
Chromatin Remodeling Is a Key Mechanism Underlying Cocaine-Induced Plasticity in Striatum
12
Neuron, Vol. 48, 303–314, October 20, 2005, Copyright ª2005 by Elsevier Inc. DOI 10.1016/j.neuron.2005.09.023 Chromatin Remodeling Is a Key Mechanism Underlying Cocaine-Induced Plasticity in Striatum Arvind Kumar, 1 Kwang-Ho Choi, 1,3 William Renthal, 1,3 Nadia M. Tsankova, 1 David E.H. Theobald, 1 Hoang-Trang Truong, 1 Scott J. Russo, 1 Quincey LaPlant, 1 Teresa S. Sasaki, 1 Kimberly N. Whistler, 1 Rachael L. Neve, 2 David W. Self, 1 and Eric J. Nestler 1, * 1 Department of Psychiatry and Center for Basic Neuroscience The University of Texas Southwestern Medical Center 5323 Harry Hines Boulevard Dallas, Texas 75390 2 Department of Psychiatry Harvard Medical School and McLean Hospital 115 Mill Street Belmont, Massachusetts 02178 Summary Given that cocaine induces neuroadaptations through regulation of gene expression, we investigated whether chromatin remodeling at specific gene promoters may be a key mechanism. We show that cocaine induces specific histone modifications at different gene pro- moters in striatum, a major neural substrate for co- caine’s behavioral effects. At the cFos promoter, H4 hyperacetylation is seen within 30 min of a single co- caine injection, whereas no histone modifications were seen with chronic cocaine, consistent with co- caine’s ability to induce cFos acutely, but not chroni- cally. In contrast, at the BDNF and Cdk5 promoters, genes that are induced by chronic, but not acute, co- caine, H3 hyperacetylation was observed with chronic cocaine only. DFosB, a cocaine-induced transcription factor, appears to mediate this regulation of the Cdk5 gene. Furthermore, modulating histone deacetylase activity alters locomotor and rewarding responses to cocaine. Thus, chromatin remodeling is an important regulatory mechanism underlying cocaine-induced neural and behavioral plasticity. Introduction Acute and chronic exposure to cocaine alters gene ex- pression in striatum, a major neural substrate for the rewarding and locomotor-activating effects of psychos- timulants and other drugs of abuse (Hyman and Mal- enka, 2001; Nestler, 2001; Yuferov et al., 2003; Kalivas, 2004; Yao et al., 2004). The Fos family of transcription factors has been implicated as important mediators of some of these drug-induced changes in gene expres- sion. Drugs of abuse rapidly but transiently induce cFos and several other Fos family proteins in striatum after acute administration (Graybiel et al., 1990; Young et al., 1991). In contrast, chronic drug exposure desensi- tizes the induction of most of these genes (Hope et al., 1992; Daunais et al., 1993; Persico et al., 1993) and leads instead to the selective accumulation of DFosB, a trun- cated splice variant of the FosB gene (Hope et al., 1994; Moratalla et al., 1996; Hiroi et al., 1997). Induction of DFosB appears to mediate enhanced sensitivity and drive for cocaine and other drugs of abuse in animal models of drug addiction (Kelz et al., 1999; Nestler et al., 2001; Colby et al., 2003; Peakman et al., 2003), and progress has been made in identifying some of the target genes through which DFosB produces this behav- ioral plasticity (Ang et al., 2001; Bibb et al., 2001; McClung and Nestler, 2003). However, the molecular mechanisms underlying the in vivo regulation of specific target genes by DFosB and other drug-induced tran- scription factors remain unexplored. The goal of this study was to examine these next levels of transcriptional mechanisms underlying chronic co- caine action. Increasing evidence from other fields sug- gests that transcription factors regulate target genes by recruiting to the gene promoters two different classes of enzymes: (1) enzymes that modify the core histones and other chromatin components by regulating their acetyla- tion, phosphorylation, methylation, etc. (e.g., histone acetyl transferases [HATs] and histone deacetylases [HDACs]), and (2) enzymes that alter the structure of chromatin by disrupting the nucleosome in an ATP- dependent manner (e.g., the SWI-SNF [mating switching and sucrose nonfermenting] complex) (for review, see Jenuwein and Allis, 2001; Turner, 2002; Felsenfeld and Groudine, 2003). These enzymes act on a target pro- moter and thereby increase or decrease expression of that gene. For example, the acetylation of histones H3 and H4, as well as phosphoacetylation of H3, at gene promoters is linked to increased gene activity, while de- acetylation is linked with suppression and silencing of gene activity. Recent evidence indicates that histone acetylation-deacetylation is regulated in adult neurons in association with learning and memory (Guan et al., 2002; Alarcon et al., 2004; Korzus et al., 2004; Levenson et al., 2004) and in response to seizures (Huang et al., 2002; Tsankova et al., 2004) and psychotropic drugs (Li et al., 2004). Moreover, HDAC inhibitors have shown promise as potential therapeutic agents in animal mod- els of Huntington’s disease (Steffan et al., 2001; Hockly et al., 2003; Mattson, 2003) and spinal muscular atrophy (Chang et al., 2001; Minamiyama et al., 2004). These data support the view that the acetylation of core histones may be a critical determinant of gene expression in brain. Here, we focused on several gene promoters known to be regulated in striatum by acute or chronic cocaine, and utilized chromatin immunoprecipitation (ChIP) assays to investigate mechanisms of chromatin remodeling asso- ciated with drug action. We chose gene promoters that represent distinct drug mechanisms: cFos is activated by acute cocaine, but desensitized by chronic cocaine, while BDNF and Cdk5 are activated chronically but not acutely. In addition, induction of the Cdk5 gene appears to be mediated by DFosB, whereas BDNF is regulated independently of DFosB by still unknown mechanisms. *Correspondence: [email protected] 3 These authors contributed equally to this work.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Chromatin Remodeling Is a Key Mechanism Underlying Cocaine-Induced Plasticity in Striatum

Neuron, Vol. 48, 303–314, October 20, 2005, Copyright ª2005 by Elsevier Inc. DOI 10.1016/j.neuron.2005.09.023
Chromatin Remodeling Is a Key MechanismUnderlying Cocaine-Induced Plasticity in Striatum
Arvind Kumar,1 Kwang-Ho Choi,1,3 William Renthal,1,3
Nadia M. Tsankova,1 David E.H. Theobald,1
Hoang-Trang Truong,1 Scott J. Russo,1
Quincey LaPlant,1 Teresa S. Sasaki,1
Kimberly N. Whistler,1 Rachael L. Neve,2
David W. Self,1 and Eric J. Nestler1,*1Department of Psychiatry
and Center for Basic NeuroscienceThe University of Texas Southwestern Medical Center5323 Harry Hines BoulevardDallas, Texas 753902Department of PsychiatryHarvard Medical School
and McLean Hospital115 Mill StreetBelmont, Massachusetts 02178
Summary
Given that cocaine induces neuroadaptations through
regulation of gene expression, we investigated whetherchromatin remodeling at specific gene promoters may
be a key mechanism. We show that cocaine inducesspecific histone modifications at different gene pro-
moters in striatum, a major neural substrate for co-caine’s behavioral effects. At the cFos promoter, H4
hyperacetylation is seen within 30 min of a single co-caine injection, whereas no histone modifications
were seen with chronic cocaine, consistent with co-caine’s ability to induce cFos acutely, but not chroni-
cally. In contrast, at the BDNF and Cdk5 promoters,
genes that are induced by chronic, but not acute, co-caine, H3 hyperacetylation was observed with chronic
cocaine only. DFosB, a cocaine-induced transcriptionfactor, appears to mediate this regulation of the Cdk5
gene. Furthermore, modulating histone deacetylaseactivity alters locomotor and rewarding responses to
cocaine. Thus, chromatin remodeling is an importantregulatory mechanism underlying cocaine-induced
neural and behavioral plasticity.
Introduction
Acute and chronic exposure to cocaine alters gene ex-pression in striatum, a major neural substrate for therewarding and locomotor-activating effects of psychos-timulants and other drugs of abuse (Hyman and Mal-enka, 2001; Nestler, 2001; Yuferov et al., 2003; Kalivas,2004; Yao et al., 2004). The Fos family of transcriptionfactors has been implicated as important mediators ofsome of these drug-induced changes in gene expres-sion. Drugs of abuse rapidly but transiently inducecFos and several other Fos family proteins in striatumafter acute administration (Graybiel et al., 1990; Younget al., 1991). In contrast, chronic drug exposure desensi-tizes the induction of most of these genes (Hope et al.,
*Correspondence: [email protected] authors contributed equally to this work.
1992; Daunais et al., 1993; Persico et al., 1993) and leadsinstead to the selective accumulation of DFosB, a trun-cated splice variant of the FosB gene (Hope et al.,1994; Moratalla et al., 1996; Hiroi et al., 1997). Inductionof DFosB appears to mediate enhanced sensitivity anddrive for cocaine and other drugs of abuse in animalmodels of drug addiction (Kelz et al., 1999; Nestleret al., 2001; Colby et al., 2003; Peakman et al., 2003),and progress has been made in identifying some of thetarget genes through which DFosB produces this behav-ioral plasticity (Ang et al., 2001; Bibb et al., 2001;McClung and Nestler, 2003). However, the molecularmechanisms underlying the in vivo regulation of specifictarget genes by DFosB and other drug-induced tran-scription factors remain unexplored.
The goal of this study was to examine these next levelsof transcriptional mechanisms underlying chronic co-caine action. Increasing evidence from other fields sug-gests that transcription factors regulate target genes byrecruiting to the gene promoters two different classes ofenzymes: (1) enzymes that modify the core histones andother chromatin components by regulating their acetyla-tion, phosphorylation, methylation, etc. (e.g., histoneacetyl transferases [HATs] and histone deacetylases[HDACs]), and (2) enzymes that alter the structure ofchromatin by disrupting the nucleosome in an ATP-dependent manner (e.g., the SWI-SNF [mating switchingand sucrose nonfermenting] complex) (for review, seeJenuwein and Allis, 2001; Turner, 2002; Felsenfeld andGroudine, 2003). These enzymes act on a target pro-moter and thereby increase or decrease expression ofthat gene. For example, the acetylation of histones H3and H4, as well as phosphoacetylation of H3, at genepromoters is linked to increased gene activity, while de-acetylation is linked with suppression and silencing ofgene activity. Recent evidence indicates that histoneacetylation-deacetylation is regulated in adult neuronsin association with learning and memory (Guan et al.,2002; Alarcon et al., 2004; Korzus et al., 2004; Levensonet al., 2004) and in response to seizures (Huang et al.,2002; Tsankova et al., 2004) and psychotropic drugs (Liet al., 2004). Moreover, HDAC inhibitors have shownpromise as potential therapeutic agents in animal mod-els of Huntington’s disease (Steffan et al., 2001; Hocklyet al., 2003; Mattson, 2003) and spinal muscular atrophy(Chang et al., 2001; Minamiyama et al., 2004). These datasupport the view that the acetylation of core histonesmay be a critical determinant of gene expression inbrain.
Here, we focused on several gene promoters known tobe regulated in striatum by acute or chronic cocaine, andutilized chromatin immunoprecipitation (ChIP) assays toinvestigate mechanisms of chromatin remodeling asso-ciated with drug action. We chose gene promoters thatrepresent distinct drug mechanisms: cFos is activatedby acute cocaine, but desensitized by chronic cocaine,while BDNF and Cdk5 are activated chronically but notacutely. In addition, induction of the Cdk5 gene appearsto be mediated by DFosB, whereas BDNF is regulatedindependently of DFosB by still unknown mechanisms.

Neuron304
Figure 1. Use of Chromatin Immunoprecipitation and Real-Time PCR to Measure Histone Modifications in Striatum of Cocaine-Treated and
Control Rats
(A) Levels of acetylated H4 (acH4) at the cFos and several other gene promoters in striatum of rats treated acutely with saline or cocaine and
analyzed 30 min later were quantified using real-time PCR, by comparing relative Ct values for the various genes of interest. Parallel experi-
ments, not shown, were carried out for acetylated and phosphoacetylated H3. Ct values of immunoprecipitated samples (saline control versus
cocaine) were normalized to Ct values obtained from ‘‘input’’ or nonimmunoprecipitated DNA. The figure shows increased levels of the cFos
promoter in anti-acetylated H4 immunoprecipitates after cocaine treatment (Ct values of 28.08 for control, and 26.68 for cocaine, indicating
a w2.5-fold increase with cocaine). Immunoprecipitation with nonimmune IgG yielded much higher Ct values (w37) indicating almost no pre-
cipitation of the cFos promoter in the absence of a specific antibody. In contrast to the cFos promoter, acute cocaine, chronic cocaine, or self-
administered cocaine (Self Adm), had no effect on levels of several other gene promoters in anti-acH4 or anti-acH3 immunoprecipitates, in-
cluding the b-tubulin promoter (A and B), the b-globin promoter (A), the tyrosine hydroxylase (TH) promoter (C), and the H4 promoter (D).
Data are expressed as mean 6 SEM (n = 6–8).
Findings from this study describe prominent alterationsin histone modifications and other mechanisms of chro-matin remodeling in striatal neurons that are associatedwith cocaine regulation of specific genes and directlyimplicate these mechanisms in cocaine-induced neuraland behavioral plasticity.
Results
Quantification of Cocaine-Induced Histone
Modifications by Chromatin ImmunoprecipitationAssays
To better understand the molecular actions of cocaine,we studied histone modifications at the promoter re-gions of the cFos, FosB, BDNF, and Cdk5 genes in ratstriatum after acute or repeated cocaine administration.To clearly differentiate between acute and chronic co-caine effects, we analyzed histone modifications at sev-eral different time points after cocaine administration.Striatal punches were fixed in 1% formaldehyde to
crosslink proteins bound to DNA in vivo. The crosslinkedchromatin was then sheared to fragments of w500 bp inlength via sonication. We then performed ChIP assayswith antibodies against acetylated H4, acetylated H3,or phosphoacetylated H3 and quantified the amount ofDNA associated with the modified histones using real-time PCR.
Several controls were performed to confirm the spec-ificity and validity of the assay (Figure 1). To control forthe specificity of antibody binding, we immunoprecipi-tated chromatin samples with nonimmune IgG, whichprecipitated negligible levels of the various genes stud-ied (Figure 1A). To ensure that our technique would allowus to measure acetylation of histones only at trans-criptionally active regions in the genome where acetyla-tion is present in vivo, we confirmed that acetylation isabsent in striatum at transcriptionally silenced genes,such as the b-globin gene (Figure 1A), which is inacti-vated within neurons early in development, and ongenes (e.g., tyrosine hydroxylase) whose expression is

Chromatin Remodeling and Cocaine Action305
Figure 2. Histone Modifications at the cFos Promoter in Striatum after Acute or Chronic Cocaine Administration
ChIP was performed on striata of cocaine-treated and control rats with anti-acetylated H4 (acH4), anti-acetylated H3 (acH3), or anti-phosphoa-
cetylated H3 (pacH3) antibodies, and levels of the cFos promoter in the immunoprecipitates were measured with real-time PCR. (A)–(C) show
acH4, acH3, and pacH3 at 30, 90, or 180 min after a single injection of cocaine or saline. (D) shows these histone modifications 1 or 24 hr after
a chronic course of cocaine or saline. Data are expressed as mean 6 SEM (n = 6–8). *p < 0.05, Student’s t test.
high in other brain regions but absent from striatum (Fig-ure 1C). Finally, we measured levels of histone acetyla-tion in striatum at the promoters of the b-tubulin (Figure1B) and core histone H4 genes (Figure 1D), which are ex-pressed in adult striatum but have expression levels thatare unchanged by acute and chronic cocaine (McClungand Nestler, 2003; Yuferov et al., 2003). As expected, lev-els of histone modifications at these two promoters didnot differ between control and acute or chronic cocaine-treated animals. These findings indicate that the ob-served changes in histone modifications at the cFos,FosB, BDNF, and Cdk5 genes, presented below, arenot global, but are limited to genes with expressionthat varies as a result of cocaine exposure.
Cocaine Regulation of Histone Modificationsat the cFos and FosB Promoters
We first assayed histone modifications at the cFos pro-moter after acute and chronic cocaine. As shown in Fig-ures 2A–2C, we found that acute cocaine causes therobust induction of both H4 acetylation and H3 phos-phoacetylation, but not of H3 acetylation alone, at thecFos promoter as early as 30 min after drug administra-tion. All of these effects reverted to control levels within3 hr and remained unchanged 24 hr after drug adminis-tration. Acute administration of amphetamine, a relatedpsychostimulant, caused a similar pattern of histone
modifications at the cFos promoter (data not shown).These findings of rapid and dramatic histone modifica-tions at the cFos gene in striatum in response to acutepsychostimulants are consistent with the fact thatcFos is an immediate early gene, which is known to beactivated very rapidly after acute administration of thesedrugs of abuse (Graybiel et al., 1990; Young et al., 1991;Hope et al., 1992).
Very different results were obtained after a course ofchronic cocaine administration. Under these conditions,levels of H4 and H3 acetylation (Figure 2D) and of H3phosphoacetylation (data not shown) were unchangedcompared to saline controls, both 1 hr and 24 hr afterthe last cocaine dose. The lack of induction of histonemodifications at the cFos promoter at the 1 hr time pointrepresents the complete desensitization of histoneacetylation after chronic exposure to cocaine and isconsistent with earlier observations that induction ofcFos mRNA is similarly desensitized after chronic psy-chostimulant exposure (Hope et al., 1992; Daunais et al.,1993; Persico et al., 1993).
We next examined cocaine regulation of histone mod-ifications at the FosB gene promoter. Like cFos, full-length FosB mRNA is induced rapidly and transientlyby acute cocaine, but unlike cFos shows only partial—not complete—desensitization after chronic cocaine ad-ministration (Hope et al., 1992, 1994; Chen et al., 1995).

Neuron306
Figure 3. Histone Modifications at the FosB Promoter in Striatum after Acute or Chronic Cocaine Administration
ChIP was performed on striata of cocaine-treated and control rats with anti-acetylated H4 (acH4), anti-acetylated H3 (acH3), or anti-phosphoa-
cetylated H3 (pacH3) antibodies, and levels of the FosB promoter in the immunoprecipitates were measured with real-time PCR. (A) shows
histone modifications 1 hr after acute cocaine or saline; (B) shows histone modifications 1 hr after the last injection of chronic cocaine or saline;
and (C) shows histone modifications 24 hr after the last cocaine or saline self administration (Self Adm). Results are not shown for pacH3, since
Ct values were virtually as high as those seen for nonimmune IgG and were unaffected by cocaine exposure. Data are expressed as mean 6
SEM (n = 6–8). *p < 0.05, Student’s t test.
The FosB gene was also of interest, because it encodesDFosB, whose mRNA is induced rapidly and transientlyby acute cocaine and shows only partial desensitizationwith chronic cocaine, but DFosB protein accumulatesuniquely during a course of chronic cocaine administra-tion due to the stability of the protein (Chen et al., 1995,1997; Nestler et al., 2001). Analysis of histone modifica-tions at the FosB promoter showed similarities to thoseobserved at the cFos promoter after acute cocaine (Fig-ure 3A), but the modifications were strikingly differentafter chronic cocaine (Figure 3B). Thus, we observed in-duction of H4 acetylation, but no change in H3 acetyla-tion or phosphoacetylation (data not shown), at theFosB promoter after acute cocaine administration. Incontrast, the FosB promoter showed significant induc-tion of H3 acetylation, but not H3 phosphoacetylationor H4 acetylation, at 1 hr after the last dose of a chroniccocaine regimen (Figure 3B). These data are consistentwith earlier observations that induction of FosB andDFosB mRNA, in contrast to cFos mRNA, only partiallydesensitizes with chronic cocaine administration (Chenet al., 1995).
To determine whether these effects of chronic investi-gator-administered cocaine on histone modifications at
the cFos and FosB promoters are relevant to cocaine ad-diction, we determined whether similar modificationsoccur after chronic self-administration of the drug. In-deed, chronic self-administration of cocaine in rats wasassociated with no histone modifications at the cFospromoter (data not shown), but with dramatic inductionof H3 acetylation, but not H4 acetylation, at the FosB pro-moter (Figure 3C).
Cocaine Regulation of Histone Modifications
at the BDNF and Cdk5 PromotersUnlike cFos, which is induced by acute, but not chronic,cocaine, BDNF and Cdk5 are examples of genes thatare induced by chronic, but not acute, cocaine adminis-tration (Bibb et al., 2001; Grimm et al., 2003; McClungand Nestler, 2003). Whereas a single dose of cocainedid not produce any significant histone modificationsat the BDNF or Cdk5 promoters (data not shown), wefound dramatic regulation at these promoters afterchronic cocaine (Figure 4). Chronic cocaine robustly in-duced levels of H3 acetylation at the BDNF and Cdk5promoters in striatum, with no changes seen in H4 acet-ylation. Strikingly, this induction of H3 acetylation wasobserved 24 hr after the last dose of cocaine (Figures
Figure 4. Histone Modifications at the BDNF
and Cdk5 Promoters in Striatum after
Chronic Cocaine Administration
ChIP was performed on striata of chronic co-
caine- and saline-treated rats (used 24 hr af-
ter the last injection) with anti-acetylated H4
(acH4), anti-acetylated H3 (acH3), or anti-
phosphoacetylated H3 (pacH3) antibodies,
and levels of the BDNF (A) or Cdk5 (B) pro-
moters in the immunoprecipitates were mea-
sured using real-time PCR. Results are not
shown for pacH3, since Ct values were virtu-
ally as high as those seen for nonimmune IgG
and were unaffected by cocaine exposure.
There was also no effect of acute cocaine on
association of the BDNF or Cdk5 promoters
with these various histone modifications
(data not shown). Data are expressed as
mean 6 SEM (n = 6–8). *p < 0.05, Student’s
t test.

Chromatin Remodeling and Cocaine Action307
Figure 5. Role of DFosB in Regulation of the Cdk5 Gene Promoter in Striatum
(A) ChIP was performed on striata of chronic cocaine-and saline-treated rats (used 24 hr after the last injection) with anti-DFosB, anti-full length
FosB, or anti-Brg1 antibodies, and levels of the Cdk5 promoter in the immunoprecipitates were measured using real-time PCR. (B) ChIP was
performed on striata of inducible bitransgenic mice either expressing (+DFosB) or not expressing (2DFosB) DFosB with anti-DFosB, anti-full
length FosB, or anti-Brg1 antibody, and levels of the Cdk5 promoter in the immunoprecipitates were measured with real-time PCR. Data are
expressed as mean 6 SEM; n = 6–8. *p < 0.05, Student’s t test.
4A and 4B), suggesting relatively long-lived adaptationsat these promoters in contrast to what was seen withacute cocaine at the cFos promoter. An even more dra-matic selective increase in H3 acetylation levels wasseen at the BDNF and Cdk5 promoters 24 hr after thelast session of chronic self-administration of cocaine(Figure 4).
To examine the longevity of increased H3 acetylationat the BDNF and Cdk5 gene promoters, we analyzedstriatal samples after 1 week of withdrawal. Interest-ingly, the level of acetylated H3 at the BDNF promoterwas strikingly high at this time point (14.1 6 3.6-fold;p < 0.05) compared to the w3-fold increase seen after1 day of withdrawal. Thus, histone modifications at theBDNF promoter are very long-lasting and even buildover the course of withdrawal. This pattern of H3 acety-lation at the BDNF gene is interesting in light of findingsthat levels of BDNF protein in this region increase furtheras a function of withdrawal (Grimm et al., 2003). In con-trast, the level of acetylated H3 at the Cdk5 promoterremained elevated after 1 week of withdrawal (1.9 60.5-fold), but this effect was no longer statistically signif-icant. This suggests that histone modifications at theCdk5 gene are beginning to return to normal at thistime point.
Role for DFosB in Cocaine Regulation
of the Cdk5 GenePrevious work has suggested that Cdk5 is one targetgene through which DFosB produces its long-lasting ef-fects on hippocampal and striatal function (Bibb et al.,2001; Chen et al., 2000; Peakman et al., 2003). This isbased partly on the observations that DFosB overex-pression induces Cdk5 in striatum, while a dominantnegative antagonist blocks cocaine induction of Cdk5.Cdk5 is a particularly interesting target gene, since re-cent evidence has implicated it in mediating the abilityof chronic cocaine to induce the outgrowth of dendriticarborizations of striatal neurons (Norrholm et al., 2003).However, direct evidence that DFosB’s regulation ofCdk5 in striatum represents a direct effect on theCdk5 gene promoter has been lacking. Indeed, there re-mains some controversy in the field as to whether
DFosB regulates gene expression directly or, rather, in-directly by interfering with the actions of other transcrip-tion factors (see Andersson et al., 2001; Nestler et al.,2001). It was, therefore, important to use ChIP assaysto gain further insight into mechanisms governing regu-lation of the Cdk5 gene in vivo. First, we carried out ChIPassays using an anti-DFosB antibody to immunoprecip-itate chromatin fragments from striata of control andchronic cocaine-treated rats. As predicted, we founda w3-fold increase in the association of DFosB withthe Cdk5 gene promoter after chronic cocaine exposure(Figure 5A). In contrast, there was no evidence ofincreased association of DFosB with the BDNF gene(data not shown), consistent with reports that BDNF ex-pression is not affected by DFosB in striatum (McClungand Nestler, 2003). We know that DFosB, and not full-length FosB, is the protein associating with the Cdk5gene promoter, since no effect was seen using an anti-body that recognizes full-length FosB but not DFosB.These findings demonstrate that, after chronic cocaineadministration, the Cdk5 gene in striatum shows signsof transcriptional activation (i.e., increased H3 acetyla-tion) as well as increased association with DFosB.
Second, we analyzed striatum from a line of bitrans-genic mouse that inducibly overexpresses DFosB withsome selectivity in striatum (Kelz et al., 1999). This mouseuses the tetracycline gene regulation system to maintainDFosB so that it is turned off during development and toturn it on in striatum of adult animals. We found that in-duction of DFosB in these mice, which induces Cdk5mRNA and protein expression (Bibb et al., 2001), wasassociated with a w3-fold increase in association ofDFosB with the Cdk5 promoter in striatum (Figure 5B).In contrast, no effect was seen at the BDNF promoter(data not shown). This further supports the notion thatDFosB interacts directly with the Cdk5 gene in striatumin vivo.
Having established evidence that the Cdk5 gene isactivated in striatum in vivo by chronic cocaine andthat this effect may be mediated, at least in part, viaDFosB, we examined whether another class of chroma-tin remodeling protein, SWI-SNF, is also involved. Brg1is the core ATPase of this multisubunit chromatin
Neuron308
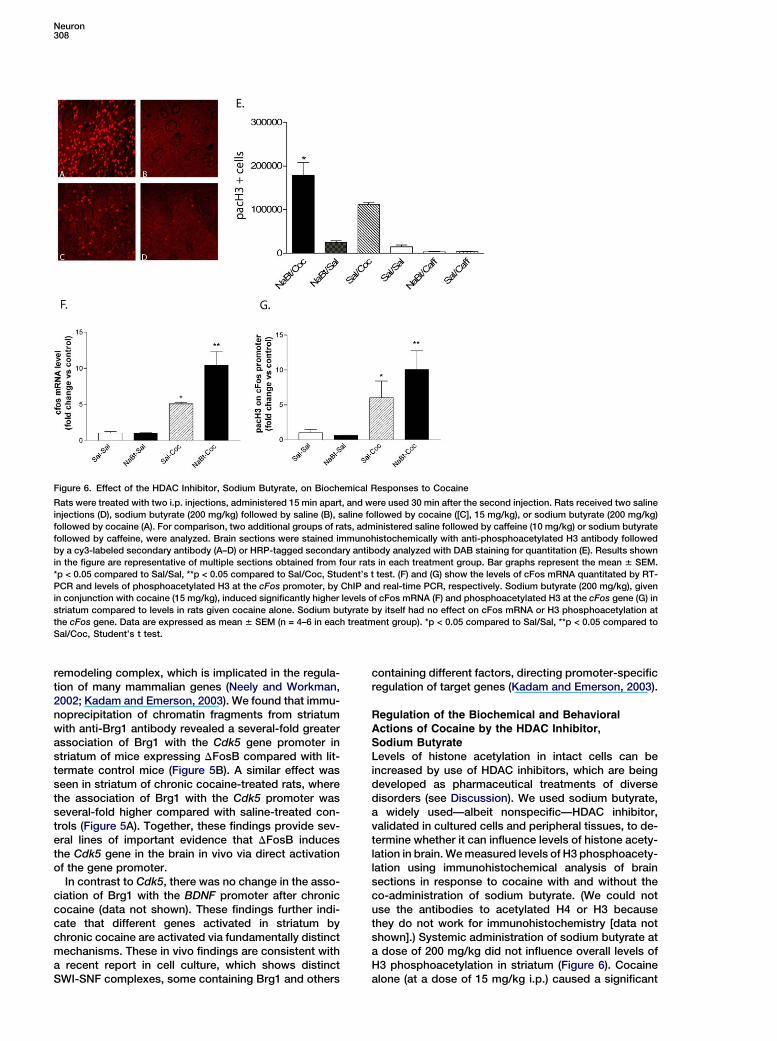
Figure 6. Effect of the HDAC Inhibitor, Sodium Butyrate, on Biochemical Responses to Cocaine
Rats were treated with two i.p. injections, administered 15 min apart, and were used 30 min after the second injection. Rats received two saline
injections (D), sodium butyrate (200 mg/kg) followed by saline (B), saline followed by cocaine ([C], 15 mg/kg), or sodium butyrate (200 mg/kg)
followed by cocaine (A). For comparison, two additional groups of rats, administered saline followed by caffeine (10 mg/kg) or sodium butyrate
followed by caffeine, were analyzed. Brain sections were stained immunohistochemically with anti-phosphoacetylated H3 antibody followed
by a cy3-labeled secondary antibody (A–D) or HRP-tagged secondary antibody analyzed with DAB staining for quantitation (E). Results shown
in the figure are representative of multiple sections obtained from four rats in each treatment group. Bar graphs represent the mean 6 SEM.
*p < 0.05 compared to Sal/Sal, **p < 0.05 compared to Sal/Coc, Student’s t test. (F) and (G) show the levels of cFos mRNA quantitated by RT-
PCR and levels of phosphoacetylated H3 at the cFos promoter, by ChIP and real-time PCR, respectively. Sodium butyrate (200 mg/kg), given
in conjunction with cocaine (15 mg/kg), induced significantly higher levels of cFos mRNA (F) and phosphoacetylated H3 at the cFos gene (G) in
striatum compared to levels in rats given cocaine alone. Sodium butyrate by itself had no effect on cFos mRNA or H3 phosphoacetylation at
the cFos gene. Data are expressed as mean 6 SEM (n = 4–6 in each treatment group). *p < 0.05 compared to Sal/Sal, **p < 0.05 compared to
Sal/Coc, Student’s t test.
remodeling complex, which is implicated in the regula-tion of many mammalian genes (Neely and Workman,2002; Kadam and Emerson, 2003). We found that immu-noprecipitation of chromatin fragments from striatumwith anti-Brg1 antibody revealed a several-fold greaterassociation of Brg1 with the Cdk5 gene promoter instriatum of mice expressing DFosB compared with lit-termate control mice (Figure 5B). A similar effect wasseen in striatum of chronic cocaine-treated rats, wherethe association of Brg1 with the Cdk5 promoter wasseveral-fold higher compared with saline-treated con-trols (Figure 5A). Together, these findings provide sev-eral lines of important evidence that DFosB inducesthe Cdk5 gene in the brain in vivo via direct activationof the gene promoter.
In contrast to Cdk5, there was no change in the asso-ciation of Brg1 with the BDNF promoter after chroniccocaine (data not shown). These findings further indi-cate that different genes activated in striatum bychronic cocaine are activated via fundamentally distinctmechanisms. These in vivo findings are consistent witha recent report in cell culture, which shows distinctSWI-SNF complexes, some containing Brg1 and others
containing different factors, directing promoter-specificregulation of target genes (Kadam and Emerson, 2003).
Regulation of the Biochemical and BehavioralActions of Cocaine by the HDAC Inhibitor,
Sodium ButyrateLevels of histone acetylation in intact cells can beincreased by use of HDAC inhibitors, which are beingdeveloped as pharmaceutical treatments of diversedisorders (see Discussion). We used sodium butyrate,a widely used—albeit nonspecific—HDAC inhibitor,validated in cultured cells and peripheral tissues, to de-termine whether it can influence levels of histone acety-lation in brain. We measured levels of H3 phosphoacety-lation using immunohistochemical analysis of brainsections in response to cocaine with and without theco-administration of sodium butyrate. (We could notuse the antibodies to acetylated H4 or H3 becausethey do not work for immunohistochemistry [data notshown].) Systemic administration of sodium butyrate ata dose of 200 mg/kg did not influence overall levels ofH3 phosphoacetylation in striatum (Figure 6). Cocainealone (at a dose of 15 mg/kg i.p.) caused a significant

Chromatin Remodeling and Cocaine Action309
Figure 7. Effect of the HDAC Inhibitor, Sodium Butyrate, on Cocaine-Induced Locomotor Activity
Rats were habituated to the locomotor chambers for 30 min, were given sodium butyrate (NaBt, 100 mg/kg i.p.), and 20 min later were given
cocaine (15 mg/kg i.p.). This regimen was repeated the following day. Line graphs represent the number of ambulatory counts recorded in 5
min bins (mean 6 SEM). Bar graphs represent total activity. *p < 0.05 compared to saline/saline, #p < 0.05 compared to saline/cocaine. In
saline-cocaine and butyrate-cocaine groups, n = 16; n = 8 in saline-saline and butyrate-saline groups. Error bars indicate the mean 6 SEM.
increase in H3 phosphoacetylation. Strikingly, adminis-tration of sodium butyrate followed by cocaine causeda more dramatic induction of H3 phosphoacetylation instriatum (Figure 6). This synergistic interaction betweensodium butyrate and cocaine was apparent not only bythe increased number of cells positive for H3 phosphoa-cetylation (Figure 6), but also by the increased intensityof staining within each positive cell. These findings dem-onstrate that sodium butyrate augments the ability ofcocaine to induce histone modifications in striatum.The effects seen in striatum were not apparent through-out the brain; for example, no obvious effect was evidentin frontal cortex, septum, or other neighboring brain re-gions (data not shown). The lack of regulation seen inthese other regions presumably represents a limit of de-tection, since cocaine is known to alter gene expressionin some of these other brain regions (see Kalivas, 2004),albeit to a lesser extent than in striatum, due to the lowerlevels of the dopamine transporter (target of cocaine) inthese various regions.
The cocaine-induced increase in H3 phosphoacetyla-tion in striatum was selective for this psychostimulant.Caffeine, at a dose (10 mg/kg) that causes the samelevel of locomotor activation as cocaine (Self et al.,1996), failed to activate H3 phosphoacetylation in stria-tum, even when it was injected in conjunction with so-dium butyrate (Figure 6E). This finding also indicatesthat increased phosphoacetylation of H3 in striatum isnot a consequence of increased locomotor activityper se.
Having established that sodium butyrate promotescocaine-induced histone modifications in striatum, itwas of interest to study the effect of this HDAC inhibitoron cocaine regulation of the cFos gene. We found thatadministration of sodium butyrate 15 min prior to co-caine administration increased the degree of inductionof cFos mRNA expression in striatum (Figure 6F). So-dium butyrate alone had no effect. Similarly, sodium bu-tyrate potentiated the ability of cocaine to induce asso-ciation of phosphoacetylated H3 with the cFos genepromoter (Figure 6G).
We next investigated whether the enhancement of co-caine’s effects on histone modifications and gene regu-lation in striatum by sodium butyrate is associated withalterations in cocaine’s behavioral effects. To addressthis question, animals were given sodium butyrate atthe lower dose of 100 mg/kg followed by cocaine15 min later. This dose of sodium butyrate by itselfhad no effect on the animals’ locomotor activity, butenhanced the locomotor-activating effects of cocaine(Figure 7). This effect was more apparent on the secondday of cocaine exposure, when sodium butyrate nearlydoubled the locomotor activation induced by cocaine(Figures 7C and 7D).
Regulation of Cocaine Reward by HDAC
Inhibition or OverexpressionWe used the conditioned place preference (CPP) para-digm, in which animals learn to prefer an environmentpaired with cocaine, to study the influence of histone

Neuron310
acetylation on cocaine reward. We first used the HDACinhibitor, trichostatin A (TSA), which is structurally dis-tinct from butyrate and has been widely validated asan HDAC inhibitor. As with butyrate, we found that sys-temic administration of TSA enhances H3 phosphoace-tylation seen in striatum in response to cocaine (datanot shown). As shown in Figure 8B, TSA administrationalso increased the rewarding effects of cocaine, as indi-cated by enhanced place conditioning to a moderatecocaine dose. These findings further substantiate ourobservations that HDAC inhibition promotes the behav-ioral actions of cocaine.
To gain further insight into these phenomena, weused a Herpes Simplex Virus (HSV) vector to overex-press a particular subtype of HDAC, HDAC4, directlyin striatum. GFP was expressed as a control. We tar-geted the ventral portion of striatum, the nucleus ac-cumbens, which is most closely associated with the re-warding effects of cocaine. We chose HDAC4 because itis highly expressed in brain and because we found inpreliminary experiments that it is highly expressed with-in striatum. We found that overexpression of HDAC4 instriatum dramatically decreased the rewarding effectsof cocaine in the place conditioning assay in compari-son with control animals that received injections ofHSV-GFP (Figure 8A). We also showed that HDAC4overexpression reduced levels of H3 acetylation in thetissue, as would be expected (data not shown). Aswith several other HSV vectors (e.g., see Carlezon et al.,1998; Barrot et al., 2002), we found high levels ofHDAC4 overexpression in a w1 mm3 area around the in-jection site. HDAC4 overexpression was not associatedwith any detectable toxicity, assessed by Nissl stainingand GFAP immunohistochemistry, beyond that seenwith injections of vehicle alone (data not shown).
Figure 8. Effect of HDAC Overexpression or Inhibition on Reward-
ing Responses to Cocaine
Mice were injected with HSV-HDAC4 or HSV-GFP into the nucleus
accumbens and then assessed for place conditioning to cocaine
(5 mg/kg i.p.). In parallel experiments, mice were treated with TSA
(0.2 mg/kg i.p.) or DMSO vehicle prior to each cocaine dose during
place conditioning to cocaine (5 mg/kg). CPP score represents the
time (in seconds) that animals spend in the cocaine-paired chamber
posttraining versus pretraining. Overexpression of HDAC4 (n = 8)
dramatically blocked cocaine place conditioning compared to find-
ings in GFP-expressing control animals (n = 8). Conversely, TSA in-
creased cocaine place conditioning compared to vehicle-treated
controls (n = 12 in each group). Bar graphs represent the mean 6
SEM. *p < 0.05, Student’s t test. Note that the same dose of cocaine
used in both experiments caused different levels of place preference
in control animals; this is due to the known sensitizing effect of intra-
cranial surgery on this measure (see Carlezon et al., 1998).
Discussion
Cocaine Induces Acetylationand Phosphoacetylation of Histones at Specific
Gene PromotersIn this study we demonstrate that acute and repeatedcocaine administration induces prominent alterationsin the N-terminal tails of H3 and H4 associated withspecific gene promoters. The clinical relevance of thechronic cocaine effects on histone modifications issupported by our observations that the chronic self-administration of cocaine results in equivalent regula-tion. Our data reveal an interesting phenomenon: theacute effects of cocaine are associated predominantlywith acetylation of H4 (cFos and FosB), whereas chroniceffects of cocaine are restricted to acetylation of H3(FosB, Cdk5, and BDNF). In addition, phosphoacetyla-tion of H3 was seen for the acute induction of thecFos promoter only. This switch from H4 acetylationacutely to H3 acetylation chronically holds even for indi-vidual genes that are induced under both conditions: forexample, acute induction of FosB involves H4 acetyla-tion, whereas chronic induction of the gene involvesH3 acetylation. These patterns of histone modificationsare generally consistent with those observed recently inhippocampus in response to acute and chronic seiz-ures: acute regulation is associated with H4 acetylationand, for cFos, with phosphoacetylation of H3 as well,while chronic regulation is associated with H3 acetyla-tion (Crosio et al., 2003; Tsankova et al., 2004). To-gether, these findings begin to provide a generalscheme of how histones are covalently modified in brainin response to acute and chronic perturbation.
The phosphoacetylation of histone H3 (i.e., phosphor-ylation at Ser10 and acetylation at Lys14) was seen onlyfor the cFos promoter. As stated above, this was thecase for cocaine in striatum, as well as for seizuresand for activation of several signaling pathways in hip-pocampus (Crosio et al., 2003; Tsankova et al., 2004).For all other promoters studied here and previously,the Ct values from samples pulled down by anti-phosphoacetylated H3 were very high, close to the Ctvalues obtained with nonimmune IgG. This is true evenfor other genes (e.g., FosB and BDNF), which, like cFos,are considered immediate early genes on the basis oftheir rapid induction. However, these other genes, whileinduced rapidly, are not induced to nearly the same de-gree or with the same rapidity as cFos. This suggeststhat phosphoacetylation of H3 may be relatively specificfor the cFos gene and related to the unique rapidity andmagnitude of its induction. Earlier work has shown de-sensitization of cFos after chronic administration of co-caine or other drugs of abuse (Hope et al., 1992; Dau-nais et al., 1993; Persico et al., 1993). We found thatthis desensitization corresponds to the loss of H4 acet-ylation and H3 phosphoacetylation seen at the cFospromoter after the first cocaine exposure. In contrast,at another Fos family promoter, FosB, cocaine contin-ues to induce histone modifications (H3 acetylation)even after prior chronic exposure to the drug, indicatingthat the FosB gene still shows evidence of gene activa-tion. This is consistent with our proposed model forDFosB: that each drug exposure continues to inducethe FosB gene, which allows its stable gene product,

Chromatin Remodeling and Cocaine Action311
DFosB, to accumulate in striatal neurons, where it be-comes a key mediator of the drug-addicted state (Nes-tler et al., 2001). The molecular switch from H4 acetyla-tion at the FosB promoter acutely to H3 acetylationchronically is a major finding of this study. The molecularbasis of this switch is not known. We found no evidencethat acute or chronic cocaine administration alterslevels of specific HDACs in striatum (data not shown),which suggests that the underlying mechanisms maybe quite complex.
Chromatin Remodeling at the BDNF and Cdk5
Promoters: Two Target Genes for Chronic CocaineThe BDNF and Cdk5 genes show evidence of activationafter chronic cocaine administration, with no regulationseen after acute cocaine. These observations are con-sistent with the induction of these genes in striatum af-ter chronic, but not acute, cocaine exposure (Bibb et al.,2001; Grimm et al., 2003; McClung and Nestler, 2003).Moreover, in contrast to FosB, where the increased lev-els of H3 acetylation that occur with chronic cocaine in-jections revert to control within a few hours, this samehistone modification at the BDNF and Cdk5 promoterspersists for at least 24 hr after the last cocaine injection.This histone modification increases significantly furtherat the BDNF gene after 1 week of withdrawal from co-caine, whereas it begins to recover toward normal levelsat the Cdk5 gene. To our knowledge, these are the mostlong-lived examples of drug-induced chromatin remod-eling in brain published to date. The data show thatchanges induced in chromatin structure persist long af-ter cocaine is removed from the system and raise thecritical question of the underlying mechanisms in-volved. We provide evidence here that, at least for theCdk5 gene, DFosB is one critical factor that may drivesome of these changes in chromatin remodeling. Suchlong-lived changes in chromatin remodeling mightbe one of the crucial mechanisms for cocaine-inducedneuroadaptations in striatum, which mediate the neu-ral and behavioral plasticity that underlies cocaineaddiction.
Direct Evidence that Cdk5 Is a Target Gene
for DFosB in Striatum In Vivo
Using ChIP, we provide direct evidence that, aftera course of chronic, but not acute, cocaine administra-tion, there is the selective association of the Cdk5gene promoter with DFosB. We show further that the in-ducible overexpression of DFosB in striatum of adult bi-transgenic mice, which is sufficient for induction of Cdk5expression in vivo (Bibb et al., 2001), causes increasedassociation of DFosB with the Cdk5 gene. Moreover, as-sociation of DFosB with the Cdk5 gene is associatedwith increased binding of Brg1-containing SWI-SNFchromatin remodeling proteins to the Cdk5 promoter.Together, these data support a scheme whereby thegradual accumulation of DFosB, in response to chroniccocaine administration, recruits chromatin remodelingfactors to its target genes like Cdk5. Such factors wouldinclude the HATs that lead to H3 acetylation as well asBrg1 and other components of the SWI-SNF complex.These factors then act to reorganize histone-DNA inter-actions to facilitate transcription (see Narlikar et al.,2002). In contrast to Cdk5, we found no evidence for
increased association of DFosB or Brg1 with the BDNFgene. This is consistent with the evidence that DFosBdoes not regulate BDNF gene expression in striatum(McClung and Nestler, 2003). Presumably, chronic co-caine leads to the long-term induction of BDNF via a dis-tinct mechanism. One possibility is the transcription fac-tor CREB, which is activated in a sustained fashion bychronic cocaine and, upon its inducible overexpressionin striatum, can induce BDNF (McClung and Nestler,2003).
It is worth emphasizing the technical advances madepossible by ChIP assays. Before the advent of this tech-nology, studies of gene regulation in the brain in vivowere severely limited. For example, it was possible todemonstrate that levels of Cdk5 mRNA expressioncould be increased by chronic cocaine and that this ef-fect could be mimicked upon overexpression of DFosB(Bibb et al., 2001). However, all studies of underlyingmechanisms would have to be performed in vitro, whereit was possible to show that the Cdk5 gene promotercontains an AP1 site that binds DFosB and that DFosBcould activate the Cdk5 promoter in cell culture (Chenet al., 2000). Now, with ChIP, it is possible for the firsttime to study such transcriptional mechanisms in thebrain in vivo and understand, with increasing complex-ity, how chronic cocaine administration leads to thelong-term regulation of its target genes.
HDAC Modulation Affects Biochemical
and Behavioral Responses to CocaineHDAC inhibitors cause increased acetylation of histonesin intact cells and are being explored as treatments forseveral diseases, such as certain cancers (Marks et al.,2001), Huntington’s disease (Steffan et al., 2001; Fer-rante et al., 2003; Hockly et al., 2003), and spinal muscu-lar atrophy (Chang et al., 2001; Minamiyama et al., 2004).We used sodium butyrate, a nonspecific HDAC inhibitor,to explore whether the changes in histone acetylationobserved in this study are relevant for cocaine’s bio-chemical and behavioral effects. We found that adminis-tration of sodium butyrate itself does not increase globallevels of histone acetylation in striatum as measured im-munohistochemically. Furthermore, while cocaine ad-ministration alone increases H3 phosphoacetylation instriatum, coadministration of sodium butyrate and co-caine results in a synergistic effect. A very similar patternwas observed for cocaine induction of both cFos mRNAand increased association of acetylated H3 with thecFos gene promoter: both showed synergistic responsesfollowing the coadministration of sodium butyrate andcocaine. This establishes a very clear correlation be-tween changes in histone acetylation and regulation ofgene expression in striatum. A similar correlation wasalso evident for the behavioral effects of cocaine: ad-ministration of sodium butyrate, at a dose that did notalter locomotor activity, almost doubled the locomotorresponse to cocaine.
The potentiation of cocaine’s behavioral effects byHDAC inhibition was also seen in the conditioned placepreference assay, which provides a measure of cocainereward. Here, we showed that TSA, a more potent andstructurally distinct HDAC inhibitor compared to buty-rate, increased the place conditioning effects of cocaine.Importantly, overexpression of HDAC4, achieved via

Neuron312
viral-mediated gene transfer, had the opposite effect:a dramatic reduction in cocaine place conditioning.Moreover, we overexpressed HSV-HDAC4 within the nu-cleus accumbens selectively. This experiment therebyprovides a critical causal connection between the regu-lation of histone acetylation in this brain region specifi-cally and an animal’s sensitivity to the behavioral effectsof cocaine. The fact that HDAC inhibition increases sen-sitivity to cocaine, and HDAC overexpression producesthe opposite effect, is not an obvious finding, since co-caine causes complex effects on gene expression,with gene induction and repression both observed(e.g., see McClung and Nestler, 2003). Our findings aretherefore interesting, because they suggest that stimu-lation of gene transcription may be the predominantmechanism for cocaine-induced behavioral plasticity.
In summary, we show here that individual histonemodifications are associated with different transcrip-tional regulatory mechanisms after acute or chronic co-caine administration. Previous work has shown that sev-eral histone modifications can take place in concert,either on the same histone tail (Cheung et al., 2000; Clay-ton et al., 2000) or on tails of different histones (Turneret al., 1992; Zeitlin et al., 2001; Sun and Allis, 2002).Thus, specific combinations of histone modificationscould correspond to various states of remodeled chro-matin and to the activation or repression of distinctsets of genes. This has been termed the ‘‘histonecode’’ (Jenuwein and Allis, 2001; Turner, 2002). Resultsof the present study raise the interesting possibilitythat acute versus chronic cocaine administration causesdistinct patterns of histone modifications, which, to-gether with ATP-dependent chromatin remodeling com-plexes, induce unique patterns of chromatin remodelingat the promoters of certain genes to regulate their tran-scription. Such regulation provides a new layer of com-plexity, at the molecular level, through which cocaineproduces neural and behavioral plasticity, and revealsmechanisms for the treatment of cocaine addictionthat involve interfering with this plasticity.
Experimental Procedures
Animals
All animal procedures were in accordance with the NIH Guide for the
Care and Use of Laboratory Animals and were approved by our IA-
CUC. Male Sprague-Dawley rats (initial weight, 250 g; Charles River,
Kingston, RI), Bl6/C57 mice (10–12 weeks old; JAX, Bar Harbor, ME),
and DFosB mutant mice off and on doxycycline (10–14 weeks old)—
under conditions that maximally induce DFosB (Kelz et al., 1999)—
were used in these studies. The animal colonies were kept on a 12 hr
light/dark cycle. Mice were housed in groups (four per cage), while
rats were housed in pairs; food and water were made available ad
libitum.
Cocaine Treatments
Acute: rats were given a single i.p. injection of 20 mg/kg cocaine or
saline in equal volumes and killed 30 min to 24 hr later. Chronic:
rats were given an i.p. injection of 20 mg/kg cocaine or saline
once daily for 7 days and were killed 1 hr, 24 hr, or 7 days later. Self-
Administration: rats were trained for cocaine self-administration ac-
cording to published procedures for 15 days (Sutton et al., 2003).
Control rats could press the same levers, but they administered
only saline. The average intake of cocaine was 40 mg/kg/day. Ani-
mals were killed 24 hr after the last self-administration session.
Brains were removed rapidly from decapitated animals and pro-
cessed immediately for ChIP.
Chromatin Immunoprecipitation Assays
ChIP assays were performed on the basis of a protocol from Up-
state Biotechnology ChIP kit (Upstate Biotechnology, Lake Placid,
NY) and modified using published methods (Johnson and Bresnick,
2002; Wells and Farnham, 2002; Tsankova et al., 2004). The follow-
ing antibodies were used: anti-acetylated H3 raised against acety-
lated K9 and 14 H3; anti-acetylated H4 raised against acetylated
K5, 8, 12, and 16 H4; phosphoacetylated H3 raised against phos-
phorylated S10 and acetylated K14 H3; anti-Brg1; and nonimmune
rabbit and mouse IgG (all from Upstate Biotechnology). For DFosB
ChIP pulldown, first, the chromatin was incubated overnight with
mouse anti-FosB (C-terminal antibody; Center for Biomedical In-
ventions, University of Texas Southwestern Medical School), which
recognizes full-length FosB only, and then with by rabbit anti-FosB
(SC048; Santa Cruz Biotechnology, Santa Cruz, CA), which recog-
nizes the middle region of both full-length FosB and DFosB. How-
ever, since virtually all of the full-length FosB is pulled down by
the C-terminal antibody, the second immunoprecipitation brings
down DFosB only.
Quantification of Chromatin Immunoprecipitation
by Real-Time PCR
Levels of specific histone modifications, Brg1, or DFosB at each
gene promoter of interest were determined by measuring the
amount of that gene in chromatin immunoprecipitates by use of
real-time PCR (ABI Prism 7700; Applied Biosystems, CA) (see Table
S1 for primers used, in the Supplemental Data available with this ar-
ticle online). Input or total DNA (nonimmunoprecipitated) and immu-
noprecipitated DNA were PCR amplified in triplicate in the presence
of SYBR Green (Applied Biosystems, CA). Ct values from each sam-
ple were obtained using the Sequence Detector 1.1 software. Rela-
tive quantification of amplified template was performed as de-
scribed earlier by (Chakrabarti et al., 2002) and by the ABI manual
and with some modifications (see Tsankova et al., 2004 for details).
Each real-time PCR reaction, run in triplicate for each brain sample,
was repeated at least twice independently.
RNA Isolation and RT-PCR Detection of cFos mRNA
Rats were treated with saline-saline, saline-cocaine, butyrate-
saline, and butyrate-cocaine using 15 mg/kg cocaine and 200 mg/kg
sodium butyrate, both administered i.p. Striatal dissections were
obtained from decapitated rats, and one punch from each rat
from each of the four groups was used for isolating total RNA (re-
maining punches were used for ChIP assays). RNA was processed,
reverse-transcribed to cDNA using a first-strand synthesis kit (Invi-
trogen, Carlsbad, CA), and quantified using real-time PCR as de-
scribed (Tsankova et al., 2004). PCR reactions were repeated at
least two times independently.
Immunohistochemistry
Rats, randomized into four groups as outlined above (saline-saline,
sodium butyrate-saline, saline-cocaine, and sodium butyrate-
cocaine) were perfused with 4% paraformaldehyde 30 min after the
last injection, fixed brains were sectioned, and coronal sections
through striatum were stained immunohistochemically with an
anti-phosphoacetylated H3 polyclonal rabbit antibody (Upstate Bio-
technology) and a fluorescent Cy3 secondary antibody. Brain sec-
tions were also stained immunohistochemically using HRP-tagged
secondary antibody and diaminobenzidine (DAB), which yielded
equivalent results. DAB-stained sections were used to quantify
the number of phosphoacetylated H3-positive nuclei. Several sec-
tions from each animal were used for quantification. Only the anti-
phosphoacetylated H3 antibody was used, since the anti-acetylated
H3 or H4 antibodies did not work satisfactorily in immunohisto-
chemistry.
Locomotor Activity Assay
Spontaneous locomotor activity was monitored in a circular corridor
as described (Rahman et al., 2003). Animals were randomized into
four groups (saline/saline, sodium butyrate/saline, saline/cocaine,
and sodium butyrate/cocaine) and habituated to the locomotor ac-
tivity apparatus for 60 min. The next day, animals were habituated to
the apparatus for 30 min; then they received their first injection (ve-
hicle or sodium butyrate 100 mg/kg i.p.), and remained in the activity

Chromatin Remodeling and Cocaine Action313
apparatus for 20 min. They were then given the second injection (ve-
hicle or cocaine 15 mg/kg) and placed in the apparatus for an addi-
tional 60 min. The process was repeated for a second day. Data
were analyzed by two-way ANOVA (sodium butyrate 3 cocaine)
for main effects and interaction effect. For multiple comparisons,
one-way ANOVA with post hoc tests was performed.
Virus-Mediated Gene Transfer
HDAC4 cDNA, a gift from Dr. Eric Olson at UT Southwestern, was
sub-cloned and packaged into a published bicistronic HSV-GFP vi-
ral vector (Clark et al., 2002; Barrot et al., 2002). Activity of the vector
was verified by demonstrating HDAC4 expression in PC12 cells by
immunoblotting (data not shown). HSV-HDAC4 or HSV-GFP (used
as a control) vectors were injected into the mouse nucleus accum-
bens by stereotaxic surgery. Mice were used because a single injec-
tion of virus can infect a much larger portion of the nucleus accum-
bens and thereby induce a greater behavioral effect. The following
coordinates were used: +1.6 mm A/P, +1.5 mm lateral, and 24.4
mm D/V from bregma (relative to dura). Mice were anesthetized, vi-
rus was delivered bilaterally using a pump (at a rate of 0.1 ml/min for
a total volume of 0.5 ml on each side), and behavior was tested 3–5
days after viral injection, when transgene expression is maximal
(Barrot et al., 2002).
Conditioned Place Preference Behavior
Place conditioning was conducted as described (Rahman et al.,
2003). For the TSA experiments, mice received either TSA (2 mg/
kg of body weight i.p.) or DMSO 30 min prior to the saline or cocaine
(5 mg/kg) injection. Conditioning involved one pairing a day for
4 days with the test conducted on day 5. TSA was formulated as
2 mg/ml in DMSO and then diluted 1:5 with phosphate-buffered sa-
line. DMSO was diluted similarly for the vehicle control group. For
the HSV experiments, HSV-HDAC4- or HSV-GFP-injected mice
were paired four times on two consecutive days, followed by testing
on days 3–5 of viral injection.
Supplemental Data
Supplemental Data include a table and can be found with this article
online at http://www.neuron.org/cgi/content/full/48/2/303/DC1/.
Acknowledgments
We would like to thank Dr. Devanjan Sikder (UT Southwestern), for
technical advice and a critical reading of the manuscript, and
Dr. Eric Olson (UT Southwestern), for providing HDAC4 cDNA.
This work was supported by grants from the NIDA and NIMH.
Received: February 3, 2005
Revised: August 4, 2005
Accepted: September 19, 2005
Published: October 19, 2005
References
Alarcon, J.M., Malleret, G., Touzani, K., Vronskaya, S., Ishii, S., Kan-
del, E.R., and Barco, A. (2004). Chromatin acetylation, memory, and
LTP are impaired in CBP+/2 mice: a model for the cognitive deficit
in Rubinstein-Taybi syndrome and its amelioration. Neuron 42,
947–959.
Andersson, M., Konradi, C., and Cenci, M.A. (2001). cAMP response
element-binding protein is required for dopamine-dependent gene
expression in the intact but not the dopamine-denervated striatum.
J. Neurosci. 21, 9930–9943.
Ang, E., Chen, J., Zagouras, P., Magna, H., Holland, J., Schaeffer,
E., and Nestler, E.J. (2001). Induction of nuclear factor-kappaB in
nucleus accumbens by chronic cocaine administration. J. Neuro-
chem. 79, 221–224.
Barrot, M., Olivier, J.D.A., Perrotti, L.I., Impey, S., Storm, D.R., Neve,
R.L., Zachariou, V., and Nestler, E.J. (2002). CREB activity in the nu-
cleus accumbens shell controls gating of behavioral responses to
emotional stimuli. Proc. Natl. Acad. Sci. USA 99, 11435–11440.
Bibb, J.A., Chen, J., Taylor, J.R., Svenningsson, P., Nishi, A.,
Snyder, G.L., Yan, Z., Sagawa, Z.K., Ouimet, C.C., Nairn, A.C.,
et al. (2001). Effects of chronic exposure to cocaine are regulated
by the neuronal protein Cdk5. Nature 410, 376–380.
Carlezon, W.A., Jr., Thome, J., Olson, V.G., Lane-Ladd, S.B., Brod-
kin, E.S., Hiroi, N., Duman, R.S., Neve, R.L., and Nestler, E.J. (1998).
Regulation of cocaine reward by CREB. Science 282, 2272–2275.
Chakrabarti, S.K., James, J.C., and Mirmira, R.G. (2002). Quantita-
tive assessment of gene targeting in vitro and in vivo by the pancre-
atic transcription factor, Pdx1. Importance of chromatin structure in
directing promoter binding. J. Biol. Chem. 277, 13286–13293.
Chang, J.G., Hsieh-Li, H.M., Jong, Y.J., Wang, N.M., Tsai, C.H., and
Li, H. (2001). Treatment of spinal muscular atrophy by sodium bu-
tyrate. Proc. Natl. Acad. Sci. USA 98, 9808–9813.
Chen, J., Kelz, M.B., Hope, B.T., Nakabeppu, Y., and Nestler, E.J.
(1997). Chronic Fos-related antigens: stable variants of deltaFosB
induced in brain by chronic treatments. J. Neurosci. 17, 4933–4941.
Chen, J., Zhang, Y., Kelz, M.B., Steffen, C., Ang, E.S., Zeng, L., and
Nestler, E.J. (2000). Induction of cyclin-dependent kinase 5 in the
hippocampus by chronic electroconvulsive seizures: role of DFosB.
J. Neurosci. 20, 8965–8971.
Chen, J.S., Nye, H.E., Kelz, M.B., Hiroi, N., Nakabeppu, Y., Hope,
B.T., and Nestler, E.J. (1995). Regulation of DFosB and FosB-like
proteins by electroconvulsive seizure (ECS) and cocaine treat-
ments. Mol. Pharmacol. 48, 880–889.
Cheung, P., Allis, C.D., and Sassone-Corsi, P. (2000). Signaling to
chromatin through histone modifications. Cell 103, 263–271.
Clark, M.S., Sexton, T.J., McClain, M., Root, D., Kohen, R., and
Neumaier, J.F. (2002). Overexpression of 5-HT1B receptor in dorsal
raphe nucleus using Herpes Simplex Virus gene transfer increases
anxiety behavior after inescapable stress. J. Neurosci. 22, 4550–
4562.
Clayton, A.L., Rose, S., Barratt, M.J., and Mahadevan, L.C. (2000).
Phosphoacetylation of histone H3 on c-fos- and c-jun-associated
nucleosomes upon gene activation. EMBO J. 19, 3714–3726.
Colby, C.R., Whisler, K., Steffen, C., Nestler, E.J., and Self, D.W.
(2003). Striatal cell type-specific overexpression of DeltaFosB en-
hances incentive for cocaine. J. Neurosci. 23, 2488–2493.
Crosio, C., Heitz, E., Allis, C.D., Borrelli, E., and Sassone-Corsi, P.
(2003). Chromatin remodeling and neuronal response: multiple sig-
naling pathways induce specific histone H3 modifications and early
gene expression in hippocampal neurons. J. Cell Sci. 116, 4905–
4914.
Daunais, J.B., Roberts, D.C., and McGinty, J.F. (1993). Cocaine self-
administration increases preprodynorphin, but not c-fos, mRNA in
rat striatum. Neuroreport 4, 543–546.
Felsenfeld, G., and Groudine, M. (2003). Controlling the double he-
lix. Nature 421, 448–453.
Ferrante, R.J., Kubilus, J.K., Lee, J., Ryu, H., Beesen, A., Zucker, B.,
Smith, K., Kowall, N.W., Ratan, R.R., Luthi-Carter, R., and Hersch,
S.M. (2003). Histone deacetylase inhibition by sodium butyrate che-
motherapy ameliorates the neurodegenerative phenotype in Hun-
tington’s disease mice. J. Neurosci. 23, 9418–9427.
Graybiel, A.M., Moratalla, R., and Robertson, H.A. (1990). Amphet-
amine and cocaine induce drug-specific activation of the c-fos
gene in striosome-matrix compartments and limbic subdivisions
of the striatum. Proc. Natl. Acad. Sci. USA 87, 6912–6916.
Grimm, J.W., Lu, L., Hayashi, T., Hope, B.T., Su, T.P., and Shaham,
Y. (2003). Time-dependent increases in brain-derived neurotrophic
factor protein levels within the mesolimbic dopamine system after
withdrawal from cocaine: implications for incubation of cocaine
craving. J. Neurosci. 23, 742–747.
Guan, Z., Giustetto, M., Lomvardas, S., Kim, J.H., Miniaci, M.C.,
Schwartz, J.H., Thanos, D., and Kandel, E.R. (2002). Integration of
long-term-memory-related synaptic plasticity involves bidirectional
regulation of gene expression and chromatin structure. Cell 111,
483–493.
Hiroi, N., Brown, J., Haile, C., Ye, H., Greenberg, M.E., and Nestler,
E.J. (1997). FosB mutant mice: Loss of chronic cocaine induction of
Fos-related proteins and heightened sensitivity to cocaine’s psy-
chomotor and rewarding effects. Proc. Natl. Acad. Sci. USA 94,
10397–10402.

Neuron314
Hockly, E., Richon, V.M., Woodman, B., Smith, D.L., Zhou, X., Rosa,
E., Sathasivam, K., Ghazi-Noori, S., Mahal, A., Lowden, P.A., et al.
(2003). Suberoylanilide hydroxamic acid, a histone deacetylase in-
hibitor, ameliorates motor deficits in a mouse model of Hunting-
ton’s disease. Proc. Natl. Acad. Sci. USA 100, 2041–2046.
Hope, B., Kosofsky, B., Hyman, S.E., and Nestler, E.J. (1992). Reg-
ulation of immediate early gene expression and AP-1 binding in the
rat nucleus accumbens by chronic cocaine. Proc. Natl. Acad. Sci.
USA 89, 5764–5768.
Hope, B.T., Nye, H.E., Kelz, M.B., Self, D.W., Iadarola, M.J., Naka-
beppu, Y., Duman, R.S., and Nestler, E.J. (1994). Induction of
a long-lasting AP-1 complex composed of altered Fos-like proteins
in brain by chronic cocaine and other chronic treatments. Neuron
13, 1235–1244.
Huang, Y., Doherty, J.J., and Dingledine, R. (2002). Altered histone
acetylation at glutamate receptor 2 and brain-derived neurotrophic
factor genes is an early event triggered by status epilepticus.
J. Neurosci. 22, 8422–8428.
Hyman, S.E., and Malenka, R.C. (2001). Addiction and the brain: the
neurobiology of compulsion and its persistence. Nat. Rev. Neuro-
sci. 2, 695–703.
Jenuwein, T., and Allis, C.D. (2001). Translating the histone code.
Science 293, 1074–1080.
Johnson, K.D., and Bresnick, E.H. (2002). Dissecting long-range
transcriptional mechanisms by chromatin immunoprecipitation.
Methods 26, 27–36.
Kadam, S., and Emerson, B.M. (2003). Transcriptional specificity of
human SWI-SNF BRG1 and BRM chromatin remodeling com-
plexes. Mol. Cell 11, 377–389.
Kalivas, P.W. (2004). Recent understanding in the mechanisms of
addiction. Curr. Psychiatry Rep. 6, 347–351.
Kelz, M.B., Chen, J., Carlezon, W.A., Jr., Whisler, K., Gilden, L.,
Beckmann, A.M., Steffen, C., Zhang, Y.J., Marotti, L., Self, D.W.,
et al. (1999). Expression of the transcription factor deltaFosB in
the brain controls sensitivity to cocaine. Nature 401, 272–276.
Korzus, E., Rosenfeld, M.G., and Mayford, M. (2004). CBP histone
acetyltransferase activity is a critical component of memory con-
solidation. Neuron 42, 961–972.
Levenson, J.M., O’Riordan, K.J., Brown, K.D., Trinh, M.A., Molfese,
D.L., and Sweatt, J.D. (2004). Regulation of histone acetylation dur-
ing memory formation in the hippocampus. J. Biol. Chem. 279,
40545–40559.
Li, J., Guo, Y., Schroeder, F.A., Youngs, R.M., Schmidt, T.W., Ferris,
C., Konradi, C., and Akbarian, S. (2004). Dopamine D2-like antago-
nists induce chromatin remodeling in striatal neurons through cy-
clic AMP-protein kinase A and NMDA receptor signaling. J. Neuro-
chem. 90, 1117–1131.
Marks, P.A., Richon, V.M., Breslow, R., and Rifkind, R.A. (2001).
Histone deacetylase inhibitors as new cancer drugs. Curr. Opin.
Oncol. 13, 477–483.
Mattson, M.P. (2003). Methylation and acetylation in nervous sys-
tem development and neurodegenerative disorders. Ageing Res
Rev 2, 329–342.
McClung, C.A., and Nestler, E.J. (2003). Regulation of gene expres-
sion and cocaine reward by CREB and DeltaFosB. Nat. Neurosci. 6,
1208–1215.
Minamiyama, M., Katsuno, M., Adachi, H., Waza, M., Sang, C.,
Kobayashi, Y., Tanaka, F., Doyu, M., Inukai, A., and Sobue, G.
(2004). Sodium butyrate ameliorates phenotypic expression in
a transgenic mouse model of spinal and bulbar muscular atrophy.
Hum. Mol. Genet. 13, 1183–1192.
Moratalla, R., Elibol, B., Vallejo, M., and Graybiel, A.M. (1996).
Network-level changes in expression of inducible Fos-Jun proteins
in the striatum during chronic cocaine treatment and withdrawal.
Neuron 17, 147–156.
Narlikar, G.J., Fan, H.Y., and Kingston, R.E. (2002). Cooperation be-
tween complexes that regulate chromatin structure and transcrip-
tion. Cell 108, 475–487.
Neely, K.E., and Workman, J.L. (2002). The complexity of chromatin
remodeling and its links to cancer. Biochim. Biophys. Acta 1603,
19–29.
Nestler, E.J. (2001). Molecular basis of long-term plasticity underly-
ing addiction. Nat. Rev. Neurosci. 2, 119–128.
Nestler, E.J., Barrot, M., and Self, D.W. (2001). DFosB: A molecular
switch for addiction. Proc. Natl. Acad. Sci. USA 98, 11042–11046.
Norrholm, S.D., Bibb, J.A., Nestler, E.J., Ouimet, C.C., Taylor, J.R.,
and Greengard, P. (2003). Cocaine-induced proliferation of den-
dritic spines in nucleus accumbens is dependent on the activity
of cyclin-dependent kinase-5. Neuroscience 116, 19–22.
Peakman, M.C., Colby, C., Perrotti, L.I., Tekumalla, P., Carle, T.,
Ulery, P., Chao, J., Duman, C., Steffen, C., Monteggia, L., et al.
(2003). Inducible, brain region-specific expression of a dominant
negative mutant of c-Jun in transgenic mice decreases sensitivity
to cocaine. Brain Res. 970, 73–86.
Persico, A.M., Schindler, C.W., O’Hara, B.F., Brannock, M.T., and
Uhl, G.R. (1993). Brain transcription factor expression: effects of
acute and chronic amphetamine and injection stress. Brain Res.
Mol. Brain Res. 20, 91–100.
Rahman, Z., Schwarz, J., Zachariou, V., Gold, S.J., Wein, M., Choi,
K.H., Kovoor, A., Chen, C.K., DiLeone, R.J., Schwarz, S.C., et al.
(2003). RGS9 modulates dopamine signaling in striatum. Neuron
38, 941–952.
Self, D.W., Barnhart, W.J., Lehman, D.A., and Nestler, E.J. (1996).
Opposite modulation of cocaine-seeking behavior by D1- and D2-
like dopamine receptor agonists. Science 271, 1586–1589.
Steffan, J.S., Bodai, L., Pallos, J., Poelman, M., McCampbell, A.,
Apostol, B.L., Kazantsev, A., Schmidt, E., Zhu, Y.Z., Greenwald,
M., et al. (2001). Histone deacetylase inhibitors arrest polyglut-
amine-dependent neurodegeneration in Drosophila. Nature 413,
739–743.
Sun, Z.W., and Allis, C.D. (2002). Ubiquitination of histone H2B reg-
ulates H3 methylation and gene silencing in yeast. Nature 418, 104–
108.
Sutton, M.A., Schmidt, E.F., Choi, K.H., Schad, C.A., Whisler, K.,
Simmons, D., Karanian, D.A., Monteggia, L.M., Neve, R.L., and
Self, D.W. (2003). Extinction-induced upregulation in AMPA recep-
tors reduces cocaine-seeking behaviour. Nature 421, 70–75.
Tsankova, N.M., Kumar, A., and Nestler, E.J. (2004). Histone mod-
ifications at gene promoter regions in rat hippocampus after acute
and chronic electroconvulsive seizures. J. Neurosci. 24, 5603–5610.
Turner, B.M. (2002). Cellular memory and the histone code. Cell
111, 285–291.
Turner, B.M., Birley, A.J., and Lavender, J. (1992). Histone H4 iso-
forms acetylated at specific lysine residues define individual chro-
mosomes and chromatin domains in Drosophila polytene nuclei.
Cell 69, 375–384.
Wells, J., and Farnham, P.J. (2002). Characterizing transcription
factor binding sites using formaldehyde crosslinking and immuno-
precipitation. Methods 26, 48–56.
Yao, W.D., Gainetdinov, R.R., Arbuckle, M.I., Sotnikova, T.D., Cyr,
M., Beaulieu, J.M., Torres, G.E., Grant, S.G., and Caron, M.G.
(2004). Identification of PSD-95 as a regulator of dopamine-
mediated synaptic and behavioral plasticity. Neuron 41, 625–638.
Young, S.T., Porrino, L.J., and Iadarola, M.J. (1991). Cocaine indu-
ces striatal c-fos immunoreactive proteins via dopaminergic D1 re-
ceptors. Proc. Natl. Acad. Sci. USA 88, 1291–1295.
Yuferov, V., Kroslak, T., Laforge, K.S., Zhou, Y., Ho, A., and Kreek,
M.J. (2003). Differential gene expression in the rat caudate putamen
after ‘‘binge’’ cocaine administration: advantage of triplicate micro-
array analysis. Synapse 48, 157–169.
Zeitlin, S.G., Barber, C.M., Allis, C.D., and Sullivan, K.F. (2001). Dif-
ferential regulation of CENP-A and histone H3 phosphorylation in
G2/M. J. Cell Sci. 114, 653–661.




















