mTORC1-mediated translational elongation limits intestinal tumour initiation and growth
Upload
independentCategory
view
1download
0
GC
MLAa
g1b
tG
Amcinttmtmsptotstaatwtktsva
*3EAm41tC7(peca1mPkpk
Neuroscience 141 (2006) 1389–1398
0d
LUTAMATE-DEPENDENT TRANSLATIONAL REGULATION IN
ULTURED BERGMANN GLIA CELLS: INVOLVEMENT OF p70S6KfcE
Km
EsG(GaamlmagIaic((G(m(ts
rgamaimoctissufc2
. E. GONZÁLEZ-MEJIA,a M. MORALES,a
. C. R. HERNÁNDEZ-KELLY,a R. C. ZEPEDA,a
. BERNABÉa,b AND A. ORTEGAa*
Departamento de Genética y Biología Molecular, Centro de Investi-ación y de Estudios Avanzados Unidad Zacatenco, Apartado Postal4-740, México DF 0300, Mexico
Unidad Académica de Ciencias Químico-Biológicas, Universidad Au-ónoma de Guerrero, Avenue Lázaro Cárdenas s/n Chilpancingo,uerrero, 39090, Mexico
bstract—Glutamate, the main excitatory amino acid trans-itter in the vertebrate brain is involved in the dynamic
hanges in protein repertoire that underlie synaptic plastic-ty. Activity-dependent differential expression patterns occurot only in neurons but also in glial cells. In fact, a membraneo nuclei signaling has been described after ionotropic glu-amate receptor stimulation in cultured chick cerebellar Berg-ann glia cells. In order to characterize other levels of pro-
ein expression regulation, we explored the effect of gluta-ate treatment in [35S]-methionine incorporation into newly
ynthesized polypeptides. A time-dependent modification inrotein synthesis was found. An important component ofranslational control is the ribosomal S6 protein kinase. Thre-nine phosphorylation renders the kinase active increasingranslation initiation. Glutamate exposure results in ribo-omal S6 protein kinase Thr389 phosphorylation in a dose andime-dependent manner that matches perfectly with the over-ll protein synthesis profile detected upon the excitatorymino acid. Pharmacological characterization of the recep-ors involved suggests the participation of both ionotropic asell as metabotropic glutamate receptors. The non-receptor
yrosine kinase Src, phosphatidylinositol 3-kinase, proteininase B and the mammalian target of rapamycin are media-ors of the glutamate effect. These results not only demon-trate that glutamate receptors activation is critically in-olved in translational control in glial cells adjacent to syn-ptic processes like cerebellar Bergmann glia cells, but also
Corresponding author. Tel: �525-55061-3769; fax: �1-525-55061-800x5317.-mail address: [email protected] (A. Ortega).bbreviations: Akt I, 1L-6-hydroxymethyl-chiro-inositol 2-(R)-2-O-ethyl-3-O-octadecylcarbonate; AMPA, amino �-3-hydroxy-5-methyl--isoaxazolepropionate; ANOVA, analysis of variance; BAPTA-AM,,2-bis(2-aminophenoxy)ethane-N,N,N=,N=-tetraacetic acid tetrace-oxy methyl ester; BCG, Bergmann glial cells; Chx, cycloheximide;NQX, 6-cyano-2,3-dihydroxy-7-nitroquinoxaline-2,3-dione; CPCCOEt,-(hydroxyimino)cyclopropa[b]chromen-1a-carboxylate ethyl ester; CPPG,RS)-�-cyclopropyl-4-phosphonophenylglycine; DHPG, (RS)-3,5-dihydroxy-henylglycine; DMEM, Dulbecco’s modified Eagle’s medium; EDTA, ethyl-nediaminetetraacetic acid; Glu, glutamate; iGluR, ionotropic glutamate re-eptor; KA, kainic acid/kainate; L-AP4, L-(�)-2-amino-4-phosphonobutyriccid; mGluRs, metabotropic glutamate receptors; MK-801, (1)-5-methyl-0,11-dihydro-5Hdibenzo[a,d]-cyclohepten-5,10-imine hydrogen maleate;TOR, mammalian target of rapamycin; NMDA, N-methyl-D-aspartate;AGE, polyacrylamide gel electrophoresis; PI3-K, phosphatidyl inositol 3inase; PKC, Ca2�/diacylglicerol protein kinase; PP2, 4-amino-5-(4-chloro-
S6K
ghenyl)-7-(t-butyl) pyrazolo[3,4-d] pyrimidine; p70 , ribosomal protein S6inase; PKB, protein kinase B; SDS, sodium dodecyl sulfate.
306-4522/06$30.00�0.00 © 2006 IBRO. Published by Elsevier Ltd. All rights reseroi:10.1016/j.neuroscience.2006.04.076
1389
urther strengthen the notion of an active participation of glialells in synaptic transmission. © 2006 IBRO. Published bylsevier Ltd. All rights reserved.
ey words: p70S6K, glutamate receptors, Bergmann glia,TOR, translational control.
xcitatory neurotransmission in the mammalian nervousystem is mediated to a large extent by glutamate (Glu).lu receptors are functionally divided into ionotropic
iGluRs) and metabotropic receptors (mGluRs). In terms oflu analogs, iGluRs have been classified into N-methyl-D-spartate (NMDA), �-amino-3-hydroxy-5-methyl-4-isoax-zolepropionate (AMPA) and kainate (KA) receptors (Holl-ann and Heinemann, 1994). Based on sequence simi-
arity, signal transduction mechanisms, and pharmacology,GluRs have been subdivided into three groups. Group Ire coupled to the stimulation of phospholipase C and theeneration of an intracellular calcium signal, while GroupsI and III regulate cyclic AMP levels through the inhibition ofdenylate cyclase. These three groups are pharmacolog-
cal activated by specific agonists such as (�)-1-aminocy-lopentane-trans-1,3-dicarboxylic acid (trans-ACPD) andRS)-3,5-dihydroxyphenylglycine (DHPG) for Group I, L-�)-2-amino-4-phosphonobutyric acid (L-AP4) activatesroup II while quisqualic acid (QA) acts upon Group III
Coutinho and Knopfel, 2002). Both iGluRs as well asGluRs are expressed in Bergmann glial cells (BGC)
Lopez et al., 1998; Gallo and Ghiani, 2000). Stimulation ofhese glial receptors are likely to have a role in long-termynaptic plasticity in the cerebellum (Iino et al., 2001).
Deficits in glutamatergic neurotransmission have beenelated to neuronal damage and development of neurode-enerative diseases (Mattson and Chan, 2003; Teismannnd Schulz, 2004). Moreover, activation of GluRs results inembrane to nuclei signaling involved in transcriptionalnd translational regulation in neurons and glial cells (Agu-
rre et al., 2002; Fields and Stevens-Graham, 2002). Whileost studies have focused into the molecular mechanismsf gene expression regulation at the transcriptional level,ovalent modifications of preexisting proteins and regula-ion of protein synthesis is fundamental in our understand-ng of gene expression regulation. In fact, dendritic proteinynthesis is believed to play a crucial role in long-termynaptic plasticity (Cammalleri et al., 2003). The fine reg-lation of the proper amount of a certain protein, mightacilitate the long-lasting structural modifications that en-ode memory (Lamprecht and LeDoux, 2004; Lang et al.,004). Previous studies in different cell types have sug-
ested that translation regulation can be mediated throughved.

pm
SgfpAtcrcgpt
pmfbsdsteTwaybtteipntorweorRpaPcaesS
wap2i
ie
sdtilat
A
CmisMu
C
TbJpsph(sac
C
C(uccfsuppnp
M
CL
Ncw1Tdn1fSaR
M. E. González-Mejia et al. / Neuroscience 141 (2006) 1389–13981390
hosphorylation of several components of the translationalachinery (Wang et al., 2001).
The serine/threonine protein kinase ribosomal protein6 kinase (p70S6K, also named S6K1) is an insulin/mito-en-activated protein kinase member of the AGC kinaseamily, whose major substrate is the 40S ribosomal subunitrotein S6 (Banerjee et al., 1990; Kozma et al., 1990;vruch, 1998). It is involved in the translational control of
he so-called 5= TOP mRNAs, a subset of mRNAs thatontain an oligopirimidine tract in their 5= untranslatedegion (Proud and Denton, 1997). TOP mRNAs encodeomponents of the translational machinery such as elon-ation factors, ribosomal proteins and poly (A)-bindingroteins and by these means play a key role in modulatingranslational efficiency.
Diverse kinases activate p70S6K through a multisitehosphorylation process directed to separated protein do-ains or modules. Roughly, p70S6K has been divided into
our domains. Module I extends from the N-terminus to theeginning of the catalytic domain and confers rapamycinensitivity to the kinase, module II contains the catalyticomain that includes a mitogen-induced phosphorylationite, module III links the catalytic domain to the carboxyerminus and contains two phosphorylation sites (Pearsont al., 1995; Weng et al., 1995; Pullen and Thomas, 1997).he fourth module comprises a pseudosubstrate domainith four phosphorylation sites (Ferrari et al., 1992). Inter-ctions of these modules lead to interdependent phosphor-lation events. Although seven phosphorylation sites haveeen identified, Thr389 phosphorylation is the main eventhrough which full p70S6K activity is accomplished, sincehe substitution of this residue with Ala fully inactivates thenzyme (Pearson et al., 1995). At least two distinct signal-
ng pathways have been identified in the activation of70S6K. One input is provided by phosphatidylinositol 3-ki-ase (PI3-K), which is recruited to the activated receptoryrosine kinases or their substrates in response to insulinr mitogens (Alessi and Downes, 1998; Wymann and Pi-ola, 1998). Treatment of cells with PI3-K inhibitors such asortmannin inhibits the activation of the enzyme (Wengt al., 1995). The other pathway contributing to activationf p70S6K described up to date, is the mammalian target ofapamycin (mTOR, also named FRAP, RAFT-1, orAPT-1) (Schmelzle and Hall, 2000). The immunosup-ressant rapamycin is known to cause dephosphorylationnd inactivation of p70S6K (Jefferies et al., 1994, 1997;earson et al., 1995). Rapamycin, in complex with theytosolic FK506-binding protein (FKBP12), binds to mTORnd inhibits its function in vivo, and previous studies havestablished that mTOR is the rapamycin-sensitive up-tream regulator of p70S6K (Harada et al., 2001; Hay andonenberg, 2004).
In chick cerebellum Bergmann glia primary cultures asell as in cerebellar slices, Glu exposure leads to thessociation of PI3-K and scaffold proteins to tyrosine-hosphorylated AMPA receptors (Millan et al., 2001,004), opening the possibility that Glu receptors could be
nvolved in translational control, as has been demonstrated t
n rat hippocampal slices and synaptoneurosomes (Bankot al., 2004).
In order to gain insight into this problem, the aim of thistudy was to establish a correlation between a Glu-depen-ent regulation of protein synthesis and Thr389 phosphoryla-ion of p70S6K. Our results strongly suggest that p70S6K isnvolved in the Glu signaling transactions that result in trans-ational control in a PI3-K/PKB/mTOR dependent mannernd demonstrate, for the first time that Glu is involved in
ranslational control in glial cells.
EXPERIMENTAL PROCEDURES
nimals
hick embryos (10 days old) were obtained from Avi-Mex andaintained at 37 °C until used. All experiments conformed to
nternational guidelines on the ethical use of animals and had thepecific approval of the Animal Ethics Committee of Cinvestav-exico. All efforts were made to reduce the number of embryossed and their suffering.
hemicals
issue culture reagents were from Gibco Invitrogen (Gaithers-urg, MD, USA). Wortmannin was obtained from Calbiochem (Laolla, CA, USA). Polyclonal anti-p-p70 S6 kinase (Thr 389), anti-70 S6 kinase and poly (ADP-ribose) polymerase (PARP) wereupplied by Santa Cruz Biotechnology (Santa Cruz, CA, USA);olyclonal anti-rabbit and anti-mouse antibodies conjugated toorseradish peroxidase were purchased from Zymed LaboratoriesSan Francisco, CA, USA). The Bradford and the sodium dodecylulfate–polyacrylamide gel electrophoresis (SDS-PAGE) re-gents were obtained from Bio-Rad (Hercules, CA, USA). All otherhemicals were from Sigma (St. Louis, MO, USA).
ell culture and stimulation protocol
hick cerebellar BGC were prepared as detailed elsewhereOrtega et al., 1991). Briefly, 14-day-old chick embryos weresed, and the cerebellum was dissected and homogenized me-hanically. Cells were plated at a density of 8�105/ml in Dulbec-o’s modified Eagle’s medium (DMEM) supplemented with 10%etal bovine serum, and 100 units/ml penicillin, and 100 �g/mltreptomycin. The cells were incubated at 37 °C in 5% CO2 andsed after 5–6 days in culture. Confluent monolayers were ex-osed to the indicated concentrations of agonists for varyingeriods of time; antagonists were added 30 min before the ago-ists. Incubation was stopped by removing the medium, and sam-les were processed as detailed below.
etabolic labeling of proteins
onfluent BGC monolayers were labeled overnight with 1 �Ci of-[35S]-methionine (Amersham Pharmacia Biotech, Piscataway,J, USA) in methionine-free DMEM. After extensive washing, theells were treated with Glu for the indicated time periods. Cellsere washed twice with ice-cold PBS (10 mM K2HPO4/KH2PO4,50 mM NaCl, pH 7.4) and lysed with ice-cold RIPA buffer (20 mMris–HCl, pH 7.5, 50 mM KCl, 5 mM MgCl2, 400 mM NaCl, 2 mMithiothreitol, 1 mM aprotinin, 1 mM leupeptin, and 100 �M phe-ylmethylsulfonyl fluoride, 20% glycerol, 1% Triton X-100). After a0 min centrifugation at top speed in a microfuge, the supernatantraction (approximately 15 �g of protein) was resolved throughDS-PAGE and visualized by silver staining. The gel was driednd exposed to X-ray films (Kodak X-Omat; Eastman Kodak,ochester, NY, USA) at �70 °C with intensifying screens. Densi-
ometric analysis was done with software Kodak 1D 3.5.4.

A
T(fiwffivasl
S
EiasePmnoatyPm
S
DaiyNtS
G[
Tpiow[fic�f[abbtoWaeuo
G
TtutaHtbf1pmeFlruTwcGpfvasc
Pr
TpBog(pp(2mpsGimwt3p1ptnD
M. E. González-Mejia et al. / Neuroscience 141 (2006) 1389–1398 1391
ssessment of protein synthesis
o measure overall protein synthesis, an aliquot of the samplesapproximately 15 �g of protein) was spotted onto GF/C microfiberlters (Whatman, Clifton, NJ, USA). The filters were air-dried andashed for 10 min in ice-cold 10% trichloroacetic acid (TCA)
ollowed by three 10-minute washes in ice-cold 5% TCA. Thelters were dried at room temperature and placed in scintillationials with 2 ml of scintillation liquid containing 10 �l glacial aceticcid. [35S]-methionine incorporation was determined via liquidcintillation counting (Beckman LS 6000 s.c.). All data were ana-yzed with the Prism, GraphPad Software (San Diego, CA, USA).
DS-PAGE and Western blots
qual amounts of protein (approximately 80 �g) were denaturizedn Laemmli’s sample buffer and resolved through 10% SDS-PAGEnd electroblotted to nitrocellulose membranes. Blots weretained with Ponceau S stain to confirm that protein content wasqual in all lanes. Membranes were soaked in PBS to remove theonceau S and incubated in TBS containing 5% dried skimmedilk and 0.1% Tween 20 for 90 min to block the excess ofonspecific protein binding sites. Membranes were then incubatedvernight at 4 °C with the primary antibodies, followed by second-ry antibodies. Immunoreactive polypeptides were detected usinghe ECL chemiluminescence Kit (Amersham). Densitometric anal-sis was performed and the data analyzed with the Prism, Graph-ad Software. The optical densities of phospho-p70S6K were nor-alized to the densities p70S6K.
tatistical analysis
ata are expressed as the mean�standard error. A one-waynalysis of variance (ANOVA) was performed to determine signif-
cant differences between the different conditions. When this anal-sis indicated significance (at the 0.05 level), post-hoc Student-ewman-Keuls test analysis was used to determine which condi-
ions were significantly different from each other Prism, GraphPadoftware.
RESULTS
lu exposure induces a biphasic effect in35S]-methionine incorporation
ranslational control has been regarded as the fine-tuningrocess in the regulation of the availability of proteins. The
mportant role that GluRs have in the plasticity propertiesf the brain prompted us to investigate if this amino acidould elicit a change in the TCA-precipitable amount of
35S]-methionine incorporation into proteins in a well-de-ned culture system: chick cerebellum BGC. To this end,onfluent monolayers were metabolically labeled with 1Ci of [35S]-methionine for 12 h and exposed to 1 mM Glu
or 5, 10, 15, 30, 60 and 120 min. A marked decrease in35S]-methionine incorporation was found after 15 min, butfter 30 min this phenomenon starts to revert returning toasal levels after 120 min. These data were indicative of aiphasic effect that necessarily involves an increase in pro-ein synthesis in its recovery phase. As depicted in panel Af Fig. 1, cycloheximide (Chx) inhibits the recovery phase.hen the protein pattern was analyzed via SDS-PAGE
nd autoradiography, it is clear that despite the overallffect of Glu in terms of TCA-precipitable proteins, individ-al polypeptides are newly synthesized even after 15 min
f Glu exposure (Fig. 1, panel B). dlu dependent phosphorylation of p70S6K
he described recovery phase in protein synthesis de-ected after 15 min of Glu treatment prompted us to eval-ate if p70S6K could be involved since it is known thatranslation of the protein synthesis machinery is favoredfter a reduction in the translational process (Thomas andall, 1997). Phosphorylation of p70S6K was determined
hrough the use of a phospho-specific anti-p70S6K anti-ody that recognizes phospho-Thr389, the critical residue
or its catalytic activity. Treatment of BGC with 1 mM Glu forh results in a marked increase in Thr389 phosphorylated
70S6K (Fig. 2, panel A). Note that Glu treatment does notodify p70S6K levels. The time dependence of the Gluffect was evaluated and as clearly shown in panel B ofig. 2, a sustained increase in p70S6K Thr389 phosphory-
ation, is detected as early as 15 min of Glu exposureeaching a maximal level after 60 min and maintaining it forp to 120 min and returning to control levels after 240 min.he dose-dependency of p70S6K Thr389 phosphorylationas established through the exposure for 60 min to in-reasing Glu concentrations. Although no saturation of thelu effect was achieved, it is clear that the increase in70S6K Thr389 phosphorylation is a receptor-mediated ef-
ect (Fig. 2, panel C). To further confirm this, the EC50
alue was calculated, an EC50 of 154 �M was obtained,nd one has to keep in mind that this value is biased by theignal amplification of the transduction components of theascade triggered by Glu.
harmacological characterization of the Glueceptors involved in p70S6K phosphorylation
he pharmacological profile of the GluRs involved in thehosphorylation of p70S6K was undertaken. ConfluentGC monolayers were exposed to a 500 �M concentrationf the ionotropic agonists NMDA (in the presence of 50 �Mlycine), AMPA and KA for 60 min. As shown in panel AFig. 3), the three analogs were able to increase p70S6K
hosphorylation, KA being the most effective. As ex-ected, the NMDA and non-NMDA antagonists, MK-801200 �M) and 6-cyano-2,3-dihydroxy-7-nitroquinoxaline-,3-dione (CNQX, 25 �M) respectively, when present 30in before Glu were able to prevent the increase in phos-horylation. The metabotropic contribution to the Glu re-ponse was also evaluated. To our surprise, Group I androup III of the mGluRs are involved in the Glu-mediated
ncrease in p70S6K Thr389 phosphorylation. The Group Ietabotropic agonist DHPG at a 25 �M concentration asell as the Group III agonist L-AP4 at a 100 �M concen-
ration increase significantly p70S6K phosphorylation (Fig.). The use of specific antagonists, 7-(hydroxyimino)cyclo-ropa[b]chromen-1a-carboxylate ethyl ester (CPCCOEt,00 �M) (Group I antagonist) and (RS)-�-cyclopropyl-4-hosphonophenylglycine (CPPG, 300 �M) (Group III an-agonist), corroborated these results. Interestingly, theon-NMDA antagonist CNQX was capable to reduce theHPG response (Fig. 3, panel B). In summary, Glu-depen-
ent p70S6K Thr389 phosphorylation is mediated through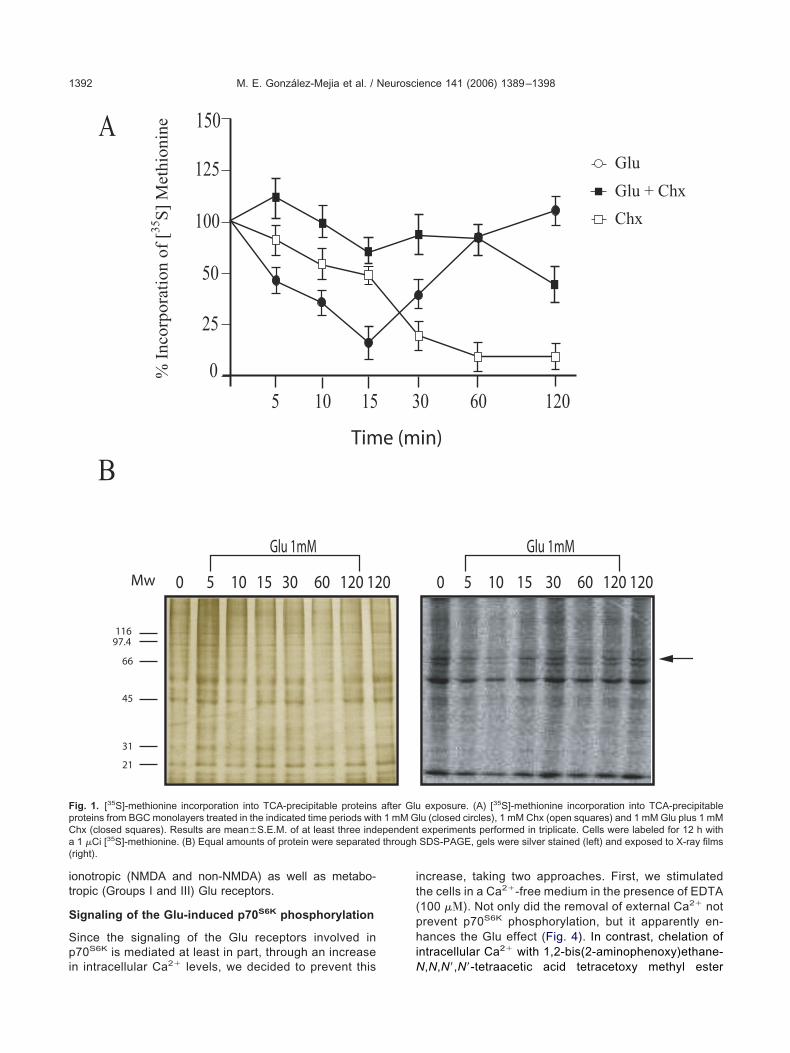
it
S
Spi
it(phi
FpCa(
M. E. González-Mejia et al. / Neuroscience 141 (2006) 1389–13981392
onotropic (NMDA and non-NMDA) as well as metabo-ropic (Groups I and III) Glu receptors.
ignaling of the Glu-induced p70S6K phosphorylation
ince the signaling of the Glu receptors involved in70S6K is mediated at least in part, through an increase
A
B
0
25
50
100
125
5 10 15
0 5 10 15 30 60 120 12
Glu 1mM
Tim
% I
ncor
pora
tion
of
[35S
] M
ethi
onin
e
Mw
11697.4
66
45
31
21
150
ig. 1. [35S]-methionine incorporation into TCA-precipitable proteinsroteins from BGC monolayers treated in the indicated time periods withhx (closed squares). Results are mean�S.E.M. of at least three ind1 �Ci [35S]-methionine. (B) Equal amounts of protein were separated
right).
n intracellular Ca2� levels, we decided to prevent this N
ncrease, taking two approaches. First, we stimulatedhe cells in a Ca2�-free medium in the presence of EDTA100 ��). Not only did the removal of external Ca2� notrevent p70S6K phosphorylation, but it apparently en-ances the Glu effect (Fig. 4). In contrast, chelation of
ntracellular Ca2� with 1,2-bis(2-aminophenoxy)ethane-
Glu
Glu + Chx
Chx
0 60 120
0 5 10 15 30 60 120 120
Glu 1mM
in)
exposure. (A) [35S]-methionine incorporation into TCA-precipitablelu (closed circles), 1 mM Chx (open squares) and 1 mM Glu plus 1 mMexperiments performed in triplicate. Cells were labeled for 12 h withSDS-PAGE, gels were silver stained (left) and exposed to X-ray films
3
0
e (m
after Glu1 mM G
ependentthrough
,N,N=,N=-tetraacetic acid tetracetoxy methyl ester

(a
s
FbipIp rmined a
M. E. González-Mejia et al. / Neuroscience 141 (2006) 1389–1398 1393
BAPTA-AM) completely prevented the Glu effect in the
ig. 2. Glu induces p70S6K phosphorylation. (A) BGC monolayers welot with the indicated antibodies. (B) BGC were treated at different t
ncreasing Glu concentrations. The membrane was incubated with aeroxidase linked goat anti-rabbit secondary antibodies. The immunopogGs using the ECL kit. Data were normalized with p70S6K levels. Aerformed using paired t-tests. * P�0.05, and EC50 values were dete
bsence of extracellular Ca2� (Fig. 4). These results, s
urprising in terms of an extracellular Ca2�-independented to 1 mM Glu for 1 h and total extracts were analyzed via Westernds with a fixed 1 mM Glu concentration. (C) BGC were treated with6K Thr389 (1:2000) or p70S6K (1:1000), following the incubation with
lypeptides were visualized with peroxidase-conjugated goat anti-rabbitdiogram of a typical experiment is shown. Statistical analysis was
fter nonlinear regression analysis with Prism 4 software (GraphPad).
re exposime perionti-p-p70S
sitive pon autora
ignaling through ionotropic receptors, are in line with

pf
pw
dCSz
Fa5fle test ana
M. E. González-Mejia et al. / Neuroscience 141 (2006) 1389–13981394
revious findings from our group (Millan et al., 2001) androm other groups (Hayashi et al., 1999).
As already mentioned in the introduction, p70S6K phos-horylation is mediated through the PI3-K/PKB pathway as
% o
f co
ntro
l p70
S6K
Pho
spho
ryla
tion
0
50
100
150
200
250
0
50
100
150
200
250
**
Ns DHPGCNQX
CNQX + DHPG
CPCCOEt
CPC
% o
f co
ntr
ol p
70S
6K P
hos
ph
oryl
atio
n
0
50
100
150
200
250
Ns Glu NMDA
NMDA + Gly
AMPA
AMPA + CNQ
KA KA
*
ig. 3. Pharmacological profile of p70S6K phosphorylation induction. (Analogues, and Glu 1 mM, NMDA 500 �M plus glycine 10 �M, AMPA0 �M, MK-801 5 �M plus glycine 10 �M, CPCCOEt (100 �M), and CPor 30 min with the antagonists CNQX 50 �M, CPCCOEt, 100 �M, andevels of p70S6K phosphorylation were detected as in Fig. 2. Values rexperiments. Data were compared by one-way ANOVA and Dunnett’s
ell as by mTOR. Since we have described a Glu-depen- c
ent PI3-K interaction with non-NMDA receptors that isa2�-independent (Millan et al., 2004) but sensitive to therc inhibitor 4-amino-5-(4-chlorophenyl)-7-(t-butyl) pyra-olo[3,4-d] pyrimidine (PP2), we decided to preincubate
p70S6K
P- p70S6K
p70S6K
P- p70S6K
NsGluNMDA + GlyNMDA + Gly + MK801AMPAAMPA + CNQXKAKA + CNQXDHPGDHPG + CPCCOEt
L-AP4 + CPPG
HPG
CPCCOEt
NQX + CPCCOEt +
DHPG
Ns
DHPG
CNQX
CNQX + DHPG
CPCCOEt
CPCCOEt + DHPG
CNQX + CPCCOEt
CNQX + CPCCOEt + DHPG
PG + CPCCOEt
L-APA
L-AP4 + C
PPG
L-AP4
onolayers were incubated for 1 h with the indicated iGluRs or mGluRsand KA 500 �M, DHPG at a 25 �M, L-AP4. The antagonists CNQX�M) were added 30 min before. (B) BGC monolayers were incubated0 �M plus CPCCOEt, before the DHPG 25 �M treatment for 1 h. Theriplicate determinations and are the mean�S.E. of three independentlysis (* P�0.05), with Prism 4 software (GraphPad).
*
COEt + D
CNQX +
C
X
+ CNQX
DHPGDH
) BGC m500 �M
PG (100CNQX 5
present t
onfluent BGC monolayers with a 100 nM concentration of

wLwfeyti(mACinm(iG
aomtrCt
Lor2atNi
cdsapOu
FmS((irrF
Fio(psa
M. E. González-Mejia et al. / Neuroscience 141 (2006) 1389–1398 1395
ortmannin an established PI3-K blocker or with 100 nMY294002 (data not shown). As shown in panel A of Fig. 5,ortmannin reverses completely the Glu effect. As expected
rom our previous work, LY294002 also prevents the Gluffect. A similar scenario is present when 1L-6-hydroxymeth-l-chiro-inositol 2-(R)-2-O-methyl-3-O-octadecylcarbonatehe AKT inhibitor (Akt I) when used at a 25 nM concentration,t also prevents the Glu-dependent p70S6K phosphorylationFig. 5, panel B). As one could expect, rapamycin, theTOR inhibitor also blocks the Glu effect (Fig. 5, panel C).t this stage, besides the role for increased intracellulara2� levels in p70S6K phosphorylation (Fig. 4), it was
mperative to evaluate other signal transduction compo-ents of the cascade triggered by Group I and Group III ofGluRs, such as the Ca2�/diacylglicerol protein kinase
PKC). Preincubation with 10 nM of the protein kinase Cnhibitor bisindolylmaleimide I (Bis I) did not prevent thelu effect (not shown).
Taken together all these results suggest that Glu iscting, at least partially, through the subsequent activationf Src, PI3-K, PKB and mTOR and that this effect might beediated through iGluRs, since it has been established
hat PI3-K associates with tyrosine phosphorylated AMPAeceptors in BGC and that this interaction, besides beinga2�-independent, results in PKB phosphorylation and
herefore activation (Morales et al., in press).
DISCUSSION
ong-term changes in synaptic strength are dependentn protein synthesis and are often mediated by NMDAeceptors as well as by Group I of mGluRs (Banko et al.,004, 2006; Page et al., in press). It has been tradition-lly assumed that this increase in protein synthesisakes place in neurons (for a review see Kandel, 2001).evertheless, increasing evidence accumulates regard-
ig. 4. Role of Ca2� in Glu-induced p70S6K phosphorylation. For exper-ments in Ca2�-free medium, BGC monolayers were treated with 500 �Mf EDTA and 25 �M BAPTA-AM for 30 min in Ca2�-free assay buffer. Glu1 mM) was added and incubation continued for 1 h. The levels of p70S6K
hosphorylation were detected as in Fig. 2. A typical autoradiogram ishown. Data were compared by one-way ANOVA and Dunnett’s testnalysis (* P�0.05), with Prism 4 software (GraphPad).
ng a plausible role of glial cells in neuronal communi-os
ation (Araque and Perea, 2004). In the cerebellum,uring parallel fiber–Purkinje cells synaptic transmis-ion, Glu evokes inward currents in BGC through thectivation of AMPA receptors and electrogenic trans-orters that face the presynaptic elements (Bellamy andgden, 2005; Matsui et al., 2005). A strong and contin-ous communication between Purkinje cells and BGC is
% o
f co
ntro
l p7
0S6K
Pho
spho
ryla
tion
% o
f co
ntro
l p7
0S6K
Pho
spho
ryla
tion
GluNs
Wort + Glu
Wort
PP2
PP2 + Glu
0
50
100
150
200
Ns GluW
ortW
ort + G
lu
PP2 + Glu
PP2
p70S6K
P- p70S6K
p70S6K
P- p70S6K
p70S6K
P- p70S6K
% o
f co
ntr
ol
p7
0S
6K
Ph
osp
ho
ryla
tio
n
Glu
Ns
Akt I + G
lu
Akt I + Glu
Akt I
Glu
Ns
Rapamycin + Glu
Rapamycin
Glu
Ns Rapam
ycin
+ Glu
Rapam
ycin
Glu
Ns
Akt I
A
B
C
*
*
*
0
25
50
75
100
125
150
175
200
DMSO
DMSO
0
25
50
75
100
125
150
175
200
225
250
ig. 5. Glu signaling involved in p70S6K phosphorylation. (A) BGConolayers were incubated with and without 100 nM wortmannin orrc kinase inhibitor PP2 100 nM for 30 min before treatment with Glu.
B) The AKT inhibitor (Akt I) 25 nM, was added 30 min before Glu1 mM) and incubation continued for 1 h. (C) Rapamycin 50 nM wasncubated for 30 min before Glu 1 mM treatment. All results shown areepresentative of at least three independent experiments with similaresults. The levels of p70S6K phosphorylation were detected as inig. 2. A typical autoradiogram is shown. Data were compared by
ne-way ANOVA and Dunnett’s test analysis (* P�0.05), with Prism 4oftware (GraphPad).
dtrnrvnrrG2ii(t2
ppptotcimste
wtmpptmSpi(tis(otaftiPd1
c(Gp
bAemwdacoFpvcGstei
iitptpicsrnppsepirbtp
IpiTtMati
AsbMf
M. E. González-Mejia et al. / Neuroscience 141 (2006) 1389–13981396
educed after the reported change in morphology thatakes place in both cell types when the BGC AMPAeceptors are modified (Watanabe, 2002). Moreover, theeurotrophic effects of Glu, dependent mainly on neu-onal and glial AMPA receptors have been recently re-iewed (Limatola, 2004). In fact, several membrane touclei signaling cascades that results in transcriptionalegulation of glia-specific genes and cytoskeletal rear-angements have been demonstrated to be triggered bylu (Lopez-Bayghen and Ortega, 2002; Millan et al.,004). Interestingly enough, one of the glial genes that
s transcriptionally regulated in a Glu-dependent fashions the Na�-dependent Glu/aspartate transporterGLAST) critically involved in the decay rate of excita-ory postsynaptic currents of Purkinje cells (Stoffel et al.,004).
Viewing gene expression in totality, translation occu-ies a position somewhere in the middle of a complexathway that begins with transcription, continues with RNArocessing and transport, and ends with protein transloca-
ion, modification, folding assembly and degradation. Eachf these steps is tightly regulated; yet, transcription and
ranslation are especially critical for the cells. Translationalontrol offers the possibility for a fine control since changes
n transcription rates are considerably greater in magnitudeost of the times. Besides, immediacy is the most con-
picuous advantage of translational control over transcrip-ional and other nuclear control mechanisms (Sonenbergt al., 2000).
The regulation of translation rates, the frequency withhich a given mRNA is translated, is primarily regulated at
he initiation phase. An important regulator of this phase isTOR, it does so primarily through its downstream targets70S6K and the elongation factor binding protein 4E-BP1.70S6K is a major regulator of translation since it promoteshe expression of several members of the translationalachinery whose mRNAs are TOP-mRNAs (Hay andonenberg, 2004). The phosphorylation state of Thr389 in70S6K is determinant for its activity and as already stated
s regulated in a complex manner by mTOR and PI3-KBalendran et al., 1999). In this context, the present con-ribution addresses a Glu-dependent translational controln cerebellar BGC. A biphasic effect in overall proteinynthesis is present in cultured BGC after Glu treatmentFig. 1), interestingly individual polypeptides such as thene marked with an arrow (�66 kDa) are augmented evenhe overall synthesis is depressed. The recovery phase,fter 15 min of Glu is sensitive to Chx, a peptidyl trans-erase inhibitor, clearly suggesting an induction of transla-ion initiation and therefore a plausible role for p70S6K, thiss further supported by the reported physical interaction ofI3-K with ionotropic Glu receptors and its agonist-depen-ent activation in neurons and glial cells (Hayashi et al.,999; Millan et al., 2004).
A time- and dose-dependent p70S6K phosphorylationould be unequivocally established after Glu exposureFigs. 2 and 3). The kinetic data strongly suggest thatlu promotes a stable (for up to 120 min) p70S6K phos-
horylation, reflecting a sustained protein synthesis aurst, which might comprise translational components.concentration-dependence of the Glu effect could be
stablished and an EC50 value of 154 �M was deter-ined. Beyond the absolute value of this constant,hich is likely to be overestimated, is the fact that Glu-ependent p70S6K phosphorylation is a receptor-medi-ted effect. Through the use of Glu analogues we couldonclude that the Glu receptors involved are ionotropicf the NMDA as well as non-NMDA subtypes (Fig. 3).urthermore, mGluRs of the Group I and Group III cou-led to phosphatidylinositol metabolism and PKC acti-ation are also involved. This broad and rather unspe-ific effect needs further addressing with more selectivelu analogues, experiments that are clearly beyond thecope of the present communication. It should be men-ioned however, that previous work from our group hasstablished the expression of iGluRs as well as mGluRs
n cultured chick BGC (Lopez et al., 1994, 1997, 1998).In terms of the signaling cascade, the effect is Ca2�-
ndependent but sensitive to the Src inhibitor PP2, which isn line with our previous findings regarding AMPA recep-ors tyrosine phosphorylation (Millan et al., 2004). As ex-ected then, if the AMPA receptors tyrosine phosphoryla-ion is blocked, the recruitment of PI3-K could not takelace and the activation of PKB would be prevented, then
n such scenario, both mTOR and p70S6K phosphorylationould not take place. In support for this interpretation is theensitivity of the Glu effect to the PKB inhibitor and toapamycin (Fig. 5). Nevertheless this interpretation doesot take in account that Group I and Group III mGluRs alsoarticipate in p70S6K phosphorylation. An attractive hy-othesis is that mGluRs could “transactivate” iGluRs, pos-ibly favoring their Tyr phosphorylation. Then one shouldxpect that a selective activation of mGluRs that results in70S6K phosphorylation (Fig. 3) would be sensitive to an
GluRs antagonist. As depicted in panel B of Fig. 3, theesponse to the selective Group I agonist DHPG is blockedy CNQX. Experiments currently under way in our labora-ory are aimed to the characterization of this novel andutative “transactivation.”
CONCLUSION
n summary, we report here a Glu-dependent regulation ofrotein synthesis in BGC in which p70S6K is likely to be
nvolved in a Src/PI3-K/PKB/mTOR dependent manner.hese results should pave the way of our understanding of
he role of glial Glu receptors in translational regulation.oreover, our data favor the hypothesis that glial cells asctive participants of synaptic plasticity, modify their pro-
ein repertoire through transcriptional and translational controln response to the main excitatory transmitter.
cknowledgments—The authors acknowledge the technical as-istance of Luis Cid and Blanca Ibarra. This work was supportedy a grant from Conacyt-México (43164-Q) to A.O. Conacyt-éxico supports M.E.G.M, M.M. and R.C.Z. through doctoral
ellowships. The critical opinions of Prof. Esther López-Bayghen
re acknowledged.
A
A
A
A
B
B
B
B
B
C
C
F
F
G
H
H
H
H
I
J
J
K
K
L
L
LL
L
L
L
M
M
M
M
M
O
P
P
P
P
S
S
S
T
T
W
M. E. González-Mejia et al. / Neuroscience 141 (2006) 1389–1398 1397
REFERENCES
guirre A, Lopez-Bayghen E, Ortega A (2002) Glutamate-dependenttranscriptional regulation of the chkbp gene: signaling mecha-nisms. J Neurosci Res 70:117–127.
lessi DR, Downes CP (1998) The role of PI3-kinase in insulin action.Biochim Biophys Acta 1436:151–164.
raque A, Perea G (2004) Glial modulation of synaptic transmission inculture. Glia 47:241–248.
vruch J (1998) Insulin signal transduction through protein kinasecascades. Mol Cell Biochem 182:31–48.
alendran A, Currie R, Armstrong CG, Avruch J, Alessi DR (1999)Evidence that 3-phosphoinositide-dependent protein kinase-1 me-diates phosphorylation of p70 S6 kinase in vivo at Thr-412 as wellas Thr-252. J Biol Chem 274:37400–37406.
anerjee P, Ahmad MF, Grove JR, Kozlosky C, Price DJ, Avruch J(1990) Molecular structure of a major insulin/mitogen-activated70-kDa S6 protein kinase. Proc Natl Acad Sci U S A 87:8550–8554.
anko JL, Hou L, Klann E (2004) NMDA receptor activation results inPKA- and ERK-dependent Mnk1 activation and increased eIF4Ephosphorylation in hippocampal area CA1. J Neurochem 91:462–470.
anko JL, Hou L, Poulin F, Sonenberg N, Klann E (2006) Regulationof eukaryotic initiation factor 4E by converging signaling pathwaysduring metabotropic glutamate receptor-dependent long-term de-pression. J Neurosci 26:2167–2173.
ellamy TC, Ogden D (2005) Short-term plasticity of Bergmann glialcell extrasynaptic currents during parallel fiber stimulation in ratcerebellum. Glia 52:325–335.
ammalleri M, Lutjens R, Berton F, King AR, Simpson C, FrancesconiW, Sanna PP (2003) Time-restricted role for dendritic activation ofthe mTOR-p70S6K pathway in the induction of late-phase long-term potentiation in the CA1. Proc Natl Acad Sci U S A 100:14368–14373.
outinho V, Knopfel T (2002) Metabotropic glutamate receptors: elec-trical and chemical signaling properties. Neuroscientist 8:551–561.
errari S, Bannwarth W, Morley SJ, Totty NF, Thomas G (1992)Activation of p70s6k is associated with phosphorylation of fourclustered sites displaying Ser/Thr-Pro motifs. Proc Natl Acad SciU S A 89:7282–7286.
ields RD, Stevens-Graham B (2002) New insights into neuron-gliacommunication. Science 298:556–562.
allo V, Ghiani CA (2000) Glutamate receptors in glia: new cells, newinputs and new functions. Trends Pharmacol Sci 21:252–258.
arada H, Andersen JS, Mann M, Terada N, Korsmeyer SJ (2001)p70S6 Kinase signals cell survival as well as growth, inactivatingthe pro-apoptotic molecule BAD. Proc Natl Acad Sci U S A 98:9666–9670.
ay N, Sonenberg N (2004) Upstream and downstream of mTOR.Genes Dev 18:1926–1945.
ayashi T, Umemori H, Mishina M, Yamamoto T (1999) The AMPAreceptor interacts with and signals through the protein tyrosinekinase Lyn. Nature 397:72–76.
ollmann M, Heinemann S (1994) Cloned glutamate receptors. AnnuRev Neurosci 17:31–108.
ino M, Goto K, Kakegawa W, Okado H, Sudo M, Ishiuchi S, Miwa A,Takayasu Y, Saito I, Tsuzuki K, Ozawa S (2001) Glia-synapseinteraction through Ca2�-permeable AMPA receptors in Bergmannglia. Science 292:926–929.
efferies HB, Reinhard C, Kozma SC, Thomas G (1994) Rapamycinselectively represses translation of the “polypyrimidine tract”mRNA family. Proc Natl Acad Sci U S A 91:4441–4445.
efferies HBJ, Fumagalli S, Dennis PB, Reinhard C, Pearson RB,Thomas G (1997) Rapamycin suppresses 5=TOP mRNA transla-tion through inhibition of p70s6k. EMBO J 16:3693–3704.
andel ER (2001) The molecular biology of memory storage: a dia-
logue between genes and synapses. Science 294:1030–1038.ozma SC, Ferrari S, Bassand P, Siegmann M, Totty N, Thomas G(1990) Cloning of the mitogen-activated S6 kinase from rat liverreveals an enzyme of the second messenger subfamily. Proc NatlAcad Sci U S A 87:7365–7369.
amprecht R, LeDoux J (2004) Structural plasticity and memory. NatRev Neurosci 5:45–54.
ang C, Barco A, Zablow L, Kandel ER, Siegelbaum SA, ZakharenkoSS (2004) Transient expansion of synaptically connected dendriticspines upon induction of hippocampal long-term potentiation. ProcNatl Acad Sci U S A 101:16665–16670.
imatola C (2004) Neurotrophic effects of AMPA. Cerebellum 3:2–10.opez-Bayghen E, Ortega A (2002) Glutamate-dependent gene ex-
pression in glial cells. Curr Topics Neurochem 3:55–68.opez T, Lopez-Colome AM, Ortega A (1994) AMPA/KA receptor
expression in radial glia. Neuroreport 5:504–506.opez T, Lopez-Colome AM, Ortega A (1997) NMDA receptors in
cultured radial glia. FEBS Lett 405:245–248.opez T, Lopez-Colome AM, Ortega A (1998) Changes in GluR4
expression induced by metabotropic receptor activation in radialglia cultures. Brain Res Mol Brain Res 58:40–46.
atsui K, Jahr CE, Rubio ME (2005) High-concentration rapid tran-sients of glutamate mediate neural-glial communication via ectopicrelease. J Neurosci 25:7538–7547.
attson MP, Chan SL (2003) Neuronal and glial calcium signaling inAlzheimer’s disease. Cell Calcium 34:385–397.
illan A, Aguilar P, Mendez JA, Arias-Montano JA, Ortega A (2001)Glutamate activates PP125(FAK) through AMPA/kainate receptorsin Bergmann glia. J Neurosci Res 66:723–729.
illan A, Arias-Montano JA, Mendez JA, Hernandez-Kelly LC, OrtegaA (2004) Alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionicacid receptors signaling complexes in Bergmann glia. J NeurosciRes 78:56–63.
orales M, González-Mejia ME, Bernabe A, Hernández-Kelly LCR,Ortega A. Glutamate activates protein kinase B (PKB/Akt) thoughAMPA receptors in cultured Bergmann glia cells. Neuroscience, inpress.
rtega A, Eshhar N, Teichberg VI (1991) Properties of kainate receptor/channels on cultured Bergmann glia. Neuroscience 41:335–349.
age G, Khidir FA, Pain S, Barrier L, Fauconneau B, Guillard O, PiriouA, Hugon J. Group I metabotropic glutamate receptors activate thep70S6 kinase via both mammalian target of rapamycin (mTOR)and extracellular signal-regulated kinase (ERK 1/2) signaling path-ways in rat striatal and hippocampal synaptoneurosomes. Neuro-chem Int, in press.
earson RB, Dennis PB, Han JW, Williamson NA, Kozma SC, Wet-tenhall RE, Thomas G (1995) The principal target of rapamycin-induced p70s6k inactivation is a novel phosphorylation site withina conserved hydrophobic domain. EMBO J 14:5279–5287.
roud CG, Denton RM (1997) Molecular mechanisms for the control oftranslation by insulin. Biochem J 328 (Pt 2):329–341.
ullen N, Thomas G (1997) The modular phosphorylation and activa-tion of p70s6k. FEBS Lett 410:78–82.
chmelzle T, Hall MN (2000) TOR, a central controller of cell growth.Cell 103:253–262.
onenberg N, Hershey JWB, Mathews MB (2000) Translational con-trol of gene expression, pp 1–88. New York: Cold Spring Harbor.
toffel W, Korner R, Wachtmann D, Keller BU (2004) Functionalanalysis of glutamate transporters in excitatory synaptic transmis-sion of GLAST1 and GLAST1/EAAC1 deficient mice. Brain ResMol Brain Res 128:170–181.
eismann P, Schulz JB (2004) Cellular pathology of Parkinson’sdisease: astrocytes, microglia and inflammation. Cell Tissue Res318:149–161.
homas G, Hall MN (1997) TOR signalling and control of cell growth.Curr Opin Cell Biol 9:782–787.
ang X, Li W, Williams M, Terada N, Alessi DR, Proud CG (2001)Regulation of elongation factor 2 kinase by p90(RSK1) and p70 S6
kinase. EMBO J 20:4370–4379.
W
W W
M. E. González-Mejia et al. / Neuroscience 141 (2006) 1389–13981398
atanabe M (2002) Glial processes are glued to synapses viaCa(2�)-permeable glutamate receptors. Trends Neurosci 25:5–6.
eng QP, Andrabi K, Klippel A, Kozlowski MT, Williams LT, Avruch J
(1995) Phosphatidylinositol 3-kinase signals activation of p70 S6kinase in situ through site-specific p70 phosphorylation. Proc NatlAcad Sci U S A 92:5744–5748.
ymann MP, Pirola L (1998) Structure and function of phosphoinosi-
tide 3-kinases. Biochim Biophys Acta 1436:127–150.(Accepted 26 April 2006)(Available online 12 June 2006)
Copyright © 2022 FDOKUMEN




















