
GABA uptake by purified avian Müller glia cells in culture
9
GABA is the main inhibitory aminoacid trans- mitter present in neurons and glial cells. Its uptake is carried out by specific high-affinity Na + /Cl - dependent transporters (GATs). It has been reported in the past that, in the avian ret- ina, [ 3 H]GABA appears to be exclusively accu- mulated by horizontal and amacrine cells in the inner nuclear layer, and also by ganglion cells. Purified chick Müller glia cultures were able to take up [ 3 H]GABA in a Na + and Cl - dependent way . Increasing GABA concentration increases GABA uptake by these cells, reaching half-max- imal transport efficiency (EC 50 ) around 0.3 mM. [ 3 H]GABA uptake by Müller glia neuronal-free cultures was not mediated by neuronal trans- porters since it was not blocked by NNC-711, but was inhibited by beta-alanine, a specific glial transporter inhibitor. Chick Müller glia in culture express both GAT-1 and GAT-3 GABA transporters. Although mixed neuron-glial dense cultures released GABA upon glutamate, high K + or veratridine stimulation, Müller glial cells did not release [ 3 H]GABA upon treatment with these agents, suggesting that different from neu- rons, transporter mediated GABA release is not a common mechanism operating in these cells. The data also suggest that Müller cells take up GABA unidirectionally , which may constitute an important mechanism of inactivating GABA activity mediated by neurons. Keywords: Müller glia; GABA uptake; retina; GABA transporter; GAT; Culture INTRODUCTION Glia represent the most abundant group of cells in the nervous system. In recent years innumerous reports have changed the view that glial cells were only a supportive system to neurons. Thus, recent evidence show that these cells participate actively in intercellular signaling through neuro-glial com- munication (Perea and Araque, 2006; Todd et al., 2006). Müller cells constitute the major macroglia cell population in the retina. These cells execute many of the functions associated with oligoden- drocytes, astrocytes and ependymal cells in other parts of the central nervous system (Newman and Reichenbach, 1996). Recently , it was demonstrated that under certain conditions avian Müller glia can transdifferentiate and generate a few neuronal types, suggesting a plastic role for these cells in the retina (Fischer and Reh, 2001, Fischer, 2005). In addition, nearly every disease of the retina is associated with Müller glial cell reactive gliosis (Bringmann et al., 2006). Müller glia span the whole length of the retina and contribute for the formation of the inner and outer limiting membranes of the tissue. Moreover they also extend processes into the inner and outer F .P. Graham Publishing Co. GABA Uptake by Purified Avian Müller Glia Cells in Culture CLARISSA DE SAMP AIO SCHITINE a , REGINA CÉLIA CUSSA KUBRUSLY a,b , RICARDO AUGUSTO DE MELO REIS a , EDNA NANAMI YAMASAKI a,c , MARIA CHRISTINA FIALHO DE MELLO a and FERNANDO GARCIA DE MELLO a,* Laboratório de Neuroquímica a e Neurobiologia da Retina c , Instituto de Biofísica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro and Laboratório de Neurofarmacologia b , Departamento de Fisiologia e Farmacologia, Instituto Biomédico, Universidade Federal Fluminense, Rio de Janeiro, Brasil. fgmello@biof.ufrj.br (Submitted 27 April 2007; Revised 9 May 2007; In final form 15 May 2007) *Corresponding author: Tel.: 00 55 21 25626594; FAX: 00 55 21 22808193; E-mail: fgmello@biof.ufrj.br ISSN 1029 8428 print/ ISSN 1476-3524 online. © 2007 FP Graham Publishing Co., www.NeurotoxicityResearch.com Neurotoxicity Research, 2007, VOL. 12(2). pp. 145-153
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of GABA uptake by purified avian Müller glia cells in culture

GABA is the main inhibitory aminoacid trans-mitter present in neurons and glial cells. Its uptake is carried out by specific high-affinityNa+/Cl- dependent transporters (GATs). It has been reported in the past that, in the avian ret-ina, [3H]GABA appears to be exclusively accu-mulated by horizontal and amacrine cells in the inner nuclear layer, and also by ganglion cells.Purified chick Müller glia cultures were able to take up [3H]GABA in a Na+ and Cl- dependent way. Increasing GABA concentration increases GABA uptake by these cells, reaching half-max-imal transport efficiency (EC50) around 0.3 mM. [3H]GABA uptake by Müller glia neuronal-free cultures was not mediated by neuronal trans-porters since it was not blocked by NNC-711, but was inhibited by beta-alanine, a specificglial transporter inhibitor. Chick Müller glia in culture express both GAT-1 and GAT-3 GABA transporters. Although mixed neuron-glial densecultures released GABA upon glutamate, highK+ or veratridine stimulation, Müller glial cellsdid not release [3H]GABA upon treatment withthese agents, suggesting that different from neu-rons, transporter mediated GABA release is not a common mechanism operating in these cells. The data also suggest that Müller cells take upGABA unidirectionally, which may constitutean important mechanism of inactivating GABA activity mediated by neurons.
Keywords: Müller glia; GABA uptake; retina; GABA transporter; GAT; Culture
INTRODUCTION
Glia represent the most abundant group of cells inthe nervous system. In recent years innumerousreports have changed the view that glial cells were only a supportive system to neurons. Thus, recent evidence show that these cells participate activelyin intercellular signaling through neuro-glial com-munication (Perea and Araque, 2006; Todd et al., 2006). Müller cells constitute the major macroglia cell population in the retina. These cells execute many of the functions associated with oligoden-drocytes, astrocytes and ependymal cells in other parts of the central nervous system (Newman and Reichenbach, 1996). Recently, it was demonstrated that under certain conditions avian Müller glia can transdifferentiate and generate a few neuronal types, suggesting a plastic role for these cells in the retina (Fischer and Reh, 2001, Fischer, 2005).In addition, nearly every disease of the retina is associated with Müller glial cell reactive gliosis (Bringmann et al., 2006). Müller glia span the whole length of the retina and contribute for the formation of the inner and outer limiting membranes of the tissue. Moreover they also extend processes into the inner and outer
F.P. Graham Publishing Co.
GABA Uptake by Purified Avian Müller Glia Cells in Culture
CLARISSA DE SAMPAIO SCHITINEa, REGINA CÉLIA CUSSA KUBRUSLYa,b, RICARDO AUGUSTO DE MELO REISa, EDNA NANAMI YAMASAKIa,c, MARIA CHRISTINA FIALHO DE MELLOa
and FERNANDO GARCIA DE MELLOa,*
Laboratório de Neuroquímicaa e Neurobiologia da Retinac, Instituto de Biofísica Carlos Chagas Filho,Universidade Federal do Rio de Janeiro and Laboratório de Neurofarmacologiab, Departamento de Fisiologia eFarmacologia, Instituto Biomédico, Universidade Federal Fluminense, Rio de Janeiro, Brasil. [email protected]
(Submitted 27 April 2007; Revised 9 May 2007; In final form 15 May 2007)
*Corresponding author: Tel.: 00 55 21 25626594; FAX: 00 55 21 22808193; E-mail: [email protected]
ISSN 1029 8428 print/ ISSN 1476-3524 online. © 2007 FP Graham Publishing Co., www.NeurotoxicityResearch.com
Neurotoxicity Research, 2007, VOL. 12(2). pp. 145-153

C. S. SCHITINE et al.146
plexiform layers (Sarthy and Ripps, 2001). Among other roles, Müller cells are thought to be involved in phagocytosis of neuronal debris, removal of toxic waste, K+ buffering, recycling of aminoacid neurotransmitters, energy control by breakdown of glycogen, and expression of a variety of voltage-gated ion channels, neurotransmitter receptors and transporters (Bringmann et al., 2006). Müller glia can also evoke Ca2+ waves, release neurotrans-mitters and other signaling molecules that, upon stimulation, can ultimately influence the activity of neuronal cells in the retina (Newman, 2004). GABAergic synapses in the avian retina are mainly represented by amacrine and horizontal cells modulating the glutamatergic radial flux of information (Calaza et al., 2006). The terminationof GABA action depends largely on membrane transporters present in the presynaptic neuronal ter-minal and/or in glial cells (Kanner, 1994). Most of the GABA uptake is done by glial transporters that exert a modulatory action on the overall activityof GABAergic synapses constituting potential tar-gets for therapeutical applications (Borden, 1996; Schousboe, 2000). To date four GABA transport-ers have been described (GAT1-4), and all types were described in glial cells, with GAT-3 being the predominant glial form (reviewed in Schousboe, 2003). However, the nomenclature related to GATsmay be confusing depending on the species stud-ied. As discussed by Wu et al. (2006), GAT1 isthe same for mouse, rat and human. Mouse GAT2corresponds to BGT-1. Mouse GAT3, is related to GAT-2 in rats and human. Finally mouse GAT4 is related to GAT-3 in rats. The GAT-1 transporter was found in rat (Brecha and Weigmann, 1994, Pow et al., 2005), cat,monkey and human (Pow et al., 2005, Casini et al., 2006), skate (Birnbaum et al., 2005), bullfrog and guinea pig retinas; GAT-2 was found in the bullfrog and GAT-3 in the guinea pig, cat, rat, mon-key, human and skate retinas (Zhao et al., 2000; Biedermann et al., 2002; Birnbaum et al., 2005; Pow et al., 2005). Previous data have shown that [3H]GABA appears to accumulate exclusively in horizontal, amacrine and in the retinal ganglion cells of the intact avian retina, but not in the chick Müller glia (Marshall and Voaden, 1974). It is known that the glial population in the avian retina is remarkably
different from other species (Won et al., 2000). Since we developed a purified chick Müller glia culture, free of neurons (Reis et al., 2002; Kubrusly et al., 2005), we decided to revisit this issue and hypothesize that perhaps glia cells in culture could take up and release GABA. Here we describe theuptake but not the release of [3H] GABA by puri-fied chick Müller glial cells in culture.
MATERIAL AND METHODS
MaterialsFertilized white Leghorn eggs were purchased from a local hatchery. The embryos were staged accord-ing to Hamburger and Hamilton (1951) and sacri-ficed by decapitation on embryonic day 9 (E9). The eyes were removed and the retinas dissected out ina Ca2+- and Mg2+-free Hanks' (CMF) solution. The retinas were then used for culture preparations.
[3H]GABA (Amersham Pharmacia Biotech,Piscataway, NJ, USA), GABA, glutamic acid,veratridine (Sigma, St. Louis, MO, USA) wereused throughout this work. GAT-1 (AB1570) and GAT-3 (AB1574) were from Chemicon. MinimumEssential Medium (MEM) and Dulbecco's modi-fied Eagle's medium (DMEM), fetal calf serum (FCS) and gentamycin were obtained from Gibco(USA). All other reagents were of analytical grade.
Retinal Müller Glia, Mixed Retinal Cultureand Neuron Enriched CulturesRetinal Müller glial cultures were generated fromE9 chick retina essentially as described before (Reiset al., 2002). Briefly, chick retina was dissociated using trypsin, and cells were seeded over 35 mmculture dishes in MEM containing 10% FCS. After a week, cell cultures were incubated with 4 mMascorbic acid for approximately 1.5 h to eliminate neurons. Cultures were subsequently washed withDMEM, and purified glial cultures were obtained after approximately 10 days and used when theyachieved confluence. Mixed retinal cultures were prepared from E8-E9
day-old retinas, seeded at approximately 30 x 106/2 ml in Minimum Essential Medium (MEM) plus5% FCS in 35-mm plastic culture dishes. Approx-imately 1 week after plating these cultures were used for the experiments. For neuron enriched cultures poly-L-lysine (10

GABA UPTAKE BY AVIAN MÜLLER GLIA 147
g/ml) coated plates were used. Cells were plated at a density of 2 x 106 cells/plate in DMEM con-taining 1% FCS to avoid glia proliferation. The medium was collected after 72 hours, filtered and used to feed purified Müller cultures.
GABA Uptake AssayAfter confluence, glial cultures or mixed dense cultures were rinsed with Hanks solution (128 mM NaCl, 4 mM KCl, 1 mM MgCl2, 3 mM CaCl2, 4 mM glucose, 20 mM HEPES) and subsequently incubated in Hanks solution with [3H]GABA (0.25
Ci/plate). The specific drugs used in the experi-ments were added 5 min before the [3H]GABA incubation: nipecotic acid (2 and 5 mM), beta alanine (10 mM), NNC-711 (50 and 100 M), zinc (100 M), amino-oxyacetic acid (AOAA, 2 and 5mM). Additional experiments were conducted in the absence of chloride (modified Hanks´ solution: 128 mM sodium acetate, 4 mM potassium acetate,1 mM magnesium sulfate, 3 mM calcium sulfate, 4mM glucose, 20 mM HEPES) or Na+ ions (modi-fied Hanks solution containing 128 mM Tris.HCl).
GABA ReleaseCultured glial or mixed retinal cells were incubated with 0.25 Ci of [3H]GABA for 90min in MEM containing 5 x 10-8 M of GABA buffered at pH 7.4with 20 mM HEPES. Non-incorporated radioactive GABA was removed by successive washings with modified Hanks solution (with 12 mM glucose). Then the cultures were superfused with 0.5 mlof Hanks solution (4 mM glucose) at 37°C and the radioactivity of each sample was determined. Greater than 90% of the radioactivity recovered inthe superfusates was characterized as GABA (doNascimento and de Mello, 1985). The radioactiv-ity released was plotted as the percentage of thetotal radioactivity taken up by the cells. The total radioactivity was estimated as that released during the superfusion plus the radioactivity remaining in the cells at the end of the procedure. The remaining cellular radioactivity was determined after disrup-tion of the cells in distilled water followed by suc-cessive freeze-thaw cycles.
Western BlotMüller glia cultures were washed twice in Phosphate Buffer Saline (PBS), homogenized in the presence
of protease inhibitors and subjected to sodium dodecyl sulfate (SDS) polyacrylamide gel electro-phoresis (PAGE). The proteins were transferred to nitrocellulose membranes, briefly stained withPonceau red, washed with an appropriate buffer and pre-incubated with 5% casein and 1% bovine serumalbumin (BSA) for 2 h at room temperature in ashaker. The membranes were rinsed with tris-buff-ered saline containing Tween and incubated with theprimary antibodies against the GABA transporters GAT-1 (1:500) and GAT-3 (1:500) at 4°C overnight under agitation. Secondary, horseradish peroxidase(HRP)-conjugated antibodies (Sigma) were used to
fdevelop the protein band corresponding to each of fthe expected molecular weights of the proteins of
interest, using ECL-plus (Pharmacia-Amersham).
Data AnalysisUnless specified otherwise, the results are expressed as mean ± standard error (SEM) of at least two sepa-rate experiments performed in triplicate. Statisticalanalysis was determined by Student's t test or tanalysis of variance (ANOVA) followed by post hoc Dunnett's test where appropriate, and statistical significance was taken at p <0.05.
RESULTS
The enrichment and purification of Müller glial cells in culture was confirmed by immunocyto-chemistry for glial markers vimentin or S100 as previously described (Reis et al., 2002). These cul-tures were also negative for Tuj1 antibody staining(a neuronal marker, data not shown). In most of the experiments performed, cultures were used at stage E9C14 (cultures prepared form 9-day-old embryosmaintained for 14 days in culture). [3H]GABA accumulation by Müller retinal glia was time dependent and reached equilibrium after 60 minutes (FIG. 1B). We also evaluated the satu-ration kinetics of GABA transport in Müller retinalglia incubating the cells with GABA concentrationsranging from 0.01 to 0.5 mM. Increasing GABA concentration resulted in increasing GABA uptake,reaching a half-maximal transport efficiency (EC50)around 0.3 mM (FIG. 1A). In all other experiments,we used a 90 min incubation period at a GABA concentration of 0.3 mM. Transporter-mediated GABA uptake in glial cells occurs in a chloride

C. S. SCHITINE et al.148
and sodium dependent manner (Amara and Arriza,1993). Accordingly, the uptake of [3H]GABA was completely inhibited in a Na+ free Hanks solution(FIGs. 1A, 1B). In order to characterize the ionic dependence of
the Müller cells [3H]GABA transporter, we inves-tigated the sodium and chloride dependence of GABA uptake. GABA uptake in control plates was approximately 10.68 ± 0.64 nmoles/mg of protein, which was reduced to about 6% of its value (0.64 ± 0.01 nmoles/mg protein) in the absence of sodium ions (FIG. 2A) and to about 10% of control values (1.07 ± 0.05 nmoles/mg of protein) in the absence of chloride ions (FIG. 2A). In order to test the temperature dependence, Müller retinal cultures were incubated at 37°C (control cul-tures) or at 0-4°C for 90 min with labeled GABA.One group of dishes was maintained at 0-4°C for 90 min, in the absence of GABA, then further incubated for an additional 90 min, at 37°C, with [3H]GABA. Approximately 10% of the uptake at 37°C was observed at 0-4°C, returning to 70% of
the control levels with the subsequent incubation at 37°C (FIG. 2B). This data also excludes cell dam-age at lower temperatures. To further characterize the GABA uptake trans-
porter in Müller glial cells in culture, nipecotic acid,a non-specific inhibitor, was incubated together with the radioactive ligand, and decreased GABA uptake to 25% of its control levels at the two differ-ent concentrations used (2 and 5 mM, FIG. 3). We also compared the effect of NNC-711 (also
referred as NO-711) on GABA uptake using mixed neuron-glia cells with the purified Müller glia cells in culture. As shown in Figure 4A, mixed
FIGURE 1 [3H]GABA uptake by Müller retinal gliafis time and concentration dependent. The uptake of
[3H]-GABA followed a temporal function reachingequilibrum after 60 min. Saturation kinetics of GABA transport by Müller retinal glia revealed a half-maximaltransport efficiency (EC50) around 0.3 mM (FIG. 1A).[3H]GABA uptake by Müller retinal glia was completelyblocked when Hanks solution had its sodium ionsexchanged for Tris (FIG. 1A, 1B).
FIGURE 2 [3H]GABA uptake by Müller glial cells issodium, chloride and temperature dependent. [3H]GABA uptake was completely blocked when Hanks solutionhad sodium ions substituted by Tris. Alternatively, Hanks solution had its chloride ions replaced by acetate(FIG. 2A). Müller retinal cell cultures were incubated at 37°C (control cultures), at 0-4°C or initially at 0-4°C and subsequently returned to 37°C, and GABA uptake was measured in these conditions (FIG. 2 B). GABA uptakein control plates was approximately 9.62 ± 0.64 nmoles/mg of protein.

GABA UPTAKE BY AVIAN MÜLLER GLIA 149
neuron-glia cultures incubated in the presence of 50 M NNC-711 decreased GABA uptake to about 10%
of its control values. On the other hand, neither 50M nor 100 M NNC-711 affected GABA uptake by
Müller retinal glia cultures (FIG. 4B). Therefore, even though purified Müller cells express GAT-1 (FIG. 5), known to be specifically blocked by NNC-711, this transporter seems not to be operating in these cells, as opposed to what was observed with neuron-glia cul-tures. Moreover the uptake of GABA was abolished when cultures were incubated in the presence of 100 μM ZnCl2 (FIG. 4C), a known inhibitor of mouse GAT 4 (GAT-3 in rats) transporter (Cohen-Kfir et al., 2005).It is worth noting that glia uptake of GABA accounted for only 20% of the uptake observed in neuron-gliamixed cultures.
Finally, beta-alanine, a specific glial inhibitor (Iversen and Kelly, 1975; Schon and Kelly, 1975; Schousboe, 2000; Schousboe et al., 2004), was tested on these cultures and as expected, [3H]GABA uptakewas inhibited in a concentration-dependent manner with 40% inhibition at 1 mM and 70% inhibition at 5mM concentration of beta-alanine (FIG. 4D). The expression of GAT-1 and GAT-3 was inves-
FIGURE 3 [3H]GABA uptake in Müller glia is blocked by nipecotic acid, a non-specific inhibitor of GABA transporters. Müller retinal cultures were incubated for 90 min in 1 ml Hanks solution with 0.3 mM [3H]GABA (0.25 Ci/plate) in the absence or presence of 2 and 5mM nipecotic acid. GABA uptake in control plates wasapproximately 9.72 ± 0.95 nmoles/mg of protein.
FIGURE 4 [3H]GABA uptake by Müller glia is blocked by different inhibitors of GABA transporters. Mixed neu-ronal-glia dense retinal cultures incubated in the presence of 50 M NNC-711 decreased GABA uptake to about 10% of its control values (FIG. 4A), whereas 50 M and 100 M NNC-711 had no effect on GABA uptake by Müller glialcultures (FIG. 4B). Beta-alanine, blocked [3H]GABA uptake in a concentration-dependent manner (FIG. 4D). GABA uptake in control plates for A, C and D was approximately 10.71 ± 0.71 nmoles/mg of protein; for B it was 48 ± 3nmoles/mg of protein.

C. S. SCHITINE et al.150
tigated using specific antibodies. Western blot analysis showed the expression of both GAT-1 and GAT-3 (FIG. 5) in Müller retinal glia that corre-sponded to the expected molecular weights. In order to evaluate whether these cultures could
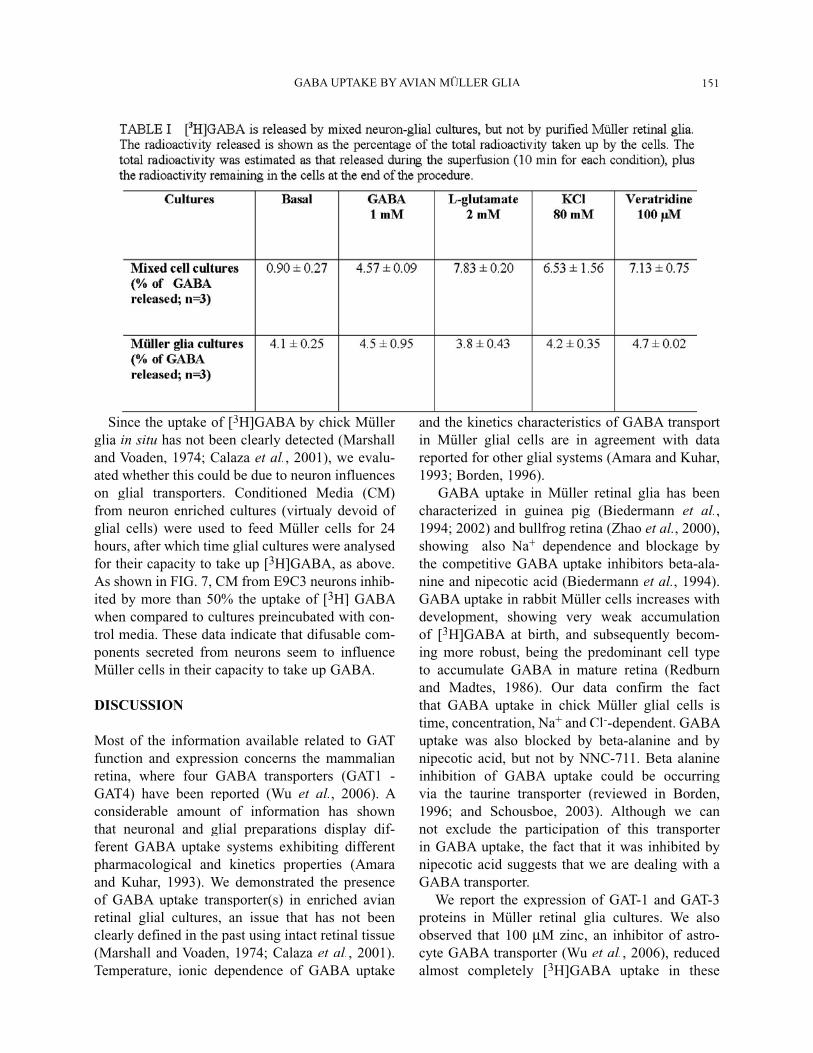
release GABA upon stimulation, we compared therelease of [3H]GABA from dense, mixed neuron-Müller cultures with the release from purified Müller cell cultures (Table I). As expected, depolariza-tion induced by KCl, glutamate or veratridine inmixed cultured cells showed a 7.0, 8.6 and 7.8-fold increase in GABA release, respectively. On the other hand, GABA-induced [3H]GABA release by homo-exchange increased 5-fold. However, none of thesestimuli elicited [3H]GABA release from Müller reti-nal glial cultures, suggesting that, different from neu-rons, induced [3H]GABA release mediated by selec-tive glial transporters is not a common mechanismoperating in the avian Müller cells. This idea wasreinforced by the observation that pre-incubation of glia cultures with 2 to 5 mM amino-oxyaceticacid (AOAA), an inhibitor of GABA transaminase, did not result in release of GABA (FIG. 6 A, 6B) promoted by either glutamate (2 mM) or GABA (3 mM). This indicates that the lack of GABA releasefrom glial cells is not due to a fast metabolization of the aminoacid inside the cells. It is worth notinghowever, that the percentage of basal, spontaneousrelease of [3H]-GABA is 4.5-fold higher in glialthan in neuron-glia mixed cultures.
FIGURE 5 Expression of GAT-1 and GAT-3 GABA transporters in cultured Müller glial cells. Representativeimmunoblots of GAT-1 (left) and GAT-3 (right). Membranes prepared from E8C13 Müller glia cultureswere evaluated by Western blot analysis using specific primary antibodies. Anti-GAT-1 and anti-GAT-3 specificantibodies recognized a strong band with an apparent MW of 66 kDa.
FIGURE 6 Amino-oxyacetic acid (AOAA) does not influence the capacity of Müller cells to release GABA.Cultures were pre-incubated with 2 or 5 mM AOAA then tested for [3H]GABA release as in Table I, always in the presence of AOAA.
FIGURE 7 Conditioned media (CM) from enriched neuronal retinal cultures decreases [3H]GABA uptake by Müller glia cells. Enriched neuron cultures (2 x 106
cells) were kept for 72 h in DMEM. CM was filtered and frozen at -70°C. Müller glia cultures were preincubated with control media or CM for 24 h. GABA uptake incontrol plates was 10.4 ± 1.0 nmoles/mg of protein.
GABA UPTAKE BY AVIAN MÜLLER GLIA 151
Since the uptake of [3H]GABA by chick Müller glia in situ has not been clearly detected (Marshall and Voaden, 1974; Calaza et al., 2001), we evalu-ated whether this could be due to neuron influenceson glial transporters. Conditioned Media (CM)from neuron enriched cultures (virtualy devoid of glial cells) were used to feed Müller cells for 24hours, after which time glial cultures were analysed for their capacity to take up [3H]GABA, as above. As shown in FIG. 7, CM from E9C3 neurons inhib-ited by more than 50% the uptake of [3H] GABA when compared to cultures preincubated with con-trol media. These data indicate that difusable com-ponents secreted from neurons seem to influenceMüller cells in their capacity to take up GABA.
DISCUSSION
Most of the information available related to GAT function and expression concerns the mammalianretina, where four GABA transporters (GAT1 - GAT4) have been reported (Wu et al., 2006). A considerable amount of information has shown that neuronal and glial preparations display dif-ferent GABA uptake systems exhibiting different pharmacological and kinetics properties (Amaraand Kuhar, 1993). We demonstrated the presenceof GABA uptake transporter(s) in enriched avianretinal glial cultures, an issue that has not beenclearly defined in the past using intact retinal tissue (Marshall and Voaden, 1974; Calaza et al., 2001). Temperature, ionic dependence of GABA uptake
and the kinetics characteristics of GABA transport in Müller glial cells are in agreement with datareported for other glial systems (Amara and Kuhar, 1993; Borden, 1996). GABA uptake in Müller retinal glia has been
characterized in guinea pig (Biedermann et al.,1994; 2002) and bullfrog retina (Zhao et al., 2000),showing also Na+ dependence and blockage by the competitive GABA uptake inhibitors beta-ala-nine and nipecotic acid (Biedermann et al., 1994). GABA uptake in rabbit Müller cells increases withdevelopment, showing very weak accumulationof [3H]GABA at birth, and subsequently becom-ing more robust, being the predominant cell type to accumulate GABA in mature retina (Redburn and Madtes, 1986). Our data confirm the fact that GABA uptake in chick Müller glial cells istime, concentration, Na+ and Cl--dependent. GABA uptake was also blocked by beta-alanine and bynipecotic acid, but not by NNC-711. Beta alanineinhibition of GABA uptake could be occurring via the taurine transporter (reviewed in Borden, 1996; and Schousboe, 2003). Although we can not exclude the participation of this transporter in GABA uptake, the fact that it was inhibited by nipecotic acid suggests that we are dealing with aGABA transporter. We report the expression of GAT-1 and GAT-3proteins in Müller retinal glia cultures. We also observed that 100 M zinc, an inhibitor of astro-cyte GABA transporter (Wu et al., 2006), reduced almost completely [3H]GABA uptake in these

C. S. SCHITINE et al.152
cells. Previous reports have shown that in primary cortical astrocyte cultures, zinc is selective for GAT-3 (Wu et al., 2006, nomenclature according to Guastella et al., 1990). Since GAT4 and GAT2 are GABA transporters known to be affected by zinc (Cohen-Kfir et al., 2005, nomenclature according to Liu et al., 1993), it is likely that GAT-3 is the predominant form of the transporter operating in our system (reviewed in Schousboe, 2003). We also show that, although Müller glia in culture can take up extracellular GABA, these cells do not release this compound after stimulation induced by glutamate, K+, veratridine or via GABA homo-exchange, suggesting that these cells are mainly buffering extracellular GABA. However, rat Müller cells do release [3H]GABA induced by high K+ and veratridine in a Ca2+ dependent manner. Also, ethylenediamine, a GABA mimetic, induces the release of GABA via a carrier-mediated exchange mechanism (Sarthy, 1983). Thus it seems that avian Müller cells behave quite differently, with respect to GABA release, as compared to mammalian retina. It is interesting to note that the chick Müller glia found in a histotypical preparation associated with retinal neurons are also incapable of taking up detectable levels of GABA as revealed by immunohistochem-istry (Calaza et al., 2001). This data points to a pos-sible neuronal regulation of the glial GAT. Recent data show that GAT is regulated through several mechanisms. GAT1 phosphorylation pro-moted by tyrosine kinase induces an increase in the number of transporters in the cell membrane and therefore, increase GABA uptake. Moreover, extra-cellular GABA decreases the level of GAT internal-ization thus increasing the amount of GABA uptake (Bernstein and Quick, 1999). On the other hand, phosphatase activators, tyrosine kinase blockers and PKC activation decrease GABA uptake (Beckman et al., 1998; Law et al., 2000; Quick, 2002). Indeed, conditioned media obtained from an enriched neu-ron retinal culture reduced in more than 50% the capacity of [3H]GABA uptake by purified Müller cells. This is consistent with the idea that, in situ, retina neurons may limit the capacity of the glial system to participate in GABA uptake. Our data also suggest that diffusable factors produced by neurons may play a role in this process. Some experimental data indicate that neuronal sig-nals influence Müller cell differentiation (reviewed
in Sarthy and Ripps, 2001). Since the control of GABA uptake by glial cells can be changed by neuronal environment in terms of excitability and excitotoxicity (Kim et al., 2003), the expression of GAT in Müller cells might be associated with the absence of neurons in the culture system used in this work. Müller glia has been shown to transdifferentiate in response to damage or to the influence of exog-enous growth factors (Fischer and Reh, 2001), sug-gesting that, perhaps, the uptake of GABA by these cells may reflect a rerouting program triggered by the purified glia cells in culture. This would be an interesting point to consider, since it can serve as a model for reactive gliosis, a process natu-rally occurring in lesioned nervous tissue. If under this process glial cells could increase its capacity for buffering GABA in lesioned sites, one could assume that gliosis could predispose excitotoxicity due to lower inhibitory environment created by the new condition.
AcknowledgementsThis work was supported by grants from FAPERJ, CNPq, PRONEX.
References
Amara S and J Arriza (1993) Neurotransmitter transporters: three distinct gene families. Curr. Opin. Neurobiol. 3, 337-344.
Amara S and MJ Kuhar (1993) Neurotransmitter transporters: recent progress. Annu. Rev. Neurosci. 16, 73-93.
Beckman ML, EM Bernstein and MW Quick (1998) Protein kinase C regulates the interaction between a GABA trans-porter and syntaxin 1A. J. Neurosci. 18, 6103-6112.
Bernstein EM and MW Quick (1999) Regulation of gamma-aminobutyric acid (GABA) transporters by extracellular GABA. J. Biol. Chem. 274, 889-895.
Biedermann B, W Eberhardt and W Reichelt (1994) GABA uptake into isolated retinal Müller glial cells of the guinea-pig detected electrophysiologically. Neuroreport 5, 438-440.
Biedermann B, A Bringmann and A Reichenbach (2002) High-affinity GABA uptake in retinal glial (Müller) cells of the guinea pig: electrophysiological characterization, immuno-histochemical localization, and modeling of efficiency. Glia 39, 217-228.
Birnbaum AD, SK Rohde, H Qian, MR Al-Ubaidi, JH Caldwell and RP Malchow (2005) Cloning, immunolocalization, and functional expression of a GABA transporter from the retina of the skate. Vis. Neurosci. 22, 211-223.
Borden LA (1996) GABA transporter heterogeneity: phar-

GABA UPTAKE BY AVIAN MÜLLER GLIA 153
macology and cellular localization. Neurochem. Int. 29, 335-356.
Brecha NC and C Weigmann (1994) Expression of GAT-1, a high-affinity gamma-aminobutyric acid plasma membrane transporter in the rat retina. J. Comp. Neurol. 345, 602-611.
Bringmann A, T Pannicke, J Grosche, M Francke, P Wiedemann, SN Skatchkov, NN Osborne and A Reichenbach (2006) Müller cells in the healthy and diseased retina. Prog. Retin. Eye Res. 25, 397-424.
Calaza KC, FG de Mello and PF Gardino (2001) GABA release induced by aspartate-mediated activation of NMDA receptors is modulated by dopamine in a selective subpopu-lation of amacrine cells. J. Neurocytol. 301, 81-93.
Calaza KC, PF Gardino and FG de Mello (2006) Transporter mediated GABA release in the retina: role of excitatory amino acids and dopamine. Neurochem. Int. 49, 769-777.
Casini G, DW Rickman and NC Brecha (2006) Expression of the gamma-aminobutyric acid (GABA) plasma mem-brane transporter-1 in monkey and human retina. Invest. Ophthalmol. Vis. Sci. 47, 1682-1690.
Cohen-Kfir E, W Lee, S Eskandari and N Nelson (2005) Zinc inhibition of gamma-aminobutyric acid transporter 4 (GAT4) reveals a link between excitatory and inhibitory neurotrans-mission. Proc. Natl. Acad. Sci. USA 102, 6154-6159.
do Nascimento JLM and FG de Mello (1985) Induced release of gamma-aminobutyric acid by a carrier-mediated, high-affinity uptake of L-glutamate in cultured chick retina cells. J. Neurochem. 45, 1820-1827.
Fischer AJ (2005) Neural regeneration in the chick retina. Prog. Retin. Eye Res. 24, 161-182.
Fischer AJ and TA Reh (2001) Müller glia are a potential source of neural regeneration in the postnatal chicken retina. Nat. Neurosci. 4, 247-252.
Guastella, J, N Nelson, H Nelson, L Czyzyk, S Keynan, MC Davidson, HA Lester and BI Kanner (1990) Cloning and expression of a rat brain GABA transporter. Science 249, 1303-1306.
Hamburger V and HL Hamilton (1951) A series of normal stages in the development of chick embryo. J. Morphol. 88, 49-92.
Iversen LL and JS Kelly (1975) Uptake and metabolism of gamma-aminobutyric acid by neurones and glial cells. Biochem. Pharmacol. 24, 933-938.
Kanner BI (1994) Sodium-coupled neurotransmitter transport: structure, function and regulation. J. Exp. Biol. 196, 237-249.
Kim D, MJ Kim, JH Lee, JO Im, YJ Won, S-Y Yoon and HN Hong (2003) Concomitant distribution shift of glial GABA transporter and S100 calcium-binding proteins in the rat retina after kainate-induced excitotoxic injury. Neurosci. Lett. 353, 17-20.
Kubrusly RCC, MCC Cunha, RAM Reis, H Soares, ALM Ventura, H Kurtenbach, MCF de Mello and FG de Mello (2005) Expression of functional receptors and transmitter enzymes in cultured Müller cells. Brain Res. 1038, 141-149.
Law RM, A Stafford and MW Quick (2000) Functional regu-lation of gamma-aminobutyric acid transporters by direct tyrosine phosphorylation. J. Biol. Chem. 275, 23986-23991.
Liu, Q-R, B López-Corcuera, S Mandiyan, H Nelson and N Nelson (1993) Molecular characterization of four pharmaco-logically distinct -aminobutyric acid transporters in mouse brain. J. Biol. Chem. 268, 2106-2112.
Marshall J and M Voaden (1974) An autoradiographic study of the cells accumulating 3H gamma-aminobutyric acid in the isolated retinas of pigeons and chickens. Invest. Ophthalmol. 13, 602-607.
Newman EA (2004) Glial modulation of synaptic transmission in the retina. Glia 47, 268-274.
Newman E and A Reichenbach (1996) The Müller cell: a func-tional element of the retina. Trends Neurosci. 19, 307-312.
Perea G and A Araque (2006) Synaptic information processing by astrocytes. J. Physiol. (Paris) 99, 92-97.
Pow DV, RKP Sullivan, SM Williams, HL Scott, PR Dodd and D Finkelstein (2005) Differential expression of the GABA transporters GAT-1 and GAT-3 in brains of rats, cats, mon-keys and humans. Cell Tissue Res. 320, 379-392.
Quick MW (2002) Substrates regulate gamma-aminobutyric acid transporters in a syntaxin 1A-dependent manner. Proc. Natl. Acad. Sci. USA 99, 5686-5691.
Redburn DA and P Madtes Jr (1986) Postnatal development of 3H-GABA-accumulating cells in rabbit retina. J. Comp. Neurol. 243, 41-57.
Reis RAM, MC Cabral da Silva, NE Loureiro dos Santos, E Bampton, JSH Taylor, FG de Mello and R Linden (2002) Sympathetic neuronal survival induced by retinal trophic factors. J. Neurobiol. 50, 13-23.
Sarthy PV (1983) Release of [3H]gamma-aminobutyric acid from glial (Müller) cells of the rat retina: effects of K+, veratridine, and ethylenediamine. J. Neurosci. 3, 2494-2503.
Sarthy V and H Ripps (2001) The Retinal Müller Cell - Structure and Function (Kluwer Academic:New York, NY) 278 pp.
Schon F and JS Kelly (1975) Selective uptake of [3H]beta-alanine by glia: association with glial uptake system for GABA. Brain Res. 86, 243-257.
Schousboe A (2000) Pharmacological and functional char-acterization of astrocytic GABA transport: a short review. Neurochem. Res. 25, 1241-1244.
Schousboe A (2003) Role of astrocytes in the maintenance and modulation of glutamatergic and GABAergic neurotrans-mission. Neurochem. Res. 28, 347-352.
Schousboe A, OM Larsson, A Sarup, and HS White (2004) Role of the betaine/GABA transporter (GAT-1/GAT2) for the control of epilepsy. Eur. J. Pharmacol. 500, 281-287.
Todd KJ, A Serrano, JC Lacaille and R Robitaille (2006) Glial cells in synaptic plasticity. J. Physiol. (Paris) 99, 75-83.
Won MH, TC Kang and SS Cho (2000) Glial cells in the bird retina: immunochemical detection. Microsc. Res. Tech. 50, 151-160.
Wu Q, M Wada, A Shimada, A Yamamoto and T Fujita (2006) Functional characterization of Zn2(+)-sensitive GABA transporter expressed in primary cultures of astrocytes from rat cerebral cortex. Brain Res. 1075, 100-109.
Zhao JW, JL Du, JS Li and XL Yang (2000) Expression of GABA transporters on bullfrog retinal Müller cells. Glia 31, 104-117.




















