
Differential distribution of estrogen receptor (ER)-α and ER-β in the midbrain raphe nuclei and...
12
DIFFERENTIAL DISTRIBUTION OF ESTROGEN RECEPTOR (ER)- AND ER- IN THE MIDBRAIN RAPHE NUCLEI AND PERIAQUEDUCTAL GRAY IN MALE MOUSE: PREDOMINANT ROLE OF ER- IN MIDBRAIN SEROTONERGIC SYSTEMS M. NOMURA, a1 K. T. AKAMA, b S. E. ALVES, b K. S. KORACH, c J.-Å. GUSTAFSSON, d D. W. PFAFF a AND S. OGAWA a * a Laboratory of Neurobiology and Behavior, The Rockefeller University, Box 275, 1230 York Avenue, New York, NY 10021, USA b Laboratory of Neuroendocrinology, The Rockefeller University, New York, NY 10021, USA c Laboratory of Reproductive and Developmental Toxicology, National Institute of Environmental Health Sciences, National Institutes of Health, Research Triangle Park, NC 27709, USA d Department of Medical Nutrition, Karolinska Institute, Huddinge S141-86, Sweden Abstract—We examined the distribution of estrogen receptor (ER)- and ER- immunoreactive (ir) cells in the dorsal (DRN) and median/paramedian (MPRN) raphe nuclei in male mice. ER- ir neurons were scattered across the three subdivisions (ventral, dorsal, and lateral) of the DRN and the MPRN. Ro- bust ER- ir cells were observed throughout the raphe nuclei, and were particularly abundant in the ventral and dorsal subdivisions of the DRN. Using dual-label immunocytochem- istry for ER- or ER- with tryptophan hydroxylase (TPH), the rate-limiting enzyme for 5-hydroxytryptamine (5-HT) synthe- sis, over 90% of ER- ir cells exhibited TPH-ir in all DRN subdivisions, whereas only 23% of ER- ir cells contained TPH. Comparisons of ER- knockout (ERKO) as well as ER- knockout (ERKO) mice with their respective wild-type (WT) littermates revealed that gene disruption of either ER- or ER- did not affect the other ER subtype expression in the raphe nuclei. In situ hybridization histochemistry revealed that there was a small but statistically significant decrease in TPH mRNA expression in the ventral DRN subdivision in ERKO mice compared with WT mice, whereas TPH mRNA levels were not affected in ERKO mice. These findings sup- port a hypothesis that ER- activation may contribute to the estrogenic regulation of neuroendocrine and behavioral functions, in part, by acting directly on 5-HT neurons in the raphe nuclei in male mice. © 2004 IBRO. Published by Elsevier Ltd. All rights reserved. Key words: serotonin, aggression, tryptophan hydroxylase, sexual behavior, knockout mouse, immunocytochemistry. It is well established that estradiol (E 2 ), an aromatized metabolite of testosterone, plays a crucial role in neuroen- docrine processes and reproduction-related behaviors in males (Meisel and Sachs, 1994; Ogawa et al., 2002; Olsen, 1992; Simon and Whalen, 1987). These estrogenic actions are mediated through at least two subtypes of estrogen receptors (ERs), ER- (Green et al., 1986) and ER- (Kuiper et al., 1996; Tremblay et al., 1997), which have similar binding affinities for the endogenous ligand, E 2 . Both ERs act as ligand-activated transcription factors that regulate the transcription of target genes. A series of behavioral studies using single (ERKO and ERKO) and double (ERKO) knockout mice for ER genes have re- vealed that activation of ER- and ER- is indeed involved in the regulation of sexual and aggressive behaviors in male mice (Imwalle et al., 2002; Nomura et al., 2002; Ogawa et al., 1997, 1998, 1999, 2000, 2002; Rissman et al., 1999; Scordalakes and Rissman, 2003; Wersinger and Rissman, 2000; Wersinger et al., 1997). These studies also collectively suggest that ER- and ER- might play different roles for the control of male sexual and aggres- sive behaviors. Neuroanatomical studies have revealed that although both types of ERs are widely distributed in brain regions responsible for sociosexual behaviors, such as the hypo- thalamus, limbic system, and midbrain, their distribution patterns are not completely overlapping (Laflamme et al., 1998; Mitra et al., 2003; Shughrue et al., 1997a). Among them, the ascending midbrain raphe nuclei, consisting of the dorsal (DRN) and median/paramedian (MPRN) raphe, and the midbrain periaqueductal gray (PAG) are particu- larly interesting. It is well established that these midbrain regions are involved in the control of male sexual and aggressive behaviors based on lesion, electrical stimula- tion, and anatomical tracing studies (File and Deakin, 1980; Murphy and Hoffman, 2001; Vergnes et al., 1988; Yamamoto and Ueki, 1977). Several studies have shown that both DRN and PAG exhibit ER protein and mRNA in a number of species including mice (Alves et al., 1998, 2000; Lu et al., 2001; Mitra et al., 2003; Murphy et al., 1999; Nomura et al., 2003; Shughrue and Merchenthaler, 2001; Turcotte and Blaustein, 1993; VanderHorst et al., 1998). These studies have consistently shown that ER- immu- noreactive (ir) cells are localized in both the raphe nucleus 1 Present address: Department of Urology, University of Occupational and Environmental Health, 1-1 Iseigaolea, Yahatanishi-ku, Kitaky- ushu-city 807-8555, Japan. *Corresponding author. Tel: 1-212-327-8667; fax: 1-212-327-8664. E-mail address: [email protected] (S. Ogawa). Abbreviations: ANOVA, analysis of variance; BNST, bed nucleus of stria terminalis; DAB, 3,3=-diaminobenzidine; DRN, dorsal raphe; ER, estrogen receptor; ir, immunoreactive; MPOA, medial preoptic area; MPRN, median/paramedian raphe; nPGi, nucleus paragigantocellu- laris; PAG, periaqueductal gray; PB, phosphate buffer; PBS, phos- phate buffer saline; PR, progesterone receptor; PVN, paraventricular nucleus; TPH, tryptophan hydroxylase; 5-HT, 5-hydroxytryptamine, serotonin. Neuroscience 130 (2005) 445– 456 0306-4522/05$30.000.00 © 2004 IBRO. Published by Elsevier Ltd. All rights reserved. doi:10.1016/j.neuroscience.2004.09.028 445
Transcript of Differential distribution of estrogen receptor (ER)-α and ER-β in the midbrain raphe nuclei and...

DEGM
MKAa
Bb
Nc
IHd
S
A(aE(basirssTE(ortT�lpefrE
1
au*EAseMlpns
Neuroscience 130 (2005) 445–456
0d
IFFERENTIAL DISTRIBUTION OF ESTROGEN RECEPTOR (ER)-� ANDR-� IN THE MIDBRAIN RAPHE NUCLEI AND PERIAQUEDUCTALRAY IN MALE MOUSE: PREDOMINANT ROLE OF ER-� IN
IDBRAIN SEROTONERGIC SYSTEMSKs
ImdmOaeEhEtbdvimOaRads
brtp1ttalrat1YtnLNTT
. NOMURA,a1 K. T. AKAMA,b S. E. ALVES,b
. S. KORACH,c J.-Å. GUSTAFSSON,d D. W. PFAFFa
ND S. OGAWAa*
Laboratory of Neurobiology and Behavior, The Rockefeller University,ox 275, 1230 York Avenue, New York, NY 10021, USA
Laboratory of Neuroendocrinology, The Rockefeller University,ew York, NY 10021, USA
Laboratory of Reproductive and Developmental Toxicology, Nationalnstitute of Environmental Health Sciences, National Institutes ofealth, Research Triangle Park, NC 27709, USA
Department of Medical Nutrition, Karolinska Institute, Huddinge141-86, Sweden
bstract—We examined the distribution of estrogen receptorER)-� and ER-� immunoreactive (ir) cells in the dorsal (DRN)nd median/paramedian (MPRN) raphe nuclei in male mice.R-� ir neurons were scattered across the three subdivisions
ventral, dorsal, and lateral) of the DRN and the MPRN. Ro-ust ER-� ir cells were observed throughout the raphe nuclei,nd were particularly abundant in the ventral and dorsalubdivisions of the DRN. Using dual-label immunocytochem-stry for ER-� or ER-� with tryptophan hydroxylase (TPH), theate-limiting enzyme for 5-hydroxytryptamine (5-HT) synthe-is, over 90% of ER-� ir cells exhibited TPH-ir in all DRNubdivisions, whereas only 23% of ER-� ir cells containedPH. Comparisons of ER-� knockout (�ERKO) as well asR-� knockout (�ERKO) mice with their respective wild-type
WT) littermates revealed that gene disruption of either ER-�r ER-� did not affect the other ER subtype expression in theaphe nuclei. In situ hybridization histochemistry revealedhat there was a small but statistically significant decrease inPH mRNA expression in the ventral DRN subdivision inERKO mice compared with �WT mice, whereas TPH mRNA
evels were not affected in �ERKO mice. These findings sup-ort a hypothesis that ER-� activation may contribute to thestrogenic regulation of neuroendocrine and behavioralunctions, in part, by acting directly on 5-HT neurons in theaphe nuclei in male mice. © 2004 IBRO. Published bylsevier Ltd. All rights reserved.
Present address: Department of Urology, University of Occupationalnd Environmental Health, 1-1 Iseigaolea, Yahatanishi-ku, Kitaky-shu-city 807-8555, Japan.Corresponding author. Tel: �1-212-327-8667; fax: �1-212-327-8664.-mail address: [email protected] (S. Ogawa).bbreviations: ANOVA, analysis of variance; BNST, bed nucleus oftria terminalis; DAB, 3,3=-diaminobenzidine; DRN, dorsal raphe; ER,strogen receptor; ir, immunoreactive; MPOA, medial preoptic area;PRN, median/paramedian raphe; nPGi, nucleus paragigantocellu-
aris; PAG, periaqueductal gray; PB, phosphate buffer; PBS, phos-hate buffer saline; PR, progesterone receptor; PVN, paraventricular
nucleus; TPH, tryptophan hydroxylase; 5-HT, 5-hydroxytryptamine,erotonin.
306-4522/05$30.00�0.00 © 2004 IBRO. Published by Elsevier Ltd. All rights reseroi:10.1016/j.neuroscience.2004.09.028
445
ey words: serotonin, aggression, tryptophan hydroxylase,exual behavior, knockout mouse, immunocytochemistry.
t is well established that estradiol (E2), an aromatizedetabolite of testosterone, plays a crucial role in neuroen-ocrine processes and reproduction-related behaviors inales (Meisel and Sachs, 1994; Ogawa et al., 2002;lsen, 1992; Simon and Whalen, 1987). These estrogenicctions are mediated through at least two subtypes ofstrogen receptors (ERs), ER-� (Green et al., 1986) andR-� (Kuiper et al., 1996; Tremblay et al., 1997), whichave similar binding affinities for the endogenous ligand,
2. Both ERs act as ligand-activated transcription factorshat regulate the transcription of target genes. A series ofehavioral studies using single (�ERKO and �ERKO) andouble (��ERKO) knockout mice for ER genes have re-ealed that activation of ER-� and ER-� is indeed involvedn the regulation of sexual and aggressive behaviors in
ale mice (Imwalle et al., 2002; Nomura et al., 2002;gawa et al., 1997, 1998, 1999, 2000, 2002; Rissman etl., 1999; Scordalakes and Rissman, 2003; Wersinger andissman, 2000; Wersinger et al., 1997). These studieslso collectively suggest that ER-� and ER-� might playifferent roles for the control of male sexual and aggres-ive behaviors.
Neuroanatomical studies have revealed that althoughoth types of ERs are widely distributed in brain regionsesponsible for sociosexual behaviors, such as the hypo-halamus, limbic system, and midbrain, their distributionatterns are not completely overlapping (Laflamme et al.,998; Mitra et al., 2003; Shughrue et al., 1997a). Amonghem, the ascending midbrain raphe nuclei, consisting ofhe dorsal (DRN) and median/paramedian (MPRN) raphe,nd the midbrain periaqueductal gray (PAG) are particu-
arly interesting. It is well established that these midbrainegions are involved in the control of male sexual andggressive behaviors based on lesion, electrical stimula-ion, and anatomical tracing studies (File and Deakin,980; Murphy and Hoffman, 2001; Vergnes et al., 1988;amamoto and Ueki, 1977). Several studies have shown
hat both DRN and PAG exhibit ER protein and mRNA in aumber of species including mice (Alves et al., 1998, 2000;u et al., 2001; Mitra et al., 2003; Murphy et al., 1999;omura et al., 2003; Shughrue and Merchenthaler, 2001;urcotte and Blaustein, 1993; VanderHorst et al., 1998).hese studies have consistently shown that ER-� immu-
oreactive (ir) cells are localized in both the raphe nucleusved.

aeefieygol2estept�r
o5rit(Q1iwT�alceidstTelltcm
M
G�aommoEfi
pawnne
I
Tsvaib4nc�
aAb2bgmU(MaaosTaoiwiiscP
D
Afmwi(ihaciItd
I
Tw�a
M. Nomura et al. / Neuroscience 130 (2005) 445–456446
nd the PAG, whereas ER-� ir as well as ER-� mRNA arexpressed in the raphe nuclei, but not in the PAG. How-ver, most of these anatomical studies are performed inemales and the detailed distribution of both ER subtypesn the male raphe nuclei and PAG along the rostro-caudalxtent or in the different subdivisions of the DRN has notet been described. Furthermore, it is not known whetherene disruption of one ER may affect the expression of thether subtype in the DRN and PAG, as found in hypotha-
amic brain regions (Nomura et al., 2003; Temple et al.,001). Therefore, in the first part of the present study, wexamined the distribution of ER-� or ER-� ir cells in threeubdivisions (i.e. dorsal, ventral, and lateral) of the DRN,he MPRN, and the PAG along the entire rostro-caudalxtent in gonadally intact male mice. In order to explore theossible effects of either ER-� or ER-� gene disruption onhe expression of the other subtype of ER, we utilizedERKO and �ERKO mice and compared results with theirespective WT siblings.
One of the targets of E2 action in the DRN for the controlf sociosexual behaviors is serotonin (5-hydroxytryptamine;-HT) synthesizing neurons. It has been shown that E2 di-ectly modulates the midbrain serotonergic system includingncreasing the expression of tryptophan hydroxylase (TPH),he rate-limiting enzyme for 5-HT synthesis, 5-HT transporterSERT), and the autoreceptor, 5-HT1A (Lu et al., 1999; Mc-ueen et al., 1997, 1999; Pecins-Thompson and Bethea,999; Pecins-Thompson et al., 1996, 1998). Recent studies
n rats and macaques have also shown that TPH co-localizesith ER-� in the DRN (Gundlah et al., 2001; Lu et al., 2001).hese findings suggest that behavioral modifications inERKO, �ERKO, and ��ERKO mice might be partly due toltered estrogenic actions on the serotonergic system at the
evel of the midbrain raphe nuclei. However, the degree ofo-localization of TPH with ER-� and that with ER-� in differ-nt subdivisions of the DRN have not been directly compared
n the same animals. Furthermore, it is still unknown howeletion of ER-� or ER-� genes may affect the TPH expres-ion in the raphe nuclei. In the second part of the study,herefore, we compared the numbers of cells co-expressingPH and ER-� ir or those co-expressing TPH and ER-� ir inach subdivision of the DRN in WT control mice, using dual-
abel immunocytochemistry. In addition, we examined theevels of TPH mRNA expression in the three subdivisions ofhe DRN and the MPRN in �ERKO and �ERKO mice andompared this expression to those of respective WT controlice.
EXPERIMENTAL PROCEDURES
ice
onadally intact adult male �ERKO (Lubahn et al., 1993) andERKO (Krege et al., 1998) mice and their respective WT (�WTnd �WT) littermates at age 9–14 weeks were used. They werebtained from the �ERKO and the �ERKO breeding coloniesaintained at the Rockefeller University by mating heterozygousale and female mice. Original breeding pairs (mixed backgroundf C57BL/6J and 129) were obtained from the National Institute ofnvironmental Health Sciences. Mice were group-housed (four to
ve mice/cage) in plastic cages (30�20�12 cm) at constant tem- Eerature (22 °C) on a 12-h light/dark cycle. Food and water werevailable ad libitum throughout the experiment. All proceduresere approved by the IACUC of the Rockefeller University andamed guidelines on the ethical use of animals. We minimized theumber of animals used and their suffering during thexperiments.
mmunohistochemistry
otal of 24 mice (n�6/genotype) were deeply anesthetized usingodium pentobarbital (75 mg/kg; ip) and perfused via the leftentricle with phosphate buffer saline (PBS) containing 0.1% hep-rin, pH 7.4, followed by 2% acrolein and 2.5% paraformaldehyde
n 0.1 M phosphate buffer (PB), pH 7.4. After postfixation, therains were equilibrated in PB containing 30% sucrose for 48 h at°C. The brain tissues were frozen with powdered dry ice, coro-
ally sectioned at 40 �m on a freezing microtome, and stored inryoprotectant (30% glycerol, 30% ethylene glycol in 0.1 M PB) at20 °C until they were used.
Free floating sections were thoroughly washed in 0.1 M PBSnd then were washed in 0.1% sodium borohydride (NaBH4).fter several washes in PBS, endogenous peroxidase activity waslocked by washing tissue in 0.1% hydrogen peroxide in PBS for0 min. Sections were washed several times in PBS and thenlocked with either 2% normal rabbit (for detection of ER-�) oroat serum (for ER-�). Sections were then incubated in a ratonoclonal ER-� antibody, H222 (1:1000; a gift of Dr. G. Greene,niversity of Chicago) or a rabbit polyclonal ER-� antibody, Z8P
1:10,000; Zymed Laboratory Inc., CA, USA; Shughrue anderchenthaler, 2001) in 0.1 M PBS containing 0.5% Triton X-100nd 2% normal rabbit (for ER-�) or goat serum (for ER-�) for 48 ht 4 °C. The sections were further incubated with a 1:200 dilutionf the biotinylated anti-rat (for ER-�) or anti-rabbit (for ER-�)econdary antibody (Vector Laboratories) in PBS containing 0.5%riton X-100 and 2% normal serum for 120 min at room temper-ture. The sections were finally incubated with avidin–biotin per-xidase complex (Vectastain ABC Elite Kit; Vector Laboratories)
n PBS for 90 min. Following several washes in PBS, the sectionsere developed with 0.03% 3,3=-diaminobenzidine (DAB) contain-
ng 0.15% nickel ammonium sulfate and 0.03% hydrogen peroxiden PBS for 3 min. Sections were then mounted on gelatin-coatedlides and air dried. The slides were dehydrated through an as-ending alcohol series, cleared with xylene, and coverslipped withermount (Fisher Scientific).
ual-label immunocytochemistry
separate set of �WT brain samples (n�4) was prepared and theree-floating sections (30 �m thickness) were processed for im-unocytochemistry as described above, except 0.3% skim milkas used instead of normal serum. Sections were first incubated
n an ER-� antibody, H222 (1:1000) or an ER-� antibody, Z8P1:10,000) for 2 days at 4 °C. The peroxide complex was visual-zed with 0.02% DAB containing 2.5% nickel sulfate and 0.01%ydrogen peroxide in a 0.175 M sodium acetate buffer for 2 min,s a dark purple punctate stain. After being washed in PBSontaining 3% skim milk overnight at 4 °C, the sections were thenncubated in a sheep polyclonal TPH antibody (1:8000; Chemiconnternational Inc.) for 2 days at 4 °C. The peroxidase complex inhe section was visualized with 0.05% DAB containing 0.1% hy-rogen peroxide for 2 min, as a light brown cytoplasmic stain.
n situ hybridization histochemistry
otal of 24 mice (n�6/genotype) were decapitated and the brainsere rapidly removed, frozen on powdered dry ice and stored at80 °C until they were used. Frozen section were cut at 12 �m oncryostat and mounted onto coated slides (Fisher Scientific). The
R-� probe was generated from a 2100-bp fragment of the mouse
EDS
so
FtEtt
M. Nomura et al. / Neuroscience 130 (2005) 445–456 447
R-� gene (Genebank No. U81451; kindly supplied byr. V. Giguere, McGill University) subcloned into pBluescript
ig. 1. The number of ER-� (�) or ER-� (B) ir cells along the rostro-cahe medial and MPRN. ER-� gene knockout (�ERKO) and their WT (�R-� gene knockout (�ERKO) and their WT (�WT) littermate mice (n�
he x axis is the distance from the Bregma determined by referring to the meanS.E.M.
K(�) plasmid vector. Antisense or sense ER-� riboprobe was [
ynthesized from the plasmid using SalI and T3 RNA polymeraser EcoRI and T7 RNA polymerase respectively, and labeled with
in the three subdivisions (dorsal, ventral, and lateral) of the DRN andrmate mice (n�6/genotype) were used for the detection of ER-� (A).pe) were used for the detection of ER-� (B). The number depicted onrd mouse brain atlas (Franklin and Paxinos, 1997). Values represent
udal axisWT) litte6/genoty
he standa
35S]UTP. To generate a plasmid DNA template for TPH riboprobe

sGTTfwT(crpGGfsNrdoscE
i15(ansaSMtsuJpSfiaAf
Fns and Paxio
M. Nomura et al. / Neuroscience 130 (2005) 445–456448
ynthesis, a cDNA fragment of the rat TPH gene (831 bp;enbank accession number X53501), which is now renamed asPH1, was first amplified by RT-PCR and then TA-cloned into theOPO-vector pCR4 (Invitrogen). Briefly, total RNA was isolated
rom the left hemisphere of an adult (approximately 350 g, frozenet weight) male rat brain (RNeasy isolation columns; Qiagen).his total RNA was then reverse-transcribed with oligo-dT primerSuperscript II reverse-transcriptase; Gibco BRL) into first-strandDNA template, which was then used in the subsequent PCReaction (AmpliTaq Gold polymerase; Perkin Elmer). The PCRrimers used were forward: 5= GTG CTG TTG TAC GGA TCCAA CTC GAC and reverse: 5= TCC AAA CGG GCG CTT CACG. The amplified product was gel purified, TOPO TA-cloned, and
urther sequenced for verification and orientation. Antisense orense TPH riboprobe was synthesized from the plasmid usingotI and T7 RNA polymerase or SpeI and T3 RNA polymerase,
espectively, and labeled with [35S]UTP. The TPH probe wasesigned by considering an avoidance of homology to c-termini ofther tetrahydropterin-dependent amino acid hydroxylases. Thepecificity of antisense probes for ER-� or TPH transcripts wasonfirmed by each sense-strand probe and the distribution of
ig. 2. Schematic diagrams showing the distribution of cells immunucleus. The distribution of ER-� and ER-� was examined in gonections at the levels of Bregma �4.48, �4.60, or �4.72 (Franklinr ER-�.
R-� ir or TPH cells in immunocytochemical study. In situ hybrid- w
zation procedures have been described previously (Ueta et al.,995). In brief, sections were fixed in 4% paraformaldehyde formin and incubated in saline containing 0.25% acetic anhydride
vol/vol) and 0.1 M triethanolamine for 10 min, then dehydratednd delipidated in chloroform. Hybridization was carried out over-ight at 52 °C (for ER-� transcripts) and 48 °C (for TPH tran-cripts) in 67.5 �l hybridization buffer containing 50% formamidend 4� SSC, 500 �g/ml sheared salmon sperm DNA (Sigma,t. Louis, MO, USA), 250 �g/ml baker’s yeast total RNA (Rocheolecular Biochemicals, Indianapolis, USA), 1� Denhart’s solu-
ion (0.02% Ficoll, 0.02% polyvinylpyrrolidone and 0.02% bovineerum albumin) and 10% dextran sulfate (500,000 mol wt; Sigma)nder a Nescofilm coverslip (Bando Chemical IMD, Ltd., Osaka,apan). Total counts of 1.0�106 CPM/slide for antisense or senserobes were used. After hybridization, sections were rinsed in 2�SC and washed for 30 min in two changes of 2� SSC/50%
ormamide at 52 °C (for ER-�) and 48 °C (for TPH) before beingncubated at 37 °C in RNase buffer (10 mM Tris–HCl, 1 mM EDTA,nd 0.5 M NaCl) containing 1 �g/ml RNase A (Sigma) for 30 min.fter a further 45 min wash in three changes of 2� SSC/50%
ormamide at 52 °C (for ER-�) and 48 °C (for TPH) and 10 min
for ER-� (A) or ER-� (B) along the rostro-caudal axis of the raphetact �WT and �WT mouse, respectively, in three representative
nos, 1997). Each dot represents one immuno-labeled cell for ER-�
o-positiveadally in
ash in two changes in 2� SSC at room temperature, the slides

wHpwUti
Q
IniatairatAE(vltai
(mMacswEoicms�aodmnm
aMCBw
(rtti
S
AcDvfalo
Ddr
EwtMyvl�Edtmdctac
Ddn
ItvEe
Fl ouse br
M. Nomura et al. / Neuroscience 130 (2005) 445–456 449
ere briefly dipped in water and 70% ethanol and air dried.ybridized sections were apposed to autoradiography films (Hy-erfilm; Amersham, Buckinghamshire, UK) for 2 weeks. Slidesere then dipped in nuclear emulsion (Kodak NTB-2; Rochester,SA) and exposed for 6 weeks. All sections for detection of each
ranscript were processed simultaneously to minimize the variabil-ty in hybridization and wash conditions.
uantitative analyses
mmunostained serial sections (40 �m thick for single-label immu-ocytochemistry and 30 �m thick for dual-label immunocytochem-
stry) were anatomically matched by referring to a mouse braintlas (Franklin and Paxinos, 1997), and those sections containing
he raphe nuclei and the PAG were selected for quantitativenalysis. The raphe nuclei were further divided into four divisions,
.e. the ventral, dorsal, and lateral subdivisions and the MPRN byeferring to the standard atlas (Franklin and Paxinos, 1997) as wells by TPH ir staining pattern (we have previously demonstratedhat the boundaries of the raphe nuclei are well defined by TPH ir;lves et al., 1998). Our previous studies revealed that ER-� andR-� cells seemed to be distributed differently within the DRN
Alves et al., 1998, 2000). Furthermore, the lateral DRN and theentromedial/interfascicular DRN may be anatomically, morpho-ogically, and neurochemically unique subdivisions of the DRN inhe gerbil (Janusonis et al., 1999). Therefore, we assumed that thenalysis for each subdivision of the DRN gave us more precise
nformation regarding the specific roles of ER-� and ER-�.For single immunocytochemistry, seven sections for the DRN
i.e. seven data points, approximately Bregma �4.24 to approxi-ately �4.96 with 120 �m intervals), and eight sections for thePRN (i.e. eight data points, approximately Bregma �4.12 topproximately �4.96 with 120 �m intervals) were selected forounting ER-� and ER-� ir expressing cells. In the PAG, 14ections (approximately Bregma �3.52 to approximately �5.02ith 120 �m intervals) were selected for cell counts of ER-� andR-� ir expressing cells, and the counts were averaged for eachf two consecutive sections (i.e. seven data points with 240 �m
ntervals). For dual-label immunocytochemistry, three sectionsontaining the DRN (approximately Bregma �4.48 to approxi-ately �4.72 with 120 �m intervals) were selected for counts of
ingle (ER-� or ER-� ir cells) or double (ER-��TPH or ER-�TPH ir cells) immunostained cells, and the counts were aver-ged over three sections. For the DRN, numbers of cells in eachf three subdivisions were counted separately for both single andouble immunocytochemistry. All cell counts were performedanually by a single experimenter who was unaware of the ge-otype of the samples, at a magnification of 200� on a Nikon lighticroscope (Eclipse E400; Tokyo, Japan).
A quantitative image analysis of in situ hybridization for ER-�nd TPH transcripts was undertaken on developed films using theCID image Analysis System (Imaging Research Inc., Ontario,anada). Five sections containing raphe nuclei (approximatelyregma �4.50 to approximately �4.70 mm, with 50 �m intervals)
ig. 3. Representative photomicrographs showing the distribution ofevels of the DRN correspond to Bregma �4.60 mm in the standard m
ere selected for each mouse by referring to the standard atlas (
Franklin and Paxinos, 1997). Optic density was measured sepa-ately in each of three subdivisions of the DRN (see above) andhe MPRN for each mouse, by a single-blind observer. In addition,he cellular localization of labeled cells was microscopically exam-ned on the emulsion-dipped sections.
tatistics
ll data are presented as a meanS.E.M. The data of the rostro-audal distribution of ER-� and ER-� in each subdivision of theRN, MRN and PAG were analyzed by a two-way analysis ofariance (ANOVA) for repeated measurements for the main ef-ects of genotype and rostro-caudal axis, as a repeated measure,nd their interaction. The data of co-localization and TPH mRNA
evels were analyzed using one-way ANOVAs for the main effectf genotype. A P0.05 was considered statistically significant.
RESULTS
istribution of ER-� ir and effects of ER-� geneisruption on the expression of ER-� in the midbrainaphe nuclei
R-� ir neurons, with weak to moderate staining intensity,ere observed scattered across the three subdivisions of
he DRN (dorsal, ventral and lateral subdivisions) and thePRN of �WT mice (Figs. 1A and 2A). Quantitative anal-
sis revealed that the number of ER-� cells in the dorsal,entral, and lateral subdivisions of the DRN peaked at the
evel of Bregma approximately �4.60 to approximately4.72 (Fig. 1A), whereas in the MPRN, the number ofR-� cells decreased toward caudal levels. Schematiciagrams of the distribution and the representative pho-omicrographs of ER-� cells in the raphe nuclei of �WTice are shown in Figs. 2A and 3A, respectively. Theisruption of the ER-� gene did not cause any significanthanges in the number and distribution of ER-� cells alonghe entire rostro-caudal axis in all subdivisions of the DRNnd the MPRN (Fig. 1A; genotype, genotype�rostro-audal axis, ns).
istribution of ER-� ir and effects of ER-� geneisruption on the expression of ER-� in the rapheuclei
ntense ER-� ir expressing neurons were observed acrosshe DRN, with particularly high expression levels in theentral subdivision of the DRN, in which the total number ofR-� ir cells was twice that of ER-� ir cells. ER-� wasxpressed moderately in the dorsal subdivision of the DRN
), ER-� ir (B) and ER-� transcripts (C) in the DRN of WT mice. Theain atlas (Franklin and Paxinos, 1997). The scale bar�100 �m.
ER-� ir (A
Figs. 1B and 2B). The number of ER-� ir cells in the dorsal

oo�tw1EnpdtmhEtsDagis
DE
Trce
toatc�llliitgDctPntt
CD
Tvls
F�t f ER-� iml dot repre
M. Nomura et al. / Neuroscience 130 (2005) 445–456450
r the ventral subdivisions of the DRN peaked at the levelsf Bregma approximately �4.60 to approximately �4.72 or4.48 to approximately �4.60, respectively (Fig. 1B). In
he lateral subdivision of the DRN, scattered ER-� ir cellsere observed along the entire rostro-caudal axis (Figs.B and 2B). In this area of the DRN, the total number ofR-� ir was about one third that of ER-� ir cells. The totalumber of ER-� ir cells was relatively low in the MPRN,articularly toward the caudal end (Fig. 1B). Schematiciagrams of the distribution and the representative pho-omicrographs of ER-� ir cells in the raphe nuclei of �WTice are shown in Figs. 2B and 3B. In situ hybridizationistochemistry revealed a similar distribution pattern ofR-� transcripts in the three subdivisions of the DRN and
he MPRN as found in the immunocytochemical study ashown in the representative photomicrographs (Fig. 3C).isruption of the ER-� gene also did not alter the numbernd distribution of ER-� ir cells (Fig. 1B; genotype,enotype�rostro-caudal axis, ns) or optical density read-
ngs of ER-� mRNA along the rostro-caudal levels of the allubdivisions of the DRN and the MPRN.
istribution of ER-� and ER-� ir cells and effects ofR-� gene disruption on ER-� ir in the PAG
he number and distribution of ER-� ir cells along theostro-caudal axis of the PAG are shown in Fig. 4. ER-� irells were detected along the entire rostro-caudal levels
ig. 4. (A) The number of ER-� ir expressing cells along the rostro-caERKO mice (n�6/genotype). The number depicted on the x axis is t
he meanS.E.M. (B) Schematic diagrams showing the distribution oevels of Bregma �3.52, �4.00, �4.48, and �4.96 are shown. Each
xamined, but the number of ER-� cells increased toward c
he caudal levels of the PAG (Fig. 4A). Within the rostralne-third of the PAG (Bregma approximately �3.52 topproximately �4.04 mm), ER-� cells were scattered inhe dorsal and lateral parts of the PAG (Fig. 4B). Within theaudal two-thirds of the PAG (Bregma approximately4.04 to approximately �4.96 mm), ER-� ir cells were
ocalized in two densely populated clusters, one in theateral and the other in the dorsal PAG (Fig. 4B). Particu-arly, within the caudal one-third (Bregma �4.48 to approx-mately �4.96 mm), the number of ER-� cells greatlyncreased in the lateral PAG (Fig. 4B). We also noticed thathe intensity of immunostaining for ER-� in the PAG wasreater than that in the raphe nuclei (data not shown).isruption of the ER-� gene did not cause any significanthanges in the total number of ER-� cell in the PAG andhe number of ER-� cell in all rostro-caudal levels of theAG (Fig. 4A; genotype, genotype�rostro-caudal axis,s). Finally, in contrast to ER-�, few ER-� ir cells and ER-�ranscripts (data not shown) were observed in any part ofhe PAG in �WT or in �ERKO mice.
o-localization of ER-� or ER-� with TPH ir in theRN of �WT mice
he level of co-localization between ER-� ir and TPH iraried among the DRN subdivisions (Table 1). The highestevel of co-localization of ER-� with TPH was in the ventralubdivision of the DRN, where 55% of ER-� cells also
of the PAG (Bregma approximately �3.52 to �4.96 mm) in �WT andce from the Bregma (Franklin and Paxinos, 1997). Values representmuno-positive cells in the PAG. Four representative sections at the
sents one immunolabeled cell.
udal axishe distan
ontained TPH ir. In the dorsal subdivision of the DRN,

alccosclacPcctFeocs
Ee
Ttgsstss6m6MiretTcd
Ttsm
mEonimpbEsvsEgttratuad
Dw
ToirewopwtsEwEc
oricc
T
EEEE
1
2
*
M. Nomura et al. / Neuroscience 130 (2005) 445–456 451
pproximately 30% of ER-� cells contained TPH ir. In theateral subdivision of the DRN, however, only 6% of ER-�ells co-expressed TPH ir. On average, 23% of ER-� cellso-localized with TPH ir in the DRN. In contrast, over 90%f the ER-� expressing cells contained TPH ir in all theubdivisions of the DRN (Table 1). The percentages ofo-localization of ER-� with TPH in the ventral, dorsal, and
ateral subdivisions of the DRN and the whole DRN werepproximately 98, 95, 93, and 96%, respectively, signifi-antly higher than for ER-� in the same regions (Table 1,0.01). It should be noted that significantly higher per-entages of co-localization between ER-� and TPH inomparison to that of ER-� and TPH were not simply dueo the differences in the number of ER-� or ER-� ir cells.or instance, in the lateral DRN, most ER-� ir cells co-xpressed TPH ir even though the cell number was onlyne third of that of ER-� ir cells. Representative photomi-rographs for co-localization of ER-� or ER-� with TPH arehown in Fig. 5.
ffects of ER-� or ER-� gene disruption on thexpression of TPH mRNA in the raphe nuclei
he distribution of TPH mRNA was in good agreement withhe immunocytochemical data. There were no significantenotype differences between �ERKO mice and their �WTiblings in the expression levels of TPH transcripts in allubdivisions of the DRN and the MPRN (Fig. 6A). In con-rast, the disruption of ER-� gene resulted in a small butignificant decrease (�17%) in the levels of TPH tran-cripts in the ventral subdivision of the DRN (P0.05; Fig.B). Further, there was a trend toward a decrease in TPHRNA in the dorsal subdivision of the DRN (P�0.08; Fig.B). However, in the lateral subdivision of the DRN and thePRN, no significant genotype differences were observed
n the expression levels of TPH transcripts (Fig. 6B). Rep-esentative autoradiographs showing the genotype differ-nces between �WT and �ERKO mice in the levels of TPH
ranscripts in the raphe nuclei are shown in Fig. 7A and B.hese genotype differences were also confirmed at theellular level by microscopic examination of emulsion-ipped sections (Fig. 7C, D).
DISCUSSION
he present study provides the first detailed report onhe distribution of ER-� and ER-� across the subdivi-ions of the DRN, the MPRN, and the PAG, in the intact
able 1. Co-localization of ER-� or ER-� with TPH in the DRN
Number of mice Dorsal
R-� n�4 24.81.3R-�/TPH 7.33.3 (29.5%)1
R-� n�4 41.93.2R-�/TPH 40.03.1 (95.5%)2**
percentage of ER-� and TPH ir cells in total number of ER-� ir cellspercentage of ER-� and TPH ir cells in total number of ER-� ir cells
* P0.01 vs. the percentage of ER-�/TPH in the same subdivision.
ale murine brain. We have found that ER-� is much d
ore abundant than ER-� in the raphe nuclei, while theR-� predominates in the PAG. Differential distributionf ER-� and ER-� in the DRN was particularly pro-ounced in serotonergic neurons, as we found that ER-�
r cells co-localized with TPH ir cells in the DRN to auch greater extent than ER-� ir cells. Thus ER-� is theredominant ER in 5-HT neurons in male mouse mid-rain raphe nuclei. Furthermore, the disruption of theR-� gene, but not of the ER-� gene, caused a small butignificant decrease in the TPH mRNA levels in theentral subdivisions of the DRN, in which ER-� expres-ion was most abundant. These results suggest thatR-� may be the primary ER subtype involved in directenomic regulation of serotonergic neurons. The fact
hat most ER-� ir cells do not co-express TPH suggestshat the ER-� may be the mediator by which estrogenegulates non-5-HT cells within the raphe nuclei. Thus,t the levels of the midbrain raphe nuclei and PAG, two
ypes of ERs may independently participate in the mod-lation of the neuroendocrine and behavioral functions,nd the activation of ER-� may play a crucial role in theirect regulation of the 5-HT system in the DRN.
istribution and co-localization of ER-� and ER-�ith TPH in the raphe nuclei
he present finding showing that a differential distributionf ER-� and ER-� ir cells in the raphe nuclei of gonadally
ntact male mice is consistent with the previous findingseported in male and female rats and female mice (Alvest al., 1998, 2000; Mitra et al., 2003). In the present study,e performed more detailed analyses of the subdivisionsf the DRN and found that ER-� ir cells were distributedredominantly in the ventral subdivisions of the DRN,hereas ER-� ir cells were distributed moderately in all
hree subdivisions of the DRN. Furthermore, we foundtriking differences between ER-� (on average, 23%) andR-� (more than 90%) cells in the extent of co-localizationith TPH ir in the DRN. These findings demonstrate thatR-� and ER-� are expressed, on the whole, in differentell types in the DRN.
One of the most well-characterized genomic effectsf E2 is the time-dependent induction of progesteroneeceptors (PR). In female macaque and mouse, E2-nduced PR ir is expressed primarily within serotonergicells of the DRN, although PR ir is seen in non-5-HTells as well (Alves et al., 2000; Bethea, 1993, 1994). In
Lateral Total
5 27.03.9 75.53.56 (55.3%)1 2.01.3 (6.2%)1 17.50.9 (23.2%)1
6 8.71.1 133.010.23 (97.3%)2** 8.11.1 (93.1%)2** 128.39.8 (96.4%)2**
Ventral
31.72.17.54.82.56.79.46.
.
.
ramatic contrast, E2-induced PR ir was identified only

wfEttitTwmmgt
Eee
EdsnfinEr(npntsidsdsemaEfAre
wtossd
Rs
TsvcTwDcmiMwIr5iNht
mtssaorahrasaap5ihsbrrtdgndio
Foibpp
M. Nomura et al. / Neuroscience 130 (2005) 445–456452
ithin cells adjacent to, but not in, serotonergic cells inemale rats (Alves et al., 1998). Furthermore, very fewR-� ir containing cells were co-localized with TPH ir in
he rat DRN (Alves et al., 1998), which is in contrast tohe present finding in mice showing that ER-� co-local-zed with a subpopulation of TPH ir cells in the DRN (6o approximately 55% depending on the subdivisions).hese differences in the distribution of PR and ER-�ithin serotonergic or non-serotonergic neurons amongacaque, mouse, and rat lead to the notion that thereay be species differences in the mechanisms by whichonadal steroids modulate the serotonergic system inhe DRN.
ffects of ER-� gene disruption on the ER-�xpression and ER-� gene disruption on the ER-�xpression in the raphe nuclei
xamining �ERKO and �ERKO mice, we found thatisruption of either ER subtype did not alter the expres-ion or distribution of the other ER in the midbrain rapheuclei or the PAG. This is in contrast to our previousndings in the medial preoptic area (MPOA) and the beducleus of stria terminalis (BNST), in which the levels ofR-� gene expression (mRNA and protein) were greatly
educed or increased, respectively, by the lack of ER-�Nomura et al., 2003). The present finding in the rapheuclei was similar to that in the medial amygdala, thearaventricular nucleus (PVN), and the ventromedialucleus of the hypothalamus, in which the disruption ofhe ER-� gene did not affect the levels of ER-� expres-ion (Nomura et al., 2003). It is likely that the differencesn the levels of co-expression of ER-a and ER-b in theifferent brain regions may be responsible for this site-pecific regulation of ER-b expression by ER-a geneisruption. Previously, Shughrue et al. (1998) demon-trated that the majority of ER-� mRNA expressing cellsxpress ER-� in the MPOA and the BNST of ovariecto-ized female rats. Recently, Greco et al. (2001) havelso shown that the majority of ER-� ir cells co-expressR-� ir in the MPOA and the BNST of ovariectomized
emale rats treated with estradiol benzoate or vehicle.lthough co-localization of ER-� and ER-� in the DRN
emains to be determined, it is expected to be less
ig. 5. Representative photomicrographs showing the co-localizationf ER-� (A) or ER-� (B) with TPH in the dorsal subdivision of the DRN
n �WT mouse brains. Black arrows indicate the DRN neurons ir foroth TPH (light brown cytoplasmic stain) and ER-� or ER-� (darkurple punctate stain). White arrows indicate the DRN neurons ex-ressing ER-� but not TPH. The scale bar�50 �m.
xtensive, since most of ER-�, but only some ER-� ir g
as detected in the serotonergic cells in the DRN. Fur-hermore, the exact molecular mechanisms (expressionf co-regulators, etc.) by which gene deletion of one ERubtype can affect the expression of the remaining ERubtype in a region specific manner also need to beetermined.
oles of ERs in the regulation of serotonergicystems
he disruption of the ER-� gene resulted in a small butignificant decrease in the level of TPH mRNA in theentral subdivision of the DRN compared with WT mice. Inontrast, disruption of the ER-� gene did not affect thePH mRNA level in any parts of the raphe nuclei. Indeed,e found that in the dorsal and ventral subdivisions of theRN, the number of ER-� ir cells was twice that of ER-� irells. Further, ER-� ir cells co-expressed TPH ir with auch greater frequency (more than 95%) than ER-� pos-
tive cells did. In the lateral subdivision of the DRN and thePRN, where a smaller number of ER-� ir cells were seen,e failed to detect any changes in the level of TPH mRNA.
t still remains to be determined whether a relatively smalleduction in TPH may translate to a similar reduction in-HT synthesis, which needs to be examined by measur-
ng the levels of 5-HT and its metabolites in the DRN.evertheless, the findings in the present study lead us toypothesize that activation of ER-� may be responsible for
he regulation of 5-HT synthesis in the DRN.Since no apparent effects of ER-� disruption on TPH
RNA levels were found in the present study, it is unlikelyhat ER-� plays a crucial role in the regulation of the tran-cripts for TPH in the raphe nuclei in male mice. However,ubstantial amounts of ER-� ir were still found in TPH neg-tive cells in the DRN and MPRN. Therefore, we cannot ruleut the possibility that ER-� may still be involved in theegulation of the serotonergic system through transsynapticction at the level of the raphe nuclei. In addition to TPH, itas also been shown in the rat and macaque DRN that E2
egulates the expression of other 5-HT related genes, suchs 5-HT1A and 5-HT2A receptors and SERT, which are re-ponsible for the overall function of the 5-HT system (Fink etl., 1999; McQueen et al., 1997, 1999; Pecins-Thompsonnd Bethea, 1999; Pecins-Thompson et al., 1998). Sinceresynaptic autoreceptor, 5-HT1A, and SERT are found in-HT neurons, it is hypothesized that ER-� plays a major role
n the E2 regulation of their gene expression. On the otherand, it is possible that the estrogenic up-regulation of post-ynaptic 5-HT2A receptor gene expression may be mediatedy ER-� localized in non-5-HT neurons in the DR. Specificoles of ER-� and ER-� in the regulation of these genes in theaphe nuclei of male mice need to be further clarified. Fur-hermore, it needs to be determined whether there are sex-ifferences in the effects of ER gene disruption on the estro-enic regulation of the serotonergic system in the rapheuclei, since there is accumulating evidence of sex-ifferences in serotonergic systems (Heninger, 1997). Finally,
t is also necessary to compare the effects of E2 replacementn the levels of TPH and other 5-HT related genes between
onadectomized WT and �ERKO mice to show more con-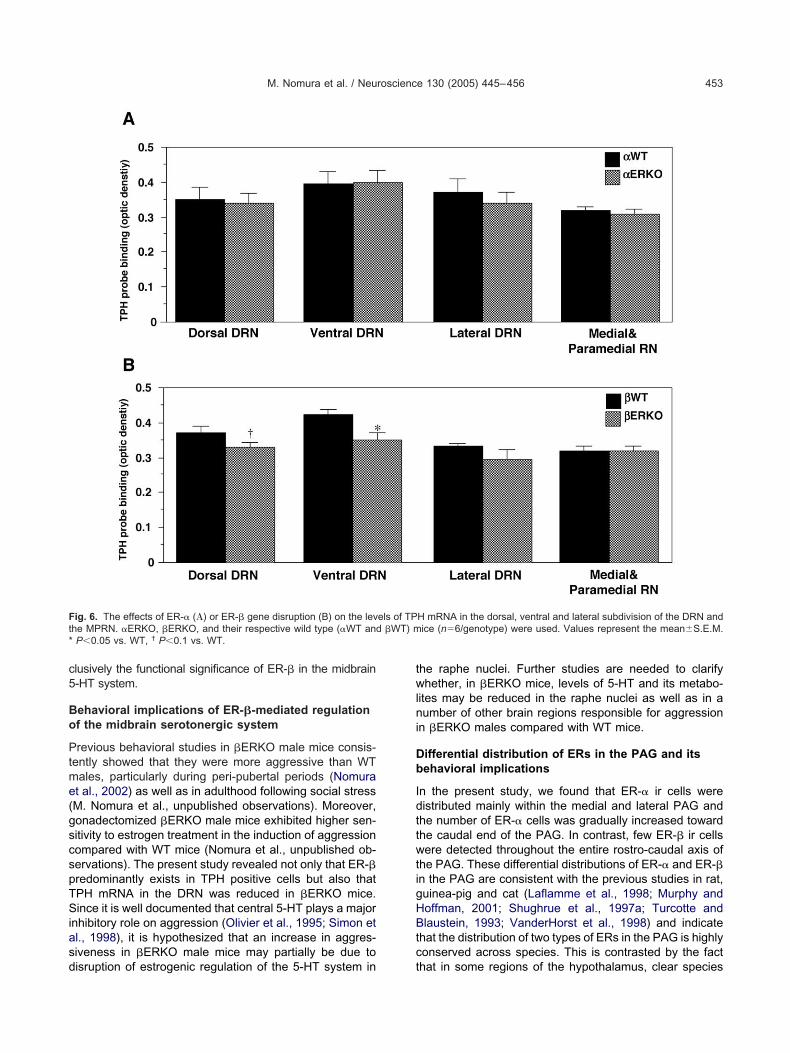
c5
Bo
Ptme(gscspTSiasd
twlni
Db
IdttwtigHBtc
Ft �WT) m*
M. Nomura et al. / Neuroscience 130 (2005) 445–456 453
lusively the functional significance of ER-� in the midbrain-HT system.
ehavioral implications of ER-�-mediated regulationf the midbrain serotonergic system
revious behavioral studies in �ERKO male mice consis-ently showed that they were more aggressive than WTales, particularly during peri-pubertal periods (Nomurat al., 2002) as well as in adulthood following social stressM. Nomura et al., unpublished observations). Moreover,onadectomized �ERKO male mice exhibited higher sen-itivity to estrogen treatment in the induction of aggressionompared with WT mice (Nomura et al., unpublished ob-ervations). The present study revealed not only that ER-�redominantly exists in TPH positive cells but also thatPH mRNA in the DRN was reduced in �ERKO mice.ince it is well documented that central 5-HT plays a major
nhibitory role on aggression (Olivier et al., 1995; Simon etl., 1998), it is hypothesized that an increase in aggres-iveness in �ERKO male mice may partially be due to
ig. 6. The effects of ER-� (�) or ER-� gene disruption (B) on the levhe MPRN. �ERKO, �ERKO, and their respective wild type (�WT andP0.05 vs. WT, † P0.1 vs. WT.
isruption of estrogenic regulation of the 5-HT system in t
he raphe nuclei. Further studies are needed to clarifyhether, in �ERKO mice, levels of 5-HT and its metabo-
ites may be reduced in the raphe nuclei as well as in aumber of other brain regions responsible for aggression
n �ERKO males compared with WT mice.
ifferential distribution of ERs in the PAG and itsehavioral implications
n the present study, we found that ER-� ir cells wereistributed mainly within the medial and lateral PAG andhe number of ER-� cells was gradually increased towardhe caudal end of the PAG. In contrast, few ER-� ir cellsere detected throughout the entire rostro-caudal axis of
he PAG. These differential distributions of ER-� and ER-�n the PAG are consistent with the previous studies in rat,uinea-pig and cat (Laflamme et al., 1998; Murphy andoffman, 2001; Shughrue et al., 1997a; Turcotte andlaustein, 1993; VanderHorst et al., 1998) and indicate
hat the distribution of two types of ERs in the PAG is highlyonserved across species. This is contrasted by the fact
H mRNA in the dorsal, ventral and lateral subdivision of the DRN andice (n�6/genotype) were used. Values represent the meanS.E.M.
els of TP
hat in some regions of the hypothalamus, clear species

dratPtepeiS
f(islPrrsniterfietrer
PEi
AMaCatsrJp
A
A
B
B
F
F
F
G
G
G
H
I
J
K
K
L
FtdaDi
M. Nomura et al. / Neuroscience 130 (2005) 445–456454
ifferences in the distribution of ER-� and ER-� have beeneported among a number of animals such as rats, micend guinea-pigs. For example, ER-� protein is present inhe PVN and the SON of guinea-pigs (Warembourg andoulain, 1991) but ER-� is absent or barely expressed in
hese nuclei of rats and mice (Greco et al., 2001; Laflammet al., 1998). ER-� protein and mRNA are abundantlyresent in the rat SON (Laflamme et al., 1998; Shughruet al., 1997a; Shughrue and Merchenthaler, 2001), but not
n the mouse SON (Mitra et al., 2003; Nomura et al., 2003;hughrue et al., 1997b).
The involvement of the PAG in E2-dependent induction ofemale reproductive behaviors is well establishedSakuma and Pfaff, 1979a,b). Although it has been much lessnvestigated, PAG is also implicated in the regulation of maleexual behavior. In male rats, it has been shown that ejacu-ation induces c-fos protein expression in a large number ofAG neurons, which generally overlapped with previously
eported distribution of ER-� (Murphy et al., 1999). In a moreecent study, Murphy and Hoffman (2001) have demon-trated that PAG serves as a relay site in the pathway con-ecting the MPOA and nucleus paragigantocellularis (nPGi)
n the brainstem, which is crucial for the initiation and main-enance of male sexual behavior, particularly erection andjaculation. They also showed that half of the PAG neuronsetrogradely labeled from the nPGi contain ER-� ir. Thesendings suggest that ER-� activation may play a major role instrogenic regulation of male sexual behavior at the level ofhe PAG. Interestingly, it has been shown that �ERKO malesarely showed ejaculation in sexual behavioral tests (Ogawat al., 1997, 1998, 2000; Wersinger et al., 1997). These
ig. 7. Representative film autoradiographs (A, B) and dark-field pho-omicrographs of emulsion dipped slides (C, D) showing the genotypeifferences in the level of TPH mRNA in the DRN between �WT (A, C)nd �ERKO (B, D) mice. Both ventral and dorsal subdivisions of theRN are shown in A and B. In C and D, the ventral division of the DRN
s shown. The scale bars�1 mm (A, B) or 100 �m (C, D).
esults raise the possibility that the expression of ER-� in the
AG may be critical for ejaculation, although the exact role ofR-� in male reproductive behaviors at the level of the PAG
s still largely unknown.
cknowledgments—The authors are thankful to Dr. V. Giguere atcGill University for his kind gift of ER-� cDNA and Dr. H. Tanakat the University of Kumamoto and Dr. S. Hayashi at Yokohamaity University for their kind gift of the ER-� probes. The authorslso thank Dr. E. G. Mirasol, Ms I. Dorfman, and Ms L. Durbak forheir technical assistance. Support and encouragement for thistudy by Dr. B. S. McEwen is greatly appreciated. MN was aecipient of postdoctoral fellowships for research abroad from theapan Society for Promotion of Science. Support for this work wasrovided by NIMH 62147 to S.O.
REFERENCES
lves SE, McEwen BS, Hayashi S, Korach KS, Pfaff DW, Ogawa S(2000) Estrogen-regulated progestin receptors are found in themidbrain raphe but not hippocampus of estrogen receptor alpha(ER alpha) gene-disrupted mice. J Comp Neurol 427:185–195.
lves SE, Weiland NG, Hayashi S, McEwen BS (1998) Immunocyto-chemical localization of nuclear estrogen receptors and progestinreceptors within the rat dorsal raphe nucleus. J Comp Neurol391:322–334.
ethea CL (1993) Colocalization of progestin receptors with serotoninin raphe neurons of macaque. Neuroendocrinology 57:1–6.
ethea CL (1994) Regulation of progestin receptors in raphe neuronsof steroid-treated monkeys. Neuroendocrinology 60:50–61.
ile SE, Deakin JF (1980) Chemical lesions of both dorsal and medianraphe nuclei and changes in social and aggressive behaviour inrats. Pharmacol Biochem Behav 12:855–859.
ink G, Sumner B, Rosie R, Wilson H, McQueen J (1999) Androgenactions on central serotonin neurotransmission: relevance formood, mental state and memory. Behav Brain Res 105:53–68.
ranklin KBJ, Paxinos G (1997) The mouse brain in stereotaxic coor-dinates. San Diego: Academic Press.
reco B, Allegretto EA, Tetel MJ, Blaustein JD (2001) Coexpression ofER beta with ER alpha and progestin receptor proteins in thefemale rat forebrain: effects of estradiol treatment. Endocrinology142:5172–5181.
reen S, Walter P, Greene G, Krust A, Goffin C, Jensen E, Scrace G,Waterfield M, Chambon P (1986) Cloning of the human oestrogenreceptor cDNA. J Steroid Biochem 24:77–83.
undlah C, Lu NZ, Mirkes SJ, Bethea CL (2001) Estrogen receptorbeta (ERbeta) mRNA and protein in serotonin neurons of ma-caques. Mol Brain Res 91:14–22.
eninger GR (1997) Serotonin, sex, and psychiatric illness. Proc NatlAcad Sci USA 94:4823–4824.
mwalle DB, Scordalakes EM, Rissman EF (2002) Estrogen receptoralpha influences socially motivated behaviors. Horm Behav42:484–491.
anusonis S, Fite KV, Foote W (1999) Topographic organization ofserotonergic dorsal raphe neurons projecting to the superior col-liculus in the Mongolian gerbil (Meriones unguiculatus). J CompNeurol 413:342–355.
rege JH, Hodgin JB, Couse JF, Enmark E, Warner M, Mahler JF,Sar M, Korach KS, Gustafsson JÅ, Smithies O (1998) Generationand reproductive phenotypes of mice lacking estrogen receptor �.Proc Natl Acad Sci USA 95:15677–15682.
uiper GGJM, Enmark E, Peltohuikko M, Nilsson S (1996) Cloning ofa novel estrogen receptor expressed in rat prostate and ovary.Proc Natl Acad Sci USA 93:5925–5930.
aflamme N, Nappi RE, Drolet G, Labrie C, Rivest S (1998) Expres-sion and neuropeptidergic characterization of estrogen receptors(ERalpha and ERbeta) throughout the rat brain: anatomical evi-
dence of distinct roles of each subtype. J Neurobiol 36:357–378.
L
L
L
M
M
M
M
M
M
N
N
O
O
O
O
O
O
O
P
P
P
R
S
S
S
S
S
S
S
S
S
T
T
T
U
V
V
W
W
W
M. Nomura et al. / Neuroscience 130 (2005) 445–456 455
u H, Ozawa H, Nishi M, Ito T, Kawata M (2001) Serotonergic neu-rones in the dorsal raphe nucleus that project into the medialpreoptic area contain oestrogen receptor beta. J Neuroendocrinol13:839–845.
u NZ, Shlaes TA, Gundlah C, Dziennis SE, Lyle RE, Bethea CL(1999) Ovarian steroid action on tryptophan hydroxylase proteinand serotonin compared to localization of ovarian steroid receptorsin midbrain of guinea pigs. Endocrine 11:257–267.
ubahn DB, Moyer JS, Golding TS, Couse JF, Korach KS, Smithies O(1993) Alteration of reproductive function but not prenatal sexualdevelopment after insertional disruption of the mouse estrogenreceptor gene. Proc Natl Acad Sci USA 90:11162–11166.
cQueen JK, Wilson H, Fink G (1997) Estradiol-17 beta increasesserotonin transporter (SERT) mRNA levels and the density ofSERT-binding sites in female rat brain. Mol Brain Res 45:13–23.
cQueen JK, Wilson H, Sumner BE, Fink G (1999) Serotonin trans-porter (SERT) mRNA and binding site densities in male rat brainaffected by sex steroids. Mol Brain Res 63:241–247.
eisel RL, Sachs BD (1994) The physiology of male sexual behavior.In: Physiology of peproduction, Vol. 1 and 2 (Knobil E, Neill JD,eds), pp 3–105. New York: Raven Press.
itra SW, Hoskin E, Yudkovitz J, Pear L, Wilkinson HA, Hayashi S,Pfaff DW, Ogawa S, Rohrer SP, Schaeffer JM, McEwen BS,Alves SE (2003) Immunolocalization of estrogen receptor beta inthe mouse brain: comparison with estrogen receptor alpha. Endo-crinology 144:2055–2067.
urphy AZ, Hoffman GE (2001) Distribution of gonadal steroid recep-tor-containing neurons in the preoptic-periaqueductal gray-brainstem pathway: a potential circuit for the initiation of malesexual behavior. J Comp Neurol 438:191–212.
urphy AZ, Shupnik MA, Hoffman GE (1999) Androgen and estrogen(alpha) receptor distribution in the periaqueductal gray of the malerat. Horm Behav 36:98–108.
omura M, Durbak L, Chan J, Smithies O, Gustafsson JÅ, Korach KS,Pfaff DW, Ogawa S (2002) Genotype/age interactions on aggres-sive behavior in gonadally intact estrogen receptor � knockout(�ERKO) male mice. Horm Behav 41:288–296.
omura M, Korach KS, Pfaff DW, Ogawa S (2003) Estrogen receptor� (ER�) protein levels in neurons depend on estrogen receptor �(ER�) gene expression and on its ligand in a brain region-specificmanner. Mol Brain Res 110:7–14.
gawa S, Chan J, Chester AE, Gustafsson JÅ, Korach KS, Pfaff DW(1999) Survival of reproductive behaviors in estrogen receptor betagene-deficient (�ERKO) male and female mice. Proc Natl Acad SciUSA 96:12887–12892.
gawa S, Chester AE, Hewitt SC, Walker VR, Gustafsson JÅ,Smithies O, Korach KS, Pfaff DW (2000) From the cover: abolitionof male sexual behaviors in mice lacking estrogen receptors � and� (��ERKO). Proc Natl Acad Sci USA 97:14737–14741.
gawa S, Korach KS, Pfaff DW (2002) Differential roles of two typesof estrogen receptors in reproductive behavior. Cuur Opin Endo-crinol Diabetes 9:224–229.
gawa S, Lubahn DB, Korach KS, Pfaff DW (1997) Behavioral effectsof estrogen receptor gene disruption in male mice. Proc Natl AcadSci USA 94:1476–1481.
gawa S, Washburn TF, Taylor J, Lubahn DB, Korach KS, Pfaff DW(1998) Modifications of testosterone-dependent behaviors by es-trogen receptor-alpha gene disruption in male mice. Endocrinology139:5058–5069.
livier B, Mos J, van Oorschot R, Hen R (1995) Serotonin receptorsand animal models of aggressive behavior. Pharmacopsychiatry28(Suppl 2):80–90.
lsen KL (1992) Genetic influences on sexual behavior differentiation.In: Sexual differentiation (Arnold GA, Howard M, Ingeborg WL,eds), pp 1–40. New York: Plenum Press.
ecins-Thompson M, Bethea CL (1999) Ovarian steroid regulation ofserotonin-1A autoreceptor messenger RNA expression in the dor-
sal raphe of rhesus macaques. Neuroscience 89:267–277.ecins-Thompson M, Brown NA, Bethea CL (1998) Regulation ofserotonin re-uptake transporter mRNA expression by ovarian ste-roids in rhesus macaques. Brain Res Mol Brain Res 53:120–129.
ecins-Thompson M, Brown NA, Kohama SG, Bethea CL (1996)Ovarian steroid regulation of tryptophan hydroxylase mRNA ex-pression in rhesus macaques. J Neurosci 16:7021–7029.
issman EF, Wersinger SR, Fugger HN, Foster TC (1999) Sex withknockout models: behavioral studies of estrogen receptor alpha.Brain Res 835:80–90.
akuma Y, Pfaff DW (1979a) Facilitation of female reproductive be-havior from mesencephalic central gray in the rat. Am J Physiol237:R278–R284.
akuma Y, Pfaff DW (1979b) Mesencephalic mechanisms for integra-tion of female reproductive behavior in the rat. Am J Physiol237:R285–R290.
cordalakes EM, Rissman EF (2003) Aggression in male mice lackingfunctional estrogen receptor alpha. Behav Neurosci 117:38–45.
hughrue PJ, Lane MV, Merchenthaler I (1997a) Comparative distri-bution of estrogen receptor-alpha and -beta mRNA in the ratcentral nervous system. J Comp Neurol 388:507–525.
hughrue PJ, Scrimo P, Lane M, Askew R, Merchenthaler I (1997b)The distribution of estrogen receptor-beta mRNA in forebrain re-gions of the estrogen receptor-alpha knockout mouse. Endocrinol-ogy 138:5649–5652.
hughrue PJ, Merchenthaler I (2001) Distribution of estrogen receptorbeta immunoreactivity in the rat central nervous system. J CompNeurol 436:64–81.
hughrue PJ, Scrimo PJ, Merchenthaler I (1998) Evidence for thecolocalization of estrogen receptor-beta mRNA and estrogenreceptor-alpha immunoreactivity in neurons of the rat forebrain.Endocrinology 139:5267–5270.
imon NG, Cologer-Clifford A, Lu SF, McKenna SE, Hu S (1998)Testosterone and its metabolites modulate 5HT1A and 5HT1Bagonist effects on intermale aggression. Neurosci Biobehav Rev23:325–336.
imon NG, Whalen RE (1987) Sexual differentiation of androgen-sensitive and estrogen-sensitive regulatory systems for aggressivebehavior. Horm Behav 21:493–500.
emple JL, Fugger HN, Li X, Shetty SJ, Gustafsson J, Rissman EF(2001) Estrogen receptor beta regulates sexually dimorphic neuralresponses to estradiol. Endocrinology 142:510–513.
remblay GB, Tremblay A, Copeland NG, Gilbert DJ, Jenkins NA,Labrie F, Giguere V (1997) Cloning, chromosomal localization, andfunctional analysis of the murine estrogen receptor �. Mol Endocrin11:353–365.
urcotte JC, Blaustein JD (1993) Immunocytochemical localization ofmidbrain estrogen receptor-containing and progestin receptor-containing cells in female guinea pigs. J Comp Neurol 328:76–87.
eta Y, Levy A, Chowdrey HS, Lightman SL (1995) S-100 antigen-positive folliculostellate cells are not the source of IL-6 gene expres-sion in human pituitary adenomas. J Neuroendocrinol 7:467–474.
anderHorst VG, Schasfoort FC, Meijer E, van Leeuwen FW,Holstege G (1998) Estrogen receptor-alpha-immunoreactive neu-rons in the periaqueductal gray of the adult ovariectomized femalecat. Neurosci Lett 240:13–16.
ergnes M, Depaulis A, Boehrer A, Kempf E (1988) Selective increaseof offensive behavior in the rat following intrahypothalamic 5,7-DHT-induced serotonin depletion. Behav Brain Res 29:85–91.
arembourg M, Poulain P (1991) Presence of estrogen receptor immuno-reactivity in the oxytocin-containing magnocellular neurons projecting tothe neurohypophysis in the guinea-pig. Neuroscience 40:41–53.
ersinger SR, Rissman EF (2000) Dopamine activates masculinesexual behavior independent of the estrogen receptor alpha.J Neurosci 20:4248–4254.
ersinger SR, Sannen K, Villalba C, Lubahn DB, Rissman EF, De
Vries GJ (1997) Masculine sexual behavior is disrupted in
Y
M. Nomura et al. / Neuroscience 130 (2005) 445–456456
male and female mice lacking a functional estrogen receptor
alpha gene. Horm Behav 32:176 –183.amamoto T, Ueki S (1977) Characteristics in aggressive behavior in-
duced by midbrain raphe lesions in rats. Physiol Behav 19:105–110.(Accepted 17 September 2004)(Available online 11 November 2004)




















