
Leucine5-enkephalin afferents to midbrain dopaminergic neurons: Light and electron microscopic...
13
THE JOURNAL OF COMPARATIVE NEUROLOGY 332269-281 (1993) Leucine5-Enkephalin Afferents to Midbrain Dopaminergic Neurons: Light and Electron Microscopic Examination CHANG-LlN LIANG, GERALD P. KOZLOWSKI, AND DWIGHT C. GERMAN Departments of Psychiatry ((3.-L.L.,D.C.G.) and Physiology (G.P.K.), University of Texas Southwestern Medical Center, Dallas, Texas 75235-9070. ABSTRACT The relationship between leucine5-enkephalin-containing nerve terminals and midbrain dopaminergic neurons was studied in the adult rat by light and electron microscopy. For light microscopy, alternate midbrain sections were immunostained with rabbit polyclonal antibodies against leucine5-enkephalin and tyrosine hydroxylase, by means of the peroxidase antiperoxi- dase technique. Leucine5-enkephalin stained fibers and terminals were observed with varying density in the retrorubral field (dopaminergic nucleus A8 region), substantia nigra pars compacta (dopaminergic nucleus A9 region), and ventral tegmental area and related nuclei (dopaminergic nucleus A10 region). For electron microscopy, midbrain sections were immuno- stained with a mouse monoclonal antibody against leucine"-enkephalin and a rabbit polyclonal antibody against tyrosine hydroxylase, by means of the peroxidase antiperoxidase technique and silver-intensified colloidal gold reactions, respectively. The nucleus A10 area was examined at the electron microscopic level, and there were a) both symmetric (75%) and asymmetric (25%) synapses made between leucine5-enkephalin axon terminals and dopaminergic dendrites, and also synaptic contacts with unlabeled dendrites; b) leucine5-enkephalin synaptic contacts with dopaminergic dendrites that were covered with astrocytic membranes; and c) leucine5- enkephalin appositions with unlabeled nerve terminals that made synaptic contacts with dopaminergic dendrites, suggestive of axo-axonic connections. These findings provide the structural basis for both direct and indirect control of A10 dopaminergic neurons by enkephalin-containing nerve terminals. Key words: opioid peptide, immunohistochemistry, tyrosine hydroxylase o 1993 Wiley-Liss, Inc. The neurotransmitter dopamine has been localized within neurons in several brain regions, and the highest concentra- tion of dopaminergic (DA)cells is found in the ventromedial midbrain (Dahlstrom and Fuxe, '64). The midbrain DA neurons have been divided into three nuclear groups. The cells in the retrorubral field were classified as nucleus A8, the cells in the substantia nigra were classified as nucleus A9, and the cells in the ventral tegmental area (VTA) were classified as nucleus A10. The nucleus A10 cells, however, are located in several nuclei in addition to the VTA, including the nucleus paranigralis, nucleus parabrachialis pigmentosus, nucleus interfascicularis, and nucleus centra- lis linearis (Phillipson, '79a; Swanson, '82). Ascending DA projections from nuclei A8, A9 and espe- cially A10 neurons innervate limbic nuclei and cerebral cortical regions, and these neurons constitute the mesolim- bic and mesocortical DA systems (Swanson, '82; Albanese and Minciacchi, '83; Loughlin and Fallon, '84; see Oades and Halliday, '87 for review). These neurons have been the focus of many physiological, behavioral, and pharmacologi- cal studies which have demonstrated that the A10 cells play a role in neural circuitry underlying locomotor behavior (Kelly et al., '75; Pijnenburg et al., '76; Fink and Smith, '84), ingestive behavior (Namand et al., '841, stress (Blanc et al., '801, intracranial self-stimulation (Crow, '72; Ger- man and Bowden, '741, drug abuse (Bozarth and Wise, '78; Di Chiara and Imperato, '88; Gysling and Wang, '83; Matthews and German, '84), and perhaps schizophrenia (e.g., Snyder et al., '74). Neuropeptides, which have neurotransmitter-like func- tion in the central nervous system, modulate the activity of mesolimbic and mesocortical DA neurons (Kalivas, '85; Kalivas and Abhold, '87). Of these, the opioid peptides have been examined extensively. Morphine or enkephalin ana- logues increase the firing frequency of A10 DA neurons and decreases the firing frequency of nearby non-DA neurons (Hommer and Pert, '83; Gysling and Wang, '83; Matthews and German, '84). The opioid-induced increase in DA Accepted January 20,1993 o 1993 WILEY-LISS, INC.
Transcript of Leucine5-enkephalin afferents to midbrain dopaminergic neurons: Light and electron microscopic...

THE JOURNAL OF COMPARATIVE NEUROLOGY 332269-281 (1993)
Leucine5-Enkephalin Afferents to Midbrain Dopaminergic Neurons: Light and Electron Microscopic Examination
CHANG-LlN LIANG, GERALD P. KOZLOWSKI, AND DWIGHT C. GERMAN Departments of Psychiatry ((3.-L.L., D.C.G.) and Physiology (G.P.K.), University of Texas
Southwestern Medical Center, Dallas, Texas 75235-9070.
ABSTRACT The relationship between leucine5-enkephalin-containing nerve terminals and midbrain
dopaminergic neurons was studied in the adult rat by light and electron microscopy. For light microscopy, alternate midbrain sections were immunostained with rabbit polyclonal antibodies against leucine5-enkephalin and tyrosine hydroxylase, by means of the peroxidase antiperoxi- dase technique. Leucine5-enkephalin stained fibers and terminals were observed with varying density in the retrorubral field (dopaminergic nucleus A8 region), substantia nigra pars compacta (dopaminergic nucleus A9 region), and ventral tegmental area and related nuclei (dopaminergic nucleus A10 region). For electron microscopy, midbrain sections were immuno- stained with a mouse monoclonal antibody against leucine"-enkephalin and a rabbit polyclonal antibody against tyrosine hydroxylase, by means of the peroxidase antiperoxidase technique and silver-intensified colloidal gold reactions, respectively. The nucleus A10 area was examined at the electron microscopic level, and there were a) both symmetric (75%) and asymmetric (25%) synapses made between leucine5-enkephalin axon terminals and dopaminergic dendrites, and also synaptic contacts with unlabeled dendrites; b) leucine5-enkephalin synaptic contacts with dopaminergic dendrites that were covered with astrocytic membranes; and c) leucine5- enkephalin appositions with unlabeled nerve terminals that made synaptic contacts with dopaminergic dendrites, suggestive of axo-axonic connections. These findings provide the structural basis for both direct and indirect control of A10 dopaminergic neurons by enkephalin-containing nerve terminals.
Key words: opioid peptide, immunohistochemistry, tyrosine hydroxylase
o 1993 Wiley-Liss, Inc.
The neurotransmitter dopamine has been localized within neurons in several brain regions, and the highest concentra- tion of dopaminergic (DA) cells is found in the ventromedial midbrain (Dahlstrom and Fuxe, '64). The midbrain DA neurons have been divided into three nuclear groups. The cells in the retrorubral field were classified as nucleus A8, the cells in the substantia nigra were classified as nucleus A9, and the cells in the ventral tegmental area (VTA) were classified as nucleus A10. The nucleus A10 cells, however, are located in several nuclei in addition to the VTA, including the nucleus paranigralis, nucleus parabrachialis pigmentosus, nucleus interfascicularis, and nucleus centra- lis linearis (Phillipson, '79a; Swanson, '82).
Ascending DA projections from nuclei A8, A9 and espe- cially A10 neurons innervate limbic nuclei and cerebral cortical regions, and these neurons constitute the mesolim- bic and mesocortical DA systems (Swanson, '82; Albanese and Minciacchi, '83; Loughlin and Fallon, '84; see Oades and Halliday, '87 for review). These neurons have been the focus of many physiological, behavioral, and pharmacologi-
cal studies which have demonstrated that the A10 cells play a role in neural circuitry underlying locomotor behavior (Kelly et al., '75; Pijnenburg et al., '76; Fink and Smith, '84), ingestive behavior (Namand et al., '841, stress (Blanc et al., '801, intracranial self-stimulation (Crow, '72; Ger- man and Bowden, '741, drug abuse (Bozarth and Wise, '78; Di Chiara and Imperato, '88; Gysling and Wang, '83; Matthews and German, '84), and perhaps schizophrenia (e.g., Snyder et al., '74).
Neuropeptides, which have neurotransmitter-like func- tion in the central nervous system, modulate the activity of mesolimbic and mesocortical DA neurons (Kalivas, '85; Kalivas and Abhold, '87). Of these, the opioid peptides have been examined extensively. Morphine or enkephalin ana- logues increase the firing frequency of A10 DA neurons and decreases the firing frequency of nearby non-DA neurons (Hommer and Pert, '83; Gysling and Wang, '83; Matthews and German, '84). The opioid-induced increase in DA
Accepted January 20,1993
o 1993 WILEY-LISS, INC.

270 C.-L. LIANG ET AL.
neuronal activity appears to be the result of disinhibition of DA neurons by non-DA neurons in the A10 region (Johnson and North, '92), with the latter neurons having the opioid receptors. Certain opioids, when injected into the A10 region, increase dopamine metabolism and release in meso- limbic terminal fields (Kalivas et al., '83; Di Chiara and Imperato, '88). Furthermore, microinjection of either en- kephalin or mu opioid receptor agonists into the nucleus A10 area results in a marked increase in spontaneous motor activity (Joyce and Iversen, '79; Kelley et al., '80; Vezina et al., '87), which is a behavioral indicator of increased dopamine release from mesolimbic nerve terminals.
Immunocytochemical studies have shown that there are enkephalin-containing fibers and terminals in both the A9 and A10 regions in the rodent (Uhl et al., '79; Johnson et al., '80; Finley et al., '81; Khachaturian et al., '85; Fallon and Leslie, '86), monkey (e.g., Haber and Elde, '82; Inagaki and Parent, '84; Inagaki et al., '861, and human (e.g., Emson et al., '80; Gaspar et al., '83; Marshall et al., '83; Pioro et al., '84; Haber and Groenewegen, '89). However, there has been no detailed mapping of the relationship between leucine5-enkephalin (LEI-containing nerve termi- nals and the three midbrain DA nuclear groups, and there has been only one recent ultrastructural study of LE nerve terminals in nucleus A10 (Sesack and Pickel, '92). The aim of the present study was to determine the relationship between LE fibers and terminals and DA neurons in the three midbrain nuclei at the light microscope level, and the relationship between LE nerve terminals and nucleus A10 neurons at the electron microscopic level. A preliminary report of these data has been published previously (Liang and German, '91).
MATERIALS AND METHODS Light microscopy
Adult male albino rats (165-300 gm) were deeplyanesthe- tized with Nembutal (60 mgikg, intraperitoneally) and perfused through the ascending aorta with 0.1 M phosphate buffered saline (PBS, pH 7.4, 5 minutes), and 10% neutral buffered formalin (20 minutes). The brains were blocked in a coronal plane and post-fixed in formalin for 5-10 hours. After immersion in a 20% sucrose solution for 12 hours, they were sectioned at 30 pm thickness on a freezing microtome. Two sets of sections were collected for immunos- taining: one for LE, and adjacent sections for tyrosine hydroxylase (TH). The sections were processed for immuno- cytochemistry by the unlabeled peroxidase anti-peroxidase (PAP) method. Different protocols were used for each antibody.
LE immunocytochemical stainingprocedure. The steps were as follows: a) The sections were incubated in 10% normal sheep serum in PBS-TSS solution (PBS, with 0.5% Triton X-100, and 1% normal sheep serum) for 30 minutes; b) incubation in rabbit polyclonal LE antiserum (from Dr. Stanley Watson, diluted 1:500-1,000) for 18-24 hours at room temperature. The sections were agitated on a shaker table for all incubation and rinse steps; c ) incubation in 1 : l O O sheep anti-rabbit IgG in PBS-TSS for 30 minutes; d) incubation in 1:300 rabbit PAP in PBS-TSS for 60 minutes. Following steps b and c, the sections were rinsed thor- oughly in PBS-TSS. The sections were then placed in PBS for 10 minutes and treated with diaminobenzidine/ hydrogen peroxide (DAB/H202) solution (50 mg DAB-4- HC1133 p1 30% HzOzilOO ml PBS). The sections were
rinsed, mounted on gelatin-coated slides, air dried, dehy- drated, and coverslipped. Control sections were processed in either a LE antiserum solution that had been pread- sorbed with 100 pg LE (Sigma Chemical Co., St. Louis, MO), or in PBS instead of the primary antibody solution. All remaining steps were similar to those used for the other sections.
The pro- cedure was as follows: a) The sections were incubated in 1% solution of H202 in PBS for 30 minutes to reduce endoge- nous peroxidase activity, and rinsed in PBS-TSS; b) incuba- tion in rabbit polyclonal TH antiserum (Eugene Tech., diluted 1:2,000-4,000) for 24-48 hours at room tempera- ture. The sections were agitated on a shaker table for a.11 incubation and rinse steps; c) incubation in 1 : l O O sheep anti-rabbit IgG in PBS-TSS for 30 minutes; d) incubation in 1:300 rabbit PAP solution for 60 minutes. Following steps b and c, the sections were rinsed thoroughly in PBS-TSS, and treated with nickeliDABIHzO2 solution (0.035% DAB/2.5% nickel ammonium sulfate/0.007% H20z in 0.1 M acetate buffer). The sections were then rinsed, mounted, air dried, dehydrated in alcohol, and coverslipped.
TH immunocytochemical staining procedure.
Electron microscopic double-labeling Adult male albino rats (150-200 gm) were deeply anesthe-
tized with Nembutal, and perfused transcardially with a ) 10-15 ml heparinized saline; b) 50 ml of 3.75% acrolein, and 2% paraformaldehyde in 0.1 M phosphate buffer (pH 7.4); and c) 250 ml2% paraformaldehyde in the same buffer. The brains were blocked in the coronal plane, and tissue contain- ing the midbrain was post-fixed in 2 8 paraformaldehyde for 1 hour. After a 1-1.5 minute rinse in 0.1 M phosphate buffer, the brains were cut at 40 pm thickness by using a Vibratome.
The basic procedure employed in this study was similar to that described by Chan et al. ('90). The sections were treated with 1% sodium borohydride in phosphate buffer for 30 minutes, washed thoroughly, and then incubated for 18 hours in a mixture of two primary antibodies: rabbit, anti-TH (Eugene Tech, 1:2,000), and mouse monoclonal. anti-LE (Sera-lab, 1:1,000). First, the LE immunoreaction was performed according to the standard ABC method (Hsu et al., '81). Horse anti-mouse biotinylated IgG (Vector, 30 minutes at 1:400 dilution), avidin biotin peroxidase com- plex (Vector, 60 minutes at 1 : l O O dilution), and DAB/H202 solutions were used at room temperature to visualize LE-positive fibers. Then the immunogold-silver labeling technique was employed to demonstrate TH neurons and fibers. One nm gold goat anti-rabbit IgG (Amersham, 2 hours at 1:50), and intenSE-M silver (Amersham, 6-8 minutes) were used. All incubations and washes were carried out with continuous agitation.
Following the immunocytochemical procedure, the tissue sections were rinsed in phosphate buffer, fixed with 2 8 osmium tetroxide for 20-60 minutes, dehydrated, and flat embedded in Poly-bed 812 (Polyscience, Inc.) between two Aclar plastic sheets. The VTA and nucleus parabrachialis pigmentosus were determined under the light microscope, and these regions were subsequently cut and embedded in Epon. Ultrathin sections were cut, picked up on copper grids, and counterstained with 2% uranyl acetate and lead citrate (Reynolds, '63). In order to maximize contrast between labeled and unlabeled profiles for electron micro- scopic (EM) examination, the counterstaining period was shorter than normal. In order to avoid false-negatives due

OPIOID-DOPAMINE INTERACTIONS IN MIDBRAIN
TH 271
LE
A 8
A 9
A10
Fig. 1. Tyrosine hydroxylase (TH) immunostained neurons were located in nucleus A8, nucleus A9, and nucleus A10 regions (left panels; brightfield illumination), as were leucine5-enkephalin (LE) immunostained neuronal processes (right panels; darkfield illumination). The LE immunoreactivity is illustrated in yellow. Arrows mark the locations of blood vessels common in the adjacent TH- and LE-immunostained sections. Dense LE immunostaining was observed in the retrorubral field (RRF), which contains the nucleus A8 dopaminergic (DA) neurons, and in the substantia nigra pars compacta (SNc), which contains the nucleus A9 DA neurons. The DA neurons in the substantia nigra pars reticulata (SNr), however, have little to no LE innervation. There is also dense LE innervation in nucleus A10 regions; in the nucleus interfascicularis (IF), nucleus paranigral (PN), and ventral tegmental area (VTA). Also notice the dense LE stainingin nerve terminals and somata in the rostra1 interpeduncular nucleus (IPR). IPRL, rostral-lateral interpeduncular nucleus; ml, medial lemniscus. Scale bar = 65 pm.

SNr
Figure 2

OPIOID-DOPAMINE INTERACTIONS IN MIDBRAIN 273
there was a moderate density of LE immunoreactive neuro- nal processes throughout the rostral-caudal extent of the substantia nigra pars compacta (SNc) (Fig. 2A-F), with dense innervation of specific regions including the substan- tia nigra pars lateralis (Fig. 2D-F). There was little to no LE labeling through much of the rostral-caudal extent of the substantia nigra pars reticulata (SNr) (Fig. 2A-G), and even in regions of the SNr where there were numerous DA somata (Fig. 2E-G; see Fig. 1, A9 panels). In nucleus A10, there was a high density of LE immunoreactive neuronal processes in the supramammillary region (Fig. 2A), throughout the rostral-caudal extent of the nucleus interfas- cicularis (Fig. 2B-E), in the VTA, nucleus parabrachialis pigmentosus, and nucleus paranigralis (Fig. 2E-F), and in the nucleus centralis linearis (Fig. 2G-H), In the remainder of the nucleus A10 regions there was a moderate density of LE immunoreactive neuronal processes, which may simply reflect the fact that many A10 regions contain few DA somata and large amounts of fiber bundles.
There was from medium to dense LE immunoreactivity in cell bodies and neuronal processes within specific subnu- clei of the interpeduncular nucleus (Figs. 1, 2E-H). There was also sparse LE immunoreactive fiber communication between the interpeduncular nucleus and the VTA, but it was difficult to determine whether these fibers project from the interpeduncular nucleus into the VTA or vice versa.
Electron microscopic findings For electron microscopy, double-immunostained tissue
was examined in the region of the VTA, at the level of the interpeduncular fossa (Fig. 2B), and in the nucleus parabra- chialis pigmentosus, at the level of the interpeduncular nucleus (Fig. 2E). Of the 248 LE immunoreactive profiles examined, 114 (46%) were identified as cross sections through the intervaricose regions of axons, and the others (54%) were through axonal varicosities (defined by an abundance of synaptic vesicles). All of the LE-labeled axons in nucleus A10 were unmyelinated, and were from 0.1-0.3 pm in diameter. Ovoid or round LE immunoreactive axon varicosities measured between 0.3-1.3 p,m in diameter.
Often the LE immunoreactive varicosities contained numerous round, small clear vesicles and LE-positive large vesicles (about 60-100 nm; see Figs. 4B, 5 0 , and a small number of LE immunostained varicosities also contained a few larger dense-cored vesicles (Fig. 3A). Very often the heavy LE-stained vesicles were located some distance from the synaptic specialization, (Fig. 3-5). About one fourth of the LE immunoreactive varicosities contained mitochon- dria (Figs. 3B, 4A-C).
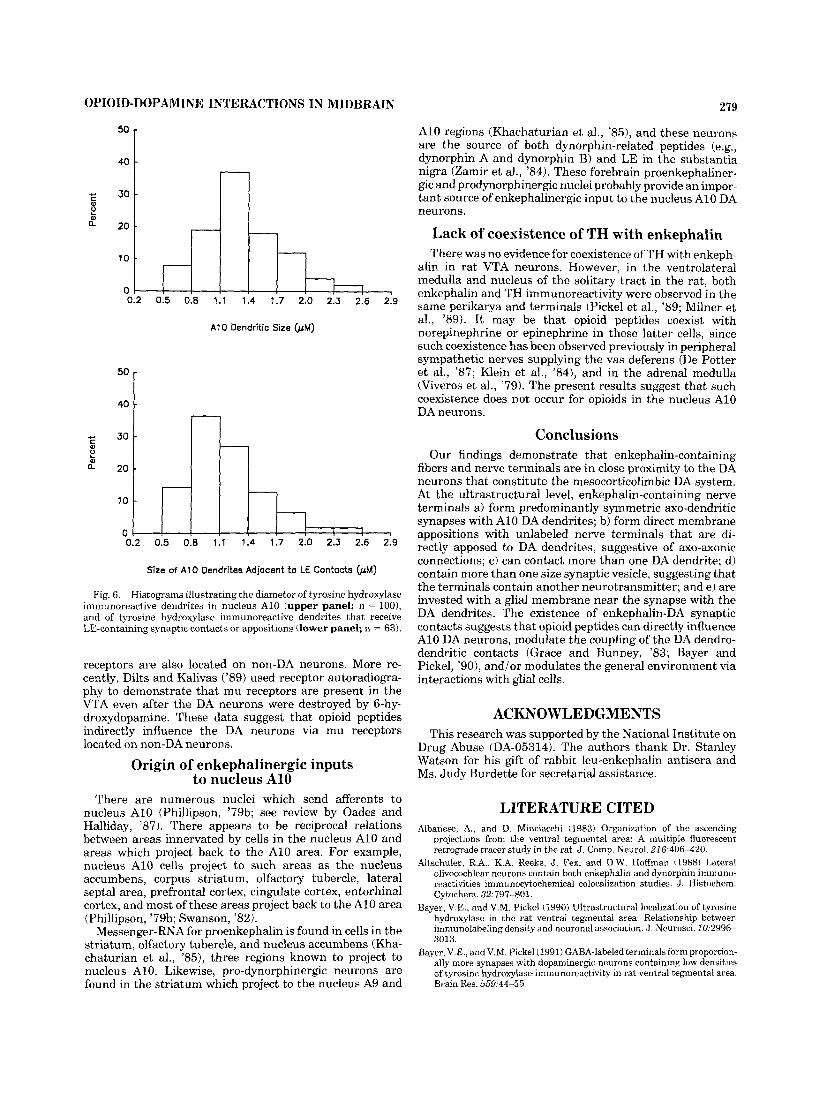
TH immunoreactive cell bodies and dendrites were la- beled with immunogold-silver particles for ultrastructural identification and analysis. The diameter of TH immunore- active dendrites varied from 0.5-2.6 km (Fig. 6). The diameter of 75% of the TH immunoreactive dendrites was from 0.8-1.7 pm, and 35% of the dendrites measured 1.1-1.4 pm. There were often dendro-dendritic appositions between the TH-labeled dendrites (Fig. 4C).
LE immunoreactive varicosities formed membrane appo- sitions, which lacked synaptic specializations, or made direct synaptic contacts with TH immunoreactive den- drites. Of 134 LE immunoreactive varicosities, 63 (47%) made appositions with TH immunoreactive dendrites, and 20 of the 63 appositions exhibited a well-defined synaptic contact. These synapses and appositions were with the thinner TH immunoreactive dendrites (Fig. 6). LE immuno-
to lack of diffusion of reagents throughout the thick sections, only sections from 0.2-1.0 p,m below the surface of the 40 km thick sections were analyzed, so that they were equally exposed to both antibodies and immunoreagents. Controls for cross-reactivity between the two antisera used in the dual-labeling technique included a) omission of either one of the primary antisera, b) omission of the colloidal gold incubation step, and c) omission of the ABC incubation steps.
The EM analysis of TH- and LE-immunostained profiles was carried out in four rats. Immunostained and related unlabeled elements were systematically scanned and photo- graphed at an initial magnification of 5,000-20,000, in a JEOL 100 CX electron microscope. The diameters of 63 TH-containing dendrites that were adjacent to LE-contain- ing nerve terminals were measured, and 100 TH-containing dendrites were measured at random. A total of 248 LE- containing profiles were analyzed.
RESULTS Light microscopic findings
TH immunoreactive cell bodies and fibers, stained with the PAP reaction, were observed in nuclei A8, A9, and A10. An example of the TH immunostaining is illustrated in Figure 1. The distribution and pattern of the DA perikarya and processes was similar to that reported previously by Dahlstrom and Fuxe ('64) using formaldehyde-induced fluorescence, and by Swanson ('821, German et al. ('831, and German and Manaye ('93) using immunocytochemistry. The same cellular distribution pattern of immunostaining of DA neurons was observed when the anti-TH antibody was conjugated to immunogold-silver particles.
Numerous LE immunoreactive varicosities were ob- served in all three midbrain DA nuclear regions. Figure 1 illustrates dense LE immunostaining in the retrorubral field, substantia nigra pars compacta, and in the ventral tegmental area, nucleus paranigralis, and nucleus interfas- cicularis. The LE immunostaining was specific as indicated by the fact that staining was not present in sections incubated with either immunoadsorbed anti-LE antibody, or when sections were incubated in PBS only, instead of the primary antibody.
Drawings were generated from adjacent sections that were stained for TH and LE, respectively (Fig. 2). A high density of LE varicosities were observed within specific regions in all three DA nuclear regions. In nucleus A8, there were numerous LE immunoreactive neuronal processes in the lateral portion, and a moderate density in the central portion of the retrorubral field (Fig. 2G,H). In nucleus A9,
Fig. 2. Maps illustrating the relationship between TH-stained somata and fibers (left side) and LE-immunoreactive neuronal pro- cesses (right side), from rostra1 (A) to caudal (H) through nuclei A8, A9, and A10. Areas with the greatest density of LE immunoreactive neuronal processes are black; areas with a medium density of LE immunoreactive neuronal processes are dark gray; and areas with little or no LE immunoreactive neuronal processes are light gray. The LE immunoreactive somata in the interpeduncular nucleus (IP) are illus- trated as white dots. CLi, nucleus centralis linearis; cp, cerebral peduncle; IF, nucleus interfascicularis; MM, nucleus mammillaris; PBP, nucleus parabrachialis pigmentosus; PN, nucleus paranigralis; RLi, nucleus rostralis linearis; RRF, retrorubral field; SNc, substantia nigra pars compacta; SNr, substantia nigra pars reticulata; SUM, nucleus supramammillaris; VTA, ventral tegmental area.

274 C.-L. LIANG ET AL.
Fig. 3. A,B: Electron micrographs illustrating synaptic contacts between LE (E) immunoreactive nerve terminals and TH (TI immuno- reactive dendrites in nucleus A10. Darkened regions in nerve terminals represent the peroxidase-antiperoxidase marker for LE, and black particles represent the location of silver-intensified gold particles in TH-containing dendrites. Both symmetric (A) and asymmetric (B) synaptic contacts were observed (arrows). Notice that the LE immuno- reactivity is unevenly distributed in the nerve terminals and is located
near dense-cored vesicles (thin arrow in A) that are some distance from the region containing the synaptic specialkation. Glial membranes were in proximity t,o the synaptic regions (stars in A and B). In B, a LE-containing nerve terminal is also apposed to an unlabeled ( u ) nerve terminal, possibly forming an axo-axonic synapse, and both are making synaptic contacts with the TH-containing dendrite. The unlabeled nerve terminal in B contains both clear vesicles and dense-cored vesicles (arrow). Scale bars = 0.5 pm.
reactive varicosities also made direct synaptic contacts with varicosities were separated from TH immunoreactive den- unlabeled dendrites (about 20%), as illustrated in Figure drites by an interposed glial membrane (Fig. 4B), or glial 4A. Examples of LE immunoreactive boutons making syn- membranes were adjacent to the LE-DA synapse (Figs. 3A, aptic contacts with TH immunoreactive dendrites are illus- 4C, 5B,C). trated in Figures 3A-B, 4C, and 5B. Sometimes LE boutons LE immunoreactive varicosities made two types of synap- made synaptic contacts and/or were apposed to two TH- tic contacts with TH immunoreactive dendrites. In about labeled dendrites (Fig. 4C). Some LE immunoreactive 75% of the cases, LE immunoreactive varicosities formed

OPIOID-DOPAMINE INTERACTIONS IN MIDBRAIN
symmetric synapses with TH immunoreactive dendrites (Figs. 3A, 4C). The symmetric synapses exhibited a thicken- ing of both the pre- and post-synaptic membranes or lack of a pronounced post-synaptic density. There were two types of vesicles in the symmetric synapses; one contained predom- inantly small clear vesicles (Fig. 3A) with a diameter of 20-40 nm and an occasional dense-cored vesicle, and the other contained predominantly larger vesicles (about 80 nm) (Fig. 5 0 . For the second type of synapse, LE immuno- reactive varicosities formed asymmetric synapses with TH immunoreactive dendrites (Fig. 3B, 5B). Asymmetric syn- apses were characterized by a pronounced post-synaptic density, and the LE immunoreactive terminals which formed these synapses contained numerous small, round vesicles (20-40 nm in diameter).
There were also examples of axo-sonic-like contacts between LE immunoreactive nerve terminals and unla- beled nerve terminals in close proximity to TH-immuno- stained dendrites. Figure 3B illustrates a LE asymmetric synapse with a DA dendrite, and the LE-containing nerve terminal is also apposed to an unlabeled nerve terminal that makes a symmetric synaptic contact with the same DA dendrite. The unlabeled nerve terminal is filled with small clear vesicles, and large dense-cored vesicles. Figure 5A illustrates a LE immunoreactive nerve terminal directly apposed to an unlabeled nerve terminal, filled with small clear vesicles, that itself is apposed to two DA dendrites.
No cell bodies or dendrites were labeled with both LE and TH immunoreactivity. This suggests that DA neurons do not colocalize with LE in nucleus A10.
2 75
and Basbaum, '84; Sasek and Elde, '86; Altshuler et al., '881, in the substantia nigra dynorphin-related peptides are found extensively in the SNr, whereas LE immunostaining is minimal in this region. Because the LE antiserum has been found to cross-react with methionine5-enkephalin (Milner et al., '891, however, the present LE immunoreactiv- ity may represent a more general enkephalin immunostain- ing.
TH is the enzyme that catalyzes the first-step in catechol- amine biosynthesis and thus is found in all catecholaminer- gic cells (i.e., those that synthesize dopamine, norepineph- rine, and epinephrine). However, since there are no adrenergic neurons in the midbrain, TH is a marker for DA neurons in this brain region (Dahlstrom and Fuxe, '64; Ungerstedt, '71; Hokfelt et al., '84).
Functions of opioid synapses in nucleus A10 At the ultrastructural level, LE is found in nerve termi-
nals that directly synapse upon DA dendrites. Both symmet- ric (75%) and asymmetric (25%) synaptic specializations were observed between enkephalin-containing axon termi- nals and the distal portions of DA dendrites (primarily 0.8-1.1 km in diameter). A recent report has found approx- imately equal percentages of symmetric and asymmetric synaptic contacts made by enkephalin-containing axon terminals and VTA DA dendrites (Sesack and Pickel, '92). The difference in the percentages of symmetric and asym- metric synaptic specializations in this latter study and the present report may be related to the fact that the latter study used a LE antibody concentration that was 20 times higher than the one used in the present study, which may have caused a greater cross-reactivity with methionine5- enkephalin. In other brain regions, LE antiserum most often labels symmetric synapses (Di Figlia et al., '82; Somogyi et al., '82; Milner et al., '891, whereas methionine5- enkephalin antiserum most often labels asymmetric syn- apses (Pickel et al., '80; Sumal et al., '82; Inagaki et al., '86).
Asymmetric synapses (Gray's type I) have classically been thought to mediate synaptic excitation, whereas sym- metric synapses (Gray's type 11) mediate synaptic inhibi- tion (Uchizono, '65; Cohen et al., '82). The present data therefore suggest that enkephalins may act to inhibit and/or excite certain A10 DA neurons. LE has been ob- served to make symmetric synaptic contacts with TH- labeled dendrites of neurons in the rostra1 ventrolateral medulla, and several lines of evidence suggest that opioid peptides inhibit the activity of these catecholaminergic neurons (see Milner et al., '89). Because enkephalins can be formed in more than one enkephalinergic precursor cell type, perhaps one synapse type represents a proenkephalin- derived enkephalin, and another synapse type represents a pro-dynorphin derived enkephalin.
In the in vitro slice preparation, opioid peptides and opioid agonists produce inhibitory effects on midbrain neurons; however, it is not the DA neuron that is inhibited but the non-DA neuron (Lacey et al., '89; Johnson and North, '92). These electrophysiological data, along with the present ultrastructural data, suggest that although there are direct synaptic contacts between enkephalin nerve terminals and A10 dendrites, there may not be opioid receptors on the DA neurons. Although mu, delta, and kappa opioid receptors are located in proximity to specific groups of midbrain DA neurons (German et al., '93; Speci- ale et al., '93), the mu and delta receptors do not appear to be located on the VTA DA neurons (Dilts and Kalivas, '89,
DISCUSSION Enkephalin-dopamine interactions in nuclei
AS, A9, and A10 The present results indicate that LE, found in both
proenkephalin and prodynorphin-containing neurons (see Rossier, '821, is located in neuronal processes in all three midbrain DA nuclear groups (nuclei AS, A9, and A10). In nucleus AS, there was a moderate to dense distribution of LE fibers and nerve terminals that overlapped with the location of the DA somata. In nucleus A9, moderate to dense LE immunoreactivity was observed throughout the rostral-caudal extent of the SNc, and the substantia nigra pars lateralis. In the SNr, however, there was very little LE immunoreactivity, even in the regions which contain DA somata (Fig. 2E-G). In nucleusA10, dense LE immunoreac- tivity was observed in the nucleus supramammillaris, nu- cleus interfascicularis, VTA, nucleus paranigralis, nucleus parabrachialis pigmentosus, and nucleus centralis linearis. These results confirm and extend those already published for LE (Uhl et al., '79; Johnson et al., '80; Finley et al., '81; Khachaturian et al., '85), and for the proenkephalin-specific peptide, met-enkephalin-arg-gly-leu (Fallon and Leslie, '86) in the rat. This pattern of innervation suggests that LE is in close proximity to the mesocorticolimbic DA neurons, as opposed to the nigrostriatal DA neurons (Fallon and Moore, '78).
Since the pattern of LE immunoreactivity at the light microscopic level is different from that exhibited by dynor- phin-related peptides in the midbrain (Fallon and Leslie, '86; Fallon and Ciofi, 'go), the present data do not appear to represent staining of dynorphin-containing neuronal pro- cesses. Although dynorphin has been co-localized within enkephalinergic neurons in certain brain regions (Guthrie

Figure 4

OPIOID-DOPAMINE INTERACTIONS IN MIDBRAIN 277
This has been also reported by Sesack and Pickel ('92) for opioid DA synapses in the VTA, and it has been reported for other peptides. In the nucleus of the solitary tract, for example, neuropeptide-Y (Pickel et al., '891, methionine5- enkephalin (Maley, '85), and substance P (Pickel et al., '77) exhibit this characteristic. These data are consistent with other evidence for colocalization of peptides with other neurotransmitters.
Axo-axonic-like contacts were observed between enkeph- alinergic nerve terminals and unlabeled nerve terminals in proximity to A10 dendrites. A small number of such synapses have been reported previously involving enkepha- lin-containing nerve terminals in the nucleus of the solitary tract (Pickel et al., '891, in the rostra1 ventrolateral medulla (Milner et al., '891, and in the VTA (Sesack and Pickel, '92). Opiate excitation of A10 DA neurons may be mediated via opioid axonic contacts with GABA nerve terminals which in turn synapse with the DA dendrites. Future studies using triple-labeling for LE, TH, and GABA will be necessary to demonstrate such synaptic arrangements.
There was often an astrocytic membrane near enkephalin- containing nerve terminals that contacted nucleus A10 dendrites. Maley ('85), Pickel et al. ('891, and Sesack and Pickel ('92) have previously observed enkephalinergic immu- nostained neuronal processes invested with astrocytic mem- brane. This finding is consistent with the observation that astrocytes have binding sites for several putative neurotrans- mitters, including opioid peptides (Hosli et al., '82; Rougon et al., '83; Bunn et al., '85). Future studies are needed to determine whether LE influences the A10 DA neurons via indirect effects on the glial cells which invest the opioid-DA synapses.
Opioid nerve terminals and opioid receptors LE is localized in the same brain region as the three
midbrain DA nuclei, but the precise cellular location of the opioid receptors has not been defined. Mu, delta, and kappa opioid receptors are differentially distributed in the region of nuclei A8, A9, and A10, as determined from receptor autoradiographic and TH immunohistochemical studies in rats (German et al., '93; Speciale et al., '93). Mu receptors are generally concentrated in higher densities (4-25-fold) than delta and kappa receptors in these brain regions. In nucleus AlO, for example, the three opioid receptor sub- types are distributed differently in specific nuclei, and the densities differ from rostral-to-caudal in these nuclei. In the VTA and nucleus parabrachial pigmentosus, the regions used for the present electron microscopic examination, all three opioid receptor subtypes are expressed in very low concentrations. Perhaps the ultrastructural relationship between opioid-containing nerve terminals and DA den- drites in brain regions containing high concentrations of opioid receptors, such as the nucleus centralis linearis, are different from those in the VTA.
Lesion studies have been controversial in that some suggest that opioid receptors are located on midbrain DA neurons, but others have not found such a relationship. It has been reported that lesions of substantia nigra DA neurons, using the neurotoxin 6-hydroxydopamine, reduce the amount of nigral and striatal opioid receptor binding (Pollard et al., '78; Llorens-Cortes et al., '791, suggesting that opioid receptors are located both on DA somata and nerve terminals. However, these investigators found that 7 0 4 0 % of the opiate binding in nucleus A9 was still present after lesioning the DA cell areas, suggesting that the opioid
'90). The functional significance of the enkephalin synaptic contacts on A10 DA neurons is unknown at the present time.
In in vivo studies, morphine, a mu opioid receptor agonist, has been found to increase the activity of midbrain DA neurons, especially the A10 neurons (Gysling and Wang, '83; Matthews and German, '84). This effect may be mediated by the axo-axonic like synapses observed in the present study. If the opioid nerve terminal presynaptically inhibits the GABA nerve terminal, which makes symmetric synapses with VTA DA neurons (Bolam and Smith, '90; Bayer and Pickel, '911, as possibly illustrated in Figure 5A, this could form the structural basis for opioid induced excitation of the midbrain DA neurons. In this case, the opioid receptor would be located on the GABA nerve terminal, not on the DA dendrite. The opioid peptide would reduce the release of the inhibitory transmitter, GABA, and thereby increase the firing rate of the DA neuron. Consis- tent with this hypothesis, enkephalin has been found to inhibit the release of GABA in the substantia nigra (Starr, '85). Furthermore, recent electrophysiological data confirm that opioids reduce GABA inhibition of A10 DA neurons (Johnson and North, '92).
Ultrastructure of enkephalinergic nerve terminals
The present study demonstrates that enkephalinergic varicosities have two distinctly different bouton types which form either synaptic contacts or membrane appositions with DA dendrites in nucleus A10. The boutons with pleomorphic vesicles and symmetric synaptic contacts may represent an inhibitory synapse, and the boutons with small round vesicles and asymmetric synaptic contacts may represent an excitatory synapse (Gray, '69). Other peptide- and monoamine-containing nerve terminals also form both symmetric and asymmetric synaptic contacts with mid- brain DA neurons in the rat; for example, substance P contacts with nucleus A9 dendrites (Chang, '88; Mahalik, '88), and serotonergic synapses with A10 cells (Herve et al., '87). Several possible explanations exist for these findings, such as a) dual actions of certain opiates, b) different cellular origination of axon terminals for the synapse types, and c) the axon terminals containing different co-transmit- ters (Rainer, '86; Mahalik, '88).
Enkephalin immunoreactivity was observed in regions containing dense-cored vesicles and was in a separate portion of the nerve terminal from the small clear vesicles.
Fig. 4. Electron micrographs illustrating synaptic contacts and membrane appositions between LE-containing (E) nerve terminals and TH-containing (T) dendrites. The markers for LE and TH are as described in Figure 3. A A symmetric synaptic contact (arrow) between a LE-containing nerve terminal and an unlabeled (u) dendrite, both of which are apposed to a TH-containing dendrite. This same dendrite has another unlabeled nerve terminal making a symmetric synaptic con- tact, and this unlabeled terminal is separated by a glial membrane (star) from another TH-containing dendrite. B: A LE-containing nerve terminal, with large synaptic vesicles, is separated from a TH- containing dendrite by an interposed glial membrane (star). C: A LE-containing nerve terminal makes a symmetric synapse with a TH-containing dendrite (arrow), which are both apposed to a second TH-containing dendrite. A glial membrane surrounds the opioid-DA synapse (star). Also notice the unlabeled (u) nerve terminal interposed between two TH-containing dendrites. The unlabeled nerve terminal contains numerous small clear vesicles and a dense-cored vesicle (arrow). Scale bars = 0.5 pm.

278 C.-L. LIANG ET AL.
Fig. 5 . Electron micrographs illustrating appositions and synaptic contacts between LE-containing nerve terminals (E) and TH-contain- ing dendrites (T). The markers for LE and TH are as described in Figure 3. A: A LE nerve terminal is apposed to an unlabeled (u) nerve terminal, suggestive of an axo-axonic contact. The unlabeled nerve terminal is apposed to two TH-containing dendrites (arrows). There is also an apposition between a LE-containing nerve terminal, and a TH-containing dendrite (upper left). B: An asymmetric synaptic con- tact (arrow) between a LE-containing nerve terminal and 3 TH-
containing dendrite, surrounded by a glial membrane (star). A second TH-containing dendrite is near the other TH-labeled dendrite. C: An apposition between a LE-containing nerve terminal and a TH- containing dendrite. Notice the large synaptic vesicles in this opioid nerve terminal. This same dendrite also receives a synaptic contact (arrow) from an unlabeled (u) nerve terminal that is also apposed to a second TH-containing dendrite. A glial membrane is located near the opioid-DA apposition (star). Scale bars = 0.5 p,m.
OPIOJD-DOPAMINE INTERACTIONS IN MIDBRAIN
10
279
A10 regions (Khachaturian et al., '851, and these neurons are the source of both dynorphin-related peptides (e.g., dynorphin A and dynorphin B) and LE in the substantia nigra (Zamir et al., '84). These forebrain proenkephaliner- gic and prodynorphinergic nuclei probably provide an impor- tant source of enkephalinergic input to the nucleus A10 DA neurons.
Lack of coexistence of TH with enkephalin There was no evidence for coexistence of TH with enkeph-
alin in rat VTA neurons. However, in the ventrolateral medulla and nucleus of the solitary tract in the rat, both enkephalin and TH immunoreactivity were observed in the same perikarya and terminals (Pickel et al., '89; Milner et al., '89). I t may be that opioid peptides coexist with norepinephrine or epinephrine in these latter cells, since such coexistence has been observed previously in peripheral sympathetic nerves supplying the vas deferens (De Potter et al., '87; Klein et al., '84), and in the adrenal medulla (Viveros et al., '79). The present results suggest that such coexistence does not occur for opioids in the nucleus A10 DA neurons.
Conclusions Our findings demonstrate that enkephalin-containing
fibers and nerve terminals are in close proximity to the DA neurons that constitute the mesocorticolimbic DA system. At the ultrastructural level, enkephalin-containing nerve terminals a) form predominantly symmetric axo-dendritic synapses with A10 DA dendrites; b) form direct membrane appositions with unlabeled nerve terminals that are di- rectly apposed to DA dendrites, suggestive of axo-axonic connections; c) can contact more than one DA dendrite; d) contain more than one size synaptic vesicle, suggesting that the terminals contain another neurotransmitter; and e) are invested with a glial membrane near the synapse with the DA dendrites. The existence of enkephalin-DA synaptic contacts suggests that opioid peptides can directly influence A10 DA neurons, modulate the coupling of the DA dendro- dendritic contacts (Grace and Bunney, '83; Bayer and Pickel, 'go), and/or modulates the general environment via interactions with glial cells.
40 t 1 L I 1.4 1.7 2.0 2.3 2.6 2.9
A1 0 Oendritic Size (pM)
40 1 I -
; 30 e a 20 '
P)
10 '
0 0.2 0.5 0.8 1.1 1.4 1.7 2.0 2.3 2.6 2.9
Size of A10 Dendrites Adjacent t o LE Contacts (pM)
Fig. 6. Histograms illustrating the diameter of tyrosine hydroxylase immunoreactive dendrites in nucleus A10 (upper panel: n = loo), and of tyrosine hydroxylase immunoreactive dendrites tha t receive LE-containing synaptic contacts or appositions (lower panel; n = 63).
receptors are also located on non-DA neurons. More re- cently, Dilts and Kalivas ('89) used receptor autoradiogra- phy to demonstrate that mu receptors are present in the VTA even after the DA neurons were destroyed by 6-hy- droxydopamine. These data suggest that opioid peptides indirectly influence the DA neurons via mu receptors located on non-DA neurons.
Origin of enkephalinergic inputs to nucleus A10
There are numerous nuclei which send afferents to nucleus A10 (Phillipson, '79b; see review by Oades and Halliday, '87). There appears to be reciprocal relations between areas innervated by cells in the nucleus A10 and areas which project back to the A10 area. For example, nucleus A10 cells project to such areas as the nucleus accumbens, corpus striatum, olfactory tubercle, lateral septa1 area, prefrontal cortex, cingulate cortex, entorhinal cortex, and most of these areas project back to the A10 area (Phillipson, '79b; Swanson, '82).
Messenger-RNA for proenkephalin is found in cells in the striatum, olfactory tubercle, and nucleus accumbens (Kha- chaturian et al., '85), three regions known to project to nucleus A10. Likewise, pro-dynorphinergic neurons are found in the striatum which project to the nucleus A9 and
ACKNOWLEDGMENTS This research was supported by the National Institute on
Drug Abuse (DA-05314). The authors thank Dr. Stanley Watson for his gift of rabbit leu-enkephalin antisera and Ms. Judy Burdette for secretarial assistance.
LITERATURE CITED Albanese, A,, and D. Minciacchi (1983) Organization of the ascending
projections from the ventral tegmental area: A multiple fluorescent retrograde tracer study in the rat. J. Comp. Neurol. 216:406-420.
Altschuler, R.A., K.A. Reeks, J. Fex, and D.W. Hoffman (1988) Lateral olivocochlear neurons contain both enkephalin and dynorphin immuno- reactivities immunocytochemical colocalization studies. J. Histochem. Cytochem .32: 79 7-80 1.
Bayer, V.E., and V.M. Pickel (1990) Ultrastructural localization of tyrosine hydroxylase in the rat ventral tegmental area: Relationship between immunolabeling density and neuronal association. J. Neurosci. I0:2996- 3013.
Bayer, V.E., and V.M. Pickel (1991) GABA-labeled terminals form proportion- ally more synapses with dopaminergic neurons containing low densities of tyrosine hydroxylase-immunoreactivity in rat ventral tegmental area. Brain Res. 559:44-55.

280 C.-L. LIANG ET AL.
Blanc, G., D. Herve, H. Simon, A. Lisoprowski, J. Glowinski, and J.P. Tassin (1980) Response to stress of mesocortico-frontal dopaminergic neurons in rats after long-term isolation. Nature 284:265-267.
Bolam, J.P., and Y. Smith (1990) The GABA and substance P input to dopaminergic neurones in the substantia nigra of the rat. Brain Res. 529:57-78.
Bozarth, M.A., and R.A. Wise (1978) Intracranial self-administration of morphine into the ventral tegmental area. Life Sci. 28:551-555.
Bunn, S.J., M.R. Hanley, and G.P. Wilkin (1985) Evidence for a kappa-opioid receptor on pituitary astrocytes: An autoradiographic study. Neurosci. Lett. 55:317-323.
Chan, J., C. Aoki, and V.M. Pickel (1990) Optimization of differential immunogold, silver and peroxidase labeling with maintenance of ultra- structure in brain sections before plastic embedding. J . Neurosci. Methods 33:113-127.
Chang, H.T. (1988) Substance P-dopamine relationship in the rat substantia nigra: A light and electron microscopy study of double immunocytochem- ically labeled materials. Brain Res. 448r391-396.
Cohen, R.S., R.K. Carlin, D.J. Grab, and P. Siekevitz (1982) Phosphopro- teins in postsynaptic densities. Prog. Brain Res. 56:49-76.
Crow, T.J. (1972) A map of the rat mesencephalon for electrical self- stimulation. Brain Res. 36.265-273.
Dahlstrom, A,, and K. Fuxe (1964) Evidence for the existence of monoamine- containing neurons in the central nervous system I. Demonstration of monoamines in the cell bodies of brain-stem neurons. Acta. Physiol. Scand. 62:l-55.
De Potter, W.P., E.P. Coen, and R.W. De Potter (1987) Evidence for the coexistence and corelease of Met-enkephalin and noradrenaline from sympathetic nerves of the bovine vas deferens. Neuroscience 20:855- 866.
Di Chiara, G., and A. Imperato (1988) Drugs abused by humans preferen- tially increase synaptic dopamine concentrations in the mesolimbic system of freely moving rats. Proc. Natl. Acad. Sci. U.S.A. 85:5274-5278.
Di Figlia, M., N. Aronin, and J.B. Martin (1982) Light and electron microscopic localization of immunoreactive leu-enkephalin in the mon- key basal ganglia. J. Neurosci. 2:303-320.
Dilts, R.P., and P.W. Kalivas (1989) Autoradiographic localization of F-opi- oid and neurotensin receptors within mesolimbic dopamine system. Brain Res. 488:311-327.
Dilts, R.P., and P.W. Kalivas (1990) Autoradiographic localization of delta opioid receptors within the mesocorticolimbic dopamine system using radioiodinated l2-D-penicillamine, 5-D-penicillaminel enkephalin. Syn- apse 6:121-132.
Emson, P.C., A. Arrequi, V. Clement-Jones, B.E.B. Sandberg. and M. Rosser (1980) Regional distribution of methionine-enkephalin and substance P-like immunoreactivity in normal human brain and in Huntington's disease. Brain Res. 199:147-160.
Fallon, J.H., and P. Ciofi (1990) Dynorphin-containing neurons. In A. Bjorklund, T. Hokfelt, and M.J. Kuhar (eds): Neuropeptides in the CNS, Handbook of Chemical Neuroanatomy. New York: Elsevier, Vol. 9, part 2, pp. 1-130.
Fallon, J.H., and F.M. Leslie (1986) Distribution of dynorphin and enkepha- lin peptides in the rat brain. J. Comp. Neurol. 249.293-336.
Fallon, J.H., and R.Y. Moore (1978) Catecholamine innervation of the basal forebrain. IV. Topography of the dopamine projection to the basal forebrain and neostriatum. J . Comp. Neurol. 180:545-580.
Fink, J.S., and G.P. Smith (1984) Mesolimbocortical dopamine terminal fields are necessary for normal locomotor and investigatory exploration in rats. Brain Res. 199:356-384.
Finley, J.C.W., J.L. Maderdrut, and P. Petrusz (1981) The immunocytochem- ical localization ofenkephalin in the central nervous system of the rat. J. Comp. Neurol. 1.98.541-565.
Gaspar, P., 8. Berger, M. Gay, M. Hamon, F. Cesselin, A. Vigny, F. Javny-Agid, and Y. Agid (1983) Tyrosine hydroxylase and methionine- enkephalin in the human mesencephalon. J. Neurol. Sci. 58947-267.
German, D.C., and D.M. Bowden (1974) Catecholamine systems as the neural substrate for intracranial self-stimulation: A hypothesis. Brain Res. 73:381419.
German, D.C., and K.F. Manaye (1993) Midbrain dopaminergic neurons (nuclei A8, A9 and A10): 3-dimensional reconstruction in rat. J. Comp. Neurol., 331297-309.
German, D.C., D.S. Schlusselberg, and D.J. Woodward (1983) Three- dimensional computer reconstruction of midbrain dopaminergic neuro- nal populations: From mouse to man. J. Neural Transni. 57.243-254.
German, D.C., S.G. Speciale, K.F. Manaye, and M. Sadeq (1993) Opioid receptors in midbrain dopaminergic regions of the rat. I. Mu receptor autoradiography. J. Neural Transm. 91:39-52.
Grace, A.A., and B.S. Bunney (1983) Intracellular and extracellular electro- physiology of nigral dopaminergic neurons. 3. Evidence for electrotonic coupling. Neuroscience 101333-348.
Gray, E.G. (1969) Electron microscopy of excitatory and inhibitory synapses: a brief review. In K. Akert and P. Waser (eds): Mechanisms of Synaptic Transmission. Prog. Brain Res. 31, Amsterdam: Elsevier, pp. 141-155.
Guthrie, J., and A.I. Basbaum (1984) Colocalization of immunoreactive proenkephalin and prodynorphin products in medullary neurons of the rat. Neuropeptides 4:437-445.
Gysling, K., and R.Y. Wang (1983) Morphine-induced activation of A10 dopamine neurons in the rat. Brain Res. 277:119-127.
Haber, S., and R. Elde (1982) The distribution of enkephalin immunoreac- tive fibers and terminals in the monkey central nervous system: An immunohistochemical study. Neuroscience 7; 1049-1095.
Haber, S.N., and H.J. Groenewegen (1989) Interrelationship of the distribu- tion of neuropeptides and tyrosine hydroxylase immunoreactivity in the human substantia nigra. J. Comp. Neurol. 290:53-68.
Herve, D., V.M. Pickel, T.H. Joh, and A. Beaudet (1987) Serotonin axon terminals in the ventral tegmental area of' the rat: Fine structure arid synaptic input to dopaminergic neurons. Brain Res. 435:71-83.
Hokfelt, T., R. Martensson, A. Bjorklund, S. Kleinan, and M. Goldstein (1984) Distributional maps of tyrosine-hydroxylase-immunoreactive neu- rons in the rat brain. In A. Bjorklund and T. Hokfelt (eds): Classical Transmitters in the CNS, Handbook of Chemical Neuroanatomy. New York: Elsevier, Vol. 2, Part 1, pp. 277-379.
Hommer, D.W., and A. Pert (1983) The actions of opiates in the rat substantia nigra: An electrophysiological analysis. Peptides 4:603-608.
Hosli, L., E. Hosli, C. Zehnter, R. Lehmann, and T.W. Lutz (1982) Evidence for the existence of alpha- and beta-adrenoceptors on cultured glia cells: An electrophysiological study. Neuroscience 7:2867-2872.
Hsu, S.M., L. Raine, and H. Fanger (1981) The use of Avidin-Biotin- Peroxidase Complex (ABC) in immunoperoxidase techniques: A compar- ison between ABC and unlabeled antibody (PAP) procedures. J. Histo- chem. Cytochem. 29r577.
Inagaki, S., Y. Kubota, and S. Kit0 (1986) Ultrastructural localization of enkephalin immunoreactivity in the substant,ia nigra of the mnnhey Brain Res. 362:171-174.
Inagaki, S., and A. Parent (1984) Distribution of substance P and enkephalin- like immunoreactivity in the substantia nigra of rat, cat and monkey. Brain Res. Bull. I3:319-329.
Johnson, R.P., M. Sar, and W.E. Stumpf (1980) A topographic localization of enkephalin on the dopamine neurons of the rat substantia nigra and ventral tegmental area demonstrated by combined histofluorescence immunocytochemistry. Brain Res. 194:506-571.
Johnson, S.W., and R.A. North (1992) Opioids excite dopamine neuron3 by hyperpolarization of local interneurons. J . Neurosci. 12:483488.
Joyce, E.M., and S.D. Iversen (1979) The effect of morphine applied locally to mesencephalic dopamine cell bodies on spontaneous motor activity in the rat. Neurosci. Lett. 14:207-212.
Kalivas, P.W. (1985) Interactions between neuropeptides and doparnine neurons in the ventromedial mesencephalon. Neurosci. Biobehav. Rev, 9t573-587.
Kalivas, P.W., and R. Abhold (1987) Enkephalin release into the ventral tegmental area in response to stress: Modulation of mesocorticolimbic dopamine. Brain Res. 414:339-348.
Kalivas, P.W., E. Widerlov, D. Stanley, G.R. Breese, and A. Prange (1983) Enkephalin action on the mesolimbic system: A dopamine-dependent and dopamine-independent increase in locomotor activity. J. Pharmacol. Exp. Ther. 227.229-237.
Kelley, A.E., L. Stinus, and S.D. Iversen (1980) Interaction between D-Ala-Met-enkephalin, A10 dopaminergic neurons, and spontaneous behavior in the rat. Behav. Brain Res. 1:3-24.
Kelly, P.H., P.W. Seviour, and S.D. Iversen (1975) Amphetamine and apomorphine responses in the rat following 6-OHDA lesions of the nucleus accumbens septi and corpus striatum. Brain Res. 94.-507-522.
Khachaturian, H., M.E. Lewis, M.H. Schafer, and S.J. Watson (1985) Anatomyof the CNS opioid systems. Trends Neurosci. 8:111-119.
Klein, R.L., S. Lemaire, A. Thureson-Klein, and R. Day (1984) Leu- enkephalin, dynorphin and bombesin contents of a highly purified large dense cored noradrenergic vesicle fraction from bovine splenic nerve. In F. Frazoli, A. Esidori, and M. Mazzetti (eds): Opioid Peptides in the Periphery. Amsterdam: Elsevier, pp. 205-212.

OPIOID-DOPAMINE INTERACTIONS IN MIDBRAIN 281
Lacey, M.G., N.B. Mercuri, and R.A. North (1989) Two cell types in rat substantia nigra zona compacta distinguished by membrane properties and the actions of dopamine and opioids. J. Neurosci. 9:1233-1241.
Liang, C.-L., and D.C. German (1991) Relationship between leu-enkephalin afferents and A10 dopaminergic neurons: Light and electron microscopic examination. SOC. Neurosci. Abstr. 17,1275.
Llorens-Cortes, C., H. Pollard, and J.C. Schwartz (1979) Localization of opiate receptors in substantia nigra: Evidence by lesion studies. Neuro- sci. Lett. 12:165-170.
Loughlin, S.E., and J.H. Fallon (1984) Suhstantia nigra and ventral tegmen- tal area projections to cortex: Topography and collateralization. Neurosci- ence 11,4254355.
Mahalik, T. (1988) Direct demonstration of interactions between sub- stance-P immunoreactive terminals and tyrosine hydroxylase immunore- active neurons in the substantia nigra of the rat: An ultrastructural study. Synapse 2,508-515.
Maley, B.E. (1985) The ultrastructural localization of enkephalin and substance P immunoreactivity in the nucleus tractus solitarii of the cat. J. Comp. Neurol. 233:490-495.
Marshall, P.E., D.M.D. Landis, and E.L. Zalneraitis (1983) Immunocytochem- ical studies of substance P and leucine-enkephalin in Huntington's disease. Brain Res. 289:ll-26.
Matthews, R.T., and D.C. German (1984) Electrophysiological evidence for excitation of ventral tegmental area dopamine neurons hy morphine. Neuroscience 11:617-625.
Milner, T.A., V.M. Pickel, and D.J. Reis (1989) Ultrastructural basis for interactions between central opioids and catecholamines. I. Rostral ventrolateral medulla. J. Neurosci. 92 114-2 130.
Namand, D., H. Simon, J.P. Herman, and M. LeMoal(1984) Contributions of the mesencephalic dopaminergic system and the trigeminal sensory pathway to the ventral tegmental aphagia syndrome in rats. Physiol. Behav. 33:879-887.
Oades, R.D., and G.M. Halliday (1987) Ventral tegmental (A101 system: Neurobiology, anatomy and connectivity. Brain Res. Rev. 12,117-165.
Phillipson, O.T. (1979a) The cytoarchitecture of the interfascicular nucleus and the ventral tegmental area of Tsai in the rat. J. Comp. Neurol. 18 7:85-98.
Phillipson, O.T. (197913) Afferent projections to the ventral tegmental area of Tsai and interfascicular nucleus: A horseradish peroxidase study in the rat. J. Comp. Neurol. 187:117-144.
Pickel, V.M., J. Chan, and T.A. Milner (1989) Ultrastructural basis for interactions between central opioids and catecholamines. 11. Nuclei of the solitary tracts. J. Neurosci. 9,2419-2535.
Pickel, V.M., D.J. Reis, and S.E. Leeman (1977) Ultrastructural localization of substance P i n neurons of rat spinal cord. Brain Res. 122534-540.
Pickel, V.M., K.K. Sumal, S.C. Beckley, R.J. Miller, and D.J. Reis (1980) Immunocytochemical localization of enkephalin in the neostriatum of rat brain: A light and electron microscopic study. J. Comp. Neurol. 189:721-740.
Pijnenburg, A.J.J., W.M.M. Honig, J.A.M. Van Der Heyden, and J.M. Van Rossum (1976) Effects of chemical stimulation of the mesolimbic dopamine system upon locomotor activity. Eur. J. Pharmacol. 35:45-58.
Pioro, E.P.J., J.T. Hughes, and A.C. Cuello (1984) Loss of substance P and enkephalin immunoreactivity in the human substantia nigra after striato-pallidal infarction. Brain Res. 292339-347.
Pollard, H., C. Llorens, J.C. Schwartz, C. Gros, and E. Dray (1978) Localization of opiate receptors and enkephalins in the rat striatum in
relationship with the nigrostriatal dopaminergic system: Lesion studies, Brain Res. 151:392-398.
Rainer, A. (1986) The co-occurrence of substance P-like immunoreactive and dynorphin-like immunoreactivity in striatopallidal and striatonigral projection neurons in birds and reptiles. Exp. Brain Res. I1:229-248.
Reynolds, E.S. (1963) The use of lead citrate a t high pH as an electron- opaque stain in electron microscopy. J. Cell Biol. 17208-212.
Rossier, J. (1982) Opioid peptides have found their roots. Nature 298:221- 222.
Rougon, G., M. Noble, and A.W. Mudge (1983) Neuropeptides modulate the beta-adrenergic response of purified astrocytes in vitro. Nature 305:715- 717.
Sasek, C.A., and R.P. Elde (1986) Coexistence of enkephalin and dynorphin immunoreactivities in neurons in the dorsal gray commissure of the sixth lumbar and first sacral spinal cord segments in rat. Brain Res. 381:8-14.
Sesack, S.R., and V.M. Pickel (1992) Dual ultrastructural localization of enkephalin and tyrosine hydroxylase immunoreactivity in the rat ventral tegmental area: Multiple substrates for opiate-dopamine interactions. J. Neuroscience 121335-1350.
Snyder, S.H., S.P. Banerjee, H.T. Yamamura, and D. Greenherg (1974) Drugs, neurotransmitters and schizophrenia. Science 184: 1243-1253.
Somogyi, P., J.V. Priestly, A.C. Cuello, A.D. Smith, and H. Takagi (1982) Synaptic connections of enkephalin-immunoreactive nerve terminals in the neostriatum: A correlated light and electron microscopic study. J. Neurocytol. 11:779-807.
Speciale, S.G., K.F. Manaye, M. Sadeq, and D.C. German 11993) Opioid receptors in midbrain dopaminergic neurons of the rat 11. Delta and kappa receptor autoradiography. J. Neural Transm. 91:53-66.
Starr, M.S. (1985) Multiple opiate receptors may be involved in suppressing 7-aminobutyrate release in substantia nigra. Life Sci. 37:2249-2255.
Sumal, K.K., V.M. Pickel, R.J. Miller, and D.J. Reis (1982) Enkephalin- containing neurons in substantia gelatinosa of spinal trigeminal com- plex: Ultrastructure and synaptic interaction with primary sensory afferents. Brain Res. 248:223-236.
Swanson, L.W. (1982) The ventral tegmental area and adjacent regions: A combined fluorescent retrograde tracer and immunofluorescence study in the rat. Brain Res. Bull. 9:321-353.
Uchizono, K. (1965) Characteristics of excitatory and inhibitory synapses in the CNS of the cat. Nature 207:642-643.
Uhl, G.R., R.R. Goodman, M.J. Kuhar, S.R. Childress, and S.H. Snyder (1979) Immunohistochemical mapping of enkephalin-containing cell, fiber and nerve terminals in the brainstem of the rat. Brain Res. 166t75-94.
Ungerstedt, U. (1971) Stereotaxic mapping of the monoamine pathways in the rat brain. Acta Physiol. Scand. (Suppl.) 367:l-48.
Vezina, P., P.W. Kalivas, and J. Stewart (1987) Sensitization occurs to the locomotor effects of morphine and the specific opioid receptor agonist, DAGO administered repeatedly to the ventral tegmental area but not to the nucleus accumbens. Brain Res. 41 7:51-58.
Viveros, O.H., E.J. Diliberto, Jr., E. Hazum, and K . J . Chang (1979) Opiate-like materials in the adrenal medulla: Evidence for storage and secretion with catecholamines. Mol. Pharmacol. 16t1101-1108.
Zamir, N., M. Palkovits, E. Weher, E. Mezey, and M.J. Brownstein (1984) A dynorphinergic pathway of leu-enkephalin production in rat substantia nigra. Nature 307:643-645.




















