
Lindane may modulate the female reproductive development through the interaction with ER-β: an in...
14
Chemico-Biological Interactions 169 (2007) 1–14 Lindane may modulate the female reproductive development through the interaction with ER-: an in vivo–in vitro approach Francesca Maranghi a,∗ , Michele Rescia a , Caterina Macr` ı a , Emma Di Consiglio b , Giovanna De Angelis b , Emanuela Testai b , Donatella Farini c , Massimo De Felici c , Stefano Lorenzetti a , Alberto Mantovani a a Department of Food Safety and Veterinary Public Health, Istituto Superiore di Sanit` a, Viale Regina Elena 299, 00161 Rome, Italy b Department of Environment and Primary Prevention, Istituto Superiore di Sanit` a, Viale Regina Elena 299, 00161 Rome, Italy c Department of Public Health and Cell Biology, University of Roma Tor Vergata, Via Montpellier 1, 00133 Rome, Italy Received 11 December 2006; received in revised form 16 April 2007; accepted 18 April 2007 Available online 22 April 2007 Abstract Lindane (-HCH) is a persistent environmental pollutant that may act as endocrine disrupter, affecting the nervous, immune and reproductive system, possibly through endocrine-mediated mechanisms. Since both estrogen receptors (ER- and -) have shown to be target for endocrine disruption, we investigated the role of -HCH on the development of female reproductive system. For an in vivo evaluation of -HCH effects during prenatal period, pregnant CD1 mice were treated p.o. on gestational days 9–16 with 15 mg/kg bw/day of -HCH and vehicle. The in vivo findings in treated F1 pups – in the absence of signs of systemic toxicity – included increase in the absolute and relative and absolute uterus weight revealed on post-natal day 22, earlier vaginal patency and reduced diameters of primary oocytes at fully sexual maturity. No effects on steroid hormone metabolism (aromatase, testosterone catabolism) were observed. Thus, -HCH elicited subtle effects on female reproductive development likely mediated by ER--mediated pathway(s), without a concurrent impairment of steroid hormone metabolism. Furthermore, to verify whether the endocrine interference of -HCH is attributable to stimulation of ER--mediated pathway(s), its effect has been evaluated in vitro on a cell line, LNCaP, expressing only functional ER-. In vitro treatments revealed a concentration- related effect on LNCaP cell viability and proliferation. Significantly, the contemporary addition of a pure anti-estrogen, the ER antagonist ICI 182,780, completely reversed -HCH effects indicating an ER--mediated action. Our findings indicate that -HCH may act as endocrine disruptor during the female reproductive system development and ER- as a potential target for this compound and other endocrine disrupting chemicals as well. © 2007 Elsevier Ireland Ltd. All rights reserved. Keywords: Lindane; Prenatal exposure; Reproduction; ER- ∗ Corresponding author. Tel.: +39 6 49902527; fax: +39 6 49902658. E-mail address: [email protected] (F. Maranghi). 1. Introduction In the last decades, attention has been focused on reproductive effects in terms of fertility, pregnancy outcomes, long-term developmental effects [1] of the persistent organic pollutants (POPs) both in wildlife and humans. Widespread in food chain and in the 0009-2797/$ – see front matter © 2007 Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.cbi.2007.04.008
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Lindane may modulate the female reproductive development through the interaction with ER-β: an in...

A
rta1
asei
era
a©
K
0
Chemico-Biological Interactions 169 (2007) 1–14
Lindane may modulate the female reproductive developmentthrough the interaction with ER-�: an in vivo–in vitro approach
Francesca Maranghi a,∗, Michele Rescia a, Caterina Macrı a, Emma Di Consiglio b,Giovanna De Angelis b, Emanuela Testai b, Donatella Farini c, Massimo De Felici c,
Stefano Lorenzetti a, Alberto Mantovani a
a Department of Food Safety and Veterinary Public Health, Istituto Superiore di Sanita, Viale Regina Elena 299, 00161 Rome, Italyb Department of Environment and Primary Prevention, Istituto Superiore di Sanita, Viale Regina Elena 299, 00161 Rome, Italy
c Department of Public Health and Cell Biology, University of Roma Tor Vergata, Via Montpellier 1, 00133 Rome, Italy
Received 11 December 2006; received in revised form 16 April 2007; accepted 18 April 2007Available online 22 April 2007
bstract
Lindane (�-HCH) is a persistent environmental pollutant that may act as endocrine disrupter, affecting the nervous, immune andeproductive system, possibly through endocrine-mediated mechanisms. Since both estrogen receptors (ER-� and -�) have showno be target for endocrine disruption, we investigated the role of �-HCH on the development of female reproductive system. Forn in vivo evaluation of �-HCH effects during prenatal period, pregnant CD1 mice were treated p.o. on gestational days 9–16 with5 mg/kg bw/day of �-HCH and vehicle.
The in vivo findings in treated F1 pups – in the absence of signs of systemic toxicity – included increase in the absolute and relativend absolute uterus weight revealed on post-natal day 22, earlier vaginal patency and reduced diameters of primary oocytes at fullyexual maturity. No effects on steroid hormone metabolism (aromatase, testosterone catabolism) were observed. Thus, �-HCHlicited subtle effects on female reproductive development likely mediated by ER-�-mediated pathway(s), without a concurrentmpairment of steroid hormone metabolism.
Furthermore, to verify whether the endocrine interference of �-HCH is attributable to stimulation of ER-�-mediated pathway(s), itsffect has been evaluated in vitro on a cell line, LNCaP, expressing only functional ER-�. In vitro treatments revealed a concentration-
elated effect on LNCaP cell viability and proliferation. Significantly, the contemporary addition of a pure anti-estrogen, the ERntagonist ICI 182,780, completely reversed �-HCH effects indicating an ER-�-mediated action.Our findings indicate that �-HCH may act as endocrine disruptor during the female reproductive system development and ER-�s a potential target for this compound and other endocrine disrupting chemicals as well.
2007 Elsevier Ireland Ltd. All rights reserved.
eywords: Lindane; Prenatal exposure; Reproduction; ER-�
∗ Corresponding author. Tel.: +39 6 49902527; fax: +39 6 49902658.E-mail address: [email protected] (F. Maranghi).
009-2797/$ – see front matter © 2007 Elsevier Ireland Ltd. All rights reservdoi:10.1016/j.cbi.2007.04.008
1. Introduction
In the last decades, attention has been focused on
reproductive effects in terms of fertility, pregnancyoutcomes, long-term developmental effects [1] of thepersistent organic pollutants (POPs) both in wildlifeand humans. Widespread in food chain and in theed.

iologica
2 F. Maranghi et al. / Chemico-Benvironment, POPs may act as Endocrine Disrupters(EDs), acting through different mechanisms. POPsinclude several chlorinated insecticides, such as DDT,methoxychlor, as well as Lindane, the � isomer of hex-achlorocyclohexane (�-HCH). �-HCH has been largelyused as insecticide and disinfectant in agriculture since1950 and also entered in the composition of some lotions,creams and shampoos used against parasites. Althoughrestrictions for �-HCH use have been introduced in manyEuropean countries and in USA (approximately start-ing from 1975), �-HCH is still used in the rest of theworld; human exposure may represents still a concernbecause of the persistence in the environment and bioac-cumulation in the feed and food chains [2]. �-HCH isa recognized reproductive and developmental toxicantwith a pattern of effects suggesting an endocrine-relatedmechanism [3,4]. �-HCH interacts with several tar-gets of the reproductive system in different stages ofthe life cycle. In the early embryo, �-HCH may inter-fere with cell-to-cell communication [5]. In male miceexposed in utero, �-HCH impairs spermatogenesis inthe absence of evident morphological abnormalities ofthe reproductive tract [3]. Exposure of adult femalerodents impairs the oestrous cycle and reduces serumestrogen and progesterone levels, sexual receptivity andlitter size [4]. In a recent two-generation study, �-HCHinduced different systemic effects, whereas the evidencefor endocrine-mediated alterations was equivocal [6].Indeed, mechanisms of reproductive effects have yetto be clarified. �-HCH does not appear to be a strongagonist or antagonist of either ER-� or AR, nor is adioxin-like agonist of the aryl hydrocarbon receptor(AhR), although a competitive binding with proges-terone receptor has been recently reported [7]. So far,limited and conflicting data exist on �-HCH/ER-� inter-action since only data based on gene reporter assayshave been published. In particular, Balaguer et al. [8]characterized the estrogenic activity of different con-taminants, including �-HCH, using stably transfectedMCF-7 and HeLa cells expressing ER-� and/or ER-�together with an ER-regulated reporter gene. In thesesystems, �-HCH did not show estrogenic activity. Lee etal. [9] performed a yeast two-hybrid assay to assess theestrogenic activity of different EDs, using the interactionbetween the human ER-� ligand-binding domain and theco-activator SRC1 nuclear receptor-binding domain asthe most effective to detect a xenoestrogen-dependentinduction of the reporter activity. In these yeast system,
�-HCH showed a detectable estrogenic activity.Overall, limited data are available on the interactionof EDs with ER-�, apart from natural plant compoundsknown as phytoestrogens. For instance, genistein, the
l Interactions 169 (2007) 1–14
main soy isoflavone, is known to interact with both ER-�and -�, with a higher affinity for ER-� [10] and tissue-specific effects [11]. Interestingly, genistein inducesreproductive effects in developing male mice with a sim-ilar pattern as �-HCH [12]. ER-� shows major biologicalroles in different tissues which have not yet fully clari-fied [13]. In both males and females, 17�-estradiol (E2)is produced by the aromatisation of dihydrotestosterone(DHT) by the cytochrome P450 aromatase enzyme com-plex. Estrogens exert their action not only regulating thefemale reproductive tract but also male gonads and, inboth sexes, the skeleton, the central nervous system aswell as the cardiovascular and immune systems [14].The basis of such a pleiotropic role relies on the broadtissue-specific expression of both ERs. ER-� and -� arenuclear hormone receptors, ligand-activated transcrip-tion factors, able to regulate gene expression in a cell-and promoter-specific way. In females, ERs are the mainregulatory factors for reproductive function, albeit withcomplementary roles. For instance, in the ovarian fol-licles ER-� mediates proliferative effects, while ER-�mediates differentiation [15]. ER-� is expressed at highlevel in tissues such as ovary, prostate, testis, salivaryglands, vascular endothelium, smooth muscle and in theimmune and nervous systems as well [14]. Interestingly,tissues with highest levels of ER-� are the granulosacells in the ovary [16] and the epithelium of the ventralprostate [17], where little or no expression of ER-� canbe detected [18]. In particular, ER-� role is critical togranulosa cell differentiation and organization of ovar-ian follicles [16]. It is noteworthy that at least in thesetissues, the main ER-� ligand is 5�-androstane-3�,17�-diol (3�Adiol) [19], the metabolite of 5�-DHT. Thus thepossible cross-talk between ER-� and AR may deservefurther attention, although conflicting findings exist [20].
In female mouse, from gestational day 9 to 16, crucialsteps for gonadal differentiation occur: establishmentand migration of primordial germ cells, colonization ofundifferentiated gonads, sexual differentiation, appear-ance of hormone receptors which all may representtargets for EDs. Furthermore, during the post-natalperiod, the development and maturation of such struc-tures proceed until the completion of sexual maturity,i.e. PND 60 in mouse [21]. Since no adequate data existon the effect on developing female reproductive systeminduced by xenobiotics through ER-� interference, inthis study were evaluated long-term effects of �-HCH onthe development of female reproductive system follow-
ing the exposure during a critical period of organogenesisin CD1 female mice followed up to sexual maturation.A dose level of 15 mg/kg bw was selected since itinduces initial reproductive effects in in utero exposed

ologica
mo3iteaHhpt[
aiftfP�pip
2
2
c
pwDFc11M7(Cw(c
cbbwP(
F. Maranghi et al. / Chemico-Bi
ale mice without signs of systemic toxicity [3]. More-ver, the selected dose level is comparable to the00 mg/kg food exposure level (corresponding to anntake of approximately 17 mg/kg bw during pregnancy)hat induced general toxicity and altered hormone lev-ls in the two-generation rat study by Matsuura etl. [6]. In order to verify the possible impact of �-CH effects on the key enzymes responsible for steroidormone metabolism, we additionally investigated theossible modulation of hepatic testosterone (TST) inac-ivation and aromatase activity in dams and F1 females22].
Concurrently, to test �-HCH endocrine interference insuitable in vitro model, we choose a cell line express-
ng only a functional ER-�. The LNCaP cell line, derivedrom human prostate adenocarcinoma, expresses a func-ional ER-� as well as AR and PRs, but it is devoid ofunctional ER-� [23–25], as confirmed also by our RT-CR data. We investigated the ER-�-mediated effect of-HCH on LNCaP cell viability/proliferation using theure anti-estrogen ICI 182,780 as a reliable tool to specif-cally distinguish �HCH-ER-� interaction from otherossible receptor-mediated pathways.
. Materials and methods
.1. Chemicals
�HCH (CAS n.58-89-9, technical grade), was pur-hased from Sigma-Aldrich (St. Louis, MO, USA).
For biochemical analysis, NADP, glucose-6-hosphate (G6P), and G6P-dehydrogenase (G6PDH)ere supplied by Roche GmBh (Mannheim, Germany).efatted bovine serum albumin (BSA) was from Serva,einbiochemica (Heidelberg, Germany). Testosterone,orticosterone, 4-androsten-3,17-dione (�4), and 2�-,6�-, 6�-, 16�-hydroxy-testosterone (2�, 16�, 6�,6�) were purchased from Sigma-Aldrich (St. Louis,O, USA). 2�-, 6�-, 7�-hydroxytestosterone (2�, 6�,
�) were supplied by Steroids Reference CollectionD.N. Kick, Department of Chemistry, Queen Maryollege, London, England). 1.2 �Ci/ml [1�-3H]-�4
as supplied from New England Nuclear-Perkin ElmerBoston, MA, USA). All other analytical or HPLC gradehemicals were obtained from commercial sources.
For cell cultures, RPMI1640 (the human prostateell lines LNCaP and C16sx) and DMEM (the humanreast cancer cell line MCF-7) w/o phenol red, fetal
ovine serum (FBS), l-glutammine and antibiotics asell as trypsin/EDTA w/o phenol red and 1× DulbeccoBS, pH 7.2 were purchased by GibcoBRL/InvitrogenCarlsbad, CA, USA). Dimethylsulfoxide (DMSO)l Interactions 169 (2007) 1–14 3
were purchased by Sigma–Aldrich (St. Louis, MO,USA). The estrogen receptor antagonist ICI 182,780was purchased by Tocris (Avonmouth, UK). The cellproliferation kit “Cell Titer 96 AQueous One SolutionCell Proliferation Assay” to perform the 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS) assay waspurchased by Promega (Wisconsin, MA, USA).
2.2. Animals and treatments: in vivo study
Experiments were carried out in compliance with theethical provisions enforced by the European Union andauthorized by the National Committee of the Italian Min-istry of Health on in vivo experimentation.
Thirty-six nulliparous pregnant CD1 female micewere purchased from Harlan Italy. They were keptunder standard laboratory conditions (22 ± 0.5 ◦C roomtemperature, 50–60% relative humidity, 12 h dark–lightalternation with 12–14 air changes/h) with water andfood (NIH-07 Open Formula diet purchased fromMucedola, Bergamo Italy) available ad libitum.
Twenty-four dams were randomly divided into twotreatment groups: 0 (vehicle only), �-HCH (15 mg/kgbw) suspended in olive oil (Sigma–Aldrich). Treatmentswere administered daily by gavage from GD 9 to 16(day 0 = positive vaginal plug). Dose level was selectedaccording to the literature and in order to avoid evidentsigns of maternal toxicity or embryotoxicity [3,6,26].The dose level was calculated on the basis of the dams’weight on the first treatment day. The concentration of �-HCH solution was calculated in order to supply volumesof 6.25 �l/g of body weight, i.e. a pregnant mouse of 32 gbw on the first day of treatment received a daily volume of200 �l. Hereafter offspring of treated dams are referredto by the material administered to their dams.
Individual body weight and food consumption ofdams were recorded on GD 6, daily during the treat-ment period, on GD 17, on the day after the parturition(PND 1) and then twice a week until the offspring wean-ing on PND 22. On PND 1, pups were counted, weighedand checked for the presence of external malformationsand/or stillbirths. Litters were weighed twice a week andexamined for the morphological and sexual developmen-tal parameters: pinna detachment, incisor appearance,ear and eye opening, vaginal patency. Dams and halfF1 females from control and treated groups were sac-rificed by intraperitoneal injection of Pentothal Sodium
followed by cervical dislocation on PND 22; the remain-ing F1 females were sacrificed on PND 60, i.e. at fullysexual maturity. Uteri, ovaries and mammary glands ofboth dams and F1 females were excised and weighted.
iologica
4 F. Maranghi et al. / Chemico-BA further group of twelve pregnant CD1 mice wasrandomly divided into two treatment groups: 0 (threedams) and 15 mg/kg bw of �-HCH (nine dams). �-HCHin olive oil or vehicle only was administered daily bygavage to dams from GD 9 to 16. Dams and F1 femaleswere sacrificed on PND 22 by cervical dislocation andtheir livers were immediately excised for microsomalpreparation.
2.3. Histology
Uteri, ovaries and mammary glands of all dams andpups from the main experiment group were fixed inBouin’s solution and stored in 80% ethyl alcohol. Theywere embedded in paraffin, cut into 5-�m sections andstained with haematoxylin and eosin for the examina-tion under a light microscopy (Nikon Microphot FX)with different lenses.
The findings of the histological evaluation were alsoutilised to target a detailed quantitative assessment per-formed by means of histomorphometry.
2.4. Histomorphometry
Slides of uteri and mammary glands were examinedby means of an image analysis system (Arkon by Nikon)applied to an optical microscope (Nikon Microphot FX).
Uteri: Using a 10× lens, the ratio between area ofthe endometrium and the myometrium was measured.To evaluate tissue maturation according to the luminalepithelial cell height, relative percentage of endometrialstroma, using a 10× lens, the rate between the relativearea occupied by glands inside the endometrium and thetotal endometrium area was measured.
Mammary glands: Using a 10× lens, the tubular den-sity as the rate between computer screen area and numberof tubuli/area was measured.
Ovaries: Follicles were scored in randomly selectedsections representing approximately 10% (five to sevensections) of each ovary. Follicle were scored on thebasis of morphological criteria according to the canoni-cal classification of Pedersen and Peters [27] as follows:“primordial/primary”, if they consisted of an intactenlarged oocyte with a visible nucleolus and a singlelayer of fusiform or cuboidal granulosa cells (types 3b,4 and 5a); “preantral”, if they contained an oocyte with avisible nucleolus, more than one layer of granulosa andthecal cells (types 5b and 6); follicle of this type contain-
ing one or more antral spaces were classified as “antral”(types 7 and 8). The diameter of oocyte was measured bya computer software using the KS 300 Image AnalysisSystem (Kontron Corp.). The mean number of differentl Interactions 169 (2007) 1–14
types of follicles and oocyte diameter were calculatedusing single ovaries from at least three different animalsfor each experimental group.
2.5. Biochemical analysis
Preparation of liver microsomes: Liver microsomeswere prepared as previously described [28]. Briefly, liv-ers of dams and F1 female mice on PND 22 from controland treated groups were homogenized in 4 volumes ofTris–Sucrose buffer (pH 7.4). Each homogenate was cen-trifuged at 8700 × g for 20 min; then the supernatant wascentrifuged at 105,000 × g for 70 min. The obtained pel-lets were homogenized in KCl buffer (pH 7.4) and thencentrifuged at 105,000 × g for 70 min. Microsomes wereresuspended in Tris–Sucrose buffer (pH 7.4) and aliquotswere stored at −80 ◦C until the enzymatic activity assayswere performed. The protein content was determinedaccording to Oyama and Eagle [29] using bovine serumalbumin as a standard.
Testosterone hydroxylase activity assay: The micro-somal activity of TST hydroxylase was determinedby the method of Sanwald et al. [30] with minormodifications. The TST hydroxylation reaction wasmeasured under the linear range of time, substrateand protein concentration. The standard incubationmixture (0.5 ml) contained: 1 mg/ml microsomal pro-tein, 2 mM G6P, 2U/ml G6PDH, 1 mM NADP in50 mM Tris buffer, 1 mM EDTA (pH 7.4). The mix-tures were kept at 37 ◦C for 3 min before startingthe reaction, by TST addition (1 mM, final concen-tration). After 10 min at 37 ◦C under gentle shaking,the reaction was stopped by adding 3.5 ml ice-colddichloromethane containing corticosterone (6.25 �Mfinal concentration) as internal standard (IS). Blankswere carried out in parallel and consisted of a stan-dard incubation mixture without NADP. The mixturewas extracted twice with dichloromethane and thenTST, its metabolites and IS were detected by HPLCanalysis (λ = 245 nm). The column (Restek C18 pin-nacle ODS ammine) was isocratically eluted with amobile phase of methanol:water (pH 4.0):tetrahydro-furan (35:55:10) at a flow-rate of 0.8 ml/min. TST,its metabolites and IS were identified by comparingtheir retention times with pure analytical standards;their amounts were determined referring to a cali-bration straight line (average correlation coefficientR2 = 0.995–0.999) prepared with known amounts of the
analytical standards of testosterone and its metabo-lites (range of tested concentrations: 10 �M–1 mM and1–75 �M, respectively). The extraction efficiency was102 ± 8% (means ± S.D.).
ologica
hf[tcbka3aNcet1manwscw
2
iacifp3rwcIs[a5a“Aivcmp3m
F. Maranghi et al. / Chemico-Bi
Aromatase activity assay: Aromatase activity inepatic microsomes was detected by measuring theormation of tritiated-water, according to You et al.31]. Microsomal samples (0.1 mg/ml, final concentra-ion) were incubated in a standard mixture (1.05 ml)ontaining: 4 mM G6P, 2 U/ml G6PDH, 143.5 �M unla-eled �4 and 1.2 �Ci/ml [1�-3H]-�4. The mixture wasept at 37 ◦C for 3 min before starting the reaction, bydding 0.5 mM NADP and then carried out for 80 min at7 ◦C under gentle shaking. Blanks were run in parallelnd consisted of a standard incubation mixture withoutADP. The reaction was stopped by the addition of 5 mlhloroform and 0.5 ml deionized water. Samples werextracted (10 min) and centrifuged (5 min at 800 × g),hen an aliquot of aqueous phase (1.0 ml) was mixed with.0 ml of charcoal, dextran-coated (5–0.5%, w/v). Theixture was vortexed (60 s) and centrifuged for 30 min
t 3000 × g. For radioactivity counting, 1 ml of super-atant was added to 10 ml of scintillation liquid. Samplesere counted for 5 min using a LS6500 Beckman liquid
cintillation counter, providing for quench correction andhecking for chemiluminescence, if necessary samplesere kept in the dark for 3 h before counting.
.6. In vitro cell viability (MTS) assays
In order to assess in a standardized way �-HCH abil-ty to interfere with the ER-�-mediated pathway, itsnti-proliferative role was tested by MTS assay. Theell line LNCaP was maintained as monolayer culturen RPMI1640 w/o phenol red, supplemented with 10%oetal bovine serum (FBS), 2 mM l-glutamine, 1 mMiruvic acid, 100 U/ml penicillin and streptomycin, at7 ◦C, 5% CO2, in water-saturated atmosphere. Theange of �-HCH concentrations tested (1 nM–100 �M)ere based on the limited data available on internal con-
entrations of �HCH in biological fluids and/or tissues.n particular, the selected concentration range was con-istent with those detected in human biological fluids32], as well as with the range of levels detected in bloodnd placenta of pregnant mice treated per os with eitheror 25 mg/kg bw of �HCH [33]. The in vitro indirect
ssessment of cell proliferation was determined using theCell Titer 96 AQueous One Solution Cell Proliferationssay” Promega kit and following the manufacturer’s
nstruction. Our experimental conditions include a star-ation period (14–16 h without FBS) to synchronizeells prior the assessment of viability/proliferation and
RNA expression. Briefly, after the synchronyzationeriod, LNCaP cells were plated at a density of× 103 cells/well in 96-well plates in 100 �l cultureedium. Treatments with different concentrations of
l Interactions 169 (2007) 1–14 5
�HCH, with or without 100 nM ICI 182,780 (both dis-solved in DMSO and diluted in 1× DPBS, pH 7.2), wereperformed in triplicate at the concentrations indicated infigure’s legends (see Section 3). After 3 days of growth,cell cultures were quenched adding 20 �l (1/5 cell cul-ture volume) of “Cell Titer 96 AQueous One SolutionReagent” and further incubated at 37 ◦C in the dark.Absorbance was measured at 490 nm using a Bio-Rad(Richmond, CA, USA) Microplate Reader, model 3550.
2.7. RNA extraction and RT-PCR
To determine hER-� gene expression levels (as wellto confirm the absence of hER-�), LNCaP cells werecultured as described above. Furthermore, as a sourceof control RNA/cDNA references for both ERs, thehuman prostate cell line C16sx, expressing solely ER-�[34], and the human breast cancer cell line MCF-7,known to express ER-�, were used. RNA from allthree-cell lines were homogenized in TRIzol reagent(Invitrogen) and total RNA was isolated accordingto the manifacturer’s instructions and further purifiedafter DNase I treatment. cDNA (from 1 �g RNA)was made using RT-PCR by means of SuperScriptTM
III First-Strand Synthesis System (Invitrogen) wasperformed according to the manifacturer’s instructions.PCR amplification conditions (2 �l cDNA out of 20)were as follows: an automatic hot-start Taq DNAPolimerase activation step at 95 ◦C for 5 min plus 35cycles of denaturation at 95 ◦C for 30 s, annealingat 58 ◦C for 30 s, and elongation at 72 ◦C for 30 s.The following primers [35] were used: for hER-�(ESR1/NR3A1, GeneBank Accession No. NM 000125;amplicon expected size: 299 nt), 5′-AGACATGAGAG-CTGCCAACC-3′ (forward) and 5′-GCCAGGCACA-TTCTAGAAGG-3′ (reverse), for hER-� (ESR2/NR3A2, GeneBank Accession No. XM 495993;amplicon expected size: 346nt), 5′-TCACATCTGTAT-GCGGAACC-3′ (forward) and 5′-CGTACCACTTCC-GAAGTCGG-3′ (reverse), and for 18S rRNA (Gene-Bank Accession No. X56974), used as control, 5′-GA-GGCCCTGTAATTGGAATGAG-3′ (forward) and 5′-GCAGCAACTTTAATATACGCTATTGG-3′ (reverse).ERs gene expression and PCR products’ size wereverified by electrophoresis on 2% agarose gel.
2.8. Statistical analysis
Differences among controls and �-HCH-treatedgroups were analysed by means of: Student’s t-test withBonferroni correction for body weight gain in dams andlitters (continuous data); Mann–Whitney U-test for his-

iologica
6 F. Maranghi et al. / Chemico-Btomorphometrical data (discontinuous data); χ2 test fordevelopmental parameters and histopathological data.
Data of follicles and oocytes were analysed usingGraphPad Instat 3.0 statistical software (GraphPad Inc.,San Diego, CA). Differences between the means numberof the different types of follicles and oocyte diameters in�-HCH treated and control ovaries were evaluated usingMann–Whitney rank test and one-way analysis of vari-ance (ANOVA), with a statistical significance assignedat P ≤ 0.05.
Data of biochemical parameters were analysed usingGraphPad Prism 4 software (version 4.01 for Windows).ANOVA followed by Bonferroni’s test were used tocompare control and treated groups, with a statisticalsignificance cut off value at P ≤ 0.05.
Results of in vitro assays were analyzed by Student’st-test.
3. Results
3.1. In vivo studies
Treatment with �-HCH during pregnancy did not pro-duce overt signs of maternal toxicity such as mortality
Table 1Litter parameters of F1 female pups treated in utero on GD 9–16 with vehicfully sexual maturity (PND 60)
Treatment group Control
Pup body weight (g)PND 1 1.68 + 0.04PND 5 3.47 + 0.19PND 12 6.78 + 0.27PND 22 12.86 + 0.54PND 60 27.1 + 1.8
Treatment group Control (n = 2
Uterus weight (�g)PND 22 absolute 24.59 + 7.5PND 22 relative 2.19 + 0.19
Treatment group Control (n = 4
Uterus weight (�g)PND 60 absolute 176.42 + 55.3PND 60 relative 6.53 + 2.07
Treatment group Control (n = 42
Vaginal opening completedPND 33 35.7%PND 34 71.4%PND 35 95.2%PND 36 100%
* Significantly different from control group (P ≤ 0.05, unpaired t-test).
l Interactions 169 (2007) 1–14
or abnormal behaviour. No significant differences wereobserved as regards body weight, body weight gain,gestation length; delivery occurred in all dams on GD18–19. The absolute and relative dam uterus weight wasnot affected by the treatment (0.25 ± 0.055 in �-HCHversus 0.26 ± 0.056 in controls). No gross malforma-tions were observed in pups on PND 1. No significantdifferences were observed concerning the average num-ber of live pups/litter (13.08 ± 0.58 in �-HCH versus11.41 ± 0.71 in controls), incidences of stillbirths orrunts, sex ratio and survival to weaning (data not shown).Mean pup weight and mean weight gain were not signif-icantly different up to fully sexual maturity (PND 60).Developmental landmarks were unaltered. Earlier vagi-nal patency was observed in �-HCH treated F1 females(average 2 days) in comparison to control (Table 1).
The absolute and relative uterus weight was increased(P < 0.05) in F1 �-HCH treated females sacrificed onPND 22 in comparison to controls. At fully sexualmaturity (PND 60), both the relative and absolute
uterus weight was not significantly increased in �-HCHtreated females although the mean values were stillslightly higher than controls (10% and 4%, respectively)(Table 1).le only (olive oil), �-HCH, sacrificed after weaning (PND 22) and at
�-HCH (15 mg/kg bw)
1.61 + 0.023.05 + 0.056.39 + 0.10
11.75 + 0.4128.68 + 2.77
3) �-HCH (n = 24)
28.68 + 5.7*
2.48 + 0.44*
5) �-HCH (n = 56)
6 194.79 + 63.346.81 + 2.17
) �-HCH (n = 56)
92.9%100%
––
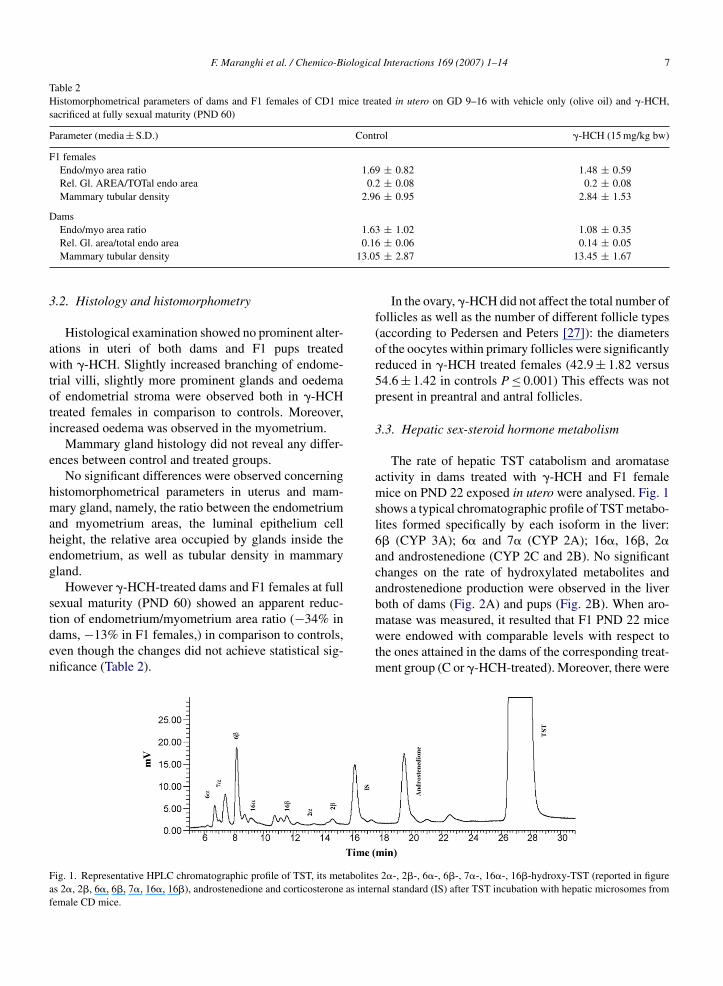
F. Maranghi et al. / Chemico-Biological Interactions 169 (2007) 1–14 7
Table 2Histomorphometrical parameters of dams and F1 females of CD1 mice treated in utero on GD 9–16 with vehicle only (olive oil) and �-HCH,sacrificed at fully sexual maturity (PND 60)
Parameter (media ± S.D.) Control �-HCH (15 mg/kg bw)
F1 femalesEndo/myo area ratio 1.69 ± 0.82 1.48 ± 0.59Rel. Gl. AREA/TOTal endo area 0.2 ± 0.08 0.2 ± 0.08Mammary tubular density 2.96 ± 0.95 2.84 ± 1.53
DamsEndo/myo area ratio 1.63 ± 1.02 1.08 ± 0.35
0.1613.05
3
awtoti
e
hmaheg
stden
Faf
Rel. Gl. area/total endo areaMammary tubular density
.2. Histology and histomorphometry
Histological examination showed no prominent alter-tions in uteri of both dams and F1 pups treatedith �-HCH. Slightly increased branching of endome-
rial villi, slightly more prominent glands and oedemaf endometrial stroma were observed both in �-HCHreated females in comparison to controls. Moreover,ncreased oedema was observed in the myometrium.
Mammary gland histology did not reveal any differ-nces between control and treated groups.
No significant differences were observed concerningistomorphometrical parameters in uterus and mam-ary gland, namely, the ratio between the endometrium
nd myometrium areas, the luminal epithelium celleight, the relative area occupied by glands inside thendometrium, as well as tubular density in mammaryland.
However �-HCH-treated dams and F1 females at fullexual maturity (PND 60) showed an apparent reduc-
ion of endometrium/myometrium area ratio (−34% inams, −13% in F1 females,) in comparison to controls,ven though the changes did not achieve statistical sig-ificance (Table 2).ig. 1. Representative HPLC chromatographic profile of TST, its metabolitess 2�, 2�, 6�, 6�, 7�, 16�, 16�), androstenedione and corticosterone as interemale CD mice.
± 0.06 0.14 ± 0.05± 2.87 13.45 ± 1.67
In the ovary, �-HCH did not affect the total number offollicles as well as the number of different follicle types(according to Pedersen and Peters [27]): the diametersof the oocytes within primary follicles were significantlyreduced in �-HCH treated females (42.9 ± 1.82 versus54.6 ± 1.42 in controls P ≤ 0.001) This effects was notpresent in preantral and antral follicles.
3.3. Hepatic sex-steroid hormone metabolism
The rate of hepatic TST catabolism and aromataseactivity in dams treated with �-HCH and F1 femalemice on PND 22 exposed in utero were analysed. Fig. 1shows a typical chromatographic profile of TST metabo-lites formed specifically by each isoform in the liver:6� (CYP 3A); 6� and 7� (CYP 2A); 16�, 16�, 2�and androstenedione (CYP 2C and 2B). No significantchanges on the rate of hydroxylated metabolites andandrostenedione production were observed in the liverboth of dams (Fig. 2A) and pups (Fig. 2B). When aro-
matase was measured, it resulted that F1 PND 22 micewere endowed with comparable levels with respect tothe ones attained in the dams of the corresponding treat-ment group (C or �-HCH-treated). Moreover, there were2�-, 2�-, 6�-, 6�-, 7�-, 16�-, 16�-hydroxy-TST (reported in figurenal standard (IS) after TST incubation with hepatic microsomes from
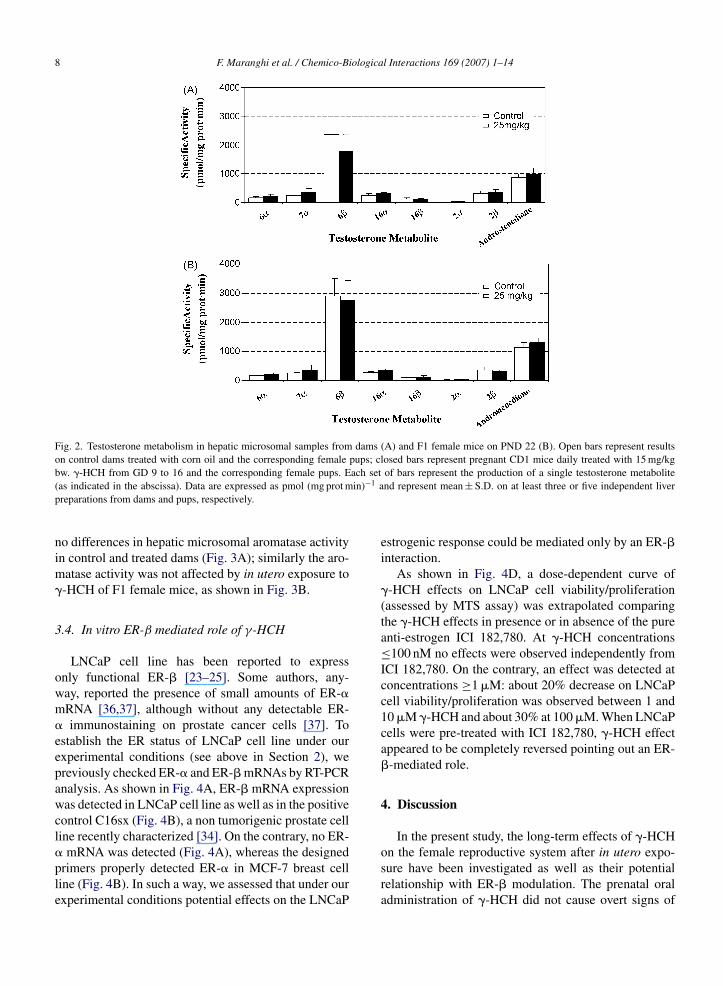
8 F. Maranghi et al. / Chemico-Biological Interactions 169 (2007) 1–14
Fig. 2. Testosterone metabolism in hepatic microsomal samples from dams (A) and F1 female mice on PND 22 (B). Open bars represent resultson control dams treated with corn oil and the corresponding female pups; closed bars represent pregnant CD1 mice daily treated with 15 mg/kg
ach setin)−1 a
bw. �-HCH from GD 9 to 16 and the corresponding female pups. E(as indicated in the abscissa). Data are expressed as pmol (mg prot mpreparations from dams and pups, respectively.
no differences in hepatic microsomal aromatase activityin control and treated dams (Fig. 3A); similarly the aro-matase activity was not affected by in utero exposure to�-HCH of F1 female mice, as shown in Fig. 3B.
3.4. In vitro ER-β mediated role of γ-HCH
LNCaP cell line has been reported to expressonly functional ER-� [23–25]. Some authors, any-way, reported the presence of small amounts of ER-�mRNA [36,37], although without any detectable ER-� immunostaining on prostate cancer cells [37]. Toestablish the ER status of LNCaP cell line under ourexperimental conditions (see above in Section 2), wepreviously checked ER-� and ER-� mRNAs by RT-PCRanalysis. As shown in Fig. 4A, ER-� mRNA expressionwas detected in LNCaP cell line as well as in the positivecontrol C16sx (Fig. 4B), a non tumorigenic prostate cellline recently characterized [34]. On the contrary, no ER-
� mRNA was detected (Fig. 4A), whereas the designedprimers properly detected ER-� in MCF-7 breast cellline (Fig. 4B). In such a way, we assessed that under ourexperimental conditions potential effects on the LNCaPof bars represent the production of a single testosterone metabolitend represent mean ± S.D. on at least three or five independent liver
estrogenic response could be mediated only by an ER-�interaction.
As shown in Fig. 4D, a dose-dependent curve of�-HCH effects on LNCaP cell viability/proliferation(assessed by MTS assay) was extrapolated comparingthe �-HCH effects in presence or in absence of the pureanti-estrogen ICI 182,780. At �-HCH concentrations≤100 nM no effects were observed independently fromICI 182,780. On the contrary, an effect was detected atconcentrations ≥1 �M: about 20% decrease on LNCaPcell viability/proliferation was observed between 1 and10 �M �-HCH and about 30% at 100 �M. When LNCaPcells were pre-treated with ICI 182,780, �-HCH effectappeared to be completely reversed pointing out an ER-�-mediated role.
4. Discussion
In the present study, the long-term effects of �-HCH
on the female reproductive system after in utero expo-sure have been investigated as well as their potentialrelationship with ER-� modulation. The prenatal oraladministration of �-HCH did not cause overt signs of
F. Maranghi et al. / Chemico-Biologica
Fig. 3. Aromatase activity in hepatic microsomes from dams (A) andF1 female mice on PND 22 (B). Open bars represent results on controldams treated with corn oil (CTRL) and the corresponding female pups;closed bars represent pregnant CD1 mice daily treated with 15 mg/kgbw. �-HCH from GD 9 to 16 and the corresponding female pups. Dataaap
ml�fpw([irttoatsp
a
re expressed as pmol (mg prot min)−1 and represent mean ± S.D. ont least three or five independent liver preparations from dams andups, respectively.
aternal or neonatal toxicity both during pregnancy andactation at the selected dose level (15 mg/kg bw/day).-HCH accelerated the time of vaginal patency in F1emales. It may be noteworthy that earlier vaginalatency has been induced in rats by other compoundsith either estrogenic (metoxychlor), antiestrogenic
tamoxifen) or antiandrogenic (flutamide) activities38,39]. Relative and absolute uterus weight wasncreased in F1 females at weaning. At fully sexual matu-ity this increase was no longer significant. According tohese parameters, �-HCH may exert effects comparableo that of an oestrogen agonist during the developmentf female reproductive system [40]. However, this effectppears to be reversible at fully sexual maturity whenhe steroid balance reaches the complete efficiency. The
ame behaviour on uterine weight is described after ratrenatal exposure to genistein in diet [41].The subtle histological and histomorphometricallterations of the uterus suggest a weak modulating activ-
l Interactions 169 (2007) 1–14 9
ity of steroid balance. It is noticeable that also genistein,both in vitro and in vivo, shows a weak estrogenic activ-ity on endometrium [42,43]. Moreover, such activity ismodulated in vitro by natural ligands of ERs, such as E2[42].
In ovaries, �-HCH did not affect the overall numberand/or type of follicles but the oocyte diameters in pri-mary follicles were significantly reduced in F1 females.Folliculogenesis in the mouse ovary begins early afterbirth with the formation of primordial follicles consist-ing of an oocyte in the meiotic prophase surrounded bya layer of undifferentiated granulosa cells. The follicu-lar development throughout primary, secondary, antraland pre-ovulatory stages and oocyte growth depends onseveral factors, including hormones and several locallyproduced growth factors synthesized by the granulosacells and the oocyte itself [44]. Coupling in the formof gap junctions between the oocyte and the surround-ing granulosa cells is a major factor promoting oocytegrowth [45]. �-HCH abolishes rat oocyte directed follicleorganizing activity in vitro [46]. Increased degenera-tion and reduced proliferation in early mouse embryohave been also attributed to �-HCH impairing gapjunction-mediated cell communication between oocyteand cumulus cells [5]. Our data on oocytes are consistentwith an impaired formation of primary follicles. Suchan effect was not yet recorded although previous dataindicate that �-HCH and POPs in general (singly or inmixture) disrupt mammalian oocyte maturation and fol-licle physiology even at very low concentration [47].�-HCH may impair gap junction function in other tis-sues such as myometrium [48]. Some data support thatestrogens are involved directly in gap junction-mediatedcommunications in in vitro systems [49]. Besides, inmammary gland of ER-�KO mice, alterations in adhe-sion molecules and ultrastructural organization wereobserved suggesting a role for ER-� in organisation andadhesion of epithelial cells [50].
It is noteworthy that �-HCH may exert differenteffects depending on the developmental stage andendocrine status. Preliminary findings on post-nataltreatment with �-HCH showed no evident effects onuterus, whereas the histomorphometry of mammaryglands was affected (F. Maranghi, personal communi-cation).
A recent rat two-generation reproductive toxicitystudy with �-HCH showed that endocrine effects wereelicited only in presence of systemic toxicity both in
F1 and F2 generations at a dose level (300 ppm in thediet corresponding to about 17 mg/kg bw during preg-nancy) comparable to that used in the in vivo sectionof the present study [6]. Nevertheless, the prolonged,
10 F. Maranghi et al. / Chemico-Biological Interactions 169 (2007) 1–14
Fig. 4. Determination of the LNCaP ER status (A) and ER-�-mediated effects of �HCH on LNCaP cell viability/proliferation (B). ER-� and ER-�gene expression has been checked by RT-PCR (A). ER-� gene expression analysis was performed in LNCaP (A, left panel) and, as positive control,in C16sx (A, middle panel). ER-� gene expression analysis was performed in LNCaP (A, left panel), in the negative control C16sx (A, middlepanel) and, as positive control, in MCF-7 (A, right panel). RT+ means that RT-PCR was done with all reagents and enzymes, whereas RT− means
y PCRay (B).
D valu
that no reverse-transcription was performed prior to the amplification blines following �HCH treatments (72 h) was determined by MTS assICI 182,780 (100 nM). CTRL+ corresponds to untreated cells. 100% Otwice in triplicate.
continuous exposure of the two-generation protocol isdifferent from our study design, targeted to administra-tion during reproductive tract organogenesis. Moreover,in our study no �HCH general toxicity was observed inpregnant and F1 mice, whereas the rat appeared morevulnerable suggesting interspecies differences. In par-ticular, pre-pubertal rats showed retardation of vaginalpatency associated with reduced body weight gain: thisis in sharp contrast with earlier vaginal patency and nor-mal growth observed in our study. However, signs ofendocrine alterations were observed in the rat study such
as the increased LH and decreased T3 and T4 serumlevels.Since a possible influence of �-HCH exposure onsteroid hormone metabolism in female rodents has not
. Cell viability and (indirect) proliferation assay of prostate cancer cellLNCaP cells were grown with (�) or without (♦) the ER antagonistes were arbitrarily assigned to CTRL+. Experiments were performed
yet been ruled out, effects on key enzymes were tested.Indeed, it has been demonstrated that in vitro �-HCHmay inhibit steroidogenesis and, in particular, that �-HCH interferes with TST synthesis in hCG-stimulatedLeydig cells [51]. It is recognized that anomalies ofreproductive development could depend on an unbal-ance of sex-steroid hormones metabolism, due to theimpairment of their biosynthesis/catabolism ratio at dif-ferent sites in the organism. Recent studies showed thatexposure to different xenoestrogens in adult rats andmice [52,53] induces changes in hepatic biotransforma-
tion capabilities, causing concomitant alterations in themetabolism of endogenous substrates such as TST. Like-wise, similar enzymatic effects in animals exposed todifferent xenobiotic during prenatal or neonatal period
ologica
hcfpd(teO�Tameh
tfsoletltb3oterndmaftpti
u[[tragoa[e
F. Maranghi et al. / Chemico-Bi
ave been reported [54,55]. The first step of TSTatabolism is its hydroxylation mainly in the liver by dif-erent cytochrome P450 isoenzymes (CYPs) at severalositions or dehydrogenation to produce androstene-ione [56]. Aromatase, a member of the CYP familyCYP19), represents a key enzyme for estrogens biosyn-hesis, which in addition to steroidogenic tissues, isxpressed in many others sites, including the liver [22].ur results showed that a prenatal exposure to 15 mg/kg-HCH produced no significant alterations on the hepaticST catabolism as well as on the biotransformation ofndrogen into estrogen, catalysed by aromatase in femaleice exposed in utero, providing evidences that �-HCH
ffects are not associated with an impairment of steroidormone metabolism.
In mammals, ER-� down-regulates ER-� transcrip-ional activity in cells that express both receptors, as theemale reproductive tissues but also e.g. in skeletal tis-ue [57]. In female mice, both ERs are expressed in thevary, but ER-� alone seems to play a direct role in fol-iculogenesis, facilitating the follicle maturation from thearly antral to the preovulatory stage [58]. In the imma-ure uterus, ER-� and ER-� are expressed at comparableevels in the epithelium and in the stroma. It appearshat ER-� keeps the uterus quiescent before the ovaryegins to secrete E2, probably via the DHT metabolite�Adiol, an ER-� ligand produced by the pre-pubertalvary [59]. In the mature uterus, ER-� plays impor-ant roles in decidualization and in cervical ripening,ssential for the implantation of the foetus and for partu-ition, respectively [14]. ER-�KO mice showed a grosslyormal mammary development and the females had noifficulties in breast feeding the offspring, suggesting aajor role of ER-� in the E2-mediated effects on suchtissue [60]. ER-� may also act as a pro-differentiation
actor in the prostatic epithelium [17] being able to coun-eract the proliferative ability of the AR [61]. Indeed,rostate growth is regulated by ER-� and 3�Adiol is therue ER-� ligand in this tissue [19]. Similarly, our datandicate a potential of �-HCH to interfere with ER-�.
The phytoestrogen genistein is considered a nat-ral Selective Estrogen Receptor Modulator (SERM)62] with high ER-� affinity and tissue-specific effects11]. As mentioned above, it is noteworthy that genis-ein may elicit similar effects on the developing femaleeproductive system as �-HCH [63]. Moreover, whendministered to male mice during the juvenile period,enistein causes a slightly decreased sperm count with-
ut relevant effects on reproductive organ weights, i.e.pattern of effect similar to that observed for �-HCH3]. However, the potent ER-� agonist DES also inducedffects similar to �-HCH on spermatogenesis [3].
l Interactions 169 (2007) 1–14 11
Our in vitro data showed, to our knowledge forthe first time, that �-HCH acts through ER-� atconcentration ≥1 �M directly affecting LNCaP cellviability/proliferation. Indeed, we demonstrated by RT-PCR that in our experimental conditions LNCaP cells donot express ER-� mRNA. Such a result is in agreementwith the reports of different authors [23–25], showingER-� as transcriptionally silenced in prostate cells byepigenetic mechanisms (promoter hypermethylation).On the other hand, few studies reported ER-� mRNAexpression in LNCaP cells [34,36,37,64,65]. Such con-troversial results might be explained taking into accountdifferent culture’s conditions that may alter ERs’ tran-scriptional regulation, i.e. serum absence (leading tostarvation, quiescence or differentiation) versus serumpresence (proliferation). In rodent ventral prostate cells,ER-� has been suggested to act as a proliferative inducerwhereas ER-� may either counteract proliferation and/ortrigger differentiation [19]. Because of our data on thedecreased LNCaP cell viability/proliferation, we mightspeculate that �HCH could interfere with cell cycle entrydecisions, namely altering the switch between prolifera-tion and differentiation. Moreover, our findings suggestthat �-HCH may interfere with ER-�, particularly inreproductive tissues where 3�Adiol is the main receptorligand, such as prostate and immature ovary [19,59].
The effect of �-HCH on the proliferation–differentiation control switch might deserve more atten-tion, since a protective role of ER-� against estrogen-dependent tumor progression in breast, ovary, colon andprostate has been suggested [66].
Interestingly, recent in vivo data on prenatal exposureof rats to 2,2′,4,4′,5-pentabromo-diphenylether (PBDE99), a halogenated compound with known endocrineactivity, was shown to modulate ERs gene expressionwithout signs of systemic toxicity, suggesting that a com-mon interfering mechanism on E2-regulated genes mightbe postulated [67].
Finally, our data point to a more detailed investiga-tion on ER-�-mediated mechanisms of xenobiotics withpotential endocrine activities.
Acknowledgments
The present study was supported by: the EU Networkof Excellence “CASCADE” (FOOD-CT-2004-506319);the Italian National Health System project “Multicentricstudy on the relationship between endocrine pathology
and environmental and occupational exposure to xeno-biotics, with the aim of creating a disease register by theinstitution of a National Reference Center for EndocrineDisrupting Chemicals” (Grant No. Fasc. 5AOC). Prof.
iologica
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
12 F. Maranghi et al. / Chemico-B
M. De Felici was supported by the EU project “GENDIS-RUPT” (QLK4-CT-2002-02403).
References
[1] T. Damstra, Potential effects of certain persistent organic pollu-tants and endocrine disrupting chemicals on the health of children,J. Toxicol. Clin. Toxicol. 40 (2002) 457–465.
[2] EFSA Scientific Panel. Opinion of the Scientific Panel on contam-inants in the food chain on a request from the Commission relatedto gamma-HCH and other hexachlorocyclohexanes as undesir-able substances in animal feed. Question No. EFSA-Q-2003-067.EFSA J. 250 (2005) 1–39.
[3] M.E. Traina, M. Rescia, E. Urbani, A. Mantovani, C. Macrı, C.Ricciardi, A.V. Stazi, P. Fazzi, E. Cordelli, P. Eleuteri, G. Leter, M.Spano, Long-lasting effects of lindane on mouse spermatogenesisinduced by in utero exposure, Reprod. Toxicol. 17 (2003) 25–35.
[4] N. Pages, M.P. Suviat, S. Bouvet, F. Goudey-Perriere, Reproduc-tive toxicity of lindane, J. Soc. Biol. 196 (2002) 325–338.
[5] M. Scascitelli, F. Pacchierotti, Effects of lindane on oocyte matu-ration and preimplantation embryonic development in the mouse,Reprod. Toxicol. 17 (2003) 299–303.
[6] I. Matsuura, T. Saitoh, E. Tani, Y. Wako, H. Iwata, N. Toyota,Y. Ishizuka, M. Namiki, N. Hoshino, M. Tsuchitani, Y. Ikeda,Evaluation of a two-generation reproduction toxicity study addingendpoints to detect endocrine disrupting activity using lindane, J.Toxicol. Sci. 30 (Spec No.) (2005) S135–S161.
[7] M.L. Scippo, C. Argiris, C. Van De Weerdt, M. Muller, P. Willem-sen, J. Martial, G. Maghuin-Rogister, Recombinant humanestrogen, androgen and progesterone receptors for detection ofpotential endocrine disruptors, Anal. Bioanal. Chem. 378 (2004)664–669.
[8] P. Balaguer, F. Francois, F. Comunale, H. Fenet, A.M. Boussioux,M. Pons, J.C. Nicolas, C. Casellas, Reporter cell lines to studythe estrogenic effects of xenoestrogens, Sci. Total Environ. 233(1999) 47–56.
[9] H.S. Lee, K. Miyauchi, Y. Nagata, R. Fukuta, S. Sasagawa, H.Endoh, S. Kato, H. Horiuchi, M. Takagi, A. Ohta, Employmentof the human estrogen receptor beta ligand-binding domain andco-activator SRC1 nuclear receptor-binding domain for the con-struction of a yeast two-hybrid detection system for endocrinedisrupters, J. Biochem. (Tokyo) 131 (2002) 399–405.
10] A. Escande, A. Pillon, N. Servant, J.P. Cravedi, F. Larrea, P. Muhn,J.C. Nicolas, V. Cavailles, P. Balaguer, Evaluation of ligand selec-tivity using reporter cell lines stably expressing estrogen receptoralpha or beta, Biochem. Pharmacol. 71 (2006) 1459–1469.
11] T. Valachovicova, V. Slivova, D. Sliva, Cellular and physiolog-ical effects of soy flavonoids, Mini Rev. Med. Chem. 4 (2004)881–887.
12] B.J. Lee, E.Y. Jung, Y.W. Yun, J.K. Kang, I.J. Baek, J.M. Yon, Y.B.Lee, H.S. Sohn, J.Y. Lee, K.S. Kim, S.Y. Nam, Effects of expo-sure to genistein during pubertal development on the reproductivesystem of male mice, J. Reprod. Dev. 50 (2004) 399–409.
13] K. Pettersson, J.A. Gustafsson, Role of estrogen receptor beta inestrogen action, Annu. Rev. Physiol. 63 (2001) 165–192.
14] K.F. Koehler, L.A. Helguero, L.A. Haldosen, M. Warner, J.A.
Gustafsson, Reflections on the discovery and significance of estro-gen receptor beta, Endocr. Rev. 26 (2005) 465–478.15] G.G. Kuiper, E. Enmark, M. Pelto-Huikko, S. Nilsson, J.A.Gustafsson, Cloning of a novel receptor expressed in rat prostateand ovary, Proc. Natl. Acad. Sci U.S.A. 93 (2006) 5925–5930.
[
[
l Interactions 169 (2007) 1–14
16] J.F. Couse, M.M. Yates, B.J. Deroo, K.S. Korach, Estrogenreceptor-beta is critical to granulosa cell differentiation and theovulatory response to gonadotropins, Endocrinology 146 (2005)3247–3262.
17] O. Imamov, A. Morani, G.J. Shim, Y. Omoto, C. Thulin-Andersson, M. Warner, J.A. Gustafsson, Estrogen receptor betaregulates epithelial cellular differentiation in the mouse ventralprostate, Proc. Natl. Acad. Sci U.S.A. 101 (2004) 9375–9380.
18] H. Sakaguchi, J. Fujimoto, B.L. Hong, T. Tamaya, Quantitativeanalysis of estrogen receptor proteins in rat ovary, J. Steroid.Biochem. Mol. Biol. 94 (2005) 83–91.
19] Z. Weihua, R. Lathe, M. Warner, J.A. Gustafsson, An endocrinepathway in the prostate, ERbeta, AR, 5alpha-androstane-3beta,17beta-diol, and CYP7B1, regulates prostate growth, Proc.Natl. Acad. Sci. U.S.A. 99 (2002) 13589–13594.
20] S. Salmi, R. Santti, J.A. Gustafsson, S. Makela, Co-localization ofandrogen receptor with estrogen receptor beta in the lower urinarytract of the male rat, J. Urol. 166 (2001) 674–677.
21] J.L. Pryor, C. Hughes, W. Foster, B.F. Hales, B. Robaire, Criticalwindows of exposure for children’s health: the reproductive sys-tem in animals and humans, Environ. Health Perspect. 108 (2000)491–503.
22] A. Conley, M. Hinshelwood, Mammalian aromatases, Reproduc-tion 121 (2001) 685–695.
23] M. Sasaki, Y. Tanaka, G. Perinchery, A. Dharia, I. Kotcherguina,S. Fujimoto, R. Dahiya, Methylation and inactivation of estrogen,progesterone, and androgen receptors in prostate cancer, J. Natl.Cancer Inst. 94 (2002) 384–390.
24] K.M. Lau, M. LaSpina, J. Long, S.M. Ho, Expression of estro-gen receptor (ER)-� and ER-� in normal and malignant prostaticepithelial cells: regulation by methylation and involvement ingrowth regulation, Cancer Res. 60 (2000) 3175–3182.
25] L.C. Li, R. Chui, K. Nakajima, B.R. Oh, H.C. Au, R. Dahiya,Frequent methylation of estrogen receptor in prostate cancer: cor-relation with tumor progression, Cancer Res. 60 (2000) 702–706.
26] M.E. Traina, E. Urbani, M. Rescia, A. Mantovani, L’insetticidalindano: identificazione dei rischi possibili per la riproduzioneumana, Rapporti Istisan 3 (2001) 1–59.
27] T. Pedersen, H. Peters, Proposal for a classification of oocytes andfollicles in the mouse ovary, J. Reprod. Fertil. 17 (1968) 555–557.
28] E. Testai, L. Vittozzi, Biochemical alterations elicited in rat livermicrosomes by oxidation and reduction products of chloroformmetabolism, Chem. Biol. Interact. 59 (1986) 157–171.
29] V.I. Oyama, H. Eagle, Measurement of cell growth in tissue cul-ture with a phenol reagent (folin-ciocalteau), Proc. Soc. Exp. Biol.Med. 91 (1956) 305–307.
30] P. Sanwald, E.A. Blankson, B.D. Dulery, J. Schoun, H.D. Hue-bert, J. Dow, Isocratic high-performance liquid chromatographicmethod for the separation of testosterone metabolites, J. Chro-matogr. B. Biomed. Appl. 672 (1995) 207–215.
31] L. You, M. Sar, E. Bartolucci, S. Ploch, M. Whitt, Induction ofhepatic aromatase by p,p′-DDE in adult male rats, Mol. Cell.Endocrinol. 178 (2001) 207–214.
32] N. Pant, N. Mathur, A.K. Banerjee, S.P. Srivastava, D.K. Saxena,Correlation of chlorinated pesticides concentration in semen withseminal vesicle and prostatic markers, Reprod. Toxicol. 19 (2004)209–214.
33] T.S. Dikshith, M.K. Srivastava, R.B. Raizada, Fetotoxicity of hex-achlorocyclohexane (HCH) in mice: morphological, biochemicaland residue evaluations, Vet. Hum. Toxicol. 32 (1990) 524–527.
34] S. Nanni, C. Priolo, A. Grasselli, M. D’Eletto, R. Merola, F.Moretti, M. Gallucci, P. De Carli, S. Sentinelli, A.M. Cianci-

ologica
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
F. Maranghi et al. / Chemico-Bi
ulli, M. Mottolese, P. Carlini, D. Arcelli, M. Helmer-Citterich,C. Gaetano, M. Loda, A. Pontecorvi, S. Bacchetti, A. Sacchi,A. Farsetti, Epithelial-restricted gene profile of primary culturesfrom human prostate tumors: a molecular approach to predictclinical behaviour of prostate cancer, Mol. Cancer Res. 4 (2006)79–92.
35] C. Roger, S. Lambard, A. Bouskine, B. Mograbi, D. Chevallier,M. Nebout, G. Pointis, S. Carreau, P. Fenichel, Estrogen-inducedgrowth inhibition of human seminoma cells expressing estro-gen receptor beta and aromatase, J. Mol. Endocrinol. 35 (2005)191–199.
36] A. Latil, I. Bieche, D. Vidaud, R. Lidereau, P. Berthon, O.Cussenot, M. Vidaud, Evaluation of androgen, estrogen (ER alphaand ER beta), and progesterone receptor expression in humanprostate cancer by real-time quantitative reverse transcription-polymerase chain reaction assays, Cancer Res. 61 (2001)1919–1926.
37] M.J. Linja, K.J. Savinainen, T.L. Tammela, J.J. Isola, T. Visakorpi,Expression of ER� and ER� in prostate cancer, Prostate 55 (2003)180–186.
38] R.E. Chapin, M.W. Harris, B.J. Davis, S. M: Ward, R.E.Wilson, M.A. Mauney, A.C. Lockhart, R.J. Smialowicz, V.C.Moser, L.T. Burka, B.J. Collins, The effects of perinatal/juvenilemethoxychlor exposure on adult rat nervous, immune and repro-ductive system function, Fundam. Appl. Toxicol. 40 (1997) 138–157.
39] H.S. Kim, J.H. Shin, H.J. Moon, T.S. Kim, I.H. Kang, J.H. Seok,I.Y. Kim, K.L. Park, S.Y. Han, Evaluation of the 20-day pubertalfemale assay in Sprague–Dawley rats treated with DES, tamox-ifen, testosterone and flutamide, Toxicol. Sci. 67 (2002) 52–62.
40] Y. Nikaido, K. Yoshizawa, N. Danbara, M. Tsujita-Kyutoku, T.Yuri, N. Uehara, A. Tsubura, Effects of maternal xenoestrogenexposure on development of the reproductive tract and mam-mary gland in female CD-1 mouse offspring, Reprod. Toxicol.18 (2004) 803–811.
41] The NTP-CERHR Expert Panel Reports on Genistein(http://cerhr.niehs.nih.gov/chemicals/genistein-soy/genistein/Genistein Report final.pdf) and Soy Formula (http://cerhr.niehs.nih.gov/chemicals/genistein-soy/soyformula/Soy-report-final.pdf), April 2006.
42] U.A. Kayisli, C.A. Aksu, M. Berkkanoglu, A. Arici, Estrogenicityof isoflavones on human endometrial stromal and glandular cells,Endocrinol. Metab. 87 (2002) 5539–5544.
43] P. Diel, R.B. Geis, A. Caldarelli, S. Schmidt, U.L. Leschowsky,A. Voss, G. Vollmer, The differential ability of the phytoestrogengenistein and of estradiol to induce uterine weight and prolifera-tion in the rat is associated with a substance specific modulationof uterine gene expression, Mol. Cell. Endocrinol. 221 (2004)21–32.
44] M.M. Matzuk, D.J. Lamb, Genetic dissection of mammalian fer-tility pathways, Nat. Cell. Biol. 4 (2002) S41–S49.
45] P.M. Wassarman, Channels of communication in the ovary, Nat.Cell. Biol. 4 (2002) S7–S9.
46] R. Li, J.P. Mather, Lindane, an inhibitory of gap junction for-mation, abolishes directed follicle organizing activity in vitro,Endocrinology 138 (1997) 4477–4480.
47] P. Pocar, T.A. Brevini, B. Fischer, F. Gandolfi, The impact of
endocrine disruptors on oocyte competence, Reproduction 125(2003) 313–325.48] R.K. Loch-Caruso, K.A. Criswell, C.M. Grindatti, K.A. Brant,Sustained inhibition of rat myometrial gap junctions and contrac-tions by lindane, Reprod. Biol. Endocrinol. 1 (2003) 62.
[
l Interactions 169 (2007) 1–14 13
49] G. Carruba, L. Cocciadiferro, V. Bellavia, S. Rizzo, C. Tsatsanis,D. Spandidos, P. Muti, C. Smith, P. Mehta, L. Castagnetta, Inter-cellular communication and human hepatocellular carcinoma,Ann. N. Y. Acad. Sci. 1028 (2004) 202–212.
50] C. Foster, S. Kietz, K. Hultenby, M. Warner, J.A. Gustafsson,Characterization of the ERbeta−/− mouse hearth, Proc. Natl.Acad. Sci. U.S.A. 101 (2002) 14234–14239.
51] A.M. Ronco, K. Valdes, D. Marcus, M. Llanos, The mechanismfor lindane-induced inhibition of steroidogenesis in cultured ratLeydig cells, Toxicology 159 (2001) 99–106.
52] M. Kawai, S.M. Bandiera, T.K. Chang, F.M. Poulet, P.M. Vancut-sem, G.D. Bellward, Modulation of hepatic CYP2A1, CYP2C11,and CYP3A9 expression in adult rats by neonatal adminis-tration of tamoxifen, Drug Metab. Dispos. 27 (1999) 1392–1398.
53] V.S. Wilson, G.A. Le Blanc, The contribution of hepatic inactiva-tion of testosterone to the lowering of serum testosterone levelsby ketoconazole, Toxicol. Sci. 54 (2000) 128–137.
54] M. Ishizuka, J. Yonemoto, H. Zaha, C. Tohyama, H. Sone,Perinatal exposure to low doses of 2,3,7,8-tetrachlorodibenzo-p-dioxin alters sex-dependent expression of hepatic CYP2C11, J.Biochem. Mol. Toxicol. 17 (2003) 278–285.
55] E.M. Laurenzana, G. Balasubramanian, C. Weis, B. Blaydes,R.R. Newbold, K.B. Delclos, Effect of nonylphenol on serumtestosterone levels and testicular steroidogenic enzyme activityin neonatal, pubertal, and adult rats, Chem. Biol. Interact. 139(2002) 23–41.
56] D.J. Waxman, Interactions of hepatic cytochromes P-450 withsteroid hormones: regioselectivity and stereoselectivity of steroidmetabolism and hormonal regulation of rat P-450 enzyme expres-sion, Biochem. Pharmacol. 37 (1988) 71–84.
57] M.K. Lindberg, S. Moverare, S. Skrtic, H. Gao, K. Dahlman-Wright, J.A. Gustafsson, C. Ohlsson, Estrogen receptor (ER)-betareduces ERalpha-regulated gene transcription, supporting a “yingyang” relationship between ERalpha and ERbeta in mice, Mol.Endocrinol. 17 (2003) 203–208.
58] J.M. Emmen, J.F. Couse, S.A. Elmore, M.M. Yates, G.E. Kissling,K.S. Korach, In vitro growth and ovulation of follicles fromovaries of estrogen receptor (ER){alpha} and ER{beta} null miceindicate a role for ER� in follicular maturation, Endocrinology146 (2005) 2817–2826.
59] Y. Omoto, R. Lathe, M. Warner, J.A. Gustafsson, Early onsetof puberty and early ovarian failure in CYP7B1 knockout mice,Proc. Natl. Acad. Sci. U.S.A. 102 (2005) 2814–2819.
60] E. Anderson, R.B. Clarke, Steroid receptors and cell cycle innormal mammary epithelium, J. Mammary Gland Biol. Neoplasia9 (2004) 3–13.
61] V. Panet-Raymond, B. Gottlieb, L.K. Beitel, L. Pinsky, M.A.Trifiro, Interactions between androgen and estrogen receptorsand the effects on their transactivational properties, Mol. Cell.Endocrinol. 167 (2000) 139–150.
62] A. Brzezinski, A. Debi, Phytoestrogens: the “natural” selectiveestrogen receptor modulators? Eur. J. Obstet. Gynecol. Reprod.Biol. 85 (1999) 47–51.
63] W.N. Jefferson, J.F. Couse, E. Padilla-Banks, K.S. Korach, R.R.Newbold, Neonatal exposure to genistein induces estrogen recep-tor (ER)alpha expression and multioocyte follicles in the maturing
mouse ovary: evidence for ERbeta-mediated and nonestrogenicactions, Biol. Reprod. 67 (2002) 1285–1296.64] T. Ito, M. Tachibana, S. Yamamoto, J. Nakashima, M. Murai,Expression of estrogen receptor (ER-alpha and ER-beta) mRNAin human prostate cancer, Eur. Urol. 40 (2001) 557–563.

iologica
[
[
551.
14 F. Maranghi et al. / Chemico-B
65] R.S. Bhattacharyya, A.V. Krishnan, S. Swami, D. Feldman,Fulvestrant (ICI 182,780) down-regulates androgen receptor
expression and diminishes androgenic responses in LNCaPhuman prostate cancer cells, Mol. Cancer Ther. 5 (2006)1539–1549.66] A. Bardin, N. Boulle, G. Lazennec, F. Mignon, P. Pujol, Lossof ERbeta expression as a common step in estrogen-dependent
[
l Interactions 169 (2007) 1–14
tumor progression, Endocr. Relat. Cancer 11 (2004) 537–
67] R. Ceccatelli, O. Faass, M. Schlumpf, W. Lichtensteiger, Geneexpression and estrogen sensitivity in rat uterus after develop-mental exposure to the polybrominated diphenylether PBDE 99and PCB, Toxicology 220 (2006) 104–116.





![Saugu[!] Asmundar, er kalladur er Kappabani :](https://static.fdokumen.com/doc/165x107/63264a17051fac18490dae0e/saugu-asmundar-er-kalladur-er-kappabani-.jpg)














