
Efficient in vivo protection of nigral dopaminergic neurons by lentiviral gene transfer of a...
12
Regular Article Efficient in vivo protection of nigral dopaminergic neurons by lentiviral gene transfer of a modified Neurturin construct Lone Fjord-Larsen, Jens Leander Johansen 1 , Philip Kusk, Jens Tornøe, Mette Grønborg, Carl Rosenblad 2,3 , Lars U. Wahlberg * ,3 NsGene A/S, Baltorpvej 154, 2750 Ballerup, Denmark Received 24 November 2004; revised 28 February 2005; accepted 21 March 2005 Available online 24 May 2005 Abstract Protein injection studies of the glial cell line derived neurotrophic factor (GDNF) family member Neurturin (NTN) have demonstrated neuroprotective effects on dopaminergic (DA) neurons, which are selectively lost during Parkinson’s disease (PD). However, unlike GDNF, NTN has not previously been applied in PD models using an in vivo gene therapy approach. Difficulties with lentiviral gene delivery of wild type (wt) NTN led us to examine the role of the pre-pro-sequence, and to evaluate different NTN constructs in order to optimize gene therapy with NTN. Results from transfected cultured cells showed that wt NTN was poorly processed, and secreted as a pro-form. A similarly poor processing was found with a chimeric protein consisting of the pre-pro-part from GDNF and mature NTN. Moreover, we found that the biological activity of pro-NTN differs from mature NTN, as pro-NTN did not form a signaling complex with the tyrosine kinase receptor Ret and GFRa2 or GFRa1. Deletion of the pro-region resulted in significantly higher secretion of active NTN, which was further increased when substituting the wt NTN signal peptide with the immunoglobulin heavy-chain signal peptide (IgSP). The enhanced secretion of active mature NTN using the IgSP-NTN construct was reproduced in vivo in lentiviral-transduced rat striatal cells and, unlike wt NTN, enabled efficient neuroprotection of lesioned nigral DA neurons, similar to GDNF. An in vivo gene therapy approach with a modified NTN construct is therefore a possible treatment option for Parkinson’s disease that should be further explored. D 2005 Elsevier Inc. All rights reserved. Keywords: Neurturin; Signal peptide; Pro-peptide processing; 6-hydroxydopamine lesion; Lentiviral delivery; Gene therapy; Parkinson’s disease Introduction Neurturin (NTN) is a member of the glial cell line derived neurotrophic factor (GDNF) family of neurotrophic factors. Recombinant NTN protein has been shown to promote the survival and maintenance of several different neuronal subtypes including autonomic and somatic periph- eral ganglion neurons, striatal neurons, and nigral dopami- nergic (DA) neurons. Signaling of GDNF family ligands is mediated through a receptor complex consisting of one type of four glycosyl phosphatidylinositol-linked ligand-binding receptors (GFRa1–4) and the transmembrane tyrosine kinase receptor Ret (Baloh et al., 2000). Although NTN preferentially binds to GFRa2, cross-binding to GFRa1 also occurs (Jing et al., 1997; Klein et al., 1997; Sanicola et al., 1997). Studies on cell cultures derived from animals lacking GFRa1 have shown that, in the nigral DA neurons, NTN appears to signal through GFRa1(Cacalano et al., 1998). This is consistent with the high expression of GFRa1 in the developing and adult nigral DA neurons while GFRa2 is only modestly expressed, if at all, in this area (Golden et al., 1998; Widenfalk et al., 1997). In studies using recombinant protein injections, NTN has been found to prevent the loss of damaged nigral dopamine neurons in an animal model of Parkinson’s disease (PD) and 0014-4886/$ - see front matter D 2005 Elsevier Inc. All rights reserved. doi:10.1016/j.expneurol.2005.03.006 * Corresponding author. Fax: +45 44608989. E-mail address: [email protected] (L.U. Wahlberg). 1 Current address: Lundbeck A/S, 2500 Valby, Denmark. 2 Current address: Department of Surgery, Lund University Hospital, Lund, Sweden. 3 Contributed equally to this work. Experimental Neurology 195 (2005) 49 – 60 www.elsevier.com/locate/yexnr
Transcript of Efficient in vivo protection of nigral dopaminergic neurons by lentiviral gene transfer of a...

www.elsevier.com/locate/yexnr
Experimental Neurology
Regular Article
Efficient in vivo protection of nigral dopaminergic neurons by
lentiviral gene transfer of a modified Neurturin construct
Lone Fjord-Larsen, Jens Leander Johansen1, Philip Kusk, Jens Tornøe, Mette Grønborg,
Carl Rosenblad2,3, Lars U. Wahlberg*,3
NsGene A/S, Baltorpvej 154, 2750 Ballerup, Denmark
Received 24 November 2004; revised 28 February 2005; accepted 21 March 2005
Available online 24 May 2005
Abstract
Protein injection studies of the glial cell line derived neurotrophic factor (GDNF) family member Neurturin (NTN) have demonstrated
neuroprotective effects on dopaminergic (DA) neurons, which are selectively lost during Parkinson’s disease (PD). However, unlike GDNF,
NTN has not previously been applied in PD models using an in vivo gene therapy approach. Difficulties with lentiviral gene delivery of wild
type (wt) NTN led us to examine the role of the pre-pro-sequence, and to evaluate different NTN constructs in order to optimize gene therapy
with NTN. Results from transfected cultured cells showed that wt NTN was poorly processed, and secreted as a pro-form. A similarly poor
processing was found with a chimeric protein consisting of the pre-pro-part from GDNF and mature NTN. Moreover, we found that the
biological activity of pro-NTN differs from mature NTN, as pro-NTN did not form a signaling complex with the tyrosine kinase receptor Ret
and GFRa2 or GFRa1. Deletion of the pro-region resulted in significantly higher secretion of active NTN, which was further increased when
substituting the wt NTN signal peptide with the immunoglobulin heavy-chain signal peptide (IgSP). The enhanced secretion of active mature
NTN using the IgSP-NTN construct was reproduced in vivo in lentiviral-transduced rat striatal cells and, unlike wt NTN, enabled efficient
neuroprotection of lesioned nigral DA neurons, similar to GDNF. An in vivo gene therapy approach with a modified NTN construct is
therefore a possible treatment option for Parkinson’s disease that should be further explored.
D 2005 Elsevier Inc. All rights reserved.
Keywords: Neurturin; Signal peptide; Pro-peptide processing; 6-hydroxydopamine lesion; Lentiviral delivery; Gene therapy; Parkinson’s disease
Introduction
Neurturin (NTN) is a member of the glial cell line
derived neurotrophic factor (GDNF) family of neurotrophic
factors. Recombinant NTN protein has been shown to
promote the survival and maintenance of several different
neuronal subtypes including autonomic and somatic periph-
eral ganglion neurons, striatal neurons, and nigral dopami-
nergic (DA) neurons. Signaling of GDNF family ligands is
0014-4886/$ - see front matter D 2005 Elsevier Inc. All rights reserved.
doi:10.1016/j.expneurol.2005.03.006
* Corresponding author. Fax: +45 44608989.
E-mail address: [email protected] (L.U. Wahlberg).1 Current address: Lundbeck A/S, 2500 Valby, Denmark.2 Current address: Department of Surgery, Lund University Hospital,
Lund, Sweden.3 Contributed equally to this work.
mediated through a receptor complex consisting of one type
of four glycosyl phosphatidylinositol-linked ligand-binding
receptors (GFRa1–4) and the transmembrane tyrosine
kinase receptor Ret (Baloh et al., 2000). Although NTN
preferentially binds to GFRa2, cross-binding to GFRa1 also
occurs (Jing et al., 1997; Klein et al., 1997; Sanicola et al.,
1997). Studies on cell cultures derived from animals lacking
GFRa1 have shown that, in the nigral DA neurons, NTN
appears to signal through GFRa1 (Cacalano et al., 1998).
This is consistent with the high expression of GFRa1 in the
developing and adult nigral DA neurons while GFRa2 is
only modestly expressed, if at all, in this area (Golden et al.,
1998; Widenfalk et al., 1997).
In studies using recombinant protein injections, NTN has
been found to prevent the loss of damaged nigral dopamine
neurons in an animal model of Parkinson’s disease (PD) and
195 (2005) 49 – 60

L. Fjord-Larsen et al. / Experimental Neurology 195 (2005) 49–6050
to enhance the function of intact DA neurons in vivo (Horger
et al., 1998; Rosenblad et al., 2003). The neuroprotective and
functional effects of NTN are equipotent to GDNF when
tested on developing ventral mesencephalic DA neurons
cultured in vitro (Cacalano et al., 1998; Horger et al., 1998)
as well as on damaged nigrostriatal DA neurons in vivo
(Horger et al., 1998). Based on the neuroprotective effects on
nigrostriatal DA neurons, NTN has been suggested as a
candidate for treatment of PD. However, the progressive
nature of PD, as well as findings from numerous preclinical
studies using GDNF, indicates that continuous administra-
tion over long periods of time may be necessary for
neuroprotection and functional recovery in the nigrostriatal
DA system. Intracerebral protein delivery is cumbersome
and inefficient, making therapeutic approaches that use
delivery by viral gene transfer attractive choices. To date,
several recombinant viral vectors have been tested for long-
term delivery of neurotrophic factors in vivo. Recombinant
adeno-associated virus (AAV) and lentivirus (rLV) are the
most extensively used in the nervous system due to their
ability to infect non-dividing cells and provide stable
expression (Bensadoun et al., 2000; Bilang-bleuel et al.,
1997; Connor et al., 2001; Georgievska et al., 2004;
Kordower, 2003; Lapchak et al., 1997; Mandel et al.,
1997; Rosenblad et al., 2000). In vivo gene therapy with
wild type (wt) GDNF has successfully been applied in
different experimental paradigms. However, data from our
lab indicate that wt NTN is not readily expressed in vivo
following transduction of cells using an rLV vector contain-
ing human wt NTN cDNA (unpublished results). The reason
for the difference in GDNF and NTN behavior is unknown
but one hypothesis could be a faulty processing of the NTN
protein. Human wt NTN encodes a 197-residue pre-pro-
protein with a predicted 19 amino acid signal peptide,
followed by a pro-region of 76 amino acids. Pro-NTN is
proteolytically cleaved at the RXXR consensus recognition
sequence for furin-like proteases to generate the mature NTN
of 102 residues.
In the present study, we investigated the role of the pre-
pro-region of human NTN in the secretion of active NTN
protein from cultured cells transfected with NTN constructs
in which different parts of the pre-pro-sequences had been
modified. We found that wt NTN was poorly processed to
the mature form and that the secreted pro-form of NTN was
not able to bind to GFRa1 or GFRa2 receptors. Deletion of
the pro-region resulted in significantly increased secretion
of active NTN, and by using a heterologous signal peptide,
the immunoglobulin heavy-chain signal peptide (IgSP)
sequence, secretion of active NTN was further increased
to more than 100-fold the levels obtained with the wt NTN
sequence. The IgSP-NTN expression construct was tested in
the intrastriatal 6-hydroxydopamine lesion model of PD via
a lentiviral in vivo gene transfer approach. Unlike wt NTN,
the IgSP-NTN gave potent neuroprotective effects demon-
strating for the first time the potential of NTN in vivo gene
therapy for PD.
Methods
DNA vectors
wt NTN constructs (pHRV-CMV.SIN.hNTN.WPRE and
pNS1n.hNTN)
Wild type human pre-pro-NTN was cut from vector
pJDM2174 (human pre-pro-NTN in pBluescript; a kind gift
from Dr. Jeffrey Milbrandt, Wash. U., St. Louis, USA) as a
BamHI, XhoI fragment and cloned into pHRV-CMV.SIN-
PLT7.WPRE, a modified derivative of pHRV-CMV-SIN-18
lentiviral transfer vector containing the Woodchuck hepatitis
virus postregulatory element (WPRE) (Zufferey et al., 1997,
1998). Similarly, the BamHI, XhoI fragment was cloned in
pNS1n (a kind gift from P. Ahring NeuroSearch A/S, DK) a
derivative of pcDNA3.1 from Invitrogen, Denmark, to yield
pNS1n.hNTN. In both vectors, the CMV promoter drives
the expression of NTN expression.
ppG-NTN construct (pNS1n.ppGDNF.hNTN)
The pre-pro-region of GDNF was PCR amplified from a
full-length human GDNF clone using the following primers:
5V primer: 5V-TATAGAATTCGCCACCATGAAGTTATGG-GATGTCG-3V and 3V primer: 5V-CCAACCGCGCCCTTTT-CAGTCTTTTAATGG-3V. The 3V primer contains 10 bases of
the 5V end of mature NTN in the 3V end. Mature human NTN
was PCR amplified from human genomic DNA using the
following primers: 5V primer: 5V-ACTGAAAAGGGCGC-GGTTGGGGGCGCGGCCT-3V and 3V primer: 5V-TAGACT-CGAGGTCGACGGTATC-3V. The 5V primer contains 10
bases of the 3V end of the pro-region of human GDNF. The
pre-pro-region of GDNF was fused to mature NTN by
overlapping PCR using the following primers: 5V primer: 5V-TATAGAATTCGCCACCATGAAGTTATGGGATGTCG-
3V and 3V primer: 5V-TAGACTCGAGGTCGACGGTATC-3V.The resulting pre-pro-GDNF-mature NTN fragment was
digested with EcoRI and XhoI and inserted between EcoRI
and XhoI sites of expression vector pNS1n (described
above).
IgSP-NTN construct (pHRV-CMV.SIN.IgSP.NTN.WPRE and
pNS1n.IgSP.NTN)
The signal peptide from mouse immunoglobulin heavy-
chain gene V-region (GenBank acc. no.: M18950) (IgSP)
was PCR amplified from pNUT-IgSP-hCNTF (ref. US
6,361,741) using the following primers: 5V primer: 5V-TAQTAGGATCCGCCACCATGAAATGCAGCTGGGTTATC-
3V, 3V primer: 5V-CCAACCGCGCCGAATTCACCCCTG-TAGAAAG-3V. The 3V primer contains 10 bases from the 5Vend of human, mature NTN sequence. Human, mature NTN
was PCR amplified from human genomic DNA using the
following primers: 5V primer: 5V-GGTGAATTCGGCGC-GGTTGGGGGCGCGGCCT-3V and 3V primer: 5V-TATACT-CGAGTCACACGCAGGCGCACTCGC-3V. The 5V primer
contains 10 bases from the 3V end of the IgSP sequence. The
IgSP-human mature NTN sequence was generated by

L. Fjord-Larsen et al. / Experimental Neurology 195 (2005) 49–60 51
overlapping PCR using the IgSP and human mature NTN
PCR fragments (described above) as templates and the
following primers: 5V primer: 5V-TATAGGATCCGCCAC-CATGAAATGCAGCTGGGTTATC-3V and 3V primer: 5V-TATACTCGAGTCACACGCAGGCGCACTCGC-3V. The
final PCR fragment containing IgSP fused to human mature
NTN was digested with BamHI and XhoI and cloned
between BamHI and XhoI sites of pHRV-CMV.SIN-
PLT7.WPRE (described above) and pNS1n (described
above).
Dpro-NTN construct (pNS1n-Dpro-NTN)The human delta-pro DNA sequence, containing the pre-
but not pro-regions of wt NTN, was generated in one PCR
reaction using human genomic DNA as template and the
following primers: 5V primer: 5V-TATAGGATCCGCCACC-ATGCAGCGCTGGAAGGCGGCGGCCTTGGCCTCAG-
TGCTCTGCAGCTCCGTGCTGTCCGCGCGG-
TTGGGGGCGCGG-3V and 3V primer: 5V-TATACTCGAGTQCACACGCAGGCGCACTCGC-3V. The delta-pro-NTN
PCR fragment was digested with BamHI and XhoI and
cloned between BamHI and XhoI sites of pNS1n (described
above).
Production of lentiviral vectors
Replication-defective lentivirus particles were generated
by co-transfection of each of the different transfer vector
constructs (described above) with pMD.G (VSV-G pseudo-
typing vector) and pBR8.91 (packaging vector) (Zufferey
et al., 1999) into 293 T cells providing the required viral
proteins in trans. Briefly, 293 T cells cultured in DMEM
with 4.5 g/l glucose and Glutamaxi (Invitrogen, DK)
supplemented with 10% FCS (Life Technologies, DK)
were seeded in T75 flasks (2 � 106 cells/flask) the day
before transfection. For each T75 flask, cells were trans-
fected with 5 Ag pMD.G, 15 Ag pBR8.91, and 20 Ag of
transfer vector using Lipofectamine+i (Invitrogen, DK)
according to the manufacturer’s instructions. Virus con-
taining cell supernatant was collected 2–3 days after the
transfection, filter-sterilized through a 0.45-Am cellulose
acetate or polysulphone filter, and concentrated by ultra
centrifugation at 50,000 � g for 90 min at 4-C. After a
second round of ultra centrifugation, the concentrated virus
pellet was resuspended in DMEM and aliquoted and stored
at �80-C. To determine virus titer, reverse transcriptase
(RT) activity was assayed (Cepko and Pear, Current
Protocols in Molecular Biology, 9.13.5-6, supplement 36)
and transducing units (TU)/ml were calculated from the
determined RT activity using an EGFP lentivirus with
known transducing activity as reference.
Cell culture
ARPE-19, a spontaneously arising human retinal pig-
ment epithelial cell line (Dunn et al., 1996), was grown in
medium consisting of DMEM/Nutrient Mix F-12 with
Glutamax (Invitrogen, DK), supplemented with 10% fetal
bovine serum (Sigma-Aldrich, DK). HiB5 (Renfranz et al.,
1991), HEK293, and CHO cells were grown in DMEM
(Invitrogen, DK) with 10% fetal bovine serum (Invitrogen,
DK), and medium for CHO cells was further supplemented
with 20 mg/l l-proline. ARPE-19, HEK293, and CHO cells
were grown at 37-C and HiB5 cells at 33-C in 5% CO2.
Transient transfection studies
Cells were seeded in 6-well plates (Corning Costar,
Biotech Line, DK) at a density of approximately 105 cells/
well. The next day, cells were transfected in triplicate wells
with the different expression plasmids. ARPE-19 cells were
transfected using Fugene6 and 3 Ag plasmid/well according
to the manufacturer’s instructions, whereas the other three
cell lines were transfected in triplicate wells using 2 Agplasmid/well and Lipofectamine Plus (Invitrogen, DK)
according to the manufacturer’s instructions. The next day,
fresh growth medium was added to the wells, and cells were
incubated for an additional 24 h before collecting condi-
tioned medium and harvesting the cells. A transfection
efficiency of more than 60% was ensured by evaluation of
EGFP expression in wells transfected in parallel with the
same vector backbone containing the cDNA for EGFP.
NTN Western blotting
Cell lysates were prepared in 96-C hot sample buffer (2%
SDS, 100 mM DTT, 60 mM Tris, pH 7.5, bromphenolblue).
5� concentrated sample buffer was added to conditioned
media. In some experiments, NTN was captured by GFRa2
from conditioned medium by incubating samples for 3 h in
ELISA plates coated with goat anti-human Fc (Jackson
ImmunoResearch, Trichem, DK) bound to a GFRa2-Ig
fusion protein (R&D Systems, UK) before hot sample buffer
was added. After boiling for 5 min, samples were loaded
onto 8–18% gradient SDS gels and electrophoresed
followed by electroblotting to PVDF membranes. NTN
was detected using polyclonal NTN antibody (#AF477,
R&D Systems, UK) diluted 1:500 followed by HRP-linked
anti-goat antibody. Bands were detected by chemolumines-
cence using the ECL+ system (Amersham Life Science).
GFRa2 ELISA
The GFRa2 ELISA detects the ternary complex for-
mation between NTN, Ret, and GFRa2 and was performed
as previously described with minor modifications (Sanicola
et al., 1997). Briefly, an Opti-plate plate (Packard Instru-
ments, Perkin Elmer, DK) was coated with 100 Al 1 Ag/ml
goat anti-human Fc (Jackson Immunoresearch Laboratories,
TriChem, DK) in 50 mM NaHCO3 (pH = 9.6). Wells were
blocked in 0.2% I-Block (Tropix, Roche, DK) in PBST
(PBS with 0.1% Tween-20) for 1 h at room temperature.

L. Fjord-Larsen et al. / Experimental Neurology 195 (2005) 49–6052
Samples from NTN-producing cells or standards (recombi-
nant human NTN from R&D Systems, UK) diluted in
ARPE-19 growth medium were added together with 1 Ag/ml
GFRa2/Fc fusion protein (R&D Systems, UK) in condi-
tioned medium from 293 EBNA cells expressing Ret-
Alkaline Phosphatase (Ret-AP) fusion protein (a kind gift
from Biogen Idec, USA). After incubation (1.5 h at room
temperature) and between steps, the wells were washed
three times with PBST. Wells were then washed in AP-
buffer [200 mM Tris (pH = 9.8), 10 mM MgCl2] and ternary
complex formation was detected using a chemiluminescent
substrate for AP by incubation for 30 min with 10%
Sapphire Enhancer (Tropix, Roche, DK) and 2% CSPD
(Tropix, Roche, DK) in AP-buffer. Luminescence was
quantified using Microbeta Trilux Counter (Perkin Elmer,
DK). The relative GFRa2 binding activity in samples was
calculated from a standard curve generated by using
recombinant NTN protein. Binding activity to GFRa1 in
conditioned medium was measured similarly, but with 1 Ag/ml GFRa1/Fc fusion protein (R&D Systems, UK) added
instead of GFRa2/Fc.
NTN ELISA
To detect immunoreactive NTN in media samples, a
sandwich ELISAwas used. Briefly, MaxiSorp plates (Nunc,
DK) were coated with 1 Ag/ml monoclonal anti-human
NTN antibody (#MAB387, R&D Systems, UK) in 2.5 mM
Na2CO3/2.5 mM NaHCO3, pH 8.2. Wells were blocked in
blocking buffer [1% bovine serum albumin (Sigma-Aldrich,
DK) in PBS] supplemented with 5% sucrose followed by
incubation with diluted media samples from NTN-produc-
ing cells or NTN standards (recombinant NTN #387-NE
from R&D Systems, UK) for 3 h at room temperature. To
detect bound NTN, wells were incubated with 1 Ag/ml
polyclonal anti-human NTN antibody (#AF387, R&D
Systems, UK) in blocking buffer and subsequently with
0.02% anti-goat-HRP (DAKO, DK) in blocking buffer
supplemented with 1% normal mouse serum (DAKO, DK).
In between each step, wells were washed three times with
PBST [0.05% Tween-20 (Sigma-Aldrich, DK) in PBS]. To
detect bound HRP-activity, 3,3V,5,5V-tetramethylbenzidine
substrate solution (Promega, Ramcon, DK) was added, and
color formation was stopped by addition of 1 N HCl after 15
min. A450 was measured using an ELX-800 plate reader
(Cambrex, DK).
Surgical procedures
A total of 24 young adult female Sprague–Dawley rats
(Møllegarden, DK) was used and housed under 12-h
light:dark cycle with free access to rat chow and water.
Virus injections and 6-hydroxydopamine lesions were
performed according to Rosenblad et al. (2000). Briefly,
isofluorane anesthetized (1.5–2%) animals were injected
with rLV vector (3 � 105 TU/animal) carrying the cDNA for
GFP, NTN, IgSP-NTN, or GDNF (n = 5–7/group). Using a
Hamilton microsyringe with a pulled glass pipette tip, four
deposits (0.75 Al/deposit) were made into the striatum along
two needle tracts at the following coordinates with respect to
bregma: (1) AP = 1.0 mm, ML = �2.6 mm; and (2) AP =
0.0 mm, ML = �3.7 mm, and DV1 = �5.0 mm, DV2 = �4.5
mm with respect to the dura. The tooth bar was set at �2.3
mm. 14 days after rLV injections, the animals were
anesthetized and a single deposit of 20 Ag 6-hydroxydo-
pamine (6-OHDA from Sigma; calculated as free base and
dissolved in 3 Al ice-cold saline supplemented with 0.02%
ascorbic acid) was injected into the right striatum with a 10-
Al Hamilton syringe at the coordinates AP = 0.5 mm; ML =
�3.4 mm relative to bregma, DV = �5.0 mm relative to the
dura and tooth bar set to 0.0 mm. The injection rate was
1 Al/min and the glass pipette was left in place for an
additional 3 min before withdrawal.
Amphetamine-induced rotation
At 10 days after rLV injections and again 4 weeks after
the 6-hydroxydopamine injections, rats were injected with
amphetamine (2.5 mg/kg, Mecobenzon, DK) and moni-
tored for turning response in automated rotometer bowls
over 90 min. Rotational asymmetry scores are expressed as
net full ipsilateral turns per minute (i.e., towards the
injected site).
Histology
Twenty-eight days after the 6-hydroxydopamine injec-
tions, the animals were deeply anesthetized with sodium
pentobarbital and transcardially perfused with saline for 1
min followed by 200 ml ice-cold 4% paraformaldehyde in
0.1 M phosphate buffer (pH 7.4). The brains were
dissected and post-fixed in the same fixative for 3–4 h
and then transferred into 25% sucrose/0.1 M phosphate
buffer for 48 h. Six series of 40-Am sections were cut on
a freezing microtome. Immunohistochemistry was per-
formed as described previously (Rosenblad et al., 1999).
In brief, sections were incubated overnight with goat-anti-
hNTN (R&D Systems, UK; 1:2000), goat-anti-hGDNF
(R&D Systems, UK; 1:2000), chicken anti-GFP (Chem-
icon; 1:2000), mouse anti-tyrosine hydroxylase (TH)
(Chemicon; 1:2000), or rabbit anti-vesicular monoamine
transporter (VMAT) (Chemicon; 1:2000) primary anti-
bodies diluted in phosphate buffered saline containing 2%
normal horse or swine serum and 0.25% Triton X-100
followed by incubation with the appropriate (anti-goat,
anti-chicken, anti-mouse, or anti-rabbit) biotinylated sec-
ondary antibody (all from Jackson Immunoresearch,
Trichem, DK and diluted 1:200) for 2 h, and avidin–
biotin complex (ABC) kit according to the manufacturer’s
instructions (Vector Laboratories, USA). Finally, the color
reaction was developed using 3V3V-diaminobenzidine as
chromogen.

L. Fjord-Larsen et al. / Experimental Neurology 195 (2005) 49–60 53
Morphometric analysis: Quantification of neurons
expressing the DA markers TH or VMAT in substantia
nigra (SN)
A person, blinded to the identity of the samples, assessed
the number of immunoreactive neurons in SN pars com-
pacta, as described previously (Sauer and Oertel, 1994). In
brief, three consecutive sections centered at the level of the
medial terminal nucleus of the accessory optic tract (MTN,
�5.3 in the atlas of Paxinos and Watson, 1997) were used
and all stained neurons lateral to the MTN were counted at
40� magnification. Cell numbers are expressed as the mean
T SEM of the percentage of the number in the intact side. For
quantification of the downregulation of TH expression, TH-
immunoreactive cells with normal as well as downregulated
expression were counted in the lesion side (see Fig. 4G for
examples of cells with different TH expression). Cells were
counted at 40� magnification in the same three consecu-
tive sections as described above and expressed as the
mean percentage T SEM of cells with downregulated TH
expression.
Striatal fiber density measurements
Striatal DA innervation was assessed by measuring
optical density (OD) of the striatum at three rostrocaudal
levels in sections stained for TH as described (Kirik et al.,
2001). Using an Olympus DP50 digital camera and a
constant illumination table, digitalized images were col-
lected. ODs on the intact and lesioned sides were measured
using ScanImage v. 4.0.2 software. Corpus callosum in each
section was used as reference for background staining.
Results
Expression of NTN in cultured cells
To characterize the role of the pre-pro-part in the
secretion of active NTN, we generated a series of cDNA
constructs encoding human NTN proteins shown in Fig. 1A.
Western blot analyses of lysates and conditioned medium
from the transfected cells were performed using an antibody
against NTN. Fig. 1B shows results obtained with HEK293
cells. In cells transfected with the wt NTN construct, a band
corresponding to the expected size of monomeric pro-NTN
(¨22 kDa) was detected in cell lysate as well as conditioned
medium under reducing conditions. A band corresponding
to the expected size of the slightly smaller NTN with the
GDNF pro-region (¨19.6 kDa) was seen in cell lysate and
conditioned medium from cells transfected with ppG-NTN.
No bands corresponding to the expected size of mature NTN
was observed in the cells or conditioned medium. Thus, the
pro-forms of NTN are expressed and secreted but not
processed at a detectable level. In cells transfected with
Dpro-NTN or IgSP-NTN, a band corresponding in size to
mature monomeric NTN (¨12.5 kDa) was observed in both
cell lysates and conditioned media. The total level of
secreted mature NTN protein appeared to be higher in cells
transfected with the IgSP-NTN construct than cells trans-
fected with Dpro-NTN. Similar Western blotting results
were obtained with ARPE-19 cells, HiB5 cells, and CHO
cells (data not shown).
We then tested the level of biologically active NTN in
conditioned medium using a functional assay where binding
of NTN to its receptor, GFRa2, is detected by formation of
a ternary complex with the GFRa co-receptor Ret. The
relative activities in this assay compared to the wt NTN
construct are shown in Fig. 1C. Conditioned medium from
cells transfected with wt NTN construct showed low levels
of active NTN (7.5 T 0.6 ng/ml, 1.5 T 0.9 ng/ml, 38.6 T 3.3
ng/ml, and 18.3 T 1.7 ng/ml for wt NTN in HEK293,
ARPE-19, HiB5, and CHO cells, respectively). Equally low
levels were observed when using the ppG-NTN construct.
However, GFRa2 binding activity in samples increased
when using the Dpro-NTN construct (90 T 19, 117 T 13,
7.5 T 1.7, and 4.1 T 0.9 fold higher NTN binding for
HEK293, ARPE-19, HiB5, and CHO cells, respectively). In
accordance with the Western blot results, NTN binding
activity was further enhanced when using the IgSP-NTN
construct (278 T 13, 771 T 50, 162 T 29, and 66 T 18 fold
higher NTN binding for HEK293, ARPE-19, HiB5, and
CHO cells, respectively). Similar results were obtained when
performing the assay with the GFRa1 co-receptor (data not
shown). Cell supernatants from cells transiently transfected
with pNS1n-EGFP showed undetectable NTN activity
confirming the specificity of the assays (data not shown).
In addition, we performed NTN Western blotting on
NTN captured from conditioned media by GFRa2-Ig
immobilized in ELISA plates prior to loading (Fig. 1D).
In contrast to what was seen in Western blot analysis in
which conditioned media were loaded directly (Fig. 1B), we
did not detect pro-forms of NTN derived from cells
transfected with wt NTN and ppG-NTN constructs, whereas
bands corresponding the expected size of mature NTN were
observed in samples from cells transfected with the IgSP-
NTN construct and to a lesser extent when using the Dpro-
NTN vector indicating poor or absent binding of pro-forms
to the GFRa2 receptor.
An NTN sandwich ELISA on conditioned medium from
cells transfected with the different NTN constructs was also
carried out. A monoclonal NTN antibody was used to
capture NTN on ELISA plates and subsequently a poly-
clonal NTN antibody was used to detect the captured NTN.
Both antibodies recognized pro-forms of NTN when used
for Western blotting under reduced conditions, but as shown
in Fig. 1E, the NTN pro-forms were not or very poorly
detected in the NTN ELISA. This finding suggests that the
pro-part of NTN inhibits binding of the antibody to native
NTN, but not to denatured NTN. The NTN sandwich
ELISA confirmed that using the IgSP-NTN construct rather
than the Dpro-NTN construct increases the level of NTN in

Fig. 1. (A) NTN expression constructs encoding wt (pre-pro) NTN, a chimeric NTN with the pre-pro-part derived from GDNF (ppG-NTN), NTN with deleted
pro-region (Dpro-NTN), and a pro-region deleted chimeric NTN (IgSP-NTN). The latter DNA sequence contains an intron. See text for further details. (B)
NTN Western blot of lysates and conditioned medium from transfected HEK293 cells. Arrows indicate the expected position of bands representing wt pro-
NTN, pro(GDNF)-NTN and mature NTN, respectively. Note that the standard is NTN-His which has a slightly higher molecular weight than wt NTN. (C)
GFRa 2 binding activity of NTN in conditioned medium from the four cell lines transfected with the NTN constructs. The GFRa 2 binding activities shown in
samples were calculated using a standard curve of recombinant NTN. Relative values are shown with activities from cells transfected with the wt NTN
construct set to 1. (D) NTN Western blot of NTN from conditioned HEK293 medium from bound to GFRa 2. Arrow indicates the expected position for band
representing mature NTN. (E) NTN sandwich ELISA on conditioned medium from the four cell lines transfected with the NTN constructs. Data in panels (C)
and (E) are expressed as mean T SEM (n = 3) from a representative experiment and * indicates a significant difference from cells transfected with the wt
construct (P < 0.05, Kruskal–Wallis one-way analysis on ranks followed by Student–Newman–Keuls Method).
L. Fjord-Larsen et al. / Experimental Neurology 195 (2005) 49–6054
conditioned medium from the transfected cells (2- to 8-fold
higher NTN levels for IgSP-NTN than Dpro-NTN, depend-
ing on cell line).
In vivo neuroprotection by IgSP-NTN in the rat
6-hydroxydopamine lesion model
Next, we wanted to investigate whether the IgSP-NTN
construct would lead to sustained secretion of active NTN in
vivo. We generated recombinant lentiviral vectors encoding
the wt NTN (rLV-wt NTN) and the IgSP-NTN (rLV-IgSP-
NTN), and tested if they were able to provide neuro-
protection in an animal model of PD where NTN protein
injections previously had been shown to prevent loss of DA
neurons (Horger et al., 1998; Rosenblad et al., 1999).
Vectors encoding green fluorescent protein (GFP; rLV-GFP)
or GDNF (rLV-GDNF) were used as negative and positive
control vectors, respectively.
Inspection of GFP-immunostained sections from rLV-
GFP-treated animals showed a column of transduced cells,
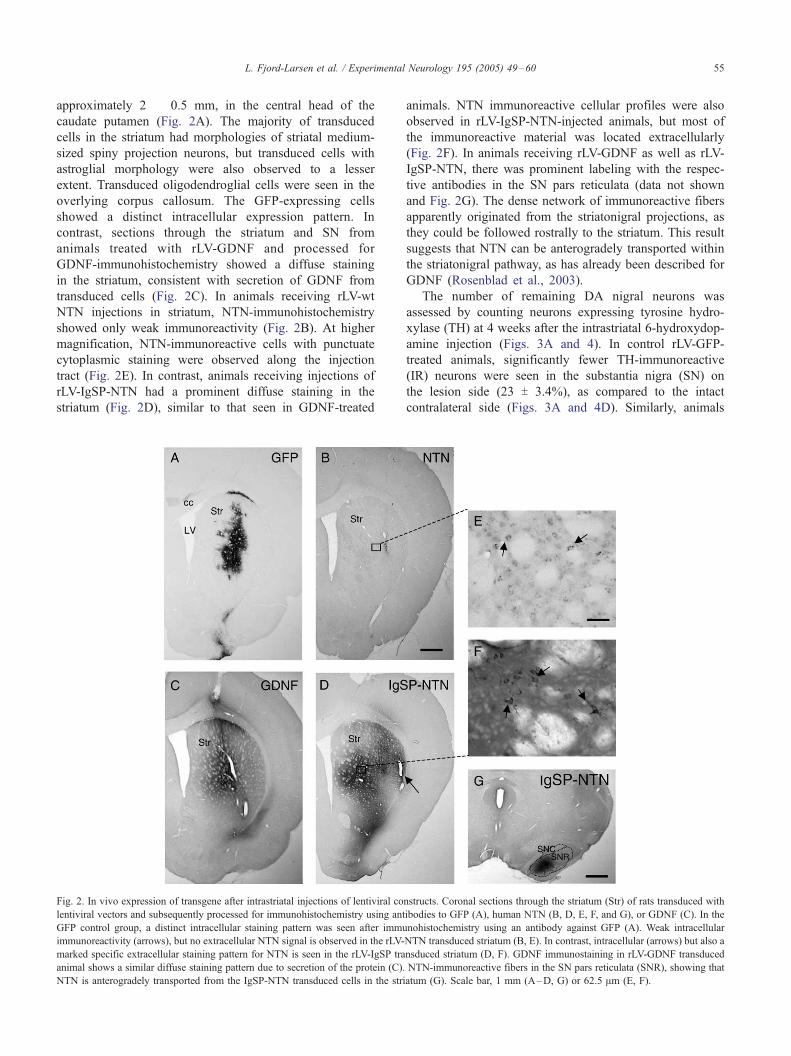
L. Fjord-Larsen et al. / Experimental Neurology 195 (2005) 49–60 55
approximately 2 � 0.5 mm, in the central head of the
caudate putamen (Fig. 2A). The majority of transduced
cells in the striatum had morphologies of striatal medium-
sized spiny projection neurons, but transduced cells with
astroglial morphology were also observed to a lesser
extent. Transduced oligodendroglial cells were seen in the
overlying corpus callosum. The GFP-expressing cells
showed a distinct intracellular expression pattern. In
contrast, sections through the striatum and SN from
animals treated with rLV-GDNF and processed for
GDNF-immunohistochemistry showed a diffuse staining
in the striatum, consistent with secretion of GDNF from
transduced cells (Fig. 2C). In animals receiving rLV-wt
NTN injections in striatum, NTN-immunohistochemistry
showed only weak immunoreactivity (Fig. 2B). At higher
magnification, NTN-immunoreactive cells with punctuate
cytoplasmic staining were observed along the injection
tract (Fig. 2E). In contrast, animals receiving injections of
rLV-IgSP-NTN had a prominent diffuse staining in the
striatum (Fig. 2D), similar to that seen in GDNF-treated
Fig. 2. In vivo expression of transgene after intrastriatal injections of lentiviral co
lentiviral vectors and subsequently processed for immunohistochemistry using ant
GFP control group, a distinct intracellular staining pattern was seen after immu
immunoreactivity (arrows), but no extracellular NTN signal is observed in the rLV-
marked specific extracellular staining pattern for NTN is seen in the rLV-IgSP tra
animal shows a similar diffuse staining pattern due to secretion of the protein (C).
NTN is anterogradely transported from the IgSP-NTN transduced cells in the stri
animals. NTN immunoreactive cellular profiles were also
observed in rLV-IgSP-NTN-injected animals, but most of
the immunoreactive material was located extracellularly
(Fig. 2F). In animals receiving rLV-GDNF as well as rLV-
IgSP-NTN, there was prominent labeling with the respec-
tive antibodies in the SN pars reticulata (data not shown
and Fig. 2G). The dense network of immunoreactive fibers
apparently originated from the striatonigral projections, as
they could be followed rostrally to the striatum. This result
suggests that NTN can be anterogradely transported within
the striatonigral pathway, as has already been described for
GDNF (Rosenblad et al., 2003).
The number of remaining DA nigral neurons was
assessed by counting neurons expressing tyrosine hydro-
xylase (TH) at 4 weeks after the intrastriatal 6-hydroxydop-
amine injection (Figs. 3A and 4). In control rLV-GFP-
treated animals, significantly fewer TH-immunoreactive
(IR) neurons were seen in the substantia nigra (SN) on
the lesion side (23 T 3.4%), as compared to the intact
contralateral side (Figs. 3A and 4D). Similarly, animals
nstructs. Coronal sections through the striatum (Str) of rats transduced with
ibodies to GFP (A), human NTN (B, D, E, F, and G), or GDNF (C). In the
nohistochemistry using an antibody against GFP (A). Weak intracellular
NTN transduced striatum (B, E). In contrast, intracellular (arrows) but also a
nsduced striatum (D, F). GDNF immunostaining in rLV-GDNF transduced
NTN-immunoreactive fibers in the SN pars reticulata (SNR), showing that
atum (G). Scale bar, 1 mm (A–D, G) or 62.5 Am (E, F).

Fig. 3. Neuroprotection of nigral DA neurons by rLV-IgSP-NTN. (A) The
number of nigral neurons expressing TH in the lesion side compared to the
intact side in the four different treatment groups. (B) The percentage of
VMAT-immunoreactive nigral neurons in lesion side compared with the
intact side. Data are expressed as mean T SEM (n = 5–7) and * indicates a
significant difference from the GFP group (P < 0.05, one-way ANOVA,
Dunnett’s Method).
L. Fjord-Larsen et al. / Experimental Neurology 195 (2005) 49–6056
receiving rLV-NTN (Figs. 3A and 4B) showed a marked
lesion-induced reduction in TH-IR neurons (30 T 7.7%
remaining). In contrast, the rLV-IgSP-NTN-treated group
had a significantly higher percentage of TH-IR neurons on
the lesion side (91 T 1.2%; P < 0.01) (Figs. 3A and 4C),
which was indistinguishable from that observed in the
group receiving GDNF treatment (86 T 3.2%; P < 0.01)
(Figs. 3A and 4H).
To confirm that the differences in the number of TH-IR
nigral neurons were due to neuroprotection rather than
regulation of TH enzyme, we quantified in adjacent sections
the number of neurons expressing another DA marker,
vesicular monoamine transporter (VMAT), since VMAT has
been shown to be less prone to regulation by neurotrophic
factors (Georgievska et al., 2002a; Kirik et al., 1998;
Rosenblad et al., 2003). As shown in Fig. 3B, transduction
with rLV-IgSP-NTN or rLV-GDNF significantly preserved
the number of VMAT-IR neurons in SN on the lesion side
(75.2 T 6.8% and 59.9 T 4.2% of that in the intact side,
respectively) compared with rLV-GFP- or rLV-NTN-treated
animals (15.5 T 2.4% and 24.9 T 6.4%, respectively),
corroborating the results seen with TH-staining.
In addition to the protection of TH-IR and VMAT-IR
neurons in the SN, it was noticeable that in specimens from
rLV-IgSP-NTN-treated animals, the TH staining intensity
was reduced in many remaining SN neurons (Fig. 4G) as
compared to TH-IR neurons on the intact side (Fig. 4E), or
lesioned side of rLV-GFP- or rLV-wt NTN-treated animals
(Fig. 4F). Reduced TH staining intensity was also observed
following treatment with GDNF, consistent with previous
reports (Georgievska et al., 2002a). In the rLV-GDNF and
rLV-IgSP-NTN groups, the percentage of cells with down-
regulated TH expression was significantly higher (79.4 T 3.5and 74.5 T 3.8, respectively) than in the rLV-NTN (16.5 T8.7) and rLV-GFP (13.9 T 7.3) groups (P < 0.001, one-way
ANOVA followed by all pairwise multiple comparison
procedures, Student–Newman–Keuls Method).
Inspection of sections through the striatum processed for
TH-immunohistochemistry showed that in all groups, the
central and lateral caudate putamen was devoid of TH-IR
fibers on the side injected with 6-hydroxydopamine.
Densitometric quantification of TH-IR innervation on the
lesioned side showed that 15–25% remained at 4 weeks
(Fig. 5). This is in agreement with earlier studies in the
intrastriatal 6-hydroxydopamine lesion model (Georgievska
et al., 2002a; Kirik et al., 2001; Rosenblad et al., 2003)
showing that recovery of TH-IR striatal innervation usually
takes longer than 4 weeks post-lesion to develop. The
reduction of TH-IR striatal innervation did not differ
between the groups. Consistently, amphetamine-induced
rotation, which can be used as a sensitive measure of
dopamine denervation in the striatum (Kirik et al., 2000),
showed no significant difference in the number of ipsilateral
turns between any of the treatment groups at 4 weeks post-
lesion (ranging from 4.5 T 1.5 to 13.4 T 3.3 net ipsilateral
turns/min; P > 0.05 two-way repeated measures ANOVA).
Amphetamine-induced turning assessed at 10 days after
viral transduction, but before the 6-hydroxydopamine
lesion, showed a slight but non-significant contralateral
turning bias in the IgSP-NTN (3.3 T 1.5) and GDNF groups
(1.9 T 1.3), compared to the rLV-GFP or rLV-NTN groups
(0.1 T 0.9 and �0.1 T 1.6, respectively), consistent with an
upregulation of the DA function on the IgSP-NTN
transduced side similar to what has been reported previ-
ously to occur after GDNF treatment (Georgievska et al.,
2002a,b).
Discussion
We investigated in vivo gene therapy to deliver NTN in a
PD animal model but found that human wt NTN, unlike its
GDNF counterpart, is inadequately expressed with this
approach. We therefore examined the role of the pre-pro-
region of human NTN in the secretion of active NTN of de

Fig. 4. Coronal sections through SN of GFP-, NTN-, IgSP-NTN-, and GDNF-treated animals. On the intact side, many TH-immunoreactive cells can be seen
(A). On the side subject to a 6-hydroxydopamine lesion, the number of remaining TH-immunoreactive neuronal profiles is significantly reduced in wt NTN- or
GFP-treated animals (B, D). By contrast, more TH-immunoreactive neurons remain in animals receiving GDNF (E) or IgSP-NTN treatment (C). Note the
reduced TH staining intensity in GDNF- and IgSP-NTN-treated animals. Higher magnification of TH-IR cells in intact side (E), lesion side of IgSP-NTN-
treated animal (G), and lesion side of wt NTN-treated animal (F). Black arrows point at neurons with downregulated TH expression in IgSP-NTN-treated
animal and white arrow points at neuron with normal TH expression. Scale bar, 1 mm (A–D, H) or 25 Am (E–G).
L. Fjord-Larsen et al. / Experimental Neurology 195 (2005) 49–60 57
novo synthesized NTN in mammalian cells by transfecting
constructs encoding modified NTN proteins into two human
and two rodent cell lines. Furthermore, the in vivo neuro-
protective effect of modified NTN protein was addressed in
the partial 6-hydroxydopamine lesion model using a gene
therapy approach.
As biologically active human GDNF is readily secreted
from various cell types after transfection, an NTN construct
encoding a chimeric protein with the pre-pro-part derived
from GDNF was generated (ppG-NTN). In addition, as the
processing of the pro-forms seemed impaired, two addi-
tional constructs encoding NTN proteins with the pro-part
deleted but with different signal peptides were made, one
containing the wt NTN signal peptide (Dpro-NTN) and one
with the IgSP (IgSP-NTN). We could not detect any
processed NTN in cell lysates or conditioned medium from
cells transfected with wt NTN or ppG-NTN constructs by
Western blot analysis. These results indicate that intra-
cellular cleavage of the pro-region is severely compromised
in the tested cell lines. Furthermore, we found that the

Fig. 5. Striatal DA fiber density measurements. The four experimental
groups had a comparable initial lesion size, and there was no significant
recovery in any group after 4 weeks. Striatal DA innervation was assessed
by measuring optical density of the striatum at three rostrocaudal levels in
sections stained for TH.
L. Fjord-Larsen et al. / Experimental Neurology 195 (2005) 49–6058
cleavage of the pro-peptide is critical for the ability of
NTN to form a signaling complex, demonstrated by the very
low binding activity to the GFRa1 or GFRa2 receptors with
the same constructs (Figs. 1B–D). The very low (but
detectable) level of binding is most likely explained by the
presence of mature NTN below the Western blot detection
limit (<1.25 ng/ml). This is also supported by the lack
of capture of the pro-forms of NTN on the GFRa2 plate
(Fig. 1D).
In contrast, a protein of the expected size of mature
NTN was detected in Western blot analysis in media
collected from cells transfected with either the Dpro-NTN
and IgSP-NTN constructs. Furthermore, high levels of
immunoreactive NTN and GFRa1 and GFRa2 binding
activities were also detected in these media. These results
indicate that the pro-sequence of NTN is not necessary for
correct protein folding in contrast to other neurotrophic
factors such as NGF, TGFh, and Activin in which the pro-
region has been shown to aid the folding and disulphide
bond formation (Gray and Mason, 1990; Suter et al.,
1991). In the case of NGF, deletion of the pro-region
results in undetectable expression, possibly due to insta-
bility and degradation of the protein in the endoplasmatic
reticulum (Suter et al., 1991). Unlike NGF, the pro-region
of NTN appears to be dispensable for intracellular stability
in our experiments.
Data obtained from the NTN sandwich ELISA indicated
significantly higher production levels (2- to 8-fold depend-
ing on cell line) of mature NTN in cells transfected with
the IgSP-NTN construct compared with Dpro-NTN (Fig.
1E) consistent with what was observed from the Western
blot analyses (Figs. 1A and D). The reason for this
increased production is unclear. Hypothetically, the IgSP
sequence could influence the stability of mRNA due to the
presence of the inherent IgSP intron. Insertion of a
heterologous intron has previously been shown to improve
expression (Palmiter et al., 1991). Other possible explan-
ations include the promotion of 3Vend formation, increased
mRNA stability in the nucleus, or improved mRNA export
to the cytoplasm (Nesic et al., 1993; Ryu and Mertz,
1989).
In this report, we provide the first demonstration of in
vivo gene delivery of NTN as a therapeutic option for PD.
In accordance with the results obtained with the cultured
cells, we observed only weak NTN immunoreactivity and
no significant neuroprotective effect in vivo when injecting
rats with lentivirus encoding human wt NTN. These results
indicate that transduction with wt NTN in rat striatum most
likely leads to expression of pro-NTN, which are poorly
recognized by the NTN antibodies as not denatured by the
fixation of tissues. These data are consistent with the in
vitro results that indicate that the native pro-NTN could not
be detected by ELISA but could be detected in a denaturing
Western blot. In contrast, a potent neuroprotective effect
was seen and a diffuse staining was observed by NTN-
immunohistochemistry after in vivo transduction with the
IgSP-NTN construct consistent with the efficient secretion
of bioactive mature NTN from the transduced cells. The
potency and efficacy of NTN in the current study is in
accordance with results obtained by Horger et al. (1998)
who showed that the effect of recombinant NTN on
survival of DA neurons in the 6-hydroxydopamine lesion
model was similar to that of GDNF when administered at
the level of SN.
The primary focus of the current study was to investigate
whether the IgSP-NTN construct would produce a molecule
with biological activity in vivo. We observed comparable
effects of our modified NTN construct and GDNF on DA
cell survival and regulation of TH expression after 4 weeks.
However, longer observations times need to be added in
order to conclude whether the NTN produced by our
modified construct is as efficient as GDNF for recovery of
striatal DA fiber innervation. Furthermore, GDNF and NTN
may show different therapeutic profiles, due to the fact that
the preferred GFRa receptors for NTN and GDNF are
differentially expressed (Akerud et al., 1999; Jing et al.,
1997; Widenfalk et al., 1997; Yu et al., 1998). For example,
in another study, GDNF delivered intracerebroventricularly
via osmotic minipumps in rats resulted in side effects,
mainly weight-loss and allodynia, whereas NTN treatment
did not lead to these symptoms (Hoane et al., 1999).
Moreover, long-term overexpression of GDNF in rat
striatum has been shown to induce aberrant sprouting of
nigrostriatal fibers (Georgievska et al., 2002b). The con-
sequence of this phenomenon is unclear, but it might be
detrimental for recovery in spontaneous motor behavior.
Interestingly, Akerud et al. (1999) showed that NTN, in
contrast to GDNF, did not induce sprouting and hyper-
trophy of adult DA neurons, though both GDNF and NTN
showed survival promoting effects. In another rat model of
PD made by medial forebrain bundle axotomy, an initial
compensatory sprouting effect induced by GDNF but not

L. Fjord-Larsen et al. / Experimental Neurology 195 (2005) 49–60 59
by NTN has been shown (Tseng et al., 1997, 1998).
Experiments to evaluate possible differences in regeneration
and behavior between NTN delivered by our construct and
GDNF are under consideration in our lab. In the described
experiments, we have used human NTN in a rodent setting.
The reason for this is that we have focused on developing
gene therapeutic application for the treatment of Parkin-
son’s disease patients and need to optimize the expression
of human NTN cDNA. Human, mouse, and rat NTN have
identical consensus recognition sites for furin-like proteases
(RRAR). In contrast, the amino acids after the cleavage site
differ between human (ARLGAR) and rat/mouse (P—GSR).
It could be speculated that these amino acids would affect the
processing of NTN. However, as we found that rodent NTN
cDNAwas also expressed as the pro-form in human as well as
rat cells in vitro (data not shown), we find it plausible that we
would have obtained a similar result by using rodent wt NTN
in vivo.
It is possible that the processing of NTN is part of an
important regulatory step for the availability of active
protein, as pro-NTN is not biologically active in the same
way as mature NTN is. The transfected cell lines used in
vitro and the transduced cells in vivo do not normally
express NTN and might not posses the processing apparatus
for NTN. Alternatively, it could be speculated that addi-
tional extracellular signals are necessary for the activation of
relevant proteases. Such signals might be present when
active NTN is needed, e.g., during development.
In summary, we show that deletion of the pro-region of
human NTN increases the fraction of biologically active
NTN. Replacing the signal peptide sequence from wt NTN
in Dpro-NTN with the heterologous IgSP sequence further
increases secretion of active NTN. The biological activity
of neurotrophic factors can be regulated at several differ-
ent levels. Our results indicate that proteolytic processing
of the NTN pro-region is an important regulatory step,
which should be taken into account when overexpressing
neurotrophic factors from transgenic constructs. We
obtained a high and stable secretion level of mature
bioactive NTN when using the IgSP-NTN construct. The
results from studies in cultured cells were reproducible in
vivo, where intrastriatal lentiviral delivery of IgSP-NTN
resulted in protection of adult DA neurons at a level
comparable to GDNF, while wt NTN had no effect. These
results indicate that in vivo gene therapy is possible with a
modified NTN gene transfer and is a potential therapy for
PD.
Acknowledgments
The authors would like to thank Hanne Fosmark and
Lise Moberg Fitting for excellent technical assistance. This
study was supported by the Fifth Framework EU Program
Quality of Life and Management of Living Resources,
QLG3-CT-2002-01000.
References
Akerud, P., Alberch, J., Eketjall, S., Wagner, J., Arenas, E., 1999.
Differential effects of glial cell line-derived neurotrophic factor and
neurturin on developing and adult substantia nigra dopaminergic
neurons. J. Neurochem. 73, 70–78.
Baloh, R.H., Enomoto, H., Johnson, Jr., E.M., Milbrandt, J., 2000. The
GDNF family ligands and receptors—Implications for neural develop-
ment. Curr. Opin. Neurobiol. 10, 103–110.
Bensadoun, J.C., Deglon, N., Tseng, J.L., Ridet, J.L., Zurn, A.D.,
Aebischer, P., 2000. Lentiviral vectors as a gene delivery system in
the mouse midbrain: cellular and behavioral improvements in a
6-OHDA model of Parkinson’s disease using GDNF. Exp. Neurol.
164, 15–24.
Bilang-bleuel, A., Revah, F., Colin, P., Locquet, I., Robert, J.J., Mallet,
J., Horellou, P., 1997. Intrastriatal injection of an adenoviral vector
expressing glial-cell-line-derived neurotrophic factor prevents dop-
aminergic neuron degeneration and behavioral impairment in a rat
model of Parkinson disease. Proc. Natl. Acad. Sci. U. S. A. 94,
8818–8823.
Cacalano, G., Farinas, I., Wang, L.C., Hagler, K., Forgie, A., Moore, M.,
Armanini, M., Phillips, H., Ryan, A.M., Reichardt, L.F., Hynes, M.,
Davies, A., Rosenthal, A., 1998. GFRalpha1 is an essential receptor
component for GDNF in the developing nervous system and kidney.
Neuron 21, 53–62.
Connor, B., Kozlowski, D.A., Unnerstall, J.R., Elsworth, J.D., Tillerson,
J.L., Schallert, T., Bohn, M.C., 2001. Glial cell line-derived neuro-
trophic factor (GDNF) gene delivery protects dopaminergic terminals
from degeneration. Exp. Neurol. 169, 83–95.
Dunn, K.C., Aotaki-Keen, A.E., Putkey, F.R., Hjelmeland, L.M., 1996.
ARPE-19, a human retinal pigment epithelial cell line with differ-
entiated properties. Exp. Eye Res. 62, 155–169.
Georgievska, B., Kirik, D., Bjorklund, A., 2002a. Aberrant sprouting and
downregulation of tyrosine hydroxylase in lesioned nigrostriatal
dopamine neurons induced by long-lasting overexpression of glial cell
line derived neurotrophic factor in the striatum by lentiviral gene
transfer. Exp. Neurol. 177, 461–474.
Georgievska, B., Kirik, D., Rosenblad, C., Lundberg, C., Bjorklund, A.,
2002b. Neuroprotection in the rat Parkinson model by intrastriatal
GDNF gene transfer using a lentiviral vector. NeuroReport 13, 75–82.
Georgievska, B., Kirik, D., Bjorklund, A., 2004. Overexpression of
glial cell line-derived neurotrophic factor using a lentiviral vector
induces time- and dose-dependent downregulation of tyrosine
hydroxylase in the intact nigrostriatal dopamine system. J. Neurosci.
24, 6437–6445.
Golden, J.P., Baloh, R.H., Kotzbauer, P.T., Lampe, P.A., Osborne, P.A.,
Milbrandt, J., Johnson, Jr., E.M., 1998. Expression of neurturin, GDNF,
and their receptors in the adult mouse CNS. J. Comp. Neurol. 398,
139–150.
Gray, A.M., Mason, A.J., 1990. Requirement for activin A and trans-
forming growth factor-beta 1 pro-regions in homodimer assembly.
Science 247, 1328–1330.
Hoane, M.R., Gulwadi, A.G., Morrison, S., Hovanesian, G., Lindner, M.D.,
Tao, W., 1999. Differential in vivo effects of neurturin and glial cell-
line-derived neurotrophic factor. Exp. Neurol. 160, 235–243.
Horger, B.A., Nishimura, M.C., Armanini, M.P., Wang, L.C., Poulsen,
K.T., Rosenblad, C., Kirik, D., Moffat, B., Simmons, L., Johnson, Jr.,
E., Milbrandt, J., Rosenthal, A., Bjorklund, A., Vandlen, R.A., Hynes,
M.A., Phillips, H.S., 1998. Neurturin exerts potent actions on survival
and function of midbrain dopaminergic neurons. J. Neurosci. 18,
4929–4937.
Jing, S., Yu, Y., Fang, M., Hu, Z., Holst, P.L., Boone, T., Delaney, J.,
Schultz, H., Zhou, R., Fox, G.M., 1997. GFRalpha-2 and GFRalpha-3
are two new receptors for ligands of the GDNF family. J. Biol. Chem.
272, 33111–33117.
Kirik, D., Rosenblad, C., Bjorklund, A., 1998. Characterization of
behavioral and neurodegenerative changes following partial lesions of

L. Fjord-Larsen et al. / Experimental Neurology 195 (2005) 49–6060
the nigrostriatal dopamine system induced by intrastriatal 6-hydro-
xydopamine in the rat. Exp. Neurol. 152, 259–277.
Kirik, D., Rosenblad, C., Bjorklund, A., Mandel, R.J., 2000. Long-term
rAAV-mediated gene transfer of GDNF in the rat Parkinson’s model:
intrastriatal but not intranigral transduction promotes functional
regeneration in the lesioned nigrostriatal system. J. Neurosci. 20,
4686–4700.
Kirik, D., Georgievska, B., Rosenblad, C., Bjorklund, A., 2001. Delayed
infusion of GDNF promotes recovery of motor function in the
partial lesion model of Parkinson’s disease. Eur. J. Neurosci. 13,
1589–1599.
Klein, R.D., Sherman, D., Ho, W.H., Stone, D., Bennett, G.L., Moffat, B.,
Vandlen, R., Simmons, L., Gu, Q., Hongo, J.A., Devaux, B., Poulsen,
K., Armanini, M., Nozaki, C., Asai, N., Goddard, A., Phillips, H.,
Henderson, C.E., Takahashi, M., Rosenthal, A., 1997. A GPI-linked
protein that interacts with Ret to form a candidate neurturin receptor
[published erratum appears in Nature 1998 Mar 12;392(6672):210].
Nature 387, 717–721.
Kordower, J.H., 2003. In vivo gene delivery of glial cell line-derived
neurotrophic factor for Parkinson’s disease. Ann. Neurol. 53 (Suppl 3),
S120–S132.
Lapchak, P.A., Araujo, D.M., Hilt, D.C., Sheng, J., Jiao, S., 1997.
Adenoviral vector-mediated GDNF gene therapy in a rodent lesion
model of late stage Parkinson’s disease. Brain Res. 777, 153–160.
Mandel, R.J., Spratt, S.K., Snyder, R.O., Leff, S.E., 1997. Midbrain
injection of recombinant adeno-associated virus encoding rat glial cell
line-derived neurotrophic factor protects nigral neurons in a progressive
6-hydroxydopamine-induced degeneration model of Parkinson’s disease
in rats. Proc. Natl. Acad. Sci. U. S. A. 94, 14083–14088.
Nesic, D., Cheng, J., Maquat, L.E., 1993. Sequences within the last intron
function in RNA 3V-end formation in cultured cells. Mol. Cell. Biol. 13,
3359–3369.
Palmiter, R.D., Sandgren, E.P., Avarbock, M.R., Allen, D.D., Brinster, R.L.,
1991. Heterologous introns can enhance expression of transgenes in
mice. Proc. Natl. Acad. Sci. U. S. A. 88, 478–482.
Paxinos, G., Watson, C., 1997. The Rat Brain in Stereotaxic Coordinates.
Academic Press, San Diego.
Renfranz, P.J., Cunningham, M.G., Mckay, R.D., 1991. Region-specific
differentiation of the hippocampal stem cell line HiB5 upon implanta-
tion into the developing mammalian brain. Cell 66, 713–729.
Rosenblad, C., Kirik, D., Devaux, B., Moffat, B., Phillips, H.S., Bjorklund,
A., 1999. Protection and regeneration of nigral dopaminergic neurons
by neurturin or GDNF in a partial lesion model of Parkinson’s disease
after administration into the striatum or the lateral ventricle. Eur. J.
Neurosci. 11, 1554–1566.
Rosenblad, C., Gronborg, M., Hansen, C., Blom, N., Meyer, M., Johansen,
J., Dago, L., Kirik, D., Patel, U.A., Lundberg, C., Trono, D., Bjorklund,
A., Johansen, T.E., 2000. In vivo protection of nigral dopamine neurons
by lentiviral gene transfer of the novel GDNF-family member
neublastin/artemin. Mol. Cell. Neurosci. 15, 199–214.
Rosenblad, C., Georgievska, B., Kirik, D., 2003. Long-term striatal
overexpression of GDNF selectively downregulates tyrosine hydro-
xylase in the intact nigrostriatal dopamine system. Eur. J. Neurosci. 17,
260–270.
Ryu, W.S., Mertz, J.E., 1989. Simian virus 40 late transcripts lacking
excisable intervening sequences are defective in both stability in the
nucleus and transport to the cytoplasm. J. Virol. 63, 4386–4394.
Sanicola, M., Hession, C., Worley, D., Carmillo, P., Ehrenfels, C., Walus, L.,
Robinson, S., Jaworski, G., Wei, H., Tizard, R., Whitty, A., Pepinsky,
R.B., Cate, R.L., 1997. Glial cell line-derived neurotrophic factor-
dependent RET activation can be mediated by two different cell-surface
accessory proteins. Proc. Natl. Acad. Sci. U. S. A. 94, 6238–6243.
Sauer, H., Oertel, W.H., 1994. Progressive degeneration of nigrostriatal
dopamine neurons following intrastriatal terminal lesions with
6-hydroxydopamine: a combined retrograde tracing and immunocyto-
chemical study in the rat. Neuroscience 59, 401–415.
Suter, U., Heymach, Jr., J.V., Shooter, E.M., 1991. Two conserved domains
in the NGF propeptide are necessary and sufficient for the biosynthesis
of correctly processed and biologically active NGF. EMBO J. 10,
2395–2400.
Tseng, J.L., Baetge, E.E., Zurn, A.D., Aebischer, P., 1997. GDNF reduces
drug-induced rotational behavior after medial forebrain bundle tran-
section by a mechanism not involving striatal dopamine. J. Neurosci.
17, 325–333.
Tseng, J.L., Bruhn, S.L., Zurn, A.D., Aebischer, P., 1998. Neurturin protects
dopaminergic neurons following medial forebrain bundle axotomy.
NeuroReport 9, 1817–1822.
Widenfalk, J., Nosrat, C., Tomac, A., Westphal, H., Hoffer, B., Olson, L.,
1997. Neurturin and glial cell line-derived neurotrophic factor receptor-
beta (GDNFR-beta), novel proteins related to GDNF and GDNFR-alpha
with specific cellular patterns of expression suggesting roles in the
developing and adult nervous system and in peripheral organs.
J. Neurosci. 17, 8506–8519.
Yu, T., Scully, S., Yu, Y., Fox, G.M., Jing, S., Zhou, R., 1998. Expression of
GDNF family receptor components during development: implications in
the mechanisms of interaction. J. Neurosci. 18, 4684–4696.
Zufferey, R., Nagy, D., Mandel, R.J., Naldini, L., Trono, D., 1997. Multiply
attenuated lentiviral vector achieves efficient gene delivery in vivo. Nat.
Biotechnol. 15, 871–875.
Zufferey, R., Dull, T., Mandel, R.J., Bukovsky, A., Quiroz, D., Naldini, L.,
Trono, D., 1998. Self-inactivating lentivirus vector for safe and efficient
in vivo gene delivery. J. Virol. 72, 9873–9880.
Zufferey, R., Donello, J.E., Trono, D., Hope, T.J., 1999. Woodchuck
hepatitis virus posttranscriptional regulatory element enhances expres-
sion of transgenes delivered by retroviral vectors. J. Virol. 73,
2886–2892.




















