
Glutamate receptor expression in rat striatum: Effect of deafferentation
11
BRAIN RESEARCH ELSEVIER Brain Research 647 (1994) 209-219 Research Report Glutamate receptor expression in rat striatum: Effect of deafferentation Ullrich Wiillner, David G. Standaert, Claudia M. Testa, G. Bernhard Landwehrmeyer, Maria Vincenza Catania, John B. Penney Jr., Anne B. Young * Neurology Sen'ice, Massachusetts General Hospital, and Harvard Medical School, Fruit Street, Boston, MA 02114, USA Accepted 15 February 1994 Abstract The cerebral cortex is the primary source of glutamatergic afferents to the neostriatum. We used in situ hybridization to examine the effect of removal of the glutamatergic input to the striatum by unilateral frontal cortical ablation on the expression of genes encoding subunits from three families of glutamate receptors: N-methyl-D-aspartate receptors (NMDAR1, NMDAR2A, and NMDAR2B); a-amino-3-hydroxy-5-methyl-4-isoxazolepropionate (AMPA) receptors (GluR1-4, flip and flop splice variants); and metabotropic receptors (mGluR1-5). Significant changes were restricted to the dorsolateral quadrant of the ipsilateral striatum, the main projection area of the sensorimotor cortex. The expression of those messages which are normally abundant, NMDAR1, NMDAR2A, GIuR1-4 flop and mGluR1, 3 and 5, was decreased in the deafferented dorsolateral striatum by 10-39% at 3 days after cortical ablation and subsequently increased to 120-165% of control at 15 and 60 days. mRNAs encoding the flip isoforms of GIuR1-4, mGluR2 and 4, and an alternatively spliced region of NMDAR1 (Insertion I) which are undetectable or present at low levels in the striatum were not induced by cortical ablation. In contrast, both glial fibrillary acid protein and /3-actin mRNA expression were markedly enhanced at 3 and 15 days, returning to near normal at 60 days. Striatal NMDA, AMPA and metabotropic type 1 ligand binding sites were increased as early as 3 days after cortical ablation, reached a peak at 15 days and remained increased for up to 60 days, while metabotropic type 2 binding was slightly but significantly reduced at 3 and 15 days and [3H]kainate binding did not change significantly. These results demonstrate that cortical ablation, and subsequent loss of glutamatergic afferents to the striatum, results in alterations in the expression of genes encoding glutamate receptor subunits in striatal neurons. The regulation of these genes appears to be coordinate, so that the relative abundance of the different messages is preserved. Key words: N-methyl-D-aspartate; AMPA; Kainate; Metabotropic; Decortication; Neostriatum 1. Introduction The striatum receives a large and highly ordered excitatory amino acid (EAA) projection from the entire cerebral cortex [9,18,43]. Ablation of a frontal hemicor- tex leads to a significant decrease in glutamate levels and [3H]glutamate uptake in the ipsilateral striatum [6,15]. Experimental data indicate that the corticostri- atal projection has an important role in the regulation of striatal neuropeptide levels; also, glutamic acid de- carboxylase (GAD) mRNA expression in the striatum is altered after a cortical lesion [30,33,40]. * Corresponding author. Fax: (1) (617) 726-2353. 0006-8993/94/$07.00 © 1994 Elsevier Science B.V. All rights reserved SSDI 0006-8993(94)00239-9 The effects of glutamate on striatal neurons are mediated via the ion channel-coupled N-methyl-D- aspartate (NMDA), a-amino-3-hydroxy-5-methyl-4-iso- xazolepropionate (AMPA) and kainate (KA) receptors, and the G-protein-coupled metabotropic (mGluR) classes of EAA receptors; high affinity binding sites for all four classes have been demonstrated in the striatum [2,44]. Ligand binding studies of striatal EAA receptors after deafferentation have yielded inconsistent results. After mechanical removal of the ipsilateral hemicortex, total [3H]glutamate binding was found to be increased to 123-134% of control [7,29] or unchanged [19], [3H]AMPA binding was either decreased by 30% [8]or unchanged [12], [3H]CPP binding (to NMDA recep- tors) was increased to 113-135% [31] while for NM-
Transcript of Glutamate receptor expression in rat striatum: Effect of deafferentation

BRAIN RESEARCH
ELSEVIER Brain Research 647 (1994) 209-219
Research Report
Glutamate receptor expression in rat striatum: Effect of deafferentation
Ullrich Wiillner, David G. Standaert, Claudia M. Testa, G. Bernhard Landwehrmeyer, Maria Vincenza Catania, John B. Penney Jr., Anne B. Young *
Neurology Sen'ice, Massachusetts General Hospital, and Harvard Medical School, Fruit Street, Boston, MA 02114, USA
Accepted 15 February 1994
Abstract
The cerebral cortex is the primary source of glutamatergic afferents to the neostriatum. We used in situ hybridization to examine the effect of removal of the glutamatergic input to the striatum by unilateral frontal cortical ablation on the expression of genes encoding subunits from three families of glutamate receptors: N-methyl-D-aspartate receptors (NMDAR1, NMDAR2A, and NMDAR2B); a-amino-3-hydroxy-5-methyl-4-isoxazolepropionate (AMPA) receptors (GluR1-4, flip and flop splice variants); and metabotropic receptors (mGluR1-5). Significant changes were restricted to the dorsolateral quadrant of the ipsilateral striatum, the main projection area of the sensorimotor cortex. The expression of those messages which are normally abundant, NMDAR1, NMDAR2A, GIuR1-4 flop and mGluR1, 3 and 5, was decreased in the deafferented dorsolateral striatum by 10-39% at 3 days after cortical ablation and subsequently increased to 120-165% of control at 15 and 60 days. mRNAs encoding the flip isoforms of GIuR1-4, mGluR2 and 4, and an alternatively spliced region of NMDAR1 (Insertion I) which are undetectable or present at low levels in the striatum were not induced by cortical ablation. In contrast, both glial fibrillary acid protein and /3-actin mRNA expression were markedly enhanced at 3 and 15 days, returning to near normal at 60 days. Striatal NMDA, AMPA and metabotropic type 1 ligand binding sites were increased as early as 3 days after cortical ablation, reached a peak at 15 days and remained increased for up to 60 days, while metabotropic type 2 binding was slightly but significantly reduced at 3 and 15 days and [3H]kainate binding did not change significantly. These results demonstrate that cortical ablation, and subsequent loss of glutamatergic afferents to the striatum, results in alterations in the expression of genes encoding glutamate receptor subunits in striatal neurons. The regulation of these genes appears to be coordinate, so that the relative abundance of the different messages is preserved.
Key words: N-methyl-D-aspartate; AMPA; Kainate; Metabotropic; Decortication; Neostriatum
1. Introduct ion
The striatum receives a large and highly ordered excitatory amino acid (EAA) projection from the entire cerebral cortex [9,18,43]. Ablation of a frontal hemicor- tex leads to a significant decrease in glutamate levels and [3H]glutamate uptake in the ipsilateral striatum [6,15]. Experimental data indicate that the corticostri- atal projection has an important role in the regulation of striatal neuropeptide levels; also, glutamic acid de- carboxylase (GAD) m R N A expression in the striatum is altered after a cortical lesion [30,33,40].
* Corresponding author. Fax: (1) (617) 726-2353.
0006-8993/94/$07.00 © 1994 Elsevier Science B.V. All rights reserved SSDI 0006-8993(94)00239-9
The effects of glutamate on striatal neurons are mediated via the ion channel-coupled N-methyl-D- aspartate (NMDA), a-amino-3-hydroxy-5-methyl-4-iso- xazolepropionate (AMPA) and kainate (KA) receptors, and the G-protein-coupled metabotropic (mGluR) classes of EAA receptors; high affinity binding sites for all four classes have been demonstrated in the striatum [2,44]. Ligand binding studies of striatal EAA receptors after deafferentation have yielded inconsistent results. After mechanical removal of the ipsilateral hemicortex, total [3H]glutamate binding was found to be increased to 123-134% of control [7,29] or unchanged [19], [3H]AMPA binding was either decreased by 30% [8]or unchanged [12], [3H]CPP binding (to NMDA recep- tors) was increased to 113-135% [31] while for NM-

21(I U. Wiillner et al. /Brain Research 647 (1994) 209-219
Table 1 Sequences of the 19 oligonucleotide probes used for in situ hybridization and the amino acid regions to which they are complementary
Receptor Amino Oligonucleotide sequence (5'-3') subunit acids
NMDAR1 138-152 AAA CCA GAC GET GGA CTG NMDAR1 InsI 197-211 CT'l" GGG TCC GCG ET'I" G'I'T NMDAR2A 567-579 AGA AGG CCC GTG GGA GCT
NMDAR2B 557-572 GGG CCT CCT GGC TCT CTG
GIuR1 flip 771-783 CAA AGC GCT GGT CTT GTC
GluR2f l ip 775-787 GAG GGC ACT GGT CTT TTC
GluR3f l ip 780-791 TAG AGC ACT GGT CTT GTC
GluR4f l ip 777-788 CAA GGC ACT CGT CTT GTC
GluR1 flop 772-783 CAA AGC GCT GGT CTT GTC
GluR2f lop 776-787 GAG GGC ACT GGT CTT TTC
G l u R 3 f l o p 780-791 TAG AGC ACT GGT CTT GTC
G l u R 4 f l o p 777-788 CAA GGC ACT CGT CTT GTC
mGluR1 484-498 CCC CAC GTG GAC ATA GTC
mGluR2 123-137 GGT GAC AGC TGT AGG AGC
mGluR3 841-855 CTG AGA ATA GGT GGT TGC
mGluR4 881-892 GGT CTC CAG GTT CTC ACA
mGluR5 124-138 GGA GCG GAA GGA AGA AGA
/3-actin 326-341 GCC GAT CCA CAC GGA GTA
GFAP 365-381 TTA CCA CGA TGT TCC TCT
GTG GGA GTA GGG CGG CAC CGT GCG AAG
GTC ATA GGA CAG TTG GTC GAG GTT TTC
TTC CCT TTG GCT AAG TTT C
CCA TCG GCT AGG CAC CTG TTG TAA CCC
CTT ACT TCC GGA GTC CTT GCT
CTT ACT TCC CGA GTC CTT GGC
CTT ACT CCC GGA GTC CTT
CTT GCT TCC CGA GTC CTT
CTT GGA GTC ACC TCC CCC
CTT GGA ATC ACC TCC CCC
CTT GGA GTC ACC GCC CCC
CTT GGA GTC ACC TCC CCC
ATA GCG ATT AGC TTC TGT GTA CTG CAG
ATC ACT GTG GGT GGC ATA GGA GCC ATC
AGT TCC GCT GAC GCT GAA CCT GTT GAG
CAG CTC TGA TTT GGC TTC CCC ATT GGG
TCC ATC TAC ACA GCG TAC CAA ACC TTC
CTT GCG CTC AGG AGG AGC AAT GAT CTT GAT
TGA GGT GGC CTT CTG ACA CGG ATT TGG TGT
DA-sensitive [3H]glutamate binding no significant change was reported [12].
Recent progress in cloning of the EAA receptor genes has revealed families of subunits or subtypes for each of the glutamate receptor classes [11,14,20,23,34, 38]. In the striatum in situ hybridization and immuno- cytochemical studies have demonstrated the expression of the NMDA receptor subunits NMDAR1, NMDAR2A, NMDAR2B, the AMPA receptor sub- units GIuR1-4, and the metabotropic receptor sub- types mGluR1-5 [17,32,35,36,39]. It is not known whether modulation of the corticostriatal input might have a differential effect on the expression of the various glutamate receptor isoforms or might induce (or suppress) alternate splicing of these isoforms by striatal neurons. To examine this question, we used both in situ hybridization with oligonucleotide probes and ligand binding methods to study the expression of these genes in the rat striatum after cortical ablation.
2. Materials and methods
2.1. Cortical ablation
Male Sprague-Dawley rats (Charles River, 200-250 g) were anes- thetized and positioned in a Kopf stereotaxic frame. A skull flap, approximately 6 mm anterior and posterior to bregma and 1 mm lateral from the midline to the lateral edge of the calvarium was removed. Decortication was performed by aspiration of the cortex and undercutt ing of the cortex lateral to the opening with a suture needle. Gelfoam and 30% H 2 0 2 were used for hemostasis. Animals were sacrificed by decapitation 3, 15 or 60 days after surgery. Brains were rapidly removed, frozen on dry ice, and 12 tzm coronal striatal sections were cut on a cryostat, mounted on poly-L-lysine coated
slides and stored at - 7 0 ° C until use. To verify the extent of the lesion, regularly spaced sections were processed for routine Nissl stain. Only those animals which had complete removal of the fronto- parietal cortex without damage to the striatum were considered further in the study; however, most of these animals had some distortion of the ipsilateral corpus callosum. For each animal (n = 4 -6 per time point) serially adjacent sections were processed for each ligand binding assay and in situ hybridization with each of the 19 probes.
2.2. In situ hybridization histochemistry
Oligodeoxyribonucleotide probes (36-48 bases in length), com- plementary to the mRNAs encoding the NMDA, A M P A and
Fig. 1. Schematic figure illustrating the region of the dorsolateral striatum which receives the major projection from the sensorimotor neocortex (*), modified according to McGeorge and Faull [18]. This region was used for quantitative analysis.

U. Wiillner et a l . / Brain Research 647 (1994) 209-219
Table 2 Summary of the excitatory amino acid receptor ligands and binding assay protocols
211
Binding Ligand Incubation buffer Blocker a Blank b site
NMDA 60 nM 50 mM Tris-acetate, pH 7.4 2.5 tzM quisqualate 1 mM receptor [ 3 H]glutamate 1 /x M kainate NMDA AMPA 30 nM 50 mM Tris-HCl, 2.5 mM - 1 mM receptor [3H]AMPA CaCI 2, 30 mM KSCN, pH 7.2 glutamate Kainate 60 nM 50 mM Tris-acetate, pH 7.4 - 100/zM receptor [ 3H]kainate kainate MET 1 100 nM 50 mM Tris-HCl, 2.5 mM 100/zM NMDA 2.5 ~M receptor [3H]glutamate CaC12, 30 mM KSCN, pH 7.2 10 ~zM AMPA quisqualate MET 2 100 nM 50 mM Tris-HCI, 2.5 mM 100/xM NMDA 500/zM receptor [3H]glutamate CaCI2, 30 mM KSCN, pH 7.2 10 p~M AMPA t-ACPD
2.5/xM quisqualate
a Drugs to block binding to other receptors. h Drugs to estimate unspecific binding. All steps were performed at 4°C.
metabotropic glutamate receptor subtypes, glial fibrillary acid pro- tein (GFAP) and /3-actin, were constructed according to published cDNA sequences (Table 1) [16,21,27,34,37,38], No significant ho- mologies other than to the target sequences were found using the
National Center for Biotechnology Information BLAST network service [3]. AMPA receptor probes were synthesized by the DNA Synthesis Core Facility (University of Michigan, Ann Arbor) on Applied Biosystems automated DNA synthesizers and HPLC puri-
(
Fig. 2. Autoradiograms of coronal sections from unlesioned animals (a, b) and animals 15 days after cortical ablation (c, d, e, f) hybridized with oligonucleotide probes complementary to the NMDA receptor subunits NMDAR1 (a, c), NMDAR1 Ins I (b, d), NMDAR2A (e), NMDAR2B (f).

212 U. Wiillner et al. / Brain Research 647 (1994) 209-219
fied using C-4, 300 ,~ columns. All other probes were synthesized using an Applied Biosystems 392 Synthesizer and purified by elec- trophoresis through a 12% polyacrylamide gel containing 8 M urea. All probes were 3' end-labeled with 3SS-adATP (specific activity > 1000 Ci /mM, Dupont-NEN) by terminal deoxynucleotidyl trans- ferase (NEP 100 kit, Dupont-NEN). Unincorporated 35S was sepa- rated using a molecular sizing column (Stratagene). For in situ hybridization, sections were fixed in 4% paraformaldehyde in 0.1 M phosphate buffer, pH 7.4, washed three times in 0.1 M phosphate buffered saline (PBS), acetylated in 0.1 M triethanolamine with 0.25%, acetic anhydride and dehydrated through graded ethanol solutions. Hybridization was carried out overnight at 37°C in 50% formamide, 0.3 M NaCI, 10 mM Tris, 1 mM EDTA, 1(J% dextran
sulfate, 1 x Denhardt 's solution (Sigma) and 100 mM dithiothreitol with 30,000 dpm/p~l of radiolabeled probe. Following hybridization the slides were washed in 2 x S S C (standard saline citrate, 0.15 M NaCI, 0.015 M sodium citrate) at room temperature (RT) for 20 min, 1 x SSC at RT for 10 min, 0.5 x SSC at 60°C for 40 min and 0.5 x SSC at RT for 10 min, rinsed in ethanol for 2 min, air dried and apposed to /3-max film (Amersham) for 3 weeks. Films were developed in Kodak D-19 and analyzed using the MCID image processing system (Imaging Research, St. Catharine's, Ont.). For data acquisition the program allows the user to define a threshold level of signal intensity for a region of interest. By using the striatal white matter bundles to define this threshold only gray matter signal in the striatum was measured. For analysis of the film autoradiograms optical density
b
Fig. 3. Autoradiograms of coronal sections from animals 15 days after cortical ablation, hybridized with oligonucleotide probes complementary to the AMPA receptor subunits GIuR1 flop (a), GIuR1 flip (b), GIuR2 flop (c), GluR2 flip (d), GIuR3 flop (el, GluR3 flip (f), GIuR4 flop (g), GluR4 flip (h).

U. Wiillner et a l . / Brain Research 647 (1994) 209-219 213
readings from four sections per animal per probe were averaged and the dorsolateral striatum (Fig. 1) ipsilateral to the lesion compared to the contralateral side using a paired Student's t-test. A change in signal intensity for a given probe was considered significant only if additional one-way analysis of variance within the time course showed a difference from the first timepoint (P < 0.01). Pretreatment with RNase A or addition of a 50-fold excess of unlabeled probe to the hybridization solution resulted in complete signal loss for all probes used. On the other hand, addition of a 50-fold excess of poly- deoxyadenosine had no effect on the hybridization signal.
binding was performed on three animals at 15 days post-lesion with 10 different concentrations of [3H]AMPA ranging from 5 nM to 5 mM. Scatchard plots of the resulting binding data were fitted using an iterative curve fitting program (LIGAND) [22].
3. Results
3.1. In situ hybridization histochemistry
2.3. Ligand binding
Autoradiographic assays for [3H]AMPA, [3H]KA, NMDA-sensi- tive [3H]glutamate and metabotropic-sensitive [3H]glutamate binding sites type 1 (quisqualate-sensitive) and type 2 (quisqualate-insensi- tive) were performed as described previously (Table 2) [4,42]. The autoradiograms were analyzed using the MC|D image processing system. For each assay, the regions of interest in each animal were analyzed on 3 sections for total binding and 2 sections for nonspecific binding. Image density was converted into bound radioactivity using a computer-generated polynomial regression curve derived from the radioactive standards; specific binding was calculated by subtracting nonspecific from total binding. A saturation analysis of [3H]AMPA
In accordance with previous studies, each EAA re- ceptor subunit probe demonstrated a unique labeling pattern. Signal intensity in the striatum varied greatly: probes for NMDAR1, GluR1-4 flop splice variants and mGluR5 gave the highest signals, whereas GluR1 flip, GluR4 flip and mGluR2 were not detectable (Figs. 2-4). MGluR3 hybridization signal was present both in the striatum and in white matter regions (Fig. 4c). Within the striatum, a lateromedial gradient of inten- sity from was observed for GluR1 and GluR3 flop and NMDAR2A, with the most intense signal laterally
8
~ g '
b
A
Fig. 4. Autoradiograms of coronal sections from animals 15 days after cortical ablation, hybridized with oligonucleotide probes complementary to the metabotropic receptors mGIuR1 (a), mGluR2 (b), mGluR3 (c), mGluR4 (d), mGluR5 (e).

214 U. Wiillner et al. /Bra in Research 647 (1994) 209-219
(Figs. 2 and 3). GluR2 flip also exhibited a gradient, with more intense labeling medially (Fig. 3c,d). GluR1-3 flip and mGluR5 gave higher signal in the nucleus accumbens than in the striatum (Figs. 3 and 4). The probe directed to the alternatively spliced region of NMDAR1, Insertion 1 (Ins I) [37] showed a re- stricted area of somewhat more intense signal in the ventrolateral striatum (Fig. 2b); in contrast, in the same region, probes for mGluR3 and mGluR4 gave a de- creased signal (Fig. 4c,d).
After fronto-parietal hemicortex ablations, changes in hybridization signals were observed only in the dor- solateral portion of the striatum, the main projection target of the neocortical sensorimotor areas. When the entire striatum was compared to the contralateral side, no significant change in the expression of any EAA receptor subunits at any timepoint was found. How- ever, subdivision of the striatum revealed consistent changes in the expression of some of the subunits in the dorsolateral portion ipsilateral to the lesion, com- pared to the corresponding region on the contralateral
side (Figs. 1-5). Signal intensity in the dorsolateral striatum contralateral to the lesion was not signifi- cantly different from values obtained with each of these probes in non-lesioned animals (Fig. 2).
Statistically significant changes in hybridization sig- nal after lesion were observed for NMDAR1 and NM- DAR2A, GluR1-4 flop and mGluR1, 3 and 5 (Fig. 5). In each case, the abundance of the message in the dorsolateral quadrant of the ipsilateral striatum was decreased at 3 days after cortical ablation, and in- creased above control levels at 15 and 60 days after the lesion (Fig. 5). NMDAR2B, GluR2 and 3 flip and mGluR4 mRNA expression exhibited a trend toward the same pattern, i.e. a decrease after 3 days and a subsequent increase; however, these changes were sig- nificant only when compared to the respective con- tralateral side with a paired Student's t-test (P < 0.05). Among the subunits that were not detectable in non-le- sioned animals, i.e. GIuR1 and 4 flip and mGluR2, GluR1 and 4 flip did exhibit a small but detectable increase. The ratios of GluR1 flop to GluR2-4 flop,
A0.4
0.3
0.2'
0.1
0
A M P A 'flip' AMPA 'flop' splice variants splice variants
B 0.4
0.3
0.~
0.1
0 GluR1 GluR2 GluR3 GluR4 GluR1 GluR2 GluR3 GluR4
C N M D A * D 0.4 mo*oh.*~.~l~ 0.2
0.3
0.1 0.2
0 .1
0 0 NMDA NM DA NMDA NMDA mGluR1 mGluR3 mGluR4 mGluR5
R1 R1 InsI R2A R2B
• CONTROL [ ] 3 DAYS • 15 DAYS • 60 DAYS
Fig. 5. Quantitative optical density measurements of the hybridization signals of oligonucleotide probes complementary to the A M P A receptor subunits GIuR1-4 flip (a), GIuR1-4 flop (b), N M D A receptor subunits NMDAR1, NMDAR1 Ins I, N M D A R 2 A and NMDAR2 B (c) and the metabotropic receptors m G l u R 1 - 5 (d) in the dorsolateral striatum 3, 15 and 60 days after cortical ablation. A change in signal intensity for a given probe was considered significant only if one-way analysis of variance within the time course showed a difference from the first t imepoint (P_< 0.01). Control refers to the intact contralateral side, control values did not differ significantly between the various t imepoints and were averaged. Data presented are mean ± S.E.M., * P < 0.01, ** P < 0.001, paired Student 's t-test, dorsolateral striatum ipsilateral to the lesion compared to the corresponding contralateral side.

U. Wiillner et al. / Brain Research 647 (1994) 209-219 215
GIuR3 flip to GluR2 flip and GluR2-3 flop to flip expression remained unchanged (data not shown). MGIuR2 remained undetectable on the film autoradio- grams after cortical ablation. The hybridization signal of the probe for the alternatively spliced region of NMDAR1, Ins I, was present only at a very low level in normal animals and did not increase after cortical ablation (Fig. 2b,d).
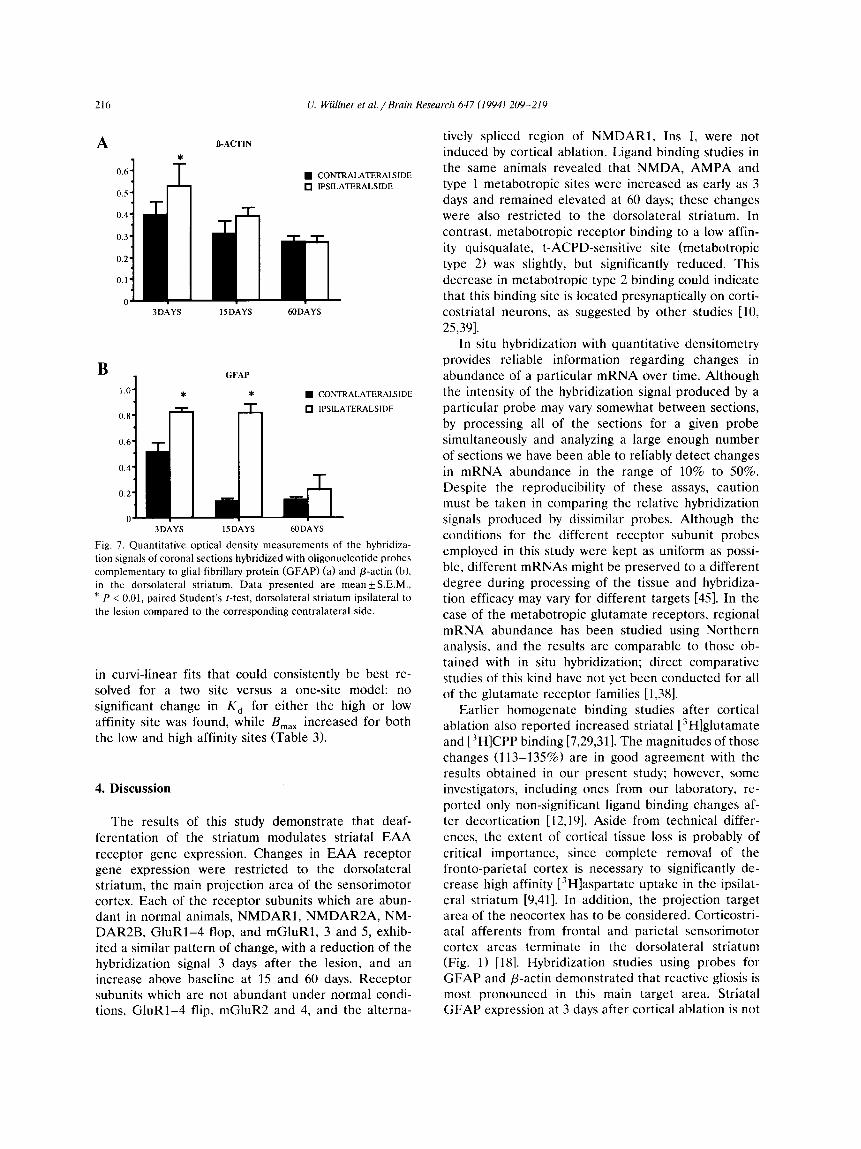
The spatial patterns and time courses of/3-actin and GFAP mRNA expression were very different from those observed for the EAA receptor subunits. The /3-actin signal was increased on the lesioned side at 3 days, especially in the dorsolateral striatum and along the margins of the cortical lesion. The intensity of the /3-actin signal decreased over time after the lesion and was comparable to that of the unlesioned side at 15 days (Figs. 6 and 7). GFAP expression in the striatum was increased 6-8-fold at 3 days, both ipsi- and con- tralateral to the lesion. On the ipsilateral side GFAP was increased throughout the striatum, most promi- nently in the dorsolateral quadrant, while on the con- tralateral side the increase was restricted to the ventro-
lateral portion of the striatum and was no longer present at 15 days. Ipsilateral GFAP was still increased at 15 days, but subsequently returned to near normal levels (Figs. 6 and 7).
3.2. Ligand binding
[3H]AMPA, [3H]kainate and [3H]glutamate (NM- DA-sensitive and metabotropic type 1 and 2) ligand binding studies were performed. Striatal NMDA, AMPA and metabotropic type 1 binding sites were increased as early as 3 days after cortical ablation, reached a peak at 15 days and remained increased for up to 60 days (Figs. 8 and 9). Metabotropic receptor binding type 2 (a low affinity quisqualate, t-ACPD sensitive site) was slightly but significantly reduced on 3 and 15 days (Figs. 8 and 9). [3H]Kainate binding did not change significantly (Figs. 8 and 9).
As with the in situ hybridization signals, changes in ligand binding were more pronounced in the dorsolat- eral striatum. The increase in [3H]AMPA binding was investigated in more detail. Saturation studies resulted
f
Fig. 6. Autoradiograms of coronal sections hybridized with oligonucleotide probes complementary to glial fibrillary protein (GFAP), (a, c, e) and /3-actin (b, d, f), in the dorsolateral striatum 3 (a, b), 15 (c, d) and 60 days (e, f) after cortical ablation.
216 U. Wiillner et al. /Brain Research 647 (1994) 209-219
A
0.6
0.5
0.4
0.3
0.2
0.1
0
B-ACTIN
3DAYS 15DAYS 60DAYS
RALATERALSIDE ~TERALSIDE
B ~ A P
1.0
0.8 T
0.6 I
0.4
0.2
• CONTRALATERALS IDE
[] IPSILATERALSIDE
0 3DAYS 15DAYS 60DAYS
Fig. 7. Quantitative optical density measurements of the hybridiza- tion signals of coronal sections hybridized with oligonucleotide probes complementary to glial fibrillary protein (GFAP) (a) and /3-actin (b), in the dorsolateral striatum. Data presented are mean+S.E.M., * P < 0.01, paired Student's t-test, dorsolateral striatum ipsilateral to the lesion compared to the corresponding contralateral side.
in curvi-linear fits that could consistently be best re- solved for a two site versus a one-site model: no significant change in K d for either the high or low affinity site was found, while Bma x increased for both the low and high affinity sites (Table 3).
4. Discussion
The results of this study demonstrate that deaf- ferentation of the striatum modulates striatal EAA receptor gene expression. Changes in EAA receptor gene expression were restricted to the dorsolateral striatum, the main projection area of the sensorimotor cortex. Each of the receptor subunits which are abun- dant in normal animals, NMDAR1, NMDAR2A, NM- DAR2B, GluR1-4 flop, and mGluR1, 3 and 5, exhib- ited a similar pattern of change, with a reduction of the hybridization signal 3 days after the lesion, and an increase above baseline at 15 and 60 days. Receptor subunits which are not abundant under normal condi- tions, GluR1-4 flip, mGluR2 and 4, and the alterna-
tively spliced region of NMDAR1, Ins I, were not induced by cortical ablation. Ligand binding studies in the same animals revealed that NMDA, AMPA and type 1 metabotropic sites were increased as early as 3 days and remained elevated at 60 days; these changes were also restricted to the dorsolateral striatum. In contrast, metabotropic receptor binding to a low affin- ity quisqualate, t-ACPD-sensitive site (metabotropic type 2) was slightly, but significantly reduced. This decrease in metabotropic type 2 binding could indicate that this binding site is located presynaptically on corti- costriatal neurons, as suggested by other studies [10, 25,39].
In situ hybridization with quantitative densitometry provides reliable information regarding changes in abundance of a particular mRNA over time. Although the intensity of the hybridization signal produced by a particular probe may vary somewhat between sections, by processing all of the sections for a given probe simultaneously and analyzing a large enough number of sections we have been able to reliably detect changes in mRNA abundance in the range of 10% to 50%. Despite the reproducibility of these assays, caution must be taken in comparing the relative hybridization signals produced by dissimilar probes. Although the conditions for the different receptor subunit probes employed in this study were kept as uniform as possi- ble, different mRNAs might be preserved to a different degree during processing of the tissue and hybridiza- tion efficacy may vary for different targets [45]. In the case of the metabotropic glutamate receptors, regional mRNA abundance has been studied using Northern analysis, and the results are comparable to those ob- tained with in situ hybridization; direct comparative studies of this kind have not yet been conducted for all of the glutamate receptor families [1,38].
Earlier homogenate binding studies after cortical ablation also reported increased striatal [3H]glutamate and [3H]CPP binding [7,29,31]. The magnitudes of those changes (113-135%) are in good agreement with the results obtained in our present study; however, some investigators, including ones from our laboratory, re- ported only non-significant ligand binding changes af- ter decortication [12,19]. Aside from technical differ- ences, the extent of cortical tissue loss is probably of critical importance, since complete removal of the fronto-parietal cortex is necessary to significantly de- crease high affinity [3H]aspartate uptake in the ipsilat- eral striatum [9,41]. In addition, the projection target area of the neocortex has to be considered. Corticostri- atal afferents from frontal and parietal sensorimotor cortex areas terminate in the dorsolateral striatum (Fig. 1) [18]. Hybridization studies using probes for GFAP and/3-actin demonstrated that reactive gliosis is most pronounced in this main target area. Striatal GFAP expression at 3 days after cortical ablation is not

U. Wiillner et al. / Brain Research 647 (1994) 209-219
f
217
Fig. 8. Autoradiograms of ligand binding receptor autoradiography of coronal sections 3 (a, b), 15 (c, d) and 60 (e, f) days after cortical ablation for [3H]AMPA (a, c, e) and [3H]kainate (b, d, f) binding. Note the increased binding on the lesioned side for [3H]AMPA but not for [3H]kainate binding.
LIGAND BINDING SITES
175
150
125
~ loo 8
~ 75
50
25
0 NMDA AMPA KAINATE MET l MET2
Fig. 9. Quantitative optical density measurements of [3H]AMPA, [3H]KA, NMDA-sensitive [3H]glutamate and metabotropic-sensitive [3H]glutamate binding sites type 1 (quisqualate-sensitive)and type 2 (t-ACPD-sensitive, quisqualate insensitive) in the dorsolateral stria- tum 3, 15 and 60 days after cortical ablation. Data are presented as percentage of the corresponding contralateral side (100%), mean ± S.E.M., * P < 0.01, ** P < 0.001, paired Student's t-test, dorsolateral striatum ipsilateral to the lesion compared to the corresponding contralateral side.
[] 3 DAYS • 15DAYS • 60DAYS
only increased ipsilaterally but also - to a lesser extent - contralaterally, confirming the existence of bilateral projections (Figs. 6 and 7). Interestingly, the contralat- eral increase was confined to a small ventrolateral area
Table 3 Saturation analysis of [3H]AMPA binding in the striatum 15 days after cortical ablation
High affinity Low affinity
K d (nM) Bma x K d low Bma x (pM/mg protein) (p.M) (pM/mg protein)
Control: 30.4±2.9 0.8±0.1 3.4±0.8 3.9±1.0 Lesion: 30.2±4.1 1.1±0.1" 3.2±0.6 9.9±3.4
Binding was performed on three animals at 15 days post-lesion with 10 different concentrations of [3H]AMPA ranging from 5 nM to 5 raM. Scatchard plots of the resulting binding data were fitted with an iterative curve fitting program (LIGAND) [22] using a two site equation and are presented as mean+S.E.M., * P > 0.05, paired Student's t-test.

218 u. Wi~llner et al. / Brain Research 647 (1994) 209-219
of the striatum, which is character ized by relatively high expression of N M D A R 1 Ins I and low expression of mGluR3 and 4 in control animals (Fig. 2).
We have observed that increases in striatal N M D A and A M P A binding occur rapidly after decortication. Three days after cortical ablation, both N M D A and A M P A sites are increased, while the intensity of the in situ hybridization signals is actually reduced, suggesting that mechanisms other than increased m R N A levels are involved in the immediate response. It is conceiv- able that the early increase in N M D A and A M P A binding sites arises f rom mobilization of preformed intracellular receptor proteins which are inserted into the postsynaptic membrane. Alternatively, the early increase in [ 3 H ] A M P A and NMDA-sens i t ive [3H] glutamate binding, at a time when the m R N A s encod- ing the subunits of these receptors are actually less abundant , could be due to reduced receptor protein turnover, or an increased rate of translation of E A A receptor m R N A .
The changes in E A A gene expression which we observed in the str iatum after cortical lesions support the concept of denervat ion supersensitivity of E A A receptors. The pat tern of change which we observed was a coordinate alteration in expression of all of the receptor subunits which are normally abundant in the striatum. Based on this data we would predict that striatal E A A supersensitivity would be manifest as a quantitative change in neural responsivity, ra ther than a qualitative change in the kinds of responses which E A A s evoke. This is in contrast to changes in E A A receptors observed after transient global ischemia in CA1 neurons of the hippocampus, which showed a selective decrease in GluR2 expression [26].
Al though denervat ion supersensitivity of certain neurot ransmit ter systems, such as dopaminergic recep- tors, is a well character ized phenomenon , similar stud- ies are rare for the glutamatergic system [5,13]. Nico- letti and associates have used measurements of PI hydrolysis in striatal slices to study the response of the metabotropic receptor system to cortical lesions [24]. They observed a sevenfold increase in ibotenic acid- st imulated PI hydrolysis after decortication, a change that clearly exceeds the magni tude of the alterations in m G l u R gene expression and metabot ropic ligand bind- ing which we have observed. These observations pro- vide evidence that supersensitivity does occur in gluta- matergic systems, at least for metabot ropic receptor media ted actions, but that mechanisms in addition to regulation of gene expression are involved, possibly including coupling to second messenger systems. It would be of part icular interest to directly investigate the effects of glutamatergic deafferenta t ion on the efficacy of ionotropic E A A receptor functions, by ex- amining glutamate gated currents or the phosphoryla- tion state of individual receptor subunits [28].
Acknowledgements
The authors would like to thank Z. Holl ingsworth for technical assistance. This study was suppor ted by USPHS grant NS19613 and D F G grant 184/1-1(UW). D.G.S. is a Howard Hughes Medical Institute Physi- cian Research Fellow and a recipient of an Amer ican Academy of Neurology Research Fellowship Award in Neuropharmacology. C.M.T. is a Fellow in the Medical Scientist Training Program of the University of Michi- gan.
References
[1] Abe, T., Sugihara, H., Nawa, H., Shigemoto, R., Mizuno, N. and Nakanishi, S., Molecular characterization of a novel metabotropic glutamate receptor mGluR5 coupled to inositol phosphate/Ca 2+ signal transduction, J. Biol. Chem., 267 (1992) 13361-13368.
[2] Albin, R.L., Makowiec, R.L., Hollingsworth, Z.R., Dure, L.S., Penney, J.B. and Young, A.B., Excitatory amino acid binding sites in the basal ganglia of the rat: a quantitative autoradio- graphic study, Neuroscience, 46 (1992) 35-48.
[3] Altschul, S., Gish, S.F., Miller, W., Myers, E. and Lipman, D., Basic local alignment search tool., Z MoL Biol., 215 (1990) 403-410.
[4] Catania, M.V., Hollingsworth, Z., Penney, J.B. and Young, A.B., Quisqualate resolves two distinct metabotropic [3H]gluta- mate binding sites, NeuroReport, 4 (1993) 311-313.
[5] Creese, I., Burr, D. and Snyder, S.H., Dopamine receptor bind- ing enhancement accompanies lesion-induced behavioural su- persensitivity, Science, 197 (1977) 596-598.
[6] Divac, I., Storm-Mathiesen, J. and Fonnum, F., High affinity uptake of glutamate in terminals of corticostriatal axons, Na- ture, 266 (1977) 377-378.
[7] Errami, M. and Nieoullion, A., Development of a micromethod to study the Na-independent L-[3H]glutamic acid binding to rat striatal membranes. Ii. Effects of selective striatal lesions and deafferentations, Brain Res., 366 (1986) 178-186.
[8] Errami, M. and Nieoullon, A., c~-[3H]amino-3-hydroxy-5-methyl - 4-isoxazolepropionic acid binding to rat striatal membranes: effect of selective brain lesions, Z Neurochem., 51 (1988) 579- 586.
[9] Fonnum, F., Storm-Mathiesen, J. and Divac, I., Biochemical evidence for glutamate as a neurotransmitter in corticostriatal and corticothalamic fibers in rat brain, Neuroscience, 6 (1981) 863-873.
[10] Fotuhi, M., Sharp, A.H., Glatt, C.E., Hwang, P.M., von Krosigk, M., Snyder, S.H. and Dawson, T.M., Differential localization of phosphoinositide-linked metabotropic glutamate receptor (mGluRl) and the inositol 1,4,5-trisphosphate receptor in rat brain, Z Neurosci., 13 (1993) 2001-2012.
[11] Gasic, G.P. and Hollmann, M., The molecular neurobiology of glutamate receptors, Annu. Ret,. Physiol., 54 (1992) 507-536.
[12] Greenamyre, J.T. and Young, A.B., Synaptic localization of striatal NMDA, quisqualate and kainate receptors, Neuroscience Lett., 101 (1989) 133 7.
[13] Heikkila, R.E., Shapiro, B.S. and Duvoisin, R.C., The relation- ship between loss of dopamine nerve terminals, striatal [3H] spiroperidol binding and rotational behaviour in unilaterally 6-hydroxydopamine-lesioned rats, Brain Res., 211 (1981) 285- 292.

U. Wiillner et al. /Brain Research 647 (1994) 209-219 219
[14] lshii, T., Moriyoshi, K., Sugihara, H., Sakurada, K., Kadotani, H., Yokoi, M., Akazawa, C., Shigemoto, R., Mizuno, N., Masu, M. and Nakanishi, S., Molecular characterization of the family of N-methyl-o-aspartate receptor subunits, J. Biol. Chem., 268 (1993) 2836-2843,
[15] Kim, J.S., Hassler, R., Haug, P. and Paik, K.S., Effect of frontal cortex ablation on striatal glutamic acid level in the rat, Brain Res., 132 (1977) 370-374.
[16] Lewis, S.A., Balcarek, J.M., Krek, V., Shelanski, M. and Cowan, N.J., Sequence of a cDNA clone encoding mouse glial fibrillary acidic protein: structural conservation of intermediate filaments, Proc. NatL Acad. Sci. USA, 81 (1984) 2743-2746.
[17] Martin, L.J., Blackstone, C.D., Levey, A.I., Hugnair, R.L. and Price, D.L., Cellular localization of AMPA glutamate receptors within the basal forebrain magnocellular complex of rat and monkey, Z Neurosci., 13 (1993) 2249-2263.
[18] McGeorge, A.J. and Faull, R.L.M., The organization of the projection from the cerebral cortex to the striatum in the rat, Neuroscience, 29 (1989) 503-537.
[19] McLennan, H., The effect of decortication on the excitatory amino acid sensitivity of striatal neurons, Neuroscience Lett., 18 (1980) 313-316.
[20] Monyer, H., Sprengel, R., Schoepfer, R., Herb, A., Higuchi, M., Lomeli, H., Burnashev, N., Sakmann, B. and Seeberg, P., Het- eromeric NMDA receptors: molecular and functional distinc- tion of subtypes, Science, 256 (1992) 1217-1221.
[21] Moriyoshi, K., Masu, M., lshii, T., Shigemoto, R., Mizuno, N. and Nakanishi, S., Molecular cloning and characterization of the rat NMDA receptor, Nature, 354 (1991) 31-37.
[22] Munson, P.J. and Rodbard, D., Ligand: a versatile computer- ized approach for characterization of ligand-binding system, AnaL Biochem., 107 (1980) 220-239.
[23] Nakanishi, S., Molecular diversity of glutamate receptors and implications for brain function, Science, 258 (1992) 597-603.
[24] Nicoletti, F., Wroblewski, J.T., Alho, H., Eva, C., Fadda, E. and Costa, E., Lesions of putative glutamatergic pathways potentiate the increase of inositol phospholipid hydrolysis elicited by exci- tatory amino acids, Brain Res., 436 (1987) 103-112.
[25] Ohishi, H., Shigemoto, R., Nakanishi, S. and Mizuno, N., Distri- bution of the m R N A for a metabotropic glutamate receptor (mGluR3) in the rat brain: an in situ hybridization study, J. Comp. Neurol., 335 (1993) 252-266.
[26] Pellegrini-Giampetro, D.E., Zukin, R.S., Bennett, M.V.L., Cho, S. and Pulsinelli, W.A., Switch in glutamate receptor subunit gene expression in CA1 subfield of hippocampus following global ischemia in rats, Proc. Natl. Acad. Sci. USA, 89 (1992) 10499-10503.
[27] Ponte, P., Ng, S., Engel, J., Gunning, P. and Kedes, L., Evolu- tionary conservation in the untranslated regions of actin mR- NAs: DNA sequence of a human /3-actin cDNA, Nucleic Acids Res., 12 (1984) 1687-1696.
[28] Raymond, L.A., Blackstone, C.D. and Huganir, R.L., Phospho- rylation of amino acid neurotransmitter receptors in synaptic plasticity, Trends Neurosci., 16 (1993) 147-153.
[29] Roberts, P.J., McBean, G.J., Sharif, N.A. and Thomas, E.M., Striatal glutamatergic functions: modifications following specific lesions, Brain Res,, 235 (1982) 83-91.
[30] Salin, P. and Chesselet, M.-F., Paradoxical increase in striatal neuropeptide gene expression following ischemic lesions of the cerebral cortex, Proc. Natl. Acad. Sci. USA, 89 (1992) 9954-9958.
[31] Samuel, D., Errami, M. and Nieoullon, A., Localization of N-methyl-D-aspartate receptors in the rat striatum: effects of specific lesion on the [3H]3-(2-carboxypiperazin-4-yl)propyl-1- phosphonic acid binding, J. Neurochem., 54 (1990) 1926-1933.
[32] Sato, K., Kiyama, H. and Tohyama, M., The differential expres- sion patterns of messenger RNAs encoding non-N-methyi-D- aspartate glutamate receptor subunits in the rat brain, Neuro- science, 52 (1993) 515-539.
[33] Somers, D.L. and Beckstead, R.M., Striatal preprotachykinin and preproenkephalin mRNA levels and the levels of nigral substance P and pallidal met [5]-enkephalin depend on corticos- tiatal axons that use the excitatory amino acid neurotransmitters aspartate and glutamate: quantitative radioimmunocytochemical and in situ hybridization evidence, Mol. Brain Res., 8 (1990) 143-158.
[34] Sommer, B., Kein~inen, K., Verdoorn, T., Wisden, W., Burna- shev, N., Herb, A., K6hler, M., Takagi, T., Sakmann, B. and Seeberg, P., Flip and flop: a cell-specific functional switch in glutamate-operated channels of the CNS, Science, 249 (1990) 1580-1585.
[35] Standaert, D.G., Testa, C.M., Penney, J.B. and Young, A.B., Alternatively spliced isoforms of the NMDAR1 glutamate re- ceptor subunit: differential expression in the basal ganglia of the rat, Neurosci. Lett., 152 (1993) 161-164.
[36] Standaert, D.G., Testa, C.M., Penney, J.B. and Young, A.B., Organization of N-methyl-D-aspartate glutamate receptor gene expression in the basal ganglia of the rat, J. Comp. Neurol., 343 (1994) 1-16.
[37] Sugihara, H., Moriyoshi, K., Ishii, T., Masu, M. and Nakanishi, S., Structures and properties of seven isoforms of the NMDA receptor generated by alternative splicing, Biochem. Biophys. Res. Commun., 185 (1992) 826-32.
[38] Tanabe, Y., Masu, M., Ishii, T., Shigemoto, R. and Nakanishi, S., A family of metabotropic glutamate receptors, Neuron, 8 (1992) 169-179.
[39] Testa, C.M,, Standaert, D.G., Young, A.B. and Penney, J.B., Differential expression of metabotropic glutamate receptor gene mRNA in the basal ganglia of the rat, J. Neurosci., 14 (1994) 3005-3018.
[40] Uhl, G.H., Navia, B. and Douglas, J., Differential expression of preproenkephalin and preprodynorphin mRNAs in striatal neu- rons: high levels of preproenkephalin expression depend on cerebral cortical afferents, ,L Neurosci., 8 (1988) 4755-4764.
[41] Walker, J.E. and Fonnum, F., Effect of regional cortical abla- tions on high-affinity D-aspartate uptake in striatum, olfactory tubercle, and pyriform cortex of the rat, Brain Res., 278 (1983) 283-286.
[42] Wiillner, U., Brouillet, E., lsacson, O., Penney, J.B. and Young, A.B., Glutamate receptor binding sites in MPTP-treated mice, Exp. NeuroL, 121 (1993) 284-287.
[43] Young, A.B., Bromberg, M.B. and Penney, J.B., Decreased glutamate uptake in subcortical areas deafferented by sensori- motor cortical ablation in the cat, J. Neurosci., 1 (1981) 214-249.
[44] Young, A.B. and Fagg, G.E., Excitatory amino acid receptors in the brain: membrane binding and receptor autoradiographic approaches, Trends Pharm. Sci., 11 (1990) 126-133.
[45] Young, W.S., III and Kuhar, M.J., Quantitative in situ hy- bridization and determination of mRNA content. In G.R. Uhl (Ed.), In Situ Hybridization in Brain, Plenum Press, New York, 1986, pp. 243-248.




















