
Activation of group I metabotropic glutamate receptors elicits pH changes in cultured rat cortical...
12
ACTIVATION OF GROUP I METABOTROPIC GLUTAMATE RECEPTORS ELICITS pH CHANGES IN CULTURED RAT CORTICAL GLIA AND NEURONS B. J. AMOS,*² A. MATHIE* and C. D. RICHARDS²‡ *Departments of Pharmacology and ²Physiology, Royal Free Hospital School of Medicine, Rowland Hill Street, London NW3 2PF, U.K. Abstract––Activation of metabotropic glutamate receptors is known to elicit a rise in intracellular Ca 2+ and the present study was undertaken to see whether they also modulate the intracellular pH (pH i ) of neurons and glia. Measurements of the pH i of neurons and astrocytes were made with the ratiometric fluorescent dye 2*,7*-biscarboxyethyl-5,6-carboxyfluorescein. In the absence of bicarbonate, stimulation with the specific metabotropic glutamate receptor agonist 1S,3R-1-aminocyclopentane-1,3-dicarboxylic acid caused a fall in pH i in both astrocytes and neurons. In the presence of bicarbonate, stimulation with 25 μM 1S,3R-1-aminocyclopentane-1,3-dicarboxylic acid elicited a rise in pH i in the astrocytes, while the neurons responded with a small acidification. The astrocytic alkalinization could also be elicited by the specific group I metabotropic glutamate receptor agonist (S)-3-hydroxyphenylglycine but not by the group II agonist (2S,1*S,2*S)-(2-carboxycyclopropyl)glycine or by the group III agonist (+)-2-amino-4- phosphonobutyric acid. The alkalinization of glial cells could be reduced by preloading the cells with BAPTA, but not by removal of extracellular Ca 2+ . Depolarization of the astrocytes with potassium elicited a small alkalinization, but stimulation with 100 μM 1S,3R-1-aminocyclopentane-1,3-dicarboxylic acid in high potassium medium elicited a further alkalinization. It is concluded that activation of group I metabotropic glutamate receptors leads to an alkalinization of astrocytes by a process that involves an elevation of intracellular Ca 2+ . The pH i changes that follow activation of the metabotropic glutamate receptors may play a role in initiation of glial proliferation following cerebral injury. ? 1998 IBRO. Published by Elsevier Science Ltd. Key words: intracellular pH, calcium, metabotropic glutamate receptor, glia, neuron. In the CNS there have been reports that neural activity is associated with changes in intracellular pH (pH i ) in both neurons and glia (see Refs 14 and 15 for reviews). For example, neurons respond to depolar- izing stimuli with an intracellular acidification, 1 while glial pH i increases both during normal electrical activity and during spreading depression. 16 Acti- vation of glutamatergic synapses in the cerebellum and hippocampus elicits an extracellular alkaliniz- ation, 13 and it has been suggested that changes in pH i may be associated with neuronal death as a consequence of glutamate excitotoxicity. 20 Since the discovery of the metabotropic glutamate receptors (mGluRs) in the mid-1980s, 29,42 the involvement of these receptors in many aspects of neuronal function has been recognized. mGluRs have been shown to be involved in neural development 29 and synaptic plasticity, 6 processes which necessarily will involve significant changes to the biochemistry of a cell. In view of this, and the known effects of receptor activation in non-neuronal tissues on pH i , we have investigated the effects of a number of selective mGluR agonists on the pH i of cultured neurons and glia. The specific questions we have sought to answer are: does activation of mGluRs lead to changes in pH i in either neurons or glia? If so, which receptors are involved and what are the mechanisms that underlie the pH i change? Preliminary accounts of some our results have been presented to the Physiological Society. 3,4 EXPERIMENTAL PROCEDURES Cell culture Neonatal Sprague–Dawley rats (three to five days old) were anaesthetized with isoflurane and then decapitated. Sections of neocortex (chiefly from the somatomotor region) were aseptically dissected and placed in HEPES- buffered minimum essential medium (MEM). The tissue was mechanically dissociated by passing the tissue through fire-polished Pasteur pipettes of decreasing bore. The cell debris was allowed to settle and the supernatant containing ‡To whom correspondence should be addressed. Abbreviations: ACPD, 1-aminocyclopentane-1,3-dicarbo- xylic acid; AM, acetoxymethyl ester; BAPTA, 1,2 bis(o- aminophenoxy)ethane NNN*N* tetra-acetate; BAPTA- AM, BAPTA tetrakis acetoxymethylester; BCECF, 2,7*-biscarboxyethyl-5,6-carboxyfluorescein; HEPES, N- 2-hydroxyethylpiperazine-N*-2-ethanesulphonic acid; 3-HPG, (S)-3-hydroxyphenylglycine; -AP4, (+)-2- amino-4-phosphonobutyric acid; -CCG-I, (2S,1*S,2*S)- (2-carboxycyclopropyl)-glycine; MEM, minimum essential medium; mGluR, metabotropic glutamate receptor; pH i , intracellular pH. Pergamon Neuroscience Vol. 86, No. 4, pp. 1109–1120, 1998 Copyright ? 1998 IBRO. Published by Elsevier Science Ltd Printed in Great Britain. All rights reserved 0306–4522/98 $19.00+0.00 PII: S0306-4522(98)00072-4 1109
Transcript of Activation of group I metabotropic glutamate receptors elicits pH changes in cultured rat cortical...

ACTIVATION OF GROUP I METABOTROPIC GLUTAMATERECEPTORS ELICITS pH CHANGES IN CULTURED RAT
CORTICAL GLIA AND NEURONS
B. J. AMOS,*† A. MATHIE* and C. D. RICHARDS†‡*Departments of Pharmacology and †Physiology, Royal Free Hospital School of Medicine,
Rowland Hill Street, London NW3 2PF, U.K.
Abstract––Activation of metabotropic glutamate receptors is known to elicit a rise in intracellular Ca2+
and the present study was undertaken to see whether they also modulate the intracellular pH (pHi) ofneurons and glia. Measurements of the pHi of neurons and astrocytes were made with the ratiometricfluorescent dye 2*,7*-biscarboxyethyl-5,6-carboxyfluorescein. In the absence of bicarbonate, stimulationwith the specific metabotropic glutamate receptor agonist 1S,3R-1-aminocyclopentane-1,3-dicarboxylicacid caused a fall in pHi in both astrocytes and neurons. In the presence of bicarbonate, stimulation with25 µM 1S,3R-1-aminocyclopentane-1,3-dicarboxylic acid elicited a rise in pHi in the astrocytes, while theneurons responded with a small acidification. The astrocytic alkalinization could also be elicited by thespecific group I metabotropic glutamate receptor agonist (S)-3-hydroxyphenylglycine but not by the groupII agonist (2S,1*S,2*S)-(2-carboxycyclopropyl)glycine or by the group III agonist (+)-2-amino-4-phosphonobutyric acid. The alkalinization of glial cells could be reduced by preloading the cells withBAPTA, but not by removal of extracellular Ca2+. Depolarization of the astrocytes with potassiumelicited a small alkalinization, but stimulation with 100 µM 1S,3R-1-aminocyclopentane-1,3-dicarboxylicacid in high potassium medium elicited a further alkalinization.
It is concluded that activation of group I metabotropic glutamate receptors leads to an alkalinization ofastrocytes by a process that involves an elevation of intracellular Ca2+. The pHi changes that followactivation of the metabotropic glutamate receptors may play a role in initiation of glial proliferationfollowing cerebral injury. ? 1998 IBRO. Published by Elsevier Science Ltd.
Key words: intracellular pH, calcium, metabotropic glutamate receptor, glia, neuron.
In the CNS there have been reports that neuralactivity is associated with changes in intracellular pH(pHi) in both neurons and glia (see Refs 14 and 15 forreviews). For example, neurons respond to depolar-izing stimuli with an intracellular acidification,1 whileglial pHi increases both during normal electricalactivity and during spreading depression.16 Acti-vation of glutamatergic synapses in the cerebellumand hippocampus elicits an extracellular alkaliniz-ation,13 and it has been suggested that changes inpHi may be associated with neuronal death as aconsequence of glutamate excitotoxicity.20
Since the discovery of the metabotropic glutamatereceptors (mGluRs) in the mid-1980s,29,42 theinvolvement of these receptors in many aspects of
neuronal function has been recognized. mGluRs havebeen shown to be involved in neural development29
and synaptic plasticity,6 processes which necessarilywill involve significant changes to the biochemistry ofa cell. In view of this, and the known effects ofreceptor activation in non-neuronal tissues on pHi,we have investigated the effects of a number ofselective mGluR agonists on the pHi of culturedneurons and glia.
The specific questions we have sought to answerare: does activation of mGluRs lead to changes inpHi in either neurons or glia? If so, which receptorsare involved and what are the mechanisms thatunderlie the pHi change? Preliminary accountsof some our results have been presented to thePhysiological Society.3,4
EXPERIMENTAL PROCEDURES
Cell culture
Neonatal Sprague–Dawley rats (three to five days old)were anaesthetized with isoflurane and then decapitated.Sections of neocortex (chiefly from the somatomotorregion) were aseptically dissected and placed in HEPES-buffered minimum essential medium (MEM). The tissue wasmechanically dissociated by passing the tissue throughfire-polished Pasteur pipettes of decreasing bore. The celldebris was allowed to settle and the supernatant containing
‡To whom correspondence should be addressed.Abbreviations: ACPD, 1-aminocyclopentane-1,3-dicarbo-
xylic acid; AM, acetoxymethyl ester; BAPTA, 1,2 bis(o-aminophenoxy)ethane NNN*N* tetra-acetate; BAPTA-AM, BAPTA tetrakis acetoxymethylester; BCECF,2,7*-biscarboxyethyl-5,6-carboxyfluorescein; HEPES, N-2-hydroxyethylpiperazine-N*-2-ethanesulphonic acid;3-HPG, (S)-3-hydroxyphenylglycine; -AP4, (+)-2-amino-4-phosphonobutyric acid; -CCG-I, (2S,1*S,2*S)-(2-carboxycyclopropyl)-glycine; MEM, minimumessential medium; mGluR, metabotropic glutamatereceptor; pHi, intracellular pH.
Pergamon
Neuroscience Vol. 86, No. 4, pp. 1109–1120, 1998Copyright ? 1998 IBRO. Published by Elsevier Science Ltd
Printed in Great Britain. All rights reserved0306–4522/98 $19.00+0.00PII: S0306-4522(98)00072-4
1109

disaggregated cells was then spun at 1000 r.p.m. for 3 min toseparate the intact cells. The supernatant was removed andthe pellet resuspended in HEPES-buffered MEM. The cellswere then plated at a density of 0.5–1#106 cells per 50-mmculture dish containing MEM supplemented with 2.5% fetalbovine serum. Each dish contained coverslips that had beencoated with poly--lysine (0.1 mg/ml). The cultures weremaintained in a humidified 5% CO2 atmosphere at 37)C forfive to seven days before use. Occasionally, cultures of10–14 days were used, but no differences in the response toagonists were evident. Neurons were identified by their sizeand morphology (phase-bright cell bodies possessing longneurites with few recurrent branches) and astrocytes wereidentified as type I by their flattened phase-dark appearance(see Fig. 3). In the following text the type I astrocytes will bereferred to simply as astrocytes.
Intracellular Ca2+ and pH measurements
pHi estimates were made using the pH-sensitive fluophore2*,7*-biscarboxyethyl-5,6-carboxyfluorescein (BCECF) andCa2+ measurements were made with Fura-2. Cells wereloaded with the indicators by incubating them at 37)C inbicarbonate buffer containing 10 µM of the acetoxymethylester (AM) of either BCECF or Fura-2 for 15 min. Follow-ing the initial incubation, the cells were washed twice andincubated for a further 15–20 min before being mounted inthe experimental chamber. Measurements were made usingdual excitation (340 and 380 nm for Fura-2 with emissionabove 420 nm; for BCECF the excitation wavelengths were440 and 490 nm and the emitted light was collected above520 nm). Both dyes were supplied by Molecular Probes(Eugene, OR, U.S.A.).
Single cell fluorescence was monitored as described pre-viously35 for cerebellar granule neurons. All experimentswere conducted at room temperature (20–22)C). For fluor-escence imaging, light was collected via a cooled CCDcamera (supplied by Digital Pixel, Brighton, U.K.) andanalysed using software supplied by Kinetic Imaging(Liverpool, U.K.). To improve the signal-to-noise ratio, thecollection times were adjusted to increase the total numberof photons accumulated at the lowest intensity of emission.Thus, for Fura-2, the collection period for 340 nm excita-tion was three times that for 380 nm excitation (the collec-tion period was generally 600 ms compared to 200 ms). ForBCECF, the collection periods were 200 ms for 490 nmexcitation and 600 ms for 440 nm excitation. The ratiovalues in the figures are not corrected for these differentcollection times.
The signals at each wavelength were corrected forautofluorescence, bound indicator and scattered light bysubtracting an image of the field after the cells had beenpermeabilized with 0.1% Triton in water. The ratio valueswere then calculated. Calibration of the ratios in terms ofCa2+ and pH was carried out in situ in several experimentsusing the ionophores 4Br-A23187 for Ca2+ and nigericin forpH (see Refs 17, 25 and 35 for further details of calibrationprocedures). The average values from these calibrationshave been used to quantify the changes in intracellular Ca2+
and pH where this was considered appropriate.For determination of the local Ca2+ or pHi from the ratio
images, specific areas of interest were chosen. The averageratio value for the designated areas was then calculated andplotted as a function of time. For each experimental pro-cedure, the ‘‘n’’ values refer to the number of cells. Ineach case these cells were taken from a minimum ofthree coverslips from at least two separate cortical culturepreparations.
Solutions
For the experiments described here, the cells were eitherbathed in Locke’s solution buffered with 16 mM HEPES orin Locke’s solution containing 16 mM bicarbonate and
equilibrated with 5% CO2 and 95% O2. The unbufferedLocke’s solution had the following composition (mM):NaCl 140, KCl 3, MgCl2 1.8, CaCl2 1.25 and glucose 5. ForHEPES-buffered Locke, the pH was adjusted to 7.3 using1 M NaOH. The agonists were diluted from concentratedstocks into the appropriate bathing solution immediatelybefore use.
Materials
Routine chemicals and tissue culture materials were sup-plied by Sigma, and the specific mGluR agonists 1S,3R-1-aminocyclopentane-1,3-dicarboxylic acid (1S,3R-ACPD),(2S,1*S,2*S)-(2-carboxycyclopropyl)glycine (-CCG-I), (S)-3-hydroxyphenylglycine (3-HPG) and (+)-2-amino-4-phosphonobutyric acid (-AP4) were all obtained fromTocris Cookson (Langford, Bristol, U.K.).
Statistical analysis
Throughout the text values are given as mean&S.E.M.unless otherwise stated. Statistical comparisons were madewith the standard t-test or by analysis of variance.
RESULTS
Steady-state pHi in neurons and astrocytes
Following loading with BCECF, as described inthe Experimental Procedures, the neurons appeareda bright apple-green when their fluorescence wasexcited by irradiation of light at either 440 or 490 nm.The signals obtained from the astrocytes followingloading with BCECF were much less intense thanobtained from the neurons. In eight individuallycalibrated fields of cells the mean steady-state pHi
values were 6.98&0.19 (n=10) for the neurons and6.97&0.17 (n=16) for the astrocytes (mean&S.D.).These values refer to the steady-state pHi inbicarbonate-buffered medium. In HEPES-bufferedmedium the values were 7.10&0.36 (n=9) for theneurons and 6.80&0.12 (n=6) for the astrocytes.
Effects of 1S,3R-1-aminocyclopentane-1,3-dicarboxy-lic acid on intracellular pH in neurons
Bath application of 1S,3R-ACPD (25–100 µM) toneurons maintained in HEPES-buffered mediumelicited an acidification in 10 of 15 cells studied bysingle-cell fluorescence photometry. This pH changewas initially rapid (0.03 pH units/min over the first2 min), reaching its maximum after 5 min, and didnot recover to its original level within the time-courseof the experiment. Examination of fluorescenceimage data showed a similar proportion of neurons(16/23) responding to 25 µM 1S,3R-ACPD with anintracellular acidosis (Fig. 1a). The magnitude of thepHi response varied from cell to cell, but the ratiosignal declined by an average of 14&2% (S.E.M.),corresponding to an average fall in pHi of about0.15 units. The data from neurons stimulated inbicarbonate-buffered medium were less clear cut. Tenof 11 neurons showed a fall in pH following appli-cation of 25 µM 1S,3R-ACPD, while one neuronshowed a small alkalinization. The overall changein pHi for all neurons was a decline of 6&2%,
1110 B. J. Amos et al.
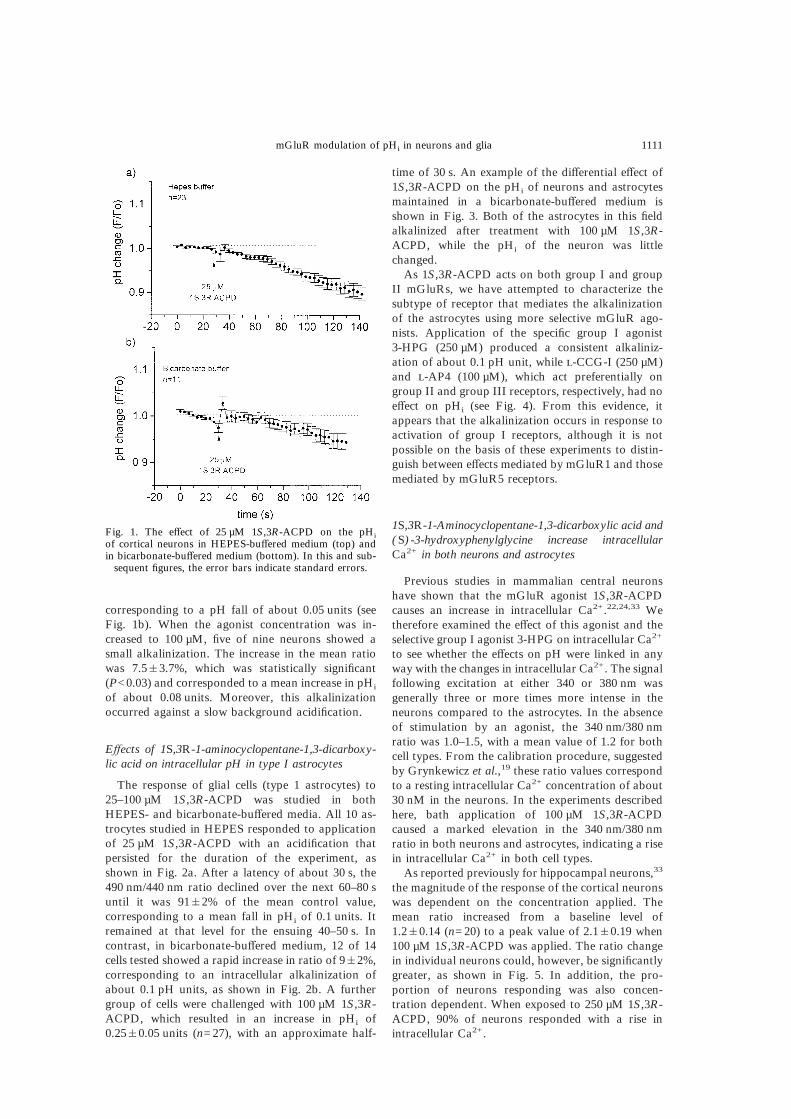
corresponding to a pH fall of about 0.05 units (seeFig. 1b). When the agonist concentration was in-creased to 100 µM, five of nine neurons showed asmall alkalinization. The increase in the mean ratiowas 7.5&3.7%, which was statistically significant(P<0.03) and corresponded to a mean increase in pHi
of about 0.08 units. Moreover, this alkalinizationoccurred against a slow background acidification.
Effects of 1S,3R-1-aminocyclopentane-1,3-dicarboxy-lic acid on intracellular pH in type I astrocytes
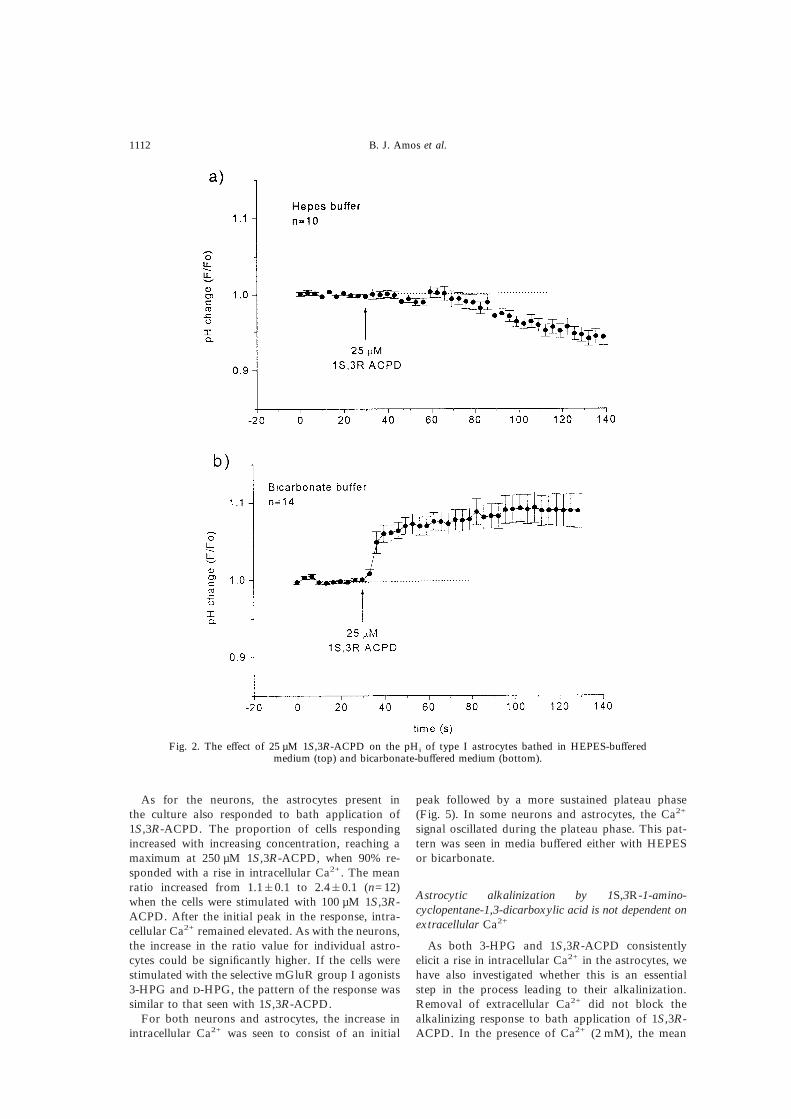
The response of glial cells (type 1 astrocytes) to25–100 µM 1S,3R-ACPD was studied in bothHEPES- and bicarbonate-buffered media. All 10 as-trocytes studied in HEPES responded to applicationof 25 µM 1S,3R-ACPD with an acidification thatpersisted for the duration of the experiment, asshown in Fig. 2a. After a latency of about 30 s, the490 nm/440 nm ratio declined over the next 60–80 suntil it was 91&2% of the mean control value,corresponding to a mean fall in pHi of 0.1 units. Itremained at that level for the ensuing 40–50 s. Incontrast, in bicarbonate-buffered medium, 12 of 14cells tested showed a rapid increase in ratio of 9&2%,corresponding to an intracellular alkalinization ofabout 0.1 pH units, as shown in Fig. 2b. A furthergroup of cells were challenged with 100 µM 1S,3R-ACPD, which resulted in an increase in pHi of0.25&0.05 units (n=27), with an approximate half-
time of 30 s. An example of the differential effect of1S,3R-ACPD on the pHi of neurons and astrocytesmaintained in a bicarbonate-buffered medium isshown in Fig. 3. Both of the astrocytes in this fieldalkalinized after treatment with 100 µM 1S,3R-ACPD, while the pHi of the neuron was littlechanged.
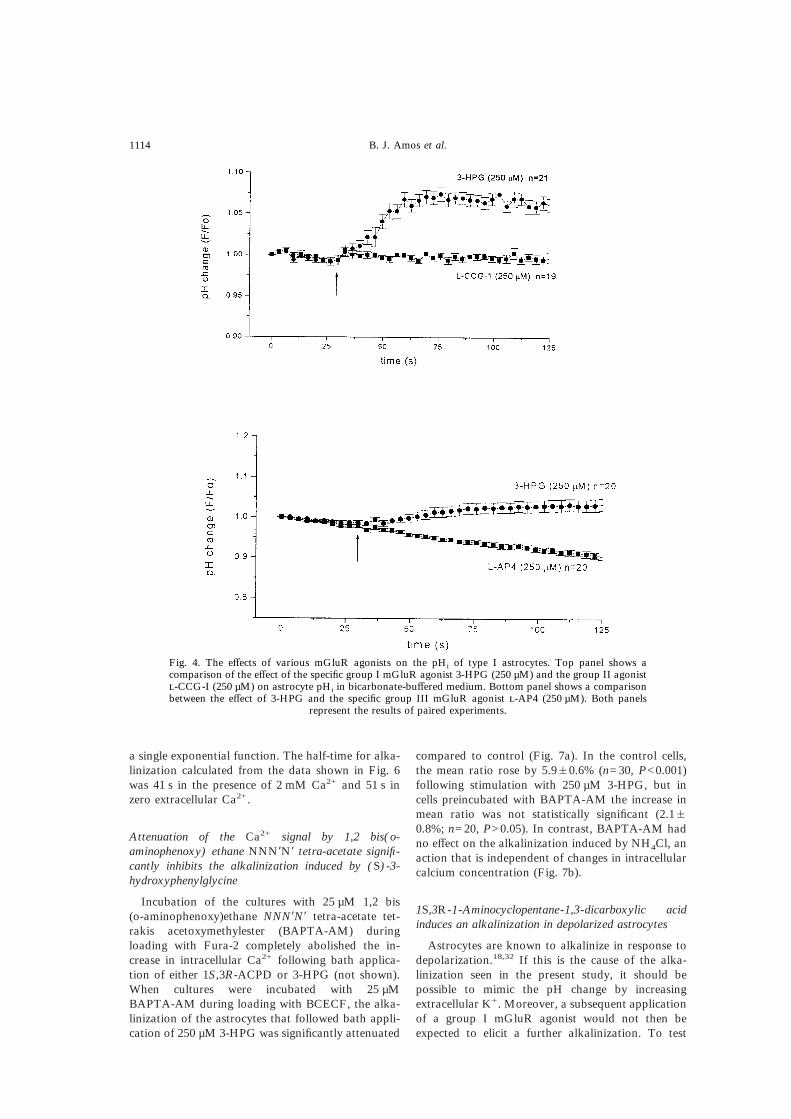
As 1S,3R-ACPD acts on both group I and groupII mGluRs, we have attempted to characterize thesubtype of receptor that mediates the alkalinizationof the astrocytes using more selective mGluR ago-nists. Application of the specific group I agonist3-HPG (250 µM) produced a consistent alkaliniz-ation of about 0.1 pH unit, while -CCG-I (250 µM)and -AP4 (100 µM), which act preferentially ongroup II and group III receptors, respectively, had noeffect on pHi (see Fig. 4). From this evidence, itappears that the alkalinization occurs in response toactivation of group I receptors, although it is notpossible on the basis of these experiments to distin-guish between effects mediated by mGluR1 and thosemediated by mGluR5 receptors.
1S,3R-1-Aminocyclopentane-1,3-dicarboxylic acid and(S)-3-hydroxyphenylglycine increase intracellularCa2+ in both neurons and astrocytes
Previous studies in mammalian central neuronshave shown that the mGluR agonist 1S,3R-ACPDcauses an increase in intracellular Ca2+.22,24,33 Wetherefore examined the effect of this agonist and theselective group I agonist 3-HPG on intracellular Ca2+
to see whether the effects on pH were linked in anyway with the changes in intracellular Ca2+. The signalfollowing excitation at either 340 or 380 nm wasgenerally three or more times more intense in theneurons compared to the astrocytes. In the absenceof stimulation by an agonist, the 340 nm/380 nmratio was 1.0–1.5, with a mean value of 1.2 for bothcell types. From the calibration procedure, suggestedby Grynkewicz et al.,19 these ratio values correspondto a resting intracellular Ca2+ concentration of about30 nM in the neurons. In the experiments describedhere, bath application of 100 µM 1S,3R-ACPDcaused a marked elevation in the 340 nm/380 nmratio in both neurons and astrocytes, indicating a risein intracellular Ca2+ in both cell types.
As reported previously for hippocampal neurons,33
the magnitude of the response of the cortical neuronswas dependent on the concentration applied. Themean ratio increased from a baseline level of1.2&0.14 (n=20) to a peak value of 2.1&0.19 when100 µM 1S,3R-ACPD was applied. The ratio changein individual neurons could, however, be significantlygreater, as shown in Fig. 5. In addition, the pro-portion of neurons responding was also concen-tration dependent. When exposed to 250 µM 1S,3R-ACPD, 90% of neurons responded with a rise inintracellular Ca2+.
Fig. 1. The effect of 25 µM 1S,3R-ACPD on the pHi
of cortical neurons in HEPES-buffered medium (top) andin bicarbonate-buffered medium (bottom). In this and sub-
sequent figures, the error bars indicate standard errors.
mGluR modulation of pHi in neurons and glia 1111
As for the neurons, the astrocytes present inthe culture also responded to bath application of1S,3R-ACPD. The proportion of cells respondingincreased with increasing concentration, reaching amaximum at 250 µM 1S,3R-ACPD, when 90% re-sponded with a rise in intracellular Ca2+. The meanratio increased from 1.1&0.1 to 2.4&0.1 (n=12)when the cells were stimulated with 100 µM 1S,3R-ACPD. After the initial peak in the response, intra-cellular Ca2+ remained elevated. As with the neurons,the increase in the ratio value for individual astro-cytes could be significantly higher. If the cells werestimulated with the selective mGluR group I agonists3-HPG and -HPG, the pattern of the response wassimilar to that seen with 1S,3R-ACPD.
For both neurons and astrocytes, the increase inintracellular Ca2+ was seen to consist of an initial
peak followed by a more sustained plateau phase(Fig. 5). In some neurons and astrocytes, the Ca2+
signal oscillated during the plateau phase. This pat-tern was seen in media buffered either with HEPESor bicarbonate.
Astrocytic alkalinization by 1S,3R-1-amino-cyclopentane-1,3-dicarboxylic acid is not dependent onextracellular Ca2+
As both 3-HPG and 1S,3R-ACPD consistentlyelicit a rise in intracellular Ca2+ in the astrocytes, wehave also investigated whether this is an essentialstep in the process leading to their alkalinization.Removal of extracellular Ca2+ did not block thealkalinizing response to bath application of 1S,3R-ACPD. In the presence of Ca2+ (2 mM), the mean
Fig. 2. The effect of 25 µM 1S,3R-ACPD on the pHi of type I astrocytes bathed in HEPES-bufferedmedium (top) and bicarbonate-buffered medium (bottom).
1112 B. J. Amos et al.

490 nm/440 nm ratio increased by 31&5% (n=27)following application of 100 µM 1S,3R-ACPD (Fig.6). In the absence of extracellular Ca2+, the meanincrease in ratio after stimulation was not signifi-
cantly different at 28&6%, corresponding to anincrease of approximately 0.25 pH units. The time-course of the pHi change was similar for both seriesof experiments. The increase in pHi could be fitted by
Fig. 3. The differential effect of 100 µM 1S,3R-ACPD on the pHi of one cortical neuron and two corticalastrocytes. (A) The left panel shows a phase-contrast image of the field of cells. The middle panel is apseudo-colour image showing uniform BCECF ratios for the astrocytes that are slightly greater than thatfor the neuron in the centre of the field. The right panel shows the same field during stimulation by 100 µM1S,3R-ACPD. An increase in BCECF ratio is indicated by an increased brightness, as indicated by thecalibration wedge (far right of panel A). (B) The time-course of the ratio change for the three cells aboveis shown. Note that the neuron shows little change in ratio, while both astrocytes show a significant
increase.
mGluR modulation of pHi in neurons and glia 1113
a single exponential function. The half-time for alka-linization calculated from the data shown in Fig. 6was 41 s in the presence of 2 mM Ca2+ and 51 s inzero extracellular Ca2+.
Attenuation of the Ca2+ signal by 1,2 bis(o-aminophenoxy) ethane NNN*N* tetra-acetate signifi-cantly inhibits the alkalinization induced by (S)-3-hydroxyphenylglycine
Incubation of the cultures with 25 µM 1,2 bis(o-aminophenoxy)ethane NNN*N* tetra-acetate tet-rakis acetoxymethylester (BAPTA-AM) duringloading with Fura-2 completely abolished the in-crease in intracellular Ca2+ following bath applica-tion of either 1S,3R-ACPD or 3-HPG (not shown).When cultures were incubated with 25 µMBAPTA-AM during loading with BCECF, the alka-linization of the astrocytes that followed bath appli-cation of 250 µM 3-HPG was significantly attenuated
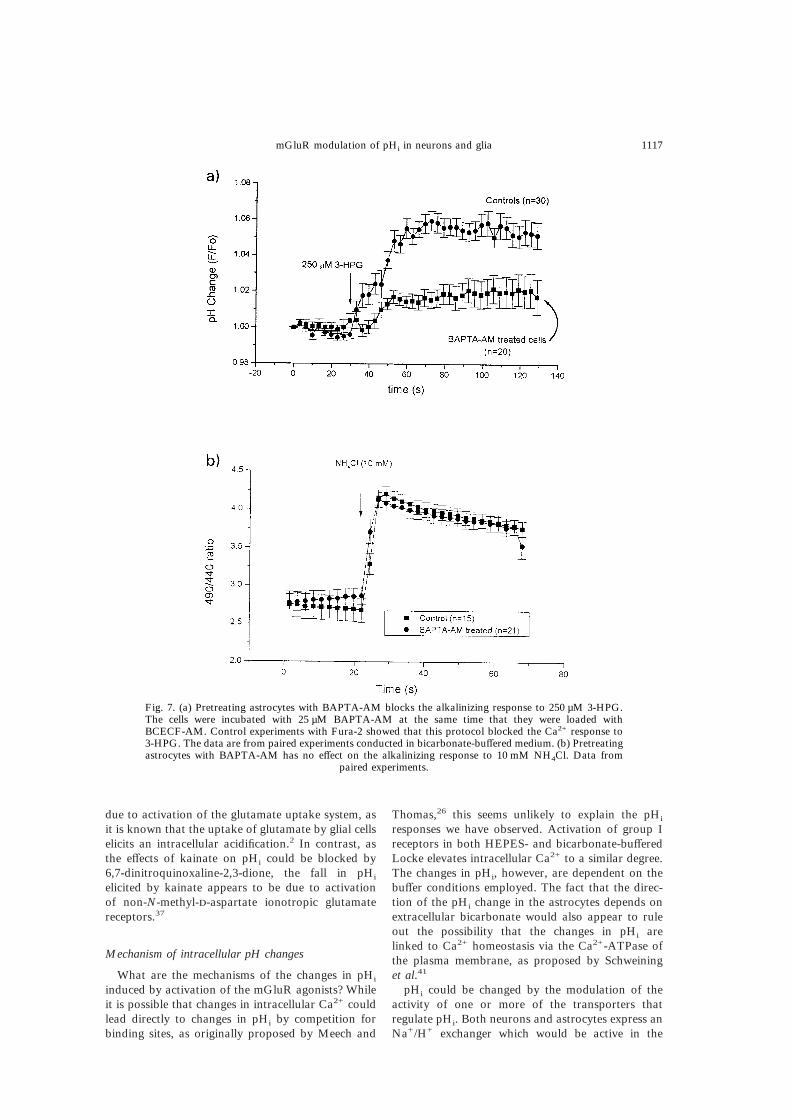
compared to control (Fig. 7a). In the control cells,the mean ratio rose by 5.9&0.6% (n=30, P<0.001)following stimulation with 250 µM 3-HPG, but incells preincubated with BAPTA-AM the increase inmean ratio was not statistically significant (2.1&0.8%; n=20, P>0.05). In contrast, BAPTA-AM hadno effect on the alkalinization induced by NH4Cl, anaction that is independent of changes in intracellularcalcium concentration (Fig. 7b).
1S,3R-1-Aminocyclopentane-1,3-dicarboxylic acidinduces an alkalinization in depolarized astrocytes
Astrocytes are known to alkalinize in response todepolarization.18,32 If this is the cause of the alka-linization seen in the present study, it should bepossible to mimic the pH change by increasingextracellular K+. Moreover, a subsequent applicationof a group I mGluR agonist would not then beexpected to elicit a further alkalinization. To test
Fig. 4. The effects of various mGluR agonists on the pHi of type I astrocytes. Top panel shows acomparison of the effect of the specific group I mGluR agonist 3-HPG (250 µM) and the group II agonist-CCG-I (250 µM) on astrocyte pHi in bicarbonate-buffered medium. Bottom panel shows a comparisonbetween the effect of 3-HPG and the specific group III mGluR agonist -AP4 (250 µM). Both panels
represent the results of paired experiments.
1114 B. J. Amos et al.
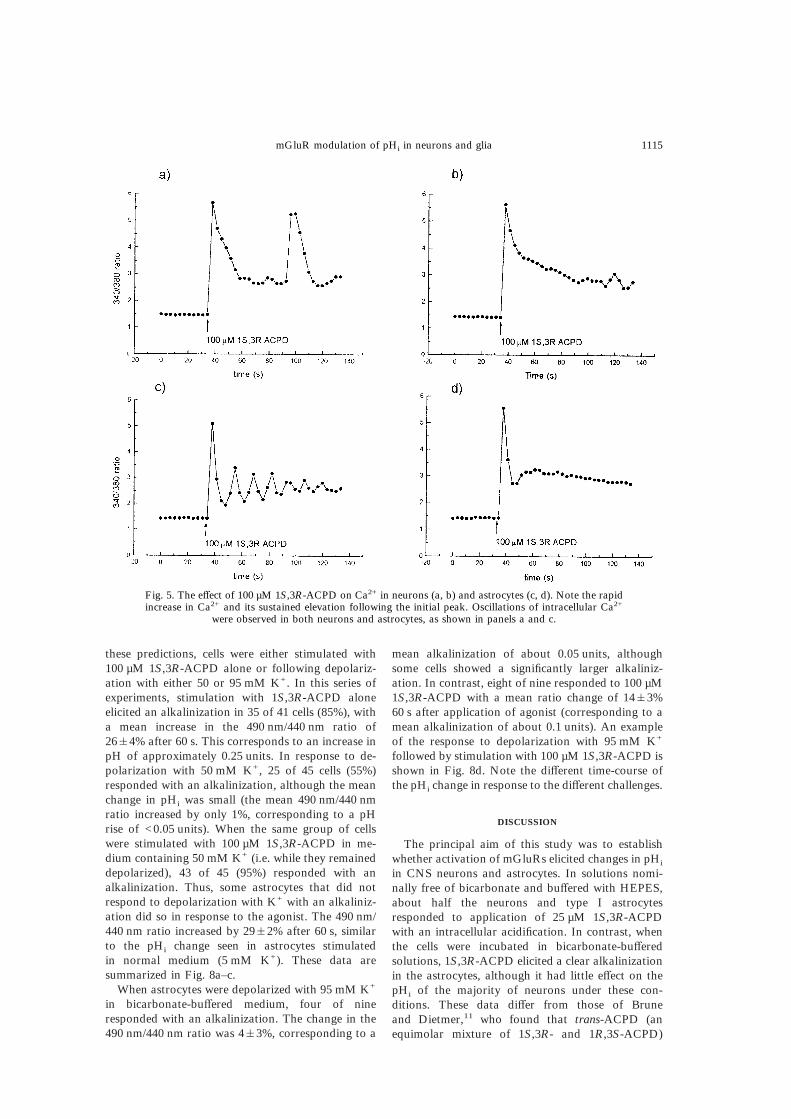
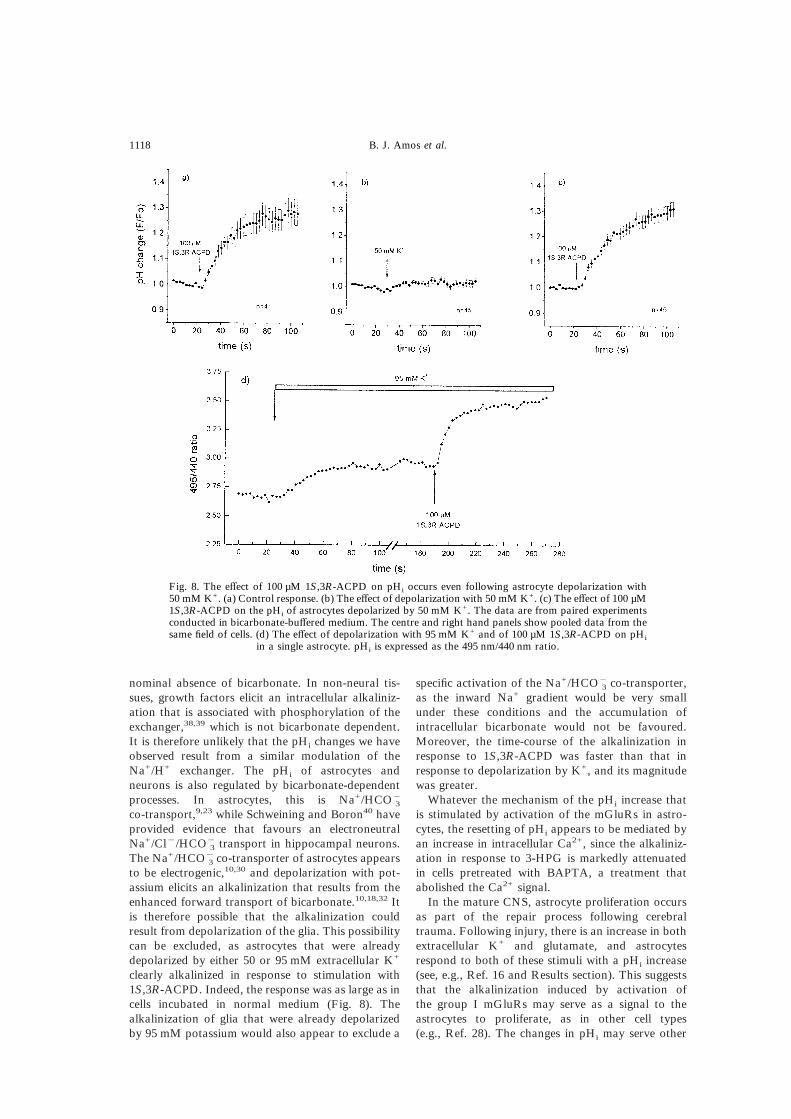
these predictions, cells were either stimulated with100 µM 1S,3R-ACPD alone or following depolariz-ation with either 50 or 95 mM K+. In this series ofexperiments, stimulation with 1S,3R-ACPD aloneelicited an alkalinization in 35 of 41 cells (85%), witha mean increase in the 490 nm/440 nm ratio of26&4% after 60 s. This corresponds to an increase inpH of approximately 0.25 units. In response to de-polarization with 50 mM K+, 25 of 45 cells (55%)responded with an alkalinization, although the meanchange in pHi was small (the mean 490 nm/440 nmratio increased by only 1%, corresponding to a pHrise of <0.05 units). When the same group of cellswere stimulated with 100 µM 1S,3R-ACPD in me-dium containing 50 mM K+ (i.e. while they remaineddepolarized), 43 of 45 (95%) responded with analkalinization. Thus, some astrocytes that did notrespond to depolarization with K+ with an alkaliniz-ation did so in response to the agonist. The 490 nm/440 nm ratio increased by 29&2% after 60 s, similarto the pHi change seen in astrocytes stimulatedin normal medium (5 mM K+). These data aresummarized in Fig. 8a–c.
When astrocytes were depolarized with 95 mM K+
in bicarbonate-buffered medium, four of nineresponded with an alkalinization. The change in the490 nm/440 nm ratio was 4&3%, corresponding to a
mean alkalinization of about 0.05 units, althoughsome cells showed a significantly larger alkaliniz-ation. In contrast, eight of nine responded to 100 µM1S,3R-ACPD with a mean ratio change of 14&3%60 s after application of agonist (corresponding to amean alkalinization of about 0.1 units). An exampleof the response to depolarization with 95 mM K+
followed by stimulation with 100 µM 1S,3R-ACPD isshown in Fig. 8d. Note the different time-course ofthe pHi change in response to the different challenges.
DISCUSSION
The principal aim of this study was to establishwhether activation of mGluRs elicited changes in pHi
in CNS neurons and astrocytes. In solutions nomi-nally free of bicarbonate and buffered with HEPES,about half the neurons and type I astrocytesresponded to application of 25 µM 1S,3R-ACPDwith an intracellular acidification. In contrast, whenthe cells were incubated in bicarbonate-bufferedsolutions, 1S,3R-ACPD elicited a clear alkalinizationin the astrocytes, although it had little effect on thepHi of the majority of neurons under these con-ditions. These data differ from those of Bruneand Dietmer,11 who found that trans-ACPD (anequimolar mixture of 1S,3R- and 1R,3S-ACPD)
Fig. 5. The effect of 100 µM 1S,3R-ACPD on Ca2+ in neurons (a, b) and astrocytes (c, d). Note the rapidincrease in Ca2+ and its sustained elevation following the initial peak. Oscillations of intracellular Ca2+
were observed in both neurons and astrocytes, as shown in panels a and c.
mGluR modulation of pHi in neurons and glia 1115

did not modulate the pHi of cultured astrocytesbathed in HEPES-buffered medium. They did, how-ever, note that the agonist evoked an increase inintracellular Ca2+.
The opposite directions of the pHi changes inHEPES- and bicarbonate-buffered Locke might beexplicable if the steady-state pHi of the astrocytesand neurons was significantly different in each situ-ation. However, the steady-state pHi values in bothHEPES- and bicarbonate-buffered solutions are verysimilar and close to previous estimates. In the presentstudy, mean steady-state pHi values in bicarbonate-buffered medium were 6.98 in neurons and 6.94 inastrocytes. In HEPES-buffered Locke the values were7.1 and 6.8, respectively. These values are close toprevious estimates of pHi in cultured CNS neurons inthe nominal absence of bicarbonate, which lie in therange 7.0–7.27.20,35 In bicarbonate-buffered Locke,neuronal pHi is very similar: mean values of 7.07 and7.17 have been reported for cultured corticalneurons5 and hippocampal neurons.36 The steady-state pHi of cultured astrocytes has been reportedpreviously12 to be 6.96, close to that seen in thepresent study. This is similar to the mean steady-statepHi of 7.04 reported for astrocytes in vivo.16
Activation of group I metabotropic glutamatereceptors alkalinizes the astrocytes
The mGluRs have been classified into three groupson the basis of their pharmacology, their sequencehomology and the transduction systems to whichthey are coupled.34 We have employed four agoniststo determine which group of receptors are involved inresetting the pHi of the glia. 1S,3R-ACPD actsselectively on group I and group II receptors and isrelatively inactive at group III receptors,21 while3-HPG is reported to be selective for group I recep-tors.8,34 Since both 1S,3R-ACPD and 3-HPG elicitedan intracellular alkalinization in the astrocytes whilethe group II agonist -CCG-I and the group IIIagonist -AP4 did not, it appears that the effects wehave observed on the pHi of neurons and astrocytesare due to activation of group I mGluRs. Whetherthe effects on pHi are mediated by mGluR1 ormGluR5, or both, remains to be established.
pHi is also modulated following activation of iono-tropic glutamate receptors. Glutamate and kainatehave both been found to elicit a decrease in the pHi ofglia maintained in HEPES-buffered media.11,37 Thisfall in pHi in response to glutamate is believed to be
Fig. 6. The effect of 100 µM 1S,3R-ACPD on both astrocyte and neuronal pHi is not abolished inCa2+-free media. The left hand panels show the effect of the agonist in the presence of 2 mM extracellularCa2+ on astrocytes (a) and neurons (c). The right hand panels show the effect of the same concentrationof agonist in Ca2+-free media for astrocytes (b) and for neurons (d). The data are from paired experiments
conducted in bicarbonate-buffered medium.
1116 B. J. Amos et al.
due to activation of the glutamate uptake system, asit is known that the uptake of glutamate by glial cellselicits an intracellular acidification.2 In contrast, asthe effects of kainate on pHi could be blocked by6,7-dinitroquinoxaline-2,3-dione, the fall in pHi
elicited by kainate appears to be due to activationof non-N-methyl--aspartate ionotropic glutamatereceptors.37
Mechanism of intracellular pH changes
What are the mechanisms of the changes in pHi
induced by activation of the mGluR agonists? Whileit is possible that changes in intracellular Ca2+ couldlead directly to changes in pHi by competition forbinding sites, as originally proposed by Meech and
Thomas,26 this seems unlikely to explain the pHi
responses we have observed. Activation of group Ireceptors in both HEPES- and bicarbonate-bufferedLocke elevates intracellular Ca2+ to a similar degree.The changes in pHi, however, are dependent on thebuffer conditions employed. The fact that the direc-tion of the pHi change in the astrocytes depends onextracellular bicarbonate would also appear to ruleout the possibility that the changes in pHi arelinked to Ca2+ homeostasis via the Ca2+-ATPase ofthe plasma membrane, as proposed by Schweininget al.41
pHi could be changed by the modulation of theactivity of one or more of the transporters thatregulate pHi. Both neurons and astrocytes express anNa+/H+ exchanger which would be active in the
Fig. 7. (a) Pretreating astrocytes with BAPTA-AM blocks the alkalinizing response to 250 µM 3-HPG.The cells were incubated with 25 µM BAPTA-AM at the same time that they were loaded withBCECF-AM. Control experiments with Fura-2 showed that this protocol blocked the Ca2+ response to3-HPG. The data are from paired experiments conducted in bicarbonate-buffered medium. (b) Pretreatingastrocytes with BAPTA-AM has no effect on the alkalinizing response to 10 mM NH4Cl. Data from
paired experiments.
mGluR modulation of pHi in neurons and glia 1117
nominal absence of bicarbonate. In non-neural tis-sues, growth factors elicit an intracellular alkaliniz-ation that is associated with phosphorylation of theexchanger,38,39 which is not bicarbonate dependent.It is therefore unlikely that the pHi changes we haveobserved result from a similar modulation of theNa+/H+ exchanger. The pHi of astrocytes andneurons is also regulated by bicarbonate-dependentprocesses. In astrocytes, this is Na+/HCO"
3
co-transport,9,23 while Schweining and Boron40 haveprovided evidence that favours an electroneutralNa+/Cl"/HCO"
3 transport in hippocampal neurons.The Na+/HCO"
3 co-transporter of astrocytes appearsto be electrogenic,10,30 and depolarization with pot-assium elicits an alkalinization that results from theenhanced forward transport of bicarbonate.10,18,32 Itis therefore possible that the alkalinization couldresult from depolarization of the glia. This possibilitycan be excluded, as astrocytes that were alreadydepolarized by either 50 or 95 mM extracellular K+
clearly alkalinized in response to stimulation with1S,3R-ACPD. Indeed, the response was as large as incells incubated in normal medium (Fig. 8). Thealkalinization of glia that were already depolarizedby 95 mM potassium would also appear to exclude a
specific activation of the Na+/HCO"3 co-transporter,
as the inward Na+ gradient would be very smallunder these conditions and the accumulation ofintracellular bicarbonate would not be favoured.Moreover, the time-course of the alkalinization inresponse to 1S,3R-ACPD was faster than that inresponse to depolarization by K+, and its magnitudewas greater.
Whatever the mechanism of the pHi increase thatis stimulated by activation of the mGluRs in astro-cytes, the resetting of pHi appears to be mediated byan increase in intracellular Ca2+, since the alkaliniz-ation in response to 3-HPG is markedly attenuatedin cells pretreated with BAPTA, a treatment thatabolished the Ca2+ signal.
In the mature CNS, astrocyte proliferation occursas part of the repair process following cerebraltrauma. Following injury, there is an increase in bothextracellular K+ and glutamate, and astrocytesrespond to both of these stimuli with a pHi increase(see, e.g., Ref. 16 and Results section). This suggeststhat the alkalinization induced by activation ofthe group I mGluRs may serve as a signal to theastrocytes to proliferate, as in other cell types(e.g., Ref. 28). The changes in pHi may serve other
Fig. 8. The effect of 100 µM 1S,3R-ACPD on pHi occurs even following astrocyte depolarization with50 mM K+. (a) Control response. (b) The effect of depolarization with 50 mM K+. (c) The effect of 100 µM1S,3R-ACPD on the pHi of astrocytes depolarized by 50 mM K+. The data are from paired experimentsconducted in bicarbonate-buffered medium. The centre and right hand panels show pooled data from thesame field of cells. (d) The effect of depolarization with 95 mM K+ and of 100 µM 1S,3R-ACPD on pHi
in a single astrocyte. pHi is expressed as the 495 nm/440 nm ratio.
1118 B. J. Amos et al.

functions. It is known that an increase in pHi stimu-lates the uptake of glutamate by the glia,7 and thiscould be important in keeping extracellular gluta-mate below the levels that may interfere with neuralsignalling, as well as preventing its neurotoxic effects.Moreover, an increase in glial pHi increases both gapjunction conductance43 and potassium channel ac-tivity in the plasma membrane,31 which may enhancethe buffering of extracellular K+ by the glia. Theactivity of high-threshold calcium channels is alsomodulated by changes in pHi.
27 Furthermore, suchchanges in pHi might be expected to lead to amplifiedpH gradients in the interstitial spaces surroundingglia and neurons due to the relatively small extra-cellular volume. Large extracellular pH signals mayhave profound effects on ion channels and transportsystems.
CONCLUSION
We have shown that activation of group I mGluRsmodulates the pHi of both astrocytes and neurons. Inthe nominal absence of bicarbonate, both cell typesacidify in response to stimulation but, in the presenceof extracellular bicarbonate (i.e. in the presence of thephysiological buffer), the astrocytes alkalinize whileneuronal pHi is less perturbed. The glial alkaliniza-tion is blocked when the intracellular Ca2+ responseto mGluR activation is blocked by BAPTA. Theseresults suggest that the pH change is mediated bya Ca2+-dependent process, although the precisemechanism for this effect remains to be elucidated.
Acknowledgements—We wish to thank the BBSRC andWellcome Trust for financial support.
REFERENCES
1. Ahmed Z. and Connor J. A. (1980) Intracellular pH changes induced by calcium influx during electrical activity inmolluscan neurones. J. gen. Physiol. 75, 403–426.
2. Amato A., Ballerini L. and Attwell D. (1994) Intracellular pH changes produced by glutamate uptake in rathippocampal slices. J. Neurophysiol. 72, 1686–1696.
3. Amos B. J. and Richards C. D. (1994) Effect of glutamate on intracellular pH of rat neurones maintained in culture.J. Physiol., Lond. 470, 43–44P.
4. Amos B. J., Mathie A. and Richards C. D. (1996) Effects of metabotropic glutamate receptor activation on theintracellular pH and calcium of rat cortical neurones and glia maintained in culture. J. Physiol., Lond. 491, 136P.
5. Amos B. J., Pocock G. and Richards C. D. (1996) On the role of bicarbonate as a hydrogen ion buffer in rat CNSneurones. Expl Physiol. 81, 623–632.
6. Bashir Z. I., Bortolotto Z. A., Davies C. H., Berreta N., Irving A. J., Seal A. J., Henley J. M., Jane D. E., WatkinsJ. C. and Collingridge G. L. (1993) Induction of LTP in the hippocampus needs synaptic activation of glutamatemetabotropic receptors. Nature 363, 347–350.
7. Billups B. and Attwell D. (1996) Modulation of non-vesicular glutamate release by pH. Nature 379, 171–174.8. Birse E. F., Eaton S. A., Jane D. E., Jones P. L., Porter R. H., Pook P. C., Sunter D. C., Udvarhelyi P. M., Wharton
B., Roberts P. J., Salt T. E. and Watkins J. C. (1993) Phenylglycine derivatives as new pharmacological tools forinvestigating the role of metabotropic glutamate receptors in the central nervous system. Neuroscience 52, 481–488.
9. Boyarsky G., Ransom B., Schule W.-R., Davis M. B. E. and Boron W. F. (1993) Intracellular pH regulation in singlecultured astrocytes from rat forebrain. Glia 8, 241–248.
10. Brookes N. and Turner R. J. (1994) K+-induced alkalinisation in mouse cerebral astrocytes mediated by reversal ofelectrogenic Na+–HCO"
3 cotransport. Am. J. Physiol. 267, C1633–C1640.11. Brune T. and Dietmer J. W. (1995) Intracellular acidification and Ca2+ transients in cultured rat cerebral astrocytes
evoked by glutamate agonists and noradrenaline. Glia 14, 153–161.12. Brune T., Fetzer S., Backus K. H. and Dietmer J. W. (1994) Evidence for electrogenic sodium–bicarbonate cotransport
in cultured rat cerebellar astrocytes. Pflugers Arch. 429, 64–71.13. Chen J. C. and Chesler M. (1992) Extracellular alkaline shifts in rat hippocampal slice are mediated by NMDA and
non-NMDA receptors. J. Neurophysiol. 68, 342–344.14. Chesler M. (1990) The regulation and modulation of pH in the nervous system. Prog. Neurobiol. 34, 401–427.15. Chesler M. and Kaila K. (1992) Modulation of pH by neuronal activity. Trends Neurosci. 15, 396–402.16. Chesler M. and Kraig R. P. (1989) Intracellular pH transients of mammalian astrocytes. J. Neurosci. 9, 2011–2019.17. Duchen M. R. (1992) Fluorescence—monitoring cell chemistry in vivo. In Monitoring Neuronal Activity—A Practical
Approach (ed. Stamford J. A.), Chap. 9, pp. 231–260. IRL, Oxford.18. Grichtchenko I. I. and Chesler M. (1994) Depolarization-induced acid secretion in gliotic hippocampal slices.
Neuroscience 62, 1057–1070.19. Grynkewicz G., Poenie M. and Tsein R. Y. (1985) A new generation of Ca2+ indicators with greatly improved
fluorescence properties. J. biol. Chem. 260, 3440–3450.20. Hartley Z. and Dubinsky J. M. (1993) Changes in intracellular pH associated with glutamate excitotoxicity. J.
Neurosci. 13, 4690–4699.21. Hayashi Y., Tanabe Y., Aramori I., Masu M., Shimamoto K., Ohfune Y. and Nakanishi S. (1992) Agonist analysis
of 2-(carboxycyclopropyl)glycine isomers for cloned metabotropic glutamate receptor subtypes expressed in Chinesehamster ovary cells. Br. J. Pharmac. 107, 539–543.
22. Irving A. J., Collingridge G. L. and Schofield J. G. (1992) Interactions between Ca2+ mobilising mechanisms incultured rat cerebellar granule cells. J. Physiol., Lond. 456, 667–680.
23. Kettenman H. and Schule W. R. (1988) Intracellular pH regulation in cultured mouse oligodendrocytes. J. Physiol.,Lond. 406, 147–162.
24. Linden D. J., Smeyne M. and Connor J. A. (1994) Trans-ACPD, a metabotropic receptor agonist, produces calciummobilisation and an inward current in cultured cerebellar Purkinje neurons. J. Neurophysiol. 71, 1992–1998.
mGluR modulation of pHi in neurons and glia 1119

25. Mason W. T., Hoyland Y., Davison I., Carew M., Somasundaram R., Tregear R., Zorec R., Lledo P. M., Shankar G.and Horton M. (1993) Quantitative real-time imaging of optical probes in living cells. In Fluorescent and LuminescentProbes for Biological Activity (ed. Mason W. T.), Chap. 12, pp. 161–195. Academic, London.
26. Meech R. W. and Thomas R. C. (1977) The effect of calcium injection on the intracellular sodium and pH of snailneurones. J. Physiol., Lond. 265, 867–879.
27. Mironov S. I. and Lux H. D. (1991) Cytoplasmic alkalinisation increases high threshold calcium current in chickdorsal root ganglion neurones. Pflugers Arch. 419, 138–143.
28. Moolenaar W. H., Tsein R. Y., van der Saag P. T. and de Laat S. W. (1983) Na+/H+ exchange and cytoplasmic pHin the action of growth factor in human fibroblasts. Nature 304, 645–648.
29. Nicoletti F., Meek J. L., Iadarola M. J., Chuang D. M., Roth B. L. and Costa E. (1986) Excitatory amino acidrecognition sites and stimulation of inositol phospholipid metabolism: developmental changes and interactions withá1-adrenoceptors. Proc. natn. Acad. Sci. U.S.A. 83, 1931–1935.
30. O’Connor E. R., Sontheim H. and Ransom B. R. (1994) Rat hippocampal astrocytes exhibit electrogenicsodium–bicarbonate co-transport. J. Neurophysiol. 72, 2580–2589.
31. Orkand R. K., Nicholls J. G. and Kuffler S. W. (1966) Effects of nerve impulses on the membrane potential of glialcells in the central nervous sytem of amphibia. J. Neurophysiol. 29, 788–806.
32. Pappas C. A. and Ransom B. R. (1994) Depolarisation-induced alkalinisation (DIA) in rat hippocampal astrocytes.J. Neurophysiol. 72, 2816–2826.
33. Phenna S., Jane S. D. and Chad J. E. (1995) Increased perinuclear Ca2+ activity evoked by metabotropic glutamatereceptor activation in rat hippocampal neurones. J. Physiol., Lond. 486, 149–161.
34. Pin J.-P. and Duvoisin R. (1995) The metabotropic glutamate receptors: structure and functions. Neuropharmacology34, 1–26.
35. Pocock G. and Richards C. D. (1992) Hydrogen ion regulation in rat cerebellar granule cells studied by single-cellfluorescence microscopy. Eur. J. Neurosci. 4, 136–143.
36. Raley-Susman K. M., Cragoe E. J. Jr, Sapolsky R. M. and Kopito R. R. (1991) Regulation of intracellular pH incultured hippocampal neurons by an amiloride-insensitive Na+/H+ exchanger. J. biol. Chem. 266, 2739–2745.
37. Rosel C. R. and Ransom B. R. (1996) Mechanisms of H+ and Na+ changes induced by glutamate, kainate and-aspartate in rat hippocampal astrocytes. J. Neurosci. 16, 5393–5404.
38. Sardet C., Counillon L., Franchi A. and Pouyssegur J. (1990) Growth factors induce phosphorylation of the Na+/H+
antiporter, a glycoprotein of 110 kD. Science 247, 723–726.39. Sardet C., Fafournoux P. and Pouyssegur J. (1991) Thrombin, epidermal growth factor and okadaic acid activate the
Na+/H+ exchanger, NHE-1, by phosphorylating a set of common sites. J. biol. Chem. 266, 19,166–19,171.40. Schweining C. J. and Boron W. F. (1994) Regulation of intracellular pH in pyramidal neurones from the rat
hippocampus by Na+ dependent Cl"–HCO"3 exchange. J. Physiol., Lond. 475, 59–67.
41. Schweining C. J., Kennedy H. J. and Thomas R. C. (1993) Calcium–hydrogen exchange by the plasma membraneCa2+-ATPase of voltage-clamped snail neurons. Proc. R. Soc. Lond., Ser. B 253, 285–289.
42. Sladeczek F., Pin J.-P., Recasens M., Bockaert J. and Weiss S. (1985) Glutamate stimulates inositol phosphateformation in striatal neurones. Nature 317, 717–719.
43. Spray D. C., Harris A. L. and Bennett M. V. L. (1981) Gap junctional conductance is a simple and sensitive functionof intracellular pH. Science 211, 712–715.
(Accepted 26 January 1998)
1120 B. J. Amos et al.




















