
Tobacco, Nicotine and Cotinine: For Memory, Neurological and Psychiatric Disorders

Chronic Exposure to Nicotine Is Associated withReduced Reward-Related Activity in the Striatum butnot the MidbrainEmma Jane Rose, Thomas J. Ross, Betty Jo Salmeron, Mary Lee, Diaa M. Shakleya, Marilyn Huestis, andElliot A. Stein
Background: The reinforcing effects of nicotine are mediated by brain regions that also support temporal difference error (TDE) processing;yet, the impact of nicotine on TDE is undetermined.
Methods: Dependent smokers (n � 21) and matched control subjects (n � 21) were trained to associate a juice reward with a visual cue ina classical conditioning paradigm. Subjects subsequently underwent functional magnetic resonance imaging sessions in which they wereexposed to trials where they either received juice as temporally predicted or where the juice was withheld (negative TDE) and later receivedunexpectedly (positive TDE). Subjects were scanned in two sessions that were identical, except that smokers had a transdermal nicotine (21mg) or placebo patch placed before scanning. Analysis focused on regions along the trajectory of mesocorticolimbic and nigrostriataldopaminergic pathways.
Results: There was a reduction in TDE-related function in smokers in the striatum, which did not differ as a function of patch manipulationbut was predicted by the duration (years) of smoking. Activation in midbrain regions was not impacted by group or drug condition.
Conclusions: These data suggest a differential effect of smoking status on the neural substrates of reward in distinct dopaminergicpathway regions, which may be partially attributable to chronic nicotine exposure. The failure of transdermal nicotine to alter reward-relatedfunctional processes, either within smokers or between smokers and control subjects, implies that acute nicotine patch administration isinsufficient to modify reward processing, which has been linked to abstinence-induced anhedonia in smokers and may play a critical role in
smoking relapse.purdwetiav
eb[hwbmriae
eotmncdt
Key Words: fMRI, nicotine, reward, smoking, striatum, temporaldifference error
H uman neuroimaging studies of reward processing suggestnetwork computation mediated through mesocorticolim-bic (MCL) dopamine (DA) pathways, i.e., those cells of the
ventral tegmental area (VTA) that project to the nucleus accumbens(NAcc) and are involved in nicotine reinforcement (1– 6). Orbito-frontal cortex and medial prefrontal cortex (mPFC) mediate positivehedonic feelings associated with reward receipt; orbitofrontal cor-tex activation also codes for reward value and is important in learn-ing from unexpected outcomes (7–10). The ventral striatum is acti-vated at earlier stages of reward processing, concomitant with theattribution of incentive salience to reward-predictive stimuli(11,12), when learning the relationship between an unconditionedstimulus and an impending reward and when temporal errors occurin the predicted receipt of a reward (13–17). Contingency-depen-dent changes in striatal activity are mediated by the activation ofmidbrain dopaminergic neurons in the VTA and substantia nigra(SN) pars compacta and their projections primarily to the ventraland dorsal striatum (18,19), suggesting that signals of this type arisefrom activity in both MCL and nigrostriatal (NS) DA pathways (i.e.,SN cell bodies with terminals predominantly in the dorsal striatum),a system involved in habit learning and postulated to play a role inreward and addiction (see [20] for review).
From the Neuroimaging Research Branch (EJR, TJR, BJS, ML, EAS) and Chem-istry and Drug Metabolism Branch (DMS, MH), National Institute on DrugAbuse, National Institutes of Health, Baltimore, Maryland.
Address correspondence to Emma Jane Rose, Ph.D., Trinity College Dublin,Department of Psychiatry, Neuropsychiatric Genetics Group, Dublin, Ire-land; E-mail: [email protected].
mReceived May 11, 2011; revised Sep 5, 2011; accepted Sep 6, 2011.
0006-3223/$36.00doi:10.1016/j.biopsych.2011.09.013
Human, nonhuman primate, and rodent studies demonstratehasic increases in DA signaling in the basal ganglia followingnexpected natural rewards, a temporal shift in DA response from
eward receipt to stimuli predicting reward (21–23), and transientecreases in signaling following the omission of anticipated re-ards (13,16,17,24). This suggests a neural basis for temporal differ-
nce (TD) learning and error prediction in the structures comprisinghe basal ganglia. Addictive drugs also produce transient increasesn DA signaling (25,26) that become conditioned in humans (27)nd may lead to positive TD error (TDE) signals that increase drugalue and reinforce drug-seeking behavior (28).
A range of nicotine-related situational states (e.g., withdrawal,xpectation, cue-induced reactivity, craving suppression) haveeen mapped onto regions supporting learning and reward (e.g.,
29 –35]). Moreover, nicotine exerts its pharmacological effects viaigh-affinity nicotinic acetylcholine receptors (nAChRs), which areidely distributed throughout the brain (36,37), including the cellodies and axon terminals of MCL and NS DA neurons, suggesting aechanism for nicotine’s reinforcing properties (38,39). Given the
ole of these pathways in a range of reward processes, nicotine’snfluence on reward processing may extend beyond motivationalspects of smoking behavior and impact on TD processing thatxtends to and modifies other (nondrug) rewards.
Using a simple classical conditioning paradigm (13), we consid-red 1) whether being a dependent smoker alters the neurobiologyf TDE processing, and 2) the impact of acute nicotine administra-
ion on the functional profile associated with TDE. Since withdrawalay have state-dependent effects on reward processing that do
ot simply reflect the impact of chronic nicotine, these issues wereonsidered in dependent smokers in the absence of a frank with-rawal state. We hypothesized that acute administration of nico-
ine and chronic exposure to nicotine in dependent smokers would
odify TDE processing of a natural reward.BIOL PSYCHIATRY 2012;71:206–213© 2012 Society of Biological Psychiatry
(F
P
plcotpnstss
T
pjpspfledtmpap
Fibtgb
E.J. Rose et al. BIOL PSYCHIATRY 2012;71:206–213 207
Methods and Materials
ParticipantsSixty-four right-handed (40) individuals were recruited from the
general population. Thirteen subjects were excluded due to dataquality issues (primarily head motion) and one subject failed tocomplete scanning. For the purposes of matching between groups,a further five participants were excluded before analysis. The anal-ysis cohort included adult (�18 years) smokers (n � 21) and non-smoking control subjects (n � 21), matched for age, gender, self-reported race, IQ (41), and years of education (Table 1). Smokerssmoked at least 15 cigarettes per day for a minimum of 1 yearbefore participation. Control subjects had no history of smoking.
Exclusion criteria included significant neurological or medicalhistory, any psychiatric history, claustrophobia, pregnancy, and anyother contraindication for magnetic resonance imaging (MRI). Withthe exception of nicotine dependence in smokers, substance abuseor dependence was exclusionary.
ProcedureThis study was approved by the National Institute on Drug
Abuse-Intramural Research Program Institutional Review Boardand written informed consent was obtained from all subjects. Par-ticipants made three study visits: task and procedural training in amock scanner and two MRI sessions. The sessions were identical forall participants, except that smokers had a 21-mg nicotine or pla-cebo patch (Nicoderm, GlaxoSmithKline Inc., Research TrianglePark, North Carolina) applied 2 hours before MRI. Session order forsmokers was single-blind, randomly determined, and counter-bal-anced between subjects (n � 10 nicotine first). Control subjectswere scanned twice without a patch manipulation. Participantswere not permitted to consume alcohol or over-the-counter medi-cations for 24 hours before each session and were limited to one-half cup of caffeinated beverage before each scan. Before MRI ses-sions, participants were tested for recent drug use (urine drug test;TRIAGE, San Diego, California), alcohol use (Alco-Sensor IV, Intoxi-meters Inc., St. Louis, Missouri), pregnancy, and expired carbonmonoxide (Vitalograph Breath CO monitor; Lenexa, Kansas). For thepurposes of characterization, participants completed measures ofattention, switching (Trail Making Test Parts A and B) (42), memory
Table 1. Participant Demographic Information
Smokers(n � 21)
Control Subjects(n � 21)
Age, Years [Mean (SD)] 32.90 (9.77) 30.29 (8.74)Gender (Male: Female) 8:13 12:9IQ
WASI [mean (SD)] 106.95 (11.06) 109.16 (13.93)Education [years; mean (SD)] 12.90 (2.71) 13.95 (2.54)
Ethnicity (AA:C:As) 7:14:0 6:14:1Cigarettes/Day [Mean (SD)] 22.55 (5.60) NAAge at First Use [Years; Min–
Max (Mean)] 9–31 (15.62) NAYears of Use Preceding Study
[Min–Max (Mean)] 2.5–38 (16.36) NAFTND [Min–Max (Mean)] 3–9 (5.76) NA
Note: There was no statistically significant difference between groups inany relevant measure.
AA, African American; As, Asian/Asian American; C, Caucasian; FTND,Fagerström Test for Nicotine Dependence; NA, not applicable; WASI,Wechsler Abbreviated Scale of Intelligence.
(43), and the Cloninger Temperament and Character Inventory (TCI)dr
44). Smokers completed a detailed smoking history, including theagerström Test for Nicotine Dependence (FTND) (45).
atch AdministrationPatches were affixed onto the upper back 30 minutes after the
articipant’s last cigarette and 2 hours before scanning. The time ofast cigarette was confirmed by research staff. A 2-hour delay washosen to allow for maximal nicotine plasma levels within the nic-tine condition and to minimize withdrawal in the placebo condi-
ion (46,47). Both patch types were manufactured by the sameharmaceutical company and were identical in appearance andonnicotine content. Participants were not debriefed regardingession order until they had completed the study. Withdrawal in-ensity, mood, and nicotine craving were queried before and aftercanning using the Parrott mood questionnaire (48) and a 12-itemhort form of the Tobacco Craving Questionnaire (TCQ) (49,50).
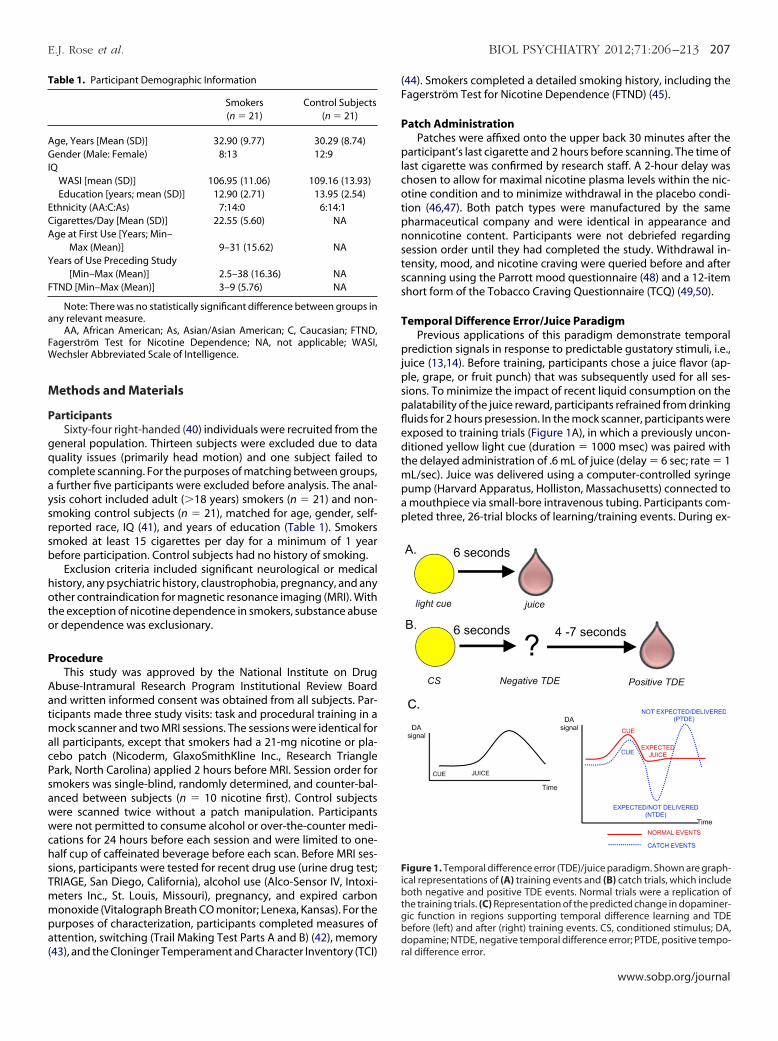
emporal Difference Error/Juice ParadigmPrevious applications of this paradigm demonstrate temporal
rediction signals in response to predictable gustatory stimuli, i.e.,uice (13,14). Before training, participants chose a juice flavor (ap-le, grape, or fruit punch) that was subsequently used for all ses-ions. To minimize the impact of recent liquid consumption on thealatability of the juice reward, participants refrained from drinkinguids for 2 hours presession. In the mock scanner, participants werexposed to training trials (Figure 1A), in which a previously uncon-itioned yellow light cue (duration � 1000 msec) was paired with
he delayed administration of .6 mL of juice (delay � 6 sec; rate � 1L/sec). Juice was delivered using a computer-controlled syringe
ump (Harvard Apparatus, Holliston, Massachusetts) connected tomouthpiece via small-bore intravenous tubing. Participants com-leted three, 26-trial blocks of learning/training events. During ex-
igure 1. Temporal difference error (TDE)/juice paradigm. Shown are graph-cal representations of (A) training events and (B) catch trials, which includeoth negative and positive TDE events. Normal trials were a replication of
he training trials. (C) Representation of the predicted change in dopaminer-ic function in regions supporting temporal difference learning and TDEefore (left) and after (right) training events. CS, conditioned stimulus; DA,
opamine; NTDE, negative temporal difference error; PTDE, positive tempo-al difference error.
www.sobp.org/journal

Fdt(w
pv
T
otsahsiebio
F
(aptfifw(
cwwaG
1oaaSAlvsasc
ndipniecn
R
P
rw[est.ct
lfaCi[tt
ss
N
n[t
208 BIOL PSYCHIATRY 2012;71:206–213 E.J. Rose et al.
perimental runs, participants were exposed to training/normal tri-als (n � 58) interspersed with randomly selected catch trials (n � 20;
igure 1B), wherein the juice reward was unpredictably and pseu-orandomly delayed by 4 to 7 seconds. It was anticipated that failing
o deliver the juice reward as predicted would generate a negative TDENTDE) signal, whereas the temporally unanticipated receipt of juice
ould engender a positive TDE (PTDE) signal (Figure 1C).To determine juice palatability, at the end of each task block,
articipants were asked to rate how much they liked the juice on aisual analog scale (range � 400).
iming ParadigmSince brain regions that might be compromised in smokers
verlap with those supporting second range timing function (51),he ability to accurately predict the time between the conditionedtimulus (CS) and juice reward was of concern. Therefore, subjectslso completed a test of timing function post-MRI. Participantseard four .5-Hz tones per trial (n � 15). The first two tones wereeparated by a 6-second interval (i.e., equal to the light cue/juicenterstimulus interval), as were the second and third tones. How-ver, the interval between tones three and four varied randomlyetween 4.5 and 7.5 seconds. Participants indicated whether the
nterval between the last two tones was shorter than, longer than,r equal to the first two intervals.
unctional ImagingWhole-brain echo planar images were acquired on a 3T Siemens
Erlangen, Germany) Allegra scanner. Thirty-nine 4-mm slices werecquired in an oblique axial plane (30° to anterior commissure-oster commissure) with the following imaging parameters: repe-
ition time [TR] � 2000 milliseconds, echo time � 27 milliseconds,eld of view � 220 � 220 mm at 64 � 64, and flip angle � 78°. Total
unctional scanning time was approximately 27 minutes. T1-eighted magnetization prepared rapid acquisition gradient echo
MPRAGE) structural imaging series with a voxel size of 1 mm3 werealso acquired.
Blood Draw and AnalysisVenous blood samples (5 mL) were collected from smokers im-
mediately following completion of each MRI session, centrifugedwithin 2 hours, and plasma stored at �20°C until analysis. A com-prehensively validated liquid chromatography tandem mass spec-trometry analysis was employed for simultaneous quantification ofnicotine, cotinine, trans-3’-hydroxycotinine, and norcotinine inplasma (52).
Data AnalysisFunctional imaging data were analyzed using AFNI (53). To cor-
rect for head motion, three-dimensional echo-planar imaging datafor each subject were registered to a base volume. The data wereinspected for motion using the censor.py application (http://brain-imaging.waisman.wisc.edu/�perlman/code/censor.py). Strict cen-soring criteria (i.e., translation �.3 mm or rotation �3° betweenconsecutive TRs) were used to remove unwanted TRs before decon-volution. Individuals with a censor rate �25% were excluded. Datatime series were analyzed using voxel-wise, multiple regression inwhich regressors were expressed as a delta function time-locked toevent onset and convolved with a hemodynamic response functionand its temporal derivative. There were four regressors of interest:light cue (CS), normal events (unconditioned stimulus [UCS]; juiceexpected/juice delivered), NTDE (juice expected/juice not deliv-ered), and PTDE (juice not expected/juice delivered). In addition, sixmotion parameters were included as regressors of no interest. For
each participant and session, a voxel-wise average amplitude dwww.sobp.org/journal
hange (�) equal to the percentage signal change from baselineas calculated for each event type. The resultant activation mapsere registered to a higher resolution (1 �L) standard space (54)
nd spatially blurred using a 4.2 mm full-width at half maximumaussian isotropic kernel.
A priori region of interest (ROI) analyses (Table S5 in Supplement) focused on the impact of participant group and drug conditionn regressor-related activity in regions along the trajectory of MCLnd NS pathways. Bilateral ROIs in the SN, striatum (NAcc, caudate,nd putamen), and mPFC (Brodmann area [BA]10 and BA32; Figure1 in Supplement 1) were defined using a Talairach template inFNI. The VTA ROI was defined as a 5 mm sphere at its anatomical
ocus (Talairach coordinates: 0 �16 �7). The mean value acrossoxels within each ROI was calculated for each participant/regres-or and subsequently used as the dependent variable in statisticalnalyses. To address variability in nicotine metabolism, compari-ons within smokers included nicotine plasma concentration as aovariate.
Behavioral data were analyzed in SPSS (SPSS Inc., Chicago, Illi-ois). Analysis of Parrott scores considered the effects of group,rug condition, session, and time (prescanning vs. postscanning;
.e., immediately preceding the first scan vs. directly following com-letion of the final scan, within session). Tobacco Craving Question-aire analysis examined four craving indices: emotionality (smok-
ng in anticipation of relief from withdrawal/negative mood);xpectancy (anticipation of positive outcomes from smoking);ompulsivity (an inability to control tobacco use); and purposeful-ess (intention/planning to smoke).
esults
arrott and TCQControl Subjects Versus Smokers. Participants were more
elaxed in session 2 versus session 1 [F (1,39) � 6.48, p � .01] andere more distracted [F (1,39) � 16.61, p .001] and hungrier
F (1,39) � 22.25, p .001] by the end of each session. While smok-rs were more dissatisfied by the end of the scanning session [ses-ion 1: t (20) � �2.88, p � .005; session 2: t (20) � �1.83, p � .04],his difference was absent in control subjects [F (1,39) � 4.39, p 05]. Smokers were also more tired at the start of the second sessionompared with control subjects [t (40) � �2.08, p � .02] but not byhe end of the session (Table S1 in Supplement 1).
Smokers: Nicotine Versus Placebo. Smokers were more re-axed [F (1,20) � 8.45, p � .009], content [F (1,20) � 5.81, p � .03],ocused [F (1,20) � 4.11, p � .05], satisfied [F (1,20) � 4.59, p � .04],nd less hungry [F (1,20) � 5.54, p � .03] in the nicotine condition.onversely, following placebo, smokers experienced higher smok-
ng expectancy [F (1,20) � 4.76, p � .04] and purposefulnessF (1,20) � 15.19, p � .001] scores on the TCQ. These data suggesthat the nicotine patch prevented a mild withdrawal state seen inhe placebo condition.
There was no interaction between time (prescanning vs. post-canning) and drug condition on any measure of mood or craving inmokers (Table S2 in Supplement 1).
icotine and Its MetabolitesBlood plasma concentrations of nicotine, cotinine, and norcoti-
ine were higher at the end of the nicotine session versus placebotNICOTINE(28) � 11.09, p .001; tCOTININE (28) � 4.29, p .001; and
(28) � 3.49, p � .002; Table S3 in Supplement 1]. Hy-
NORCOTININEroxycotinine concentrations did not differ between conditions.
psgrl
J
cctb
F
sptsat
atee
aNePdvta
S
M
m
nv
E.J. Rose et al. BIOL PSYCHIATRY 2012;71:206–213 209
TCIThe TCI provides indices of novelty seeking, harm avoidance,
reward dependence, persistence, self-directedness, cooperative-ness, and self-transcendence, which purportedly reflect underlyingneurobiology (e.g., dopaminergic, serotonergic, or noradrenergicactivity) (55). Control subjects and smokers were matched on allthese aspects of temperament and character (p � .05).
Cognitive MeasuresPerformance on all prescanning cognitive measures was equiv-
alent between groups (p � .05). Control subjects and smokers alsoerformed equally well on the timing task, and accuracy was con-istent across conditions for smokers (p � .05). Thus, irrespective ofroup or drug condition, participants were equally able to accu-
ately determine the duration of timing intervals approximating theight/juice interstimulus interval (Table S4 in Supplement 1).
uice PalatabilityJuice palatability did not vary as a function of group or drug
ondition (p � .05; mean [SD]: nicotine � 165.90 [177.51]; pla-ebo � 187.24 [156.82]; control subjects � 238.79 [106.65]) andhere was no session, group, or drug effect on the juice rating
Table 2. Summary Statistics from the ROI Analysis of Reward-Related Activ
Group A Priori
triatumCaudate F(1,120) � 4.69c
(smokers control subjects)F(3,120) � 11.45a
Putamen F(1,120) � 6.94c
(smokers control subjects)F(3,120) � 34.14a
NAcc F(1,120) � 5.76c
(smokers control subjects)F(3,120) � 5.18c
idbrainVTA nil F(3,120) � 10.29a
SN nil F(3,120) � 28.78a
PFCBA10 nil F(3,120) � 2.87c
BA32 F(1,120) � 5.38c
(smokers control subjects)F(3,120) � 7.72a
BA, Brodmann area; CS, conditioned stimulus; MCL, mesocorticolimbic; megative temporal difference error; PTDE, positive temporal difference erroentral tegmental area.
ap .001.bp .005.cp .05.
etween the start and end of sessions (p � .05). s
unctional ImagingEffect of Acute Nicotine. Acute nicotine administration in
mokers did not alter activity in any a priori ROI compared with thelacebo condition. There was also no session effect in control par-
icipants. Therefore, our results focus on the average activity acrossessions in control subjects compared with that of smokers aver-ged across patch conditions. To enhance readability, the results ofhese between-group analyses are presented in Table 2.
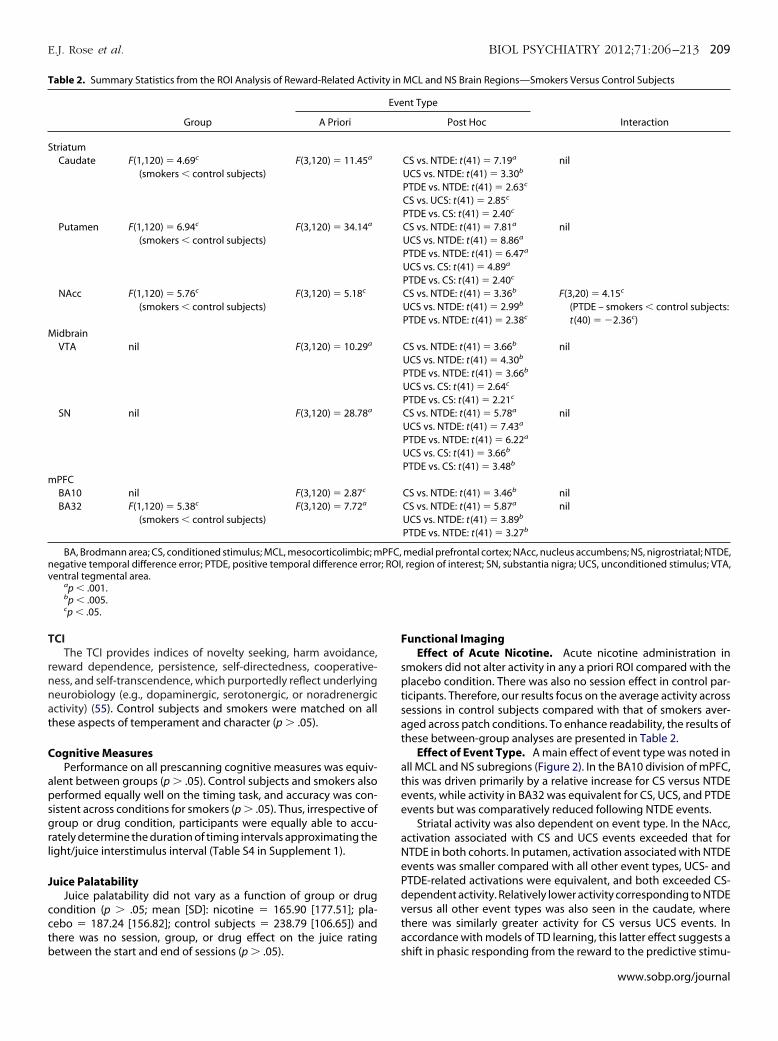
Effect of Event Type. A main effect of event type was noted inll MCL and NS subregions (Figure 2). In the BA10 division of mPFC,his was driven primarily by a relative increase for CS versus NTDEvents, while activity in BA32 was equivalent for CS, UCS, and PTDEvents but was comparatively reduced following NTDE events.
Striatal activity was also dependent on event type. In the NAcc,ctivation associated with CS and UCS events exceeded that forTDE in both cohorts. In putamen, activation associated with NTDEvents was smaller compared with all other event types, UCS- andTDE-related activations were equivalent, and both exceeded CS-ependent activity. Relatively lower activity corresponding to NTDEersus all other event types was also seen in the caudate, wherehere was similarly greater activity for CS versus UCS events. Inccordance with models of TD learning, this latter effect suggests a
MCL and NS Brain Regions—Smokers Versus Control Subjects
nt Type
InteractionPost Hoc
CS vs. NTDE: t(41) � 7.19a
UCS vs. NTDE: t(41) � 3.30b
PTDE vs. NTDE: t(41) � 2.63c
CS vs. UCS: t(41) � 2.85c
PTDE vs. CS: t(41) � 2.40c
nil
CS vs. NTDE: t(41) � 7.81a
UCS vs. NTDE: t(41) � 8.86a
PTDE vs. NTDE: t(41) � 6.47a
UCS vs. CS: t(41) � 4.89a
PTDE vs. CS: t(41) � 2.40c
nil
CS vs. NTDE: t(41) � 3.36b
UCS vs. NTDE: t(41) � 2.99b
PTDE vs. NTDE: t(41) � 2.38c
F(3,20) � 4.15c
(PTDE – smokers control subjects:t(40) � �2.36c)
CS vs. NTDE: t(41) � 3.66b
UCS vs. NTDE: t(41) � 4.30b
PTDE vs. NTDE: t(41) � 3.66b
UCS vs. CS: t(41) � 2.64c
PTDE vs. CS: t(41) � 2.21c
nil
CS vs. NTDE: t(41) � 5.78a
UCS vs. NTDE: t(41) � 7.43a
PTDE vs. NTDE: t(41) � 6.22a
UCS vs. CS: t(41) � 3.66b
PTDE vs. CS: t(41) � 3.48b
nil
CS vs. NTDE: t(41) � 3.46b nilCS vs. NTDE: t(41) � 5.87a
UCS vs. NTDE: t(41) � 3.89b
PTDE vs. NTDE: t(41) � 3.27b
nil
medial prefrontal cortex; NAcc, nucleus accumbens; NS, nigrostriatal; NTDE,region of interest; SN, substantia nigra; UCS, unconditioned stimulus; VTA,
ity in
Eve
PFC,r; ROI,
hift in phasic responding from the reward to the predictive stimu-
www.sobp.org/journal

cpc
Vve
si
dpblnBcc.aeerpa
D
ietTsbld
N
prtdptpinssee
trifpmcttc
( ntal cP itione
210 BIOL PSYCHIATRY 2012;71:206–213 E.J. Rose et al.
lus. However, due to experimental design limitations, we cannotconfirm this. CS-related activity in the caudate was also greater thanthat in the putamen [t (41) � 3.19, p � .003] and NAcc [t (41) � 3.95,p .001]. These data indicate a partial dissociation in reward pro-
essing in striatal subregions, with reward receipt being processedredominantly in putamen and TD learning effects primarily in theaudate.
The main effect of event type was identical in midbrain SN andTA, where it was manifest as greater activity for CS, UCS, and PTDEersus NTDE events and greater activity for UCS and PTDE versus CSvents.
Effect of Group. There was a main effect of group in all striatalubregions and BA32, resulting from comparatively reduced activ-ty in smokers (Figure 2).
Group � Event Type Interaction. Only the NAcc demon-strated a group � event type interaction, which was manifest as lessactivity associated with PTDE events in smokers and a relative re-duction in activity for PTDE versus NTDE events in control subjectsonly.
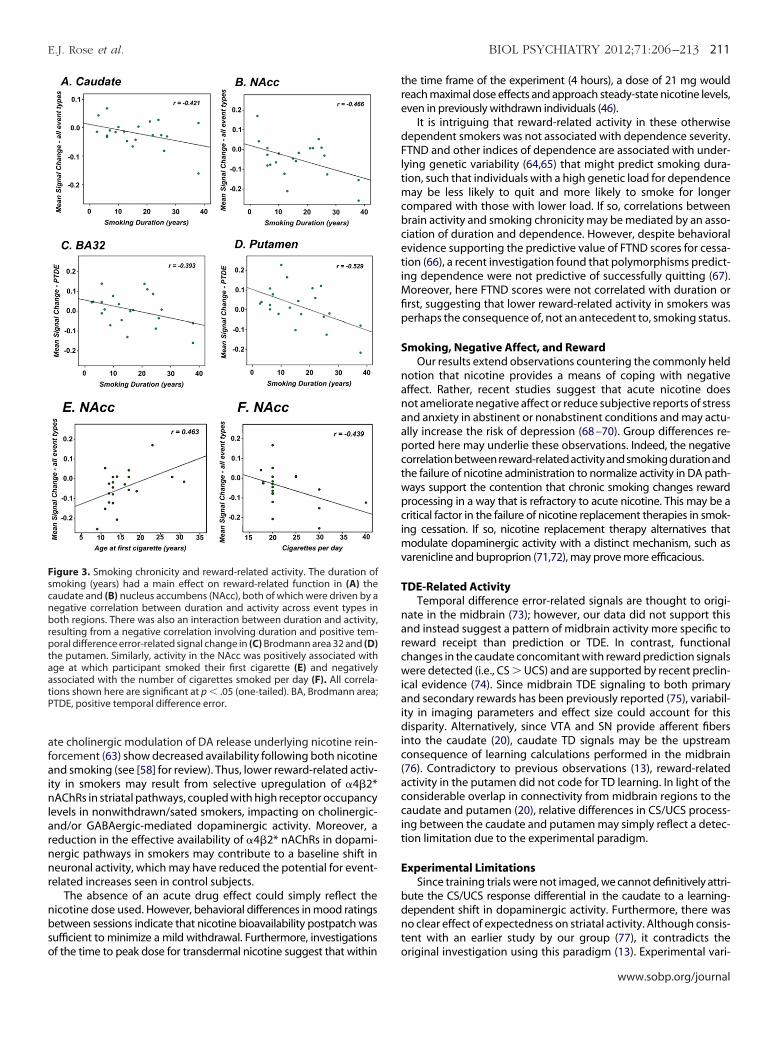
Smoking History. To delineate the influence of smoking char-acteristics, we considered the impact of 1) duration of smoking(years), 2) age at first cigarette (first), 3) number of cigarettes per day(CPD), and (4) FTND on activity associated with each event type inall ROIs. There was a main effect of duration in the NAcc [F (1,19) �5.28, p .05] and caudate [F (1,19) � 4.09, p � .05] and event type �
uration interaction effects in BA32 [F (1,19) � 11.02, p .01] andutamen [F (1,19) � 3.94, p .01]. Linear contrasts and post hocivariate correlations (p .05; one-tailed) confirmed that TDE-re-
ated activity in the caudate (Figure 3A) and NAcc (Figure 3B) wasegatively correlated with duration and that interaction effects inA32 (Figure 3C) and putamen (Figure 3D) were due to a negativeorrelation between duration and PTDE events. CPD was negativelyorrelated with event-related activity in NAcc [F (1,19) � 4.54, p
05; Figure 3E], whereas first was positively associated with NAccctivity [F (1,19) � 5.17, p .05; Figure 3F]. FTND did not mediatevent-related activity in any ROI and none of these factors influ-nced activity in BA10 or midbrain. These data suggest that inegions where activity was relatively reduced in smokers, functionalrocessing was most consistently influenced by smoking chronicity
Figure 2. The impact of event type (conditioned stimulus/unconditioned stimgroup (control subjects vs. smokers) on the mean signal change in a priori anigrostriatal dopamine pathways (i.e., medial prefrontal cortex [BA 10 and B(substantia nigra and ventral tegmental area). Note: error bars show � 1 stp .05). BA, Brodmann area; CS, conditioned stimulus; MPFC, medial prefroTDE, positive temporal difference error; SN, substantia nigra; UCS, uncond
nd not nicotine dependence. d
www.sobp.org/journal
iscussion
Using a classical conditioning paradigm, we observed outcomesndicative of trait effects of being a dependent smoker but not stateffects of acute nicotine administration in TDE/reward-related ac-ivity in regions along the trajectory of MCL and NS DA pathways.his effect was manifest as lower activity in smokers (vs. controlubjects) in striatal and mPFC/BA32 subregions but not the mid-rain. Moreover, smoking-related reductions in activity were corre-
ated with the duration of smoking (years) but not the severity ofependence (FTND).
icotine and TDE-Related ActivityPreclinical observations indicate that nicotine acts as both a
rimary reinforcer and enhances the incentive motivational andeinforcing effects of accompanying stimuli (56). However, nico-ine’s influence on reward processing in humans may rely less onirect rewarding effects than its ability to modulate the rewardingroperties of other stimuli, which may be integral to nicotine addic-
ion and underlie behavioral phenomena such as the increasedropensity to smoke while drinking (57). Yet, a direct pharmacolog-
cal effect of acute nicotine when using a natural (juice) reward wasot observed. Rather, differences in TDE-related activity were onlyeen as a function of group, i.e., smokers versus nonsmokers. Thisuggests that this stimulus-independent property of nicotine’s influ-nce on reward/reinforcement might be more related to chronic drugxposure-induced neuroplasticity rather than acute nicotine, per se.
The lack of an effect of acute nicotine administration implies arait-like influence of chronic nicotine exposure/smoking status oneward processing. That group differences in reward-related activ-ty were associated with smoking longevity raises the issue of howunctional changes in dopaminergic pathways, especially striatalathways, may be mediated by repeated nicotine exposure. Hu-an and animal models indicate upregulation of nAChRs following
hronic nicotine, an effect perhaps related to desensitization ofhese receptors (see [58,59] for review). Chronic nicotine enhances4* receptors in striatal pathways (60,61), and selective upregula-
ion of these subunits on gamma-aminobutyric acid (GABA)ergicells plus chronic stimulation by nicotine engenders decreased
/negative temporal difference error/positive temporal difference error) andmical regions of interest along the trajectory of the mesocorticolimbic and), striatum (i.e., nucleus accumbens, caudate, and putamen), and midbraind error; †significant main effect of group (p .05); *significant interactionortex; NAcc, nucleus accumbens; NTDE, negative temporal difference error;d stimulus; VTA, ventral tegmental area.
ulusnatoA 32]
andar
opaminergic function (62). Moreover, 4�2* receptors that medi-
tre
dFltmcbcetiMfip
S
nanaapctwpcimv
T
narcwiaidic(accit
E
bdnt
cnbrptaatP
E.J. Rose et al. BIOL PSYCHIATRY 2012;71:206–213 211
ate cholinergic modulation of DA release underlying nicotine rein-forcement (63) show decreased availability following both nicotineand smoking (see [58] for review). Thus, lower reward-related activ-ity in smokers may result from selective upregulation of 4�2*nAChRs in striatal pathways, coupled with high receptor occupancylevels in nonwithdrawn/sated smokers, impacting on cholinergic-and/or GABAergic-mediated dopaminergic activity. Moreover, areduction in the effective availability of 4�2* nAChRs in dopami-nergic pathways in smokers may contribute to a baseline shift inneuronal activity, which may have reduced the potential for event-related increases seen in control subjects.
The absence of an acute drug effect could simply reflect thenicotine dose used. However, behavioral differences in mood ratingsbetween sessions indicate that nicotine bioavailability postpatch wassufficient to minimize a mild withdrawal. Furthermore, investigations
Figure 3. Smoking chronicity and reward-related activity. The duration ofsmoking (years) had a main effect on reward-related function in (A) the
audate and (B) nucleus accumbens (NAcc), both of which were driven by aegative correlation between duration and activity across event types inoth regions. There was also an interaction between duration and activity,
esulting from a negative correlation involving duration and positive tem-oral difference error-related signal change in (C) Brodmann area 32 and (D)
he putamen. Similarly, activity in the NAcc was positively associated withge at which participant smoked their first cigarette (E) and negativelyssociated with the number of cigarettes smoked per day (F). All correla-ions shown here are significant at p .05 (one-tailed). BA, Brodmann area;TDE, positive temporal difference error.
of the time to peak dose for transdermal nicotine suggest that within o
he time frame of the experiment (4 hours), a dose of 21 mg wouldeach maximal dose effects and approach steady-state nicotine levels,ven in previously withdrawn individuals (46).
It is intriguing that reward-related activity in these otherwiseependent smokers was not associated with dependence severity.TND and other indices of dependence are associated with under-
ying genetic variability (64,65) that might predict smoking dura-ion, such that individuals with a high genetic load for dependence
ay be less likely to quit and more likely to smoke for longerompared with those with lower load. If so, correlations betweenrain activity and smoking chronicity may be mediated by an asso-iation of duration and dependence. However, despite behavioralvidence supporting the predictive value of FTND scores for cessa-ion (66), a recent investigation found that polymorphisms predict-ng dependence were not predictive of successfully quitting (67).
oreover, here FTND scores were not correlated with duration orrst, suggesting that lower reward-related activity in smokers waserhaps the consequence of, not an antecedent to, smoking status.
moking, Negative Affect, and RewardOur results extend observations countering the commonly held
otion that nicotine provides a means of coping with negativeffect. Rather, recent studies suggest that acute nicotine doesot ameliorate negative affect or reduce subjective reports of stressnd anxiety in abstinent or nonabstinent conditions and may actu-lly increase the risk of depression (68 –70). Group differences re-orted here may underlie these observations. Indeed, the negativeorrelation between reward-related activity and smoking duration andhe failure of nicotine administration to normalize activity in DA path-ays support the contention that chronic smoking changes rewardrocessing in a way that is refractory to acute nicotine. This may be aritical factor in the failure of nicotine replacement therapies in smok-
ng cessation. If so, nicotine replacement therapy alternatives thatodulate dopaminergic activity with a distinct mechanism, such as
arenicline and buproprion (71,72), may prove more efficacious.
DE-Related ActivityTemporal difference error-related signals are thought to origi-
ate in the midbrain (73); however, our data did not support thisnd instead suggest a pattern of midbrain activity more specific toeward receipt than prediction or TDE. In contrast, functionalhanges in the caudate concomitant with reward prediction signalsere detected (i.e., CS � UCS) and are supported by recent preclin-
cal evidence (74). Since midbrain TDE signaling to both primarynd secondary rewards has been previously reported (75), variabil-
ty in imaging parameters and effect size could account for thisisparity. Alternatively, since VTA and SN provide afferent fibers
nto the caudate (20), caudate TD signals may be the upstreamonsequence of learning calculations performed in the midbrain76). Contradictory to previous observations (13), reward-relatedctivity in the putamen did not code for TD learning. In light of theonsiderable overlap in connectivity from midbrain regions to theaudate and putamen (20), relative differences in CS/UCS process-
ng between the caudate and putamen may simply reflect a detec-ion limitation due to the experimental paradigm.
xperimental LimitationsSince training trials were not imaged, we cannot definitively attri-
ute the CS/UCS response differential in the caudate to a learning-ependent shift in dopaminergic activity. Furthermore, there waso clear effect of expectedness on striatal activity. Although consis-
ent with an earlier study by our group (77), it contradicts the
riginal investigation using this paradigm (13). Experimental vari-www.sobp.org/journal

1
1
1
1
1
1
1
1
1
1
2
2
2
2
2
2
2
2
2
2
3
212 BIOL PSYCHIATRY 2012;71:206–213 E.J. Rose et al.
ability, such as differences in the delay between CS and PTDE and inthe number/ratio of catch events, may account for this difference.Unique trial types involving the omission of juice reward (NTDE-only trials) or temporally uncoupling juice from the CS (PTDE-onlytrials) may better delineate expectedness.
Pharmacological manipulation studies using blood-oxygenlevel dependent require additional consideration with respect tononspecific signal transduction interactions. However, since smok-ing was associated with regionally specific/task-dependent effectsin the absence of global perturbations, between-group differencesare unlikely to reflect disease-related vascular alterations (78) butrather functional consequences of alterations in receptor and/orneurotransmitter function at specific locations, especially the stria-tum. Nonetheless, regionally nonspecific effects cannot be completelyprecluded and while smoking-related differences are attributed tochronic nicotine intake, cigarettes contain thousands of compounds(79), any number of which could contribute to the observed differ-ences. Yet, contaminants (e.g., carbon monoxide, tars) might also beexpected to have a more global action, arguing for particular nAChR-induced neuroplasticity rather than global neurotoxicity.
Importantly, our experimental paradigm does not allow for thedisambiguation of the effects of acute nicotine and withdrawalalleviation. Considering reward function at different time pointssince last cigarette, using more substantially withdrawn partici-pants, or administering acute nicotine to nicotine-naïve individualswould help clarify this issue.
Finally, despite the comparatively robust analysis of covarianceresults, post hoc correlations used to confirm the directionality ofthe relationship between smoking characteristics and activity werepotentially underpowered due to the relatively small number ofsubjects and should be cautiously interpreted.
In sum, reduced TDE-related/reward-related processing wasseen in striatum and mPFC but not in the midbrain of smokers.These effects were related to chronic nicotine exposure and werenot amenable to acute, transdermal nicotine delivery. The impact ofchronic nicotine exposure on TD-related activity may have implica-tions for the hedonic consequences of smoking cessation and maybe highly relevant for the efficacy of treatment strategies.
This study was supported by the National Institute on Drug Abuse(NIDA), Intramural Research Program.
We thank Loretta Spurgeon, Kimberley Slater, Eliscia Smith, andNIDA nursing and NIDA recruitment staff for their invaluable assis-tance in running this protocol. We also thank Drs. J. Waltz and J. Gold(Maryland Psychiatric Research Centre), S. McClure (Stanford Univer-sity), and J. Schweitzer (University of California Davis) for their contri-butions to the design of this version of the temporal difference error/juice paradigm.
The authors report no biomedical financial interests or potentialconflicts of interest.
Supplementary material cited in this article is available online.
1. Di Chiara G (2000): Role of dopamine in the behavioural actions ofnicotine related to addiction. Eur J Pharmacol 393:295–314.
2. Gotti C, Guiducci S, Tedesco V, Corbioli S, Zanetti L, Moretti M, et al.(2010): Nicotinic acetylcholine receptors in the mesolimbic pathway:Primary role of ventral tegmental area alpha6beta2* receptors in medi-ating systemic nicotine effects on dopamine release, locomotion, andreinforcement. J Neurosci 30:5311–5325.
3. Janhunen S, Ahtee L (2007): Differential nicotinic regulation of the ni-
grostriatal and mesolimbic dopaminergic pathways: Implications fordrug development. Neurosci Biobehav Rev 31:287–314.3
www.sobp.org/journal
4. Zoli M, Moretti M, Zanardi A, McIntosh JM, Clementi F, Gotti C (2002):Identification of the nicotinic receptor subtypes expressed on dopami-nergic terminals in the rat striatum. J Neurosci 22:8785– 8789.
5. Diekhof EK, Falkai P, Gruber O (2008): Functional neuroimaging of re-ward processing and decision-making: A review of aberrant motiva-tional and affective processing in addiction and mood disorders. BrainRes Rev 59:164 –184.
6. McClure SM, York MK, Montague PR (2004): The neural substrates ofreward processing in humans: The modern role of fMRI. Neuroscientist10:260 –268.
7. O’Doherty JP (2004): Reward representations and reward-related learn-ing in the human brain: Insights from neuroimaging. Curr Opin Neuro-biol 14:769 –776.
8. Elliott R, Dolan RJ, Frith CD (2000): Dissociable functions in the medialand lateral orbitofrontal cortex: Evidence from human neuroimagingstudies. Cereb Cortex 10:308 –317.
9. Montague PR, Berns GS (2002): Neural economics and the biologicalsubstrates of valuation. Neuron 36:265–284.
0. Knutson B, Fong GW, Bennett SM, Adams CM, Homme D (2003): Aregion of mesial prefrontal cortex tracks monetarily rewarding out-comes: Characterization with rapid event-related fMRI. Neuroimage 18:263–272.
1. Knutson B, Fong GW, Adams CM, Varner JL, Hommer D (2001): Dissoci-ation of reward anticipation and outcome with event-related fMRI. Neu-roreport 12:3683–3687.
2. O’Doherty JP, Deichmann R, Critchley HD, Dolan RJ (2002): Neural re-sponses during anticipation of a primary taste reward. Neuron 33:815–826.
3. McClure SM, Berns GS, Montague PR (2003): Temporal prediction errorsin a passive learning task activate human striatum. Neuron 38:339 –346.
4. Berns GS, McClure SM, Pagnoni G, Montague PR (2001): Predictabilitymodulates human brain response to reward. J Neurosci 21:2793–2798.
5. Pagnoni G, Zink CF, Montague PR, Berns GS (2002): Activity in humanventral striatum locked to errors of reward prediction. Nat Neurosci5:97–98.
6. Montague PR, Dayan P, Sejnowski TJ (1996): A framework for mesen-cephalic dopamine systems based on predictive hebbian learning.J Neurosci 16:1936 –1947.
7. Schultz W, Dayan P, Montague PR (1997): A neural substrate of predic-tion and reward. Science 275:1593–1599.
8. Hollerman JR, Schultz W (1998): Dopamine neurons report an error in thetemporal prediction of reward during learning. Nat Neurosci 1:304–309.
9. Schultz W (1998): Predictive reward signal of dopamine neurons. J Neu-rophysiol 80:1–27.
0. Wise RA (2009): Roles for nigrostriatal–not just mesocorticolimbic–dopamine in reward and addiction. Trends Neurosci 32:517–524.
1. Schultz W (2000): Multiple reward signals in the brain. Nat Rev Neurosci1:199 –207.
2. Schultz W (2002): Getting formal with dopamine and reward. Neuron36:241–263.
3. Schultz W (2007): Behavioral dopamine signals. Trends Neurosci 30:203–210.
4. Montague PR, Hyman SE, Cohen JD (2004): Computational roles fordopamine in behavioural control. Nature 431:760 –767.
5. Koob GF, Sanna PP, Bloom FE (1998): Neuroscience of addiction. Neuron21:467– 476.
6. Zhou FM, Liang Y, Dani JA (2001): Endogenous nicotinic cholinergicactivity regulates dopamine release in the striatum. Nat Neurosci4:1224 –1229.
7. Boileau I, Dagher A, Leyton M, Welfeld K, Booij L, Diksic M, Benkelfat C(2007): Conditioned dopamine release in humans: A positron emissiontomography [11c]raclopride study with amphetamine. J Neurosci 27:3998 – 4003.
8. Redish AD (2004): Addiction as a computational process gone awry.Science 306:1944 –1947.
9. Azizian A, Monterosso J, O’Neill J, London ED (2009): Magnetic reso-nance imaging studies of cigarette smoking. In: Henningfield JE, Lon-don ED, Pogun S, editors. Nicotine Psychopharmacology. Berlin: Springer-Verlag, 113–143.
0. Brody AL, Mandelkern MA, Olmstead RE, Jou J, Tiongson E, Allen V, et al.(2007): Neural substrates of resisting craving during cigarette cue expo-sure. Biol Psychiatry 62:642– 651.
1. David SP, Munafo MR, Johansen-Berg H, Smith SM, Rogers RD,Matthews PM, Walton RT (2005): Ventral striatum/nucleus accumbens

5
5
6
6
6
6
6
6
6
6
6
6
7
7
7
7
7
7
7
7
7
7
E.J. Rose et al. BIOL PSYCHIATRY 2012;71:206–213 213
activation to smoking-related pictorial cues in smokers andnonsmokers: A functional magnetic resonance imaging study. BiolPsychiatry 58:488 – 494.
32. Due DL, Huettel SA, Hall WG, Rubin DC (2002): Activation in mesolimbicand visuospatial neural circuits elicited by smoking cues: Evidence fromfunctional magnetic resonance imaging. Am J Psychiatry 159:954 –960.
33. McBride D, Barrett SP, Kelly JT, Aw A, Dagher A (2006): Effects of expectancyand abstinence on the neural response to smoking cues in cigarette smok-ers: An fMRI study. Neuropsychopharmacology 31:2728–2738.
34. Smolka MN, Buhler M, Klein S, Zimmermann U, Mann K, Heinz A,Braus DF(2006): Severity of nicotine dependence modulates cue-induced brainactivity in regions involved in motor preparation and imagery. Psycho-pharmacology (Berl) 184:577–588.
35. Zhang X, Salmeron BJ, Ross TJ, Gu H, Geng X, Yang Y,Stein EA (2011):Anatomical differences and network characteristics underlying smok-ing cue reactivity. Neuroimage 54:131–141.
36. Koob GF, Nestler EJ (1997): The neurobiology of drug addiction. J Neu-ropsychiatry Clin Neurosci 9:482– 497.
37. Clementi F, Fornasari D, Gotti C (2000): Neuronal nicotinic acetylcholinereceptors: From structure to therapeutics. Trends Pharmacol Sci 21:35–37.
38. Picciotto MR, Caldarone BJ, King SL, Zachariou V (2000): Nicotinic recep-tors in the brain. Links between molecular biology and behavior. Neu-ropsychopharmacology 22:451– 465.
39. Picciotto MR (2003): Nicotine as a modulator of behavior: Beyond theinverted u. Trends Pharmacol Sci 24:493– 499.
40. Oldfield RC (1971): The assessment and analysis of handedness: TheEdinburgh Inventory. Neuropsychology 9:97–113.
41. Wechsler D (2007): Wechsler Abbreviated Scale of Intelligence (WASI). SanAntonio, TX: Pearson/PsychCorp.
42. Reitan RM, Wolfson D (1993): The Halstead-Reitan NeuropsychologicalTest Battery. Tuscan: Neuropsychology Press.
43. Wechsler D (1987): Wecshler Memory Scale - Revised Manual. San Anto-nio: The Psychological Corporation.
44. Cloninger CR, Przybeck TR, Svrakic DM, Wetzel RD (1994): The Temperamentand Character Inventory (TCI): A Guide to its Development and Use. St. Louis:Washington University: Center for Psychobiology of Personality.
45. Heatherton TF, Kozlowski LT, Frecker RC, Fagerstrom KO (1991): TheFagerström test for nicotine dependence - a revision of the Fagerstromtolerance questionnaire. Br J Addict 86:1119-1127.
46. Gorsline J, Gupta SK, Dye D, Rolf CN (1993): Steady-state pharmacoki-netics and dose relationship of nicotine delivered from nicoderm® (nic-otine transdermal system). J Clin Pharmacol 33:161–168.
47. Hendricks PS, Ditre JW, Drobes DJ, Brandon TJ (2006): The early timecourse of smoking withdrawal effects. Psychopharmacology (Berl) 187:385–396.
48. Parrott AC, Garnham NJ, Wesnes K, Pincock C (1996): Cigarette smokingand abstinence: Comparative effects upon cognitive task performanceand mood state over 24 hours. Hum Psychopharmacol 11:391– 400.
49. Heishman SJ, Singleton EG, Moolchan ET (2003): Tobacco Craving Ques-tionnaire: Reliability and validity of a new multifactorial instrument.Nicotine Tob Res 5:645– 654.
50. Heishman SJ, Singleton EG, Pickworth WB (2007): Reliability and validityof a short form of the Tobacco Craving Questionnaire. Nicotine Tob Res10:643– 651.
51. Harrington DL, Boyd LA, Mayer AR, Sheltraw DM, Lee RR, Huang M, RaoSM (2004): Neural representation of interval encoding and decisionmaking. Brain Res Cogn Brain Res 21:193–205.
52. Shakleya DM, Huestis MA (2009): Optimization and validation of a liquidchromatography-tandem mass spectrometry method for the simulta-neous quantification of nicotine, cotinine, trans-3’-hydroxycotinine andnorcotinine in human oral fluid. Anal Bioanal Chem 395:2349 –2357.
53. Cox RW (1996): AFNI software for analysis and visualization of functionalmagnetic resonance neuroimages. Comput Biomed Res 29:162–173.
54. Talairach J, Tournoux P (1988): Co-Planar Stereotaxic Atlas of the HumanBrain. New York: Thieme.
55. Cloninger CR (1986): A unified biosocial theory of personality and its rolein the development of anxiety states. Psychiatr Dev 4:167–226.
56. Caggiula AR, Donny EC, Palmatier MI, Liu X, Chaudhri N, Sved AF (2009):The role of nicotine in smoking: A dual-reinforcement model. Nebr SympMotiv 55:91–109.
57. Glautier S, Clements K, White JA, Taylor C, Stolerman IP (1996): Alcohol and
the reward value of cigarette smoking. Behav Pharmacol 7:144–154.8. Sharma A, Brody AL (2009): In vivo brain imaging of human exposure tonicotine and tobacco. Handb Exp Pharmacol 192:145–171.
9. Penton RE, Lester RA (2009): Cellular events in nicotine addiction. SeminCell Dev Biol 20:418 – 431.
0. Xiao C, Nashmi R, McKinney S, Cai H, McIntosh JM, Lester HA (2009):Chronic nicotine selectively enhances alpha4beta2* nicotinic acetyl-choline receptors in the nigrostriatal dopamine pathway. J Neurosci29:12428 –12439.
1. McCallum SE, Parameswaran N, Bordia T, Fan H, Tyndale RF, LangstonJW, et al. (2006): Increases in alpha4* but not alpha3*/alpha6* nicotinicreceptor sites and function in the primate striatum following chronicoral nicotine treatment. J Neurochem 96:1028 –1041.
2. Nashmi R, Xiao C, Deshpande P, McKinney S, Grady SR, Whiteaker P, et al.(2007): Chronic nicotine cell specifically upregulates functional alpha4*nicotinic receptors: Basis for both tolerance in midbrain and en-hanced long-term potentiation in perforant path. J Neurosci 27:8202–8218.
3. Champtiaux N, Gotti C, Cordero-Erausquin M, David DJ, Przybylski C,Lena C, et al. (2003): Subunit composition of functional nicotinic recep-tors in dopaminergic neurons investigated with knock-out mice. J Neu-rosci 23:7820 –7829.
4. Wessel J, McDonald SM, Hinds DA, Stokowski RP, Javitz HS, Kennemer M,et al. (2010): Resequencing of nicotinic acetylcholine receptor genesand association of common and rare variants with the Fagerström Testfor Nicotine Dependence. Neuropsychopharmacology 35:2392–2402.
5. Baker TB, Weiss RB, Bolt D, von Niederhausern A, Fiore MC, Dunn DM, etal. (2009): Human neuronal acetylcholine receptor a5-a3-b4 haplotypesare associated with multiple nicotine dependence phenotypes. NicotineTob Res 11:785–796.
6. Piper ME, McCarthy DE, Bolt DM, Smith SS, Lerman C, Benowitz N, et al.(2008): Assessing dimensions of nicotine dependence: An evaluation ofthe Nicotine Dependence Syndrome Scale (NDSS) and the WisconsinInventory of Smoking Dependence Motives (WISDM). Nicotine Tob Res10:1009 –1020.
7. De Ruyck K, Nackaerts K, Beels L, Werbrouck J, De Volder A, Meysman M,et al. (2010): Genetic variation in three candidate genes and nicotinedependence, withdrawal and smoking cessation in hospitalized pa-tients. Pharmacogenomics 11:1053–1063.
8. Perkins KA, Karelitz JL, Conklin CA, Sayette MA, Giedgowd GE (2010):Acute negative affect relief from smoking depends on the affect situa-tion and measure but not on nicotine. Biol Psychiatry 67:707–714.
9. Boden JM, Fergusson DM, Horwood LJ (2010): Cigarette smoking anddepression: Tests of causal linkages using a longitudinal birth cohort.Br J Psychiatry 196:440 – 446.
0. Hajek P, Taylor T, McRobbie H (2010): The effect of stopping smoking onperceived stress levels. Addiction 105:1466 –1471.
1. Benowitz NL (2009): Pharmacology of nicotine: Addiction, smoking-induced disease, and therapeutics. Annu Rev Pharmacol Toxicol 49:57–71.
2. Crunelle CL, Miller ML, Booij J, van den Brink W (2010): The nicotinicacetylcholine receptor partial agonist varenicline and the treatment ofdrug dependence: A review. Eur Neuropsychopharmacol 20:69 –79.
3. Niv Y, Schoenbaum G (2008): Dialogues on prediction errors. TrendsCogn Sci 12:265–272.
4. Oyama K, Hernadi I, Iijima T, Tsutsui K (2010): Reward prediction errorcoding in dorsal striatal neurons. J Neurosci 30:11447–11457.
5. D’Ardenne K, McClure SM, Nystrom LE, Cohen JD (2008): Bold responsesreflecting dopaminergic signals in the human ventral tegmental area.Science 319:1264 –1267.
6. Logothetis NK, Pauls J, Augath M, Trinath T, Oeltermann A (2001): Neu-rophysiological investigation of the basis of the fMRI signal. Nature412:150 –157.
7. Waltz JA, Schweitzer JB, Gold JM, Kurup PK, Ross TJ, Salmeron BJ, et al.(2009): Patients with schizophrenia have a reduced neural response toboth unpredictable and predictable primary reinforcers. Neuropsycho-pharmacology 34:1567–1577.
8. Corfield DR, Murphy K, Josephs O, Adams L, Turner R (2001): Doeshypercapnia-induced cerebral vasodilation modulate the hemody-namic response to neural activation? Neuroimage 13:1207–1211.
9. Hoffmann D, Hoffmann I (1997): The changing cigarette, 1950 –1995. J
Toxicol Environ Health 50:307–364.www.sobp.org/journal
Copyright © 2022 FDOKUMEN




















