
Radial basis function networks for modeling marine electromagnetic survey

Organization of Radial and Non-RadialGlia in the Developing Rat Thalamus
CAROLINA FRASSONI,1* ALIDA AMADEO,2 BARBARA ORTINO,2
ANITA JARANOWSKA,1AND ROBERTO SPREAFICO1
1Dipartimento di Neurofisiologia Sperimentale, Istituto Nazionale Neurologico “C. Besta,”20133 Milano, Italy
2Dipartimento di Fisiologia e Biochimica Generali, Sezione di Istologia ed AnatomiaUmana, Universita degli Studi di Milano, 20122 Milano, Italy
ABSTRACTThe organization of glia and its relationship with migrating neurons were studied in the
rat developing thalamus with immunocytochemistry by using light, confocal, and electronmicroscopy. Carbocyanine labeling in cultured slice of the embryonic diencephalon was alsoused. At embryonic day (E) 14, vimentin immunoreactivity was observed in radial fasciclesspanning the neuroepithelium and extending from the ventricular zone to the lateral surfaceof the diencephalic vesicle. Vimentin-immunopositive fibers orthogonal to the radial oneswere also detected at subsequent developmental stages. At E16, radial and non-radialprocesses were clearly associated with migrating neurons identified by the neuronal markerscalretinin and g-aminobutyric acid. Non-radial glial fibers were no longer evident by E19.Radial fibers were gradually replaced by immature astrocytes at the end of embryonicdevelopment. In the perinatal period, vimentin immunoreactivity labeled immature astro-cytes and then gradually decreased; vimentin-immunopositive cells were only found in theinternal capsule by the second postnatal week. Glial fibrillary acidic protein immunoreactiv-ity appeared at birth in astrocytes of the internal capsule, but was not evident in most of theadult thalamic nuclei. Confocal and immunoelectron microscopy allowed direct examinationof the relationships between neurons and glial processes in the embryonic thalamus, showingthe coupling of neuronal membranes with both radial and non-radial glia during migration.Peculiar ultrastructural features of radial glia processes were observed. The occurrence ofnon-radial migration was confirmed by carbocyanine-labeled neuroblasts in E15 cultured slices.The data provide evidence that migrating thalamic cells follow both radial and non-radial glialpathways toward their destination. J. Comp. Neurol. 428:527–542, 2000. © 2000 Wiley-Liss, Inc.
Indexing terms: vimentin; astrocytes; diencephalon; neuronal migration; electron microscopy;
confocal microscopy
In the central nervous system (CNS), cell migration isusually restricted to specific developmental periods andoccurs mainly radially: radial glia forms a scaffold alongwhich neurons climb from the sites of generation towardtheir final position (Hatten, 1990, 1999; see also for reviewRakic, 1995a). Vimentin (VIM) is the primary structuralprotein of radial glia (Dhal et al., 1981) and characterizesthe peculiar state of immaturity associated with neuronalmigration. During the maturation of glia, VIM is replacedby glial fibrillary acidic protein (GFAP) and radial gliatransforms into astrocytes (Levitt and Rakic, 1980; Dhalet al., 1981; Pixley and DeVellis, 1984; McDermott andLantos 1989; Voigt, 1989; Culican et al., 1990).
VIM-immunoreactive (ir) radial glia is a common fea-ture in the developing mammalian brain, and a wealth of
immunocytochemical studies has described radial glia indistinct brain areas of different species during develop-ment (Levitt and Rakic, 1980; Bignami et al., 1982; Choi,1986; Misson et al., 1988, 1991; Hutchins and Casa-grande, 1989; McDermott and Lantos, 1989; Voigt, 1989;Stichel et al., 1991; Elmquist et al., 1994; Missler et al.,1994; Pulido-Caballero et al., 1994; Botchkina and Morin,
Grant sponsor: Italian Ministry of Health.*Correspondence to: Dr. Carolina Frassoni, Dipartimento di Neurofisio-
logia Sperimentale, Istituto Nazionale Neurologico “C: Besta,” Via Celoria11, 20133 Milano, Italy. E-mail: [email protected]
Received 24 March 2000; Revised 30 August 2000; Accepted 30 August2000
THE JOURNAL OF COMPARATIVE NEUROLOGY 428:527–542 (2000)
© 2000 WILEY-LISS, INC.

1995). Although the distribution and time of appearanceof VIM and GFAP in glial cells vary between species, theshift in the expression of these proteins is a consistentfeature. This implies a key role of the two intermediatefilament proteins during development and confirms theirimportance as markers of glial differentiation.
Despite the increasing number of immunocytochemicalstudies on glia during brain development, only a few in-vestigations so far have been devoted to the analysis ofradial glia in the diencephalon. In the developing ferretand rat thalamus, the early organization of glial cells(radial glia, astrocytes, and microglia), and chondroitinsulfate proteoglycan expression in these cells, were inves-tigated during axonal ingrowth and blood vessel invasion(Mitrofanis et al., 1997; Earle and Mitrofanis, 1998).These studies focused on the glial environment in thereticular and perireticular thalamic nuclei, two regions ofthe ventral thalamus in which corticofugal and corticope-tal axons undergo a major developmental reorganization(Mitrofanis and Baker, 1993). A detailed topographicalanalysis of radial glia, its ultrastructural features and itsrelation with migrating cells in the developing dorsal tha-lamic nuclei is, however, still lacking.
On this basis, the aim of the present study was toexamine, during the embryonic and postnatal develop-ment of the rat thalamus: (1) the morphological and topo-graphical organization of radial glia, identified by VIMimmunostaining; (2) the temporal sequence of disappear-ance of radial glia and appearance of mature astrocytes,identified by GFAP immunostaining; (3) the relationshipbetween glia and migrating thalamic neurons, identifiedby neuronal markers expressed during the prenatal pe-riod; and (4) the ultrastructure of thalamic glia, to revealthe milieu in which neurons migrate, as well as the con-tacts between migrating neurons and glial processes. Pre-liminary results have been presented in abstract form(Regondi et al., 1999). In addition, on the basis of thefindings observed in the thalamus during prenatal devel-opment, dynamic data on migrating neurons were alsoobtained in cultured slices of the embryonic diencephalonby means of carbocyanine labeling.
MATERIALS AND METHODS
Experiments were performed on 38 rats (Wistar,Charles River Italia, Calco, LC, Italy) ranging from em-bryonic day (E) 14 to postnatal day (P) 40. Brain sliceswere prepared from two additional E15 embryos for the invitro part of the study. The day of detection of a vaginalplug was designated as E1 and the day of birth as P0. Theexperiments were undertaken in accordance with theguidelines established in the Principles of Laboratory An-imal Care (NIH Publication no. 86-23, revised 1985). Allefforts were made to minimize animal suffering and toreduce the number of animals used.
Perfusion and histology
For the immunohistochemical part of the study, a totalof 20 embryos (E14, n 5 4; E15, n 5 3; E16, n 5 4; E17, n 53; E18, n 5 3; E19, n 5 3) were obtained from pregnantrats anesthetized with chloral hydrate (4%; 1 ml/100 gbody weight, i.p.). The embryos were quickly removedfrom the uterus and perfused transcardially with mixedaldehydes by using a glass capillary. A solution of 4%paraformaldehyde and 0.4% glutaraldehyde in 0.1 M
phosphate buffer (PB), pH 7.2, was used for light micros-copy, whereas a solution of 2.5% glutaraldehyde and0.5–1% paraformaldehyde in 0.1 M cacodylate buffer,pH.7.2, containing 0.01% CaCl2 , was used for ultrastruc-tural investigation. Embryos at E18–E19 were anesthe-tized with ether before perfusion. The heads were im-mersed in 4% paraformaldeyde for 24 hours at 4°C; thebrains were then removed and stored from 5 to 10 days inthe same fresh fixative. After embedding in 6% agarose,50- to 60-mm-thick serial sections were cut by means of aVibratome VT1000S (Leica, Heidelberg, Germany) andcollected in cold PB, pH 7.2.
A total of 18 postnatal rats (P1, n 5 1; P2, n 5 2; P4, n 53; P5, n 5 2; P7, n 5 2; P9, n 5 1; P11, n 5 2; P15, n 5 2;P21, n 5 1; P40, n 5 2) were anesthetized with chloralhydrate (4%; 1–2 ml/100 g body weight, i.p.) and perfusedthrough the heart with a buffered solution of 4% parafor-maldehyde and 0.4–1% glutaraldehyde. The brains wereremoved from the skull and soaked overnight in 4% para-formaldehyde at 4°C. Serial 50-mm-thick coronal sectionswere cut by using a Vibratome through the rostrocaudalextent of the thalamus and collected in PB, pH 7.2.
Immunocytochemical procedures
The sections destined for immunocytochemistry wereincubated for 30 minutes in 0.05 M NH4Cl in PB to quenchfree aldehyde groups, rinsed in 0.1 M phosphate-bufferedsaline pH 7.4 (PBS), incubated in 1–3% H2O2 in PBS for10 minutes to inactivate endogenous peroxidase, andrinsed again in PBS.
Single immunocytochemistry. All sections throughthe embryonic diencephalon and every third sectionthrough the diencephalon of postnatal rats were preincu-bated free-floating in PBS containing 10% normal horseserum (NHS) and 0.2–0.4% Triton X-100 to mask non-specific adsorption sites and increase the penetration ofantibodies. The sections were then incubated overnight at4°C in anti-VIM (DAKO, Glostrup, Denmark) and anti-GFAP (Boehringer Mannheim, Mannheim, Germany)monoclonal antibodies diluted 1:200 and 1:500, respec-tively, in PBS containing 1% NHS. After several rinses inPBS, the sections were incubated for 75 minutes in bio-tinylated horse anti-mouse IgG (Vector Laboratories, Bur-lingame, CA) diluted 1:200 in PBS/1% NHS. The sectionswere then rinsed in PBS, and incubated for 75 minutes inavidin-biotin peroxidase complex (ABC kit, Vector), di-luted 1:100 in PBS/1% NHS. A solution of 0.075% 3-3’-diaminobenzidine tetrahydrochloride (DAB; Sigma, St.Louis, MO) and 0.002% H2O2 in 0.05 M Tris-HCl buffer,pH 7.6, was used as chromogen. Finally, the sections weremounted onto gelatin-coated slides, dehydrated and cov-erslipped with DPX. To prevent tissue damage during airdrying, the sections from the embryonic specimens wereimmersed in a solution of 2% gelatin and 0.2% chrome-alum in PBS before mounting. In order to control thespecificity of the antibodies, series of sections were pro-cessed omitting the primary antibody or replacing it withnormal serum (diluted 1:100). No specific staining wasdetected in these sections. For the cytoarchitectonic anal-ysis in embryonic rats, immunoreacted sections were com-pared with standard series of 10-mm-thick sections fromparaffin-embedded brains of matching ages stained withthionin (0.1% in distilled water), previously prepared inour laboratory. In the material derived from postnatalanimals, the sections adjacent to those processed for im-
528 C. FRASSONI ET AL.

munocytochemistry were stained with thionin. The at-lases of Altman and Bayer (1995) and Paxinos et al. (1991)were used for the identification of the developing thalamicnuclei.
Double immunocytochemistry. In order to examinethe relationship between glia and migrating thalamic neu-rons, double immunoperoxidase and immunofluorescentlabeling were performed on free-floating sections obtainedfrom E16 embryos. To this purpose, antibodies againstg-aminobutyric acid (GABA) and calretinin (CR) wereused as neuronal markers, because both these moleculesare intensely expressed in young neurons during embry-onic life (Lauder et al., 1986; Frassoni et al., 1998). In thedouble immunoperoxidase experiments, sections wereprocessed for VIM detection following the procedure out-lined above. In the final step of the procedure, Vip sub-strate (Vector), which results in a purple reaction product,was used as chromogen. After several rinses in PBS, thesesections were incubated with 10% normal goat serum inPBS for 45 minutes and subsequently incubated for 16hours at 4°C with either rabbit polyclonal anti-GABAantibodies (Sigma-Aldrich, Milan, Italy) diluted 1:7,000,or anti-CR antibodies (Swant, Bellinzona, Switzerland)diluted 1:5,000 in PBS/1% NGS. The avidin-biotin protocol(ABC kit, Vector) was used in the subsequent steps of theimmunohistochemical procedure, and DAB, which resultsin a brown reaction product, was used as chromogen. Thesections were then mounted onto gelatin-coated slides,dehydrated, and coverslipped.
For double immunofluorescent labeling, free-floatingsections were first incubated for 30 minutes in PBS con-taining 1% bovine serum albumin and 0.2% Triton X-100,and then in a mixture of monoclonal VIM (1:200) andpolyclonal GABA (1:2,000) or CR (1:2,500) antibodies.These sections were subsequently incubated in a mixtureof lyssamine rhodamine sulphonyl chloride (LRSC)-conjugated goat anti-mouse IgG and fluorescein (FITC)-conjugated goat anti-rabbit antibodies (Jackson ImmunoResearch Lab, West Grove, PA), which were preabsorbedagainst the non-corresponding primary antibody. The sec-tions were then mounted in PBS-glycerol (1:2) and exam-ined by using a TCS NT confocal laser-scanning micro-scope (Leica Lasertecknik GmbH, Heidelberg, Germany)equipped with a 75-mW Krypton/Argon mixed gas laser.FITC and LRSC were excited with the 488 and 568 nmlines, respectively, imaged separately, and merged withLeica Power Scan software.
Electron microscopy
The ultrastructural analysis was performed at selecteddevelopmental stages in the thalamic region extendingfrom the reticular neuroepithelium to the pial surface fora total of 11 embryos (E14, E15, E17, E18). The internalcapsule (IC) and the reticular thalamic nucleus (Rt) wereexamined in 5 postnatal (P4, P8, P15) rats and one adult(P40) rat. The material was processed for immunoperoxi-dase as outlined above with anti-VIM and anti-GFAPantibodies. Triton X-100 was not used in the processing ofthis material and the Vibratome sections were permeabil-ized either with ethanol (10%, 25%, 10% in PB, 5 minuteseach) or by freeze-thawing according to Pinault et al.(1997). After completion of the immunoenzymatic proce-dure, the sections were washed in PB, postfixed for 15minutes in 2.5% glutaraldehyde in PB, washed in PB, andpostfixed for 1 hour in 1–2% OsO4. After dehydration, the
sections were cleared in propylene oxide, flat-embedded inEpon-Spurr between acetate foils (Aclar, Ted Pella, Red-ding, CA) and polymerized at 60°C for 36 hours. Embed-ded sections were then examined under a dissecting mi-croscope. Small areas of the embryonic thalamusextending from the reticular neuroepithelium to the pialsurface, and areas within the IC and Rt from thalamicpostnatal sections, were excised with razor blades andglued to cured resin blocks. Semithin (1 mm) sections werecut with a Reichert ultramicrotome and collected on glassslides without counterstaining. Ultrathin sections werecut from the surface, counterstained with lead citrate, andexamined with a Jeol T8 electron microscope.
Fluorescent dye labeling in brain slices
Migrating cells in living slices were labeled with the car-bocyanine dye DiI (1,1’-dioctadecyl-3,3,3’,3’-tetrametylindo-carbocyanine perchlorate; Molecular Probes, Inc., Eugene,OR), by using a published procedure (O’Rourke et al.,1992). Brain slices from E15 embryos were prepared fororganotypic culture (Stoppini et al., 1991; Roberts at al.,1993) and DiI crystals were then placed in the anteriorregion of the thalamic neuroepithelium. The slices wereincubated for 24 hours to allow the migration of labeledcells away from the injection site and then fixed in 4%paraformaldehyde in PB. The sections were then mountedin PBS-glycerol (1:2) and examined by using the TCS NTconfocal laser-scanning microscope.
RESULTS
Light and confocal microscopy
VIM immunoreactivity during prenatal development
Embyonic day 14. At this stage of development, theportion of diencephalic vescicle which gives rise to thethalamus was almost exclusively constituted by neuroep-ithelium clearly identifiable in the thionin-stained sec-tions (Fig. 1A). VIM immunoreactivity was evident in glialfibers extending radially from the margin of the ventricu-lar germinative zone to the pial surface of the diencephalicvescicle (Fig. 1B). The glial processes were grouped infascicles which followed nearly straight radial trajectories(Fig. 1B,C). The density of fibers was particularly highclose to the pial surface, where they appeared very tortu-ous (Fig. 1D). Very few fine longitudinal VIM-ir fiberswere also detected (Fig.1C). The intense immunolabelingat the pial surface prevented a clear visualization of theglial endfeet, which were instead identifiable in ultrathinsections (see the ultrastructural findings below). Also, thecell bodies of radial glia could be clearly identified only inthe electron micrographs.
Embryonic day 16. At E16, the neuroepithelium wasstill prominent and mitotically active but some thalamicnuclei, such as the ventrolateral nucleus and Rt, wereclearly detectable in the thionin-stained sections (Fig. 2A).The VIM-ir radial glial fibers arising from the ventriculargerminal zone coursed parallel through the dorsal tha-lamic primordium (Fig. 2B). These radial processes termi-nated with endfeet varicosities on the lateral surface,where some VIM-stained glial cells were also identified(Fig. 2C). In the ventricular region, glial fibers appearedaggregated in fascicles (Fig. 2D). At E16, VIM-ir glialfibers showed a peculiar arrangement in the lateral por-tion of the thalamus, where a set of fibers, running paral-
529RADIAL AND NON-RADIAL GLIA IN DEVELOPING THALAMUS

lel to the lateral brain surface, extended perpendicular tothe plane of the radial glial processes (Fig. 2B,E).
In double-labeled material examined in light and confo-cal microscopy, CR-ir young Rt neurons showing the mor-phology of migrating cells were associated with the VIM-ir
radial glial path ventral to the dorsal thalamic primor-dium (Fig. 3A). Following the migratory stream, someCR-ir cells were seen to reach the lateral border of thethalamus along the radial glia (Fig. 3B). In addition to theradial migration of cells, GABA immunolabeling showed
Fig. 1. Photomicrographs of mid-diencephalic coronal sections in therat at embryonic day (E)14. A portion of the diencephalic vescicle stainedwith thionin is shown at low power in A and vimentin (VIM) immuno-reactivity in the adjacent section is shown in B. C: At higher magni-fication, VIM immunoreactivity is clearly detected in radial fibers
grouped in fascicles (large arrowheads); a few fine longitudinal VIM-immunoreactive fibers are also evident (small arrowheads). D: Highermagnification photomicrograph showing VIM-immunoreactive thickfibers at the pial surface (arrowheads). 3V, third ventricle; Ne, neu-roepithelium. Scale bars 5 75 mm in A,B; 45 mm in C; 25 mm in D.
530 C. FRASSONI ET AL.

at this age neurons which were not radially oriented (Fig.3C–E), and climbed VIM-ir glial processes in a plane per-pendicular to radial glia (Fig. 3E). The net of mediolateral
and longitudinal glial fibers, first observed in immunoper-oxidase preparations (Fig. 2B,E), could thus be clearlyidentified in confocal microscopy (Fig. 3C–E).
Fig. 2. Photomicrographs of mid-diencephalic coronal sections inthe rat at embryonic day (E)16. A: Dorsal thalamic nuclei and theregion of migration of reticular thalamic neurons are shown in athionin-stained section. B: Vimentin (VIM) immunoreactivity is evi-dent in parallel radial glial fibers extending radially from the ventric-ular germinative zone to the lateral pial surface; note that in thelateral portion of the thalamus the fibers are arranged perpendicularto the radial glia fibers (arrowheads). C: VIM-immunoreactive fibers
contact the pial surface with endfeet (arrows). D: High magnificationof the ventricular region in which glial fibers are concentrated in astream of fascicles (arrowheads). E: Higher magnification of the areamarked by the asterisk in B, showing glial VIM-immunoreactive fibersperpendicular (arrows) to the plane of radial glial fibers (arrowheads).Hb, habenular complex; Ne, neuroepithelium; Rt, reticular thalamicnucleus; Rtm, reticular thalamic neuron migration; VL, ventrolateralthalamic nucleus. Scale bars 5 125 mm in A,B; 30 mm in C–E.
531RADIAL AND NON-RADIAL GLIA IN DEVELOPING THALAMUS

Figure 3
532 C. FRASSONI ET AL.

Embryonic day 19. At this age, most of the thalamicnuclei were detectable in thionin-stained sections (Fig.4A). The VIM immunoreactivity was still intense in radialglial fibers across the entire thalamus (Fig. 4B). However,the fibers perpendicular to the radial glia, seen at E16,were no longer detectable at E19. Some VIM-ir glial ele-ments were observed at the pial surface (Fig. 4C) and thecell bodies of radial glia were displaced from the ventric-ular zone to a more lateral location in the thalamus (Fig.4C,E). The fascicles of fibers appeared to be reduced inthickness, especially near the ventricular germinativezone, where a few fine strands of VIM-immunostainedprocesses were observed (Fig. 4D).
DiI labeling in vitro. Focal deposit of DiI crystals inE15 cultured brain slices resulted in brilliant diffuse fill-ing of the anterior region of the thalamic neuroepithelium.In every case, DiI-labeled cells were observed outside theinjection site. Numerous DiI-labeled neurons were seenafter 24 hours, packed along a radial migratory stream onthe lateral side of the injection site (Fig. 3F). In addition,vertically oriented brilliantly labeled neurons showing thecharacteristic morphology of migratory cells with fusiformshape and leading process, were clearly seen in the dorsalthalamic primordium (Fig. 3F).
Postnatal development. At birth, VIM immunoreac-tivity was much less evident in the thalamus than atprevious developmental stages, and regional differenceswere clearly visible (Fig. 5). In the intralaminar and mid-line nuclei, the intensity of immunolabeling had markedlydeclined in respect to prenatal stages. Along the thalamicmidline, VIM immunoreactivity was detected only in thinprocesses scattered through the paraventricular and cen-tral medial nuclei (Fig. 5A,C). In the dorsal and lateralthalamic domains, regional variation in glial immunola-beling was very obvious: VIM-ir fibers filled the laterodor-sal thalamic nucleus and immature astrocytes werestrongly VIM-positive (Fig. 5B,E), whereas less intenselylabeled fibers were scattered throughout the ventroposte-rior and ventral lateral geniculate nuclei (Fig. 5D,F).
The VIM staining pattern did not exhibit major changesin the thalamus during the first postnatal days (P0–P4).On the other hand, darkly stained star-shaped GFAP-ircells were seen by P4 in the internal medullary laminaand IC (Fig. 6A,B). At the end of the first postnatal week,VIM immunoreactivity further decreased (Fig. 6C),whereas GFAP immunoreactivity began to outline theboundaries of some thalamic nuclei, such as the borderbetween the ventral lateral and dorsal lateral geniculate(DLG) nuclei. In addition, GFAP-stained cells became ev-ident in the dorsal part of the lateroposterior nucleus (Fig.6D). A marked decrease of VIM immunoreactivity oc-curred in the thalamus during the second postnatal week.
In the adult thalamus, VIM staining was virtually ab-sent and a few immunolabeled elements were only seen inthe IC (Fig. 6E). The principal thalamic nuclei seemed inadulthood to be devoid also of GFAP immunoreactivity,with the exception of Rt. However, faintly GFAP-stainedcells and fine processes were detectable at the midline,especially in the thalamic paraventricular nucleus ( Fig.6G), as well as in the zones bordering other nuclei.GFAP-ir elements were seen in the IC in adulthood (Fig.6F).
Electron microscopy
The ultrastructural immunolocalization of glial anti-gens was investigated in embryos in the thalamic regionextending from the reticular protuberance of the neuroep-ithelium to the pial surface, and in postnatal and adultrats in the IC and Rt.
VIM immunoreactivity during prenatal development.
At all the developmental ages, VIM immunoreactivity wasfound in cell bodies, in radial glial processes, and in nu-merous enlarged endfeet at the pial surface. At E14, ovoidVIM-ir cells were detected in the thalamic neuroepithe-lium, whereas bipolar VIM-ir cell bodies were frequentlydistributed at all levels of the germinative zone, as well asclose to the pial surface among a dense row of labeled glialendfeet. The VIM-ir endfeet arose from radial glial fibersof different caliber, and they contained mainly clusters ofribosomes, mitochondria, and vacuoles; their attachmentto the pial surface appeared flat and sometimes consider-ably expanded (Fig. 7A).
VIM immunoreactivity was especially intense in theglial fibers of the thalamic primordium at all stages ofprenatal development (Fig. 7B–F). The VIM-ir radial glialprocesses became progressively thinner from E14 to E18and their cytoplasm was filled with polysomes, microtu-bules, vacuoles, and clear vesicles of different sizes (Fig.7B,C,E,F). The vesicles were sometimes grouped at thelevel of large appositions between VIM-ir processes andunlabeled cells (Fig. 7B). At E14, when large extracellularspaces were observed between radial glia and migratingcells, non-specialized small contacts were seen betweenVIM-ir fibers and VIM-immunonegative somatic protru-sions (Fig. 7C). Immunolabeled radial glial fibers weregenerally parallel and frequently found along both sides ofbipolar migrating cells (Fig. 7D). VIM-positive processesperpendicular to the radial glia were sometimes observedat E17 (Fig. 7E).
Finally, the ultrastructural analysis at E18 pointed outa considerable reduction of the extracellular spaces andthe appearance of several puncta adhaerentia betweenVIM-ir processes and neuronal cell bodies (Fig. 7F).
Fig. 3. Photomicrographs of double labeling experiments per-formed in embryonic thalamus at embryonic day (E)16 with immuno-peroxidase staining (A,B) or immunofluorescence examined with con-focal laser scanning microscopy (C–F), showing migrating thalamiccells along radial and non-radial glial fibers. A: Fusiform calretinin(CR)-immunoreactive neurons (brown, arrows) in the region of thereticular thalamic migration in association with radial vimentin(VIM)-immunoreactive glial fibers (purple, arrowheads). B: CR-immunoreactive cell (arrow) that reaches the lateral border of thethalamus (asterisk). C: Two vertically oriented g-aminobutyric acid(GABA)-immunoreactive cells (green, arrows): note the cell on theright side juxtaposed to a VIM-immunoreactive longitudinal glialfiber (red, arrowheads) D: On the left, a GABA-immunoreactive neu-ron (large arrow) along one longitudinal glial fiber; on the right, aradially oriented GABA-immunoreactive neuron (small arrow); VIM-immunoreactive fibers (thin arrowhead) orthogonal to the radial ones(large arrowhead) are evident. E: Illustrates the same GABAergic cellindicated by the asterisk in C; the image derives from different depthsin the section. F: Photomicrographs of 1,19-dioctadecyl-3,3,39,39-tetrametylindocarbocyanine perchlorate (DiI) labeling in E15 tha-lamic slice examined with confocal laser scanning microscopy, 24hafter DiI crystal placement in the anterior thalamic neuroepithelium:note the vertically oriented neurons (arrows) in a plane perpendicularto the radially migrating cells (asterisk). The orientation of the figureis defined in the upper right side (d, dorsal; l, lateral). Scale bars 5 7.5mm in A,B; 30 mm in D (also applies to C); 12 mm in E; 16 mm in F.
533RADIAL AND NON-RADIAL GLIA IN DEVELOPING THALAMUS
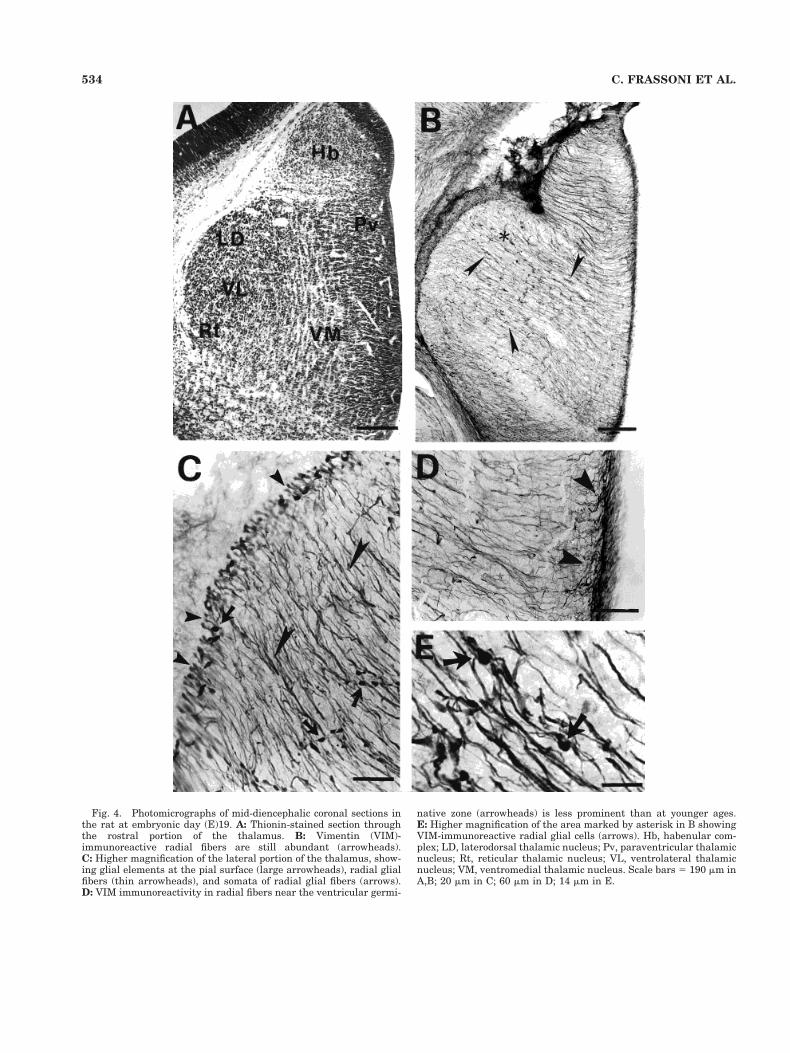
Fig. 4. Photomicrographs of mid-diencephalic coronal sections inthe rat at embryonic day (E)19. A: Thionin-stained section throughthe rostral portion of the thalamus. B: Vimentin (VIM)-immunoreactive radial fibers are still abundant (arrowheads).C: Higher magnification of the lateral portion of the thalamus, show-ing glial elements at the pial surface (large arrowheads), radial glialfibers (thin arrowheads), and somata of radial glial fibers (arrows).D: VIM immunoreactivity in radial fibers near the ventricular germi-
native zone (arrowheads) is less prominent than at younger ages.E: Higher magnification of the area marked by asterisk in B showingVIM-immunoreactive radial glial cells (arrows). Hb, habenular com-plex; LD, laterodorsal thalamic nucleus; Pv, paraventricular thalamicnucleus; Rt, reticular thalamic nucleus; VL, ventrolateral thalamicnucleus; VM, ventromedial thalamic nucleus. Scale bars 5 190 mm inA,B; 20 mm in C; 60 mm in D; 14 mm in E.
534 C. FRASSONI ET AL.
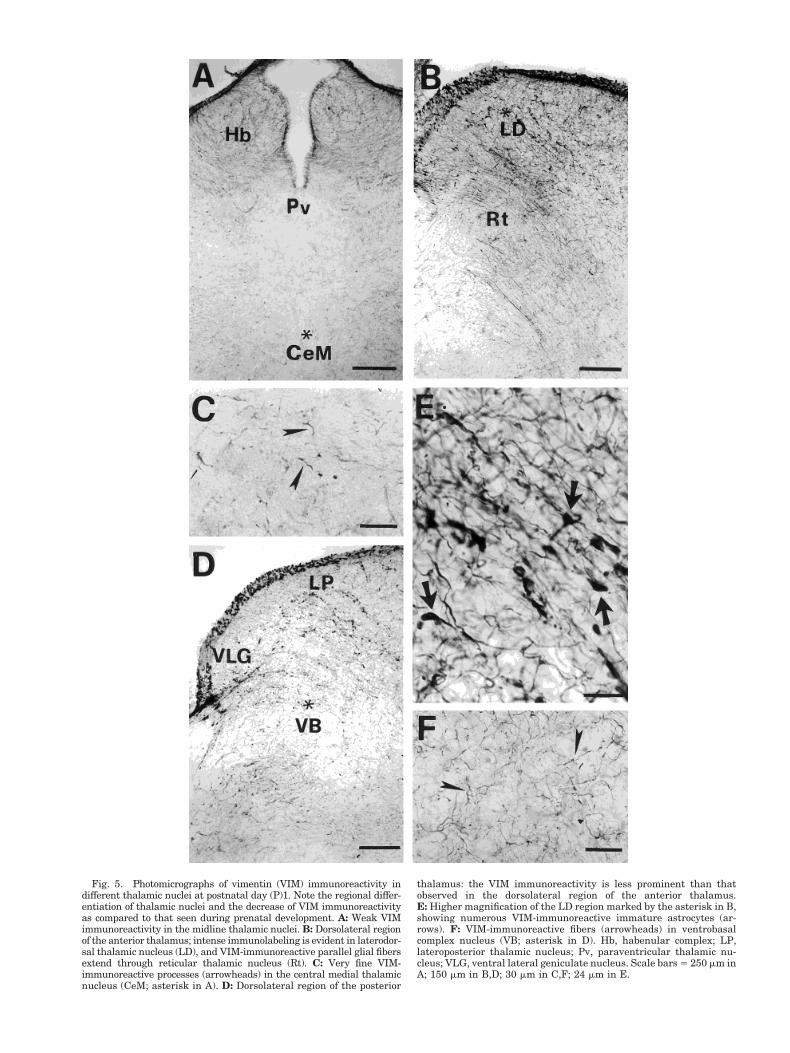
Fig. 5. Photomicrographs of vimentin (VIM) immunoreactivity indifferent thalamic nuclei at postnatal day (P)1. Note the regional differ-entiation of thalamic nuclei and the decrease of VIM immunoreactivityas compared to that seen during prenatal development. A: Weak VIMimmunoreactivity in the midline thalamic nuclei. B: Dorsolateral regionof the anterior thalamus; intense immunolabeling is evident in laterodor-sal thalamic nucleus (LD), and VIM-immunoreactive parallel glial fibersextend through reticular thalamic nucleus (Rt). C: Very fine VIM-immunoreactive processes (arrowheads) in the central medial thalamicnucleus (CeM; asterisk in A). D: Dorsolateral region of the posterior
thalamus: the VIM immunoreactivity is less prominent than thatobserved in the dorsolateral region of the anterior thalamus.E: Higher magnification of the LD region marked by the asterisk in B,showing numerous VIM-immunoreactive immature astrocytes (ar-rows). F: VIM-immunoreactive fibers (arrowheads) in ventrobasalcomplex nucleus (VB; asterisk in D). Hb, habenular complex; LP,lateroposterior thalamic nucleus; Pv, paraventricular thalamic nu-cleus; VLG, ventral lateral geniculate nucleus. Scale bars 5 250 mm inA; 150 mm in B,D; 30 mm in C,F; 24 mm in E.

Fig. 6. Photomicrographs of glial fibrillary acidic protein (GFAP;A,B,D,F,G) and vimentin (VIM; C,E) immunoreactivities in the thal-amus at different postnatal stages. A,B: Postnatal day (P)4; darklyGFAP-stained star-shaped astrocytes in the internal medullary lam-ina (A) and internal capsule (IC; B). C,D: P8; mid-diencephalic coronalsections show VIM (C) and GFAP (D) immunoreactivity. At this age,VIM immunoreactivity is intense only in IC, where GFAP immuno-labeling is also detectable; in addition, GFAP immunoreactivity out-
lines some thalamic nuclei (arrowheads). E–G: P40; a few VIM-immunoreactive elements can be seen in IC (E); GFAPimmunoreactivity is evident in the IC and in reticular thalamic nu-cleus (Rt; F) and fine GFAP-immunoreactive processes are seen in theparaventricular thalamic nucleus (G). DLG, dorsal lateral geniculate;iml, internal medullary lamina; LP, lateroposterior thalamic nucleus;VB, ventrobasal complex; VLG, ventral lateral geniculate nucleus.Scale bars 5 250 mm in A; 50 mm in B,F,G; 500 mm in C,D.
536 C. FRASSONI ET AL.

VIM and GFAP immunoreactivities during postnatal
development. The distribution of VIM-ir structureschanged after birth due to the appearance of mature as-trocytes and the lack of radially oriented glial processes.VIM immunoreactivity was mainly observed in fine pro-cesses surrounding the blood vessel walls (Fig. 8A) and instar-shaped cell bodies rich in organelles (Fig. 8B). Onlyafter P15 VIM immunoreactivity was associated with bun-dles of intermediate filaments (Fig. 8C). At the ultrastruc-tural level GFAP immunoreactivity showed a distributionsimilar to that described for VIM (Fig. 8D).
DISCUSSION
This study provides a detailed account of the morpho-logical and topographical organization of glial fibers in thethalamus during embryonic and postnatal development.In addition, our findings point out novel data on neuronalmigration phenomena in thalamic structures. At thesesites, we show that migrating cells follow both radial andnon-radial pathways toward their final destination, asunequivocally confirmed by the direct observation of DiI-labeled migrating cells in living brain slices. The electronmicroscopic study allowed the examination of the relation-ships between neurons and glial processes revealing thecellular milieu in which neurons migrate.
Radial glia and neuronal migration
In the present investigation, the double CR and VIMimmunolabeling provided evidence that early generatedthalamic cells, which arise from the reticular neuroepithe-lium (Altman and Bayer, 1988a,b), reach their final des-tination in association with glial fibers. In a previousstudy, we demonstrated that CR immunoreactivity re-veals the migratory pattern of the early generated Rtneurons (Frassoni et al., 1998). Thus, the present dataindicate that cells of the Rt migratory stream, exhibitinglong leading and trailing processes and ellipsoid shape,climbed along the radial glial fibers toward the lateralsurface of the thalamus. A similar pattern of lateral mi-gration and postmigratory destination was described inthe hamster thalamus, where neurons identified by spe-cific neuropeptide immunoreactivity were seen to migrateto their final destination along the zona limitans, at theborder between the dorsal and ventral thalamus, in asso-ciation with radial glia (Botchkina and Morin, 1995).
The intimate relationship between neurons and radialglial processes was previously investigated in microculturesystems (Edmonson and Hatten, 1987; Hatten, 1990,1999).These studies provided a detailed analysis of the cellularinteractions and molecular basis of glial-guided neuronalmigration in vitro. The present double labeling experiments,based on immunoperoxidase staining or immunofluores-cence using confocal laser scanning microscopy, revealedneurons migrating in tight association with VIM-ir fibers,and these neurons showed features, ellipsoid shape andleading process, similar to those described in culture systemsand observed also in our study on brain slices.
Non-radial migration
Although the long processes of radial glia support andguide the migration of a large proportion of neurons, it hasbeen established in recent years that tangential migrationrepresents an important developmental feature in the ro-dent cerebellum and olfactory bulb (Sotelo et al., 1990;
Wichterle et al., 1997; Peretto et al., 1998). Non-radialpathways of cell migration also have been described indistinct brain regions of different species (Martinez et al.,1992; Rakic, 1995b; Golden et al., 1997). Tangential mi-gration also plays a crucial role in the normal corticaldevelopment, contributing to the dispersion of clonallyrelated cells, although the substrate of this type of migra-tion is still unclear (O’Rourke et al., 1995, 1997; Rakic,1995b; Tamamaki et al., 1997; Pearlman et al., 1998;Seong-Seng Tan et al., 1998). In the lateral cortex, thepresence of non-radially oriented glial fibers has beensuggested (Misson et al., 1991; O’Rourke et al., 1995,1997), and axonal processes have been supposed to pro-vide a substrate for non-radial migration in other brainregions (Gray et al., 1990; Kawano et al. 1995, Rakic,1995b). It has also been suggested that cell adhesion mol-ecules responsible for neuronal development are involvedin tangential migration (Ono et al., 1994; Hu et al., 1996;Ohyama et al., 1998).
The present findings revealed that thalamic neuronsfollow both radial and non-radial pathways. In particular,cells identified in our experiments as GABAergic followednon-radial trajectories. In the neocortex, the migratingcells moving along a non-radial trail have been suggestedto represent GABAergic interneurons arising from theventricular and subventricular zones. In addition, early-born neurons invade the neocortical marginal zone arisingfrom the lateral ganglionic eminence and following a tan-gential route (De Diego et al., 1994; De Carlos et al., 1996;Tamamaki et al., 1997; Meyer et al., 1998; Lavdas et al.,1999). Our results indicate instead that in the embryonicthalamus, some migrating neurons climb glial fibers or-thogonal to the radial ones, pointing out that both radialand orthogonal migrating thalamic neurons are linkedwith a glial system. No evidence of such organization hasbeen reported in the cortex, in which the cohort of tangen-tially migrating cells is apparently unrelated to glialguidelines. We observed that radial fibers extended fromthe ventricular surface to the lateral border of the thala-mus, whereas the origin and termination of the longitudi-nally oriented glial fibers could not be identified in ourmaterial. Other substrates, such as axonal processes orextracellular matrix components, may serve non-radialmigration in the thalamus, and the full range of theseevents require further clarifications.
Because in the present study non-radial migrating neu-rons were recognized with an anti-GABA serum, we can-not rule out that other unidentified non-GABAergic neu-rons could also climb non-radial glial fibers. Theoccurrence of non-radially migrating thalamic GABAergiccells raises questions on their origin and final destination.In the adult rat thalamus, GABAergic interneurons ac-count for less than 1% of the total thalamic cell populationwith the exception of Rt and DLG nuclei (for review, seeArcelli et al., 1997). During development, Rt and DLG arelocated outside the trajectory of orthogonal fibers, indicat-ing that the non-radial migrating GABAegic cells may notbelong to the cohort of interneurons destined for thesenuclei. They could therefore represent a neuronal popula-tion expressing GABA only transiently during migration.Alternatively, these cells could represent prospective in-terneurons, which are then dispersed in a low proportionwithin the adult thalamus outside Rt and DLG.
It is interesting to note that orthogonal glial fibers wereclearly detected in a more restricted time window, i.e., be-
537RADIAL AND NON-RADIAL GLIA IN DEVELOPING THALAMUS

Figure 7
538 C. FRASSONI ET AL.

tween E15 and E18, than the radial ones. The cells of theanterior, intralaminar, and midline structures are generatedduring this embryonic period (Altman and Bayer, 1988b).Tangential fibers could therefore contribute to the migrationof neuroblasts that colonize these nuclear groups, but theseevents remain to be fully elucidated.
Ultrastructural features
Our ultrastructural immunolocalization of VIM in thedeveloping thalamus allowed the analysis of radial glialfibers and their mutual interrelations, of VIM-ir endfeetat the pial surface, and of the sites of apposition betweenVIM-ir glial processes and neuronal migrating cells.
The occurrence of VIM-ir fibers perpendicular to theVIM-ir radial glia at early stages of prenatal developmentwas also documented in electron microscopy. The mostfrequent organelles consistently detected in the VIM-irfibers were clear vesicles of different sizes and microtu-bules. The presence of vesicles in glial processes, reportedin several in vitro and in vivo studies (Gregory et al., 1988;
Fig. 8. Electron micrographs of vimentin (VIM)-immunoreactive(A–C) and glial fibrillary acidic protein (GFAP)-immunoreactive (D)structures in the internal capsule of postnatal rats. A: Postnatal day(P)4; a VIM-immunoreactive glial profile in close apposition to anendothelial cell (E) of a blood vessel. B: P15; a VIM-immunoreactive
astrocyte rich in organelles extends its processes (arrows) betweenmyelinated fibers (f). C: P15; a glial process containing a cross-sectioned bundle of VIM-immunoreactive intermediate filaments (ar-rowheads). D: P8; the cell body of a GFAP-immunoreactive astrocyte.N, nucleus. Scale bars 5 0.5 mm in A,C; 1 mm in B,D.
Fig. 7. Electron micrographs showing the distribution of vimentin(VIM) immunoreactivity in some areas of the dorsal thalamic primor-dium at different prenatal stages. A: Embryonic day (E)14; at the pialsurface small (S) and large (L) VIM-immunoreactive end feet contact(arrows) the basal membrane (BM) without any junctional specializa-tion. Radial VIM-immunoreactive fibers, containing several vacuoles(asterisks), are sometimes juxtaposed (arrowheads). B: E14; a VIM-immunoreactive process contains several clear vesicles of differentsizes (arrowhead) grouped at the site of its close apposition with anunlabeled cell body (U). C: E14; a small contact (arrow) between avesicle-rich VIM-immunoreactive fiber (F) and an unlabeled somaticprotrusion (SP). D: E15; two heavily VIM-labeled radial processesfollow the lateral surfaces of an unlabeled cell body (CB). N, nucleus.E: E17; two VIM-immunoreactive fibers perpendicular to each othercontain several microtubules (arrowheads); the vertical process alsocontains a large group of vacuoles and vesicles (arrow). F: E18; aVIM-immunoreactive profile rich in vacuoles (asterisks) connects withtwo unlabeled cells (U) by means of puncta adhaerentia (arrows). N,nuclei. Scale bars 5 0.5 mm in A–C,F; 1 mm in D,E.
539RADIAL AND NON-RADIAL GLIA IN DEVELOPING THALAMUS

Mason et al.,1988; Gadisseux et al., 1990; Yuasa et al.,1996), was correlated with the neuron-glia interactionsoccurring during contact guidance in migration (Yuasa etal., 1996). On the other hand, our study provided demon-stration in vivo of a large amount of microtubules in radialglial fibers, in agreement with in vitro studies that haveunderlined the important role of this cytoskeletal compo-nent during migration (Gregory et al., 1988; Cameron andRakic, 1994).
The present observations also provided data on the ul-trastructure of radial glial endfeet during prenatal devel-opment. In embryos, the endfeet contained several or-ganelles, such as ribosomes, mitochondria, and vacuoles,but they were not closely packed one to the other, andtheir membranes were infrequently juxtaposed and didnot show any specialization. In contrast, the endfeet ofmature astrocytes were generally organized in a compactstack of expansions rich in gliofilaments and forming junc-tions between them and with the basal lamina of the pialsurface, thus supporting previous findings (Nakazawa andIshikawa, 1998). Moreover, in embryos the glial contactwith the pial surface was devoid of the undercoats ofelectron-dense material typical of the mature astrocyticendfeet (Nakazawa and Ishikawa, 1998).
The present ultrastructural investigation showed atE14 a few non-specialized contacts between radial VIM-irfibers and VIM-immunonegative neuronal cell bodies. AtE18, these contacts where no longer evident, but numer-ous puncta adhaerentia interconnected VIM-ir processesto adjacent neurons. The ultrastructure of neuroglial in-terrelations and of the neuronal and glial sides of closeappositions, defined as “interstitial” or “migration” junc-tions, have been repeatedly investigated (Gregory et al.,1988; Mason et al., 1988; Gasser and Hatten, 1990; Hat-ten, 1990; Fishell and Hatten, 1991; Cameron and Rakic,1994; O’Rourke et al., 1995; Kawano et al., 1995; Anton etal., 1996). These dynamic junctions were described in cul-ture (Gregory et al., 1988; Fishell and Hatten,1991), butwere not identified in vivo (Gadisseux et al., 1990; Kawanoet al., 1995; Yuasa et al., 1996). Following the block ofmigration with antibodies against membrane components,several puncta adhaerentia were found to appear betweenglial fibers and neurons (Fishell and Hatten, 1991; Antonet al., 1996). The membrane associations observed in ourmaterial at E14 could represent migration junctions, be-cause they were detected only at the time of migratoryevents, defined by Altman and Bayer (1988b). However, inour in vivo analysis, these neuroglial contacts appearedshorter and less numerous than those previously de-scribed in vitro (Gregory et al., 1988). Technical problemsin handling embryonic tissue for ultrastructural investi-gation could account for such differences. On the otherhand, the high number of neuroglial puncta adhaerentiawe observed at E18 could correlate temporally with theend of migration defined by Altman and Bayer (1988b).
The VIM/GFAP shift of expression
VIM immunoreactivity was found to be transient in thedeveloping thalamus, because VIM was expressed at earlyembryonic stages and almost disappeared by the end ofthe first postnatal week. The appearance of radial glia,identified by VIM immunolabeling, fits the time course ofthalamic neurogenesis, and we found that the transforma-tion of radial glial cells in astrocytic form was initiated inthe rat thalamus at E19, when neurogenesis is completed
in most of the dorsal thalamic nuclei (Altman and Bayer,1979, 1988b). Our results indicate that the VIM/GFAPtransition occurred when the migration was complete andradial glial fibers were no longer required for the guidanceof neurons. In mammals, the elimination of radial glia isparalleled by the appearance of GFAP-positive astrocytes(Levitt and Rakic, 1980; Misson et al., 1991; Stichel et al.,1991). In particular, in rodents, the involution of the ra-dial fiber system and the development of astrocytes occurpostnatally (Pixley and DeVellis, 1984; Misson et al.,1988, 1991; Stichel et al., 1991).
Regional differences in VIM immunoreactivity werefound in the rat thalamic nuclei at birth. In the intralami-nar, midline, and principal nuclei, the immunolabelinghad markedly decreased, whereas in the lateral region ofthe dorsal thalamus, VIM immunoreactivity was stillprominent at birth and was associated with immatureastrocytes. Star-shaped GFAP-ir astrocytes appearedaround birth in the IC, whereas, in agreement with aprevious study (Kalman and Hajos, 1989), the main tha-lamic nuclei of the adult rat were poor in GFAP-labeledelements, with the exception of Rt. Thus, the presentfindings point out a distinct developmental program indifferent thalamic domains. Altogether, our data supportthe general idea that, as in other CNS regions (Hutchinsand Casagrande, 1989; Misson et al., 1991), extensiveremodeling of VIM-ir glial cells constitutes a key event inthe normal maturation of the thalamus.
ACKNOWLEDGMENTS
The authors are deeply grateful to Prof. Marina Ben-tivoglio for her help and suggestions in the preparation ofthe manuscript. Many thanks are also due to Prof. SilviaDe Biasi for her comments on the electron microscopicdata, to Prof. Laura Vitellaro-Zuccarello for her help inconfocal microscopy, and to Dr. Mariateresa Selvaggio forher contribution to the early phase of this study. Theauthors also thank Dr. Alfonso Fairen for his help insetting up the technique of organotypic cultures of embry-onic nervous tissue, and Dr. Javier Morante-Oria for tech-nical support.
LITERATURE CITED
Altman J, Bayer SA 1979. Development of the diencephalon in the rat. VI.Re-evalutation of the embryonic development of the thalamus on thebasis of thymidine-radiographic datings. J Comp Neurol 188:501–524.
Altman J, Bayer SA. 1988a. Development of the rat thalamus: III. Timeand site of origin and settling pattern of neurons of the reticularnucleus. J Comp Neurol 275:406–428.
Altman J, Bayer SA.1988b. Development of the rat thalamus: I. Mosaicorganization of the thalamic neuroepithelium. J Comp Neurol 275:346–377.
Altman J, Bayer SA. 1995. Atlas of prenatal rat brain development. BocaRaton, FL: CRC Press.
Anton ES, Cameron RS, Rakic P. 1996. Role of neuron-glial junctionaldomain proteins in the maintenance and termination of neuronal mi-gration across the embryonic cerebral wall. J Neurosci 16:2283–2293.
Arcelli P, Frassoni C, Regondi MC, De Biasi S, Spreafico R. 1997. GABAer-gic neurons in mammalian thalamus: a marker of thalamic complexity?Brain Res Bull 42:27–37.
Bignami A, Raju T, Dahl D. 1982. Localization of vimentin, the non-specificintermediate filament protein, in embryonal glia and in early differen-tiating neurons. Dev Biol 91:286–295.
Botchkina GI, Morin LP. 1995. Specialized neuronal and glial contribu-tions to development of the hamster lateral geniculate complex andcircadian visual system. J Neurosci 15:190–201.
540 C. FRASSONI ET AL.

Cameron RS, Rakic P. 1994. Identification of membrane proteins thatcomprise the plasmalemmal junction between migrating neurons andradial glial cells. J Neurosci 14:3139–3155.
Choi BH. 1986. Glial fibrillary acid protein in radial glia of early humanfetal cerebrum: a light and electron microscopic immunoperoxidasestudy. J Neuropathol Exp Neurol 45:408–418.
Culican M, Baumrind N, Yamamoto M, Pearlman A. 1990. Cortical radialglia: identification in tissue culture and evidence for their transforma-tion into astrocytes. J Neurosci 10:684–692.
De Carlos JA, Lopez-Mascaraque L, Valverde F. 1996. Dynamic of cellmigration from the lateral eminence in the rat. J Neurosci 16:6146–6156.
De Diego I, Smith-Fernandez A, Fairen A. 1994. Cortical cells that migratebeyond area boundaries: characterization of an early neuronal popula-tion in the lower intermediate zone of prenatal rats. Eur J Neurosci6:983–997.
Dhal D, Rueger DC, Bignami A. 1981. Vimentin, the 57000 molecularweight protein of fibroblast filaments, is major cytoscheleton compo-nent in immature glia. Eur J Cell Biol 24:191–196.
Earle KL, Mitrofanis J. 1998. Development of glia and blood vessels in theinternal capsule of the rats. J Neurocytol 27:127–139.
Edmondson JC, Hatten ME. 1987. Glial-guided granule neuron migrationin vitro: a high-resolution time-lapse video microscopic study. J Neu-rosci 7:1928–1934.
Elmquist JK, Swanson JJ, Sakaguchi DS, Ross LR, Jacobson CD. 1994.Development distribution of GFAP and vimentin in Brazilian opossumbrain. J Comp Neurol 344:283–296.
Fishell G, Hatten ME. 1991. Astrotactin provides a receptor system forCNS neuronal migration. Development 112:755–765.
Frassoni C, Arcelli P, Selvaggio MT, Spreafico R. 1998. Calretinin immu-noreactivity in the developing thalamus of the rat: a marker of earlygenerated thalamic cells. Neuroscience 83:1203–1219.
Gadisseux JF, Kadhim HJ, van den Boch de Aguilar P, Caviness VS,Evrard P. 1990. Neuron migration within the radial glial fiber systemof the developing murine cerebrum: an electron microscopic autoradio-graphic analysis. Brain Res Dev Brain Res 52:39–56.
Gasser UE, Hatten ME. 1990. Neuron-glia interactions of art hippocampalcells in vitro: glail-guided neuronal migration and neronal regulation ofglial differentiation. J Neurosci 10:1276–1285.
Golden JA, Zitz JC, McFadden K, Cepko CL. 1997. Cell migration in thedeveloping chick diencephalon. Development 124: 3525–3533.
Gray GE, Leber SM, Sanes JR. 1990. Migratory patterns of clonally relatedcells in the developing central nervous system. Experimentia 46:929–940.
Gregory WA, Edmondson JC, Hatten ME, Mason CA. 1988. Cytology andneuron-glial apposition of migrating cerebellar granule cells in vitro.J Neurosci 8:1728–1738.
Hatten ME. 1990. Riding the glial monorail: a common mechanism forglial-guided neuronal migration in different regions of the developingmammalian brain. Trends Neurosci 13:179–188.
Hatten ME. 1999. Central nervous system neuronal migration. Ann RevNeurosci 22:511–539.
Hu H, Tomasiewicz H, Magnuson, Rutishauser U. 1996. The role of poly-sialic acid in migration of olfactory bulb interneuron precursors in thesubventricular zone. Neuron 16:735–743.
Hutchins JB, Casagrande VA. 1989. Vimentin: change in distributionduring brain development. Glia 2:55–66.
Kalman M, Hajos F. 1989. Distribution of glial fibrillary acidic protein(GFAP)-immunoreactive astrocytes in the rat brain. Exp Brain Res78:147–163.
Kawano H, Ohyama K, Kawamura K, Nagatsu I. 1995. Migration ofdopaminergic neurons in the embryonic mesencephalon of mice. BrainRes Dev Brain Res 86:101–113.
Lauder JM, Han UKM, Henderson P,Verdoon T, Towle AC. 1986. Prenatalontogeny of the GABAergic system in the rat brain: an immunocyto-chemical study. Neurosci 19:465–493.
Lavdas AA, Grigoriou M, Pachnis V, Parnavelas JG. 1999. The medialganglionic eminence gives rise to a population of early neurons in thedeveloping cerebral cortex. J Neurosci 15:7881–7888.
Levitt P, Rakic P. 1980. Immunoperoxidase localization of glial fibrillaryacidic protein in radial glial cells and astrocytes of the developingrhesus monkey brain. J Comp Neurol 193:815–840.
Martinez S, Puelles L, Alvarado-Mallart RM. 1992. Tangential neuronal
migration in the avian tectum: cell type identification and mapping ofregional differences with quail/chick homotopic transplants. Brain ResDev Brain Res 66: 153–163.
Mason CA, Edmondson JC, Hatten ME. 1988. The extending astroglialprocesses: development of glial cell shape. The growing tip and inter-actions with neurons. J Neurosci 8:3124–3134.
McDermott KW, Lantos PL. 1989 The distribution of glial fibrillary acidprotein and vimentin in postnatal marmoset (Callithrix jacculus)brain. Brain Res Dev Brain Res 45:169–177.
Meyer G, Soria JM, Martınez-Galan JR, Martın-Clemente B, Fairen A.1998. Different origins and developmental history of transient neuronsin the marginal zone of the fetal and neonatal rat cortex. J CompNeurol 397:493–518.
Missler M, Eins S, Bottcher H, Wolff J. 1994. Postnatal development ofglial fibrillary acidic protein in monkey visual cortex: evidence for atransient reduction of GFAP immunoreactivity. Brain Res Dev BrainRes 82:103–117.
Misson JP, Edwards M, Yamamoto M, Caviness Jr. 1988. Identification ofradial glial cells within the developing murine central nervous system:Studies based on a new immunohistochemical marker. Brain Res DevBrain Res 44:95–108.
Misson JP, Takahashi T, Caviness VS Jr. 1991. Ontogeny of radial andastroglial cells in murine cerebral cortex. Glia 4:138–148.
Mitrofanis J, Baker GE. 1993. Development of the thalamic reticular andperireticular nuclei in rats and their relationship to the course ofgrowing corticofugal and corticopetal axons. J Comp Neurol 388:575–587.
Mitrofanis J, Earle KL, Reese BE. 1997. Glial organization and chondroitinsulfate proteoglycan expression in the developing thalamus. J Neuro-cytol 26:83–100.
Nakazawa E, Ishikawa H. 1998. Ultrastructural observations of astrocyteendfeet in the rat central nervous system. J Neurocytol 27:431–440.
Ohyama K, Kawano H, Asou H, Fukuda T, Oohira A, Uyemura K,Kawamura K. 1998. Coordinate expression of L1 and 6B4 proteoglycan/phosphacan is correlated with the migration of mesencephalic dopami-nergic neurons in mice. Brain Res Dev Brain Res 107:219–226.
Ono K, Tomasiewicz H, Magnuson T, Rutishauser U. 1994. N-CAM muta-tion inhibits tangential migration and is phenocopied by enzymaticremoval of polysialic acid. Neuron 13:595–609.
O’Rourke NA, Dailley ME, Smith SJ, McConnell SK. 1992. Diverse migra-tory pathways in the developing cerebral cortex. Science 258:299–302.
O’Rourke NA, Sullivan DP, Kaznowski CE, Jacobs AA, McConnell SK.1995. Tangential migration of neurons in the developing cerebral cor-tex. Development 121:2165–2176.
O’Rourke N, Chenn A, McConnell SK. 1997. Postmitotic neurons migratetangentially in the cortical ventricular zone. Development 124:997–1005.
Paxinos G, Turk I, Teott LH, Velentino KL. 1991. Atlas of the developingrat brain. New York: Academic Press.
Pinault D, Smith V, Dechene M. 1997. Dendrodentritic and axoaxonicsynapses in the thalamic reticular nucleus of adult rat. J Neurosci17:3215–3233.
Pearlman AL, Phyllis LF, Hatten ME, Brunstrom JE. 1998. New directionfor neuronal migration. Curr Opin Neurobiol 8:45–54.
Peretto P, Bonfanti L, Merighi A., Fasolo A. 1998. Carnosine-like immu-noreactivity in astrocytes of glial tubes and in newly-generated cellswithin the tangential part of rostral migratory stream of rodents.Neuroscience 85:527–542.
Pixley SR, DeVellis J. 1984. Transition between immature radial glia andmature astrocytes studied with monoclonal antibody to vimentin.Brain Res Dev Brain Res 15:201–209.
Pulido-Caballero J, Jimenez-Sampedro F, Echevarria-Aza D, Martınez-Millan L. 1994. Postnatal development of vimentin-positive cells in therabbit superior colliculus. J Comp Neurol 343:102–112.
Rakic P. 1995a. Radial glial cells: scaffolding for brain construction. In:Ketterman H, Ransom BR, editors. Neuroglia. New York: Oxford Uni-versity Press. p 746–762.
Rakic P. 1995b. Radial versus tangential migration of neuronal clones inthe developing cerebral cortex. Proc Natl Acad Sci 92:11323–11327.
Regondi MC, Jaranowska A, Amadeo A, Ortino B, De Biasi S, Spreafico R,Frassoni C. 1999. Topographical glial organization in developing dorsalthalamus: an immunocytochemical and ultrastructural study. NeurosciLett Suppl 52:S43.
541RADIAL AND NON-RADIAL GLIA IN DEVELOPING THALAMUS

Roberts JS, O’Rourke NA, McConnell SK. 1993. Cell migration in culturedcerebral cortical slices. Dev Biol 155:396–408.
Seong-Seng T, Kalloniatis M, Strurm K, Tam PL, Reese BE, Faulkner-Jones B. 1998. Separate progenitors for radial and tangential celldispersion during development of the cerebral neocortex. Neuron 21:295–304.
Sotelo C, Alvarado-Mallart RM, Gardette R, Crepel F. 1990. Fate of graftedembryonic Purkinije cells in the cerebellum of the adult “Purkinje celldegeneration” mutant mouse. I. Development of reciprocal greft-hostinteractions. J Comp Neurol 295:165–87.
Stichel CC, Muller CM, Zilles K. 1991. Distribution of glial fibrillary acidicprotein and vimentin immunoreactivity during rat visual cortex devel-opment. J Neurocytol 20:97–108.
Stoppini L, Buchs PA, Muller D. 1991. A simple method for organotypiccultures of nervous tissue. J Neurosci Methods 37:173–182.
Tamamaki N, Fujimori KE, Takauji R. 1997. Origin and route of tangen-tially migrating neurons in the developing neocortical intermediatezone. J Neurosci 17:8313–23.
Voigt T. 1989. Development of glial cells in the cerebral wall of ferrets:direct tracing of their transformation from radial glia into astrocytes.J Comp Neurol 289:74–88.
Wichterle H, Garcia-Verdugo JM, Alvarez-Buylla A. 1997. Direct evidence forhomotypic, glia-independent neuronal migration. Neuron 18:779–791.
Yuasa S, Kawamura K, Kuwano R, Ono K. 1996. Neuron-glia interrela-tions during migration of Purkinje cells in the mouse embryonic cere-bellum. Int J Dev Neurosci 14:429–438.
542 C. FRASSONI ET AL.
Copyright © 2022 FDOKUMEN




















