
Hemiparkinsonian rats rotate toward the side with the weaker dopaminergic neurotransmission
9
Available online at www.sciencedirect.com Behavioural Brain Research 189 (2008) 364–372 Research report Hemiparkinsonian rats rotate toward the side with the weaker dopaminergic neurotransmission Claudio Da Cunha a,∗ , Evellyn Claudia Wietzikoski a , Marcelo Machado Ferro b , Glaucia Regina Martinez b , Maria Aparecida Barbato Fraz˜ ao Vital a , D´ ebora Hip ´ olide c , Sergio Tufik c , Newton Sabino Canteras d a Laborat´ orio de Fisiologia e Farmacologia do Sistema Nervoso Central, Departamento de Farmacologia, UFPR, C.P. 19.031, 81.531-980 Curitiba PR, Brazil b Departamento de Bioqu´ ımica e Biologia Molecular, UFPR, Curitiba, Brazil c Departamento de Psicobiologia, UNIFESP, S˜ ao Paulo, Brazil d Departamento de Anatomia, Instituto de Ciˆ encias Biom´ edicas-3, USP, S˜ ao Paulo, Brazil Received 4 December 2007; received in revised form 23 January 2008; accepted 24 January 2008 Available online 5 February 2008 Abstract Rats with unilateral lesion of the substantia nigra pars compacta (SNpc) have been used as a model of Parkinson’s disease. Depending on the lesion protocol and on the drug challenge, these rats rotate in opposite directions. The aim of the present study was to propose a model to explain how critical factors determine the direction of these turns. Unilateral lesion of the SNpc was induced with 6-hydroxydopamine (6-OHDA) or 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP). Separate analysis showed that neither the type of neurotoxin nor the site of lesion along the nigrostriatal pathway was able to predict the direction of the turns these rats made after they were challenged with apomorphine. However, the combination of these two factors determined the magnitude of the lesion estimated by tyrosine-hydroxylase immunohistochemistry and HPLC-ED measurement of striatal dopamine. Very small lesions did not cause turns, medium-size lesions caused ipsiversive turns, and large lesions caused contraversive turns. Large-size SNpc lesions resulted in an increased binding of [ 3 H]raclopride to D2 receptors, while medium-size lesions reduced the binding of [ 3 H]SCH-23390 D1 receptors in the ipsilateral striatum. These results are coherent with the model proposing that after challenged with a dopamine receptor agonist, unilaterally SNpc-lesioned rats rotate toward the side with the weaker activation of dopamine receptors. This activation is weaker on the lesioned side in animals with small SNpc lesions due to the loss of dopamine, but stronger in animals with large lesions due to dopamine receptor supersensitivity. © 2008 Elsevier B.V. All rights reserved. Keywords: Substantia nigra pars compacta; Turning behaviour; Dopamine; Basal ganglia; Dopamine receptor; Parkinson’s disease; MPTP; 6-Hydroxydopamine 1. Introduction In 1968, Ungerstedt proposed that the turning behaviour of rats with unilateral lesion of the substantia nigra pars compacta (SNpc) induced by intracerebral infusion of 6-hydroxydopamine (6-OHDA) which are challenged with dopaminergic drugs can be used as a model of Parkinson’s disease (PD). Since then, it has been one of the most popular models to study drugs poten- tially useful to treat this disease [4,16,41,49]. However, there is no consensual explanation for the direction of the turns made ∗ Corresponding author. Tel.: +55 41 3361 1717; fax: +55 41 3266 2042. E-mail address: [email protected] (C. Da Cunha). by these rats. Most studies have shown that rats with unilat- eral lesions induced by 6-OHDA present a contraversive turning behaviour (in the direction opposite to the lesioned side) when challenged with apomorphine. However, ipsiversive or few turns in both directions have also been reported in animals with uni- lateral lesions induced by 6-OHDA [2,7,15,35,38,44,46], and other neurotoxins (e.g., rotenone, MPP + ) [22,24,42]. Turning behaviour seems to be caused by a misbalance between the right and left side drive to initiate movements due to an asymmetrical modulation of midbrain dopaminergic neurons on corticostriatal synapses [12,32]. The site of infusion of 6-OHDA has been reported to be crit- ical to explain the direction of the turns [38,40,42]. The medial forebrain bundle (MFB) has been indicated as the infusion 0166-4328/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.bbr.2008.01.012
Transcript of Hemiparkinsonian rats rotate toward the side with the weaker dopaminergic neurotransmission

A
lh1tcmctwad©
K
1
r((bhtn
0d
Available online at www.sciencedirect.com
Behavioural Brain Research 189 (2008) 364–372
Research report
Hemiparkinsonian rats rotate toward the side with theweaker dopaminergic neurotransmission
Claudio Da Cunha a,∗, Evellyn Claudia Wietzikoski a, Marcelo Machado Ferro b,Glaucia Regina Martinez b, Maria Aparecida Barbato Frazao Vital a,
Debora Hipolide c, Sergio Tufik c, Newton Sabino Canteras d
a Laboratorio de Fisiologia e Farmacologia do Sistema Nervoso Central, Departamento de Farmacologia,UFPR, C.P. 19.031, 81.531-980 Curitiba PR, Brazil
b Departamento de Bioquımica e Biologia Molecular, UFPR, Curitiba, Brazilc Departamento de Psicobiologia, UNIFESP, Sao Paulo, Brazil
d Departamento de Anatomia, Instituto de Ciencias Biomedicas-3, USP, Sao Paulo, Brazil
Received 4 December 2007; received in revised form 23 January 2008; accepted 24 January 2008Available online 5 February 2008
bstract
Rats with unilateral lesion of the substantia nigra pars compacta (SNpc) have been used as a model of Parkinson’s disease. Depending on theesion protocol and on the drug challenge, these rats rotate in opposite directions. The aim of the present study was to propose a model to explainow critical factors determine the direction of these turns. Unilateral lesion of the SNpc was induced with 6-hydroxydopamine (6-OHDA) or-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP). Separate analysis showed that neither the type of neurotoxin nor the site of lesion alonghe nigrostriatal pathway was able to predict the direction of the turns these rats made after they were challenged with apomorphine. However, theombination of these two factors determined the magnitude of the lesion estimated by tyrosine-hydroxylase immunohistochemistry and HPLC-EDeasurement of striatal dopamine. Very small lesions did not cause turns, medium-size lesions caused ipsiversive turns, and large lesions caused
ontraversive turns. Large-size SNpc lesions resulted in an increased binding of [3H]raclopride to D2 receptors, while medium-size lesions reduced3
he binding of [ H]SCH-23390 D1 receptors in the ipsilateral striatum. These results are coherent with the model proposing that after challengedith a dopamine receptor agonist, unilaterally SNpc-lesioned rats rotate toward the side with the weaker activation of dopamine receptors. Thisctivation is weaker on the lesioned side in animals with small SNpc lesions due to the loss of dopamine, but stronger in animals with large lesionsue to dopamine receptor supersensitivity.
2008 Elsevier B.V. All rights reserved.
al gan
bebcilo
eywords: Substantia nigra pars compacta; Turning behaviour; Dopamine; Bas
. Introduction
In 1968, Ungerstedt proposed that the turning behaviour ofats with unilateral lesion of the substantia nigra pars compactaSNpc) induced by intracerebral infusion of 6-hydroxydopamine6-OHDA) which are challenged with dopaminergic drugs cane used as a model of Parkinson’s disease (PD). Since then, it
as been one of the most popular models to study drugs poten-ially useful to treat this disease [4,16,41,49]. However, there iso consensual explanation for the direction of the turns made∗ Corresponding author. Tel.: +55 41 3361 1717; fax: +55 41 3266 2042.E-mail address: [email protected] (C. Da Cunha).
bams
if
166-4328/$ – see front matter © 2008 Elsevier B.V. All rights reserved.oi:10.1016/j.bbr.2008.01.012
glia; Dopamine receptor; Parkinson’s disease; MPTP; 6-Hydroxydopamine
y these rats. Most studies have shown that rats with unilat-ral lesions induced by 6-OHDA present a contraversive turningehaviour (in the direction opposite to the lesioned side) whenhallenged with apomorphine. However, ipsiversive or few turnsn both directions have also been reported in animals with uni-ateral lesions induced by 6-OHDA [2,7,15,35,38,44,46], andther neurotoxins (e.g., rotenone, MPP+) [22,24,42]. Turningehaviour seems to be caused by a misbalance between the rightnd left side drive to initiate movements due to an asymmetricalodulation of midbrain dopaminergic neurons on corticostriatal
ynapses [12,32].The site of infusion of 6-OHDA has been reported to be crit-
cal to explain the direction of the turns [38,40,42]. The medialorebrain bundle (MFB) has been indicated as the infusion

l Brai
sctra
firtStrlmatifimf
2
2
ttopoaU
2
wepLSdao(pc
2
awhwairosstfrmt±M
Ff
C. Da Cunha et al. / Behavioura
ite that causes more contraversive turns after an apomorphinehallenge. In contrast, ipsiversive behaviour is observed whenhese animals are challenged with amphetamine [46]. Dopamineeceptor supersensitivity in the striatum has also been suggesteds an important factor inducing contraversive turns [44].
In the present study, rather than simply describing the criticalactors that determine the occurrence and direction of the turn-ng behaviour in unilaterally SNpc-lesioned rats, we propose aational model to explain this phenomenon. The hypothesis thathe direction of the turns is determined by the site of lesion in theNpc, its size or magnitude or by dopamine receptor supersensi-
ivity in the striatum was tested in 6-OHDA- and MPTP-lesionedats. These neurotoxins were chosen to induce large and smallesions in the SNpc, respectively [9,35]. The hypothesis that a
isbalance between dopamine receptor activation in the rightnd left striatum determines the direction of the turns was alsoested. This hypothesis does not rule out the site of the lesion andts magnitude or dopamine receptor supersensitivity as importantactors determining the direction of turning behaviour. Instead,t proposes that the right/left striatal dopaminergic neurotrans-
ission misbalance is determined by the combination of theseactors.
. Materials and methods
.1. Animals
Adult male Wistar rats from our own breeding stock weighing 280–310 g athe beginning of the experiments were used. The animals were maintained in aemperature-controlled room (22 ± 2 ◦C) on a 12-h dark:12-h light cycle (lights
n at 7:00 a.m.), with food and water available ad libitum. All experimentalrocedures were in compliance with the guidelines of the National Institutesf Health and the Brazilian Society for Neuroscience and Behaviour and werepproved by the Institutional Animal Care and Use Committee of the Federalniversity of Parana State.iiaos
ig. 1. Schematic drawing of coronal slices of the rat brain showing the sites of injectioorebrain bundle; SNpc-L, SNpc-C, and SNpc-M = lateral, central, and medial portio
n Research 189 (2008) 364–372 365
.2. Chemicals
The drugs and other fine chemical compounds used in these experimentsere purchased from the following sources: chloral hydrate (Reagen, Brazil),
thanol, acetonitrile, tetrahydrofuran and perchloric acid (Merck, Germany),enicillin G-procaine (Bristol-Myers Squibb, USA), sodium thiopental (Abbottaboratories, USA), [3H]SCH-23390 and [3H]raclopride (PerkinElmer Lifecience, USA), and atropine sulfate, acetaldehyde, apomorphine, citric acid,opamine, desipramine, DOPAC, EDTA, HVA, MPTP, HCl, heptanesulfoniccid, butaclamol, sulpiride and 6-OHDA (Sigma Chemical Co., USA). The mon-clonal antibody against TH raised in mice was purchased from Incstar Corp.USA), and the ABC Elite kit to localize the antigen–antibody complex wasurchased from Vector Laboratories (USA). The Synergi C-18 reverse-phaseolumn was purchased from Phenomenex (Torrance, CA, USA).
.3. Surgeries
The animals received atropine sulfate (0.4 mg/kg, i.p.) to suppress salivationnd penicillin G-procaine (20,000 U in 0.1 ml, i.m.) to prevent infection, andere anesthetized with 3 ml/kg equitesin (1% sodium thiopental, 4.25% chloralydrate, 2.13% magnesium sulfate, 42.8% propylene glycol, and 3.7% ethanol inater). Animals lesioned with MPTP received three i.p. injections of 120 mg/kg
cetaldehyde 10 min before, at the beginning, and immediately after surgery, toncrease the effectiveness of the neurotoxin. Animals lesioned with 6-OHDAeceived 25 mg/kg desipramine (i.p.) 10 min before surgery to prevent the lossf noradrenergic neurons. MPTP (100 �g/�l in saline) or 6-OHDA (8 �g/�l inaline supplemented with 0.2% ascorbic acid) was infused through a 30-gaugetainless needle at 0.25 �l/min. The needle was maintained in place for an addi-ional 2 min to avoid reflux. The following volumes and coordinates were usedor neurotoxin infusion: medial part of the SNpc (SNpc-M): 1 �l, anteroposte-ior (AP), −5.9 mm from the bregma; mediolateral (ML), ±1.2 mm from theidline; dorsoventral (DV), −8.6 mm from the skull; nose bar, −3.3 mm from
he interaural line; central part of the SNpc (SNpc-C): 1 �l, AP, −5.0 mm; ML,2.1 mm; DV, −7.7 mm; lateral part of the SNpc (SNpc-L): 1 �l, AP, −6.1 mm;L, ±2.6 mm; DV, −7.3 mm. Fig. 1 illustrates the sites of MPTP or 6-OHDA
nfusion. The following parameters were used for animals that received MPTPnto the three sites of the SNpc (SNpc-M,C,L): the same coordinates as describedbove, 1 �l into the medial, 2 �l into the central, and 2 �l into the lateral partf the SNpc. MFB: 2 �l: AP, −1.9 mm; ML, +1.9 mm; DV, −7.2 mm. Thesetereotaxic coordinates were adapted from the Atlas of Paxinos and Watson
n of MPTP or 6-OHDA. Adapted from Paxinos and Watson [36]. MFB = medialns of the substantia nigra pars compacta, respectively.

3 l Brai
[ibsawsd
2
apht1
2
d(woawaawte(brP−
2
3wT(tTme(3pcas
2
(s−−iaao
iCi9(w(iKdprwbdtg
2
wrbfc
3
3it
iigpiSi(fae((
isgNoits
66 C. Da Cunha et al. / Behavioura
36]. Sham-operated animals were submitted to the same surgical procedure,ncluding the hole in the skull, but the infusion needle was not lowered into therain. Another set of animals (Saline) was submitted to the same procedure butaline was infused instead of the neurotoxins. After surgery, the animals werellowed to recover from the anesthesia in a temperature-controlled chamber andere then returned to their home cage. The animals were fed a pasty diet con-
isting of a mixture of crumbled chow and water during the first 5 postoperativeays. This procedure reduced body weight loss and mortality.
.4. Turning behaviour
One week after surgery the animals were challenged by injecting 1 mg/kgpomorphine, s.c. Immediately after the injection, the rats were individuallylaced in a round plastic container measuring 28 cm in diameter and 25 cm ineight, and the number of 360◦ turns toward the lesioned side (ipsiversive) andoward the opposite side (contraversive) were video-recorded over a period ofh [47].
.5. Tyrosine hydroxylase immunohistochemistry
After the behavioural test, the animals were killed by decapitation and theirorsal striata were removed for the determination of dopamine concentrationsee below). The posterior part of the rat brain was preserved in formalin for 1eek and then placed in 20% sucrose formalin 48 h before sectioning. Four seriesf 30-�m thick sections were cut with a sliding microtome on the frontal planend collected from the caudal diencephalon to the caudal midbrain. The sectionsere immunostained for tyrosine hydroxylase (TH) with a monoclonal antibody
gainst TH (1:5000 dilution). The antigen–antibody complex was localized withn ABC Elite kit (Vector, UK). Slides were then dehydrated and coverslippedith DPX. The number of TH-immunoreactive neurons was determined using
he 40× objective of an Axiophot Stereo HB050 (Zeiss, Germany) microscopequipped with a camera lucida. The UTHSCSA Image Tool version 2.0 softwareUniversity of Texas, San Antonio, USA) was used to count these neurons onoth sides of the midbrain, including the ventral tegmental area, SNpc, andetrorubral nucleus in eight series of coronal sections based on the Atlas ofaxinos and Watson [37]: −4.8 mm, −5.2 mm, −5.6 mm, −5.8 mm, −6.04 mm,6.3 mm, −6.72 mm, and −7.04 mm from the bregma.
.6. Determination of dopamine and metabolites by HPLC-ED
Endogenous levels of dopamine and its non-conjugated metabolites,4-dihydroxyphenylacetic acid (DOPAC) and homovanillic acid (HVA)ere assayed by reverse-phase HPLC with electrochemical detection (ED).he system consisted of a Synergi Fusion-RP C-18 reverse-phase column
150 × 4.6 mm i.d., 4-�m particle size, Phenomenex, USA), an L-ECD-6A elec-rochemical detector (Shimadzu, Japan), and an LC-10AD pump (Shimadzu).he column was maintained inside a temperature-controlled oven (30 ◦C, Shi-adzu). The oxidation potential was fixed at +0.80 V using an Ag/AgCl working
lectrode. The tissue samples were homogenized with an ultrasonic cell disrupterSonics, USA) in 0.1 M perchloric acid. After centrifugation at 15,000 × g for0 min, 20 �l of the supernatant was injected into the chromatograph. The mobilehase, used at a flow rate of 1 ml/min, had the following composition: 15.7 gitric acid, 471.5 ml HPLC-grade water, 78 mg heptane sulfonic acid, 20 mlcetonitrile, and 10 ml tetrahydrofuran, pH 3.0. The peak areas of the externaltandards were used to quantify the sample peaks.
.7. Autoradiographic binding assays
Seven days after surgery, rats from the saline-M,C,L (n = 8), saline-MFBn = 8), MPTP-M,C,L lesion (n = 8) and 6-OHDA-MFB (n = 8) groups wereacrificed. The brains were removed quickly, frozen over dry ice and stored at80 ◦C. Twenty-micrometer coronal sections were cut with a Leica cryostat at
20 ◦C and mounted on washed gelatin-coated slides at approximately 0.3 mmntervals along the anterior–posterior extent of the brain. The slides were storedt −80 ◦C until assayed. Binding assays of [3H]SCH-23390 to D1 receptors [34]nd of [3H]raclopride to D2 receptors [39] were performed as described previ-usly. Briefly, slices were brought to room temperature and then preincubated
odta
n Research 189 (2008) 364–372
n 50 mM Tris–HCl, 120 mM NaCl, 4 mM MgCl2, 1 mM EDTA, and 1.5 mMaCl2 2H2O buffer, pH 7.4, for 30 min (D1) or 15 min (D2). Sections were then
ncubated in fresh buffer containing 2 nM [3H]SCH-23390 (85.0 Ci/mmol) for0 min at room temperature (D1), or in buffer containing 2 M [3H]raclopride60.1 Ci/mmol) for 120 min at room temperature (D2). Additional sets of slicesere incubated in the presence of 2 �M butaclamol (D1) or 10 �M sulpiride
D2) for the determination of nonspecific binding. Sections were then washedn buffer and allowed to dry at room temperature. The slices were exposed toodak BioMax MR films in tungsten cassettes together with calibrated stan-ards for 4 weeks. The films were developed and densitometric analyses wereerformed using an M2 MCID system (Imaging Research, Canada). Anatomicalegions were defined according to Paxinos and Watson [36]. Multiple readingsere obtained by an investigator who was unaware to which group the animalselonged and averaged over serial sections for each brain. Density values ofifferent regions of the striatum region were first averaged within each section,hen across sections within each brain, and then across subjects within eachroup.
.8. Statistics
All results are reported as the mean ± S.E.M. Differences among groupsere analyzed by one-way ANOVA, followed by the Newman–Keuls test. Data
egarding the time course of apomorphine-induced turns were scored as the num-er of ipsiversive–contraversive turns and were analyzed by two-way ANOVAor repeated measures, followed by the Newman–Keuls test. Differences wereonsidered to be statistically significant when p ≤ 0.05.
. Results
.1. Effects of the unilateral infusion of 6-OHDA or MPTPnto different sites of the nigrostriatal pathway on theurning behaviour of rats
The effects of these treatments on apomorphine-induced turn-ng behaviour are shown in Fig. 2. The infusion of saline per sento the SNpc induced ipsiversive turns compared to the shamroup (Fig. 2A; F(6,65) = 37.99, p ≤ 0.001, one-way ANOVA;≤ 0.05, Newman–Keuls post hoc test). The infusion of MPTP
nto the central, lateral, or central + medial + lateral sites of theNpc, but not into the medial part of the SNpc or into the MFB,
nduced more ipsiversive turns compared to the saline groupFig. 2A; p ≤ 0.05, Newman–Keuls test). Equivalent results wereound when comparing the time-course curves of the effect ofpomorphine (Fig. 2B). Two-way ANOVA showed significantffects for site of infusion (F(6,65) = 37.99, p ≤ 0.001), timeF(11,715) = 17.43, p ≤ 0.001), and site versus time interactionF(66,715) = 3.80, p ≤ 0.001).
As can be seen in Fig. 2A′, rats lesioned with 6-OHDAnfused into the MFB or into the medial part of the SNpc pre-ented contraversive turns significantly different from the salineroup (F(5,52) = 14.65, p ≤ 0.001, one-way ANOVA; p ≤ 0.05,ewman–Keuls test). Contraversive turning behaviour was alsobserved in most, but not all, animals that received 6-OHDAnto the central part of the SNpc (Fig. 2A′); the scores ofhis group did not differ significantly from those obtained forham or saline animals (p > 0.2, Newman–Keuls test). More-
ver, infusion of 6-OHDA into the lateral part of the SNpcid not cause contraversive behaviour (p > 0.2, Newman–Keulsest), but ipsiversive turns that differed significantly from shamnimals (Fig. 2A′; F(5,52) = 3.43, p ≤ 0.01, one-way ANOVA;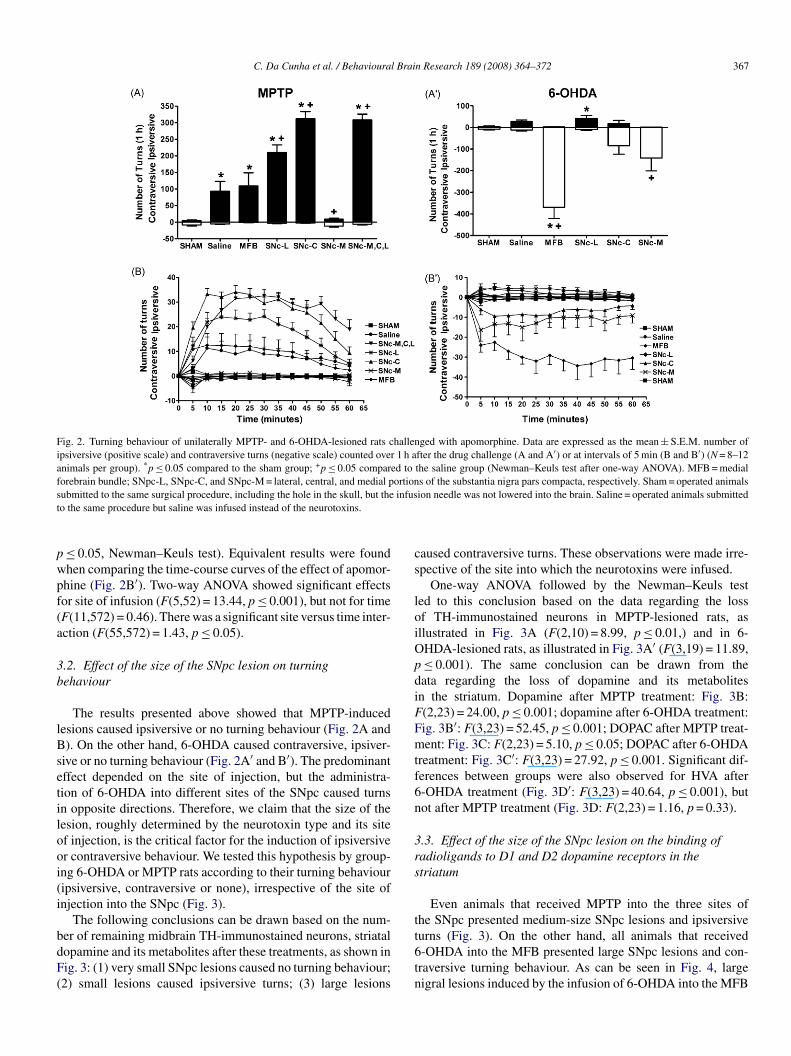
C. Da Cunha et al. / Behavioural Brain Research 189 (2008) 364–372 367
Fig. 2. Turning behaviour of unilaterally MPTP- and 6-OHDA-lesioned rats challenged with apomorphine. Data are expressed as the mean ± S.E.M. number ofipsiversive (positive scale) and contraversive turns (negative scale) counted over 1 h after the drug challenge (A and A′) or at intervals of 5 min (B and B′) (N = 8–12animals per group). *p ≤ 0.05 compared to the sham group; +p ≤ 0.05 compared to the saline group (Newman–Keuls test after one-way ANOVA). MFB = medialforebrain bundle; SNpc-L, SNpc-C, and SNpc-M = lateral, central, and medial portions of the substantia nigra pars compacta, respectively. Sham = operated animalss infust
pwpf(a
3b
lBsetilooi(i
bdF(
cs
loiOpdiFFmtf6n
3rs
t
ubmitted to the same surgical procedure, including the hole in the skull, but theo the same procedure but saline was infused instead of the neurotoxins.
≤ 0.05, Newman–Keuls test). Equivalent results were foundhen comparing the time-course curves of the effect of apomor-hine (Fig. 2B′). Two-way ANOVA showed significant effectsor site of infusion (F(5,52) = 13.44, p ≤ 0.001), but not for timeF(11,572) = 0.46). There was a significant site versus time inter-ction (F(55,572) = 1.43, p ≤ 0.05).
.2. Effect of the size of the SNpc lesion on turningehaviour
The results presented above showed that MPTP-inducedesions caused ipsiversive or no turning behaviour (Fig. 2A and). On the other hand, 6-OHDA caused contraversive, ipsiver-
ive or no turning behaviour (Fig. 2A′ and B′). The predominantffect depended on the site of injection, but the administra-ion of 6-OHDA into different sites of the SNpc caused turnsn opposite directions. Therefore, we claim that the size of theesion, roughly determined by the neurotoxin type and its sitef injection, is the critical factor for the induction of ipsiversiver contraversive behaviour. We tested this hypothesis by group-ng 6-OHDA or MPTP rats according to their turning behaviouripsiversive, contraversive or none), irrespective of the site ofnjection into the SNpc (Fig. 3).
The following conclusions can be drawn based on the num-
er of remaining midbrain TH-immunostained neurons, striatalopamine and its metabolites after these treatments, as shown inig. 3: (1) very small SNpc lesions caused no turning behaviour;2) small lesions caused ipsiversive turns; (3) large lesionst6tn
ion needle was not lowered into the brain. Saline = operated animals submitted
aused contraversive turns. These observations were made irre-pective of the site into which the neurotoxins were infused.
One-way ANOVA followed by the Newman–Keuls tested to this conclusion based on the data regarding the lossf TH-immunostained neurons in MPTP-lesioned rats, asllustrated in Fig. 3A (F(2,10) = 8.99, p ≤ 0.01,) and in 6-HDA-lesioned rats, as illustrated in Fig. 3A′ (F(3,19) = 11.89,≤ 0.001). The same conclusion can be drawn from theata regarding the loss of dopamine and its metabolitesn the striatum. Dopamine after MPTP treatment: Fig. 3B:(2,23) = 24.00, p ≤ 0.001; dopamine after 6-OHDA treatment:ig. 3B′: F(3,23) = 52.45, p ≤ 0.001; DOPAC after MPTP treat-ent: Fig. 3C: F(2,23) = 5.10, p ≤ 0.05; DOPAC after 6-OHDA
reatment: Fig. 3C′: F(3,23) = 27.92, p ≤ 0.001. Significant dif-erences between groups were also observed for HVA after-OHDA treatment (Fig. 3D′: F(3,23) = 40.64, p ≤ 0.001), butot after MPTP treatment (Fig. 3D: F(2,23) = 1.16, p = 0.33).
.3. Effect of the size of the SNpc lesion on the binding ofadioligands to D1 and D2 dopamine receptors in thetriatum
Even animals that received MPTP into the three sites ofhe SNpc presented medium-size SNpc lesions and ipsiversive
urns (Fig. 3). On the other hand, all animals that received-OHDA into the MFB presented large SNpc lesions and con-raversive turning behaviour. As can be seen in Fig. 4, largeigral lesions induced by the infusion of 6-OHDA into the MFB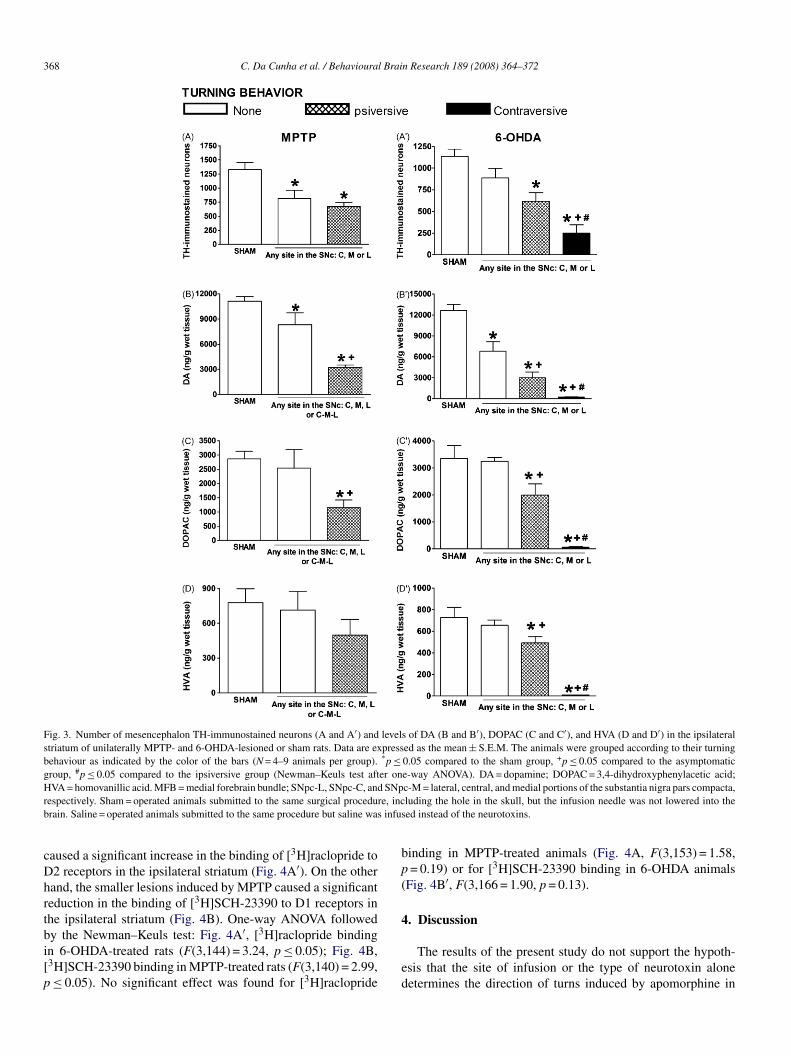
368 C. Da Cunha et al. / Behavioural Brain Research 189 (2008) 364–372
Fig. 3. Number of mesencephalon TH-immunostained neurons (A and A′) and levels of DA (B and B′), DOPAC (C and C′), and HVA (D and D′) in the ipsilateralstriatum of unilaterally MPTP- and 6-OHDA-lesioned or sham rats. Data are expressed as the mean ± S.E.M. The animals were grouped according to their turningbehaviour as indicated by the color of the bars (N = 4–9 animals per group). *p ≤ 0.05 compared to the sham group, +p ≤ 0.05 compared to the asymptomaticgroup, #p ≤ 0.05 compared to the ipsiversive group (Newman–Keuls test after one-way ANOVA). DA = dopamine; DOPAC = 3,4-dihydroxyphenylacetic acid;H SNpr re, incb infus
cDhrtbi[p
bp(
4
VA = homovanillic acid. MFB = medial forebrain bundle; SNpc-L, SNpc-C, andespectively. Sham = operated animals submitted to the same surgical procedurain. Saline = operated animals submitted to the same procedure but saline was
aused a significant increase in the binding of [3H]raclopride to2 receptors in the ipsilateral striatum (Fig. 4A′). On the otherand, the smaller lesions induced by MPTP caused a significanteduction in the binding of [3H]SCH-23390 to D1 receptors inhe ipsilateral striatum (Fig. 4B). One-way ANOVA followed
y the Newman–Keuls test: Fig. 4A′, [3H]raclopride bindingn 6-OHDA-treated rats (F(3,144) = 3.24, p ≤ 0.05); Fig. 4B,3H]SCH-23390 binding in MPTP-treated rats (F(3,140) = 2.99,≤ 0.05). No significant effect was found for [3H]racloprideed
c-M = lateral, central, and medial portions of the substantia nigra pars compacta,luding the hole in the skull, but the infusion needle was not lowered into theed instead of the neurotoxins.
inding in MPTP-treated animals (Fig. 4A, F(3,153) = 1.58,= 0.19) or for [3H]SCH-23390 binding in 6-OHDA animals
Fig. 4B′, F(3,166 = 1.90, p = 0.13).
. Discussion
The results of the present study do not support the hypoth-sis that the site of infusion or the type of neurotoxin aloneetermines the direction of turns induced by apomorphine in

C. Da Cunha et al. / Behavioural Brain Research 189 (2008) 364–372 369
F o stri( tissue
SnwHtttaOlt6Otsnp
mrrospahlTcaamMot
iacdt
bipMippBdntaootmisftiObst
ig. 4. Binding of [3H]raclopride (A and A′) and [3H]SCH-23390 (B and B′) tN = 8–9 animals per group). Values are expressed as means ± S.E.M. in �Ci/g
Npc-hemilesioned rats. It is true that the infusion of a specificeurotoxin into some specific sites along the nigrostriatal path-ay causes a predominant pattern of turning behaviour (Fig. 2).owever, the direction of the turns cannot be predicted only by
he site of infusion. The present findings clearly demonstrate thathe infusion of 6-OHDA, but not of MPTP, into the MFB and intohe medial part of the SNpc caused contraversive turns. MPTPlways caused ipsiversive or no turning behaviour, whereas 6-HDA caused predominantly contraversive turns. However, the
atter may also cause ipsiversive turns, like in rats infused intohe lateral part of the SNpc. In addition, even when infused with-OHDA into the medial or central part of the SNpc, some 6-HDA rats presented ipsiversive behaviour. This inconsistent
urning behaviour of 6-OHDA rats has been reported in sometudies [7,20,35,38,41,44]. In order to obtain a more homoge-eous sample, many authors excluded 6-OHDA rats that did notresent contraversive turns [26,41,43].
The apparently chaotic pattern of results described aboveakes sense if explained not as a result of the site of lesion, but
ather as a function of the loss of midbrain dopaminergic neu-ons and striatal depletion of dopamine, irrespective of the sitef lesion (Fig. 3). Before discussing the evidence in favor of thisecond hypothesis, we will see whether the pattern of lesion canrovide a clue regarding the neurotoxic mechanisms of MPTPnd 6-OHDA. When 6-OHDA was infused into a site with aigh density of dopaminergic fibers, the neurotoxin caused largeesions in neurons from which dopaminergic fibers originate.his was the case of infusion of 6-OHDA into the MFB. In thisase, most of the midbrain dopaminergic neurons were lost [seelso 28, 45, 49]. The fibers that arise from neurons of the medialnd central parts of the SNpc leave it by passing through the
ost medial and ventral parts of the SNpc and then enter theFB [14,30]. Therefore, the more medial and ventral the sitef 6-OHDA infusion, the more the neurotoxin will be in con-act with dopaminergic fibers. The present study and previous
rtt
atal slices of rat brain one week after the administration of MPTP or 6-OHDA. *p ≤ 0.05 compared to the non-lesioned side (right striatum).
nvestigations have found that infusion of 6-OHDA into medialnd ventral parts of the SNpc caused a more marked and robustontraversive turning behaviour than infusion into lateral andorsal parts of the SNpc [7,35,38,41]. These findings suggesthat 6-OHDA is mainly taken up by dopaminergic fibers.
On the other hand, MPTP did not cause a large loss of mid-rain dopaminergic neurons when it was infused into sites richn dopaminergic fibers, such as the MFB and the ventromedialart of the SNpc. It is unlikely that the toxic metabolite of MPTP,PP+, is taken up preferentially by dopaminergic neuron bod-
es/dendrites compared to axons, since it is taken up by the samerotein that also takes up 6-OHDA, i.e., the dopamine trans-orter (DAT) [27]. It is also possible that monoamine oxidase(MAO-B), the enzyme that transforms MPTP into MPP+, is
ifferentially expressed in fibers and bodies of dopaminergiceurons. In addition, the brain capillaries of rats contain excep-ionally high levels of MAO-B compared to primates [23]. Asconsequence, rats are more resistant to the neurotoxic effectsf MPTP than primates [5,19]. Therefore, even the simultane-us infusion of a highly concentrated solution of MPTP intohree sites throughout the SNpc did not cause the same loss of
idbrain dopaminergic neurons than the infusion of 6-OHDAnto the MFB. This treatment did not mimic the magnitude andpreading loss of midbrain dopaminergic neurons that resultedrom the infusion of 6-OHDA into the MFB. The ipsiversiveurning behaviour of rats infused with MPTP into the three sites,n contrast to the contraversive turning behaviour of MFB 6-HDA rats, strongly suggests that it was not the site of infusionut the degree of dopaminergic neuron loss and the consequenttriatal dopamine depletion which determined the direction ofhe turns.
This hypothesis was further tested by grouping animalseceiving 6-OHDA or MPTP into different sites according toheir turning behaviour (Fig. 3). This criterion suggested thathere are two critical thresholds that determine the direction of
3 l Brai
trwnltb
egrttr
t(Ditasbd
S
For
70 C. Da Cunha et al. / Behavioura
urning behaviour: Animals with midbrain dopaminergic neu-on loss below a certain level did not present turning behaviour,hereas those with higher, but partial, loss of dopaminergiceurons presented ipsiversive turning behaviour and those thatost almost all midbrain dopaminergic neurons presented con-raversive turning behaviour. Only 6-OHDA rats showed thisehaviour.
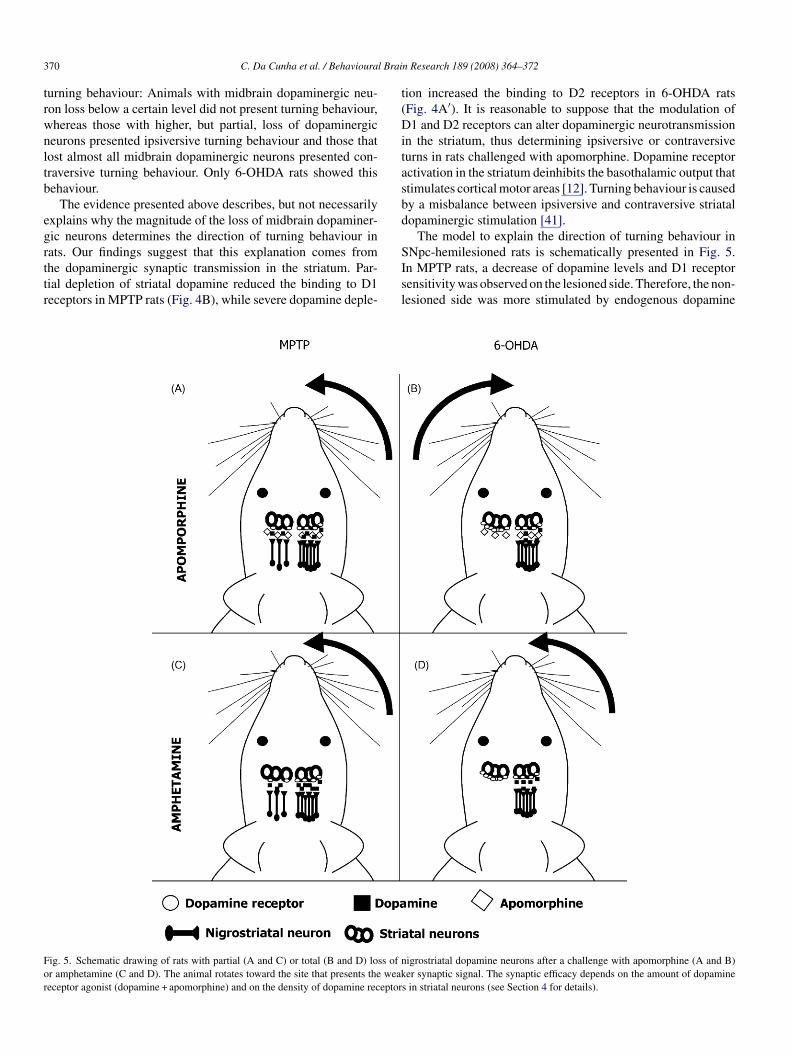
The evidence presented above describes, but not necessarilyxplains why the magnitude of the loss of midbrain dopaminer-ic neurons determines the direction of turning behaviour inats. Our findings suggest that this explanation comes from
he dopaminergic synaptic transmission in the striatum. Par-ial depletion of striatal dopamine reduced the binding to D1eceptors in MPTP rats (Fig. 4B), while severe dopamine deple-Isl
ig. 5. Schematic drawing of rats with partial (A and C) or total (B and D) loss of nr amphetamine (C and D). The animal rotates toward the site that presents the weakeceptor agonist (dopamine + apomorphine) and on the density of dopamine receptor
n Research 189 (2008) 364–372
ion increased the binding to D2 receptors in 6-OHDA ratsFig. 4A′). It is reasonable to suppose that the modulation of1 and D2 receptors can alter dopaminergic neurotransmission
n the striatum, thus determining ipsiversive or contraversiveurns in rats challenged with apomorphine. Dopamine receptorctivation in the striatum deinhibits the basothalamic output thattimulates cortical motor areas [12]. Turning behaviour is causedy a misbalance between ipsiversive and contraversive striatalopaminergic stimulation [41].
The model to explain the direction of turning behaviour inNpc-hemilesioned rats is schematically presented in Fig. 5.
n MPTP rats, a decrease of dopamine levels and D1 receptorensitivity was observed on the lesioned side. Therefore, the non-esioned side was more stimulated by endogenous dopamineigrostriatal dopamine neurons after a challenge with apomorphine (A and B)er synaptic signal. The synaptic efficacy depends on the amount of dopamine
s in striatal neurons (see Section 4 for details).

l Brai
p(iolFi[bid
rtdtwM
esntdmitaeb
csdstctfpiul
A
FsCa
R
[
[
[
[
[
[
[
[
[
[
C. Da Cunha et al. / Behavioura
lus the exogenous dopamine receptor agonist apomorphineFig. 5A). 6-OHDA rats presented D2 receptor supersensitiv-ty on the lesioned side, which compensates for the decreasef dopamine and, therefore, the lesioned side was more stimu-ated than the non-lesioned side in the presence of apomorphineig. 5B). Similar findings have been reported in previous stud-
es on 6-OHDA rats [13,21,31,33,44,48] and MPTP monkeys1]. Therefore, the balance of dopaminergic neurotransmittersetween the lesioned and non-lesioned sides was the oppositen MPTP and 6-OHDA rats, which would explain the oppositeirections of their turning behaviour.
The ipsiversive turning behaviour of 6-OHDA and MPTPats challenged with amphetamine can also be explained byhis model. Since amphetamine induces a higher release ofopamine on the non-lesioned side in MPTP and 6-OHDA rats,his side is more stimulated in both cases, a fact explaininghy amphetamine induces ipsiversive turning behaviour in bothPTP- and 6-OHDA-hemilesioned rats (Fig. 5C and D).In summary, the present study suggests that rats with unilat-
ral lesion of the SNpc rotate toward the side with the weakertriatal dopaminergic neurotransmission. The dopaminergiceurotransmission misbalance is determined by the magni-ude of dopamine depletion and by up- or down-regulation ofopamine receptors. When challenged with apomorphine, ani-als with small unilateral lesion/depletion of dopamine present
psiversive turning behaviour due to a weaker stimulation ofhe lesioned side (caused by the loss of dopamine), whereasnimals with an almost total unilateral loss of midbrain dopamin-rgic neurons/striatal dopamine present contraversive turningehaviour due to dopamine receptor supersensitization.
It is possible that the mechanisms that convert ipsiversive intoontraversive turning behaviour in SNpc-hemilesioned rats areimilar to those that support the motor abnormalities observeduring the early and late phases of PD. Nowadays, some studiesuggest that one of the most marked disabilities observed duringhe late phase of the disease, i.e., dyskinesia, is closely related toontraversive turns in 6-OHDA rats [25,26]. Previous investiga-ions from our group showed that MPTP rats are a good modelor the study of cognitive impairments that occur during the earlyhase of PD [3,6,8–11,17,18,29,37]. Therefore, we suggest thatt would be worth studying MPTP and 6-OHDA rats to betternderstand the mechanisms that underlie the characteristics andimits of the early and late stages of PD.
cknowledgements
This research was supported in part by CNPq, CAPES,APESP, Instituto do Milenio, and Fundacao Araucaria. Thekillful assistance of Mariza Bortolanza, Vinicius A. Lima, Jeffhandler, Scheila M. G. Takayasu, and Michele L. Gregorio iscknowledged.
eferences
[1] Alexander GM, Schwartzman RJ, Grothusen JR, Brainard I, Gordon SW.Changes in brain dopamine-receptors in MPTP parkinsonian monkeys fol-lowing L-dopa treatment. Brain Res 1993;625:276–82.
[
n Research 189 (2008) 364–372 371
[2] Annett LE, Wannerton LA, Watt CA, Thiemann G, Hasenohrl RU. Behav-ioral responses directed into contralateral compared with ipsilateral spacespare dopamine in the striatum following a unilateral 6-OHDA lesion ofthe medial forebrain bundle in rats. Cell Transplant 2006;15:549–50.
[3] Bellissimo MI, Kouzmine I, Ferro MM, Oliveira BH, Canteras NS, DaCunha C. Is the unilateral lesion of the left substantia nigra pars compactasufficient to induce working memory impairment in rats? Neurobiol LearnMem 2004;82:150–8.
[4] Blandini F, Levandis G, Bazzini E, Nappi G, Armentero MT. Time-courseof nigrostriatal damage, basal ganglia metabolic changes and behaviouralalterations following intrastriatal injection of 6-hydroxydopamine in therat: new clues from an old model. Eur J Neurosci 2007;25:397–405.
[5] Blum D, Torch S, Lambeng N, Nissou MF, Benabid AL, Sadoul R, et al.Molecular pathways involved in the neurotoxicity of 6-OHDA, dopamineand MPTP: contribution to the apoptotic theory in Parkinson’s disease.Prog Neurobiol 2001;65:135–72.
[6] Braga R, Kouzmine I, Canteras NS, Da Cunha C. Lesion of the substantianigra, pars compacta impairs delayed alternation in a Y-maze in rats. ExpNeurol 2005;192:134–41.
[7] Costall B, Marsden CD, Naylor RJ, Pycock CJ. Relationship betweenstriatal and mesolimbic dopamine dysfunction and nature of circlingresponses following 6-hydroxydopamine and electrolytic lesions of ascend-ing dopamine systems of rat-brain. Brain Res 1976;118:87–113.
[8] Da Cunha C, Angellucci MEM, Canteras NS, Wonnacott S, Takahashi RN.The lesion of the rat substantia nigra pars compacta dopaminergic neuronsas a model for Parkinson’s disease memory disabilities. Cell Mol Neurobiol2002;22:227–37.
[9] Da Cunha C, Gevaerd MS, Vital MABF, Miyoshi E, Andreatini R, Sil-veira R, et al. Memory disruption in rats with nigral lesions induced byMPTP: a model for early Parkinson’s disease amnesia. Behav Brain Res2001;124:9–18.
10] Da Cunha C, Wietzikoski S, Wietzikoski EC, Miyoshi E, Ferro MM,Anselmo-Franci JA, et al. Evidence for the substantia nigra pars com-pacta as an essential component of a memory system independent of thehippocampal memory system. Neurobiol Learn Mem 2003;79:236–42.
11] Da Cunha C, Wietzikoski S, Wietzikoski EC, Silva MHC, Chandler J,Ferro MM, et al. Pre-training to find a hidden platform in the Morris watermaze can compensate for deficit to find a cued platform in a rat model ofParkinson’s disease. Neurobiol Learn Mem 2007;87:451–63.
12] DeLong MR, Wichmann T. Circuits and circuit disorders of the basalganglia. Arch Neurol 2007;64:20–4.
13] Dewar KM, Paquet M, Reader TA. Alterations in the turnover rate ofdopamine D1 but not D2 receptors in the adult rat neostriatum after aneonatal dopamine denervation. Neurochem Int 1997;30:613–21.
14] Fallon JH, Loughlin SE. Substantia nigra. In: Paxinos G, editor. The ratnervous system. San Diego: Academic Press; 1985. p. 353–74.
15] Gardner EL, Mishra RK, Iordanidis P, Katzman R, Makman MH. Catalyticand receptor components of enhanced dopamine-stimulated adenylate-cyclase in 2 behaviorally distinct striatal denervation syndromes. Fed Proc1976;35:456–1456.
16] Gerlach M, Riederer P. Animal models of Parkinson’s disease: An empiricalcomparison with the phenomenology of the disease in man. J Neural Transm1996;103:987–1041.
17] Gevaerd MS, Miyoshi E, Silveira R, Canteras NS, Takahashi RN, DaCunha C. L-DOPA restores striatal dopamine level but fails to reverseMPTP-induced memory deficits in rats. Int J Neuropsychopharmacol2001;4:361–70.
18] Gevaerd MS, Takahashi RN, Silveira R, Da Cunha C. Caffeine reverses thememory disruption induced by intra-nigral MPTP-injection in rats. BrainRes Bull 2001;55:101–6.
19] Giovanni A, Sieber BA, Heikkila RE, Sonsalla PK. Studies on speciessensitivity to the dopaminergic neurotoxin 1-methyl-4phenyl-1,2,3-6-tetrahidropyridine. Part 1. Systemic administration. J Pharmacol Exp Ther
1994;270:1000–7.20] Hirschhorn ID, Hittner D, Gardner EL, Cubells J, Makman MH. Evidencefor a role of endogenous opioids in the nigrostriatal system—influenceof naloxone and morphine on nigrostriatal dopaminergic super-sensitivity.Brain Res 1983;270:109–17.

3 l Brai
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
Brain Res 2005;1038:163–70.
72 C. Da Cunha et al. / Behavioura
21] Inaji M, Okauchi T, Ando K, Maeda J, Nagai Y, Yoshizaki T, et al.Correlation between quantitative imaging and behaviour in unilaterally6-OHDA-lesioned rats. Brain Res 2005;1064:136–45.
22] Jasso-Lopez D, Tapia R. Neurotoxic effect of intranigral injection of 1-methyl-4-phenylpyridinium on gaba-containing neurons and its relation tocircling behaviour. J Neurochem 1995;64:794–801.
23] Kalaria RN, Mitchell MJ, Harik SI. Correlation of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine neurotoxicity with blood-brain barriermonoamine-oxidase activity. Proc Natl Acad Sci USA 1987;84:3521–5.
24] Kondo J, Kitanura Y, Inden M, Taniguchi T. Hemiparkinsonian rat mod-els: different sensitivity of dopaminergic neurotoxins. Int Congr Ser2004;1260:281–5.
25] Konitsiotis S, Tsironis C. Levodopa-induced dyskinesia and rotationalbehaviour in hemiparkinsonian rats: Independent features or componentsof the same phenomenon? Behav Brain Res 2006;170:337–41.
26] Lane EL, Cheetham SC, Jenne P. Does contraversive circling in the6-OHDA-lesioned rat indicate an ability to induce motor complica-tions as well as therapeutic effects in Parkinson’s disease? Exp Neurol2006;197:284–90.
27] Lehmensiek V, Tan EM, Liebau S, Lenk T, Zettlmeisl H, Schwarz J, etal. Dopamine transporter-mediated cytotoxicity of 6-hydroxydopamine invitro depends on expression of mutant alpha-synucleins related to Parkin-son’s disease. Neurochem Int 2006;48:329–40.
28] Marin C, Rodriguez-Oroz MC, Obeso JA. Motor complications in Parkin-son’s disease and the clinical significance of rotational behaviour in the rat:have we wasted our time? Exp Neurol 2006;197:269–74.
29] Miyoshi E, Wietzikoski S, Camplessei M, Silveira R, Takahashi RN, DaCunha C. Impaired learning in a spatial working memory version and in acued version of the water maze in rats with MPTP-induced mesencephalicdopaminergic lesions. Brain Res Bull 2002;58:41–7.
30] Mori S, Crain BJ, Chacko VP, Van Zijl PCM. Three-dimensional trackingof axonal projections in the brain by magnetic resonance imaging. AnnNeurol 1999;45:265–9.
31] Newman-Tancredi A, Cussac D, Brocco M, Rivet JM, Chaput C,Touzard M, et al. Dopamine D receptor-mediated G-protein activationin rat striatum: 2 functional autoradiography and influence of uni-lateral 6-hydroxydopamine lesions of the substantia nigra. Brain Res2001;920:41–54.
32] Nicola SM, Surmeier DT, Malenka RC. Dopaminergic modulation of neu-ronal excitability in the striatum and nucleus accumbens. Ann Rev Neurosci2000;23:185–215.
33] Nikolaus S, Larisch R, Beu M, Vosberg H, Muller-Gartner HW.Bilateral increase in striatal dopamine D-2 receptor density in the 6-hydroxydopamine-lesioned rat: a serial in vivo investigation with small
animal pet. Eur J Nucl Med Mol Imaging 2003;30:390–5.34] Nobrega JN, Richter A, Tozman N, Jiwa D, Loscher W. Quantitativeautoradiography reveals regionally selective changes in dopamine D1 andD2 receptor binding in the genetically dystonic hamster. Neuroscience1996;71:927–36.
[
n Research 189 (2008) 364–372
35] Olds ME, Jacques DB, Kopyov O. Relation between rotation in the 6-OHDA, lesioned rat and dopamine loss in striatal and substantia nigrasubregions. Synapse 2006;59:532–44.
36] Paxinos G, Watson C. The rat brain in stereotaxic coordinates. 5th ed. SanDiego: Academic Press; 2005. p. 205.
37] Perry JC, Hipolide DC, Tufik S, Martins RD, Da Cunha C, AndreatiniR, et al. Intra-nigral MPTP lesion in rats: behavioral and autoradiographystudies. Exp Neurol 2005;195:322–9.
38] Redgrave P, Mitchell I. Functional validation of projection topogra-phy in the nigrostriatal dopamine system. Neuroscience 1982;7:885–94.
39] Sasaki T, Kennedy JL, Nobrega JN. Localized changes in the GABA-gatedchloride channel in rat brain after long-term haloperidol: relation to vacuouschewing movements. Synapse 1997;25:73–9.
40] Schwarting RK, Huston JP. Unilateral 6-hydroxydopamine lesions of themeso-striatal dopaminergic neurons and their physiological sequelae. ProgNeurobiol 1996;49:215–66.
41] Schwarting RK, Huston JP. The unilateral 6-hydroxydopamine lesionmodel in behavioral brain research: analysis of functional deficits, recoveryand treatments. Prog Neurobiol 1996;50:275–331.
42] Sindhu KM, Banerjee R, Senthilkumar KS, Saravanan KS, Raju BC, RaoJM, et al. Rats with unilateral median forebrain bundle, but not striatal ornigral, lesions by the neurotoxins MPP+ or rotenone display differentialsensitivity to amphetamine and apomorphine. Pharmacol Biochem Behav2006;84:321–9.
43] Takeda R, Ikeda T, Tsuda F, Abe H, Hashiguchi H, Ishida Y, et al. Unilat-eral lesions of mesostriatal dopaminergic pathway alters the withdrawalresponse of the rat hind paw to mechanical stimulation. Neurosci Res2005;52:31–6.
44] Thal L, Mishra RK, Gardner EL, Horowitz SG, Varmuza S, Makman MH.Dopamine antagonist binding increases in 2 behaviorally distinct striataldenervation syndromes. Brain Res 1979;170:381–6.
45] Truong L, Allbutt H, Kassiou M, Henderson JM. Developing a preclinicalmodel of Parkinson’s disease: a study of behaviour in rats with graded6-OHDA lesions. Behav Brain Res 2006;169:1–9.
46] Ungerstedt U. Postsynaptic supersensitivity after 6-hydroxydopamineinduced degeneration of nigro-striatal dopamine system. Acta PhysiolScand 1971;367:69–93.
47] Ungerstedt U, Arbuthnott GW. Quantitative recording of rotationalbehaviour in rats after 6-hydroxydopamine lesions of the nigrostriataldopamine system. Brain Res 1970;24:485–93.
48] Xu ZC, Ling G, Sahr RN, Neal-Beliveau BS. Asymmetrical changes ofdopamine receptors in the striatum after unilateral dopamine depletion.
49] Yuan H, Sarre S, Ebinger G, Michotte Y. Histological, behaviouraland neurochemical evaluation of medial forebrain bundle and striatal 6-OHDA lesions as rat models of Parkinson’s disease. J Neurosci Methods2005;144:35–45.




















