
DNA-Induced Dimerization of Poly(ADP-ribose) Polymerase1 Triggers Its Activation

Receptor-Like Activity Evoked by Extracellular ADP inArabidopsis Root Epidermal Plasma Membrane1
Vadim Demidchik, Zhonglin Shang, Ryoung Shin, Renato Colaco, Anuphon Laohavisit,Sergey Shabala, and Julia M. Davies*
Department of Biology, University of Essex, Colchester CO4 3SQ, United Kingdom (V.D.); College of LifeScience, Hebei Normal University, Shijiazhang 050016, Hebei, China (Z.S.); Riken Plant Science Center,Yokohama City, Kanagawa 230–0045, Japan (R.S.); Department of Plant Sciences, University of Cambridge,Cambridge CB2 3EA, United Kingdom (R.C., A.L., J.M.D.); and School of Agricultural Science, University ofTasmania, Hobart, Tasmania 7001, Australia (S.S.)
Extracellular purine nucleotides are implicated in the control of plant development and stress responses. While extracellularATP is known to activate transcriptional pathways via plasma membrane (PM) NADPH oxidase and calcium channelactivation, very little is known about signal transduction by extracellular ADP. Here, extracellular ADP was found to activatenet Ca2+ influx in roots of Arabidopsis (Arabidopsis thaliana) and transiently elevate cytosolic free Ca2+ in root epidermalprotoplasts. An inward Ca2+-permeable conductance in root epidermal PM was activated within 1 s of ADP application andrepeated application evoked a smaller current. Such response speed and densitization are consistent with operation ofequivalents to animal ionotropic purine receptors, although to date no equivalent genes for such receptors have been identifiedin higher plants. In contrast to ATP, extracellular ADP did not evoke accumulation of intracellular reactive oxygen species.While high concentrations of ATP caused net Ca2+ efflux from roots, equivalent concentrations of ADP caused net influx.Overall the results point to a discrete ADP signaling pathway, reliant on receptor-like activity at the PM.
Extracellular purine nucleotides have been shownto be involved in the regulation of plant cell viability,membrane permeability, immunity, symbiosis, stressresponses, and growth (Lew and Dearnaley, 2000; Tanget al., 2003; Chivasa et al., 2005, 2009; Kim et al., 2006,2009; Roux and Steinebrunner, 2007; Wu et al., 2007;Riewe et al., 2008a; Wu and Wu, 2008; Yi et al., 2008;Demidchik et al., 2009; Govindarajulu et al., 2009; Kimet al., 2009; Clark et al., 2010a, 2010b; Tanaka et al.,2010a, 2010b; Terrile et al., 2010; Tonon et al., 2010).Purine nucleotide release from plant cells may occurthrough wounding, exocytosis, or through the activityof plasmamembrane (PM) ATP-binding cassette trans-porters (Thomas et al., 2000; Kim et al., 2006). Growthpoints are hotspots of extracellular ATP (Kim et al.,2006) while touch-stimulated ATP release is mediatedby heterotrimeric G-protein activity (Weerasinghe et al.,2009). Levels of extracellular ATP and ADP can be reg-ulated by the concerted action of ecto-apyrases, phos-
phatases, and adenosine nucleosidases with retrieval ofadenine (the ultimate hydrolytic product) by PM purinepermeases (Komoszynski, 1996; Steinebrunner et al.,2003; Wu et al., 2007; Riewe et al., 2008a, 2008b;Govindarajulu et al., 2009; Clark et al., 2010b; Lianget al., 2010). Manipulation of ecto-apyrase levels re-sults in abnormal growth in Arabidopsis (Arabidopsisthaliana), cotton (Gossypium hirsutum), and potato (So-lanum tuberosum; Wu et al., 2007; Riewe et al., 2008a;Clark et al., 2010a, 2010b) while nodulation by Brady-rhizobium japonicum is impaired in ecto-apyrase-deficientsoybean (Glycine max; Govindarajulu et al., 2009).Current models suggest that poise of the cell’s statebetween death, stress adaptation, and growth involvessignaling governed by the level of extracellular ATP(Chivasa et al., 2005, 2009; Roux and Steinebrunner,2007; Wu et al., 2007; Wu and Wu, 2008; Kim et al.,2009; Clark et al., 2010a, 2010b; Terrile et al., 2010;Tonon et al., 2010).
The mechanism of purine nucleotide perception isunclear as higher plant genomes do not encode forhomologs of animal PM purinoceptors (Kim et al.,2006; Fountain et al., 2008). The latter are either cou-pled to heterotrimeric G proteins or are ionotropicreceptors (activated by nanomolar or even 0.1 mM
ATP) that permit Ca2+ influx to transduce the purinenucleotide signal (for review, see Khakh and North,2006). It remains possible that plants evolved struc-turally different systems for purine sensing to animalsbut antagonists of animal purinoceptors are effectiveagainst plant extracellular ATP responses. Despite the
1 This work was supported by the Leverhulme Trust (grant no.F/09 741/C), the Biotechnology and Biological Sciences ResearchCouncil, the University of Cambridge Frank Smart Trust andBrookes Fund, the Royal Society (China Incoming Fellowship), theAustralian Research Council, and The Monsanto Company.
* Corresponding author; e-mail [email protected] author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policydescribed in the Instructions for Authors (www.plantphysiol.org) is:Julia M. Davies ([email protected]).
www.plantphysiol.org/cgi/doi/10.1104/pp.111.174722
Plant Physiology�, July 2011, Vol. 156, pp. 1375–1385, www.plantphysiol.org � 2011 American Society of Plant Biologists 1375

apparent absence of classical purinoceptors in higherplants, extracellular purine nucleotides can cause tran-sient elevations of cytosolic free Ca2+ ([Ca2+]cyt), incommon with animals (Demidchik et al., 2003a, 2009;Jeter et al., 2004; Tanaka et al., 2010b). In Arabidop-sis roots, extracellular ATP most likely triggers intra-cellular Ca2+ release, resulting in stimulation of PMNADPH oxidase activity (Demidchik et al., 2009). Theresultant reactive oxygen species (ROS) activate a PMCa2+ influx pathway that governs a transcriptionalresponse. In Arabidopsis root hairs, extracellular ATPalso causes intracellular ROS accumulation by activat-ing NADPH oxidases (Clark et al., 2010b). Leaves alsosupport extracellular ATP-induced transcription viaPM NADPH oxidases (Song et al., 2006). Nitric oxideis now also established as a product of extracellularpurine nucleotide perception. Nitric oxide evolutionby plants has been observed on application of extra-cellular ATP or extracellular ADP and linked to influxof Ca2+ or production of phosphatidic acid (Foresiet al., 2007; Wu and Wu, 2008; Reichler et al., 2009;Clark et al., 2010b; Sueldo et al., 2010; Tonon et al.,2010).
In this study, the effects of extracellular ADP on iontransport at the plant PM have been examined. Extra-cellular ADP elevates [Ca2+]cyt in both roots and leavesof Arabidopsis and it can be equally or more effectivethan extracellular ATP (Demidchik et al., 2003a, 2009;Jeter et al., 2004; Tanaka et al., 2010b). In common withextracellular ATP, extracellular ADP can elicit oscilla-tions of [Ca2+]cyt in Arabidopsis seedlings (Tanakaet al., 2010b). At the physiological level, extracellularADP stimulates nodule development (Govindarajuluet al., 2009) and was shown to stimulate root hairelongation (Lew and Dearnaley, 2000). In commonwith extracellular ATP, it is now known that extracel-lular ADP has a biphasic effect on cotton fiber and roothair elongation with high concentrations being inhib-itory (Clark et al., 2010a, 2010b). Arabidopsis rootepidermal cells have been used here as these are wellcharacterized electrophysiologically and are amongthe first cell types to sense the pathogen attack andenvironmental change that could involve purine sig-naling. By using a fast-perfusion system in patch-clampelectrophysiology experiments, extracellular ADP hasbeen delivered to the extracellular PM face of proto-plasts within 1 s, permitting the fastest resolution todate of rapid activation of plant ion transport eventsby an extracellular purine nucleotide. These events ap-pear to be independent of heterotrimeric G proteins.Epidermal protoplasts expressing the [Ca2+]cyt re-porter aequorin have been used to explore further theability of extracellular ADP to evoke [Ca2+]cyt elevation.In further contrast to extracellular ATP, extracellularADP does not elicit intracellular ROS accumulation.Using whole roots, it has also been possible to resolveextracellular ADP-induced net Ca2+ and K+ fluxes; highconcentrations of ATP but not ADP evoke net Ca2+ efflux.Overall the results show rapid activation of PM Ca2+—and K+—conductances by extracellular ADP, with
initial events supported by PM ionotropic receptor-like activity. The differential responses of intracellularROS and net Ca2+ flux to extracellular ADP and ex-tracellular ATP could provide the basis for distinctsignaling outcomes.
RESULTS
Extracellular ADP Activates a PM Ca2+
Influx Conductance
To establish whether the PM could respond to extra-cellular ADP, protoplasts from mature epidermal rootcells were assayed using the whole-cell patch-clampconfiguration to resolve activity of channel populationsas a function of applied membrane voltage and extra-cellular agonist. Application of 20 mM extracellular ADPto protoplasts (using conventional peristaltic pumpdelivery to the recording chamber) transiently activateda hyperpolarization-activated (i.e. activated by increas-ingly negative voltage) Ca2+-permeable conductance(Fig. 1A), very similar to that characterized in responseto 20 mM extracellular ATP in this cell type (Demidchiket al., 2009). The concentration used is representative ofwounding levels (Song et al., 2006). The increase ininwardly directed current at hyperpolarized (insidenegative) voltage was observed 1 to 3 min after extra-cellular ADP addition (a similar time course to the ex-tracellular ATP response reported by Demidchik et al.,2009; Shang et al., 2009) and was evident over thephysiological voltage range for this cell type (Maathuisand Sanders, 1993). Mean 6 SE current-voltage (IV)relationships are shown in Figure 1B; negative voltagesare physiologically relevant. At 2200 mV, extracellularADP induced a peak mean 6 SE current increase from2163 6 36 pA to 2382 6 63 pA at 2200 mV (n = 4);current then declined over a 5 to 15 min period.Negative current signifies influx of positive charge intothe cytosol or efflux of negative charge. The peak cur-rent response is not significantly different (Student’st test) to that found for 20 mM extracellular ATP(Demidchik et al., 2009).
Rapid, Reversible, and Desensitizing Response to
Extracellular ADP at the PM
Having found that the PMwas electrically responsiveto extracellular ADP on slow perfusion and long-termexposure, a fast-perfusion system was used to deliverextracellular ADP to the PM within 1 s. In contrast toexperiments using peristaltic pump delivery of theagonist to the recording chamber (with a waiting timefor flow from the chamber edge to the protoplast), thefast-perfusion system used a primed multichannel de-livery pipette placed near the protoplast (describedfurther in “Materials and Methods”). In these experi-ments, the pipette solution (PS; equivalent to the cyto-sol) contained a nonhydrolyzable GDP analog (0.5 mM
GDPbS) to minimize G-protein-mediated channel acti-vation (Li and Assmann, 1993). In preliminary experi-
Demidchik et al.
1376 Plant Physiol. Vol. 156, 2011

ments, this increased the K+ efflux current, as describedby Li and Assmann (1993), indicative of effectiveG-protein inhibition. It has been proposed that extracel-lular purine nucleotides can relay a signal via hetero-trimeric G proteins in plants (Song et al., 2006; Rouxand Steinebrunner, 2007; Wu and Wu, 2008), so theirinhibition could delineate possible direct extracellularADP effects on a receptor-like channel. Experimentswereperformed within the first 20 to 30 min of recording sothat extracellular ADP-activated currents could be dis-tinguished from the time-dependent developmentof the constitutive hyperpolarization-activated Ca2+-permeable conductance. The latter becomes the dominantCa2+ conductance in this cell type after approximately1 h (Demidchik et al., 2002).Fast perfusion proved problematic as in approxi-
mately 80% of trials (65 out of 85), the recording
configuration was disturbed or lost by distortion of theprotoplast. Given the high failure rate, a higher con-centration of extracellular ADP (0.3 mM) was used inthese trials to ensure activation. In the remaining 15trials in which seal resistance was unaffected by fastperfusion, extracellular ADP evoked a rapid and re-versible change in PM conductance (Fig. 2A). With thePM clamped at a holding potential (HP) of 290 mV,fast perfusion with control bathing solution (BS) waswithout effect (Fig. 2A, top trace). Addition of 0.3 mM
extracellular ADP caused a downward current deflec-tion, consistent with entry of positive charge into theprotoplast (Fig. 2A, middle trace). Current activationoccurred immediately after exposure to extracellularADP. Fast removal of extracellular ADP resulted inimmediate recovery to control conditions. As can beseen from the middle trace of Figure 2A, a repeatedapplication of 0.3 mM extracellular ADP to the sameprotoplast again caused rapid and reversible currentactivation. However in this case, recovery to baselinecurrent began in the presence of extracellular ADP.This was typical of all protoplasts where extracellularADP activation was observed.
Figure 1. Extracellular ADP activates a PM influx conductance in rootepidermal protoplasts. A, Effect of 20 mM extracellular ADP on whole-cell currents from a representative protoplast, recording conditionswere as used previously by Demidchik et al. (2009). B, Mean 6 SE IVrelationships of control and peak ADP peak response (n = 4). Control BSwas as described in “Materials and Methods” (20 mM CaCl2, 0.1 mM
KCl, and Na+ equimolar to test BS, pH 6). PS comprised (mM): 40K-gluconate, 10 KCl, 1 1,2-bis(2-aminophenoxy)ethane-N,N,N’,N’-tetraacetic acid (free Ca2+ adjusted to 100 nM with 0.4 CaCl2), pH 7.2adjusted with Tris/MES.
Figure 2. Rapid activation of epidermal PM cation conductances byextracellular ADP. The results are from whole-cell recordings onprotoplasts from the root mature epidermis. A, Top trace: two sequen-tial applications of control BS (green bars) to a protoplast from themature epidermis failed to elicit a current; HP across the PM was 290mV inside negative. Middle trace (same protoplast): two sequentialapplications to the same protoplast of 0.3 mM ADP (red bars) by fastperfusion evoked inwardly directed current (downward deflection).Baseline current was recovered on removal of the ADP stimulus.Bottom trace (different protoplast); inhibiting effect of 0.3 mM Gd3+,added together with ADP in the second application. BS was as in Figure1. Test BS included 0.3 mM NaADP 6 0.3 mM GdCl3 as indicated. PSincluded 0.5 mM GDPbS. B, Current in response to voltage ramp (40-slong started from +100 mV) recorded from a single protoplast, approx-imately 1 min after the addition of control BS (green), 0.3 mM ADP (red),or 0.3 mM ADP with 0.3 mM Gd3+ (blue). Solutions as in A.
Receptor-Like Activity Evoked by Extracellular ADP
Plant Physiol. Vol. 156, 2011 1377

Mean 6 SE values of extracellular ADP-evokedcurrents at HP = 290 mV are given in Table I. Inexperimental set A, the first application of 0.3 mM
extracellular ADP (event 1) caused a mean 89% in-crease in inwardly directed current. After removal ofthe stimulus and recovery to baseline current, a secondapplication of 0.3 mM extracellular ADP (event 2)caused a mean 54% increase. All differences are sta-tistically significant (Student’s t test; P , 0.05). Inexperimental set B, event 1 (0.3 mM extracellular ADP)evoked amean 101% increase in inwardly directed currentat HP = 290 mV; in event 2, application of 0.3 mM extra-cellularADPand 0.3mMGd3+ to the sameprotoplasts onlycaused a mean 11% increase. Thus, the greater part of theextracellular ADP-induced current was sensitive to thecation channel blocker, Gd3+. To explore cation channelactivity further, IV relationships were determined.
Extracellular ADP Increases PM Ca2+ Influx and K+
Efflux Conductance
As Figure 2B shows, fast delivery of 0.3 mM extra-cellular ADP increased both inwardly and outwardlydirected currents across the PM of a single protoplast,resulting in a positive shift of reversal potential (Erev)away from the equilibrium potentials for K+ and Cl2
(EK =2158 mV; ECl =236mV) and toward that for Ca2+
(ECa = 155 mV). The mean Erev of the extracellular ADP-induced current (estimated by subtracting the controlIV from that determined in the presence of extracellularADP; Fig. 3, inset) was 4.7 6 8.4 mV (n = 4), indicatingCa2+ permeation. Extracellular ADP approximatelydoubled the Gd3+-sensitive inward current (Figs. 2Band 3) over the physiological voltage range for this celltype (2100 to 2150 mV; Maathuis and Sanders, 1993);thus, Ca2+ could be delivered to the cell to act as a signal
in response to the extracellular purine nucleotide. Forexample, from the mean response shown in Figure 3, at2120 mV inward Ca2+ current increased from2696 23mA m22 to 2159 6 25 mA in response to 0.3 mM
extracellular ADP (n = 4; mean 6 SE). This large Gd3+-sensitive Ca2+ influx current helps explain an extracel-lular ADP-induced elevation in whole root and rootepidermal protoplast [Ca2+]cyt found in aequorin lumi-nometry assays (Demidchik et al., 2003a, 2009). In ourexperimental conditions only K+ efflux (50 mM K+ in thePS) could mediate the outward current (Shabala et al.,2006). Both inward and outward currents were alsosensitive to the cation channel blocker Gd3+. It may bethat both Ca2+ influx and K+ efflux conductances werecaused by the activation of one cation channel type orthat Ca2+ influx activated K+ efflux.
Extracellular ADP Elevates Root Epidermal [Ca2+]cyt
Epidermal protoplasts were isolated from the ma-ture epidermis of plants constitutively expressingcytosolic apoaequorin to examine extracellular ADP-induced [Ca2+]cyt elevation. Previously, such proto-plasts were found to sustain transient [Ca2+]cyt elevationsin response to 0.1 mM extracellular ATP, its poorlyhydrolyzable analog adenosine 5#-(a,b-methylene)triphosphate (abATP), and ADP. All these were inhib-ited by Gd3+ and the purinoceptor antagonist, suramin(Demidchik et al., 2009). AMP (as an ADP breakdownproduct) up to 1 mM has been shown to be ineffective(Demidchik et al., 2009). A test concentration of 0.1 mM
ADP was used in this study as it is a feasible concen-tration to simulate a wound response (Song et al.,2006) and was found to cause Ca2+ current activationin four out of nine whole-cell patch-clamp trials us-ing slow perfusion (data not shown). Application of0.1 mM extracellular ADP at pH 6.0 caused a transientincrease in [Ca2+]cyt (Fig. 4A). Overall the mean peak
Table I. The effect of sequential extracellular ADP addition on PMcurrent
Control values were taken 1 to 3 s immediately preceding theapplication of ADP. Event 1 refers to the first application of ligand andEvent 2 to the subsequent application to the same protoplast. Record-ing conditions as in Figure 1A. HP = 290 mV.
Procedure Mean 6 SE Maximal Currents
mA m22
Set AEvent 1 (n = 4)Control 240.63 6 3.450.3 mM ADP 276.59 6 4.95
Event 2 (n = 3)Control 236.74 6 2.000.3 mM ADP 256.51 6 5.3
Set BEvent 1 (n = 3)Control 248.36 6 19.830.3 mM ADP 297.01 6 12.90
Event 2 (n = 3)0.3 mM ADP 243.4 6 15.230.3 mM ADP + 0.3 mM Gd3 248.33 6 21.30
Figure 3. Extracellular ADP activates Ca2+ influx and K+ efflux inepidermal protoplasts. Mean 6 SE IV relationships for control condi-tions and in the presence of 0.3 mM extracellular ADP, n = 4; solutionsas in Figure 2. Current values were taken from voltage ramps applied1 to 3 min after application of control BS or activation by ADP. Insert:difference IV relationship (control currents were subtracted from thevalues obtained after ADP activation).
Demidchik et al.
1378 Plant Physiol. Vol. 156, 2011
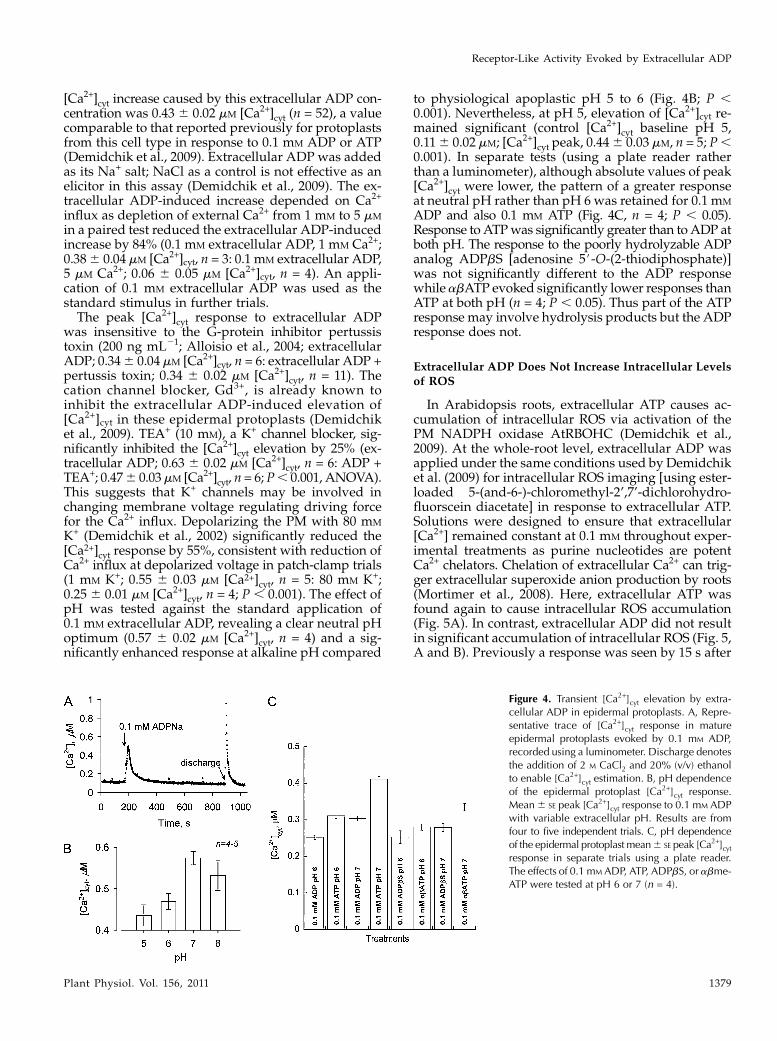
[Ca2+]cyt increase caused by this extracellular ADP con-centration was 0.43 6 0.02 mM [Ca2+]cyt (n = 52), a valuecomparable to that reported previously for protoplastsfrom this cell type in response to 0.1 mM ADP or ATP(Demidchik et al., 2009). Extracellular ADP was addedas its Na+ salt; NaCl as a control is not effective as anelicitor in this assay (Demidchik et al., 2009). The ex-tracellular ADP-induced increase depended on Ca2+
influx as depletion of external Ca2+ from 1 mM to 5 mM
in a paired test reduced the extracellular ADP-inducedincrease by 84% (0.1 mM extracellular ADP, 1 mM Ca2+;0.386 0.04 mM [Ca2+]cyt, n = 3: 0.1 mM extracellular ADP,5 mM Ca2+; 0.06 6 0.05 mM [Ca2+]cyt, n = 4). An appli-cation of 0.1 mM extracellular ADP was used as thestandard stimulus in further trials.The peak [Ca2+]cyt response to extracellular ADP
was insensitive to the G-protein inhibitor pertussistoxin (200 ng mL21; Alloisio et al., 2004; extracellularADP; 0.346 0.04mM [Ca2+]cyt, n = 6: extracellular ADP +pertussis toxin; 0.34 6 0.02 mM [Ca2+]cyt, n = 11). Thecation channel blocker, Gd3+, is already known toinhibit the extracellular ADP-induced elevation of[Ca2+]cyt in these epidermal protoplasts (Demidchiket al., 2009). TEA+ (10 mM), a K+ channel blocker, sig-nificantly inhibited the [Ca2+]cyt elevation by 25% (ex-tracellular ADP; 0.63 6 0.02 mM [Ca2+]cyt, n = 6: ADP +TEA+; 0.476 0.03mM [Ca2+]cyt, n = 6; P, 0.001, ANOVA).This suggests that K+ channels may be involved inchanging membrane voltage regulating driving forcefor the Ca2+ influx. Depolarizing the PM with 80 mM
K+ (Demidchik et al., 2002) significantly reduced the[Ca2+]cyt response by 55%, consistent with reduction ofCa2+ influx at depolarized voltage in patch-clamp trials(1 mM K+; 0.55 6 0.03 mM [Ca2+]cyt, n = 5: 80 mM K+;0.25 6 0.01 mM [Ca2+]cyt, n = 4; P , 0.001). The effect ofpH was tested against the standard application of0.1 mM extracellular ADP, revealing a clear neutral pHoptimum (0.57 6 0.02 mM [Ca2+]cyt, n = 4) and a sig-nificantly enhanced response at alkaline pH compared
to physiological apoplastic pH 5 to 6 (Fig. 4B; P ,0.001). Nevertheless, at pH 5, elevation of [Ca2+]cyt re-mained significant (control [Ca2+]cyt baseline pH 5,0.116 0.02 mM; [Ca2+]cyt peak, 0.446 0.03 mM, n = 5; P,0.001). In separate tests (using a plate reader ratherthan a luminometer), although absolute values of peak[Ca2+]cyt were lower, the pattern of a greater responseat neutral pH rather than pH 6 was retained for 0.1 mM
ADP and also 0.1 mM ATP (Fig. 4C, n = 4; P , 0.05).Response toATPwas significantly greater than toADP atboth pH. The response to the poorly hydrolyzable ADPanalog ADPbS [adenosine 5#-O-(2-thiodiphosphate)]was not significantly different to the ADP responsewhile abATP evoked significantly lower responses thanATP at both pH (n = 4; P , 0.05). Thus part of the ATPresponse may involve hydrolysis products but the ADPresponse does not.
Extracellular ADP Does Not Increase Intracellular Levelsof ROS
In Arabidopsis roots, extracellular ATP causes ac-cumulation of intracellular ROS via activation of thePM NADPH oxidase AtRBOHC (Demidchik et al.,2009). At the whole-root level, extracellular ADP wasapplied under the same conditions used by Demidchiket al. (2009) for intracellular ROS imaging [using ester-loaded 5-(and-6-)-chloromethyl-2’,7’-dichlorohydro-fluorscein diacetate] in response to extracellular ATP.Solutions were designed to ensure that extracellular[Ca2+] remained constant at 0.1 mM throughout exper-imental treatments as purine nucleotides are potentCa2+ chelators. Chelation of extracellular Ca2+ can trig-ger extracellular superoxide anion production by roots(Mortimer et al., 2008). Here, extracellular ATP wasfound again to cause intracellular ROS accumulation(Fig. 5A). In contrast, extracellular ADP did not resultin significant accumulation of intracellular ROS (Fig. 5,A and B). Previously a response was seen by 15 s after
Figure 4. Transient [Ca2+]cyt elevation by extra-cellular ADP in epidermal protoplasts. A, Repre-sentative trace of [Ca2+]cyt response in matureepidermal protoplasts evoked by 0.1 mM ADP,recorded using a luminometer. Discharge denotesthe addition of 2 M CaCl2 and 20% (v/v) ethanolto enable [Ca2+]cyt estimation. B, pH dependenceof the epidermal protoplast [Ca2+]cyt response.Mean6 SE peak [Ca2+]cyt response to 0.1 mM ADPwith variable extracellular pH. Results are fromfour to five independent trials. C, pH dependenceof the epidermal protoplast mean6 SE peak [Ca2+]cytresponse in separate trials using a plate reader.The effects of 0.1 mM ADP, ATP, ADPbS, or abme-ATP were tested at pH 6 or 7 (n = 4).
Receptor-Like Activity Evoked by Extracellular ADP
Plant Physiol. Vol. 156, 2011 1379

extracellular ATP application (0.01–1 mM) with 0.1 mM
extracellular ATP causing over a 3-fold increase insignal intensity (Demidchik et al., 2009). Here, thesame concentration range for extracellular ADP underidentical conditions did not evoke intracellular ROSproduction, even over an extended observation pe-riod. Under control conditions, mean6 SE fluorescenceintensity of single primary roots was 19 6 0.9 fluores-cence per pixel (n = 23) and there were no significantincreases on application of extracellular ADP (Tukey’smultiple comparison test; Fig. 5B). Separate trialsusing a different imaging system (Leica) and withextracellular Ca2+ maintained at 0.1 or 1 mM resulted inhigher control values than recorded previously, show-ing the variable nature of ROS accumulation (Fig. 5, Cand D). Extracellular ATP (1 mM) caused an increase influorescence intensity with 1 mM Ca2+ but 1 mM ADPdid not (Fig. 5, C and D; n = 11). Fluorescence intensitywith ADP was significantly lower than with ATP(Student’s t test; P , 0.02). Overall the results suggestthat the extracellular ADP/[Ca2+]cyt pathway is inde-pendent of intracellular ROS.
Extracellular ADP Regulates Net Ca2+ and K+ Fluxes fromExcised Roots
To test whether extracellular ADP can activate cat-ion fluxes in intact cells, we monitored net fluxes at theepidermis of excised roots using the MIFE system for
noninvasive microelectrode Ca2+- and K+-flux mea-surements. This permits fine spatio-temporal mappingof net fluxes. Constancy of extracellular Ca2+ (0.1 mM
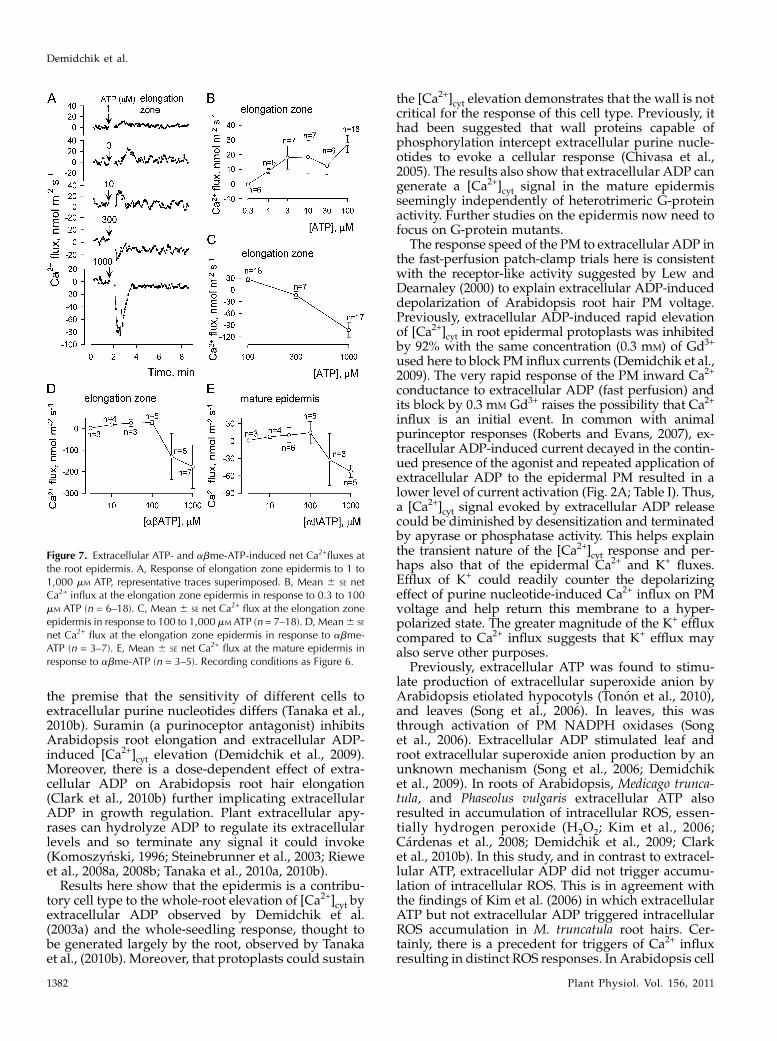
Ca2+) ensured that changes in flux were due to extra-cellular ADP perception alone (rather than changes in[Ca2+]) and also aided electrode function. Applicationof extracellular ADP (Na+ salt) to the elongation zoneepidermis caused transient increases in net Ca2+ influx(i.e. more positive flux values) in a dose-dependentmanner (Fig. 6, A and B; n = 3–6 for 1 mM to 1 mM
extracellular ADP). A significant increase above basallevel was first detected at 3 mM (P , 0.02; Student’st test). Duration of transients was up to 4 min. Controlflux levels were recovered. In control experimentsfor the effect of Na+, 4 mM NaCl had no effect on netflux (n = 4).
Net Ca2+ influx in response to extracellular ADPwasless pronounced in mature zone epidermis and [ADP]below 10 mM did not evoked significant changes (Fig.6C, n = 3–7). The mature zone has been found previ-ously to be less receptive to exogenous ROS than theelongation zone (Demidchik et al., 2003b, 2009). Dif-ferences between elongation zone and mature zonewere clearer for extracellular ADP-induced K+ efflux(Fig. 6D), with elongation zone epidermis (Fig. 6E, n =3–6) supporting greater net K+ efflux than the maturezone (Fig. 6F, n = 3–7) across the concentration rangetested. Significant changes in K+ flux relative to basalwere observed at 1 mM ADP in both zones. Extracel-
Figure 5. Extracellular ADP does not cause intracellular ROS accumulation. Arabidopsis roots were ester loaded with 5-(and-6)-chloromethyl-2#,7#-dichlorodihydrofluorescein (CM-H2DCF) and exposed to extracellular ADP or ATP. A, Top sections, rootsbathed in medium (0.1 mM KCl, 0.1 mM CaCl2, pH 6.0) with 0, 0.1 mM, 1 mM extracellular ADP, or 0.1 mM extracellular ATP withfluorescence false color mapping applied using the same scaling used for the positive extracellular ATP response reported byDemidchik et al. (2009). Bottom sections, corresponding bright-field images. Scale bar = 2 mm. B, Mean 6 SE fluorescence perpixel for control (n = 23), 0.01mM extracellular ADP (n = 24), 0.1 mM extracellular ADP (n = 25), and 1mM extracellular ADP (n =23) exposure. C, In separate trials, roots were exposed to 1 mM ADP or ATP but with Ca2+ maintained at 1 mM. Scale bar = 0.2mm, fluorescence false color-mapping scale is shown on the right. D, Mean 6 SE fluorescence per pixel for 0.1 mM Ca2+ buffer(n = 8), 1 mM Ca2+ buffer (n = 17), 1 mM ADP with 1 mM Ca2+ (n = 11), 1 mM ATP with 1 mM Ca2+ (n = 11).
Demidchik et al.
1380 Plant Physiol. Vol. 156, 2011

lular AMP was found previously to be a very weakinducer of root Ca2+ influx (Demidchik et al., 2009) andseedling [Ca2+]cyt elevation (Tanaka et al., 2010b). Here,AMP did not elicit a K+ efflux response even at 1 mM,in either the elongation zone or mature epidermis(elongation zone control; 237 6 34 nmol m22 s21, n = 3:elongation zone + AMP, 231 6 26 nmol m22 s21, n = 3;mature zone control;2196 13 nmolm22 s21, n = 3: AMP,210 6 18 nmol m22 s21, n = 3). This suggests thatbreakdown of extracellular ADP to AMP would helpterminate the cell’s response.Extracellular ATP and its poorly hydrolyzable ana-
log adenosine 5#-(a,b-methylene)triphosphate (abATP)were also tested. The pattern of greater net Ca2+ and K+
fluxes at the elongation zone was retained for bothagonists (Figs. 7 and 8). In the elongation zone, bothextracellular ATP and abATP up to 100 mM causedtransient net Ca2+ influx comparable to that evoked byextracellular ADP (Fig. 7, A–D). The threshold of thedetectable ATP response lay between 0.3 and 1 mM (n =6). However while at 300 mM and 1 mM extracellularADP promoted net Ca2+ influx, ATP and abATP causednet Ca2+ efflux (n = 5–17; Fig. 7, C and D). The meanpeak net Ca2+ efflux caused by 1 mM abATPwas almostdouble that caused by 1 mM ATP (Fig. 7, C and D). Atthe mature epidermis, while 1 mM ADP caused net Ca2+
influx, both 1 mM ATP and abATP again caused netCa2+ efflux with abATP eliciting the greater response(Fig. 7E; 1 mM ATP,256 21 nmol m22 s21, n = 8). ThusATP hydrolysis appears necessary to limit the Ca2+
efflux response. At both elongation zone and matureepidermis, both extracellular ATP and abATP causeddose-dependent net K+ efflux in common with ADP(Fig. 8). Thus while at high concentrations of ADPnet Ca2+ influx and K+ efflux appear as a coupledresponse, net Ca2+ flux is uncoupled from K+ efflux athigh concentrations of ATP or abATP.
DISCUSSION
Evidence is now accumulating that extracellularATP is a regulator of plant physiology and develop-ment (Lew and Dearnaley, 2000; Tang et al., 2003;Chivasa et al., 2005, 2009; Kim et al., 2006, 2009; Rouxand Steinebrunner, 2007; Wu et al., 2007; Riewe et al.,2008a, 2008b; Wu and Wu, 2008; Yi et al., 2008;Demidchik et al., 2009; Reichler et al., 2009; Clarket al., 2010a, 2010b; Tanaka et al., 2010a, 2010b; Terrileet al., 2010; Tonon et al., 2010). In common with ATP, itis likely that ADP is released on wounding and couldperhaps also be released with ATP by exocytosis or theactivity of ATP-binding cassette transporters (Thomaset al., 2000; Kim et al., 2006; Tanaka et al., 2010a).Extracellular ATP hotspots have been found in elon-gating root cells (Kim et al., 2006) and ADP may alsobe released there. This would be consistent with thefinding here that the Arabidopsis root elongation zoneepidermis supported a greater flux response to extra-cellular ADP than the mature zone. Such data support
Figure 6. Extracellular ADP-induced elevation of net Ca2+ influx and K+ efflux at the root epidermis. A, Response of elongationzone epidermal net Ca2+ flux to 3 mM, 100 mM, or 1 mM ADP, representative traces superimposed. Positive values signify influx. B,Mean6 SE net Ca2+ influx at the elongation zone epidermis (n = 3–6). C, Mean6 SE net Ca2+ influx at the mature epidermis (n =3–7). D, Response of mature and elongation zone epidermis to 300mM ADP, representative traces superimposed. Negative valuessignify efflux. E, Mean 6 SE net K+ efflux at the elongation zone epidermis (n = 3–6). F, Mean 6 SE net K+ efflux at the matureepidermis (n = 3–7). Basal assay solution comprised 0.1 mM KCl, 0.1 mM CaCl2, pH 6.0.
Receptor-Like Activity Evoked by Extracellular ADP
Plant Physiol. Vol. 156, 2011 1381
the premise that the sensitivity of different cells toextracellular purine nucleotides differs (Tanaka et al.,2010b). Suramin (a purinoceptor antagonist) inhibitsArabidopsis root elongation and extracellular ADP-induced [Ca2+]cyt elevation (Demidchik et al., 2009).Moreover, there is a dose-dependent effect of extra-cellular ADP on Arabidopsis root hair elongation(Clark et al., 2010b) further implicating extracellularADP in growth regulation. Plant extracellular apy-rases can hydrolyze ADP to regulate its extracellularlevels and so terminate any signal it could invoke(Komoszynski, 1996; Steinebrunner et al., 2003; Rieweet al., 2008a, 2008b; Tanaka et al., 2010a, 2010b).
Results here show that the epidermis is a contribu-tory cell type to the whole-root elevation of [Ca2+]cyt byextracellular ADP observed by Demidchik et al.(2003a) and the whole-seedling response, thought tobe generated largely by the root, observed by Tanakaet al., (2010b). Moreover, that protoplasts could sustain
the [Ca2+]cyt elevation demonstrates that the wall is notcritical for the response of this cell type. Previously, ithad been suggested that wall proteins capable ofphosphorylation intercept extracellular purine nucle-otides to evoke a cellular response (Chivasa et al.,2005). The results also show that extracellular ADP cangenerate a [Ca2+]cyt signal in the mature epidermisseemingly independently of heterotrimeric G-proteinactivity. Further studies on the epidermis now need tofocus on G-protein mutants.
The response speed of the PM to extracellular ADP inthe fast-perfusion patch-clamp trials here is consistentwith the receptor-like activity suggested by Lew andDearnaley (2000) to explain extracellular ADP-induceddepolarization of Arabidopsis root hair PM voltage.Previously, extracellular ADP-induced rapid elevationof [Ca2+]cyt in root epidermal protoplasts was inhibitedby 92% with the same concentration (0.3 mM) of Gd3+
used here to block PM influx currents (Demidchik et al.,2009). The very rapid response of the PM inward Ca2+
conductance to extracellular ADP (fast perfusion) andits block by 0.3 mM Gd3+ raises the possibility that Ca2+
influx is an initial event. In common with animalpurinceptor responses (Roberts and Evans, 2007), ex-tracellular ADP-induced current decayed in the contin-ued presence of the agonist and repeated application ofextracellular ADP to the epidermal PM resulted in alower level of current activation (Fig. 2A; Table I). Thus,a [Ca2+]cyt signal evoked by extracellular ADP releasecould be diminished by desensitization and terminatedby apyrase or phosphatase activity. This helps explainthe transient nature of the [Ca2+]cyt response and per-haps also that of the epidermal Ca2+ and K+ fluxes.Efflux of K+ could readily counter the depolarizingeffect of purine nucleotide-induced Ca2+ influx on PMvoltage and help return this membrane to a hyper-polarized state. The greater magnitude of the K+ effluxcompared to Ca2+ influx suggests that K+ efflux mayalso serve other purposes.
Previously, extracellular ATP was found to stimu-late production of extracellular superoxide anion byArabidopsis etiolated hypocotyls (Tonon et al., 2010),and leaves (Song et al., 2006). In leaves, this wasthrough activation of PM NADPH oxidases (Songet al., 2006). Extracellular ADP stimulated leaf androot extracellular superoxide anion production by anunknown mechanism (Song et al., 2006; Demidchiket al., 2009). In roots of Arabidopsis, Medicago trunca-tula, and Phaseolus vulgaris extracellular ATP alsoresulted in accumulation of intracellular ROS, essen-tially hydrogen peroxide (H2O2; Kim et al., 2006;Cardenas et al., 2008; Demidchik et al., 2009; Clarket al., 2010b). In this study, and in contrast to extracel-lular ATP, extracellular ADP did not trigger accumu-lation of intracellular ROS. This is in agreement withthe findings of Kim et al. (2006) in which extracellularATP but not extracellular ADP triggered intracellularROS accumulation in M. truncatula root hairs. Cer-tainly, there is a precedent for triggers of Ca2+ influxresulting in distinct ROS responses. In Arabidopsis cell
Figure 7. Extracellular ATP- and abme-ATP-induced net Ca2+fluxes atthe root epidermis. A, Response of elongation zone epidermis to 1 to1,000 mM ATP, representative traces superimposed. B, Mean 6 SE netCa2+ influx at the elongation zone epidermis in response to 0.3 to 100mM ATP (n = 6–18). C, Mean 6 SE net Ca2+ flux at the elongation zoneepidermis in response to 100 to 1,000 mM ATP (n = 7–18). D, Mean6 SE
net Ca2+ flux at the elongation zone epidermis in response to abme-ATP (n = 3–7). E, Mean 6 SE net Ca2+ flux at the mature epidermis inresponse to abme-ATP (n = 3–5). Recording conditions as Figure 6.
Demidchik et al.
1382 Plant Physiol. Vol. 156, 2011

suspension cells, both hyper- and hypoosmotic stresstrigger Ca2+ influx but only hypoosmotic stress resultsin extracellular H2O2 accumulation (Beffagna et al.,2005). Extracellular superoxide anions (resulting fromperception of extracellular purine nucleotides) couldgenerate H2O2 that could then be translocated tothe cytosol by PM aquaporins (Bienert et al., 2007;Dynowski et al., 2008). One possible explanation forthe disparity between the intracellular H2O2 accumu-lation response to extracellular ATP and extracellularADP is that in the extracellular ADP pathway, H2O2translocation is blocked. This could be achieved byclosure and/or retrieval of H2O2-permeable aquapor-ins (Boursiac et al., 2008). Alternatively, extracellularADP perception could inhibit an intracellular ROS-generating system, stimulate an intracellular antioxi-dant activity, or extracellular ADP perception in theapoplast could result in suppression of extracellularROS production downstream of superoxide anions.Given the importance of ROS in plant cell signaling,these possibilities merit further investigation particu-larly in the context of nodule development. Extracel-lular ADP has been found to regulate this process thatis known to involve complex spatio-temporal changesin ROS (Chang et al., 2009; Govindarajulu et al., 2009).Whether ATP-dependent intracellular ROS accumu-lation is the cause of net Ca2+ efflux from the rootepidermis at high [ATP] now needs to be determinedas this may be relevant to the suppression of defenseresponses noted when exogenous ATP is increased(Chivasa et al., 2009).
The pH dependence of animal ionotropic purinore-ceptors varies with isoform (King et al., 1997; Wildmanet al., 1999; Wirkner et al., 2008). Here, the extracellularADP-induced [Ca2+]cyt response was operative over awide pH range but activity at more neutral to alkalinepH suggests that extracellular ADP (and ATP) sig-naling could operate during stress responses in whichthe cytosol acidifies and apoplastic pH elevates aboveits normal acid value, such as hypoosmotic stress orelicitor challenge (Mathieu et al., 1996; Pugin et al.,1997; Beffagna et al., 2005). Additionally, pH de-pendence of ADP- or ATP-induced Ca2+ and K+ fluxcould be relevant at growth points, such as the root hairapex, at which apoplastic pH oscillates and pH oscil-lations are linked to nitrogen availability (Monshausenet al., 2007; Bloch et al., 2011). Overall, extracellular ADPis competent to act as a trigger of [Ca2+]cyt-dependentsignaling with the potential to evoke responses dis-tinct to those produced by ATP. Its initial perceptionat the PM appears reliant on ionotropic receptor-likeactivity.
CONCLUSION
Perception of extracellular ADP at the root epider-mal PM is consistent with the operation of functionalequivalents of ionotropic purine nucleotide receptors.In contrast to extracellular ATP, downstream events donot include accumulation of intracellular ROS thusindicating disparate signaling pathways.
Figure 8. Extracellular ATP- and abme-ATP-induced net K+ fluxes at the root epidermis. A, Response of elongation zoneepidermis to 0.3 to 1,000 mM ATP, representative traces superimposed. B, Mean 6 SE net K+ efflux at the elongation zoneepidermis in response to 0.3 to 1,000 mM ATP (n = 6–37). C, Mean6 SE net K+ efflux at the mature epidermis in response to 10 to1,000 mM ATP (n = 4–13). D, Response of elongation zone and mature epidermis to 300 mM abme-ATP, representative tracessuperimposed. E, Mean6 SE net K+ efflux at the elongation zone in response to 3 to 1,000 mM abme-ATP (n = 3–7). F, Mean6 SE
net K+ efflux at the mature epidermis in response to 3 to 1,000 mM abme-ATP (n = 3–6). Recording conditions as Figure 6.
Receptor-Like Activity Evoked by Extracellular ADP
Plant Physiol. Vol. 156, 2011 1383

MATERIALS AND METHODS
Plant Growth
Arabidopsis (Arabidopsis thaliana Columiba-0; Columbia-0 expressing
apoaequorin under a 35s promoter; Demidchik et al., 2009) from laboratory
stock was grown vertically on full-strength Murashige and Skoog (Duchefa;
www.duchefa.com) medium with 1% (w/v) Suc and 0.3% (w/v) Phytagel
(Sigma; www.sigmaaldrich.com). Growth conditions were 22�C and 16 h day
length, 100 mmol m22 s21 irradiance.
Patch-Clamp Electrophysiology
Protoplasts were isolated from root mature epidermis (10–15 d plants)
using the protocol described by Demidchik et al. (2003b). Protoplasts used for
patch clamping were 20 6 1.5 mm in diameter. The PS contained (mM): 40
K-gluconate, 10 KCl, 1 1,2-bis(2-aminophenoxy)ethane-N,N,N’,N’-tetraacetic
acid (free Ca2+ adjusted to 100 nM with 0.4 CaCl2), pH 7.2 adjusted with Tris/
MES. The G-protein antagonist GDPbS (0.5 mM) was incorporated into the PS
as indicated. Control BS comprised (mM): 20 CaCl2, 0.1 KCl, 0.3 NaCl (varied
to supply equimolar Na+ to that in purine-containing BS), pH 6 with 2 Tris/4
MES. BS containing purine agonist was freshly prepared directly before
application and the pH of the solution was adjusted where necessary (Tris). BS
and PS were adjusted to 290 to 300 mOsM with D-sorbitol. Delivery of BS +
ADP to the recording chamber was either by gravity feed or by a fast-flow
system comprising an AutoMate perfusion system with four pinch valves,
ValveBank controller, and four-barrel delivery pipette (barrel diameter 0.1
mm; AutoMate Scientific Inc.). This system allowed switching to test BS
in 0.01 s. Average delivery time was less than 1 s. The delivery pipette was
placed at 30 to 40 mm from the protoplast surface and delivery speed was 0.5
to 1 mL s21. Two barrels of the delivery pipette were used—one for control BS
and another for purine-containing BS. Solution changes were controlled by the
programmable ValveBank controller. Recording equipment was as described
in Demidchik et al. (2003b). Ion activities were calculated using Geochem
(Parker et al., 1995). Liquid junction potentials were measured and corrected
as described previously (Demidchik et al., 2003b).
[Ca2+]cyt Measurement
The protocol described in Demidchik et al. (2007) was used to isolate
protoplasts from root mature epidermis of Arabidopsis (10–15 d old) express-
ing cytosolic (apo)aequorin (cauliflower mosaic virus 35S promoter) with the
modification that incubation with protoplasting enzymes was increased from
50 to 60 min. Protoplasts were suspended for 3 to 4 h (dark, 28�C) in recording
medium (mM: 1 CaCl2, 1 KCl, 1 MgCl2, 10 Glc; pH 6.0 adjusted by 2 Tris/4
MES; 290–300 mOsM with D-sorbitol) with 10 mM coelenterazine (NanoLight
Technology; www.nanolight.com) to match the protocol used by Demidchik
et al. (2009) for ATP and ADP studies. After washing, recording medium
(without coelentrazine) and standard luminometry procedures were used.
The discharging solution enabling estimation of [Ca2+]cyt contained 2 M CaCl2and 20% (w/v) ethanol. Luminescence was recorded using a luminometer or
FLUOstar plate reader.
Imaging of ROS
Imaging of intracellular ROS used 50 mM 5-(and-6-)-chloromethyl-2’,7’-
dichlorohydrofluorscein diacetate, acetyl ester (Molecular Probes; www.
invitrogen.com) with dye loading as described previously (Demidchik et al.,
2009). Assay medium was as described by Demidchik et al. (2009) and
comprised (mM): 0.1 KCl, 0.1 or 1 CaCl2, 4 MES, and 2 Tris base (pH 6.0).
Images of 5-d-old roots were acquired either with a Nikon SMZ 1500
microscope and a QImaging Retiga cooled 12-bit camera or a Leica M165
FC microscope and a Leica DFC310 FX 1.4-megapixel camera. Ca2+ was
maintained at 0.1 or 1 mM in the presence of purines by appropriate additions
of CaCl2 (calculated using MaxChelator and Geochem programs; Parker et al.,
1995; Patton et al., 2004).
MIFE Recordings of Net Cation Fluxes
Excised roots (5–15 d old) were fixed into a recording chamber by an agar
drop and immersed in 0.5 mL control solution comprising (in mM) 0.1 KCl, 0.1
CaCl2, 4 MES, 2 Tris base (pH 6.0; Demidchik et al., 2009). Chamber design
permitted rapid mixing of test solution and resumption of flux recording 20 to
30 s after addition of 0.5 mL test solution. Ca2+ was maintained at 0.1 mM in the
presence of purines by appropriate additions of CaCl2. Levels were confirmed
experimentally in a cell-free assay using a Ca2+-selective microelectrode.
Purines were added as Tris, Na+, or Li+ salts. NaCl or LiCl at equimolar
concentrations did not perturb K+ efflux (tested experimentally). Measure-
ments of mature epidermal flux were taken 1 to 1.5 mm from the root apex and
for the elongation zone, 100 to 150 mm from the apex (Demidchik et al., 2007).
Net Ca2+ influx and K+ efflux were determined using the noninvasive slowly
vibrating microelectrode (MIFE) technique. K+- and Ca2+-selective microelec-
trodes (external tip diameter approximately 3 mm) were made and calibrated
as described previously (Shabala and Shabala, 2002). Electrodes were cali-
brated before and after use. Agonists (tested up to 1 mM) had no effect on
calibration or electrode response time. The microelectrode was placed 20 mm
above the root surface and moved between two positions, 20 and 50 mm above
the root surface in a square wave manner (5-s half-cycle).
ACKNOWLEDGMENTS
We thank Daniel Schachtmann for advice and Adeeba Dark for help with
plant cultivation.
Received February 18, 2011; accepted May 6, 2011; published May 11, 2011.
LITERATURE CITED
Alloisio S, Cugnoli C, Ferroni S, Nobile M (2004) Differential modulation
of ATP-induced calcium signalling by A1 and A2 adenosine receptors in
cultured cortical astrocytes. Br J Pharmacol 141: 935–942
Beffagna N, Buffoli B, Busi C (2005) Modulation of reactive oxygen species
production during osmotic stress in Arabidopsis thaliana cultured cells:
involvement of the plasma membrane Ca2+-ATPase and H+-ATPase.
Plant Cell Physiol 46: 1326–1339
Bienert GP, Møller ALB, Kristiansen KA, Schulz A, Møller IM,
Schjoerring JK, Jahn TP (2007) Specific aquaporins facilitate the diffusion
of hydrogen peroxide across membranes. J Biol Chem 282: 1183–1192
Bloch D, Monshausen G, Singer M, Gilroy S, Yalovsky S (2011) Nitrogen
source interacts with ROP signalling in root hair tip-growth. Plant Cell
Environ 34: 76–88
Boursiac Y, Boudet J, Postaire O, Luu D-T, Tournaire-Roux C, Maurel C
(2008) Stimulus-induced downregulation of root water transport in-
volves reactive oxygen species-activated cell signalling and plasma
membrane intrinsic protein internalization. Plant J 56: 207–218
Cardenas L, Martınez A, Sanchez F, Quinto C (2008) Fast, transient and
specific intracellular ROS changes in living root hair cells responding to
Nod factors (NFs). Plant J 56: 802–813
Chang C, Damiani I, Puppo A, Frendo P (2009) Redox changes during the
legume-rhizobium symbiosis. Mol Plant 2: 370–377
Chivasa S, Murphy AM, Hamilton JM, Lindsey K, Carr JP, Slabas AR
(2009) Extracellular ATP is a regulator of pathogen defence in plants.
Plant J 60: 436–448
Chivasa S, Ndimba BK, Simon WJ, Lindsey K, Slabas AR (2005) Extra-
cellular ATP functions as an endogenous external metabolite regulating
plant cell viability. Plant Cell 17: 3019–3034
Clark G, Torres J, Finlayson S, Guan X, Handley C, Lee J, Kays JE, Chen
ZJ, Roux SJ (2010a) Apyrase (nucleoside triphosphate-diphosphohy-
drolase) and extracellular nucleotides regulate cotton fiber elongation in
cultured ovules. Plant Physiol 152: 1073–1083
Clark G, Wu M, Wat N, Onyirimba J, Pham T, Herz N, Ogoti J, Gomez D,
Canales AA, Aranda G, et al (2010b) Both the stimulation and inhibition
of root hair growth induced by extracellular nucleotides in Arabidopsis
are mediated by nitric oxide and reactive oxygen species. Plant Mol Biol
74: 423–435
Demidchik V, Bowen HC, Maathuis FJ, Shabala SN, Tester MA, White PJ,
Davies JM (2002) Arabidopsis thaliana root non-selective cation channels
mediate calcium uptake and are involved in growth. Plant J 32: 799–808
Demidchik V, Nichols C, Oliynyk M, Dark A, Glover BJ, Davies JM
(2003a) Is ATP a signaling agent in plants? Plant Physiol 133: 456–461
Demidchik V, Shabala SN, Coutts KB, Tester MA, Davies JM (2003b) Free
Demidchik et al.
1384 Plant Physiol. Vol. 156, 2011

oxygen radicals regulate plasma membrane Ca2+- and K+-permeable
channels in plant root cells. J Cell Sci 116: 81–88
Demidchik V, Shabala SN, Davies JM (2007) Spatial variation in H2O2
response of Arabidopsis thaliana root epidermal Ca2+ flux and plasma
membrane Ca2+ channels. Plant J 49: 377–386
Demidchik V, Shang Z, Shin R, Thompson E, Rubio L, Laohavisit A,
Mortimer JC, Chivasa S, Slabas AR, Glover BJ, et al (2009) Plant
extracellular ATP signalling by plasma membrane NADPH oxidase and
Ca2+ channels. Plant J 58: 903–913
Dynowski M, Schaaf G, Loque D, Moran O, Ludewig U (2008) Plant
plasma membrane water channels conduct the signalling molecule
H2O2. Biochem J 414: 53–61
Foresi NP, Laxalt AM, Tonon CV, Casalongue CA, Lamattina L (2007)
Extracellular ATP induces nitric oxide production in tomato cell sus-
pensions. Plant Physiol 145: 589–592
Fountain SJ, Cao L, YoungMT, North RA (2008) Permeation properties of a
P2X receptor in the green algae Ostreococcus tauri. J Biol Chem 283:
15122–15126
Govindarajulu M, Kim SY, Libault M, Berg RH, Tanaka K, Stacey G,
Taylor CG (2009) GS52 ecto-apyrase plays a critical role during soybean
nodulation. Plant Physiol 149: 994–1004
Jeter CR, Tang W, Henaff E, Butterfield T, Roux SJ (2004) Evidence of a
novel cell signaling role for extracellular adenosine triphosphates and
diphosphates in Arabidopsis. Plant Cell 16: 2652–2664
Khakh BS, North RA (2006) P2X receptors as cell-surface ATP sensors in
health and disease. Nature 442: 527–532
Kim S-H, Yang SH, Kim T-J, Han J-S, Suh J-W (2009) Hypertonic stress
increased extracellular ATP levels and the expression of stress-responsive
genes in Arabidopsis thaliana seedlings. Biosci Biotechnol Biochem 73:
1252–1256
Kim SY, Sivaguru M, Stacey G (2006) Extracellular ATP in plants: visu-
alization, localization, and analysis of physiological significance in
growth and signaling. Plant Physiol 142: 984–992
King BF, Wildman SS, Ziganshina LE, Pintor J, Burnstock G (1997) Effects
of extracellular pH on agonism and antagonism at a recombinant P2X2
receptor. Br J Pharmacol 121: 1445–1453
Komoszynski MA (1996) Comparative studies on animal and plant apy-
rases (ATP diphosphohydrolase EC 3.6.1.5) with application of immu-
nological techniques and various ATPase inhibitors. Comp Biochem
Physiol B Biochem Mol Biol 113: 581–591
Lew RR, Dearnaley JDW (2000) Extracellular nucleotide effects on electrical
properties of growing Arabidopsis thaliana root hairs. Plant Sci 153: 1–6
Li WW, Assmann SM (1993) Characterization of a G-protein-regulated
outward K+ current in mesophyll cells of Vicia faba L. Proc Natl Acad Sci
USA 90: 262–266
Liang CY, Tian J, Lam HM, Lim BL, Yan XL, Liao H (2010) Biochemical and
molecular characterization of PvPAP3, a novel purple acid phosphatase
isolated from common bean enhancing extracellular ATP utilization.
Plant Physiol 152: 854–865
Maathuis FJM, Sanders D (1993) Energisation of potassium uptake in
Arabidopsis thaliana. Planta 191: 302–307
Mathieu Y, Lapous D, Thomine S, Lauriere C, Guern J (1996) Cytoplasmic
acidification as an early phosphorylation-dependent response of to-
bacco cells to elicitors. Planta 199: 416–424
Monshausen GB, Bibikova TN, Messerli MA, Shi C, Gilroy S (2007)
Oscillations in extracellular pH and reactive oxygen species modulate tip
growth of Arabidopsis root hairs. Proc Natl Acad Sci USA 104: 20996–21001
Mortimer JC, Laohavisit A, Miedema H, Davies JM (2008) Voltage, reactive
oxygen species and the influx of calcium. Plant Sig Behav 3: 698–699
Parker DR, Norvell WA, Chaney RL (1995) GEOCHEM-PC: a chemical speci-
ation program for IBM and compatible computers. In RH Loeppert, AP
Schwab, S Goldberg, eds, Chemical Equilibrium andReactionModels, Special
Publication 42. Soil Science Society of America, Madison, WI, pp 253–269
Patton C, Thompson S, Epel D (2004) Some precautions in using chelators
to buffer metals in biological solutions. Cell Calcium 35: 427–431
Pugin A, Frachisse JM, Tavernier E, Bligny R, Gout E, Douce R, Guern J
(1997) Early events induced by elicitor cryptogein in tobacco cells:
involvement of a plasma membrane NADPH oxidase and activation of
glycolysis and the pentose phosphate pathway. Plant Cell 9: 2077–2091
Reichler SA, Torres J, Rivera AL, Cintolesi VA, Clark G, Roux SJ (2009)
Intersection of two signalling pathways: extracellular nucleotides reg-
ulate pollen germination and pollen tube growth via nitric oxide. J Exp
Bot 60: 2129–2138
Riewe D, Grosman L, Fernie AR, Wucke C, Geigenberger P (2008a)
The potato-specific apyrase is apoplastically localized and has influ-
ence on gene expression, growth and development. Plant Physiol 146:
1579–1598
Riewe D, Grosman L, Fernie AR, Zauber H, Wucke C, Geigenberger P
(2008b) A cell wall-bound adenosine nucleosidase is involved in the
salvage of extracellular ATP in Solanum tuberosum. Plant Cell Physiol 49:
1572–1579
Roberts JA, Evans RJ (2007) Cysteine substitution mutants give structural
insight and identify ATP binding and activation sites at P2X receptors.
J Neurosci 27: 4072–4082
Roux SJ, Steinebrunner I (2007) Extracellular ATP: an unexpected role as a
signaler in plants. Trends Plant Sci 12: 522–527
Shabala S, Demidchik V, Shabala L, Cuin TA, Smith SJ, Miller AJ, Davies
JM, Newman IA (2006) Extracellular Ca2+ ameliorates NaCl-induced K+
loss from Arabidopsis root and leaf cells by controlling plasma mem-
brane K+-permeable channels. Plant Physiol 141: 1653–1665
Shabala S, Shabala L (2002) Kinetics of net H+, Ca2+, K+, Na+, NH4+, and Cl-
fluxes associated with post-chilling recovery of plasma membrane
transporters in Zea mays leaf and root tissues. Physiol Plant 114: 47–56
Shang Z, Laohavisit A, Davies JM (2009) Extracellular ATP activates an
Arabidopsis plasma membrane Ca2+-permeable conductance. Plant
Signal Behav 4: 989–991
Song CJ, Steinebrunner I, Wang X, Stout SC, Roux SJ (2006) Extracellular
ATP induces the accumulation of superoxide via NADPH oxidases in
Arabidopsis. Plant Physiol 140: 1222–1232
Steinebrunner I, Wu J, Sun Y, Corbett A, Roux SJ (2003) Disruption of
apyrases inhibits pollen germination in Arabidopsis. Plant Physiol 131:
1638–1647
Sueldo DJ, Foresi NP, Casalongue CA, Lamattina L, Laxalt AM (2010)
Phosphatidic acid formation is required for extracellular ATP-mediated
nitric oxide production in suspension-cultured tomato cells. New Phytol
185: 909–916
Tanaka K, Gilroy S, Jones AM, Stacey G (2010a) Extracellular ATP
signaling in plants. Trends Cell Biol 20: 601–608
Tanaka K, Swanson SJ, Gilroy S, Stacey G (2010b) Extracellular nucleo-
tides elicit cytosolic free calcium oscillations in Arabidopsis. Plant
Physiol 154: 705–719
Tang W, Brady SR, Sun Y, Muday GK, Roux SJ (2003) Extracellular ATP
inhibits root gravitropism at concentrations that inhibit polar auxin
transport. Plant Physiol 131: 147–154
Terrile MC, Tonon C, Iglesias MJ, Lamattina L, Casalongue CA (2010)
Extracellular ATP and nitric oxide signaling pathways regulate redox-
dependent responses associated to root hair growth in etiolated Arabi-
dopsis seedlings. Plant Sig Behav 5: 698–701
Thomas C, Rajagopal A, Windsor B, Dudler R, Lloyd A, Roux SJ (2000) A
role for ectophosphatase in xenobiotic resistance. Plant Cell 12: 519–533
Tonon C, Cecilia Terrile M, Jose Iglesias M, Lamattina L, Casalongue CA
(2010) Extracellular ATP, nitric oxide and superoxide act coordinately to
regulate hypocotyl growth in etiolated Arabidopsis seedlings. J Plant
Physiol 167: 540–546
Weerasinghe RR, Swanson SJ, Okada SF, Garrett MB, Kim SY, Stacey G,
Boucher RC, Gilroy S, Jones AM (2009) Touch induces ATP release in
Arabidopsis roots that is modulated by the heterotrimeric G-protein
complex. FEBS Lett 583: 2521–2526
Wildman SS, King BF, Burnstock G (1999) Modulation of ATP-responses
at recombinant rP2X4 receptors by extracellular pH and zinc. Br J
Pharmacol 126: 762–768
Wirkner K, Stanchev D, Milius D, Hartmann L, Kato E, Zadori ZS, Mager
PP, Rubini P, Norenberg W, Illes P (2008) Regulation of the pH
sensitivity of human P2X receptors by N-linked glycosylation. J Neuro-
chem 107: 1216–1224
Wu J, Steinebrunner I, Sun Y, Butterfield T, Torres J, Arnold D, Gonzalez
A, Jacob F, Reichler S, Roux SJ (2007) Apyrases (nucleoside triphos-
phate-diphosphohydrolases) play a key role in growth control in
Arabidopsis. Plant Physiol 144: 961–975
Wu SJ, Wu JY (2008) Extracellular ATP-induced NO production and its
dependence on membrane Ca2+ flux in Salvia miltiorrhiza hairy roots.
J Exp Bot 59: 4007–4016
Yi C, Qu HX, Jiang YM, Shi J, Duan XW, Joyce DC, Li YB (2008) ATP
induces changes in energy status and membrane integrity of harvested
litchi fruit and its relation to pathogen resistance. J Phytopathol 156:
365–371
Receptor-Like Activity Evoked by Extracellular ADP
Plant Physiol. Vol. 156, 2011 1385
Copyright © 2022 FDOKUMEN




















