
Piccolo modulation of Synapsin1a dynamics regulates synaptic vesicle exocytosis
Upload
independentCategory
view
0download
0
This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Group I metabotropic glutamate autoreceptors induce abnormal glutamateexocytosis in a mouse model of amyotrophic lateral sclerosis
Francesco Giribaldi a,1, Marco Milanese a,1, Tiziana Bonifacino a, Pia Irene Anna Rossi b,c, Silvia Di Prisco a,Anna Pittaluga a,d, Carlo Tacchetti e, Aldamaria Puliti b,c,d, Cesare Usai f, Giambattista Bonanno a,d,*
aDepartment of Experimental Medicine Pharmacology and Toxicology Unit, University of Genoa, Genoa, ItalybMolecular Genetics and Cytogenetics Unit, Gaslini Institute, Genoa, ItalycDepartment of Pediatric Sciences, University of Genoa, Genoa, ItalydCenter of Excellence for Biomedical Research, University of Genoa, Genoa, ItalyeDepartment of Experimental Medicine, Human Anatomy Unit, University of Genoa, Genoa, Italyf Institute of Biophysics, National Research Council, Genoa, Italy
a r t i c l e i n f o
Article history:Received 11 January 2012Received in revised form26 April 2012Accepted 13 May 2012
Keywords:Amyotrophic lateral sclerosisGlutamate releasePre-synaptic mGlu1 receptorsPre-synaptic mGlu5 receptorsNeurotoxicitySOD1/G93A mice
a b s t r a c t
Glutamate-mediated excitotoxicity plays a major role in ALS and reduced astrocytic glutamate transportwas suggested as a cause. Based on previous workwe have proposed that abnormal release may representanother source of excessive glutamate. In this line, herewe studied themodulation of glutamate release inALS by Group I metabotropic glutamate (mGlu) receptors, that comprise mGlu1 and mGlu5 members.Synaptosomes from the lumbar spinal cord of SOD1/G93A mice, a widely used murine model forhuman ALS, and controls were used in release, confocal or electron microscopy and Western blot experi-ments. Concentrations of themGlu1/5 receptor agonist 3,5-DHPG>0.3 mMstimulated the release of [3H]D-aspartate, used to label the releasing pools of glutamate, both in control and SOD1/G93Amice. At variance,�0.3 mM3,5-DHPG increased [3H]D-aspartate release in SOD1/G93Amice only. Experiments with selectiveantagonists indicated the involvement of both mGlu1 and mGlu5 receptors, mGlu5 being preferentiallyinvolved in the high potencyeffects of 3,5-DHPG.High 3,5-DHPG concentrations increased IP3 formation inbothmouse strains,whereas low3,5-DHPGdid it in SOD1/G93Amice only. Release experiments confirmedthat 3,5-DHPG elicited [3H]D-aspartate exocytosis involving intra-terminal Ca2þ release through IP3-sensitive channels. Confocal microscopy indicated the co-existence of both receptors presynaptically inthe same glutamatergic nerve terminal in SOD1/G93A mice. To conclude, activation of mGlu1/5 receptorsproduced abnormal glutamate release in SOD1/G93A mice, suggesting that these receptors are implicatedin ALS and that selective antagonists may be predicted for new therapeutic approaches.
This article is part of a Special Issue entitled ‘Metabotropic Glutamate Receptors’.� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Glutamate, the principal excitatory neurotransmitter in thecentral nervous system, exerts its actions through activation ofa receptor family that includes ionotropic and metabotropicmembers. Ionotropic receptors are divided into N-methyl-D-aspartate (NMDA), a-amino-3-hydroxy-5-methyl-4-isoxazolepropionate (AMPA) and kainate receptors that exist as multipleheteromers formed by the co-assembly of different subunits(Dingledine et al., 1999). Metabotropic glutamate (mGlu) receptorsare also heterogeneous comprising eight receptors classified intothree groups, based on their sequence homology, signalling andpharmacology (Conn and Pin, 1997; Pin et al., 2003; Nicoletti et al.,2011). Group I, including mGlu1 and mGlu5 receptors, is positivelycoupled to phosphatidylinositol breakdown and intracellular Ca2þ
Abbreviations: ALS, amyotrophic lateral sclerosis; AMPA, a-amino-3-hydroxy-5-methyl-4-isoxazole propionate; 2-APB, 2-aminoethoxydiphenylborane; [3H]D-Asp,[3H]D-Aspartate; BAPTA-AM, 1,2-bis-(2-aminophenoxy)-ethane-N,N,N0 ,N0-tetra-acetic acid, tetraacetoxymethyl ester; CPCCOEt, 7-(hydroxyimino)cyclopropa[b]chrome n-1a-carboxylate ethyl ester; 3,5-DHPG, (S)-3,5-dihydroxyphenylglycine;fenobam, N-(3-chlorophenyl)-N0-(4,5-dihydro-1-methyl-4-oxo-1H-imidazol-2-yl)urea; LY367385, (S)-(þ)-a-amino-4-carboxy-2-methylbenzeneacetic acid; mGluR,metabotropic glutamate receptor; MAP2, microtubule associated proteine 2; MNs,motor neurons; MPEP, 2-Methyl-6-(phenylethynyl)pyridine hydrochloride; NMDA,N-methyl-D-aspartate; SOD1, superoxide dismutase type 1; SOD1/G93A, superoxidedismutase type 1 with a Gly93Ala substitution; U73122, 1-(6-[([17b]-3-methoxyestra-1,3,5[10]-trien-17-yl)amino]hexyl)-1H-pyrrole-2,5-dione; vGluT1,vesicular Glutamate Transporter 1.* Corresponding author. Department of Experimental Medicine, Pharmacology
and Toxicology Unit, University of Genoa, Viale Cembrano 4, 16148 Genoa, Italy.Tel.: þ39 010 3532657; fax: þ39 010 3993360.
E-mail address: [email protected] (G. Bonanno).1 These authors equally contributed to the manuscript.
Contents lists available at SciVerse ScienceDirect
Neuropharmacology
journal homepage: www.elsevier .com/locate/neuropharm
0028-3908/$ e see front matter � 2012 Elsevier Ltd. All rights reserved.doi:10.1016/j.neuropharm.2012.05.018
Neuropharmacology 66 (2013) 253e263

Author's personal copy
mobilization; Group II, including mGlu2 and mGlu3 receptors, andGroup III, including mGlu4 and mGlu6-8 receptors, are negativelycoupled to adenylyl cyclase activity (Conn and Pin, 1997; De Blasiet al., 2001; Ferraguti et al., 2011). There is plentiful evidence thatglutamate receptors exhibit both post- and pre-synaptic localiza-tion and at the pre-synaptic level they generally mediate control ofneurotransmitter release (see, for reviews, MacDermott et al., 1999;Cartmell and Schoepp, 2000; Schoepp, 2001; Raiteri, 2008). Inparticular, Group I mGlu receptors have been proposed to belocated also presynaptically at glutamatergic terminals, even if thenature of the receptor(s) involved in pre-synaptic effect has beenmatter of debate. Indeed, although most of the works favored thepresence of mGlu5 receptors subtypes (Herrero et al., 1992; Sistiagaet al., 1998; Wang and Sihra, 2004), evidence also were providedsupporting the existence of pre-synaptic mGlu1 receptors (Lujanet al., 1996; Moroni et al., 1998; Reid et al., 1999; Fazal et al.,2003) and, more recently, co-localization of low potency ligandbinding mGlu1 and high potency ligand binding mGlu5 autor-eceptors at mouse cortical glutamatergic terminals, which stimu-late glutamate release, has been demonstrated (Musante et al.,2008, 2010).
Group I mGlu receptors have been implicated as contributing toneurodegeneration in a number of neurological disorders (Breysseet al., 2003; Wang and Tseng, 2004; Risterucci et al., 2006; Vernonet al., 2007) including amyotrophic lateral sclerosis (ALS; Rossiet al., 2008; D’Antoni et al., 2011). ALS is a chronic neuromusculardisorder characterized by muscle wasting, weakness and spasticity,reflecting a progressive degeneration of upper and lower motorneurons (MNs) (Brown, 1995). The pathogenesis of ALS is largelyunknown, although vulnerability of MNs in ALS has been ascribedto numerous causes, including protein misfolding, mitochondrialdysfunction, oxidative damage, defective axonal transport,glutamate-mediated excitotoxicity, insufficient growth factorsignaling and inflammation (Cleveland et al., 1996; Morrison andMorrison, 1999; Ferraiuolo et al., 2011). Damage within MNs isenhanced by damage incurred by non-neuronal neighboring cellsaccelerating disease progression (Boillèe et al., 2006; Ilieva et al.,2009; Haidet-Phillips et al., 2011).
In spite of the depicted etiological heterogeneity of the disease,glutamate-mediated excitotoxicity, based on the high levels ofextracellular glutamate detected in an elevated percentage ofsporadic and familiar ALS patients (Perry et al., 1990), has beensuggested to play a major role. Impaired glutamate transport,founded on reduced expression of glial glutamate transporter oftype 1, has been proposed as the reason (Rothstein et al., 1995).However, due to the complex interplay of multiple mechanisms,glutamate transport defects may not be the only origin of excito-toxicity and other causes should be considered, including increaseof glutamate release. Indeed, utilizing a mouse model of humanALS expressing high copy number of mutant human superoxidedismutase type 1 (SOD1) with a Gly93Ala substitution (SOD1/G93A; Gurney et al., 1994), we have previously detected abnormalrelease of glutamate induced by GABA and glycine transportersheterologously expressed at spinal cord glutamatergic nerveterminals and astrocytes (Raiteri et al., 2003, 2004; Milanese et al.,2010) or by stimuli leading to glutamate exocytosis, such as highKCl, ionomycin or hypertonic sucrose (Milanese et al., 2011). In thepresent work we investigated on the modulation of glutamaterelease provoked by activation of Group I mGlu receptors in thespinal cord of either control and SOD1/G93A mice and found thatboth mGlu1 and mGlu5 receptor activation stimulated the basalrelease of the excitatory amino acid. Interestingly, this receptor-mediated effect was abnormal in SOD1/G93A mice, being detect-able at much lower concentrations of agonists than in controlmice.
2. Materials and methods
2.1. Animals
Adult B6SJL-TgN SOD1/G93A(þ)1Gur mice expressing high copy number ofmutant human SOD1 with a Gly93Ala substitution [SOD1/G93A] and B6SJL-TgN(SOD1)2Gur mice expressing wild-type human SOD1 [SOD1] (Gurney et al., 1994)were originally obtained from Jackson Laboratories (Bar Harbor, ME, USA) and bredat the animal facility of the Pharmacology and Toxicology Unit of the Department ofExperimental Medicine in Genoa, where they were kept until experiments werecarried out. Transgenic animals have been crossed with background-matched B6SJLwild-type female and selective breeding maintained each transgene in the hemi-zygous state. Non-transgenic littermate mice were used as controls [WT]. Femaleand male mice were utilized; sexes were equally distributed in each series ofexperiments. All transgenic mice were identified analyzing tissue extracts from tailtips. In SOD1/G93A mice death usually occurs between 120 and 140 days. Forexperimental use animal were killed at a late stage of disease, scored according tomotor impairment severity. Animals were housed at constant temperature(22 � 1 �C) and relative humidity (50%) with a regular 12 he12 h light cycle (light 7AMe7 PM), throughout the experiments. Food and water were freely available. Allexperiments were carried out in accordance with the European CommunitiesCouncil Directive of 24 November 1986 (86/609/EEC). All efforts were made tominimize animal suffering and to use only the number of animals necessary toproduce reliable results. A total number of 60 SOD1/G93A, 43 SOD1 and 20 WTmice were used in this study.
2.2. Synaptosomes purification
Animals were sacrificed and the spinal cord was rapidly removed and main-tained at 4 �C; only the thoracic-lumbar (T13-L6) region was used for experimentalprocedures. Synaptosomes were prepared essentially as previously described(Stigliani et al., 2006). The tissue was homogenized in 10 volumes (1 g in 10 mL) of0.32 M sucrose, buffered at pH 7.4 with TriseHCl, using a glass-teflon tissue grinder(clearance 0.25 mm). The homogenate was centrifuged (5 min, 1000 � g at 4 �C) toremove nuclei and debris and the supernatant was gently stratified on a discontin-uous Percoll� gradient (2, 6, 10 and 20% v/v in Tris-buffered sucrose). After centri-fugation at 33,500 � g for 5 min, the layer between 10 and 20% Percoll�
(synaptosomal fraction) was collected, washed by centrifugation at 20,000 � g for15 min and then resuspended in physiological medium having the followingcomposition (mM): NaCl, 140; KCl, 3; MgSO4 1.2; NaH2PO4, 1.2; NaHCO3 5; CaCl2 1.2;HEPES 10; glucose, 10; pH 7.4. Proteins were measured according to Bradford (1976)using bovine serum albumin as a standard.
2.3. Release experiments
Synaptosomes were incubated at 37 �C for 15 min in the presence of 0.05 mM[3H]D-Aspartate ([3H]D-Asp; a non-metabolizable analogue of glutamate used tolabel the synaptosomal glutamate-releasing pools). Aliquots of synaptosomalsuspension (5e10 mg protein) were equally distributed onmicroporous filters placedat the bottom of a set of parallel superfusion chambers maintained at 37 �C(Superfusion System, Ugo Basile, Comerio, Varese, Italy; Raiteri et al., 1984). Super-fusionwas then started with standardmedium at a rate of 0.5 ml/min and continuedfor 48 min. After 36 min of superfusion to equilibrate the system, five 3-min frac-tions were collected. (S)-3,5-Dihydroxyphenylglycine (3,5-DHPG; 0.001e30 mM)was introduced at the end of the first sample collected (t ¼ 39 min) and main-tained until the end of the experiment; all other drugs were introduced att ¼ 30 min. When appropriate, the Ca2þ-free mediumwas introduced at t ¼ 20 min.In some experiments, synaptosomes were incubated 30 min (15 min before andduring [3H]D-Asp labelling) in the presence of 100 mM 1,2-bis-(2-aminophenoxy)-ethane-N,N,N0 ,N0-tetraacetic acid, tetraacetoxymethyl ester (BAPTA-AM) or with0.1 mM bafilomycin A1. Collected samples and superfused synaptosomes werecounted for radioactivity. Tritium released in each sample was calculated as frac-tional rate � 100 (percentage of the total synaptosomal neurotransmitter content atthe beginning of the respective collection period). Drug effects were evaluated bycalculating the ratio between the efflux in the fourth sample collected (in which themaximum effect of 3,5-DHPG was generally reached) and the efflux of the firstfraction (basal efflux). This ratio was compared to the corresponding ratio obtainedunder resting conditions. Appropriate controls were always run in parallel.
2.4. Cytosolic IP3 quantification
Suspensions of spinal cord synaptosomes (50 mg protein) were equilibrated at37 �C for 15 min. Each sample was exposed to 0.3 or 30 mM 3,5-DHPG for 9 min. Thereaction was stopped by cooling in ice water and sonication and the cytosolicfraction was separated from membranes by centrifugation (16,000 � g, 3 min).Cytosolic IP3 content was determined by using a commercially available radio-receptor assay kit (Inositol-1,4,5-trisphosphate [3H] radioreceptor assay kit, PerkinElmer Life Sciences, Boston, MA, USA). The detection limit of the assay was 0.1 pmol(standard curve range 0.19e25 pmol).
F. Giribaldi et al. / Neuropharmacology 66 (2013) 253e263254
Author's personal copy
2.5. Confocal microscopy
Purified synaptosomes (40 mg protein) were stratified onto coverslips pre-treated with poly-L-lysine and maintained 45 min at room temperature to allowsetting and sticking to the surface. The preparations were fixed with 2% para-formaldehyde (15 min), washed with PBS (3 � 5 min) and permeabilized (5 min)with 0.05% triton X-100. After washing (3 � 5 min) with PBS containing 0.5% serumalbumin, the preparations were overnight incubated at 4 �C with the primaryantibodies diluted in PBS containing 3% albumin. The following antibodies wereused: guinea pig anti-vesicular Glutamate Transporter 1 (vGluT1, 1:500, Millipore,Billerica, MA, USA) mouse anti-mGlu1 receptors, (1:250, BD Biosciences, NJ, USA)and rabbit anti-mGlu5 receptors, (1:250, Abcam, Cambridge, UK), chicken anti-Microtubule associated protein 2 (MAP2, 1:1000, Millipore, Billerica, MA, USA),mouse anti-PSD95 (1:300, SigmaeAldrich, St Louis, MO, USA) and rabbit anti-PSD95receptors, (1:300, Novus Biologicals, Littleton, CO, USA). After washing (3 � 5 min)with PBS containing 0.5% serum albumin, the preparations were incubated 45 minwith the appropriate secondary goat anti-guinea pig Alexa Fluor A488-conjugated,donkey anti-mouse AlexaFluor A647-conjugated, goat anti-rabbit AlexaFluor A555-conjugated, donkey anti-rabbit AlexaFluor A488-conjugated and donkey anti-chicken AlexaFluor A488-conjugated (Molecular Probes Europe, Leiden, TheNetherlands) antibodies, diluted 1:2000 in PBS containing 3% albumin. Fluorescenceimage (512 � 512 � 8 bit) acquisition was performed by a three-channel Leica TCSSP5 laser-scanning confocal microscope, equipped with 458, 476, 488, 514, 543 and633 nm excitation lines, through a plan-apochromatic oil immersion objective 63�/1.4. Light collection configuration was optimized according to the combination ofchosen fluorochromes and sequential channel acquisition was performed to avoidcross-talk. Leica “LAS AF” software package was used for image acquisition. Theestimation of co-localized proteins was performed calculating the “co-localizationcoefficients” (Manders et al., 1992).
2.6. Electron microscopy
For ultrastructural analysis, purified lumbar spinal cord synaptosomes werefixed in 2.5% glutaraldehyde in cacodylate buffer 0.1 M, pH 7.2, postfixed in 1%osmium tetroxide in cacodylate buffer 0.1 M, pH 7.2, en bloc stained with a 1%aqueous solution of uranyl acetate, dehydrated through a graded ethanol series.Samples were then embedded in LX112 (Polysciences Inc., Warrington, PA, USA),polymerized for 12 h at 42 �C, followed by 48 h at 60 �C. Grayesilver ultrathinsections, obtained using a Leica Ultracut E microtome, were stained with uranylacetate and lead citrate, and analyzed with a FEI CM10 electron microscopy. Toquantify the number of post-synaptic densities, 10 electron micrographs were ob-tained at 8,900� from ultrathin sections of each sample. Micrographs were selectedin order to include the maximum number of structures.
2.7. Immunoblotting
Spinal cord synaptosomal membranes were obtained (Barbiero et al., 2007) andused in Western blot analyses. Proteins were separated on 7% gel by means ofSDSepolyacrylamide gel electrophoresis. The concentration of proteins in eachsample fell on the linear portion of the curve. Electroblotted proteins were moni-tored using Naphthol blue black staining (SigmaeAldrich, MO, USA). Membraneswere then incubated with the following antibodies: mouse monoclonal anti-mGlu1receptor (1:2500; BD Biosciences, San Jose, CA, USA); rabbit monoclonal anti-mGlu5receptor (1:10000, Epitomics, CA, USA); mouse monoclonal Anti-b-tubulin III(1:2000; SigmaeAldrich, MO, USA); rabbit monoclonal anti-PSD95 (diluted 1:500,Novus Biologicals, Littleton, Colorado, USA) and mouse monoclonal anti-sintaxin(1:3000, Chemicon, Temecula, CA, USA). After incubation with peroxidase-coupledsecondary antibodies, protein bands were detected by using a Western blottingdetection system (ECL Advance�; Amersham Biosciences, Piscataway, NJ, USA).Bands were detected and analyzed for density using an enhanced chem-iluminescence system (Versa-Doc 4000 and Quantity One software; Bio-Rad,Hercules, CA, USA). All of the protein bands used were normalized for b-tubulin IIIlevel in the same membrane.
2.8. Statistics
Data are expressed as means � SEM throughout. Two-tailed Student’s t test wasused for the analysis of two populations of means. Comparison of more than twomean populations was performed using two-way ANOVA followed byNewmaneKeuls, Bonferroni or Dunnett’s test where appropriate.
2.9. Materials
[3H]D-Aspartate ([3H]D-Asp, specific activity: 11.3 Ci/mmol) were purchasedfrom Perkin Elmer Life Sciences, Boston, MA, USA; Percoll, Bafilomycin A1 andBAPTA-AM and 1-(6-[([17b]-3-methoxyestra-1,3,5[10]-trien-17-yl)amino]hexyl)-1H-pyrrole-2,5-dione (U73122) were obtained from Sigma Chemical Co. (St. Louis,MO, USA); 3,5-DHPG, 7-(hydroxyimino)cyclopropa[b]chrome n-1a-carboxylateethyl ester (CPCCOEt), 2-Methyl-6-(phenylethynyl)pyridine hydrochloride (MPEP),
(S)-(þ)-a-Amino-4-carboxy-2-methylbenzeneacetic acid (LY367385) and N-(3-chlorophenyl)-N0-(4,5-dihydro-1-methyl-4-oxo-1H-imidazol-2-yl)urea (fenobam)and 2-aminoethoxydiphenylborane (2-APB) was obtained from Tocris Bioscience(Bristol, UK). All the other reagents were of laboratory grade.
3. Results
3.1. Effects of 3,5-DHPG on [3H]D-Asp release
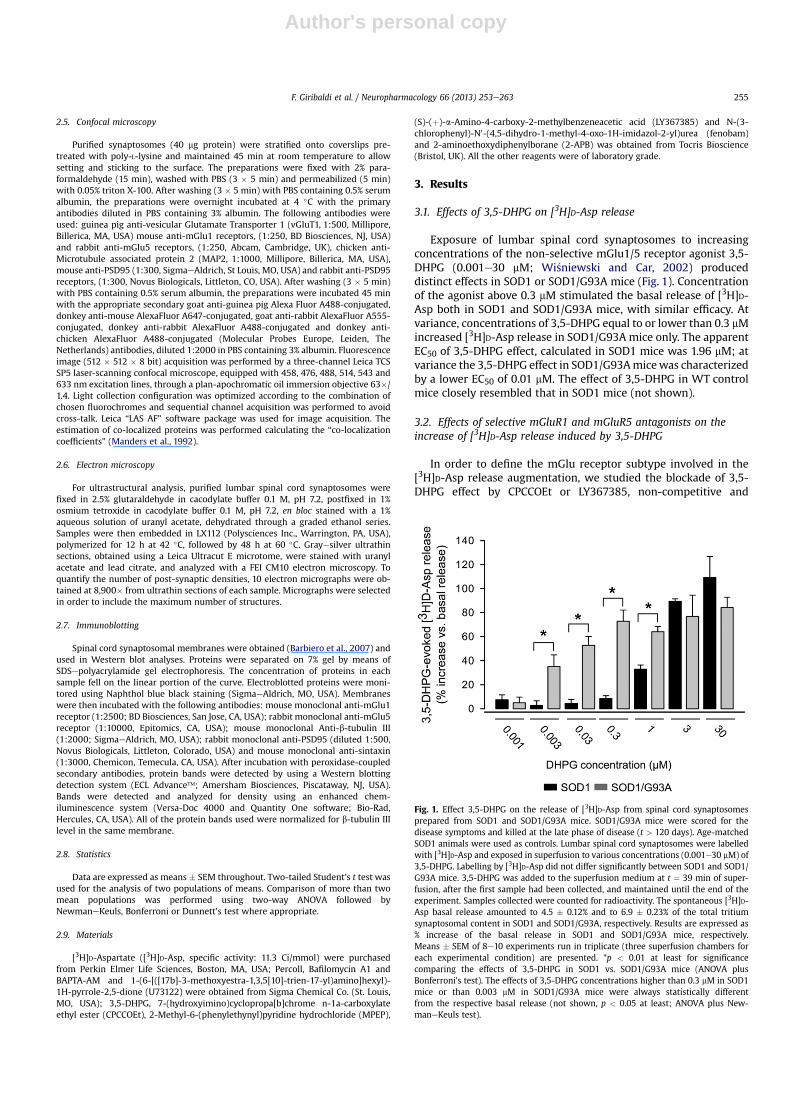
Exposure of lumbar spinal cord synaptosomes to increasingconcentrations of the non-selective mGlu1/5 receptor agonist 3,5-DHPG (0.001e30 mM; Wi�sniewski and Car, 2002) produceddistinct effects in SOD1 or SOD1/G93A mice (Fig. 1). Concentrationof the agonist above 0.3 mM stimulated the basal release of [3H]D-Asp both in SOD1 and SOD1/G93A mice, with similar efficacy. Atvariance, concentrations of 3,5-DHPG equal to or lower than 0.3 mMincreased [3H]D-Asp release in SOD1/G93A mice only. The apparentEC50 of 3,5-DHPG effect, calculated in SOD1 mice was 1.96 mM; atvariance the 3,5-DHPG effect in SOD1/G93Amicewas characterizedby a lower EC50 of 0.01 mM. The effect of 3,5-DHPG in WT controlmice closely resembled that in SOD1 mice (not shown).
3.2. Effects of selective mGluR1 and mGluR5 antagonists on theincrease of [3H]D-Asp release induced by 3,5-DHPG
In order to define the mGlu receptor subtype involved in the[3H]D-Asp release augmentation, we studied the blockade of 3,5-DHPG effect by CPCCOEt or LY367385, non-competitive and
Fig. 1. Effect 3,5-DHPG on the release of [3H]D-Asp from spinal cord synaptosomesprepared from SOD1 and SOD1/G93A mice. SOD1/G93A mice were scored for thedisease symptoms and killed at the late phase of disease (t > 120 days). Age-matchedSOD1 animals were used as controls. Lumbar spinal cord synaptosomes were labelledwith [3H]D-Asp and exposed in superfusion to various concentrations (0.001e30 mM) of3,5-DHPG. Labelling by [3H]D-Asp did not differ significantly between SOD1 and SOD1/G93A mice. 3,5-DHPG was added to the superfusion medium at t ¼ 39 min of super-fusion, after the first sample had been collected, and maintained until the end of theexperiment. Samples collected were counted for radioactivity. The spontaneous [3H]D-Asp basal release amounted to 4.5 � 0.12% and to 6.9 � 0.23% of the total tritiumsynaptosomal content in SOD1 and SOD1/G93A, respectively. Results are expressed as% increase of the basal release in SOD1 and SOD1/G93A mice, respectively.Means � SEM of 8e10 experiments run in triplicate (three superfusion chambers foreach experimental condition) are presented. *p < 0.01 at least for significancecomparing the effects of 3,5-DHPG in SOD1 vs. SOD1/G93A mice (ANOVA plusBonferroni’s test). The effects of 3,5-DHPG concentrations higher than 0.3 mM in SOD1mice or than 0.003 mM in SOD1/G93A mice were always statistically differentfrom the respective basal release (not shown, p < 0.05 at least; ANOVA plus New-maneKeuls test).
F. Giribaldi et al. / Neuropharmacology 66 (2013) 253e263 255

Author's personal copy
competitive mGlu1 receptor antagonist, respectively (Bruno et al.,1999; Litschig et al., 1999), by the non-competitive mGlu5receptor antagonist MPEP (Gasparini et al., 1999) and by the non-competitive antagonist/negative allosteric modulator of the mGlu5receptor fenobam (Porter et al., 2005).
The 30 mM 3,5-DHPG-induced increase of [3H]D-Asp release inSOD1 mice was partly antagonized by 1 mM MPEP, 1 mM LY367385or 5 mM CPCCOEt while it was almost completely abated by 10 mMMPEP or 10 mM LY367385 (Fig. 2A). The concomitant presence ofboth 1 mM MPEP and 5 mM CPCCOEt or 1 mM MPEP and 1 mMLY367385 did not further affect the 3,5-DHPG-mediated increase of[3H]D-Asp release. Fig. 2A also shows that 1 mM MPEP, 1 mMLY367385 or 5 mM CPCCOEt only partially antagonized 30 mM 3,5-DHPG in SOD1/G93A mice and that the antagonists did not proveadditive effects when added together. Again, 10 mMMPEP or 10 mMLY367385 completely antagonized DHPG.
As shown inFig. 2B, also theeffectof 0.3mM3,5-DHPG, thehighestconcentration which stimulated [3H]D-Asp release selectively inSOD/G93A mice, was largely or completely prevented by 1 or 10 mM
MPEP or by 1 or 10 mM LY367385 respectively. Once more, theconcomitant application of the two antagonists, at the concentrationof 1 mM, did not show additive effect in dampening the 0.3 mM 3,5-DHPG-induced stimulation of the [3H]D-Asp basal release. Notably,the effects of 0.03 mM 3,5-DHPG was completely counteracted by1 mM MPEP (Fig. 2A) or 0.3 mM fenobam (Table 1), whereas 1 mMLY367385 was ineffective. Table 1 also shows that 0.3 mM fenobampowerfully counteracted also 0.3 mM or 30 mM 3,5-DHPG in SOD1/G93A mice. The antagonists utilized did not significantly modify bythemselves the spontaneous release of [3H]D-Asp (not shown).
3.3. Inositol-3-phosphate and internal Ca2þ involvement in themodulation of [3H]D-Asp release induced by 3,5-DHPG
Since Group I mGluRs are positively linked to phosphatidyli-nositol breakdown and IP3 production, we measured the intra-synaptosomal levels of IP3 after exposure to 3,5-DHPG. Data ob-tainedwere in linewith the above release experiments. In fact, high3,5-DHPG (30 mM) induced IP3 formation in both mouse strains,whereas 0.3 mM 3,5-DHPG induced a significant increase of thebasal content of IP3 in synaptosomes purified from the spinal cordof SOD1/G93A mice but not from SOD1 mice (Fig. 3A).
In order to verify the impact of the above IP3 changes on themodulation of [3H]D-Asp release, we exposed mouse lumbar spinalcord synaptosomes to 0.3 or 30 mM 3,5-DHPG in the absence ofexternal Ca2þ, in the presence of external Ca2þ plus cytoplasmicBAPTA, in the presence of the phospholipase C inhibitor U73122(Bleasdale et al., 1990) or in the presence of the IP3 receptor blocker2-APB (Maruyama et al., 1997). Omission of Ca2þ in the superfusionmedium did not significantly reduce the increase of [3H]D-Asprelease brought about by 0.3 mM 3,5-DHPG in SOD1/G93A mice; onthe contrary, a significant reduction was obtained in synaptosomesincubated in the presence of 100 mM BAPTA-AM prior releaseexperiments or exposed in superfusion to 1 mMU73122 or 10 mM2-APB (Fig. 3B). Fig. 3B also shows that the 0.3 mM 3,5-DHPG-induced[3H]D-Asp release was likely of vesicular origin, being completelyoccluded by incubation of synaptosomes with 0.1 mM bafilomycinA1, that depletes the vesicular neurotransmitter content, priorrelease experiments. Also the effect of 30 mM 3,5-DHPG wasU73122, 2-APB or bafilomycin A1 sensitive, both in SOD1 and SOD1/G93A mice (Fig. 3C).
3.4. Expression of mGlu1 and mGlu5 receptors in spinal cordsynaptosomes
Confocal microscopy experiments were performed by concom-itantly labelling lumbar spinal cord synaptosomes from SOD1 andSOD1/G93A mice with a vGluT1 antibody (blue), to highlight glu-tamatergic nerve terminals, as well as with antibodies for mGlu1(red (in the web version)) and mGlu5 (green) receptors (Fig. 4A).Synaptosomal preparations efficiently stained for all the antibodiestested and co-localization of vGluT1 and mGlu1 receptors, vGluT1and mGlu5 receptors and mGlu1 and mGlu5 receptors was
Fig. 2. Effects of mGluR1 and mGluR5 receptor antagonists on the release of [3H]D-Aspevoked by 30 mM 3,5-DHPG from SOD1 or SOD1/G93A mouse spinal cord synapto-somes (A) or by 0.3 mM 3,5-DHPG from SOD1/G93A mouse spinal cord synaptosomes(B). SOD1/G93A mice were scored for the disease symptoms and killed at the latephase of disease. Age-matched SOD1 animals were used as controls. Lumbar spinalcord synaptosomes were labelled with [3H]D-Asp and exposed in superfusion to 3,5-DHPG from t ¼ 39 min of superfusion. CPCCOEt, MPEP and LY367385 were intro-duced 9 min before 3,5-DHPG. Results are expressed as % increase of the basal release.Means � SEM of 10e12 experiments run in triplicate are presented. *p < 0.01 at leastvs. the respective effect of 3,5-DHPG alone in SOD1 or SOD1/G93A mice, as appropriate(ANOVA plus Dunnett’s test).
Table 1Effects of fenobam on the release of [3H]D-Asp evoked by 3,5-DHPG from SOD1/G93A mouse spinal cord synaptosomes.
Drugs % increase n
0.03 mM 3,5-DHPG 47.39 � 7.07 50.03 mM 3,5-DHPG þ 0.3 mM fenobam 6.60 � 0.51a 50.3 mM 3,5-DHPG 81.56 � 7.98 40.3 mM 3,5-DHPG þ 0.3 mM fenobam 10.45 � 0.88a 430 mM 3,5-DHPG 86.50 � 9.81a 330 mM 3,5-DHPG þ 0.3 mM fenobam 22.45 � 1.79 3
a p < 0.001 at least vs. the respective effect of 3,5-DHPG alone (Two-tailedStudent’s t test).
F. Giribaldi et al. / Neuropharmacology 66 (2013) 253e263256

Author's personal copy
analyzed. Merging of the appropriate image pairs revealed thata large proportion of Glu-storing terminals expressed mGlu1 andmGlu5 receptors. In particular, 61 � 3.4% and 66.1 � 2.2% of thevGluT1-positive synaptosomes from SOD1 mice were also positivefor mGlu1 and mGlu5 receptors, respectively. Co-existence valuesobtained in SOD1/G93A mice were very similar to those reportedabove for SOD1mice (Fig. 4A). Moreover, in SOD1mice, 47� 2.0% ofmGlu1 receptor-expressing synaptosomes also expressed mGlu5receptors and 65 � 2.4% of mGlu5 receptor-expressing synapto-somes were mGlu1 receptor-positive. Again, no relevant differ-ences could be appreciated between SOD1 and SOD1/G93A mice(Fig. 4A).
We also performed confocal microscopy experiments aimed toanalyze the co-existence of mGlu1 and mGlu5 immunoreactivitywith the post-synaptic density marker PSD95, in order to excludethat the two receptors were localized at the post-synaptic densitiesattached to purified synaptosomes. Fig. 4B shows that only13.7 � 3.1% mGlu1 or 13.0 � 1.2% mGlu5 receptor-positive punctawere also positive for PSD95 in SOD1/G93A mouse preparations.Similar results were obtained in SOD1 mice (Fig. 4B).
The images clearly show that the scarce co-localization of mGlureceptors with PSD95 mainly is due to the minimal PSD95 immu-noreactivity present in spinal cord synaptosomal preparations. Tofurther investigate on this aspect, we executed experiments bydouble-labelling spinal cord synaptosomes using a specific anti-body for the synaptosomal marker MAP2, to stain the wholesynaptosomal population, and for PSD95. Synaptosomes wereextensively labelled by the MAP2 antibody used and, as illustratedin Fig. 5A, quantification of MAP2/PSD95 co-localization indicatedthat only 12.2 � 2.3% and 10.9 � 2.8% of the MAP2-positivesynaptosomal particles were also PSD95-positive in SOD1/G93Aand SOD1 mice, respectively. As a control, we found that the PSD95antibody used in the above experiments extensively labelledmousecerebral cortex synaptosomes prepared from SOD1/G93Amice (norshown), a results that we have previously obtained also using ratcerebral cortex synaptosomes (Stigliani et al., 2006). Accordingly,Western blot experiments indicated that, differently from cerebralcortex, PDS95 cannot be revealed in synaptosomal spinal cordmembranes (Fig. 5B). To confirm the above observations, wedirectly counted the number of post-synaptic density-endowedspinal cord synaptosomes by electron microscopy. The results ob-tained indicated that attached post-synaptic elements werepresent in 5 over 56 synaptosomes in SOD1 mice and in 5 over 52synaptosomes in SOD1/G93A mice (Fig. 5C).
Lastly in order to quantify changes in the expression of mGlu1and mGlu5 receptors in SOD1/G93A respect to SOD1 and WT mice,we carried out Western blot experiments using protein extractsfrom membranes obtained from lumbar spinal cord synaptosomes.The mGlu1 (Fig. 6A) and mGlu5 (Fig. 6B) receptors expression wasnotmodified inWTand SOD1mice. On the contrary, the expressionlevels of mGlu5 receptors were significantly higher in SOD1/G93Amice, compared to the expression in control animals. mGlu1receptors were only slightly, although not significantly, augmented.
4. Discussion
The first finding of the present investigation is that gluta-matergic nerve endings isolated from the thoracic-lumbar region ofthe spinal cord of the adult mouse posses pre-synaptic mGlureceptors belonging to the mGlu1 or mGlu5 subtype, whose
Fig. 3. Effect of 3,5-DHPG on IP3 synaptosomal content (A) and effects of Ca2þ
deprivation, U73122, 2-APB or bafilomycin on the 0.3 mM (B) or 30 mM (C) 3,5-DHPG-evoked [3H]D-Asp release in spinal cord synaptosomes from SOD1 or SOD1/G93A mice.SOD1/G93A mice were scored for the disease symptoms and killed at the late phase ofdisease. Age-matched SOD1 animals were used as controls. In IP3 measurement,lumbar spinal cord synaptosomes were equilibrated at 37 �C for 15 min and exposed to3,5-DHPG for 9 min. IP3 was determined by using a commercially available radio-receptor assay. Results are expressed as pmol IP3 per mg of synaptosomal protein.Means � SEM of 4 experiments run in triplicate are presented. *p < 0.05 vs. the IP3basal content measured in SOD1 or SOD1/G93A mice, as appropriate; #p < 0.05 vs. the0.3 mM 3,5-DHPG-evoked IP3 content measured in SOD1 mice (ANOVA plus New-maneKeuls test). In release experiments, lumbar spinal cord synaptosomes werelabelled with [3H]D-Asp and exposed in superfusion to 3,5-DHPG from t ¼ 39 min ofsuperfusion. 1 mM U73122 or 10 mM 2-APB was introduced from t ¼ 30 min; Ca2þ wasomitted from t ¼ 20 min; synaptosomes were incubated 30 min at 37 �C in thepresence of 100 mM BAPTA-AM or 0.1 mM bafilomycin A1 for 30 min before releaseexperiments. The presence of BAPTA-AM or bafilomycin A1 did not affect significantly
synaptosomal labelling by [3H]D-Asp. Results are expressed as % increase of the basalrelease. Means � SEM of 8e10 independent experiments run in triplicate are pre-sented. *p < 0.001 at least vs. the effect of 3,5-DHPG alone in SOD1 or SOD1/G93Amice, as appropriate (ANOVA plus Dunnett’s test).
F. Giribaldi et al. / Neuropharmacology 66 (2013) 253e263 257

Author's personal copy
Fig. 4. Identification by immunocytochemistry of vGluT1, of mGlu1, and mGlu5 receptors, and of PSD95 in spinal cord synaptosomes from SOD1 and SOD1/G93A mice. (A)Expression and co-existence of vGluT1, mGlu1 and mGlu5 receptors; (B) Expression and co-existence of PSD95, mGlu1 and mGlu5 receptors. SOD1/G93A mice were killed at the latephase of disease and age-matched SOD1 animals were used. Synaptosomes were layered onto coverslips, fixed with paraformaldehyde, permeabilized with triton X-100 andincubated with the proper primary and secondary antibodies. Samples were analysed by laser confocal microscopy. Images show immune-positivity for Alexa Fluor 488-taggedanti-vGluT1, Alexa Fluor 647-tagged anti-mGlu1 receptors, for Alexa Fluor 555-tagged anti-mGlu5 receptors and for Alexa Fluor 647-tagged anti-PSD95. Merge represents theco-expression (A) of vGluT1, mGluR1 and mGluR5 and (B) of mGlu1 or mGlu5 receptor with PDS95. Bar plots indicate the quantification of the above protein couple co-localization.Images are representative of 3 separate experiments in triplicate. Scale bar ¼ 100 mM.
F. Giribaldi et al. / Neuropharmacology 66 (2013) 253e263258

Author's personal copy
activation elicited the release of glutamate. The second finding isthat low concentrations (�0.3 mM) of the mixed mGlu1/5 receptoragonist 3,5-DHPG potentiated the basal release of glutamate inSOD1/G93A mice while only high (�1 mM) 3,5-DHPG stimulatedglutamate release also in WT and SOD1 mice. The activity of mGlu1and mGlu5 selective receptor antagonists evidenced that the highpotency effect of 3,5-DHPG was mostly supported by mGlu5receptors, whereas both receptors contributed to the low potency3,5-DHPG effect.
Here, neurotransmitter release has been monitored utilizingpurified synaptosomes in superfusion, according to a technique setup in our laboratory and considered the method of choice for thistype of studies (Raiteri et al., 1984). According to this technique,
synaptosomes are layered on large microporous filters in a smallamount that constitutes less than a monolayer and up-downsuperfused. By this way all the released endogenous transmitters/modulators are immediately removed by the superfusion mediumbefore they can be taken up by transporters or they can interactwith receptors or other molecules expressed at the pre-synapticlevel. The absence of a biophase surrounding synaptosomes isdemonstrated by the observation that uptake inhibitors or receptorantagonists added to the superfusion medium do not modify bythemselves the release of neurotransmitters. Therefore, duringsuperfusion, all the pre-synaptic targets (transporters, receptors,ion channels, enzymes, etc.) can be considered virtually free ofendogenous ligands and can be studied separately by adding the
Fig. 5. Characterization post-synaptic membrane presence in spinal cord synaptosomes by confocal microscopy, Western blot and electron microscopy. (A) Identification byimmunocytochemistry of MAP2 and PSD95 in spinal cord synaptosomes from SOD1 and SOD1/G93A mice. SOD1/G93A mice were killed at the late phase of disease and age-matched SOD1 animals were used. Synaptosomes were layered onto coverslips, fixed with paraformaldehyde, permeabilized with triton X-100 and incubated with the properprimary and secondary antibodies. Samples were analysed by laser confocal microscopy. Images show immune-positivity for Alexa Fluor 488-tagged anti-MAP2 and Alexa Fluor647-tagged anti-PSD95. Merge represents co-expression of MAP2 with PDS95. Bar plots indicate the quantification of protein couple co-localization. Images are representative of 3separate experiments in triplicate. Scale bar ¼ 100 mM. (B) Quantification of PSD95, syntaxin, b-tubulin III content in synaptosomes obtained from spinal cord or cerebral cortex ofSOD1 mice. Representative immunoreactive bands for PSD95, syntaxin and b-tubulin III were obtained by SDS-10% PAGE and Western blot onto nitrocellulose membranes, using20 mg (lane 1) or 40 mg (lane 2) of detergent soluble membrane proteins of synaptosomes purified from the lumbar spinal cord. (C) Electron micrograph of lumbar spinal cordsynaptosomes (arrows) prepared from SOD1/G93A mice. Purified synaptosomes were fixed, dehydrated and embedded in LX112. Ultrathin sections were stained with uranyl acetateand lead citrate, and analyzed with a FEI CM10 electron microscopy. Bar plots indicate the number of post-synaptic densities over the total number of counted synaptosomes inSOD1 and SOD1/G93A mice. Scale bar ¼ 780 nm.
F. Giribaldi et al. / Neuropharmacology 66 (2013) 253e263 259

Author's personal copy
appropriate ligands to the superfusion medium at the desiredconcentration. Under these experimental conditions, drug-inducedchanges of the release of one neurotransmitter reasonably can beattributed to direct actions at the terminals storing and releasingthe neurotransmitter at issue. As a consequence, we can argue thatthe group I mGlu receptor-mediated modifications of glutamaterelease here measured are due to activation/blockade of thesereceptor subtypes, confined presynaptically to glutamate-releasingnerve terminals of spinal cord. In line with this hypothesis, confocalmicroscopy experiments unveiled that mGlu1 andmGlu5 receptorswere markedly expressed in preparations of purified synaptosomesand co-localized at glutamatergic particles. Concomitantly, confocalmicroscopy demonstrated a scant co-existence of mGlu1 andmGlu5 receptors with the post-synaptic marker PSD95, both inSOD1 and SOD1/G93A mice. This event based on the apparent verylow immunoreactivity for PSD95 in spinal cord synaptosomalpreparation; an occurrence also confirmed by electron microscopyexperiments, that hardly could reveal post-synaptic densitiesattached to spinal cord synaptosomal particles. Taken together, all
the experimental evidence presented strongly indicate that the twoGroup I mGlu receptors, that modulate glutamate release, arepresent presynaptically at the glutamatergic nerve terminals. Asa technical note, the rareness of synaptosome-attached post-synaptic densities makes purified spinal cord synaptosomes anunique preparation to investigate on the pre-synaptic localizationand function of a given target.
Group 1 mGlu receptors are expressed in astrocytes and mGlu5receptors expression is higher in astrocytes of SOD1/G93Amice andin human individuals with ALS (Aronica et al., 2001; Rossi et al.,2008). Thus, in order to exclude that increased release of gluta-mate could be due to stimulation of receptors expressed at higherlevels in SOD1/G93A mouse astrocytes rather than in pre-synapticterminals, the pureness of the synaptosomal preparation fromSOD1 and SOD1/G93A mice, with respect to astroglial contamina-tion, turns out to be a relevant point. We have already verified thepurity of synaptosomes isolated by homogenization and separationon Percoll� gradient from rat cerebral cortex (Stigliani et al., 2006),hippocampus and cerebellum (Pedrazzi et al., 2006) tissues.Moreover, we already assessed in a previous recent paper thepureness of mouse spinal cord synaptosomes prepared in the verysame way of the present experiments (Milanese et al., 2010). Thedata collected by means of confocal microscopy and Western blotanalyses showed that the Percoll�-purified spinal cord synapto-somes are devoid of significant astrocyte, microglia or oligoden-drocyte contamination. Furthermore, the degree of synaptosomespurification did not vary between control and SOD1/G93A mice. Inaddition, we repeatedly tested the contamination of our prepara-tions also during the present experiments and found resultssuperimposable to those described above.
The complete block of receptor-induced glutamate releasebrought about by either selective mGlu1 or mGlu5 receptorantagonists indicates that the 3,5-DHPG-induced facilitation ofglutamate release founds on the activation of both receptorsubtypes and suggests the existence of a functional interplaybetween the two receptors. The data so far available in literature asto the release-regulating Group I mGlu receptors suggest a complexscenario, wherein mGlu1 and mGlu5 receptor-induced changes ofneurotransmitter release imply different mGlu1/mGlu5receptorereceptor interactions, depending on the brain region and/or on the population of nerve terminals investigated. Indeed, thetwo receptors were found to compensate each other (Volk et al.,2006; Luccini et al., 2007), to be functionally exclusive (Benquetet al., 2002; Feligioni et al., 2003) or to bind agonists withdifferent affinities (Reid et al., 1999; Fazal et al., 2003; Musanteet al., 2008). Since the existence of mGlu1/mGlu5 heterodimershas been recently demonstrated (Doumazane et al., 2011), onelikely hypothesis, on the basis of the present results, is that thespinal cord glutamatergic nerve terminals co-express mGlu1 andmGlu5 receptors that functionally cooperate as heterodimers, beingboth receptors essential to elicit glutamate release, at least atconcentration of 3,5-DHPG � 0.3 mM. If this is the case, antagonistsacting at one mGlu receptor subtype may also reduce the proba-bility of binding of the orthosteric agonist to the other functionallycoupled receptor (Milligan and Smith, 2007). Accordingly to thishypothesis, partial blockade of both receptors should not produceadditive effects. Actually, selective block of mGlu5 receptors bysubmaximal MPEP concentrations caused inhibition of the 3,5-DHPG-induced facilitation of glutamate release that could not befurther reinforced when submaximal concentrations of the mGlu1receptor antagonist LY367385 were concomitantly added to thesuperfusion medium.
The 3,5-DHPG-mediated facilitation of glutamate release hasbeen observed in glutamatergic terminals isolated from the spinalcord of WT mice, of mice overexpressing the unmodified human
Fig. 6. Quantification of mGlu1(A) and mGlu5 (B) receptor content in synaptosomesobtained from the spinal cord of WT, SOD1 and SOD1/G93A mice. SOD1/G93A micewere killed at the late phase of disease. Age-matched SOD1 and WT animals were usedas controls. mGlu1 and mGlu5 receptor amounts were measured by SDSe7%PAGE andWestern blot onto nitrocellulose membranes, using 10 mg of detergent soluble proteinextracts from membranes of synaptosomes purified from the lumbar spinal cord.Monomeric mGlu1 and mGlu5 receptor expression levels were normalized for b-tubulin III in the same blotted membrane and quantization reported in the bar plots.Representative immunoreactive bands are also shown. Mouse monoclonal antibodiesanti-mGlu1 receptor or anti-b-tubulin III and rabbit monoclonal antibody anti-mGlu5receptor were used. Results are expressed as relative density. Means � SEM of 6 micefor each group run in triplicate are presented. *p < 0.05 vs. WT or SOD1 mice (ANOVAplus Bonferroni test).
F. Giribaldi et al. / Neuropharmacology 66 (2013) 253e263260

Author's personal copy
SOD1 and of mice carrying the human SOD1 with the G93Amutation. In this context we found that 3,5-DHPG became effica-cious in releasing glutamate in SOD1/G93A animals also atconcentrations �0.3 mM, that were devoid of activity in WT andSOD1mice.When applied at concentrations lower than 0.3 mM, 3,5-DHPG caused a marked release of glutamate that was almost totallyprevented by antagonists of the mGlu5 receptor subtype, butinsensitive to mGlu1 receptor antagonists, suggesting that thereleasing effect can be ascribed mostly to the binding of the agonistto mGlu5 receptors. In other words, it seems that the presence ofthe G93A-mutated SOD1 allows the membrane insertion of pre-synaptic mGlu5 autoreceptors which bind submicromolarconcentrations of the agonist. Noteworthy, we found that mGlu5receptors are overexpressed in synaptosomal membranes isolatedfrom the spinal cord of SOD1/G93A mice. It should be noted,however, that also SOD1 or WT animals are endowed with func-tional mGlu5 receptors, that co-exist with mGlu1 receptors on thesame nerve terminal. Although it is difficult to conclude from theWestern blot experiments whether the increase of mGlu5 signal isdue to an increase of functional receptors located at the nerveterminal external membrane, on the basis of the above evidence itcould be speculated that the mere over-expression of the mGlu5receptor linked to mutated SOD1 allows the assembly of anincreased number of active membrane receptors, the stechiometryof which could disclose their ability to bind glutamate with highpotency in SOD1/G93A mice.
Since the 0.3 mM 3,5-DHPG effect on glutamate release, evi-denced in SOD1/G93A mice, could not be observed in healthyanimals, it seems plausible to hypothesize that this occurrencemight be relevant to the development of ALS symptoms. Ourexperiments were conducted in animals at a late stage of thepathology: further investigation is needed in presymptomatic mice,to strengthen the importance of the described mechanisms in thedisease progress. In any case, considering that submicromolarconcentration of exogenous agonist became efficacious in SOD1/G93Amice, it can be envisaged that these mGlu receptors can senseconcentration of glutamate that are usually inactive, triggeringa positive feed-back mechanism that, in turn, can favor anabnormal availability of the excitatory amino acid in the synapticcleft and excitotoxicity. The expression of Group I mGlu receptors inthe dorsal horn of spinal cord has been previously reported by othergroups (Alvarez et al., 2000; Aronica et al., 2001) and it has beensuggested that their differential expression might providea possible clue to the selective vulnerability of different MNsubpopulations in ALS (Anneser et al., 1999; Laslo et al., 2001), eventhough very few studies examined the protective activity of Group ImGlu receptor ligands against degeneration of cultured spinal cordMNs (Pizzi et al., 2000; Anneser et al., 2006; D’Antoni et al., 2011).
There is increasing evidence that MN death can be mostconvincingly explained assuming that mutated SOD1 expression innon-neuronal cells is required to drive disease progression (Ilievaet al., 2009; Foran et al., 2011; Li et al., 2011). In other words, theunique vulnerability of neuronal phenotypes in ALSwould need thecoincidental convergence of multiple causes involving neighbour-ing cell types. In this scenario, astrocytes promise to play a pivotalrole by influencing disease progression (Yamanaka et al., 2008) andMN vulnerability to excitotoxicity (Van Damme et al., 2007).Interestingly, mGlu5 receptor expression dramatically increases inastrocytes of SOD1/G93A mice and human individuals affected byALS (Aronica et al., 2001; Rossi et al., 2008). Stimulation of glialmGlu5 receptors may in turn contribute to astrogliosis (Anneseret al., 2004) and to aberrant intracellular Ca2þ elevation and celldegeneration (Martorana et al., 2012). In addition, a recent paperreported that a prolonged pharmacological blockade of mGlu5receptors present in astrocytes protects co-cultured MNs against
AMPA toxicity through a mechanism that involves reduction ofBDNF production/release from astrocytes and down-regulation ofthe GluR1 subunit of AMPA receptors present in MNs (D’Antoniet al., 2011). At variance, our data suggest that also glutamate-releasing neurons in the lumbar spinal cord overexpress thisreceptor subtype, which may account for over-release of glutamateand, possibly, neurotoxicity. Therefore, also neuronal mGlu5receptor blockade could be of utility in the therapy of ALS.
It has been recently shown that chronic treatment with theprototypic mGlu5 receptor antagonist MPEP attenuates cell death,delays the onset of motor symptoms, and prolongs survival inSOD1/G93A mutant mice, although only of few days (Rossi et al.,2008). Interestingly, new, selective and potent mGlu5 receptorantagonists are now under clinical trials. One of these, fenobam,which showed elevated efficacy in blocking both the high and lowconcentration-mediated 3,5-DHPG glutamate-releasing effects inour experiments, possesses anxiolytic and analgesic properties inclinic (Porter et al., 2005; Montana et al., 2009) and it was recentlytested also for fragile X syndrome (Berry-Kravis et al., 2009),exhibiting each time a good therapeutic profile without significantadverse effects. A more recent compound, AFQ056, is under clinicalevaluation in phase II for Parkinson disease dyskinesias (Levandiset al., 2008) and fragile X syndrome (Levenga et al., 2011) andshowed good safety profile and efficacy in already closed trials.Once tested in vivo in the ALS animal models and in case of morecompelling data respect to those described for MPEP, fenobam andAFQ056 may allow a direct translation of animal studies to theclinic.
In conclusion, we propose here a possible new mechanism thatmakes stronger the significance of the Group I metabotropicglutamate receptors as possible targets in ALS, since they producean abnormal stimulation of glutamate release. Accordingly, mGlu1and mGlu5 receptor antagonists may be envisaged as beneficialdrugs in the therapy of this neurodegenerative disease. Moreover,due to the role attributed to astrocytes in the developing of thedisease and of the possible importance of astrocytic mGlu1/5receptors in ALS, the blockade of these receptors my cause favor-able synergistic effects by interfering with both neuronal and glialbiochemistry.
Conflict of interest
The authors state no conflict of interest.
Acknowledgments
This work was supported by grants from Ministero dell’Uni-versità e Ricerca (PRIN project n. n. 2006058401 to GB) and byMinistero della Salute, Progetti Ordinari 2008 to GB. The authorsare grateful to Ms. Maura Agate for her excellent secretarialassistance.
References
Alvarez, F.J., Villalba, R.M., Carr, P.A., Grandes, P., Somohano, P.M., 2000. Differentialdistribution of metabotropic glutamate receptors 1a, 1b, and 5 in the rat spinalcord. J. Comp. Neurol. 422, 464e487.
Anneser, J.M., Borasio, G.D., Berthele, A., Zieglgänsberger, W., Tölle, T.R., 1999.Differential expression of group I metabotropic glutamate receptors in ratspinal cord somatic and autonomic motoneurons: possible implications for thepathogenesis of amyotrophic lateral sclerosis. Neurobiol. Dis. 6, 140e147.
Anneser, J.M., Chahli, C., Ince, P.G., Borasio, G.D., Shaw, P.J., 2004. Glial proliferationand metabotropic glutamate receptor expression in amyotrophic lateral scle-rosis. J. Neuropathol. Exp. Neurol. 63, 831e840.
Anneser, J.M., Chahli, C., Borasio, G.D., 2006. Protective effect of metabotropicglutamate receptor inhibition on amyotrophic lateral sclerosis-cerebrospinalfluid toxicity in vitro. Neuroscience 141, 1879e1886.
F. Giribaldi et al. / Neuropharmacology 66 (2013) 253e263 261

Author's personal copy
Aronica, E., Catania, M.V., Geurts, J., Yankaya, B., Troost, D., 2001. Immunohisto-chemical localization of group I and II metabotropic glutamate receptors incontrol and amyotrophic lateral sclerosis human spinal cord: upregulation inreactive astrocytes. Neuroscience 105, 509e520.
Barbiero, V.S., Giambelli, R., Musazzi, L., Tiraboschi, E., Tardito, D., Perez, J., Drago, F.,Racagni, G., Popoli, M., 2007. Chronic antidepressants induce redistribution anddifferential activation of alphaCaM kinase II between presynaptic compart-ments. Neuropsychopharmacology 32, 2511e2519.
Benquet, P., Gee, C.E., Gerber, U., 2002. Two distinct signaling pathways upregulateNMDA receptor responses via two distinct metabotropic glutamate receptorsubtypes. J. Neurosci. 22, 9679e9686.
Berry-Kravis, E., Hessl, D., Coffey, S., Hervey, C., Schneider, A., Yuhas, J., Hutchison, J.,Snape, M., Tranfaglia, M., Nguyen, D.V., Hagerman, R., 2009. A pilot open label,single dose trial of fenobam in adults with fragile X syndrome. J. Med. Genet.46, 266e271.
Bleasdale, J.E., Thakur, N.R., Gremban, R.S., Bundy, G.L., Fitzpatrick, F.A., Smith, R.J.,Bunting, S., 1990. Selective inhibition of receptor-coupled phospholipase C-dependent processes in human platelets and polymorphonuclear neutrophils.J. Pharmacol. Exp. Ther. 255, 756e768.
Boillée, S., Yamanaka, K., Lobsiger, C.S., Copeland, N.G., Jenkins, N.A., Kassiotis, G.,Kollias, G., Cleveland, D.W., 2006. Onset and progression in inherited ALSdetermined by motor neurons and microglia. Science 312, 1389e1392.
Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of micro-gram quantities of protein utilizing the principle of protein dye binding. Anal.Biochem. 72, 248e254.
Breysse, N., Amalric, M., Salin, P., 2003. Metabotropic glutamate 5 receptor blockadealleviates akinesia by normalizing activity of selective basal-ganglia structuresin parkinsonian rats. J. Neurosci. 23, 8302e8309.
Brown, R.H., 1995. Amyotrophic lateral sclerosis: recent insights from genetic andtransgenic mice. Cell 80, 687e692.
Bruno, V., Battaglia, G., Kingston, A., O’Neill, M.J., Catania, M.V., Di Grezia, R.,Nicoletti, F., 1999. Neuroprotective activity of the potent and selective mGlu1ametabotropic glutamate receptor antagonist, (þ)-2-methyl-4 carbox-yphenylglycine (LY367385): comparison with LY357366, a broader spectrumantagonist with equal affinity for mGlu1a and mGlu5 receptors. Neurophar-macology 38, 199e207.
Cartmell, J., Schoepp, D.D., 2000. Regulation of neurotransmitter release bymetabotropic glutamate receptors. J. Neurochem. 75, 889e907.
Cleveland, D.W., Bruijn, L.I., Wong, P.C., Marszalek, J.R., Vechio, J.D., Lee, M.K.,Xu, X.S., Borchelt, D.R., Sisodia, S.S., Price, D.L., 1996. Mechanisms of selectivemotor neuron death in transgenic mouse models of motor neuron disease.Neurology 47 (Suppl. 2), S54eS61. Discussion S61e62.
Conn, P.J., Pin, J.-P., 1997. Pharmacology and functions of metabotropic glutamatereceptors. Annu. Rev. Pharmacol. Toxicol. 37, 205e237.
D’Antoni, S., Berretta, A., Seminara, G., Longone, P., Giuffrida-Stella, A.M.,Battaglia, G., Sortino, M.A., Nicoletti, F., Catania, M.V., 2011. A prolonged phar-macological blockade of type-5 metabotropic glutamate receptors protectscultured spinal cord motor neurons against excitotoxic death. Neurobiol. Dis.42, 252e264.
De Blasi, A., Conn, P.J., Pin, J., Nicoletti, F., 2001. Molecular determinants of metab-otropic glutamate receptor signaling. Trends Pharmacol. Sci. 22, 114e120.
Dingledine, R., Borges, K., Bowie, D., Traynelis, S.F., 1999. The glutamate receptor ionchannels. Pharmacol. Rev. 51, 7e61.
Doumazane, E., Scholler, P., Zwier, J.M., Eric, T., Rondard, P., Pin, J.P., 2011. A newapproach to analyze cell surface protein complexes reveals specific hetero-dimeric metabotropic glutamate receptors. FASEB J. 25, 66e77.
Fazal, A., Parker, F., Palmer, A.M., Croucher, M.J., 2003. Characterisation of theactions of group I metabotropic glutamate receptor subtype selective ligands onexcitatory amino acid release and sodium-dependent re-uptake in rat cere-brocortical minislices. J. Neurochem. 86, 1346e1358.
Feligioni, M., Raiteri, L., Pattarini, R., Grilli, M., Buzzone, S., Cavazzani, P., Raiteri, M.,Pittaluga, A., 2003. The human immunodeficiency virus-1 protein Tat and itsdiscrete fragments evoke selective release of acetylcholine from human and ratcerebrocortical terminals through species-specific mechanisms. J. Neurosci. 23,6810e6888.
Ferraguti, G., Pierandrei, S., Bruno, S.M., Ceci, F., Strom, R., Lucarelli, M., 2011.A template for mutational data analysis of the CFTR gene. Clin. Chem. Lab. Med.49, 1447e1451.
Ferraiuolo, L., Kirby, J., Grierson, A.J., Sendtner, M., Shaw, P.J., 2011. Molecularpathways of motor neuron injury in amyotrophic lateral sclerosis. Nat. Rev.Neurol. 7, 616e630.
Foran, E., Bogush, A., Goffredo, M., Roncaglia, P., Gustincich, S., Pasinelli, P., Trotti, D.,2011. Motor neuron impairment mediated by a sumoylated fragment of the glialglutamate transporter EAAT2. Glia 59, 1719e1731.
Gasparini, F., Lingenhöhl, K., Stoehr, N., Flor, P.J., Heinrich, M., Vranesic, I., Biollaz, M.,Allgeier, H., Heckendorn, R., Urwyler, S., Varney, M.A., Johnson, E.C., Hess, S.D.,Rao, S.P., Sacaan, A.I., Santori, E.M., Veliçelebi, G., Kuhn, R., 1999. 2-Methyl-6-(phenylethynyl)-pyridine (MPEP), a potent, selective and systemically activemGlu5 receptor antagonist. Neuropharmacology 38, 1493e1503.
Gurney, M.E., Pu, H., Chiu, A.Y., Dal Canto, M.C., Polchow, C.Y., Alexander, D.D.,Caliendo, J., Hentati, A., Kwon, Y.W., Deng, H.X., et al., 1994. Motor neurondegeneration in mice that express a human Cu, Zn superoxide dismutasemutation. Science 264, 1772e1775.
Haidet-Phillips, A.M., Hester, M.E., Miranda, C.J., Meyer, K., Braun, L., Frakes, A.,Song, S., Likhite, S., Murtha, M.J., Foust, K.D., Rao, M., Eagle, A., Kammesheidt, A.,
Christensen, A., Mendell, J.R., Burghes, A.H., Kaspar, B.K., 2011. Astrocytes fromfamilial and sporadic ALS patients are toxic to motor neurons. Nat. Biotechnol.29, 824e828.
Herrero, I., Miras-Portugal, M.T., Sanchez-Prieto, J., 1992. Positive feedback ofglutamate exocytosis by metabotropic presynaptic receptor stimulation. Nature360, 163e166.
Ilieva, H., Polymenidou, M., Cleveland, D.W., 2009. Non-cell autonomous toxicity inneurodegenerative disorders: ALS and beyond. J. Cell Biol. 187, 761e772.
Laslo, P., Lipski, J., Funk, G.D., 2001. Differential expression of Group I metabotropicglutamate receptors in motoneurons at low and high risk for degeneration inALS. NeuroReport 12, 1903e1908.
Levandis, G., Bazzini, E., Armentero, M.T., Nappi, G., Blandini, F., 2008. Systemicadministration of an mGluR5 antagonist, but not unilateral subthalamic lesion,counteracts l-DOPA-induced dyskinesias in a rodent model of Parkinson’sdisease. Neurobiol. Dis. 29, 161e168.
Levenga, J., Hayashi, S., de Vrij, F.M., Koekkoek, S.K., van der Linde, H.C.,Nieuwenhuizen, I., 2011. AFQ056, a new mGluR5 antagonist for treatment offragile X syndrome. Neurobiol. Dis. 42, 311e317.
Li, Q., Spencer, N.Y., Pantazis, N.J., Engelhardt, J.F., 2011. ALSIN and SOD1G93Aregulate endosomal ROS production by glial cells and pro-inflammatory path-ways responsible for neurotoxicity. J. Biol. Chem. 286, 40151e40162.
Litschig, S., Gasparini, F., Rueegg, D., Stoehr, N., Flor, P.J., Vranesic, I., Prézeau, L.,Pin, J.P., Thomsen, C., Kuhn, R., 1999. CPCCOEt, a noncompetitive metabotropicglutamate receptor 1 antagonist, inhibits receptor signaling without affectingglutamate binding. Mol. Pharmacol. 55, 453e461.
Luccini, E., Musante, V., Neri, E., Brambilla Bas, M., Severi, P., Raiteri, M., Pittaluga, A.,2007. Functional interactions between presynaptic NMDA receptors andmetabotropic glutamate receptors co-expressed on rat and human noradren-ergic terminals. Br. J. Pharmacol. 151, 1087e1094.
Lujan, R., Nusser, Z., Roberts, J.D., Shigemoto, R., Somogyi, P., 1996. Perysinapticlocation of metabotropic glutamate receptors mGluR1 and mGluR5 ondendrites and dendritic spines in the rat hippocampus. Eur. J. Neurosci. 8,1488e1500.
MacDermott, A.B., Role, L.W., Siegelbaum, S.A., 1999. Presynaptic ionotropicreceptors and the control of transmitter release. Annu. Rev. Neurosci. 22,443e485.
Manders, E.M., Stap, J., Brakenhoff, G.J., van Driel, R., Aten, J.A., 1992. Dynamics ofthree-dimensional replication patterns during the S-phase, analysed by doublelabelling of DNA and confocal microscopy. J. Cell Sci. 103, 857e862.
Martorana, F., Brambilla, L., Valori, C.F., Bergamaschi, C., Roncoroni, C., Aronic, a E.,Volterra, A., Bezzi, P., Rossi, D., 2012. The BH4 domain of Bcl-X(L) rescuesastrocyte degeneration in amyotrophic lateral sclerosis by modulating intra-cellular calcium signals. Hum. Mol. Genet. 21, 826e840.
Maruyama, T., Kanaji, T., Nakade, S., Kanno, T., Mikoshiba, K., 1997. 2APB, 2-aminoethoxydiphenyl borate, a membrane-penetrable modulator of Ins(1,4,5)P3-induced Ca2þ release. J. Biochem. 122, 498e505.
Milanese, M., Zappettini, S., Jacchetti, E., Bonifacino, T., Cervetto, C., Usai, C.,Bonanno, G., 2010. In-vitro activation of GAT1 transporters expressed in spinalcord gliosomes stimulates glutamate release that is abnormally elevated in theSOD1/G93A(þ) mouse model of amyotrophic lateral sclerosis. J. Neurochem.113, 489e501.
Milanese, M., Zappettini, S., Onori, F., Musazzi, L., Tardito, D., Bonifacino, T.,Messa, M., Racagni, G., Usai, C., Benfenati, F., Popoli, M., Bonanno, G., 2011.Abnormal exocytotic release of glutamate in a mouse model of amyotrophiclateral sclerosis. J. Neurochem. 116, 1028e1042.
Milligan, G., Smith, N.J., 2007. Allosteric modulation of heterodimeric G-protein-coupled receptors. Trends Pharmacol. Sci. 28, 615e620.
Montana, M.C., Cavallone, L.F., Stubbert, K.K., Stefanescu, A.D., Kharasch, E.D.,Gereau 4th, R.W., 2009. The metabotropic glutamate receptor subtype 5antagonist fenobam is analgesic and has improved in vivo selectivity comparedwith the prototypical antagonist 2-methyl-6-(phenylethynyl)-pyridine.J. Pharmacol. Exp. Ther. 330, 834e843.
Moroni, F., Cozzi, A., Lombardi, G., Sourtcheva, S., Leonardi, P., Carfì, M., Pellicciari, R.,1998. Presynaptic mGlu1 type receptors potentiate transmitter output in the ratcortex. Eur. J. Pharmacol. 347, 189e195.
Morrison, B.M., Morrison, J.H., 1999. Amyotrophic lateral sclerosis associated withmutations in superoxide dismutase: a putative mechanism of degeneration.Brain Res. Rev. 29, 121e135.
Musante, V., Neri, E., Feligioni, M., Puliti, A., Pedrazzi, M., Conti, V., Usai, C.,Diaspro, A., Ravazzolo, R., Henley, J.M., Battaglia, G., Pittaluga, A., 2008.Presynaptic mGlu1 and mGlu5 autoreceptors facilitate glutamate exocy-tosis from mouse cortical nerve endings. Neuropharmacology 55,474e482.
Musante, V., Summa, M., Neri, E., Puliti, A., Godowicz, T.T., Severi, P., Battaglia, G.,Raiteri, M., Pittaluga, A., 2010. The HIV-1 viral protein Tat increases glutamateand decreases GABA exocytosis from human and mouse neocortical nerveendings. Cereb. Cortex 20, 1974e1984.
Nicoletti, F., Bockaert, J., Collingridge, G.L., Conn, P.J., Ferraguti, F., Schoepp, D.D.,Wroblewski, J.T., Pin, J.P., 2011. Metabotropic glutamate receptors: from theworkbench to the bedside. Neuropharmacology 60, 1017e1041.
Pedrazzi, M., Raiteri, L., Bonanno, G., Patrone, M., Ledda, S., Passalacqua, M.,Milanese, M., Melloni, E., Raiteri, M., Pontremoli, s., Sparatore, B., 2006. Stim-ulation of excitatory amino acid release from adult mouse brain glia sub-cellular particles by high mobility group box 1 protein. J. Neurochem. 99,827e838.
F. Giribaldi et al. / Neuropharmacology 66 (2013) 253e263262

Author's personal copy
Perry, T.L., Krieger, C., Hansen, S., Eisen, A., 1990. Amyotrophic lateral sclerosisamino acid levels in plasma and cerebrospinal fluid. Ann. Neurol. 28, 12e17.
Pin, J.-P., Galvez, T., Prézeau, L., 2003. Evolution, structure, and activation mecha-nism of family 3/C G protein-coupled receptors. Pharmacol. Ther. 98, 325e354.
Pizzi, M., Benarese, M., Boroni, F., Goffi, F., Valerio, A., Spano, P.F., 2000. Neuro-protection by metabotropic glutamate receptor agonists on kainate-induceddegeneration of motor neurons in spinal cord slices from adult rat. Neuro-pharmacology 39, 903e910.
Porter, R.H., Jaeschke, G., Spooren, W., Ballard, T.M., Büttelmann, B., Kolczewski, S.,Peters, J.U., Prinssen, E., Wichmann, J., Vieira, E., Mühlemann, A., Gatti, S.,Mutel, V., Malherbe, P., 2005. Fenobam: a clinically validated non-benzodiazepine anxiolytic is a potent, selective, and noncompetitive mGlu5receptor antagonist with inverse agonist activity. J. Pharmacol. Exp. Ther. 315,711e721.
Raiteri, M., Bonanno, G., Marchi, M., Maura, G., 1984. Is there a functional linkagebetween neurotransmitter uptake mechanisms and presynaptic receptors?J. Pharmacol. Exp. Ther. 231, 671e677.
Raiteri, L., Paolucci, E., Prisco, S., Raiteri, M., Bonanno, G., 2003. Activation ofa glycine transporter on spinal cord neurons causes enhanced glutamate releasein a mouse model of amyotrophic lateral sclerosis. Br. J. Pharmacol. 138,1021e1025.
Raiteri, L., Stigliani, S., Zappettino, S., Mercuri, N.B., Raiteri, M., Bonanno, G., 2004.Excessive and precocious glutamate release in a mouse model of amyotrophiclateral sclerosis. Neuropharmacology 46, 782e792.
Raiteri, M., 2008. Presynaptic metabotropic glutamate and GABAB receptors. Handb.Exp. Pharmacol. 184, 373e407.
Reid, M.E., Toms, N.J., Bedingfield, J.S., Roberts, P.J., 1999. Group I mGlu receptorspotentiate synaptosomal [3H]glutamate release independently of exogenouslyapplied arachidonic acid. Neuropharmacology 38, 477e485.
Risterucci, C., Coccurello, R., Banasr, M., Stutzmann, J.M., Amalric, M., Nieoullon, A.,2006. The metabotropic glutamate receptor subtype 5 antagonist MPEP and theNaþ channel blocker riluzole show different neuroprotective profiles inreversing behavioral deficits induced by excitotoxic prefrontal cortex lesions.Neuroscience 137, 211e220.
Rossi, D., Brambilla, L., Valori, C.F., Roncoroni, C., Prugnola, A., Yokota, T.,Bredesen, D.E., Volterra, A., 2008. Focal degeneration of astrocytes in amyo-trophic lateral sclerosis. Cell Death Differ. 15, 1691e1700.
Rothstein, J.D., Van Kammen, M., Levey, A.I., Martin, L.J., Kuncl, R.W., 1995. Selectiveloss of glial glutamate transporter GLT-1 in amyotrophic lateral sclerosis. Ann.Neurol. 38, 73e84.
Schoepp, D.D., 2001. Unveiling the functions of presynaptic metabotropic glutamatereceptors in the central nervous system. J. Pharmacol. Exp. Ther. 299, 12e20.
Sistiaga, A., Herrero, I., Conquet, F., Sanchez-Prieto, J., 1998. The metabotropicglutamate receptor 1 is not involved in the facilitation of glutamate release incerebrocortical nerve terminals. Neuropharmacology 37, 1485e1492.
Stigliani, S., Zappettini, S., Raiteri, L., Passalacqua, M., Melloni, E., Venturi, C.,Tacchetti, C., Diaspro, A., Usai, C., Bonanno, G., 2006. Glia re-sealed particlesfreshly prepared from adult rat brain are competent for exocytotic release ofglutamate. J. Neurochem. 96, 656e668.
Van Damme, P., Bogaert, E., Dewil, M., Hersmus, N., Kiraly, D., Scheveneels, W.,Bockx, I., Braeken, D., Verpoorten, N., Verhoeven, K., Timmerman, V.,Herijgers, P., Callewaert, G., Carmeliet, P., Van Den Bosch, L., Robberecht, W.,2007. Astrocytes regulate GluR2 expression in motor neurons and theirvulnerability to excitotoxicity. Proc. Natl. Acad. Sci. USA 104, 14825e14830.
Vernon, A.C., Zbarsky, V., Datla, K.P., Croucher, M.J., Dexter, D.T., 2007. Subtypeselective antagonism of substantia nigra pars compacta Group I metabotropicglutamate receptors protects the nigrostriatal system against 6-hydroxydopamine toxicity in vivo. J. Neurochem. 103, 1075e1091.
Volk, L.J., Daly, C.A., Huber, K.M., 2006. Differential roles for group 1 mGluRsubtypes in induction and expression of chemically induced Hippocampal long-term depression. J. Neurophysiol. 95, 2427e2438.
Wang, S.-J., Sihra, T.S., 2004. Noncompetitive metabotropic glutamate5 receptorantagonist (E)-2-methyl-6-styryl-pyridine (SIB1893) depresses glutamaterelease through inhibition of voltage-dependent Ca2þ entry in rat cere-brocortical nerve terminals (synaptosomes). J. Pharmacol. Exp. Ther. 309,951e958.
Wang, Y.J., Tseng, G.F., 2004. Spinal axonal injury transiently elevates the level ofmetabotropic glutamate receptor 5, but not 1, in cord-projection centralneurons. J. Neurotrauma 21, 479e489.
Wi�sniewski, K., Car, H., 2002. (S)-3,5-DHPG: a review. CNS Drug Rev. 8, 101e116.Yamanaka, K., Chun, S.J., Boillee, S., Fujimori-Tonou, N., Yamashita, H.,
Gutmann, D.H., Takahashi, R., Misawa, H., Cleveland, D.W., 2008. Astrocytes asdeterminants of disease progression in inherited amyotrophic lateral sclerosis.Nat. Neurosci. 11, 251e253.
F. Giribaldi et al. / Neuropharmacology 66 (2013) 253e263 263
Copyright © 2022 FDOKUMEN




















![In vivo positron emission tomography imaging with [ 11 C]ABP688: binding variability and specificity for the metabotropic glutamate receptor subtype 5 in baboons](https://static.fdokumen.com/doc/165x107/6316b26ad18b031ae106426d/in-vivo-positron-emission-tomography-imaging-with-11-cabp688-binding-variability.jpg)