
Effects of the dopamine D-1 antagonist SCH 23390 microinjected into the accumbens, amygdala or...
10
BRAIN RESEARCH ELSEVIER Brain Research 692 (1995) 47-56 Research report Effects of the dopamine D-1 antagonist SCH 23390 microinjected into the accumbens, amygdala or striatum on cocaine self-administration in the rat S. Barak Caine a, Stephen C. Heinrichs a, Vicki Lynn Coffin b, George F. Koob a,, Department of Neuropharmacology, The Scripps Research Institute, CVN- 7, 10666 N Torrev Pines Rd., La Jolla, CA 9203Z USA b Schering-Plough Research, Kenilworth. N J, USA Accepted 2 May 1995 Abstract This study tested the hypothesis that blockade of D-1 dopamine receptors in the nucleus accumbens shell, central nucleus of the amygdala or dorsal striatum by intracerebral microinjection of the dopamine antagonist SCH 23390 produces an attenuation of the effects of self-administered cocaine. Microinjection of SCH 23390 (0-4.0 p.g total dose) into any of the three brain regions dose-dependently increased the rate of cocaine self-administration, consistent with a partial attenuation of the effects of cocaine under these conditions (0.25 mg cocaine i.v.; fixed-ratio 5 timeout 20 s). The regional rank order potency of SCH 23390 was accumbens > amygdala > striatum, striatal injections being equipotent with subcutaneous administration. Moreover, SCH 23390 produced rapid effects on cocaine self-administration only when injected into the accumbens or amygdala. The time course of this regional selectivity was consistent with the rate of diffusion of SCH 23390 from the site of injection as measured by quantitative autoradiography, demonstrating that the regional selectivity of intracerebral injections of SCH 23390 is time-dependent. These results support a role for D-1 dopamine receptors in the nucleus accumbens and amygdala in the effects of self-administered cocaine, and suggest that D-1 receptors in certain portions of the 'extended amygdala' may be an important substrate for the reinforcing actions of cocaine. Keywords: Accumbens; Amygdala; Cocaine; Dopamine D-l; Reinforcement; SCH 23390; Self-administration; Striatum 1. Introduction It has been hypothesized that cocaine produces its rein- forcing effects by inhibiting the reuptake of dopamine and thereby increasing the extracellular concentration of dopamine in terminal regions of the mesocorticolimbic dopaminergic pathways [24,35,38]. Destruction of dopaminergic elements in the nucleus accumbens or ven- tral tegmental area by infusion of the catecholaminergic neurotoxin 6-hydroxydopamine decreased cocaine self-ad- ministration [39-41], but not responding maintained by other reinforcers such as food or heroin [8,34]. In some cases this decrease in cocaine self-administration may have been indicative of an attenuation of the reinforcing proper- ties of cocaine, since some animals exhibited patterns of behavior resembling extinction, where a high rate of re- sponding was followed by cessation of responding [40]. * Corresponding author. Fax: (44) (1223) 333564. 0006-8993/95/$09.50 © 1995 Elsevier Science B.V. All rights reserved SSDI 0006-8993(95)00598-6 Moreover, blockade of dopamine receptors by systemic administration of dopamine receptor antagonists has been shown to shift the cocaine self-administration dose-effect function to the right [5]. Thus blockade of dopamine receptors increases or decreases cocaine self-administra- tion in a manner similar to reducing the dose of cocaine under the dosage and scheduling conditions in effect [5,9,42,52,54]. The nucleus accumbens may be a critical neuroanatomical substrate for these effects, since local administration of dopamine receptor antagonists into the nucleus accumbens increased cocaine self-administration under dosage and scheduling conditions where such an increase would be consistent with an attenuation of the effects of cocaine [28,32,36]. More recently, the relative roles of dopamine receptors in the amygdala and accumbens in cocaine self-administra- tion have been compared. Local administration of the D-1 dopamine antagonist SCH 23390 into the amygdala in- creased the rate of cocaine (1.5 mg/kg) self-administration under a fixed-ratio schedule, an effect consistent with an attenuation of the effects of cocaine under those condi-
Transcript of Effects of the dopamine D-1 antagonist SCH 23390 microinjected into the accumbens, amygdala or...

BRAIN RESEARCH
E L S E V I E R Brain Research 692 (1995) 47-56
Research report
Effects of the dopamine D-1 antagonist SCH 23390 microinjected into the accumbens, amygdala or striatum on cocaine self-administration in the rat
S. Barak Caine a, Stephen C. Heinrichs a, Vicki Lynn Coffin b, George F. Koob a,, Department of Neuropharmacology, The Scripps Research Institute, CVN- 7, 10666 N Torrev Pines Rd., La Jolla, CA 9203Z USA
b Schering-Plough Research, Kenilworth. N J, USA
Accepted 2 May 1995
Abstract
This study tested the hypothesis that blockade of D-1 dopamine receptors in the nucleus accumbens shell, central nucleus of the amygdala or dorsal striatum by intracerebral microinjection of the dopamine antagonist SCH 23390 produces an attenuation of the effects of self-administered cocaine. Microinjection of SCH 23390 (0-4.0 p.g total dose) into any of the three brain regions dose-dependently increased the rate of cocaine self-administration, consistent with a partial attenuation of the effects of cocaine under these conditions (0.25 mg cocaine i.v.; fixed-ratio 5 timeout 20 s). The regional rank order potency of SCH 23390 was accumbens > amygdala > striatum, striatal injections being equipotent with subcutaneous administration. Moreover, SCH 23390 produced rapid effects on cocaine self-administration only when injected into the accumbens or amygdala. The time course of this regional selectivity was consistent with the rate of diffusion of SCH 23390 from the site of injection as measured by quantitative autoradiography, demonstrating that the regional selectivity of intracerebral injections of SCH 23390 is time-dependent. These results support a role for D-1 dopamine receptors in the nucleus accumbens and amygdala in the effects of self-administered cocaine, and suggest that D-1 receptors in certain portions of the 'extended amygdala' may be an important substrate for the reinforcing actions of cocaine.
Keywords: Accumbens; Amygdala; Cocaine; Dopamine D-l; Reinforcement; SCH 23390; Self-administration; Striatum
1. Introduction
It has been hypothesized that cocaine produces its rein- forcing effects by inhibiting the reuptake of dopamine and thereby increasing the extracellular concentration of dopamine in terminal regions of the mesocorticolimbic dopaminergic pathways [24,35,38]. Destruction of dopaminergic elements in the nucleus accumbens or ven- tral tegmental area by infusion of the catecholaminergic neurotoxin 6-hydroxydopamine decreased cocaine self-ad- ministration [39-41], but not responding maintained by other reinforcers such as food or heroin [8,34]. In some cases this decrease in cocaine self-administration may have been indicative of an attenuation of the reinforcing proper- ties of cocaine, since some animals exhibited patterns of behavior resembling extinction, where a high rate of re- sponding was followed by cessation of responding [40].
* Corresponding author. Fax: (44) (1223) 333564.
0006-8993/95/$09.50 © 1995 Elsevier Science B.V. All rights reserved SSDI 0006-8993(95)00598-6
Moreover, blockade of dopamine receptors by systemic administration of dopamine receptor antagonists has been shown to shift the cocaine self-administration dose-effect function to the right [5]. Thus blockade of dopamine receptors increases or decreases cocaine self-administra- tion in a manner similar to reducing the dose of cocaine under the dosage and scheduling conditions in effect [5,9,42,52,54]. The nucleus accumbens may be a critical neuroanatomical substrate for these effects, since local administration of dopamine receptor antagonists into the nucleus accumbens increased cocaine self-administration under dosage and scheduling conditions where such an increase would be consistent with an attenuation of the effects of cocaine [28,32,36].
More recently, the relative roles of dopamine receptors in the amygdala and accumbens in cocaine self-administra- tion have been compared. Local administration of the D-1 dopamine antagonist SCH 23390 into the amygdala in- creased the rate of cocaine (1.5 mg/kg) self-administration under a fixed-ratio schedule, an effect consistent with an attenuation of the effects of cocaine under those condi-

48 S.B. Caine et al. / Brain Research 692 (1995) 47-56
tions, however the same treatment failed to alter the break- ing point for this dose of cocaine under a progressive-ratio schedule [32]. In contrast to intra-amygdala SCH 23390, intra-accumbens SCH 23390 significantly decreased the breaking point under the progressive-ratio schedule, an effect also consistent with an attenuation of the effects of cocaine, but produced lesser effects on cocaine self-admin- istration under the fixed-ratio schedule [32]. Based on these results, the authors proposed different roles for amygdala and accumbens in cocaine self-administration behavior, such that dopamine receptors in the amygdala may be involved in the interoceptive stimulus properties of cocaine, while dopamine receptors in the accumbens are preferentially involved in the reinforcing effects of cocaine [32].
Though intriguing, the behavioral endpoints measured in that study often occurred over several hours following intracerebral administration. Moreover, injections of SCH 23390 into either site produced equivalent decreases in locomotor activity, and there was no demonstration of a neuroanatomical site completely insensitive to the effects of SCH 23390 administration [32]. These issues raise concern about the regional selectivity and therefore inter- pretation of the effects of intracerebral administration of a highly lipophilic compound such as SCH 23390 on co- caine self-administration behavior.
The present study therefore examined the effects of local administration of SCH 23390 into the accumbens, amygdala or dorsal striatum on cocaine self-administration under a fixed-ratio schedule. The shell of the accumbens and central nucleus of the amygdala were chosen as targets for several reasons. First, these are the subregions of densest dopaminergic innervation within these structures [12,14,15,51]. Second, the accumbens shell has received considerable attention with regard to converging afferent inputs from limbic structures [17,21,22,45] as well as significant efferent connections to limbic forebrain regions [18] and mesencephalic dopaminergic cell groups [4]. Fi- nally, it has been proposed that the accumbens shell and central amygdala represent the borders of a large forebrain continuum termed the extended amygdala [1,18,20] and that they may share, by virtue of anatomical and neuro- chemical similarities, functional attributes.
The dorsal striatum was chosen as a neuroanatomical control equidistant between the accumbens shell and cen- tral amygdala, since it also has a rich dopaminergic inner- vation [26] but has not been implicated in the reinforcing properties of cocaine [28,36,41]. A fixed-ratio schedule was chosen so that the time course of alterations in behav- ior maintained under constant schedule conditions could be evaluated. In addition to comparing the effects of SCH 23390 in different brain regions, the regional selectivity of intracerebral SCH 23390 administration was further evalu- ated by measuring the time course of diffusion of locally administered SCH 23390 using quantitative autoradio- graphic methods.
2. Materials and methods
2.1. Subjects
Thirty male albino Wistar rats (Charles River, Kingston, NY) weighing 260-300 g upon arrival were used for this study. Eighteen subjects were used to generate the self-ad- ministration studies presented. An additional twelve ani- mals were used for quantitative autoradiography studies. Upon arrival animals were group-housed (2-3), main- tained in a temperature- and light-controlled environment (12:12 h L / D ) and handled daily. All behavioral testing occurred during the dark phase of the light/dark cycle. The rats had free access to food and water except during daily 3 h test sessions.
2.2. Catheterization
One week after arrival, rats were anesthetized with a halothane-oxygen vapor mixture and implanted with chronic intravenous (i.v.) jugular catheters as described previously [7]. Briefly, the catheters consisted of poly- ethylene tubing glued to a guide cannula bent at a right angle and encased in dental cement anchored with a 1-inch square of durable mesh. The tubing was passed subcuta- neously from the animals's mid-scapular region to the right external jugular vein, where it was inserted and secured gently with suture thread. All animals were al- lowed to recover for a minimum of 4 days before given access to cocaine. Catheters were flushed with saline con- taining heparin (30 USP U/ml) daily, as well as before and after self-administration sessions. When not in use, a threaded stylet cap was affixed to the guide cannula. The integrity of the catheter was tested whenever an animal not receiving drug pretreatments displayed behavior outside baseline parameters. Briefly, 0.1 ml of the ultrashort-acting barbiturate anesthetic Brevital Sodium (1% methohexital sodium, Eli Lilly, Indianapolis, IN) was administered through the catheter. Animals with patent catheters exhibit prominent signs of anesthesia (pronounced loss of muscle tone) within 3 s of i.v. injection. Animals with faulty catheters were recatheterized on the opposite jugular vein. Pretreatment tests were repeated when recatheterization was necessary in order to reinstate baseline behavior after a pretreatment test.
2.3. Drugs
Cocaine HC1 was obtained from Sigma Chemical Co. (St. Louis, MO) and dissolved in saline (2.5 mg/ml). SCH 23390 maleate was supplied by Schering-Plough (Kenil- worth, NJ) and dissolved in methanol to insure dissolution, then diluted with saline to a minimum 20:1 saline/methanol ratio. A 20:1 saline/methanol solution was used for vehicle treatments (zero dose, Fig. 1).

S.B. Caine et al. /Brain Research 692 (1995) 47-56 49
[3H]SCH 23390 (specific activity 72.8 Ci /mmol) was obtained from Dupont NEN Products (Boston, MA).
2.4. Self-administration
The rats were given access to cocaine (0.25 mg/0 .1 m l / 4 s) under a fixed-ratio (FR) 1 timeout (TO) 20 s schedule for 3 h daily. Each session was preceded by the automated delivery of two reinforcers prior to availability of the lever. The completion of the FR requirement re- suited in activation of a white cue light above the lever which remained on during the 4 s infusion and for the remainder of the 20 s timeout period, during which re- sponses were recorded but were not reinforced. Animals that did not respond at least once per hour were given four noncontingent injections at the beginning of each hour on alternate days during training. When reliable responding was observed, the fixed-ratio was increased to a value of 5. After baseline levels of self-administration were achieved under this schedule (criterion of < 10% deviation from the mean of the total reinforcers earned in three consecutive sessions for each rat) the animals were prepared with chronic indwelling intracerebral cannulae.
2.5. lntracerebral cannulations
Animals were anesthetized with a halothane/oxygen vapor mixture and placed in a Kopf stereotaxic instrument with the incisor bar set 3.3 mm below the interaural line (skull flat), and coordinates were chosen with the aid of a standard brain atlas [33]. Stainless steel 23 gauge intracere- bral cannulae 10 mm in length were aimed 3 mm above the target sites and implanted bilaterally using the follow- ing coordinates from Bregma (AP), midline (ML) and skull surface (DV): Nucleus accumbens shell, AP + 1.7 ML + 1.0 DV -5 .0 ; central nucleus of the amygdala, AP +2.5 ML +4.2 DV -5 .1 ; dorsal striatum, AP - 0 . 9 ML +_4.4 DV -3 .4 . Intracerebral cannulae were secured to the skull using dental cement anchored with four skull screws. Removable stylet wire (10 mm) maintained pa- tency of the intracerebral cannulae between injections. After 4 days recovery from surgery, the animals were again given access to cocaine 3 h daily, beginning approxi- mately 1-2 h after the start of the dark phase of the l ight/dark cycle.
2.6. Intracerebral microinjections
Local administrations of SCH 23390 were made by replacing the stylet wire with a 30 gauge needle fashioned to extend 3 mm beyond the tip of the intracerebral cannula. Injection volume was 0.33 /xl per side, infused over 28 s using a Hamilton microsyringe connected to the needle via polyethylene tubing. Needles were left in place for 30 s following the infusion, and then replaced with the wire stylet, after which the animal was immediately placed in
the self-administration chamber for testing. After the in- tracerebral cannulation surgery animals were re-baselined (at least two consecutive days _+ 10% pre-surgery baseline or three consecutive days + 10% total reinforcers earned per session) prior to pretreatment tests. The animals were re-tested in this fashion (with a return to baseline or establishment of a new baseline criterion for a new pre- treatment test) until all doses (0, 0.5, 1.0, 2 .0 /xg/s ide = 0, 1.0, 2.0, 4.0 /zg total dose administered) were tested in a Latin square within-subjects design (i.e., each subject re- ceived each dose once).
2.7. Quantitati~e autoradiography - - intracerebral [3H]SCH 23390
Eighteen animals were used for quantitative autoradiog- raphy. Ten animals with intracerebral cannulae aimed at the central nucleus of the amygdala were used 1 week after they had been used for a separate study where they were injected once with purified water (0.5 /xl/side) and tested as a control group in an anxiolytic evaluation behavioral paradigm. These animals were injected with 1.0 /xM [3H]SCH 23390 (0.33 /xl/side) as described above, and sacrificed at 2, 10, 20, 60, 120 or 180 min in order to estimate the diffusion of [3H]SCH 23390 from the injec- tion site by quantitative autoradiography. Briefly, the ani- mals were decapitated and the brains rapidly removed and fresh-frozen in powdered dry ice. They were then stored at - 7 0 ° C until sections (30 /.~m) were cut on a cryotome ( - 1 6 ° C ) and thaw-mounted on gelatin-chromate coated slides. The slides were air-dried and apposed to tritium- sensitive film (Amersham, Arlington Heights, IL) and placed in light-tight cassettes for 2 weeks, then re-apposed to new film for 3 months. The autoradiographs were developed with standard chemicals and the optical densi- ties analyzed using a microcomputer based image analysis system.
2.8. Quantitati~,e autoradiography - - in tracenous [3H]SCH 23390
Because the intracerebrally administered [3H]SCH 23390 used above for estimating diffusion was signifi- cantly lower in concentration than that used for the behav- ioral studies, an additional complementary approach was employed to measure diffusion. Immediately following intracerebral injection of SCH 23390 (1.0 /xg/0.33 /xl/side) into the accumbens shell, central amygdala or dorsal striatum, 33 /zCi of [3H]SCH 23390 was adminis- tered i.v. to label D-1 receptors in vivo. Previous work in rats suggested that the specific binding of the radioligand would peak at approximately 1 h post-injection [23]. Thus in order to estimate the diffusion of intracerebrally admin- istered SCH 23390 via the local protection of D-1 recep- tors from binding i.v. [3H]SCH 23390, the diffusion could not be measured at time points shorter than 1 h.

50 S.B. Caine et al. / Brain Research 692 (1995) 47-56
Six of the animals used for self-administration (two from each brain region group) were subsequently used for estimating the diffusion of intracerebral SCH 23390 from the injection site. Animals were injected intracerebrally with SCH 23390 (1.0 /xg /0 .33 /x l / s ide ) as in the behav- ioral tests, and immediately thereafter injected i.v. with 33 /xCi of [3H]SCH 23390 to label D-1 dopamine receptors in vivo as previously described [2,3,23]. Two additional ani- mals with similar cocaine self-administration experience, but lacking intracerebral cannulae, were also used for control labeling of D-1 receptors by i.v. [3H]SCH 23390 and to evaluate the effects of s.c. pretreatment with SCH 23390 ( 2 0 / z g / k g ) on the in vivo labeling of D-1 receptors by i.v. [3H]SCH 23390. Al l animals used for in vivo labeling of D-1 receptors with i.v. [3H]SCH 23390 were sacrificed 60 rain fol lowing radioligand administration. Ten tritium standards were made by homogenizing 0.6 ml aliquots of rat brain mash with increasing concentrations of tritium and fresh-freezing the mashes for sectioning at
B
ccSh
CeA
A SCH23390: FIRST 20 MIN
~ Z
m PUJ z z I,LI--
w O
250
E
0 1 2 4
TOTAL DOSE SCH23390 (!~g i.c.)
SCH23390: ENTIRE 3 HR SESSION
.k
,Ic f_. ~= 2o0 * ~ Z
Z ~
u lU, ,
mm
I - ~ 1 1 0 0 z z
0 1 2 4
T O T A L DOSE SCH23390 (p-g i.c.)
[ ] AccSh [ ] CeA [] CPu
[] AccSh [] CeA • CPu
CPu
Fig. 1. A. Effects of SCH 23390 (0, 0.5, 1.0, 2.0 p,g/0.33 /.d/side = 0, 1.0, 2.0, 4.0 total dose) microinjected into the accumbens shell (AccSh), central amygdala (CeA) or dorsal striatum (CPu) on cocaine self-adminis- tration (0.25 mg i.v.; FR 5 TO 20 sec) in separate groups of rats (values are group means and standard errors, n = 6/brain region). Top, effects on self-administration in the first 20 min of the session. Bottom, effects on self-administration over the entire 3 h session. Asterisks indicate signifi- cantly different from vehicle injection (* p < 0.05; * * p < 0.01) by Neuman Keuls a posteriori test following significant main effect of dose by ANOVA. B. Top three panels, location of ventral most extent of the injector tracks in the animals used for the data in Fig. 1A. Bottom panel, nissl stained section showing the injector tracks (see arrows) and associ- ated tissue damage for a representative animal in the accumbens shell group.
Fig. 1 (continued).
30 /xm. After scintillation counting of these samples, an image analysis system was used to generate an optical density-concentration function from the autoradiographic images produced by the standards that were included in each film cassette. Optical densities of all autoradiographic images were determined from three measurements of each image on the image analysis system.

S.B. Caine et al. / Brain Research 692 (1995) 47-56 51
2.9. Histology
Animals not used for quantitative autoradiography were euthanized by i.p. injection of pentobarbital sodium (100 mg/kg) followed by intracardiac infusion with 50 ml of 4% formalin. Brains were removed and stored in 4% formalin and subsequently sectioned coronally (52 /zm thickness) to localize the ventral most extent of the injector tracks for recording on a brain map (Fig. 1B).
2.10. Analyses
Self-administration data from pretreatment tests were calculated as the percent of the immediately preceding baseline rate of cocaine self-administration, and each ex- perimental group (accumbens, amygdala, striatum) was analyzed using ANOVA with repeated measures on dose (SCH 23390). Following a significant main effect of dose, individual comparisons of single doses with the vehicle were performed using Neuman-Keul's a posteriori test. Potency comparisons for different routes of administration were calculated using a computer program that calculates relative potencies and 95% confidence limits using a paral- lel line assay of the dose-effect functions of the individual data points [49]. Quantitative autoradiographic data (Sec- tion 3.3) were analyzed by two-way ANOVA on intracere- bral injection site and brain region measured, and signifi- cant main effects were further explored using Dunnett's test for comparing all treatments with a control.
3. Results
3.1. Self-administration studies
When microinjected into the accumbens shell or central amygdala, SCH 23390 produced a dose-dependent increase in cocaine self-administration (0.25 mg i.v.) which was observable within the first 20 min of the session (Fig. 1, top: Accumbens, F3,15 = 11.3, P < 0.001; amygdala, F3,15
= 3.8, P < 0.05). In contrast, injections into the dorsal striatum failed to alter cocaine self-administration in the first 20 rain of the session (F3,15 = 0.9, P > 0.5). Injec- tions into all three brain regions dose-dependently in- creased the rate of cocaine self-administration over 3 h (Fig. 1, bottom: Accumbens, F3,t5 = 14.4, P<0 .0 0 1 ; amygdala, F3.L~ = 7.0, P < 0.01; striatum, F3,15 = 8.2, P < 0.01). However, a higher dose of SCH 23390 (4.0 k~g total) was necessary to produce this effect when adminis- tered into the dorsal striatum (4.0 /zg total, P < 0.01) compared with the accumbens or amygdala (2.0 /xg total, P < 0.05). The localization of the ventral most extent of the injector tracks, and a photomicrograph of the typical extent of tissue damage produced during the study are shown in Fig. lB.
A Exposure Time 2 wk 3 mos
2 min ~!iii!i;iil
l O m i n i ~ , ~
20 min iii i~i i I
60 min
120 rain
180 min
B
rr
I . - -
t U
o a . J
ta t l
z O m
O f t
i i
i i
E O
o B
DIFFUSION OF [3H]-SCH23390 F R O M INTRACRANIAL INJECTION SITE
3 -
~ a ........................................ :'~'~ ........................... U'""ie size of the
central nucleus of the amygdala
, i • i • i , i • | • m , i
20 40 60 80 1 O0 120 140 TIME AFTER INJECTION (min)
C O N C E N T R A T I O N OF [3H]-SCH23390 AT INTRACRANIAL INJECTION SITE 0 . 5 - -z 0.4-
O o.3
5 , I"' 0.2- Z I,U o Z 0.1 o o
0.0 . ,o 2'0 ; , o ~ ' o ~ ; ~ o
TIME AFTER INJECTION (rain)
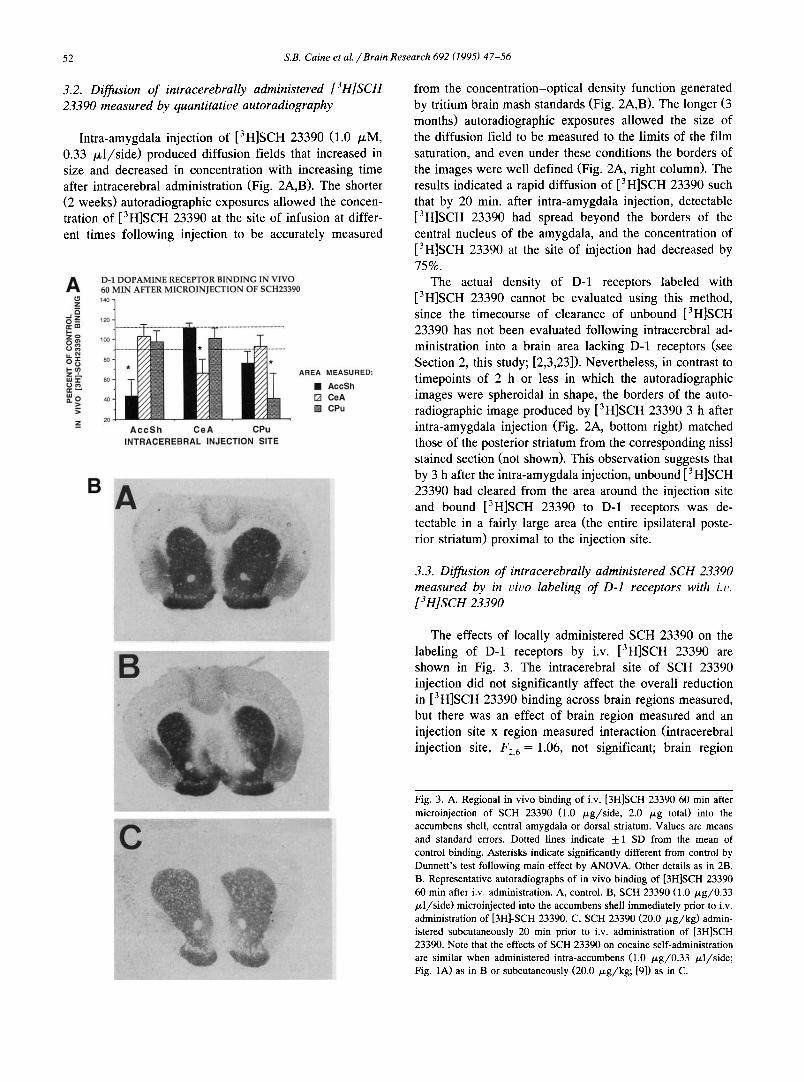
Fig. 2. A. Autoradiographs showing diffusion of [3H]SCH 23390 at different timepoints following intra-amygdala administration (1.0 ~M, 0.33 txl/28 sec). Note detection of the radioligand throughout the posterior striatum 3 hr after injection (bottom right). Shown actual size. B. Diffusion (ram 3) and concentration of [3H]SCH 23390 (~M) at different timepoints following intra-amygdala administration• Autoradio- graphs were analyzed using a microcomputer based image processor and molar values were calculated using tritium brain mash standards included in each film cassette. Other details as in 2A.
52 S.B. Caine et al. /Brain Research 692 (1995) 47-56
3.2. Diffusion of intracerebrally administered [3H]SCH 23390 measured by quantitative autoradiography
Intra-amygdala injection of [3H]SCH 23390 (1.0 /zM, 0.33 /zl /s ide) produced diffusion fields that increased in size and decreased in concentration with increasing time after intracerebral administration (Fig. 2A,B). The shorter (2 weeks) autoradiographic exposures allowed the concen- tration of [3H]SCH 23390 at the site of infusion at differ- ent times following injection to be accurately measured
14o
12o ...................................................................................................
1oo * .......................
8o * .
2O _z A c c S h CeA CPu
INTRACEREBRAL INJECTION SITE
D-1 DOPAMINE RECEPTOR BINDING IN VIVO 60 MIN AFTER MICROINJECTION OF SCFI23390
A R E A M E A S U R E D :
[] AccSh [] CeA [] CPu
from the concentration-optical density function generated by tritium brain mash standards (Fig. 2A,B). The longer (3 months) autoradiographic exposures allowed the size of the diffusion field to be measured to the limits of the film saturation, and even under these conditions the borders of the images were well defined (Fig. 2A, right column). The results indicated a rapid diffusion of [3H]SCH 23390 such that by 20 min. after intra-amygdala injection, detectable [3H]SCH 23390 had spread beyond the borders of the central nucleus of the amygdala, and the concentration of [3H]SCH 23390 at the site of injection had decreased by 75%.
The actual density of D-1 receptors labeled with [3H]SCH 23390 cannot be evaluated using this method, since the timecourse of clearance of unbound [3H]SCH 23390 has not been evaluated following intracerebral ad- ministration into a brain area lacking D-1 receptors (see Section 2, this study; [2,3,23]). Nevertheless, in contrast to timepoints of 2 h or less in which the autoradiographic images were spheroidal in shape, the borders of the auto- radiographic image produced by [3H]SCH 23390 3 h after intra-amygdala injection (Fig. 2A, bottom right) matched those of the posterior striatum from the corresponding nissl stained section (not shown). This observation suggests that by 3 h after the intra-amygdala injection, unbound [3H]SCH 23390 had cleared from the area around the injection site and bound [3H]SCH 23390 to D-1 receptors was de- tectable in a fairly large area (the entire ipsilateral poste- rior striatum) proximal to the injection site.
3.3. Diffusion of intracerebrally administered SCH 23390 measured by in civo labeling of D-1 receptors with i.~,. [3H]SCH 23390
B
C
The effects of locally administered SCH 23390 on the labeling of D-1 receptors by i.v. [3H]SCH 23390 are shown in Fig. 3. The intracerebral site of SCH 23390 injection did not significantly affect the overall reduction in [3H]SCH 23390 binding across brain regions measured, but there was an effect of brain region measured and an injection site x region measured interaction (intracerebral injection site, F2, 6 = 1.06, not significant; brain region
Fig. 3. A. Regional in vivo binding of i.v. [3H]SCH 23390 60 min after microinjection of SCH 23390 (1.0 /xg/side, 2.0 /.Lg total) into the accumbens shell, central amygdala or dorsal striatum. Values are means and standard errors. Dotted lines indicate 4-1 SD from the mean of control binding. Asterisks indicate significantly different from control by Dunnett's test following main effect by ANOVA. Other details as in 2B. B. Representative autoradiographs of in vivo binding of [3H]SCH 23390 60 min after i.v. administration. A, control. B, SCH 23390 (1.0 /xg/0.33 /xl/side) microinjected into the accumbens shell immediately prior to i.v. administration of [3H]-SCH 23390. C, SCH 23390 (20.0 /xg/kg) admin- istered subcutaneously 20 min prior to i.v. administration of [3H]SCH 23390. Note that the effects of SCH 23390 on cocaine self-administration are similar when administered intra-accumbens (1.0 /~g/0.33 /xl/side; Fig. 1A) as in B or subcutaneously (20.0 ~g/kg; [9]) as in C.

S.B. Caine et al. /Bra in Research 692 (1995) 47-56 53
measured, F2, 6 = 6.23, P < 0.01; injection site × region measured interaction, F4,12 = 72.09, P < 0.001). More- over, the binding of i.v. administered [3H]SCH 23390 was selectively protected in the region injected without altering radiolabeling in the other regions (see Fig. 3A, asterisks indicate significantly different from control by independent comparison, P < 0.05).
The diffusion of intracerebral SCH 23390 (1.0/xg/0.33 /xl/side) 1 h following injection (2.2 _+ 0.5 mm3/side) was similar to that estimated using intracerebral adminis- tration of [3H]SCH 23390 (see Section 3.2). It is notable that the diffusion of SCH 23390 1 h after intracerebral administration is markedly restricted compared with the effect of s.c. pretreatment with SCH 23390 (20 /xg/kg) on the in vivo binding of i.v. [3H]SCH 23390 (Fig. 3B), given that these two treatments produce comparable effects on cocaine self-administration (Fig. 1, this study; [9]).
3.4. Relative potency of SCH 23390 by intracerebral ad- ministration t, ersus subcutaneous administration
The dose-effect data for SCH 23390 administered into the three brain regions was compared to previously re- ported data on the effects of SCH 23390 administered subcutaneously on cocaine self-administration under iden- tical conditions (0.25 mg i.v.; FR 5 TO 20 sec) [9]. The linear regression plots for the dose-effect data by each route of administration were subjected to a parallel line assay and relative potency values were calculated [49]. The potency of SCH 23390 administered s.c. [9] was assigned a relative potency value of 1.0, against which the other routes of administration were compared. The relative po- tency values in Table 1 therefore reflect the ratio of the amount of SCH 23390 by each route of administration required to produce an equivalent effect to that of a dose given by subcutaneous administration over the entire 3 h session. Intra-accumbens or intra-amygdala but not intra- striatal administrations were significantly more potent than subcutaneous administration (P < 0.05), although the max- imum difference (accumbens versus subcutaneous) repre- sented only an approximate twofold increase in potency.
Table 1 Regional potency of intracerebral SCH 23390 relative to subcutaneous administration a
Region Potency ratio Significance
Accumbens shell 0.45 P < 0.05 Central amygdala 0.69 P < 0.05 Caudate-putamen 0.96 P > 0.1
a The potency ratio shown refers to the ratio of the amount of drug necessary to produce an equivalent effect to some standard assigned a value of unity [49]. Thus 0.45 /~g in the accumbens shell or 0.69 p,g in the central amygdala produces an effect equivalent to that produced by 1.0 ,u,g administered subcutaneously.
Thus intra-accumbens administration of 4.0 /zg total (2.0 /zg/side) of SCH 23390 produced an effect identical to that observed after administration of 20.0 /zg/kg s.c. (8.0 /xg total for the average bodyweight of 0.4 kg/subject).
4. Discussion
Intra-accumbens or intra-amygdala administration of the D-1 antagonist SCH 23390 produced a rapid increase in the rate of cocaine self-administration with a potency slightly but significantly greater than systemic administra- tion. In contrast, intra-striatal SCH 23390 did not increase cocaine self-administration within the first 20 min of the session, and the potency of intra-striatal SCH 23390 was equivalent to subcutaneous administration of the antago- nist. In addition to the behavioral data, autoradiographic measures indicated that the regional selectivity of D-1 receptor blockade following intracerebral administration of SCH 23390 is time-dependent (e.g., exceeding the borders of the central amygdala within 20 min). Taken together, these data support the hypothesis that i.v. cocaine self-ad- ministration in the rat can be modulated through D-1 dopamine receptors in the nucleus accumbens or amyg- dala.
Increased cocaine self-administration after intra-accu- mbens or intra-amygdala administration of a dopamine antagonist have been observed previously [28,32,36]. Such an increase in self-administration rate is consistent with a shift to the right of the cocaine dose-effect function under these conditions [9,25]. Indeed a shift to the right in the entire cocaine self-administration dose-effect function has been demonstrated following systemic administration of SCH 23390 [5,10]. Nevertheless it remains to be conclu- sively determined that blockade of D-1 receptors in the accumbens or amygdala produces a surmountable antago- nism of the effects of self-administered cocaine using a range of cocaine doses. Such a demonstration may be particularly important, since lesion studies of specific ter- minal regions of the mesocorticolimbic dopamine path- ways, such as the medial prefrontal cortex and the amyg- dala, have identified effects on cocaine self-administration only at lower or higher doses of cocaine, respectively [29,31,44].
It also remains to be determined that changes in cocaine self-administration under rate-dependent schedules reflect changes specifically in the reinforcing effects of cocaine. Cocaine produces changes in schedule-controlled behavior maintained by other reinforcers [48,53], and such changes are reversed by dopamine receptor antagonists [46]. More- over, and of particular relevance to the present study, others have proposed that dopaminergic receptor blockade in the amygdala alters the rate of cocaine self-administra- tion by altering the interoceptive stimulus, but not the reinforcing, effects of cocaine [32].
Several additional issues warrant further consideration

54 S.B. Caine et al. / Brain Research 692 (1995) 47-56
regarding interpretation of the present data. First, it is perhaps surprising that intracerebral SCH 23390 in this study had a maximum potency only two fold greater than that of subcutaneously administered SCH 23390. One pos- sible explanation is that the critical neuroanatomical loci for the effects of SCH 23390 on cocaine self-administra- tion have yet to be identified. An alternative explanation is that multiple neuroanatomical loci of D-1 receptors are involved in cocaine self-administration, and that the simul- taneous blockade of these sites by systemic administration of SCH 23390 contributes to attenuate the behavioral effects of cocaine at relatively low doses of the systemi- cally administered antagonist. A third possibility comes from previous work demonstrating a mutual antagonism between dopamine (indirectly increased by cocaine) and dopamine receptor antagonists in cocaine self-administra- tion [5,10] as well as in other behavioral assays [19,46,47,50]. Thus it is possible that significant concentra- tions of SCH 23390, even locally, may be necessary to compete with the effects of cocaine to indirectly activate D-1 receptors. It should be noted that the dose-related effects of intracerebral SCH 23390 observed here are in good agreement with previous studies [28,32].
A further consideration regards the measurement of diffusion of SCH 23390 by the autoradiographic methods employed here. Although the time-related diffusion of [3H]SCH 23390 following direct intracerebral administra- tion suggested a local action (< 2.0 mm 3) of SCH 23390 in the first hour following microinjections of the antago- nist, the radioligand concentration was significantly lower than that used in the behavioral studies. However, when the appropriate concentration of SCH 23390 was microin- jected locally, a regionally selective blockade of the bind- ing of i.v. [3H]SCH 23390 in vivo was observed at 1 h post-injection with a similar diffusion area (approximately 2.0 mm3). Taken together, these data might suggest that intracerebral SCH 23390 produces rapid regionally selec- tive effects within a distance of approximately 2.0 mm 3 in the first hour. Nevertheless, given the rapid decrease in the concentration of [3H]SCH 23390 at the site of injection following intracerebral administration, caution is still war- ranted in interpreting results without adequate neu- roanatomical controls. Such caution must also be taken in drawing conclusions from the present results about the involvement of D-1 receptors in subregions of the accum- bens (i.e., shell vs. core) or specific nuclei of the amygdala (i.e., central) in cocaine self-administration.
Finally, these results suggest a role for dopaminergic mechanisms in the amygdala in cocaine self-administra- tion, but the process by which D-1 receptor blockade in the amygdala might modulate cocaine self-administration is unclear. It has been suggested that, by virtue of common neurochemical and anatomical characteristics, the central nucleus of the amygdala and nucleus accumbens shell adjoin a forebrain continuum termed the extended amyg- dala, and thus may share functional attributes [1,18]. Sup-
port for a role for the extended amygdala in cocaine self-administration comes from the observation that lesions of projection areas from the shell of the accumbens to the sublenticular extended amygdala produce more robust ef- fects on cocaine self-administration than do lesions of projection areas from the core region of the accumbens to the subcommissural ventral pallidum [43].
Other evidence of amygdala involvement in the behav- ioral effects of cocaine is provided by observations that blockade of D-1 receptors in the amygdala attenuates the discriminative stimulus properties of cocaine [11], and that combined 6-hydroxydopamine lesions of the amygdala and accumbens more effectively disrupt cocaine self-adminis- tration than accumbens lesions alone (Koob et al., unpub- lished observations). Nevertheless, removal of dopaminer- gic terminals in the amygdala with 6-hydroxydopamine does not produce robust effects on cocaine self-administra- tion (Koob et al., unpublished observations; [31]) except for high doses of cocaine [31]. Moreover, dopaminergic activation in the nucleus accumbens is modulated by dopaminergic activity in the amygdala [27,31], and this effect may be responsible for the facilitated acquisition of self-administration of low doses of amphetamine observed following 6-hydroxydopamine lesions of the amygdala [13]. Thus it may be that, as has been proposed for the medial prefrontal cortex [16,37], dopaminergic mechanisms in the amygdala are involved in cocaine self-administration by virtue of their influence on dopaminergic tone in the nucleus accumbens.
The precise nuclei within the amygdala that may play a role in cocaine self-administration remain unclear. The central nucleus of the amygdala contains a significant amount of dopamine and is associated with the extended amygdala. The basolateral nucleus provides the major efferent pathway from the amygdala to the accumbens [22], and contains a density of D-1 receptors comparable to that of the central nucleus [6]. In addition, within the amygala information flows from the basolateral nucleus to the central nucleus without a reciprocal innervation [30]. Despite the regional selectivity of effects produced by intracerebral SCH 23390 observed here, identifying the involvement of specific amygdala nuclei in the behavioral effects of self-administered cocaine, as well as the role of accumbens dopaminergic activity in these effects, may require manipulations more regionally selective than those employed in the present study.
Acknowledgements
This work was supported by grants DA05278, DA04398 and DA07352 from the National Institute on Drug Abuse. The authors thank Bob Lintz for assistance with hardware and software, and Claire E. Williams for her valuable contribution to these studies. This is publication 8993-NP from The Scripps Research Institute.

S.B. Caine et al. / Brain Research 692 (1995) 47-56 55
References
[1] Alheid, G.F. and Heimer, L., New perspectives in basal forebrain organization of special relevance for neuropsychiatric disorders: the striatopallidal, amygdaloid, and corticopetal components of the sub- stantia innominata, Neuroscience, 27 (1988) 1-39.
[2] Anderson, P.H., Comparison of the pharmacological characteristics of [3H]raclopride and [3H]SCH 23390 binding to dopamine recep- tors in vivo in mouse brain, Eur. J. Pharmacol., 146 (1988) 113-120.
[3] Anderson, P.H. and Gronvald, F.C., Specific binding of 3H-SCH 23390 to dopamine D1 receptors in vivo, Life Sci., 38 (1986) 1527-1524.
[4] Berendse, H.W., Groenewegen, H.J. and Lohman, A.H.M., Com- partmental distribution of ventral striatal neurons projecting to the mesencephalon in the rat, J. Neurosci., 12 (1992) 2079-2103.
[5] Bergman, J., Kamien, J.B. and Spealman, R.D., Antagonism of cocaine self-administration by selective dopamine D1 and D2 antag- onists, BehaL,. Pharmacol., 1 (1990) 355-363.
[6] Boyson, S.J., McGonigle, P. and Molinoff, P.B., Quantitative auto- radiographic localization of the D1 and D2 subtypes of dopamine receptors in rat brain, J. Neurosci., 6 (1986) 3167-3178.
[7] Caine, S.B., Lintz, R. and Koob, G.F., Intravenous drug self-admin- istration techniques in animals. In A. Sahgal (Ed.), Behavioural Neuroscience: A Practical Approach, Vol. 2, Oxford University, Oxford, 1993, pp. 116-143.
[8] Caine, S.B. and Koob, G.F., Effects of mesolimbic dopamine deple- tion on responding maintained by cocaine or food, J. Exp. Anal, Behac., 61 (1994) 213-221.
[9] Caine, S.B. and Koob, G.F., Effects of dopamine D-1 and D-2 antagonists on cocaine self-administration under different schedules of reinforcement in the rat, J. PharmacoL Exp. Ther., 270 (1994) 209-217.
[10] Caine, S.B. and Koob, G.F., Pretreatment with the dopamine agonist 7-OH-DPAT shifts the cocaine self-administration dose-effect func- tion to the left under different schedules in the rat, Behm,. Pharma- col., 6 (1995) 333-347.
[11] Callahan, P.M.. Bryan, S.K. and Cunningham, K.A., Discriminative stimulus effects of cocaine: antagonism by dopamine D1 receptor blockade in thc amygdala, Pharmacol. Biochem. Behav., 51 (1995) 759-766.
[12] Chronister, R.B., Sikes, R.W., Wood, J. and DeFrance, J.F., The pattern and termination of ventral tegmental afferents into the nu- cleus accumbens: an anterograde HRP analysis, Neurosci. Lett., 16 (1980) 231-235.
[13] Deminiere, J.M., Taghzouti, K., Tassin, J.P., Le Moal, M. and Simon, H., Increased sensitivity to amphetamine and facilitation of amphetamine self-administration after 6-hydroxydopamine lesions of the amygdala, Psychopharmacology, 94 (1988) 232-236.
[14] Fallon, J.H. and Ciofi, P., Distribution of monoamines in within the amygdala. In J.P. Aggleton (Ed.), The Amygdala: Neurobiological Aspects of Emotion, Memory and Mental Dysfunction, Wiley-Liss, New York, 1992, pp. 97-114.
[15] Fallon, J.H., Koziell. D.A. and Moore, R.Y., Catecholamine innerva- tion of the basal forebrain, J. Comp. Neurol., 170 (1978) 529-532.
[16] Goeders, N.E. and Smith, J.E., Intracranial cocaine self-administra- tion into the medial prefrontal cortex increases dopamine turnover in the nucleus accumbens, J. PharmacoL Exp. Ther., 265 (1993) 592-600.
[17] Groenewegen, H.J., Vermeulen-Van der Zee, E., Te Kortschot, A. and Witter, M.P., Organization of the projections from the subicu- lure to the ventral striatum in the rat. A study using anterograde transport of Phaseolus t,ulgaris leucoagglutinin, Neuroscience, 23 (1987) 103-120.
[18] Heimer. L., Zahm, D.S., Churchill, L., Kalivas, P.W. and Wohlt-
mann, C., Specificity in the projection patterns of accumbal core and shell in the rat, Neuroscience, 41 (1991) 89-125.
[19] Herling, S., Downs, D.A. and Woods, J.H., Cocaine, d-ampheta- mine, and pentobarbital effects on responding maintained by food or cocaine in rhesus monkeys, Psychopharmacology, 64 (1979) 261- 269.
[20] Johnston, J.B., Further contributions to the study of evolution of thc forebrain, J. Comp. Neurol., 35 (1923) 337-521.
[21] Kelley, A.E. and Domesick, V.B., The distribution of the projection from the hippocampal formation to the nucleus accumbens in the rat: an anterograde and retrograde horseradish peroxidase study, Neuro- science, 7 (1982) 2321-2335.
[22] Kelley, A.E., Domesick, V.B. and Nauta, W.J.H., The amygdalostri- atal projection in the rat - an anatomical study by anterograde and retrograde tracing methods, Neuroscience, 7 (1982) 615-630.
[23] Kohler, C., Hall, H. and Gawell, L., Regional in vivo binding of the substituted benzamide [3H]-Eticlopride in the rat brain: evidence for selective labelling of dopamine receptors, Eur. J. Pharmacol., 120 (1986) 216-226.
[24] Koob, G.F. and Bloom, F.E., Cellular and molecular mechanisms of drug dependence, Science, 242 (1988) 715-723.
[25] Koob, G.F., Vaccarino, F.J., Amalric, M. and Bloom, F.E., Positive reinforcement properties of drugs: Search for neural substrates. In J. Engel and L. Oreland (Eds.), Brain Reward Systems and Abase, Raven, New York, 1987, pp. 35-52.
[26] Lindvall, O. and Bjorklund, A. Monoamine lnnervation of Cerebral Cortex, Alan Liss, New York, 1984, pp. 9-40.
[27] Louilot, A., Simon, H., Taghzouti, K. and Le Moal, M., Modulation of dopaminergic activity in the nucleus accumbens following facili- tation or blockade of the dopaminergic transmission in the amyg- dala: a study by in vivo differential pulse voltammetry, Brain Res., 346 (1985) 141-145.
[28] Maldonado, R., Robledo, P., Chover, A.J., Caine, S.B. and Koob, G.F., D1 dopamine receptors in the nucleus accumbens modulate cocaine self-administration in the rat, PharmacoL Biochem. Behat'.. 45 (1993) 239-242.
[29] Martin-Iverson, M.T., Szostak, C. and Fibiger, H.C., 6-hydroxy- dopamine lesions of the medial prefrontal cortex fail to influence intravenous self-administration of cocaine, Psychopharmacology, 88 (1986) 310-314.
[30] McDonald, A.J., Cell types and intrinsic connections of the amyg- dala. In J.P. Aggleton (Ed.), The Amygdala: Neurobiological As- pects of Emotion, Memory and Mental Dysfunction, Wiley-Liss, New York, 1992, pp. 67-96.
[31] McGregor, A., Baker, G. and Roberts, D.C.S., Effect of 6-hydroxy- dopamine lesions of the amygdala on intravenous cocaine self-ad- ministration under a progressive ratio schedule of reinforcement, Brain Res., 646 (1994) 273-278.
[32] McGregor, A. and Roberts, D.C.S., Dopaminergic antagonism within the nucleus accumbens or the amygdala produces differential effects on intravenous cocaine self-administration under fixed and progres- sive ratio schedules of reinforcement, Brain Res., 624 (1993) 245- 252.
[33] Paxinos, G. and Watson, C. The Rat Brain in Stereotaxic Coordi- nates, Academic Press, Orlando, FL, 1986, 2nd edn.
[34] Pettit, H.O., Ettenberg, A., Bloom, F.E. and Koob, G.F., Destruction of dopamine in the nucleus accumbens selectively attenuates cocaine but not heroin self-administration in rats, P.Lvchopharmacology, 84 (1984) 167-163.
[35] Pettit, H.O. and Justice, J.B., Dopamine in the nucleus accumbens during cocaine self-administration as studied by in vivo microdialy- sis, Pharmacol. Biochem. Behat~., 34 (1989) 890-904.
[36] Phillips, A.G., Broekkamp, C.L. and Fibiger, H.C., Strategies for studying the neurochemical substrates of drug reinforcement in rodents, Prog. Neuropsychopharmacol. Biol. P.~vchiat~, 7 (1983) 585-590.

56 S.B. Caine et al. / Brain Research 692 (1995) 47-56
[37] Pycock, C.J., Carter, C.J. and Kerwin, R.W., Effect of 6-hydroxy- dopamine lesions of the medial prefrontal cortex on neurotransmitter systems in subcortical sites in the rat, J. Neurochem., 34 (1980) 91-99.
[38] Ritz, M.C., Lamb, R.J., Goldberg, S.R. and Kuhar, M.J., Cocaine receptors on dopamine transporters are related to self-administration of cocaine, Science, 237 (1987) 1218-1223.
[39] Roberts, D.C.S., Corcoran, M.E. and Fibiger, H.C., On the role of ascending catecholaminergic systems in intravenous self-administra- tion of cocaine, Pharmacol. Biochem. Behav., 6 (1977) 615-620.
[40] Roberts, D.C.S., Koob, G.F., Klonoff, P. and Fibiger, H.C., Extinc- tion and recovery of cocaine self-administration following 6-hy- droxydopamine lesions of the nucleus accumbens, Pharmacol. Biochem. Behav., 12 (1980) 781-787.
[41] Roberts, D.C.S. and Koob, G.F., Disruption of cocaine self-adminis- tration following 6-hydroxydopamine lesions of the ventral tegmen- tal area in rats, Pharmacol. Biochem. Behav., 16 (1982) 901-904.
[42] Roberts, D.C.S. and Vickers, G., Atypical neuroleptics increase cocaine self-administration of cocaine: An evaluation of a be- havioural screen for antipsychotic activity, Psychopharmacology, 82 (1984) 135-139.
[43] Robledo, P. and Koob, G.F., Two discrete nucleus accumbens projection areas differentially mediate cocaine self-administration in the rat, Behav. Brain Res., 55 (1993) 159-166.
[44] Schenk, S., Horger, B.A., Peltier, R. and Shelton, K., Supersensitiv- ity to the reinforcing effects of cocaine following 6-hydroxydopa- mine lesions to the medial prefrontal cortex in rats, Brain Res., 523 (1991) 227-235.
[45] Sesack, S.R. and Pickel, V.M., In the rat medial nucleus accumbens, hippocampal catecholaminergic terminals converge on spiny neurons and are in apposition to each other, Brain Res., 527 (1990) 83-91.
[46] Spealman, R.D., Antagonism of behavioral effects of cocaine by selective dopamine receptor blockers, Psychopharmacology, 101 (1990) 142-145.
[47] Spealman, R.D., Bergman, J., Madras, B.K. and Melia, K.F., Dis- criminative stimulus effects of cocaine in squirrel monkeys: Involve- ment of dopamine receptor subtypes, J. Pharmacol. Exp. Ther., 258 (1991) 945-953.
[48] Spealman, R.D. and Kelleher, Behavioral effects of self-adminis- tered cocaine: responding maintained alternately by cocaine and electric shock in squirrel monkeys, J. Pharmacol. Exp. Ther., 210 (1979) 206-214.
[49] Tallarida, R.J. and Murray, R.B., Manual of Pharmacologic Calcu- lations with Computer Programs, Springer Verlag, New York, 1987, pp. 35-44.
[50] Vanover, K.E., Kleven, M.S. and Woolverton, W.L., Blockade of the discriminative stimulus effects of cocaine in rhesus monkeys with the D1 dopamine antagonists SCH 39166 and A-66359, Behav. Pharmacol., 2 (1991) 152-159.
[51] Voorn, P., Jorritsma-Byham, B., Van Dijk, C. and Buijs, R.M., The dopaminergic innervation of the ventral striatum in the rat: a light- and electron-microscopical study with antibodies against dopamine, J. Comp. Neurol., 252 (1986) 84-99.
[52] Wilson, M.C. and Schuster, C.R., The effects of chlorpromazine on psychomotor stimulant self-administration in the rhesus monkey, Psychopharmacologia, 26 (1972) 115-126.
[53] Wilson, M.C. and Schuster, C.R., Interactions between atropine, chlorpromazine and cocaine on food reinforced behavior, Pharma- col Biochem. Behav., 3 (1975) 363-375.
[54] Woolverton, W.L., Effects of a D1 and a D2 antagonist on the self-administration of cocaine and piribedil by rhesus monkeys, Pharmacol. Biochem. Behav., 24 (1986) 531-535.




















