
Caudate evoked synaptic activities in nigral neurons
17
EXPERIMENTAL NEUROLOGY 49, 123-139 (1975) Caudate Evoked Synaptic Activities in Nigral Neurons TAMAS L. FRIGYESI AND JANOS SZABO r Dcpartmcnt of Physiology, New York Medical College, Valhalla, New York Recrived March 21,1975; rcz~isiou received April 22,1975 Intracellular recordings from neurons in the substantia nigra have re- vealed three varieties of monosynaptic PSPs in response to stimulation of the ipsilateral head of the caudate nucleus : short (3-5 msec) latency EPSPs ; short (3-5 msec) latency IPSPs; and long (15-20 msec) latency IPSPs. The data indicate that at least three efferent fiber systems link the caudate to the nigra: two fast conducting axonal systems of comparable diameters mediate the short-latency PSPs whereas a slow conducting axonal system mediates the long-latency PSPs. The caudate evoked long-latency IPSPs in nigral neurons were preceded by antidromic and orthodromic potentials in the motor cortex; these cortical potentials are regarded as epiphenomena. The dual, facilitatory-inhibitory control of caudate on nigral neurons is con- sonant with the proposal that the caudate self-regulates its input from the nigra. The caudate-evoked EPSPs in nigral neurons are sine qua uoyt for the operation of the caudato-nigrothalamic projection system. INTRODUCTION The head of the caudate nucleus projects to both segmentsof the globus pallidus and the substantia nigra in the primate ( 11, 28, 34, 36, 56, 57)) and to the entopeduncular nucleus which is the feline homologue of the primate internal pallidal segment (12) ; the caudate does not project onto the motor cortex (49). Yet, it has been conclusively established that caudato-fugal effects elicit synaptic potentials in neurons in the ventro- lateral and medial thalamus (20) as well as in the motor cortex (39). Thus, the corollary question arises as to what are the trajectories of the caudate-dorsal thalamus (and hence the motor cortex) (8, 32, 55) pro- jection activities: inasmuch as the caudate does not project to the dorsal 1 This work was supported in part by grants from the National Institute of Neurological Diseases and Stroke, N.I.H., (NS-0989%02), the Eric Slack-Gyr Foundation, and the Medical Research Council of Canada. Dr. Szabo was a Visiting Scientist on leave from the Department of Anatomy, University of Ottawa. We gratefully acknowledge the many helpful discussions with Dr. Giinter Baumgartner and Dr. Manfred Klee during the course of these studies. 123 Copyright 0 1975 by AcademicPress, Inc. All rights of reproduction in any form reserved.
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Caudate evoked synaptic activities in nigral neurons

EXPERIMENTAL NEUROLOGY 49, 123-139 (1975)
Caudate Evoked Synaptic Activities in Nigral Neurons
TAMAS L. FRIGYESI AND JANOS SZABO r
Dcpartmcnt of Physiology, New York Medical College, Valhalla, New York Recrived March 21,1975; rcz~isiou received April 22,1975
Intracellular recordings from neurons in the substantia nigra have re- vealed three varieties of monosynaptic PSPs in response to stimulation of the ipsilateral head of the caudate nucleus : short (3-5 msec) latency EPSPs ; short (3-5 msec) latency IPSPs; and long (15-20 msec) latency IPSPs. The data indicate that at least three efferent fiber systems link the caudate to the nigra: two fast conducting axonal systems of comparable diameters mediate the short-latency PSPs whereas a slow conducting axonal system mediates the long-latency PSPs. The caudate evoked long-latency IPSPs in nigral neurons were preceded by antidromic and orthodromic potentials in the motor cortex; these cortical potentials are regarded as epiphenomena. The dual, facilitatory-inhibitory control of caudate on nigral neurons is con- sonant with the proposal that the caudate self-regulates its input from the nigra. The caudate-evoked EPSPs in nigral neurons are sine qua uoyt for the operation of the caudato-nigrothalamic projection system.
INTRODUCTION
The head of the caudate nucleus projects to both segments of the globus pallidus and the substantia nigra in the primate ( 11, 28, 34, 36, 56, 57)) and to the entopeduncular nucleus which is the feline homologue of the primate internal pallidal segment (12) ; the caudate does not project onto the motor cortex (49). Yet, it has been conclusively established that caudato-fugal effects elicit synaptic potentials in neurons in the ventro- lateral and medial thalamus (20) as well as in the motor cortex (39). Thus, the corollary question arises as to what are the trajectories of the caudate-dorsal thalamus (and hence the motor cortex) (8, 32, 55) pro- jection activities: inasmuch as the caudate does not project to the dorsal
1 This work was supported in part by grants from the National Institute of Neurological Diseases and Stroke, N.I.H., (NS-0989%02), the Eric Slack-Gyr Foundation, and the Medical Research Council of Canada. Dr. Szabo was a Visiting Scientist on leave from the Department of Anatomy, University of Ottawa. We gratefully acknowledge the many helpful discussions with Dr. Giinter Baumgartner and Dr. Manfred Klee during the course of these studies.
123
Copyright 0 1975 by Academic Press, Inc. All rights of reproduction in any form reserved.

124 FRIGYESI AND SZABO
thalamus either (49), caudato-fugal activities may reach the thalamus either by relaying through the pallidum or substantia nigra or both. Anatomical data show that the internal segment of globus pallidus (48, 49), as well as the nigra (7, 47), project onto the ventrolateral and medial thalamus. Electrophysiological studies have shown that stimula- tion of both globus pallidus and substantia nigra elicit monosynaptic excitatory postsynaptic potentials (EPSPs) in ventrolateral and medial thalamic neurons (13, 19, 20, 24). That is, in principle, both globus pallidus and substantia nigra could equally relay caudato-fugal activities to dorsal thalamic neurons via the ventral and dorsal divisions of ansa lenticularis and the nigrothalamic pathway (7, 18, 46). However, the concensus among electrophysiologists is that caudate generates inhibitory postsynaptic po- tentials (IPSPs) in neurons of the internal pallidal segment [dominantly
(23, 42) ; exclusively (60) 1. Furthermore, caudate-evoked synaptic po- tentials were demonstrated in the thalamus of squirrel monkeys whose pallidum was destroyed and in cats whose entopeduncular nucleus was destroyed by prior intranuclear administration of high doses of 6-hydroxy- dopamine ( 13, 26). These observations indicate that even if ansa lenticularis subserves transmitting functions for caudato-fugal effects to the thalamus, this projection system is not the primary one, (40, 45) but is an ancillary one for establishing caudate-thalamus functional relationships. In this formulation, ansa lenticularis plays a subsidiary role which directs atten- tion to the nigra as the paramount relay structure between the caudate and the thalamus (and hence to the motor cortex) (8, 14, 31, 39).
In previous studies, on unanesthetized cats, we reported that stimula- tion of the caudate elicited monsynaptic EPSPs in nigral neurons (22) ; and nigral stimulation, in turn, elicited monosynaptic EPSPs in ventro- lateral and medial thalamic neurons (20, 24). Based on these and above data, it was postulated that there is functional connectivity along the caudate-nigral and nigro-thalamic pathways; i.e. the operation of a caudato-nigro-thalamic projection system was proposed (18, 20). This hypothesis was corroborated by McKenzie and Bendrups (6) who re- ported that caudate and nigral stimulation had similar effects which ap- peared to be exerted on a similar pool of medial thalamic neurons.
Fundamentally different observations were made by Yoshida et al. (58, 60) in the barbiturate anesthetized cat. They observed that caudate stimulation elicited monosynaptic IPSPs, not only in pallidal but also in nigral neurons. These data cannot be reconciled with the demonstration of caudato-fugal excitatory effects reaching neurons in the dorsal thalamus unless the conclusiveness of the anatomical studies on the caudate efferents (vi& surpa) is questioned; alternatively, McNair et al. (44) proposed that barbiturates favor the appearance of inhibitory responses in the nigra following stimulation of the caudate. On the other hand, since the nigra

NIGRAL NEURONS 125
contains high concentrations of a putative inhibitory transmitter, gamma- aminobutiric acid (4), it was tempting to conjecture that this property of the nigra and the demonstration of caudate-evoked inhibitory potentials in nigral neurons provide mutual corroboration to one another (10, 33, 37). Powerful as this argument may be, it still leaves the question of the trajectory of caudate-thalamus (and caudate-motor cortex) excitatory effects unanswered. Therefore, it was deemed desirable to reinvestigate the intracellular potentials in nigral neurons during caudate stimulation. The data here seem to reconcile the apparently conflicting observations showing that both monosynaptic EPSPs and monosynaptic IPSPs are demonstrable in nigral neurons in response to caudate stimulation. Pre- liminary reports have already been published (15, 16).
MATERIALS AND METHODS
The experiments were performed on 30 volatile liquid (Methoxyfluran) anesthetized, succinylcholine paralyzed, encephale isole cats prepared under initial sodium thiamilal anesthesia in a manner similar to that described in intracellular studies of thalamic (19) and nigral (22) neurons. The gist of the procedure is that hemicraniectomy was performed which permitted partial decortication, callosectomy, and severance of the fornix by suc- tion-ablation ; these procedures produced direct access to the motor cortex and the dorsal surfaces of the head of the caudate nucleus and of the mesencephalon. Arrays of concentrically bipolar electrodes with inter- polar distances of 0.2-0.3 mm were introduced into the caudate under direct visual control. Similar arrays were positioned in the entopeduncular nucleus and internal capsule under stereotactic guidance. Brief (0.1-0.5 msec) rectangular pulses were delivered through an isolation transformer at frequencies of l-100 Hz. The experimental position of the stimulating electrodes were determined by the electrographic properties of evoked responses in the motor cortex (19). Glass micropipettes, which were filled with 2~ K+-citrate and had OS-O.2 pm tip diameters and lo-40 MO resistances, were lowered through the pretectum or superior colliculus into the substantia nigra. Intracellular potentials from nigral neurons were fed through a transistorized, neutralized input capacity amplifier and displayed on a DC channel of a 4 trace oscilloscope. Surface activities from the motor cortex (precruciate gyrus) were recorded by Ag-AgCl ball electrodes (0.5 mm in diameter) against a reference electrode embedded into the temporal muscle after amplification by an R.C. coupled amplifier with long time constant (0.2-l set) and displayed on a channel of the oscilloscope. Further details of the stimulating and recording techniques have been published (19). All stimulating and recording sites were verified histo- logically ( 19).

126 FRIGYESI AND SZABO
FIG. 1. Photomicrographs show electrode tracks in the caudate (A) and mesence- phalon (B). Unstained frozen sections. The dorsal surface of caudate and hippo- campus are exposed. (A) Two stimulating electrodes in the caudate; one pair in the lateral third (black arrow), another pair in the medial third (white arrow). (B)

KIGRAL NEURONS 127
RESULTS
Identification of Nigrtrl Ncztro~. Cell locations were identified by apply- ing measurements from the scale of microelectrode drive to histologically determined tracts of all recording micropipettes. All electrode tracts were found between anterior planes 3 and 5, and lateral planes 3 and 8. During the experiments, while the micropipettes were lowered into the mesen- cephalon, cells were readily encountered and successfully impaled in the tectum and upper half of the tegmentum (during the initial 7-8 mm pas- sage of the micropipette). Thereafter, while the micropipettes traversed the epinigral bundles of myelinated axons (medial lemniscus and brachium conjunctivum) (Fig. l), the passage was characterized by reiterative plugging of the tips of the micropipettes. Approximately 805% of the micro- pipettes were irreversibly plugged at this region, and only 20% reached the substantia nigra in good recording condition. The latter group of micro- pipettes, during further advance, once again exhibited characteristics of reiterative plugging of their tips (Fig. 1) ; this indicated that the micro- pipette had passed through the nigra and reached the myelinated axonal bundles of the cerebral peduncle. Under these conditions, intracellular recordings were obtained from 51 nigral neurons ; however, only 17 of these are included in this report because, in the remainder, either caudate stimu- lation was not employed or stable membrane potentials were not attained. The neurons included in this report exhibited spike potentials of 40-60 mV and resting potentials in excess of these values for periods of S-20 min. These attributes of the intracellular recordings have provided evidence that postsynaptic potentials observed were not significantly affected by the traumatic effects of impalements.
Eleven out of these 17 nigral neurons exhibited postsynaptic potentials in response to stimulation of the ipsilateral head of the caudate nucleus.
Evoked En-c&tory Spapfic Activities. Low-frequency (8 Hz) stimu- lation of the rostromed,iul third of the head of the caudate (Fig. 1) elicited EPSPs in two out of the 17 neurons in the substantia nigra. The evoked EPSPs showed 33 msec latencies and 15-25 msec durations (Fig. 2). In each cell, the evoked EPSPs had unvarying latencies and durations irre- spective of whether or not the EPSP triggered a spike discharge (Fig. 2C). Single discharges were superimposed on the summit of the evoked EPSPs at invariant latencies ; repetitive firing was not observed (Fig. 2A). Evoked EPSPs exhibited variations in magnitude ; those which were of low amplitude (< 8 mV) failed to trigger spike discharges (Fig. 2B). Significant differences were noticeable on the EPSPs associated with the
The long, tapered shaft of a glass micropipette is seen in the tegmentum; open arrow head points to the epinigral myelinated axonal systems. The track penetrates the nigra and enters the cerebral peduncle (black arrow).

128 FRIGYESI AND SZABO
MC
A
S>/,
.Cd
I /
50
/Lb- mV
. . 20 msec,
B -- A
C-,223/,6
L/j& h
. %d 10 msec
FIG. 2. Caudate evoked EPSPs in a neuron in the substantia nigra. In this and all subsequent figures-upper channel : monopolarly recorded surface responses from the motor cortex, upward deflections signal negativities ; lower channel : intracellular recordings from nigral neurons; the closed triangles point at the shock artifacts of 8 Hz stimulation of the caudate. (A) and (B) Continuous recordings. Rostro- medial caudate stimulation evokes 3-4 msec latency EPSPs in the nigral neuron. Single spike discharges are superimposed on the EPSPs elicited by the first two stimuli of the iterative stimulation of the caudab. Caudate evoked EPSPs uncom- plicated by spike discharges are seen at end of (A) and in (B). (C) Superimposi- tion of two EPSPs show that the time course of caudate evoked EPSPs is unaltered by the action potential triggered by one of them. In (A), between the second and third stimuli, a spontaneous spike discharge, arising from a slow-rising EPSP, is noticeable (cf. the rising phases of the spontaneous and evoked EPSPs). Evoked hyperpolarizations are not observed. Note that the surface recording is void of ‘evoked potentials.
driven and nondriven spikes ; the former arose from fast-rising whereas the latter arose from slow-rising EPSPs (Fig. 2A, following the second stimulus). Episodes of hyperpolarizations (40-60 msec durations) were seen in these cells, but they were temporally unlocked to the caudate stimuli.
Stimulation of the distinct sites in the rostromedial head of the caudate nucleus, which yielded the short-latency EPSPs without following IPSPs in the nigral neurons, was unassociated with detectable evoked potentials in surface recordings from the motor cortex (Fig. 2).
Evoked Inhibitory Synupdic Activities. Hyperpolarizing postsynaptic potentials were evoked by caudate stimulation in nine of the 17 nigral neurons. The latencies of the caudate-evoked IPSPs exhibited a bimodal distribution.
a. Short-latency IPSPs. Low-frequency (8 Hz) stimulation of the rostromedial third of the head of the caudate nucleus (Fig. 1) elicited short-latency (3-5 msec) IPSPs in four of the 17 nigral neurons (Figs. 3-SB). The latencies were unvarying in each of three cells (Figs. 3, 5A, 5B) ; this can be readily appreciated by the superimposition of intracellular recordings on Figs. 5A and B. Gradual and progressive prolongation of

NIGRAL NEURONS 129
FIG. 3. Caudate evoked short-latency IPSPs in a nigral neuron. (A-C) Continuous recordings. (A) Background activity of the neuron (spike discharges and prominent synaptic noise) is shown. (B, C) Rostromedial caudate stimulation elicits 3-4 msec latency IPSPs in this neuron. The dashed horizontal lines are drawn through the putative baseline level to emphasize that the caudate stimulation elicits only IPSPs without prior EPSPs in this neuron. Spontaneous activity is significantly reduced in the nigral neuron during the entire period of caudate stimulation. Observe the “double-negativities” on the motor cortical recordings.
MC!. ,,, ,, ,., 1, a
*
4 c - 6
- 56 mV
FIG. 4. Lack of convergence between caudato-fugal and entopedunculo-projection activities in a nigral neuron. (A) Background activity of the neuron. Spike dis- charge arise from slow-rising EPSPs. (B) Rostromedial caudate stimulation elicits 3-4 msec latency, prolonged IPSPs in this nigral neuron. (C) Entopeduncular stimu- lation (8 Hz, at open circles) is unassociated with evoked synaptic activities in this neuron. Surface recordings : (B) caudate evoked “double-negativities” are seen; (C) entopeduncular evoked short-latency positivities are noticeable (cf. the entopeduncu- lar evoked positivity in the motor cortex with those in Figs. SC and D; only the latter two are succeeded by long-latency, evoked IPSPs in nigral neurons).
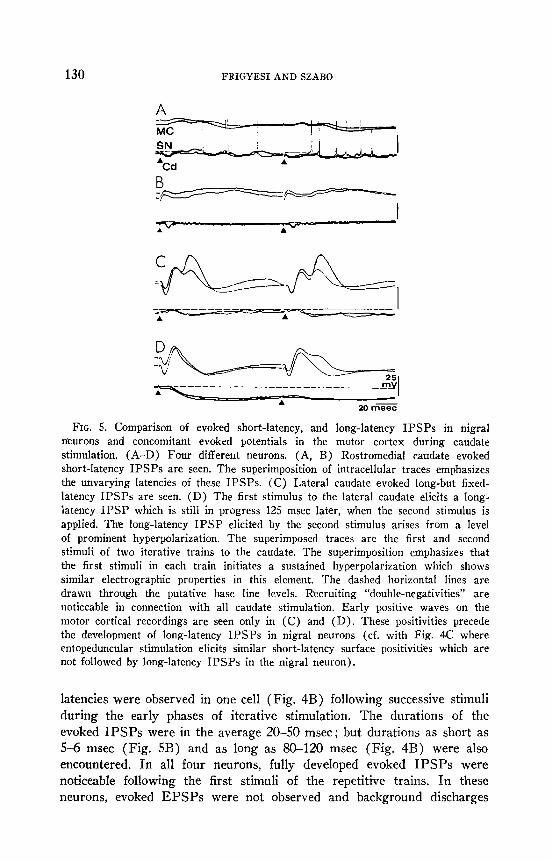
130 FRIGYESI AND SZABO
A Gr=?-=-
I I !
--- %d
AI) *
FIG. 5. Comparison of evoked short-latency, and long-latency IPSPs in nigral neurons and concomitant evoked potentials in the motor cortex during caudate stimulation. (A-D) Four different neurons. (A, B) Rostromedial caudate evoked short-latency IPSPs are seen. The superimposition of intracellular traces emphasizes the unvarying latencies of these IPSPs. (C) Lateral caudate evoked long-but fixed- latency IPSPs are seen. (D) The first stimulus to the lateral caudate elicits a long- latency IPSP which is still in progress 125 msec later, when the second stimulus is applied. The long-latency IPSP elicited by the second stimulus arises from a level of prominent hyperpolarization. The superimposed traces are the first and second stimuli of two iterative trains to the caudate. The superimposition emphasizes that the first stimuli in each train initiates a sustained hyperpolarization which shows similar electrographic properties in this element. The dashed horizontal lines are drawn through the putative base line levels. Recruiting “double-negativities” are noticeable in connection with all caudate stimulation. Early positive waves on the motor cortical recordings are seen only in (C) and (D). These positivities precede the development of long-latency IPSPs in nigral neurons (cf. with Fig. 4C where entopeduncular stimulation elicits similar short-latency surface positivities which are not followed by long-latency IPSPs in the nigral neuron).
latencies were observed in one cell (Fig. 4B) following successive stimuli during the early phases of iterative stimulation. The durations of the evoked IPSPs were in the average 20-50 msec ; but durations as short as 5-6 msec (Fig. 5B) and as long as 80-120 msec (Fig. 4B) were also encountered. In all four neurons, fully developed evoked IPSPs were noticeable following the first stimuli of the repetitive trains. In these neurons, evoked EPSPs were not observed and background discharges

NIGRAL NEURONS 131
were significantly reduced (Fig. 3A, B, C) or abolished (Fig. 4A, B) during iterative stimulation of the caudate.
b. Long-latency IPSPs. Low-frequency stimulation of the lateral third of the head of the caudate nucleus (Fig. 1) elicited long-latency (15-20 msec) IPSPs in five of the 17 nigral neurons (Figs. 5C, D). In each cell, the latencies of the evoked IPSPs were unvarying. The duration of these IPSPs ranged from 40 to 100 msec. In all neurons, fully developed IPSPs were noticeable following the first stimulus of a repetitive train. In the neuron illustrated in Fig. SD, the first stimulus of an 8 Hz train induced a prominent, long-latency (18 msec), long-duration (> 100 msec) IPSP which blocked cell discharges. The second and subsequent stimuli elicited similar long-latency, long-duration IPSPs which arose from the level of sustained hyperpolarization initiated by the first stimulus. The super- imposition of intracellular traces shows a remarkable degree of similarity in the development and sustenance of the prolonged hyperpolarizations in this neuron. When high-frequency (So-100 Hz) stimulation of the caudate was employed, the initial stimuli of those trains elicited IPSPs similar to those by the first shocks in the 8 Hz trains. Under these condi- tions, the evoked IPSPs summated, resulted in prolonged hyperpolariza- tions which attenuated gradually ; spike generations recurred after 3-500 msec.
The Relationship of Evoked IPSPs in Nigral Neurons to Evoked Poten- tials in the Motor Cortex during Caudate Stimulation. Low-frequency (6- 12 Hz) stimulation of the caudate either failed to elicit responses in the motor cortex (Figs. 2A, B, upper traces) or evoked two, long latency (20-40 msec) negative waves (Figs. 3B, C ; Fig. 4B ; Figs. SA, B, upper traces) ; the second waves of these “double negativities” frequently exhibited recruiting characteristics (19, 21). It was a common finding in this study that slight (> 1 mm) displacement of the stimulating electrodes resulted in either complete disappearance or sudden development of the caudate evoked potentials in the motor cortex. Development of caudate evoked EPSPs in nigral neurons was not dependent on the presence of evoked potentials in the motor cortex (Fig, 2). Caudate evoked “double-neg- ativities” were invariably present on the surface recordings when evoked short-latency IPSPs were noticeable in nigral neurons (Figs. 3-5B). Stimulating parameters, sufficient to induce such short-latency IPSPs from the rostrornedial third of the caudate, were insufficient to induce the long-latency IPSPs from the lateral third of the caudate. For the latter, several folds higher stimulus intensities were required. Stimulation of the lateral third of the caudate with such stimuli, however, was associated with early positive deflections which preceded the “double-negativities” in the motor cortex (Figs. 5C, D), Attention is directed at this point to the

132 FRIGYESI AND SZABO
records on Fig. 4C where the effects of stimulation of the entopeduncular nucleus are shown. Such stimulation was unavoidably associated with spread of stimulating current to capsular-peduncular long axons, i.e. entopeduncular stimulation was virtually always associated with early positivities in the cortical recording (Fig. 4C, upper trace). However, under these conditions, these short-latency positivities on the surface recordings were unassociated with long-latency IPSPs in nigral neurons (Fig. 4C, lower trace).
DISCUSSION
On the one hand, these data corroborate previous observations; on the other hand, they have disclosed several properties of caudate-nigra synaptic relations which were not apparent in previous intracellular studies of nigral neurons (22, 58). The foregoing results indicate that three distinct varieties of synaptic potentials are recordable from nigral neurons during low-frequency stimulation of various parts of the ipsilateral head of the caudate nucleus : short-latency EPSPs ; short-latency IPSPs ; and long- latency IPSPs. The different effects of caudate stimulation on the elec- tronigrograms appear to be determined by the involvement of different caudate neurons engaging different synaptic junctions in the nigra, rather than by the partial involvement of extracaudate pathways impinging on nigral neurons and of intranigral multineuronal chains (44) ; this view is consonant to that proposed for caudato-pallidal synaptic relations (41).
The latencies of the caudate evoked EPSPs and short-latency IPSPs in nigral neurons were comparable. The values observed in this study (3-5 msec) are in good agreement with those of the caudate evoked monosynaptic EPSPs in nigral neurons found in a previous study (22). Since the largest diameter of axons leaving the caudate are less than 1.6 pm in diameter (52), and since the stimulating-recording distance between the caudate and nigra was a minimum of 20 mm, considerations of latencies alone necessitate that the caudate-evoked EPSPs and short-latency IPSPs in this study are designated as monosynaptic in origin. It is implicit in these observations that these two types of synaptic potentials in nigral neurons are generated by two types of caudate neurons (35) which have axons of similar diameters. Further, necessarily, two types of transmitters (29, 30) and two types of synaptic junctions in nigra (3, 4) are involved as essential conditions for caudato-fugal volleys to induce the EPSPs and short-latency IPSPs in nigral neurons.
The most parsimonious explanation for the bimodal distribution of latencies of the caudate-evoked IPSPs in nigral neurons is that the short- latency IPSPs were generated monosynaptically whereas the long-latency IPSPs were generated polysynaptically. Structure-functional considera-

NIGRAL NEURONS 133
tions indeed support the contention that the short-latency IPSPs are mono- synaptic (vi& supra) . However, several mutually exclusive explanations are applicable to account for the origin of the long-latency IPSPs. Their latencies are sufficiently long (15-20 msec) to include transmission through a number of synapses either in the caudate, or in the entopeduncular nucleus or within the nigra, However, these alternatives are treated here with convenient vagueness because the long-latency IPSPs in each cell exhibited stereotypic electrographic properties ; this phenomenon is incon- sistent with transmission through polysynaptic chains; rather, it is indica- tive of monosynaptic generation of the IPSPs. In considering the mono- synaptic origin of the caudate-evoked long-latency IPSPs in nigral neurons, three apposite alternatives are canvassed: the pathway which mediated the nigropetal activity (1) originated in the neocortex, (2) was an axon-reflex, or (3) originated in the caudate. First. the caudate evoked long-latency IPSPs in nigral neurons were preceded by short-latency positivities on the surface recordings from the motor cortex. These positive waves have been established to be generated by long axons traversing the internal capsule (21). When such positivities are detected during caudate stimulation, stimulating current spread from the caudate to the internal capsule (21, 22). Such ineluctable activation of long axons in the internal capsule spreads rostrafly and thereby elicits antidromic and orthodromic activation of motor cortical neurons, and also spreads caudally and thereby should elicit antidromic and orthodromic activation of nigral neurons if corticonigral and nigrocortical fibers would pass through the internal capsule. However, experimental anatomical data indicate that such fiber systems are virtually absent in the cat and monkey (31,46, 47, 53, 54). The respective studies were not only performed with meticulous care, but also provided corollary data that gives considerable credence to these negative findings. Therefore, it is reasoned, that activation of motor cortex-substantia nigra axons traveling through the internal capsule were not involved in the generation of the long-latency IPSPs in nigral neurons during caudate stimulation. Second, some brachium conjunctivum fibers reach the caudate and could also gain access to nigral neurons (17) ; corticospinal fibers give off axon collaterals to both caudate and nigra ; nigroocaudate axons give off intranigral collaterals; thus, all of these systems could form the struc- tural basis for axon-reflexes between the caudate and the nigra. However, these fiber systems have been shown to be excitatory at their main termina- tions (2, 17, 22) ; @so facto, they are poor candidates for generating IPSPs in nigral neurons. Third, activities, which arose in caudate neurons and traveled through slow conducting pathways, initiated the long-latency IPSPs in nigral neurons. Computations of conduction velocities and cor- responding fiber diameters of such caudate-nigra axons are jettisoned

134 FRIGYESI AND SZABO
despite the availability of data necessary to make rule of thumb estimates (distances between stimulating and recording sites and latencies of evoked PSPs). This is because such computations are vitiated by that the vast majority of caudate efferents are medullated until they reach the “comb system,” but there they abruptly lose their myelin sheath and reach the substantia nigra as unmyelinated axons (11). Inasmuch as the range of diameter of myelinated axons within the caudate is 0.3-1.6 pm (52) and most of the caudate efferents in the “comb system” are slightly more than 0.1 pm in diameter ( 11) , it is not disconcerting to find latencies of caudate evoked IPSPs in nigral neurons as long as 15-20 msec. Yoshida and Precht (59), though through different lines of evidence and reasoning, also reached the same conclusion: they discerned monosynaptic IPSPs of 18.2 msec mean latencies as the unique synaptic potentials in nigral neurons induced by caudate stimulation. (These studies, unlike ours, were per- formed on barbiturate anesthetized cats; McNair et al. (44) observed that the caudate evoked inhibitory effects in the nigra were related to the level of barbiturate anesthesia). That is, the short-latency positivities on the surface recordings from the motor cortex during caudate stimulation may merely reflect that so much stimulating current was required to acti- vate the small diameter caudato-nigral axons that the larger diameter, thus lower threshold, heterogenous capsular axons could have inevitably been activated by the spread of current. Implicit in the foregoing are that the short-latency positivities in the motor cortex are epiphenomena, and heterogenous axons were not involved in the generation of long-latency IPSPs in nigral neurons during stimulation of the caudate.
The three types of reputedly monosynaptic PSPs elicited in nigral neurons during caudate stimulation raise important questions about the nature of pathways and synaptic junctions involved in their production. The caudate-evoked EPSPs and short-latency IPSPs, because of their similar latencies, can be readily accounted for by caudato-fugal activities which arise in caudate neurons of comparable sizes, travel in caudato-nigral axons of comparable diameters, and terminate in different synaptic junc- tions in the substantia nigra. The caudate evoked long-latency IPSPs could be accounted for by caudato-fugal activities which arise in smaller caudate neurons, travel in thinner caudato-nigral axons, and terminate on synaptic junctions not necessarily different from those which generate the short-latency IPSP. However, Fox et al. (11) have found that there is only one type of efferent neuron in the caudate. But Grofova and Rinvik (27) have recognized three types of synaptic endings in the nigra, one of which undoubtedly, and another suspectedly, originated in the caudate. The experiments of Kemp and Powell (35) showed that there are six cell types in the caudate, two of which (one larger and one smaller) were

NIGRAL XEURONS 135
tentatively identified as the source of the efferent fibers of the head of the caudate nucleus. Moreover, Bak (3) described two types of synaptic junc- tions in the substantia nigra of rodents : one (“encircled type”) is composed of two types of terminals, thus “it might be controlled by dual chemical mechanism”, whereas the other (“sandwich type”) comprises only one of these two types of terminals ; thus one of the chemical mechanisms is replicated in both types of junctions. The anatomical and electro-physio- logical data combined suggest that there are at least three efferent systems from the head of the caudate nucleus to the substantia nigra: two fast conducting, and one slow conducting, fiber pathways. It is tempting to conjecture that the “encircled type of synaptic junctions” are involved in the generation of the two types of short-latency PSPs whereas the “sand- wich type junction” of the long-latency IPSPs in the nigral neurons. It is noted that separate excitatory and inhibitory caudate neurons have been proposed to operate on the pallidum also (41) .
The recognition of the coexistence of excitatory and inhibitory caudato- fugal axon terminals in the nigra imparts that two types of transmitters are released by caudato-fugal volleys in the nigra. This is corroborated by the fact that two putative transmitters have been shown to operate in the nigra : cholinergic (SO) and GABAergic (37) pathways have been de- scribed linking the caudate to the nigra as well as cholinergic (30)) and GABAergic (3) receptors have been identified in the nigra. In seeming contradiction, the existence of both of these pathways has been challenged [cholinergic (43) ; GABAergic (29) 1. In view of the topographical arrangement of caudato-nigral axons (56) and the exactness of placements of stimulating electrodes in the caudate for eliciting specific potentials in the nigra (vi& sz~pra), these converse histochemical findings are readily explicable by the criticalness of sampling.
The experimental findings in this study also conform to the view that the major trajectory of caudate effects to the motor cortex involves relays first in the substantia nigra and then in the dorsal thalamus (18, 20). Albeit in numerous anatomical studies the ansa lenticularis has been regarded as the major discharge route of the strio-lenticular system (40, 45, 51), electrophysiological studies seemed to have denuded this concept of its innate meaning: The dominant or preponderant caudate-evoked synaptic effects in ansa lenticularis projection neurons in the globus pallidus are inhibitory (18, 42, 58, 60) : and caudate-evoked potentials in the dorsal thalamus and the motor cortex were demonstrated in pallido- ansectomized animals (13, 26). Such findings have been interpreted as ansa lenticularis may transmit pallido-thalamic excitatory effects from sources mainly other than the caudate (18). These findings make the issue of trajectories for caudate-motor cortex projection activities pungently

136 FRIGYESI AND SZABO
perceptible. The averment that caudate generates not only inhibitory but also excitatory synaptic potentials in nigral neurons supports the hypoth- esis of the caudato-nigro-thalamic projection system.
A germane issue is the self-regulation by the caudate of its nigral input (5, 30). From the available information, it is difficult to draw firm con- clusions whether the nigra exerts a dual, facilitatory-inhibitory control over the caudate neurons. The plausibility of such a mechanism is sup- ported by that in addition to the dopaminergic ( 1) , a nondopaminergic (9, 12) nigro-caudate pathway has also been suggested. Although current electrophysiological data (18, 38) do not support the notion of a direct strio-nigro-striatal loop (5, 7), the existence of a multisynaptic functional loop cannot be excluded. In this formulation, the caudate-evoked EPSPs in nigral neurons might be also involved in maintaining functional relation- ships between the caudate and the nigra by constituting an essential mech- anism in the operation of the reciprocally linked synaptic pathways appended to the direct caudato-nigral and nigro-caudate (22, 25) path- ways.
REFERENCES
1. ANDEN, N.-E., A. CARLSSON, A. DAHLSTR~M, K. FUXE, N.-A. HILLARP, and K. LARSSON. 1964. Demonstration and mapping out of nigro-neostriatal dopamine neurons. Life Sci. 3 : 523-530.
2. ASANUMA, H. 1973. Cerebral cortical control of movement. Physiologist 16: 143- 166.
3. BAK, I. J. 1967. The ultrastructure of the substantia nigra and caudate nucleus of the mouse and the cellular localization of catecholamines. Exp. Brain Res. 3 : 40-57.
4. BAK, I. J. 1975. Some observations on the ultrastructure of the substantia nigra and the striatum with and without destruction of the strio-nigral pathways in the cat, pp. 121-136. In “Subcortical Mechanisms and Sensorimotor Activities.” T. L. Frigyesi [Ed.]. Hans Huber, Bern.
5. BEDARD, P., L. LAROCHELLE, A. PARENT, and L. J. POIRIER. 1969. The nigro- striatal pathway: A correlative study based on neuroanatomical and neuro- chemical criteria in the cat and the monkey. Ex#. Newel. 25: 365-377.
6. BENDRUPS, A. P., and J. S. MCKENZIE. 1974. Suppression of extralemniscal thalamic unit responses by substantia nigra stimulation. Bruin Res. 80: 131- 134.
7. CARPENTER, M. B., and P. PETER. 1972. Nigrostriatal and nigrothalamic fibers in the rhesus monkey. J. Camp. Netkrol. 144: 93-116.
8. DUSSER DE BARENNE, J. G., and W. S. MCCULLOCH. 1938. Sensorimotor cortex, nucleus caudatus and thalamus opticus. J. Neakrophysiol. 1: 364377.
9. FELTZ, P., and J. DECHAMPLAIN. 1972. Persistence of coudate unitary responses to nigral stimulation after destruction and functional impairment of striatal dopaminergic terminals. Bruin Res. 43 : 595600.
10. FONNUM, F., I. GOROFOVA, E. RINVIK, J. STORM-MATHISEN, and F. WALBERG. 1974. Origin and distribution of glutamate decarboxylase in substantia nigra of the cat. Bra& Res. 71: 77-92.

NIGRAL NEURONS 137
11. Fox, C. A., A. N. ANDRADE, R. C. SCRWYN, and J. A. RAFOLS. 1971/1972. The aspiny neurons and the glia in the primate striatum: A Golgi and electro- microscopic study. J. Hirtiforsch. 13 : 341-362.
12. Fox, C. A., D. E. HILLMAN, K. A. SIEGESMUND, and L. A. SETHER. 1966. The primate globus pallidus and its feline and avian homologues: A Golgi and electronmicroscopic study, pp. 237-248. IR “Evolution of the Forebrain, Phylo- genesis and Ontogenesis of the Forebrain.” R. Hassler and H. Stephan [Eds.]. Georg Thieme Verlag, Stuttgart.
13. FRIGYESI, T. L. 1975. Organization of synaptic pathways linking the head of caudate nucleus to the dorsal thalamus. Int. J. Nez~ol. 8 : 111-138.
14. FRIGYESI, T. L. 1972. Intracellular recordings from neurons in the dorsalateral thalamic reticular nucleus during capsular, basal ganglia and midline thalamic stimulation. Bruin Rex. 48 : 1.57-172.
15. FRIGYESI, T. L. 1974. Caudate evoked IPSPs in nigral neurons. ZRCS (Neuro- biol. Physiol.) 2 : 1720.
16. FRIGYESI, T. L. 1975. Caudate evoked PSPs in nigral neurons. rlnat. Rec. 181: 530-531.
17. FRIGYESI, T. L. 1975. Basal ganglia-thalamus interface, pp. 251-288. Itr “Sub- cortical Mechanisms and Sensorimotor Activities.” T. L. Frigyesi [Ed.]. Hans Huber, Bern.
18. FRIGYESI, T. L. 1975. Structure-function relationships of the interconnections be- tween the caudate nucleus, globus pallidus, substantia nigra and thalamus, pp. 13-45. In “Subcortical Mechanisms and Sensorimotor Activities.” T. L. Frigyesi [Ed.]. Hans Huber, Bern.
19. FRIGYESI, T. L., and J. MACHEK. 1970. Basal ganglia-diencephalon synaptic rela- tions in the cat. I. An intracellular study of dorsal thalamic neurons during capsular and basal ganglia stimulation. Braitt RES. 20: 201-217.
20. FRIGYESI, T. L., and J. MACHEK. 1971. Basal ganglia-diencephalon synaptic rela- tions in the cat. II. Intracellular recordings from dorsal thalamic neurons during low frequency stimulation of the caudatothalamic projection system and the nigrothalamic pathway. Brain REX 27 : 59-78.
21. FRIGYESI, T. L., and D. P. PURPURA. 1964. Functional properties of synaptic pathways influencing transmission in the specific cerebella-thalamocortical pro- jection system. Exp. Neural. 10 : 305-324.
22. FRIGYESI, T. L., and D. P. PURPURA. 1967. Electrophysiological analysis of reciprocal caudato-nigral relations. Brais RES. 6 : 440-456.
23. FRIGYESI, T. L., and A. RABIN. 1971. Basal ganglia-diencephalon synaptic rela- tions in the cat. III. An intracellular study of ansa lenticularis, lenticular fasciculus and pallidosubthalamic projection activities. Brairt Res. 35 : 67-78.
24. FRIGYESI, T. L., and R. SCHWARTZ. 1972. Cortical control of thalamic sensori- motor relay activities in the cat and the squirrel monkey, pp. 161-196. In “Corticothalamic Projections and Sensorimotor Activities.” T. L. Frigyesi, E. Rinvik, and M. D. Yahr [Eds.]. Raven Press, New York.
25. FRIGYESI, T. L., and J. SZABO. 1974. Dendritic spike generation in caudate neurons induced by nigral stimulation in the adult cat. ZRCS (Amt. Newobiol.) 2: 1692.
26. FRIGYESI, T. L., Y. TSUKAMOTO, A. IGE, J. SZABO, and L. COHEN. 1974. Sub- stantia nigra and the sensorimotor system. (Chemonigrectomies induced by intrategmental administration of the paraquinone of 6-hydroxydopamine and 6, 7-dihydroxytryptamine in the cat and monkey.) Int. J. Nczwol. 10: 98-114.

138 FRIGYESI AND SZABO
27. GROFOVA. I., and E. RINVIK. 1970. An experimental electron-microscopic study on the striato-nigral projection in the cat. Exp. Bruin Res. 11 : 249-262.
28. HAJDU, F., R. HASSLER, and I. J. BAK. 1973. Electronmicroscopic study of the substantia nigra and the strio-nigral projection in the cat. 2. Zellforschzing. 146 : 207-221.
29. HATTORI, T., P. L. MCGEER, H. C. FIBIGER, and E. G. MCGEER. 1973. On the source of GABA-containing terminals in the substantia nigra. Electron- microscopic, autoradiographic and biochemical studies. Brain Rcs. 54 : 103-114.
30. JAVOY, F., Y. AGID, D. BOUVET, and J. GLOWINSKI. 1974. Changes in neostriatal DA metabolism after carbachol or atropine microinjections into the substantia nigra. Brailt Rcs. 68 : 253-260.
31. JUNG, R., and R. HASSLER. 1960. The extrapyramidal system, pp. 863-928. In “Handbook of Physiology.” Section I. Vol. II. J. Field, H. W. Magoun, and V. E. Hall [Eds.]. Amer. Physiol. Sot., Washington, D.C.
32. JUNG, R., and J. F. TGNNIES. 1950. Hirnelektrische Untersuchungen iiber Entste- hung und Erhaltung von Krampfentladung: Die Vorginge am Reizort und die Bremsfahigkeit des Gehirns. Arch. Psychiat. Ncrvenkr. 18.5: 701-735.
33. KATAOKA, K., I. J. BAK, R. HASSLER, J. S. KIM, and A. WAGNER. 1974. L- glutamate decarboxylase and choline acetyltransferase activity in the sub- stantia nigra and the striatum after surgical interruption of the strio-nigral fibers of the baboon. Exp. Brain Res. 19 : 217-227.
34. KEMP, J. M. 1970. The termination of strio-pallidal and strio-nigral fibers. Brain Res. 17 : 125-128.
35. KEMP, J. M. and T. P. S. POWELL. 1971. The structure of the caudate nucleus of the cat: light and electronmicroscopy. Phil. Traw. Roy. Sot. Lond. B. 262: 383-401.
36. KEMP, J. M. and T. P. S. POWELL. 1971. The connexion of the striatum and globus pallidus: Synthesis and speculation. Phil. Trans. Roy. Sot. Loud. B. 262 : 441-457.
37. KIM, J. S., I. J. BAK, R. HASSLER, and Y. OKADA. 1971. Role of y-aminobutyric acid (GABA) in the extra-pyramidal motor system. 2. Some evidence for the existence of a type of GABA-rich strio-nigral neurons. Exp. Brain Res. 14: 95-104.
38. KITAI, S. T., W. PRECHT, T. OHNO, and A. WAGNER. 1974. Antidromic and orthodromic activation of the caudate neurons. Abstracts, Sot. for Neuro- science, Fourth Ann. Meeting. p. 285.
39. KLEE, M. R., and H.-D. Lux. 1962. Intracellulire Untersuchungen iiber den Einfluss hemmender Potentiale im motorischen Cortex. II. Die Wirkungen elektrischer Reizung des Nucleus Caudatus. 2. GES. Neural. Psychiat. 203: 667-689.
40. Kuo, J.-S., and M. B. CARPENTER. 1973. Organization of pallidothalamic projec- tions in the rhesus monkey. J. Camp. Nezbrol. 151 : 201-236.
41. LEVINE, M. S., C. D. HULL, and N. A. BUCHWALD. 1974. Pallidal and ento- peduncular intracellular responses to striatal, cortical, thalamic, and sensory inputs. Exp. New. 44 : 44W60.
42. MALLIANI, A. and D. P. PURPURA. 1967. Intracellular studies of the corpus striatum. II. Patterns of synaptic activities in lenticular and entopeduncular neurons. Brain Res. 6: 341-354.
43. MCGREER, E. G., H. C. FIBIGER, P. L. MCGREER, and S. BROOKE. 1973. Temporal changes in amino synthetizing enzymes of rat extrapyramidal structures after hemitranssections or 6-hydroxydopamine administration. Brain Res. 52 : 28% 300.

NIGRAL NEURONS 139
44. MCNAIR, J. L., J. SUTIN, and T. TSUBOKAWA. 1972. Suppression of cell firing in the substantia nigra by caudate nucleus stimulation. Exp. New. 37 : 395411.
45. METTLER, F. A. 1957. Anatomy and physiology of the extrapyramidal passage. Acta. Med. Belg. 11-37.
46. METTLER, F. A. 1969. Anatomy of the basal ganglia, pp. I-55. In “Handbook of Clinical Neurology.” P. J. Vinken and G. W. Bruyn [Eds.]. North-Holland, Amsterdam.
47. MEITLFX, F. A. 1970. Nigrofugal connections in the primate brain. J. COUI~. Neatrol. 138 : 291-320.
48. NAUTA, W. J. H., and W. R. MEHLER. 1966. Projections of the lentiform nucleus in the monkey. Brain Rcs. 1: 3-42.
49. NAUTA, H. J. W., M. B. PRITZ, and R. J. LASEK. 1974. Afferents to the rat caudoputamen studied with horseradish peroxidase. An evaluation of retro- grade neuroanatomical research method. Br& Rcs. 67: 219-238.
50. OLIVIER, A., A. PARENT, H. SIMARD. and L. J. POIRIER. 1970. Cholinesterasic striatopallidal and striatonigral efferents in the cat and the monkey. Brain Rcs. 18 : fl3-282.
51. PAPEZ, J. W. 1942. A summary of fiber connections of the basal ganglia with each other and with other portions of the brain. .gRNMD 21: 21-68.
52. PURPURA, D. P., T. L. FRIGYESI, and A. MALLIANI. 1967. Intrinsic synaptic organization and relations of the corpus striatum, pp. 177-214. “Neurophysio- logical Basis of Normal and Abnormal Motor Activities.” M. D. Yahr and D. P. Purpura [Eds.]. Raven Press, Hewlett, New York.
53. RINVIK, E. 1966. The co&co-nigral projection in the cat. J. Conzp. Nrzlr. 126:
241-254. 54. RINVIK, E., and F. WALBERG. 1969. Is there a corticospinal tract? A comment
based on expprimental electronmicroscopic observations in the cat. Brairz Res. 14 : 742-744.
55. SHIMAMOTO, T., and M. VERZEANO. 1954. Relation between caudate and dif- fusely projecting thalamic nuclei. J. Nellroplzysiol. 17 : 278-288.
56. SZABO, J. 1962. Topical distribution of striatal efferents in the monkey. Exp. Ncakrol. 5 : 21-36.
57. SZABO, J. 1971. A silver impregnation study of nigrostriate projections in the cat. Amt. Rec. 69 : 441.
58. YOSHIDA, M. 1975. Role of GABA and other substances in, and functional aspects of, the basal ganglia, pp. 156-176. ZK “Subcortical Mechanisms and Sensori- motor Activities.” T. L. Frigyesi [Ed.]. Hans Huber, Bern.
59. YOSHIDA, M., and W. PRECHT. 1971. Monosynaptic inhibition of neurons of the substantia nigra by caudatonigral fibers. Brak Rcs. 32: 225-228.
60. YOSHIDA, M., A. RABIN, and M. ANDERSON. 1972. Monosynaptic inhibition of pallidal neurons by axon collaferals of caudato-nigral fibers. Exp. Brain RCS. 15 : 333-347.




















