
The unconditioned fear produced by morphine withdrawal is regulated by μ- and κ-opioid receptors...
7
Behavioural Brain Research 204 (2009) 140–146 Contents lists available at ScienceDirect Behavioural Brain Research journal homepage: www.elsevier.com/locate/bbr Research report The unconditioned fear produced by morphine withdrawal is regulated by - and -opioid receptors in the midbrain tectum Jana De Ross a,b , Milton A.V. Ávila a,b , Rafael N. Ruggiero a,b , Manoel J. Nobre a,b , Marcus L. Brandão a,b , Vanessa M. Castilho a,b,∗ a Laboratory of Psychobiology, FFCLRP, University of São Paulo, Ribeirão Preto 14040-901, SP, Brazil b Institute of Neuroscience and Behavior, Campus USP, Ribeirão Preto 14040-901, SP, Brazil article info Article history: Received 6 February 2009 Received in revised form 26 May 2009 Accepted 27 May 2009 Available online 9 June 2009 Keywords: Anxiety Dorsal periaqueductal gray Fear Inferior colliculus Morphine withdrawal Opioid receptors abstract We have recently shown that morphine withdrawal sensitizes the neural substrates of fear in the midbrain tectum structures—the dorsal periaqueductal gray (dPAG) and inferior colliculus (IC). In the present study, we investigated the role of - and -opioid receptors in the mediation of these effects. Periadolescent rats chronically treated with morphine (10mg/kg; s.c.) twice daily for 10 days were implanted with an electrode glued to a guide-cannula into the dPAG or the IC. Forty-eight hours after the interruption of this treatment, the effects of intra-dPAG or intra-IC microinjections of [D-Ala 2 ,N-Me-Phe 4 ,Gly 5 -ol]-enkephalin (DAMGO; 0.6 and 1 nmol/0.2 l) – a selective -receptor agonist – or nor-binaltorphimine (BNI; 2.5 and 5 g/0.2 l) – a selective -receptor antagonist with tardive action – on the freezing and escape thresholds determined by electrical stimulation of the dPAG and the IC were examined. For both structures, morphine withdrawal produced pro-aversive effects. DAMGO and BNI had antiaversive effects when injected into the dPAG and IC of non-dependent rats. In morphine-withdrawn rats, only BNI continued to promote antiaversive effects in both structures. Whereas DAMGO lost its antiaversive efficacy when injected into the dPAG, only its highest dose promoted antiaversive effects in the IC of morphine-withdrawn rats, suggesting the development of an apparent tolerance. Thus, the enhanced reactivity of the midbrain tectum in morphine-withdrawn periadolescent rats may be due, at least partially, to an impairment of the inhibitory influence of mechanisms mediated by -receptors on the neural substrates of fear in this region. © 2009 Elsevier B.V. All rights reserved. 1. Introduction The development of drug dependence is characterized by the emergence of an abstinence syndrome when chronic drug use is interrupted. In addition to its peculiar somatic signs [31,45], opiate withdrawal promotes disturbances in affective states and emotion- ality [29,46,53]. Neurochemical and immunohistochemical studies on opiate dependence have indicated that two different neural systems underlie the physical and affective aspects of opiate with- drawal. In general, it has been suggested that affective signs, such as conditioned place aversion, are mainly mediated by the central nucleus of the amygdala, the shell region of the nucleus accum- bens and the bed nucleus of the stria terminalis [10,24,54]. On the other hand, the main neural substrates mediating the somatic aspects of opiate dependence are proposed to be localized in the ∗ Corresponding author at: Laboratory of Psychobiology, FFCLRP, University of São Paulo, Ribeirão Preto 14040-901, SP, Brazil. Tel.: +55 1636023788; fax: +55 1636024830. E-mail address: [email protected] (V.M. Castilho). hypothalamus, the locus coeruleus and the periaqueductal gray (PAG) [10,24,31], particularly in its lateral and ventrolateral columns [7,27]. Anxiety is one of the main affective symptoms of withdrawal from opiates [29] and other drugs of abuse [15,25,32]. The avoidance of this unpleasant symptom is hypothesized to negatively reinforce continuous drug use and to be one of the main components of crav- ing for the drug [30,44]. In this context, withdrawal from drugs of abuse, such as morphine, may function as an unconditioned stres- sor, and as such can promote its negative emotional effects through the activation of midbrain structures involved with the organiza- tion of fear states. Possible loci for the origin of these aversive effects are the dorsal PAG (dPAG) and the inferior colliculus (IC) [12,14]. Electrical or chemical stimulation of these structures elicits behav- ioral and autonomic responses similar to reactions displayed by animals facing dangerous stimuli commonly found in their natu- ral environment [12,16,19]. A recent study has demonstrated that even in the absence of intense somatic withdrawal symptoms, anx- iety induced by morphine-withdrawal goes along with pronounced neural activation of the dPAG, as detected by Fos-protein expression [20]. 0166-4328/$ – see front matter © 2009 Elsevier B.V. All rights reserved. doi:10.1016/j.bbr.2009.05.033
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The unconditioned fear produced by morphine withdrawal is regulated by μ- and κ-opioid receptors...

R
Tb
JMa
b
a
ARRAA
KADFIMO
1
eiwaosdanbta
S1
0d
Behavioural Brain Research 204 (2009) 140–146
Contents lists available at ScienceDirect
Behavioural Brain Research
journa l homepage: www.e lsev ier .com/ locate /bbr
esearch report
he unconditioned fear produced by morphine withdrawal is regulatedy �- and �-opioid receptors in the midbrain tectum
ana De Ross a,b, Milton A.V. Ávila a,b, Rafael N. Ruggiero a,b, Manoel J. Nobre a,b,arcus L. Brandão a,b, Vanessa M. Castilho a,b,∗
Laboratory of Psychobiology, FFCLRP, University of São Paulo, Ribeirão Preto 14040-901, SP, BrazilInstitute of Neuroscience and Behavior, Campus USP, Ribeirão Preto 14040-901, SP, Brazil
r t i c l e i n f o
rticle history:eceived 6 February 2009eceived in revised form 26 May 2009ccepted 27 May 2009vailable online 9 June 2009
eywords:nxietyorsal periaqueductal grayearnferior colliculus
a b s t r a c t
We have recently shown that morphine withdrawal sensitizes the neural substrates of fear in the midbraintectum structures—the dorsal periaqueductal gray (dPAG) and inferior colliculus (IC). In the present study,we investigated the role of �- and �-opioid receptors in the mediation of these effects. Periadolescentrats chronically treated with morphine (10 mg/kg; s.c.) twice daily for 10 days were implanted with anelectrode glued to a guide-cannula into the dPAG or the IC. Forty-eight hours after the interruption of thistreatment, the effects of intra-dPAG or intra-IC microinjections of [D-Ala2,N-Me-Phe4,Gly5-ol]-enkephalin(DAMGO; 0.6 and 1 nmol/0.2 �l) – a selective �-receptor agonist – or nor-binaltorphimine (BNI; 2.5 and5 �g/0.2 �l) – a selective �-receptor antagonist with tardive action – on the freezing and escape thresholdsdetermined by electrical stimulation of the dPAG and the IC were examined. For both structures, morphinewithdrawal produced pro-aversive effects. DAMGO and BNI had antiaversive effects when injected into
orphine withdrawalpioid receptors
the dPAG and IC of non-dependent rats. In morphine-withdrawn rats, only BNI continued to promoteantiaversive effects in both structures. Whereas DAMGO lost its antiaversive efficacy when injected intothe dPAG, only its highest dose promoted antiaversive effects in the IC of morphine-withdrawn rats,suggesting the development of an apparent tolerance. Thus, the enhanced reactivity of the midbraintectum in morphine-withdrawn periadolescent rats may be due, at least partially, to an impairment ofthe inhibitory influence of mechanisms mediated by �-receptors on the neural substrates of fear in this
region.. Introduction
The development of drug dependence is characterized by themergence of an abstinence syndrome when chronic drug use isnterrupted. In addition to its peculiar somatic signs [31,45], opiate
ithdrawal promotes disturbances in affective states and emotion-lity [29,46,53]. Neurochemical and immunohistochemical studiesn opiate dependence have indicated that two different neuralystems underlie the physical and affective aspects of opiate with-rawal. In general, it has been suggested that affective signs, suchs conditioned place aversion, are mainly mediated by the central
ucleus of the amygdala, the shell region of the nucleus accum-ens and the bed nucleus of the stria terminalis [10,24,54]. Onhe other hand, the main neural substrates mediating the somaticspects of opiate dependence are proposed to be localized in the∗ Corresponding author at: Laboratory of Psychobiology, FFCLRP, University ofão Paulo, Ribeirão Preto 14040-901, SP, Brazil. Tel.: +55 1636023788; fax: +55636024830.
E-mail address: [email protected] (V.M. Castilho).
166-4328/$ – see front matter © 2009 Elsevier B.V. All rights reserved.oi:10.1016/j.bbr.2009.05.033
© 2009 Elsevier B.V. All rights reserved.
hypothalamus, the locus coeruleus and the periaqueductal gray(PAG) [10,24,31], particularly in its lateral and ventrolateral columns[7,27].
Anxiety is one of the main affective symptoms of withdrawalfrom opiates [29] and other drugs of abuse [15,25,32]. The avoidanceof this unpleasant symptom is hypothesized to negatively reinforcecontinuous drug use and to be one of the main components of crav-ing for the drug [30,44]. In this context, withdrawal from drugs ofabuse, such as morphine, may function as an unconditioned stres-sor, and as such can promote its negative emotional effects throughthe activation of midbrain structures involved with the organiza-tion of fear states. Possible loci for the origin of these aversive effectsare the dorsal PAG (dPAG) and the inferior colliculus (IC) [12,14].Electrical or chemical stimulation of these structures elicits behav-ioral and autonomic responses similar to reactions displayed byanimals facing dangerous stimuli commonly found in their natu-
ral environment [12,16,19]. A recent study has demonstrated thateven in the absence of intense somatic withdrawal symptoms, anx-iety induced by morphine-withdrawal goes along with pronouncedneural activation of the dPAG, as detected by Fos-protein expression[20].
rain R
iMIsaas�
eirraitfto�rtI
2
2
(RimftTrwG
2
(i1ts
2
mmCatwm
2
Dta((aittba
J. De Ross et al. / Behavioural B
It has been suggested that the defensive behavior integratedn the midbrain tectum is modulated by opioid mechanisms [12].
icroinjections of low doses of morphine into the dPAG and theC promote antiaversive effects, whereas high doses induce defen-ive responses [18,34]. Several studies have demonstrated that thentiaversive effects of low doses of opioids injected into the dPAGre mediated by the activation of �-receptors, whereas the aver-ive effects of high doses are probably due to the activation of-receptors [2,35,38,43].
We have recently shown that morphine withdrawal producesnhanced anxiety levels associated with an increase in the reactiv-ty of the dPAG and the IC to electrical stimulation in periadolescentats [3]. These findings suggested that the sensitization of the neu-al substrates of fear in these structures may underlie the increasednxiety induced by morphine withdrawal. In the present study, wenvestigated the role of �- and �-opioid receptors in the media-ion of the effects of morphine withdrawal on the unconditionedear responses induced by activation of the dPAG and the IC. Tohis end, we have studied the influence of local microinjectionsf [D-Ala2,N-Me-Phe4,Gly5-ol]-enkephalin (DAMGO) – a selective-receptor agonist – and nor-binaltorphimine (BNI) – a selective �-
eceptor antagonist with tardive action – on the freezing and escapehresholds determined by the electrical stimulation of the dPAG andC in non-dependent and morphine-withdrawn periadolescent rats.
. Materials and methods
.1. Subjects
One hundred and ninety-two male Wistar rats, ranging in age from 35 to 38 days180–210 g) at the beginning of treatment, from the animal house of the campus ofibeirão Preto, University of São Paulo, were used. They were housed in groups of four
n Plexiglas-walled cages lined with wood shavings that were changed every 3 days,aintained in a 12:12 dark/light cycle (lights on 07:00 h) at 24 ± 1 ◦C, and were given
ree access to food and water throughout the experiment. Before the beginning of thereatments, the animals had a 3-day habituation period to the lodging conditions.he experiments reported in this article were performed in compliance with theecommendations of the SBNeC (Brazilian Society for Neuroscience and Behavior),hich are in accordance with the rules of the U. S. National Institutes of Healthuidelines for Care and Use of Laboratory Animals.
.2. Drugs
Morphine sulfate (Cristalia, Brazil), [D-Ala2,N-Me-Phe4,Gly5-ol]-enkephalinDAMGO, Sigma, USA) and nor-binaltorphimine (BNI, Sigma, USA) were dissolvedn physiological saline (0.9%). The doses used were: 10 mg/kg morphine, 0.6 and.0 nmol DAMGO, 2.5 and 5 �g BNI. Physiological saline served as the vehicle con-rol. The drug doses and the time of administration were chosen based on previoustudies [3,35,39].
.3. Induction of morphine dependence and withdrawal
Rats received twice daily (09:00 and 19:00 h) subcutaneous (s.c.) injections oforphine (10 mg/kg) for 10 consecutive days. The last injection was given in theorning of the 10th day. This treatment regimen was based on a previous study [3].
ontrol animals received saline injections (untreated group). During treatment, thenimals were gently handled daily for 3 min immediately before drug administrationo reduce the stressful effects of the injection procedure. Withdrawal from morphineas achieved 48 h after the last injection. Two groups of animals (untreated andorphine) were assessed in all experiments in this study.
.4. Surgery
Surgery was performed 5 days before the end of the treatments (on day 5).ay 5 was chosen to allow the animals to recover from surgery for a week before
he beginning of the behavioral tests (after 48 h withdrawal). The animals werenesthetized with tribromoethanol (250 mg/kg, ip) and fixed in a stereotaxic frameInsight, São Paulo, Brazil). A chemitrode made of a stainless steel guide-cannulao.d. 0.6 mm, i.d. 0.4 mm) glued to an electrode was implanted in the midbrain,
imed at the dPAG or the IC. The electrode was made of stainless steel wire, 160 �mn diameter, insulated except at the cross-section of the tip reaching 1 mm belowhe lower end of the cannula. At the time of surgery, the rats weighed 220–250 g. Forhis weight range, the placement of the upper incisor bar was adjusted to 2.5 mmelow the interaural line so that the skull was horizontal between the bregmand lambda. The chemitrode was introduced using the following coordinates [40]esearch 204 (2009) 140–146 141
with the lambda serving as the reference for each plane: dPAG (with an angle of16◦) – anteroposterior = 0.1 mm, mediolateral = 1.9 mm, dorsoventral = 5.1 mm; IC –anteroposterior = −0.9 mm, mediolateral = 1.2 mm, and dorsoventral = 4.5 mm. Thechemitrode was fixed to the skull by means of acrylic resin and two stainless steelscrews. At the end of surgery, each guide-cannula was sealed with a stainless-steelwire to protect it from congestion. Each animal received an intramuscular injection(0.2 ml) of a veterinary pentabiotic (penicillin—120,000 UI plus 100 mg/ml of dihy-drostreptomycin sulfate and streptomycin sulfate) followed by an injection of theanti-inflammatory and analgesic banamine (flunixin meglumine, 2.5 mg/kg).
2.5. Measurement of the aversive thresholds
Forty-eight hours following the last saline or morphine injection, rats wereplaced in an experimental box (25 cm × 20 cm × 20 cm) with walls made of Plexi-glas. The floor was made of stainless steel bars of 5 mm in diameter, spaced 1.5 cmapart. This box was placed inside a sound attenuation chamber made of plywood,illuminated from within by a red 5 W lamp, with 65-dB permanent background noisedelivered by a fan. The rats were allowed a 5-min period of habituation in the appara-tus at the beginning of each experimental session. After that, the dPAG or IC electricalstimulation procedure was started. The brain was stimulated electrically by meansof a sine wave stimulator (Insight, São Paulo, Brazil), as previously described [19].The stimulation current was monitored by measuring the voltage drop across a 1Kresistor using an oscilloscope (Philips, USA). The brain is resistant to the passage ofelectrical current in the same way as a resistor is. 1K means one thousand ohms. Brainstimuli (alternate current—AC, 60 Hz, 15 s) were presented at 30-s intervals with thecurrent intensity increasing by steps of 5 �A for determination of the aversive thresh-olds. The freezing threshold was defined as the lowest intensity producing completeimmobility except for respiratory movements. The escape threshold was defined asthe lowest current intensity that produced running or jumping in two successiveascending series of electrical stimulation. Animals with a basal escape thresholdabove 150 �A (peak-to-peak) were discarded from the experiment. The behavior ofthe animals was recorded by a video-camera positioned beside the experimentalbox and monitored via a closed circuit TV-camera. The box was cleaned with a 5%ammonium solution immediately before each animal was placed within it.
2.6. Drug microinjection
The animals were gently wrapped in a cloth, held by hand, and a thin dentalneedle (o.d., 0.3 mm) was introduced though the guide-cannula until its lower endwas 1 mm below the guide-cannula, reaching the same depth as the electrode tip.The injection needle was linked to a 5-�l Hamilton syringe by means of polyethylenetubing (PE-10). A volume of 0.2 �l of DAMGO, BNI or vehicle was injected over 30 swith the aid of an infusion pump (Insight, Brazil) and the needle was held in place foran additional 30 s. The displacement of an air bubble inside the polyethylene tubingwas used to monitor the microinjection procedure.
2.7. Experimental procedure
On the 10th day, chronic treatment with morphine or saline was inter-rupted. At the end of the 48-h withdrawal period, the aversive thresholds weremeasured (baseline). Each dPAG or IC group (morphine and untreated) was sub-divided into saline and drug-treated groups. Fifteen minutes after intra-dPAG orintra-IC microinjections of saline or DAMGO (0.6 and 1 nmol/0.2 �l), the aver-sive thresholds were again determined. For each structure (dPAG or IC), thegroups assessed were: untreated × saline, untreated × DAMGO, morphine × saline,and morphine × DAMGO. In additional groups, the baseline of aversive thresholdswas measured at the end of a 24-h withdrawal period. Next, each dPAG or IC group(morphine and untreated) was treated with microinjections of saline or BNI (2.5 and5 �g/0.2 �l). Twenty-four hours later, the aversive thresholds were redetermined.The timing of BNI administration was based on previous evidence that although BNIacts as a non-selective antagonist of opioid receptors during the first hour after injec-tion, it selectively blocks �-receptors after 24 h [9]. For each structure (dPAG or IC),the groups assessed were: untreated × saline, untreated × BNI, morphine × saline,and morphine × BNI.
2.8. Histology
Upon completion of the experiments, the animals were deeply anesthetized withurethane and perfused intracardially with saline (0.9%) followed by buffered forma-lin solution (4%). The brains were removed, immersed in formalin solution for 2 hand then kept in a sucrose solution (30%). Three days later the brains were frozen.Serial 60-�m brain sections were cut using a cryostat (Leica, Wetzlar, Germany) andstained with Nissl to localize the positions of the electrode tips according to thePaxinos and Watson’s atlas [40]. Data from rats with electrode tips located outsidethe dPAG or IC were not included in the statistical analysis.
2.9. Statistical analysis
The results obtained are shown as mean + S.E.M. For each structure (dPAG or IC),the data were subjected to a three-way mixed design ANOVA with groups (untreatedvs. morphine) and agonist/antagonist treatment (DAMGO vs. saline and BNI vs. saline)
1 rain R
atil
3
3
wabsta
3
o
Fid
42 J. De Ross et al. / Behavioural B
s between-subject factors, and condition (baseline vs. test measures of the aversivehresholds) as the within-subject repeated factor. Newman–Keuls’ post hoc compar-sons were carried out whenever significant p-values were obtained. A probabilityevel of p < 0.05 was considered to be significant.
. Results
.1. Electrical stimulation of the dPAG and IC
As the intensity of the current applied to the dPAG or the ICas increased, the animals suddenly stopped, became immobile
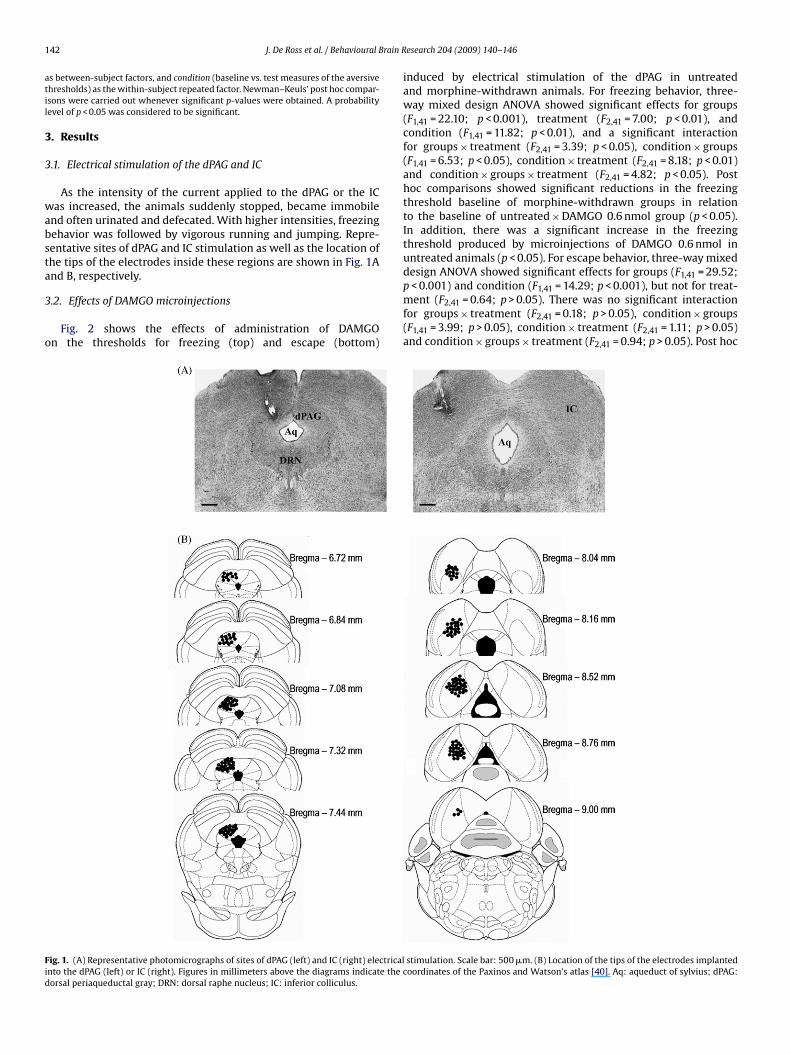
nd often urinated and defecated. With higher intensities, freezingehavior was followed by vigorous running and jumping. Repre-entative sites of dPAG and IC stimulation as well as the location ofhe tips of the electrodes inside these regions are shown in Fig. 1And B, respectively.
.2. Effects of DAMGO microinjections
Fig. 2 shows the effects of administration of DAMGOn the thresholds for freezing (top) and escape (bottom)
ig. 1. (A) Representative photomicrographs of sites of dPAG (left) and IC (right) electricalnto the dPAG (left) or IC (right). Figures in millimeters above the diagrams indicate the corsal periaqueductal gray; DRN: dorsal raphe nucleus; IC: inferior colliculus.
esearch 204 (2009) 140–146
induced by electrical stimulation of the dPAG in untreatedand morphine-withdrawn animals. For freezing behavior, three-way mixed design ANOVA showed significant effects for groups(F1,41 = 22.10; p < 0.001), treatment (F2,41 = 7.00; p < 0.01), andcondition (F1,41 = 11.82; p < 0.01), and a significant interactionfor groups × treatment (F2,41 = 3.39; p < 0.05), condition × groups(F1,41 = 6.53; p < 0.05), condition × treatment (F2,41 = 8.18; p < 0.01)and condition × groups × treatment (F2,41 = 4.82; p < 0.05). Posthoc comparisons showed significant reductions in the freezingthreshold baseline of morphine-withdrawn groups in relationto the baseline of untreated × DAMGO 0.6 nmol group (p < 0.05).In addition, there was a significant increase in the freezingthreshold produced by microinjections of DAMGO 0.6 nmol inuntreated animals (p < 0.05). For escape behavior, three-way mixeddesign ANOVA showed significant effects for groups (F1,41 = 29.52;
p < 0.001) and condition (F1,41 = 14.29; p < 0.001), but not for treat-ment (F2,41 = 0.64; p > 0.05). There was no significant interactionfor groups × treatment (F2,41 = 0.18; p > 0.05), condition × groups(F1,41 = 3.99; p > 0.05), condition × treatment (F2,41 = 1.11; p > 0.05)and condition × groups × treatment (F2,41 = 0.94; p > 0.05). Post hocstimulation. Scale bar: 500 �m. (B) Location of the tips of the electrodes implantedoordinates of the Paxinos and Watson’s atlas [40]. Aq: aqueduct of sylvius; dPAG:

J. De Ross et al. / Behavioural Brain Research 204 (2009) 140–146 143
Fig. 2. Freezing (top) and escape (bottom) thresholds determined by the proce-dure of electrical stimulation of the dorsal periaqueductal gray (dPAG) in untreatedand morphine-withdrawn animals. Columns represent mean + S.E.M. of the aversivethresholds measured before (baseline) and after (test) microinjections of saline orDAMGO (0.6 or 1 nmol) into the dPAG. *p < 0.05 in relation to the untreated group;#
t0m
cboib
ttastp(bdpbltb(stp(ptp
Fig. 3. Freezing (top) and escape (bottom) thresholds determined by the procedureof electrical stimulation of the inferior colliculus (IC) in untreated and morphine-withdrawn animals. Columns represent mean + S.E.M. of the aversive thresholdsmeasured before (baseline) and after (test) microinjections of saline or DAMGO (0.6
#
p < 0.05 in relation to the baseline of the same group, according to Newman–Keuls’est. Mor: morphine. Saline: n = 8 for untreated and n = 7 for morphine; DAMGO.6 nmol: n = 8 for both groups; DAMGO 1 nmol: n = 7 for untreated and n = 9 fororphine.omparisons showed significant reductions in the escape thresholdaseline of morphine-withdrawn groups in relation to the baselinef untreated groups (p < 0.05). Furthermore, there were significant
ncreases in the escape threshold produced by microinjections ofoth doses of DAMGO in untreated animals (p < 0.05).
Fig. 3 shows the effects of the administration of DAMGO on thehresholds for freezing (top) and escape (bottom) induced by elec-rical stimulation of the IC in untreated and morphine-withdrawnnimals. For freezing behavior, three-way mixed design ANOVAhowed significant effects for groups (F1,48 = 25.99; p < 0.001),reatment (F2,48 = 4.31; p < 0.05), and condition (F1,48 = 15.69;< 0.001), and a significant interaction for groups × treatment
F2,48 = 4.31; p < 0.05) and condition × groups (F1,48 = 4.14; p < 0.05),ut not for condition × treatment (F2,48 = 2.69; p > 0.05) or con-ition × groups × treatment (F2,48 = 2.23; p > 0.05). Post hoc com-arisons showed significant reductions in the freezing thresholdaseline of morphine-withdrawn groups in relation to the base-
ine of untreated × 1 nmol DAMGO group (p < 0.05). In addition,here were significant increases in the freezing threshold producedy microinjection of both doses of DAMGO in untreated animalsp < 0.05). For escape behavior, three-way mixed design ANOVAhowed significant effects for groups (F1,48 = 34.63; p < 0.001),reatment (F2,48 = 4.34; p < 0.05), and condition (F1,48 = 32.02;
< 0.001), and a significant interaction for condition × treatmentF2,48 = 11.00; p < 0.001), but not for groups × treatment (F2,48 = 0.07;> 0.05), condition × groups (F1,48 = 0.02; p > 0.05) or condi-
ion × groups × treatment (F2,48 = 2.52; p > 0.05). Post hoc com-arisons showed significant reductions in the escape threshold
or 1 nmol) into the IC. *p < 0.05 in relation to the untreated group; p < 0.05 in relationto the baseline of the same group, according to Newman–Keuls’ test. Mor: morphine.Saline: n = 11 for untreated and n = 12 for morphine; DAMGO 0.6 nmol: n = 7 for bothgroups; DAMGO 1 nmol: n = 8 for untreated and n = 9 for morphine.
baseline of morphine-withdrawn groups in relation to the baselineof untreated groups (p < 0.05). Furthermore, there were significantincreases in the escape thresholds produced by microinjectionsof both doses of DAMGO in untreated animals (p < 0.05), and bymicroinjections of 1 nmol DAMGO in morphine-withdrawn rats(p < 0.05).
3.3. Effects of BNI microinjections
Fig. 4 shows the effects of administration of BNI on thethresholds for freezing (top) and escape (bottom) induced byelectrical stimulation of the dPAG in untreated and morphine-withdrawn animals. For freezing behavior, three-way mixeddesign ANOVA showed significant effects for groups (F1,39 = 14.01;p < 0.001) and condition (F1,39 = 16.24; p < 0.001), but not fortreatment (F2,39 = 0.04; p > 0.05). There was a significant interac-tion for condition × treatment (F2,39 = 3.77; p < 0.05), but not forgroups × treatment (F2,39 = 1.71; p > 0.05), condition × groups(F1,39 = 1.21; p > 0.05), or condition × groups × treatment(F2,39 = 1.38; p > 0.05). Post hoc comparisons showed that morphinewithdrawal did not significantly change the freezing thresholdbaselines (p > 0.05). On the other hand, there were significantincreases in the freezing threshold produced by microinjec-
tions of 5 �g BNI in morphine-withdrawn animals (p < 0.05).For escape behavior, three-way mixed design ANOVA showedsignificant effects for groups (F1,39 = 21.67; p < 0.001) and condition(F1,39 = 48.68; p < 0.001), but not for treatment (F2,39 = 1.69; p > 0.05).There was a significant interaction for condition × treatment
144 J. De Ross et al. / Behavioural Brain Research 204 (2009) 140–146
Fig. 4. Freezing (top) and escape (bottom) thresholds determined by the proce-dure of electrical stimulation of the dorsal periaqueductal gray (dPAG) in untreatedand morphine-withdrawn animals. Columns represent mean + S.E.M. of the aversivethresholds measured before (baseline) and 24 h after (test) microinjections of salineor nor-binaltorphimine (BNI; 2.5 or 5 �g) into the dPAG. *p < 0.05 in relation to theuntreated group; #p < 0.05 in relation to the baseline of the same group, accordingtmu
(pdcboib(
ttasmT(ptichp(smap
Fig. 5. Freezing (top) and escape (bottom) thresholds determined by the pro-cedure of electrical stimulation of the inferior colliculus (IC) in untreated andmorphine-withdrawn animals. Columns represent mean + S.E.M. of the aversivethresholds measured before (baseline) and 24 h after (test) microinjections of salineor nor-binaltorphimine (BNI; 2.5 or 5 �g) into the IC. *p < 0.05 in relation to the
ulation of the dPAG and IC elicited defensive responses, such as
o Newman–Keuls’ test. Mor: morphine. Saline: n = 8 for untreated and n = 10 fororphine; BNI 2.5 �g: n = 7 for untreated and n = 5 for morphine; BNI 5 �g: n = 7 for
ntreated and n = 8 morphine.
F2,39 = 5.94; p < 0.01), but not for groups × treatment (F2,39 = 0.06;> 0.05), condition × groups (F1,39 = 0.83; p > 0.05), or con-ition × groups × treatment (F2,39 = 1.04; p > 0.05). Post hocomparisons showed significant reductions in the escape thresholdaseline of morphine-withdrawn groups in relation to the baselinef untreated groups (p < 0.05). In addition, there were significant
ncreases in the escape threshold produced by microinjections ofoth doses of BNI in untreated and morphine-withdrawn animalsp < 0.05).
Fig. 5 shows the effects of the administration of BNI on thehresholds for freezing (top) and escape (bottom) induced by elec-rical stimulation of the IC in untreated and morphine-withdrawnnimals. For freezing behavior, three-way mixed design ANOVAhowed significant effects for groups (F1,40 = 12.19; p < 0.01), treat-ent (F2,40 = 9.74; p < 0.001) and condition (F1,40 = 20.64; p < 0.001).
here was a significant interaction for condition × treatmentF2,40 = 7.59; p < 0.01), but not for groups × treatment (F2,40 = 0.73;> 0.05), condition × groups (F1,40 = 2.18; p > 0.05), or condi-
ion × groups × treatment (F2,40 = 1.36; p > 0.05). Post hoc compar-sons showed that morphine withdrawal did not significantlyhange the freezing threshold baselines (p > 0.05). On the otherand, there was a significant increase in the freezing thresholdroduced by microinjections of 5 �g BNI in untreated animalsp < 0.05). For escape behavior, three-way mixed design ANOVA
howed significant effects for groups (F1,40 = 35.21; p < 0.001), treat-ent (F2,40 = 5.12; p < 0.05), and condition (F1,40 = 67.63; p < 0.001),nd a significant interaction for condition × groups (F1,40 = 5.33;< 0.05), condition × treatment (F2,40 = 25.02; p < 0.001) and con-
untreated group; #p < 0.05 in relation to the baseline of the same group, accordingto Newman–Keuls’ test. Mor: morphine. Saline: n = 10 for untreated and n = 11 formorphine; BNI 2.5 �g: n = 6 for untreated and n = 5 morphine; BNI 5 �g: n = 7 forboth groups.
dition × groups × treatment (F2,40 = 3.28; p < 0.05), but not forgroups × treatment (F2,40 = 0.63; p > 0.05). Post hoc comparisonsshowed significant reductions in the escape threshold baselineof morphine-withdrawn groups in relation to the baseline ofuntreated groups (p < 0.05). Furthermore, there were significantincreases in the escape thresholds produced by microinjections of5 �g BNI in untreated and morphine-withdrawn animals (p < 0.05).
4. Discussion
It has been proposed that midbrain tectum structures, suchas the dPAG and the IC, are critically involved in the regulationof unconditioned defensive responses to impending danger stim-uli [12]. These structures act in concert with prosencephalic areassuch as the hypothalamus and the amygdala in the elaboration ofdefensive behavior and it has been proposed that malfunctioningof this system is involved in the genesis of certain anxiety disorders[12,26]. In this respect, electrical or chemical stimulation of dPAGpromotes intense fear in humans [37] and has been considered tobe an experimental model of panic attacks [26]. The aversive natureof IC stimulation has been confirmed by studies showing that ratssubmitted to avoidance paradigms quickly learn to switch-off suchstimulation [13,33]. In accordance with previous studies, our resultsshowed that stepwise increases in the intensity of electrical stim-
freezing and escape behaviors [12,16,19].It has been shown that microinjections of opioid antagonists
into the PAG precipitate somatic symptoms of withdrawal inmorphine-dependent animals [10,31]. These symptoms include

rain R
l[epsaiumnMtawbfo
iatssat2it
aOatwtatsosIhb
ittdddosoawr�r
aiesBar
J. De Ross et al. / Behavioural B
ocomotor hyperactivity, rearing, piloerection and escape attempts31], responses that resemble the fear-like behavior evoked bylectrical or chemical stimulation of the dPAG [6,12,22]. In theresent study, morphine withdrawal significantly reduced the aver-ive thresholds determined by electrical stimulation of the dPAGnd the IC. These findings are in line with a recent study show-ng that withdrawal from the same morphine treatment regimensed here produced enhanced anxiety levels in the elevated plus-aze and open field tests associated with the sensitization of the
eural substrates of fear in these midbrain tectum structures [3].oreover, there is immunohistochemical evidence showing that
he anxiety elicited by morphine withdrawal correlates with thectivation of the dPAG in rats displaying few somatic symptoms ofithdrawal [20]. In fact, the activation of the midbrain tectum may
e a common process involved in the elaboration of unconditionedear responses induced by withdrawal from several classes of drugsf abuse, such as ethanol, benzodiazepines and opiates [3,16,20,23].
Microinjections of DAMGO into the dPAG or IC significantlyncreased the aversive thresholds in non-dependent (untreated)nimals, suggesting that the activation of �-receptors inhibitshe neural substrates of aversion in these structures. Antiaver-ive effects were also produced by microinjections of BNI in bothtructures. It is known that although BNI acts as a non-selectiventagonist of opioid receptors during the first hour post-injection,his drug promotes a long-lasting selective blockade on �-receptors4 h after injection [9]. Accordingly, the antiaversive effects of BNI
n the present study were due to the blockade of such receptors inhe dPAG and the IC.
It has been demonstrated that the defensive behavior integratedt the midbrain tectum is modulated by opioid mechanisms [12].ur results are consistent with several studies showing that thectivation of �-receptors or the blockade of �-receptors [2,35,38] inhe dPAG promotes antiaversive effects in the elevated plus-maze,hereas the local blockade of �-receptors [38,43] or the activa-
ion of �-receptors [35,43] induces conditioned and unconditionedversive responses. Low doses of morphine administered directly inhe IC produce antiaversive effects, whereas high doses elicit aver-ive responses similar to those obtained after electrical stimulationf this structure [18]. The present results suggest that the antiaver-ive effects of low doses of morphine in the dPAG and also in theC may be due to the activation of �-receptors, for which morphineas higher affinity, whereas the aversive effects of high doses maye due to the activation of �-receptors.
The highest doses of DAMGO and BNI used in the present studynhibited the sensitization produced by morphine withdrawal onhe neural substrates of fear in the IC. These results are consis-ent with previous evidence showing that �- and �-receptors haveistinct roles in processes of positive reinforcement and opiateependence. In fact, the activation of �-receptors promotes con-itioned place preference, whereas the activation of �-receptorsr the blockade of �-receptors produces conditioned place aver-ion [4,5,36]. It has been proposed that the action of morphinen �-receptors in midbrain tectum structures could underlie theversive responses produced by naloxone-precipitated morphineithdrawal [11], which would be the result of the blockade of �-
eceptors by naloxone and the concomitant action of morphine on-receptors, for which naloxone has lower affinity [8]. The presentesults give support to this hypothesis.
Chronic treatment with opiates has been shown to promoteltered signal transduction of �-receptors in several brain areasncluding the dPAG [48,56]. This process contributes to the tol-
rance during treatment and to the occurrence of abstinenceymptoms if drug use is abruptly interrupted [30,55]. DAMGO andNI had similar antiaversive effects when injected into the dPAGnd IC of untreated rats. These findings confirm that �- and �-eceptors have opposite modulatory roles on the neural substratesesearch 204 (2009) 140–146 145
of aversion in the midbrain tectum [2,35,38,43]. In contrast, onlyBNI continued to promote antiaversive effects in the dPAG andIC of morphine-withdrawn rats. In these animals, DAMGO lostits antiaversive efficacy when injected into the dPAG although itcontinued to be effective when injected into the IC at a higherdose (1 nmol) than that necessary in untreated rats (0.6 nmol).These results suggest that an apparent tolerance for the anti-aversive effects of DAMGO developed in these midbrain tectumstructures, mainly at the dPAG level. Indeed, whereas the high-est dose of DAMGO significantly increased the aversive thresholdsdetermined by IC stimulation in morphine-withdrawn rats, thisagonist did not promote antiaversive effects in the dPAG of theseanimals.
In this study, animals ranged in age from 35 to 38 days at thebeginning of the chronic morphine treatment. In rats, this agerange corresponds to adolescence, which extends from postna-tal days 28 to 42 [50,51]. Since drug treatment continued for 10days, animals ranged from 47 to 50 days of age at the testingsession (after 48 h withdrawal). This period represents a “transi-tion zone” between true adolescence and adulthood and is usuallyreferred to as periadolescence, which extends up to postnatal day56 [50]. Clinical and experimental evidence suggests that periado-lescence represents a period of heightened vulnerability to theaddictive properties of drugs of abuse [21,49]. Periadolescent ani-mals are more sensitive to the positive reinforcing effects of ethanoland nicotine than adult animals [49]. In addition, novelty seek-ing and amphetamine sensitization are greater in periadolescents[1]. In terms of opioids, Spear et al. [52] reported that morphine-induced locomotion is more pronounced in periadolescents thanin adult rats. Moreover, there is evidence that periadolescent ani-mals may have an enhanced susceptibility to developing toleranceto repeated intermittent injections of morphine compared to adultrats [28]. In this context, it is possible that the effects of with-drawal from chronic morphine treatment observed in our studyare specific for periadolescent rats. On the other hand, it has beendemonstrated that certain parameters of responses to drugs ortheir withdrawal effects are similar in periadolescent and adultanimals [41,42,47]. Campbell et al. [17] reported no differencein the magnitude of place conditioning between periadolescentsand adults conditioned with either cocaine or morphine. Thus,further studies involving a comparison between the effects of mor-phine withdrawal in rats of different ages would be useful todetermine whether the results obtained in the present study arespecific for periadolescent animals or can be extended to adultrats.
In summary, the data obtained in the present study confirmedprevious evidence that morphine withdrawal promotes enhancedreactivity of the midbrain tectum to its electrical stimulation inperiadolescent rats [3]. This finding supports the hypothesis thatwithdrawal from chronic treatment with opiates leads to a fear statepossibly engendered by the activation of the dPAG and the IC. Ourresults also showed that, in non-dependent animals, the activationof �-receptors or the blockade of �-receptors inhibit the neuralsubstrates of aversion in the dPAG and the IC. On the other hand,in morphine-withdrawn rats, only the antagonism of �-receptorsmaintains its antiaversive efficacy in both structures. The neces-sity for a higher dose of DAMGO to promote antiaversive effects inthe IC and the absence of these effects in the dPAG of morphine-withdrawn rats suggest that chronic opiate treatment promotes anapparent tolerance to the antiaversive effects of the activation of �-receptors in the midbrain tectum. This process appears to be more
pronounced in the dPAG than in the IC. Thus, the enhanced reactiv-ity of the midbrain tectum in morphine-withdrawn periadolescentrats may be due, at least partially, to an impairment of the inhibitoryinfluence of mechanisms mediated by �-receptors on the neuralsubstrates of fear in this region.
1 rain R
A
0
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
1990;37:767–73.
46 J. De Ross et al. / Behavioural B
cknowledgement
This research was supported by grants from FAPESP (Proc. no.4/02859-0).
eferences
[1] Adriani W, Laviola G. Windows of vulnerability to psychopathology andtherapeutic strategy in the adolescent rodent model. Behav Pharmacol2004;15:341–52.
[2] Anseloni VC, Coimbra NC, Morato S, Brandao ML. A comparative study of theeffects of morphine in the dorsal periaqueductal gray and nucleus accumbensof rats submitted to the elevated plus-maze test. Exp Brain Res 1999;129:260–8.
[3] Ávila MA, Ruggiero RN, Cabral A, Brandão ML, Nobre MJ, Castilho VM. Involve-ment of the midbrain tectum in the unconditioned fear promoted by morphinewithdrawal. Eur J Pharmacol 2008;590:217–23.
[4] Bals-Kubik R, Ableitner A, Herz A, Shippenberg TS. Neuroanatomical sites medi-ating the motivational effects of opioids as mapped by the conditioned placepreference paradigm in rats. J Pharmacol Exp Ther 1993;264:489–95.
[5] Bals-Kubik R, Herz A, Shippenberg TS. Evidence that the aversive effects ofopioid antagonists and kappa-agonists are centrally mediated. Psychopharma-cology (Berl) 1989;98:203–6.
[6] Bandler R, Shipley MT. Columnar organization in the midbrain periaqueductalgray: modules for emotional expression? Trends Neurosci 1994;17:379–89.
[7] Bellchambers CE, Chieng B, Keay KA, Christie MJ. Swim-stress but not opioidwithdrawal increases expression of c-fos immunoreactivity in rat periaque-ductal gray neurons which project to the rostral ventromedial medulla.Neuroscience 1998;83:517–24.
[8] Bigelow GE, Preston KL, Opioids. In: Bloom FE, Kupfer DJ, editors. Psychophar-macology: the fourth generation of the progress. New York: Raven; 1995. p.1731–44.
[9] Birch PJ, Hayes AG, Sheehan MJ, Tyers MB. Norbinaltorphimine: antagonistprofile at kappa opioid receptors. Eur J Pharmacol 1987;144:405–8.
10] Bozarth MA, Wise RA. Anatomically distinct opiate receptor fields mediatereward and physical dependence. Science 1984;224:516–7.
11] Brandão ML. Involvement of opioid mechanisms in the dorsal periaqueductalgray in drug abuse. Rev Neurosci 1993;4:397–405.
12] Brandão ML, Anseloni VZ, Pandossio JE, De Araujo JE, Castilho VM. Neurochem-ical mechanisms of the defensive behavior in the dorsal midbrain. NeurosciBiobehav Rev 1999;23:863–75.
13] Brandão ML, de Aguiar JC, Graeff FG. GABA mediation of the anti-aversive actionof minor tranquilizers. Pharmacol Biochem Behav 1982;16:397–402.
14] Brandão ML, Tomaz C, Borges PC, Coimbra NC, Bagri A. Defense reaction inducedby microinjections of bicuculline into the inferior colliculus. Physiol Behav1988;44:361–5.
15] Busto UE, Sellers EM. Anxiolytics and sedative/hypnotics dependence. Br JAddict 1991;86:1647–52.
16] Cabral A, Isoardi N, Salum C, Macedo CE, Nobre MJ, Molina VA, et al. Fear stateinduced by ethanol withdrawal may be due to the sensitization of the neuralsubstrates of aversion in the dPAG. Exp Neurol 2006;200:200–8.
17] Campbell JO, Wood RD, Spear LP. Cocaine and morphine-induced place condi-tioning in adolescent and adult rats. Physiol Behav 2000;68:487–93.
18] Cardoso SH, Melo L, Coimbra NC, Brandão ML. Opposite effects of low and highdoses of morphine on neural substrates of aversion in the inferior colliculus.Behav Pharmacol 1992;3:489–95.
19] Castilho VM, Avanzi V, Brandão ML. Antinociception elicited by aversive stim-ulation of the inferior colliculus. Pharmacol Biochem Behav 1999;62:425–31.
20] Castilho VM, Borelli KG, Brandão ML, Nobre MJ. Anxiety-like symptoms inducedby morphine withdrawal may be due to the sensitization of the dorsal periaque-ductal grey. Physiol Behav 2008;94:552–62.
21] Chambers RA, Taylor JR, Potenza MN. Developmental neurocircuitry of motiva-tion in adolescence: a critical period of addiction vulnerability. Am J Psychiatry2003;160:1041–52.
22] Depaulis A, Bandler R, Vergnes M. Characterization of pretentorial periaque-ductal gray matter neurons mediating intraspecific defensive behaviors in therat by microinjections of kainic acid. Brain Res 1989;486:121–32.
23] Fontanesi LB, Ferreira R, Cabral A, Castilho VM, Brandão ML, Nobre MJ. Brain-stem areas activated by diazepam withdrawal as measured by Fos-proteinimmunoreactivity in rats. Brain Res 2007;1166:35–46.
24] Frenois F, Cador M, Caille S, Stinus L, Le Moine C. Neural correlates of themotivational and somatic components of naloxone-precipitated morphinewithdrawal. Eur J Neurosci 2002;16:1377–89.
25] Gawin FH. Cocaine addiction: psychology and neurophysiology. Science
1991;251:1580–6.26] Graeff FG. Serotonin, the periaqueductal gray and panic. Neurosci Biobehav Rev2004;28:239–59.
27] Hacker J, Pedersen NP, Chieng BC, Keay KA, Christie MJ. Enhanced Fos expressionin glutamic acid decarboxylase immunoreactive neurons of the mouse peri-aqueductal grey during opioid withdrawal. Neuroscience 2006;137:1389–96.
[
[
esearch 204 (2009) 140–146
28] Ingram SL, Fossum EN, Morgan MM. Behavioral and electrophysiological evi-dence for opioid tolerance in adolescent rats. Neuropsychopharmacology2007;32:600–6.
29] Jaffe JH. Drug addiction and abuse. In: Gilman AG, Rall TW, Nies AS, Taylor P,editors. Goodman and Gilman’s the pharmacological basis of therapeutics. NewYork: Pergamon Press; 1990. p. 522–73.
30] Koob GF, Le Moal M. Drug abuse: hedonic homeostatic dysregulation. Science1997;278:52–8.
31] Maldonado R, Stinus L, Gold LH, Koob GF. Role of different brain structures inthe expression of the physical morphine withdrawal syndrome. J PharmacolExp Ther 1992;261:669–77.
32] Markou A, Kosten TR, Koob GF. Neurobiological similarities in depression anddrug dependence: a self-medication hypothesis. Neuropsychopharmacology1998;18:135–74.
33] Melo LL, Cardoso SH, Brandão ML. Antiaversive action of benzodiazepines onescape behavior induced by electrical stimulation of the inferior colliculus.Physiol Behav 1992;51:557–62.
34] Motta V, Brandão ML. Aversive and antiaversive effects of morphine in thedorsal periaqueductal gray of rats submitted to the elevated plus-maze test.Pharmacol Biochem Behav 1993;44:119–25.
35] Motta V, Penha K, Brandão ML. Effects of microinjections of mu and kappareceptor agonists into the dorsal periaqueductal gray of rats submitted to theplus maze test. Psychopharmacology (Berl) 1995;120:470–4.
36] Narita M, Funada M, Suzuki T. Regulations of opioid dependence by opioidreceptor types. Pharmacol Ther 2001;89:1–15.
37] Nashold Jr BS, Wilson WP, Slaughter DG. Sensations evoked by stimulation inthe midbrain of man. J Neurosurg 1969;30:14–24.
38] Nobre MJ, Ribeiro dos SN, Aguiar MS, Brandão ML. Blockade of mu- and activa-tion of kappa-opioid receptors in the dorsal periaqueductal gray matter producedefensive behavior in rats tested in the elevated plus-maze. Eur J Pharmacol2000;404:145–51.
39] Osaki MY, Castellan-Baldan L, Calvo F, Carvalho AD. Felippotti TT, de OR et al.Neuroanatomical and neuropharmacological study of opioid pathways in themesencephalic tectum: effect of mu(1)- and kappa-opioid receptor blockadeon escape behavior induced by electrical stimulation of the inferior colliculus.Brain Res 2003;992:179–92.
40] Paxinos G, Watson C. The rat brain in stereotaxic coordinates. San Diego: Else-vier; 2005.
41] Philpot RM, Kirstein CL. The effects of repeated alcohol exposure on theneurochemistry of the periadolescent nucleus accumbens septi. Neuroreport1998;9:1359–63.
42] Philpot RM, Kirstein CL. Repeated cocaine exposure: effects on catecholaminesin the nucleus accumbens septi of periadolescent animals. Pharmacol BiochemBehav 1999;62:465–72.
43] Sante AB, Nobre MJ, Brandão ML. Place aversion induced by blockade of mu oractivation of kappa opioid receptors in the dorsal periaqueductal gray matter.Behav Pharmacol 2000;11:583–9.
44] Schulteis G, Koob GF. Reinforcement processes in opiate addiction: a homeo-static model. Neurochem Res 1996;21:1437–54.
45] Schulteis G, Markou A, Gold LH, Stinus L, Koob GF. Relative sensitivity to nalox-one of multiple indices of opiate withdrawal: a quantitative dose–responseanalysis. J Pharmacol Exp Ther 1994;271:1391–8.
46] Schulteis G, Yackey M, Risbrough V, Koob GF. Anxiogenic-like effects of sponta-neous and naloxone-precipitated opiate withdrawal in the elevated plus-maze.Pharmacol Biochem Behav 1998;60:727–31.
47] Shram MJ, Siu EC, Li Z, Tyndale RF, Le AD. Interactions between age andthe aversive effects of nicotine withdrawal under mecamylamine-precipitatedand spontaneous conditions in male Wistar rats. Psychopharmacology (Berl)2008;198:181–90.
48] Sim-Selley LJ, Selley DE, Vogt LJ, Childers SR, Martin TJ. Chronic heroin self-administration desensitizes mu opioid receptor-activated G-proteins in specificregions of rat brain. J Neurosci 2000;20:4555–62.
49] Smith RF. Animal models of periadolescent substance abuse. Neurotoxicol Ter-atol 2003;25:291–301.
50] Spear LP. The adolescent brain and age-related behavioral manifestations. Neu-rosci Biobehav Rev 2000;24:417–63.
51] Spear LP, Brake SC. Periadolescence: age-dependent behavior and psychophar-macological responsivity in rats. Dev Psychobiol 1983;16:83–109.
52] Spear LP, Horowitz GP, Lipovsky J. Altered behavioral responsivity to morphineduring the periadolescent period in rats. Behav. Brain Res 1982;4:279–88.
53] Stinus L, Caille S, Koob GF. Opiate withdrawal-induced place aversion lasts forup to 16 weeks. Psychopharmacology (Berl) 2000;149:115–20.
54] Stinus L, Le Moal M, Koob GF. Nucleus accumbens and amygdala are possiblesubstrates for the aversive stimulus effects of opiate withdrawal. Neuroscience
55] Van Ree JM, Gerrits MA, Vanderschuren LJ. Opioids, reward and addic-tion: an encounter of biology, psychology, and medicine. Pharmacol Rev1999;51:341–96.
56] Waldhoer M, Bartlett SE, Whistler JL. Opioid receptors. Annu Rev Biochem2004;73:953–90.




















