Schlesinger, R., Koch, K.W., 2004. Irregular dimerization of guanylatecyclase-activating protein 1 mutants causes loss of target activation. Eur. J.Biochem. 271, 3785–3793.
Ishihara, H., Shibasaki, Y., Kizuki, N., Katagiri, H., Yazaki, Y., Asano, T., Oka, Y., 1996.Cloning of cDNAs encoding two isoforms of 68-kDa type I phosphatidylinositol-4-phosphate 5-kinase. J. Biol. Chem. 271, 23611–23614.
Ishihara, H., Shibasaki, Y., Kizuki, N., Wada, T., Yazaki, Y., Asano, T., Oka, Y., 1998.Type I phosphatidylinositol-4-phosphate 5-kinases. Cloning of the third isoformand deletion/substitution analysis of members of this novel lipid kinase family.J. Biol. Chem. 273, 8741–8748.
Jackson, C.L., Casanova, J.E., 2000. Turning on ARF: the Sec7 family of guanine-nucleotide-exchange factors. Trends Cell Biol. 10, 60–67.
Jones, D.H., Bax, B., Fensome, A., Cockcroft, S., 1999. ADP ribosylation factor 1mutants identify a phospholipase D effector region and reveal thatphospholipase D participates in lysosomal secretion but is not sufficient forrecruitment of coatomer I. Biochem. J. 341 (Pt 1), 185–192.
Jones, D.H., Morris, J.B., Morgan, C.P., Kondo, H., Irvine, R.F., Cockcroft, S., 2000. TypeI phosphatidylinositol 4-phosphate 5-kinase directly interacts with ADP-ribosylation factor 1 and is responsible for phosphatidylinositol 4,5-bisphosphate synthesis in the Golgi compartment. J. Biol. Chem. 275, 13962–13966.
Kawamoto, F., Alejo-Blanco, R., Fleck, S.L., Kawamoto, Y., Sinden, R.E., 1990. Possibleroles of Ca2+ and cGMP as mediators of the exflagellation of Plasmodium bergheiand Plasmodium falciparum. Mol. Biochem. Parasitol. 42, 101–108.
Kunz, J., Wilson, M.P., Kisseleva, M., Hurley, J.H., Majerus, P.W., Anderson, R.A., 2000.The activation loop of phosphatidylinositol phosphate kinases determinessignaling specificity. Mol. Cell 5, 1–11.
Kunz, J., Fuelling, A., Kolbe, L., Anderson, R.A., 2002. Stereo-specific substraterecognition by phosphatidylinositol phosphate kinases is swapped by changinga single amino acid residue. J. Biol. Chem. 277, 5611–5619.
Kyes, S., Pinches, R., Newbold, C., 2000. A simple RNA analysis method shows varand rif multigene family expression patterns in Plasmodium falciparum. Mol.Biochem. Parasitol. 105, 311–315.
Lee, F.J., Patton, W.A., Lin, C.Y., Moss, J., Vaughan, M., Goldman, N.D., Syin, C., 1997.Identification and characterization of an ADP-ribosylation factor in Plasmodiumfalciparum. Mol. Biochem. Parasitol. 87, 217–223.
Martin, S.K., Jett, M., Schneider, I., 1994. Correlation of phosphoinositide hydrolysiswith exflagellation in the malaria microgametocyte. J. Parasitol. 80, 371–378.
Martin, T.F., 2001. PI(4,5)P(2) regulation of surface membrane traffic. Curr. Opin.Cell Biol. 13, 493–499.
Nijhout, M.M., Carter, R., 1978. Gamete development in malaria parasites:bicarbonate-dependent stimulation by pH in vitro. Parasitology 76, 39–53.
Noe, A.R., Fishkind, D.J., Adams, J.H., 2000. Spatial and temporal dynamics of thesecretory pathway during differentiation of the Plasmodium yoelii schizont. Mol.Biochem. Parasitol. 108, 169–185.
Ogwan’g, R.A., Mwangi, J.K., Githure, J., Were, J.B., Roberts, C.R., Martin, S.K., 1993.Factors affecting exflagellation of in vitro-cultivated Plasmodium falciparumgametocytes. Am. J. Trop. Med. Hyg. 49, 25–29.
Pachebat, J.A., Kadekoppala, M., Grainger, M., Dluzewski, A.R., Gunaratne, R.S., Scott-Finnigan, T.J., Ogun, S.A., Ling, I.T., Bannister, L.H., Taylor, H.M., Mitchell, G.H.,Holder, A.A., 2007. Extensive proteolytic processing of the malaria parasitemerozoite surface protein 7 during biosynthesis and parasite release fromerythrocytes. Mol. Biochem. Parasitol. 151, 59–69.
Powner, D.J., Payne, R.M., Pettitt, T.R., Giudici, M.L., Irvine, R.F., Wakelam, M.J., 2005.Phospholipase D2 stimulates integrin-mediated adhesion viaphosphatidylinositol 4-phosphate 5-kinase Igamma b. J. Cell Sci. 118, 2975–2986.
Promeneur, D., Liu, Y., Maciel, J., Agre, P., King, L.S., Kumar, N., 2007.Aquaglyceroporin PbAQP during intraerythrocytic development of the malariaparasite Plasmodium berghei. Proc. Natl. Acad. Sci. USA 104, 2211–2216.
Rajebhosale, M., Greenwood, S., Vidugiriene, J., Jeromin, A., Hilfiker, S., 2003.Phosphatidylinositol 4-OH kinase is a downstream target of neuronal calciumsensor-1 in enhancing exocytosis in neuroendocrine cells. J. Biol. Chem. 278,6075–6084.
Rao, V.D., Misra, S., Boronenkov, I.V., Anderson, R.A., Hurley, J.H., 1998. Structure oftype IIbeta phosphatidylinositol phosphate kinase: a protein kinase foldflattened for interfacial phosphorylation. Cell 94, 829–839.
Renner, M., Danielson, M.A., Falke, J.J., 1993. Kinetic control of Ca(II) signaling:tuning the ion dissociation rates of EF-hand Ca(II) binding sites. Proc. Natl. Acad.Sci. USA 90, 6493–6497.
Salcedo, E., Cortese, J.F., Plowe, C.V., Sims, P.F., Hyde, J.E., 2001. A bifunctionaldihydrofolate synthetase–folylpolyglutamate synthetase in Plasmodiumfalciparum identified by functional complementation in yeast and bacteria.Mol. Biochem. Parasitol. 112, 239–252.
Siddhanta, A., Backer, J.M., Shields, D., 2000. Inhibition of phosphatidic acidsynthesis alters the structure of the Golgi apparatus and inhibits secretion inendocrine cells. J. Biol. Chem. 275, 12023–12031.
Sinden, R.E., Canning, E.U., Spain, B., 1976. Gametogenesis and fertilization inPlasmodium yoelii nigeriensis: a transmission electron microscope study. Proc. R.Soc. Lond. B Biol. Sci. 193, 55–76.
Skippen, A., Jones, D.H., Morgan, C.P., Li, M., Cockcroft, S., 2002. Mechanism of ADPribosylation factor-stimulated phosphatidylinositol 4,5-bisphosphate synthesisin HL60 cells. J. Biol. Chem. 277, 5823–5831.
Stafford, W.H., Stockley, R.W., Ludbrook, S.B., Holder, A.A., 1996. Isolation,expression and characterization of the gene for an ADP-ribosylation factorfrom the human malaria parasite, Plasmodium falciparum. Eur. J. Biochem. 242,104–113.
Strahl, T., Huttner, I.G., Lusin, J.D., Osawa, M., King, D., Thorner, J., Ames, J.B., 2007.Structural insights into activation of phosphatidylinositol 4-kinase (pik1) byyeast frequenin (Frq1). J. Biol. Chem. 282, 30949–30959.
Takimoto, K., Yang, E.K., Conforti, L., 2002. Palmitoylation of KChIP splicing variantsis required for efficient cell surface expression of Kv4.3 channels. J. Biol. Chem.277, 26904–26911.
Topolska, A.E., Lidgett, A., Truman, D., Fujioka, H., Coppel, R.L., 2004.Characterization of a membrane-associated rhoptry protein of Plasmodiumfalciparum. J. Biol. Chem. 279, 4648–4656.
Trager, W., Jensen, J.B., 1976. Human malaria parasites in continuous culture.Science 193, 673–675.
Weber, J.H., Vishnyakov, A., Hambach, K., Schultz, A., Schultz, J.E., Linder, J.U., 2004.Adenylyl cyclases from Plasmodium, Paramecium and Tetrahymena are novel ionchannel/enzyme fusion proteins. Cell Signal. 16, 115–125.
White, N.J., Nosten, F., Looareesuwan, S., Watkins, W.M., Marsh, K., Snow, R.W.,Kokwaro, G., Ouma, J., Hien, T.T., Molyneux, M.E., Taylor, T.E., Newbold, C.I.,Ruebush 2nd, T.K., Danis, M., Greenwood, B.M., Anderson, R.M., Olliaro, P., 1999.Averting a malaria disaster. Lancet 353, 1965–1967.
Wickham, M.E., Rug, M., Ralph, S.A., Klonis, N., McFadden, G.I., Tilley, L., Cowman,A.F., 2001. Trafficking and assembly of the cytoadherence complex inPlasmodium falciparum-infected human erythrocytes. EMBO J. 20, 5636–5649.
Wiek, S., Cowman, A.F., Lingelbach, K., 2004. Double cross-over gene replacementwithin the sec 7 domain of a GDP-GTP exchange factor from Plasmodiumfalciparum allows the generation of a transgenic brefeldin A-resistant parasiteline. Mol. Biochem. Parasitol. 138, 51–55.
Wrenger, C., Luersen, K., Krause, T., Muller, S., Walter, R.D., 2001. The Plasmodiumfalciparum bifunctional ornithine decarboxylase, S-adenosyl-L-methioninedecarboxylase, enables a well balanced polyamine synthesis without domain–domain interaction. J. Biol. Chem. 276, 29651–29656.
Zhao, X., Varnai, P., Tuymetova, G., Balla, A., Toth, Z.E., Oker-Blom, C., Roder, J.,Jeromin, A., Balla, T., 2001. Interaction of neuronal calcium sensor-1 (NCS-1)with phosphatidylinositol 4-kinase beta stimulates lipid kinase activity andaffects membrane trafficking in COS-7 cells. J. Biol. Chem. 276, 40183–40189.
Copyright © 2022 FDOKUMEN




















