
Interleukin1β mediates GDNF up-regulation upon dopaminergic injury in ventral midbrain cell cultures

JOURNAL OF NEUROBIOLOGY, VOL. 6, NO. 1, PP. 3-22
OCULOMOTOR AREAS IN THE RABBIT’S MIDBRAlN AND PRETECTUM
H. COLLEWIJN
Department of Physiology, Medical Fucult.y, Erasmus Unixrs i ty , Rotterdam. The Netherlands
SUMMARY
The role of some meso- and diencephalic structures in eye movements was investigated by ablation and stimulation experiments. Optokinetic nystagmus was abolished by small lesions in the lateral pretectum, but not by complete removal of the superior colliculi. Stimulation of the superior colliculus and other visual centers was effective in eliciting nystagmus (slow phase ipsilateral) , but the most efficient trigger zones are found in the lateral pretectum and the midbrain tegmentum. Only from these areas could nystagmus still be elicited after degeneration of the primary optic fibers. The lateral pretectal trigger zone is probably identical with the nucleus of the optic tract. It is postulated that this nucleus is an essential station for horizontal optokinetic reactions. Saccades were obtained by stimulation of the mesencephalic central grey, but not for any visual centers such as the superior colliculus.
INTRODUCTION
In recent years, several investigators have demonstrated a subtle mechanism for the generation of fast eye movements (saccades) in the superior colliculus of the monkey (Robinson, 1972; Schiller and Stryker, 1972). Brief, low intensity stimulation of a point in the superior col- liculus induced a fast eye movement that aimed the fovea at that point of the visual field represented by the retinotopical coordinates of the stimulated site. Typically, the size and direction of these movements were independent of initial eye position and fully determined by the position of the stimulus. Thus this fixation or “foveation” system operates in a coordinate system locked to visual space and not to the head. This concept has been strengthened by the findings of appro- priately located single unit discharges prior to saccades in the deeper collicular layers (Wurtz and Goldberg, 1972).
In the cat, a somewhat similar but less well developed system has been demonstrated by Straschill and Rieger (1973). In the cat, the end point of the saccade appears to be fixed relative to the head rather than to visual space. As pointed out by Straschill and Rieger, such a reference system is more appropriate to auditory than to visual cues, although the visual and auditory perceptual space can be brought in functional register
3 3 1975 by John Wiley & Sons, Inc.

4 COLLEWIJN
if the cat usually keeps his eyes in the primary position. Differences in the visual system and behavior of cat and monkey may make these differences undei-standable. The monkey is a foveate animal with a highly developed saccadic scanning behavior, like man. The cat is essentially a noc.turna1 animal with a poorly developed central area; precisely directed visual searching, independent of head movements, may not be of primary importance to this animal.
The question arose whether anything like a fixation mechanism could be demonstrated by collicular stimulation in the rabbit. This animal’s visual system is organized much more diffusely than that of the cat. The rabbit’s retina contains a n elongated “visual streak” which is roughly aligned with the horizon; the density of visual and neuronal elements is considerably higher there than in the periphery (Hughes, 1971). Such a n organization suggests a relatively low need for precisely organized search movements, and a dominance of reflectory stabilizing systems, such as vestibulo-ocular and op tokinetic reflexes. The latter have been investi- gated in detail in recent years (Collewijn, 1969, 1972a,b). Spontaneous eye movements, not combined with head movements, are rarely seen in the rabbit, although they may occur (Hughes, 1971).
The rabbit’s optokinetic system stabilizes the eyes on the resting (or moving) surroundings as a whole. It has no role in the visual pursuit of isolated moving objects; this latter type of oculomotor behavior cannot be demonstrated in the rabbil. Some evidence has been provided for the hypothesis that direction selective retinal units are involved in this reflex (Oyster, Takahashi, and Collewijn, 1972). However, the pathway of the optokinet ic reflex is still obscure. Earlier assertions concerning the role of the superior colliculus (Hobbelen and Collewijn, 1971) were based upon lesions immediately prior to the measurements; therefore the functional extent of these lesions may have been underestimated.
On the basis of degeneration studies of primary optic fibers in the rabbit (Giolli and Guthrie, 1969; Scalia, 1972) the following main path- ways may be distinguished: ( I ) geniculo-cortical; 12) superior collicular; (3) pretectal, and (4) accessory.
I n accordance with earlier investigations, Hobbeleii and Collewijn (1971) conclusively demonstrated that even immediately after massive decortication optokinetic reactions in the rabbit are perfectly normal. Therefore, one or more of the remaining three pathways must mediate this reflex.
The present work, consisting of stimulation and ablation experiments, concerned itself with the role of the subcortical visual projections in the rabbit in optokinelic or any other oculomotor phenomena.
METHODS
Animal prepurcction
Young adult Dutch belted rahhits (weight about 2 kg) were used. Under halothane (Fluothane I.C.I.) anesthesia, the trachea was cannulated, a dental bite board was

RABBIT OCULOMOTOR AREAS 5
fixed with acrylic to the upper jaw and a craniotomy was done. The dural memhrane was opened and the midbrain and caudal diencephalon carefully exposed hy removing most o t the hemispheres, using suction. The brainstem was covered with paraffin oil. A scleral induction coil was sutured to the limbus of the right eye. The spinal cord was sectioned at C, and artificial ventilation was provided. A t this stage, anesthesia was terminated. The head was fixed to a stereotaxic apparatus, using the bite hoard, which was always inclined 30" below the horizontal. The enc6phale isole preparation was kept a t a temperature of 38 & 0.5"C. All cerebral surgery and electrode placement was done using a Zeiss surgical microscope. The preparation usually remained in good condition during the whole day of the experiment, at the end of which a n overdose of barbiturate was administered, after which the brain was fixated with formaldehyde by perfusion from the left ventricle.
Collicu7ar ablation
In a number of animals, complete removal of both superior colliculi was attempted under aseptic conditions, several months prior to the final measurements. Under halo- thane anesthesia, the skull was opened. The dura was incised parasagittally and re- flected. Major branches to the sagittal venous sinus were coagulated by heat and divided. Thereafter, the sagittal sinus between the coronal suture and the lambda was removed. The hemispheres were separated as gently as possible and the superior collic- ulus exposed. Significant bleeding did usually not occur and the superior colliculus could be completely removed & uue. Pretectal area and optic tract were carefully spared, although damage to the caudal pretectum could not be completely avoided, since it extends below the colliculus. The hemispheres were replaced and covered by a piece of gel foam. Recovery was usually prompt and uneventful. The skin was closed.
Optic nerve sectioning
I n a number of animals, one or both optic nerves were sectioned 8-30 days before the The optic nerves were approached extradurally through a frontal
Under direct visual control, one or hoth nerves Such animals were of course uni-
Recovery
final experiment. craniotomy, by gently lifting the brain. were divided completely, rostra1 to the chiasma. laterally or completely blind, but their orbital contents were undamaged. was usually undisturbed.
E y e movement recording
The scleral coil method was used, which has been described before (Collewijn, 1969). Provisions were made for the simultaneous measurement of horizontal and vertical eye movements, which were written on a Grass Polygraph (Model 7) pen recorder.
Stimulation
Optokinetic reactions were elicited by rotating a striped drum around the animal, as has been described before (Collewijn, 1969).
Electrical stimuli were generated by a constant current stimulator (Nuclear Chicago Model 7152) and in most cases administered through monopolar stainless steel electrodes (tip about 0.5 m m long and 0.2 m m thick), which were lowered into the brain using a n stereotaxic electrode carrier, under visual control from a surgical microscope. T h e electrode was always a cathode, while the dental bite board served as the positive reference electrode. Typically, trains were delivered of 20 impulses (2 msec duration) per second, with a n intensity of 0.1 mA. For marking purposes, the polarity was in- verted and a positive current of 1 mA dc was passed during 10 sec.
Histology
All brains of interest were serially sectioned and stained by the Kluver-Rarera T h e method. Transversal sections were prepared, parallel to the electrode tracks.

6 COLLEWIJN
stained sections were projected on a screen. The stimulation sites were reconstructed from the visible electrode tracks arid electrolytical marks anti transferred onto standard drawings. In general, the nomenclature of Gerhard (1968) was followed, except in case of the pretectal nuclei, for which the naming by Giolli and Guthrie (1969) and Scalia (19721 was preferred. In particular, the “posterin, thalamic nucleus” of Gerhard is named the anterior pretectal nucleus. However, the “lateral Imsterior nucleus” of Gerhard and Scalia is recognized as such and it is not named “pulvinar,” as done by Giolli and Guthrie. (For a discussion of the nomenclature, see Stewart, Towns and Birt (1973).)
RESULTS
Optokinetic reactions after collicular ablation
Nine rabbits with intended complete bilateral removal of the superior colliculus promptly recovered from surgery and survived for several months. Within 24 hr the animals were behaving and eating normally; posture and locomotion were undisturbed. The only apparent neu- rological defect consisted usually of wide pupils, which were unresponsive to light. A simple visual discrimination task (horizontal vs. vertical stripes) was easily learned by all animals (Van Hof and Collewijn, in preparation).
Optokinetic reactions in these animals were measured about 10 months after the collicular ablation. All animals were in excellent condition a t that time. All of them showed positive and practically normal opto- kinetic reactions, except for one animal which developed a vigorous spontaneous nystagmus (slow phase to the left) which could not be in- fluenced by optical stimuli. This animal had shown episodes of running in circles to the left side. I n some animals, after the presence of opto- kinetic reactions had been confirmed, a n additional removal of the hemi- spheres was performed. The optokinetic reactions were not abolished or diminished by this procedure.
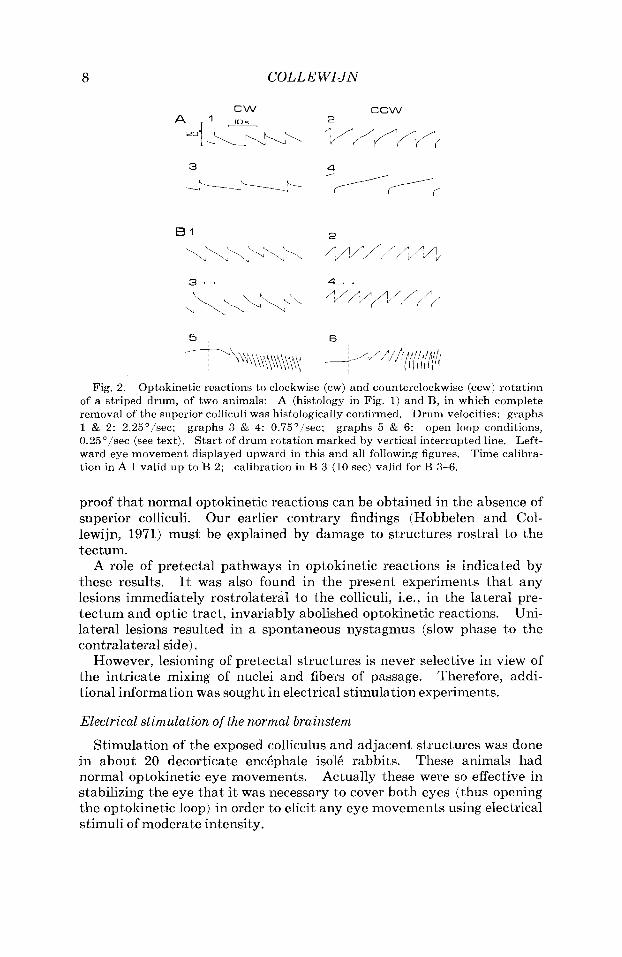
Histological analysis showed that in the animal with spontaneous nystagmus the lesion on the right side extended forward into the optic tract. I n the other animals, optic tract and pretectum were largely in- tact. Some damage to the medial edge of the hemispheres was usually evident. The superior colliculus was almost totally removed, but in most animals some small rests (usually at the latero-ventral side) were still p~esent . Only in two rabbits the collicular lesion proved to be com- plete. Figure 2 shows the optokinetic reactions of this animal (A) and the other one with a complete lesion (B).
To further confirm that the optokinetic system was indeed functioning normally, “open loop” conditions were tested (Fig. 2B). In this situa- tion, the left eye is seeing but immobilized, while the right eye can move but is covered (Collewijn, 1969). Under these conditions, slow drum movemeiits (e.g. 0.25’1sec) elicit a vigorous nystagmus of the right eye with a slow phase velocity many times higher than the stimulus velocity, which is a measure of the open loop gain of the system (Collewijn, 1969).
No spontaneous nystagmus was ever seen.
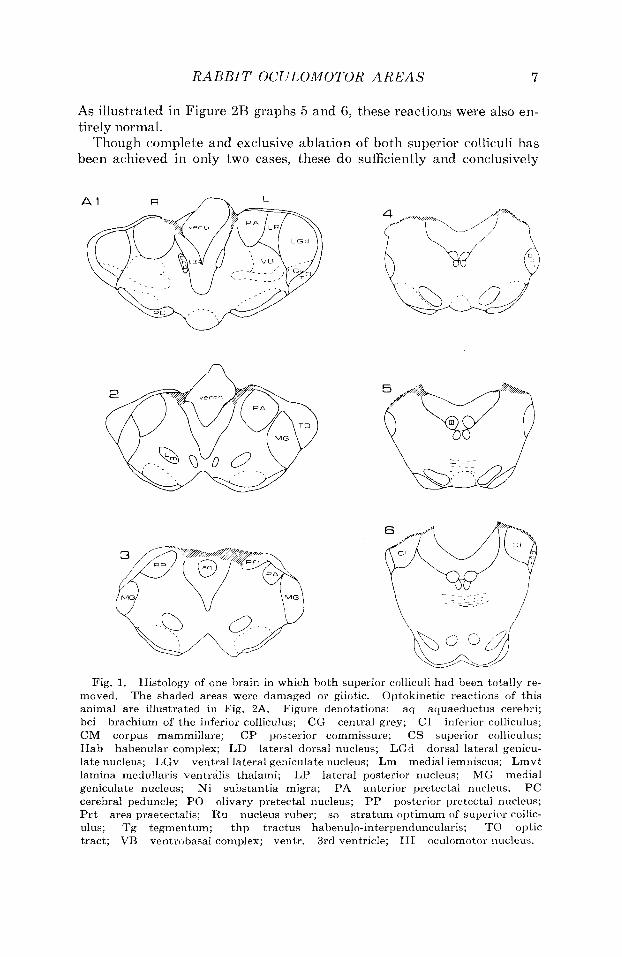
The histology of one of these is illustrated in Figure 1.
R A B B I T OCULOMOTOR A R E A S 7
As illustrated in Figure 2R graphs 5 and 6, these reactions were also en- tirely normal.
Though complete and exclusive ablation of' both superior colliculi has been achieved in only two cases, these do sufficiently and conclusively
Fig, 1. Histology of one brain in which both superior colliculi had been totally re- moved. Optokinetic reactions of this animal are illustrated in Fig. 2A. Figure denotations: aq aquaeductus cerebri; bci brachium of the inferior colliculus; CG central grey; CI inferior colhculus; CM corpus mammillare; CP posterior commissure; CS superior colliculus; Hab habenular complex; LD lateral dorsal nucleus; LGd dorsal lateral genicu- late nucleus; LGv ventral lateral geniculate nucleus; Lm medial lemniscus; Lmvt lamina medullaris ventralis thalami; L P lateral posterior nucleus; MG medial geniculate nucleus; Ni substaiitia migra; PA anterior pretectal nucleus; PC cerebral peduncle; PO olivary pretectal nucleus; PP posterior pretectal nucleus; P r t area praetectalis; Ru nucleus ruber; so stratum optimum of superior collic- ulus; Tg tegmentum; thp tractus habenulo-interpenduncularis; TO optic tract; VB ventrobasal complex; ventr. 3rd ventricle; 111 oculomotor nucleus.
The shaded areas were damaged or gliotic.
8 COLLE WIJN
B1
3 -
5
C C W 2
4
6
Fig. 2. Optokinetic reactions to clockwise (cw) and counterclockwise (ccw) rotation of a striped drum, of two animals: A (histology in Fig. 1) and B, in which complete removal of the superior colliculi was histologically confirmed. Drum velocities: graphs 1 & 2: 2.25"/sec; open loop conditions, 0.25'/sec (see text). Start of drum rotation marked by vertical interrupted line. Left- ward eye movement displayed upward in this and all following figures. Time calibra- tion in A 1 valid up to B 2; calibration in B 3 (10 sec) valid for B 3-6.
graphs 3 & 4: 0.75"/sec; graphs 5 & 6:
proof that normal optokinetic reactions can be obtained in the absence of superior colliculj. Our earlier contrary findings (Hobbelen and Col- lewijn, 1971) must be explained by damage to structures rostra1 to the tec tum.
A role of pretectal pathways in optokinetic reactions is indicated by these results. It was also found in the present experiments that any lesions immediately rostrolateral to the colliculi, i.e., in the lateral pre- tectum and optic tract, invariably abolished optokinetic reactions. Uni- lateral lesions resulted in a spontaneous nystagmus (slow phase to the contralateral side).
However, lesioning of pretectal structures is never selective in view of the intricate mixing of nuclei and fibers of passage. Therefore, addi- tional information was sought in electrical stimulation experiments.
Electrical stimulation of the normal brainstem
Stimulation of the exposed colliculus and adjacent structures was done in about 20 decorticate enckphale isolk rabbits. These animals had normal optokinetic eye movements. Actually these were so effective in stabilizing the eye that it was necessary to cover both eyes (thus opening the optokinetic loop) in order to elicit any eye movements using electrical stimuli of moderate intensity.

RABBIT OCULOMOTOR AREAS 9
13 ,----,/ill;
L4
Fig. 3. Typical horizontal eye movements elicited by electrical stimulation of the The inset shows the stimulus positions in the colliculus, seen
20 pulses (2 msec duration' Depth of stim-
left superior colliculus. from ahove (rostra1 edge upward). Stimulus parameters: per second, intensity 0.1 mA, train duration marked by horizontal bars. ulus electrode about 1 mm.
In preliminary experiments, stimulus trains containing about 20 im- pulses (2 msec duration) per second with an intensity of 0.1 mA proved to be usually quite effective, and sometimes even much lower intensities could be used. Brief stimulus trains (shorter than 1 sec) had very little effect and certainly did not systematically elicit saccades. Trains with a duration of many seconds gave typical reac tioiis.
The dominating part of the response was a slow conjugate horizontal ipsilateral deviation, interrupted by contralateral fast movements, which together formed a nystagmus. These reactions are illustrated in Figure 3. Apart from the direction of the nystagmus the responses show little specificity for electrode position. It was found that the threshold was usually minimal about 1 mm below the collicular surface. Stimulation of

10 COLLEWIJN
Fig. 4. Same animal as in Fig. 3. Simultaneous horizontal (Hor.) and vertical (Vert.) responses elicited by collicular stimulation a t position 13. Upper trace: stimu- lus train duration: second trace: time in seconds.
the caudal area of the superior colliculus were always less effective than stimulation rostrally. It was consistently found that a site a t the rostra1 collicular boundary, 4-5 mm lateral from the midline was much more sen- sitive and effective than any other location (Fig. 3). From this site, nystagmus could be easily elicited even when the eyes were opened. Thus, the effect was so strong that it could not be blocked by optokinetic stabilization. The meaning of this finding will be discussed later. In all cases, the primary response to stimulation was a slow deviation. A fast phase was only triggered as a reset of the slow movement. Isolated saccades, unaccompanied by a slow deviation, were not obtained by collicular stimulation.
A second component of the response, illustrated in Figure 4, consisted of vertical eye movements. These were clearly very different in nature from the horizontal movements. The vertical deviations were not nys- tagmic, but rather steady and they tended to move the eye to a certain end position, which was somewhat specific for the transversal coordinates of the stimulus electrode. During stimulation of medial collicular areas the contralateral eye moved upward and the ipsilateral eye downward. From the lateral parts of the superior colliculus, opposite reactions were elicited (Fig. 5). In view of the projection of the visual field upon the superior colliculus (Hughes, 1971) this means that the (horizontal) visual streak of the retina was moved towards the area of the visual field corre- sponding to the stimulus site in the colliculus. Thus, the direction of these vertical movements is related to those found in the monkey, but they are extremely slow in comparison and it remains to be determined whether this finding has behavioral significance.
As can be seen in Figure 3, the slow phase velocity (and therefore the frequency of beats) of the evoked nystagmus gradually built up to a max- imum, which could be maintained indefinitely by continued stimulation. Similar effects are known in optokinetic reactions (see Fig. 2R, graphs 5-6). The intensity of this steady state (expressed in slow phase veloc- ity) was dependent upon the stimulus intensity. However, if the fre- quency and/or current strength of the stimulus train was increased be- yond a certain optimum (usually around 0.2 mA, 40 'sec) the response

RARHIT OCULOMOTOR AREAS 11
10.sc
6
I L . l 1- Y
I I
A
+ I , 5 ---- r-i- r-
Fig. 5. Vertical eye movements as a function of stimulus location. The horizontal stimulus positions in superior colliculi seen bars mark the stimulus duration.
from above; rostra1 edge upward. Inset:
changed into a tonic deviation of erratic character without fast reset movements. With still higher intensities, even ipsilaterslly directed saccades were sometimes elicited. In view of further findings these re- sults are considered to be due to overstimulation and current spread.
After termination of the stimulus, the horizontal nystagmus and the vertical displacement subsided gradually. A rebound effect (nystagmus in opposite direction) was sometimes seen.
The results of collicular siimulatioii in the rabbit thus did not reveal a fixation system such as found in the monkey and to a certain degree in the cat, with the possible exception of a weak vertical effect.
Horizontal nystagmus such as described above has been elicited in rabbits before by many authors from many structures, iiicludiiig the superior colliculus. I explored a number of locations and can confirm that in the normal braiiistem similar horizontal nystagmus (slow phase ipsilateral) can be elicited a t least from the following structures: superior colliculus, lateral pretecium, optic tract, lateral geiiiculate nucleus. However, since all these structures are pervaded by primary optic

12 COLL E W I J N
afferent fibers, the question arises whether the effects are really specific for these nuclei.
To obtain a straightforward answer to this question, it was decided to repeat the stirnulation experiments and to explore the brainstem sys- tematically in depth in animals of which one or both optic nerves had been sectioned before. The resulting degeneration of all primary fibers will abolish nonspecific effects due to stimulation of these fibers.
Stimulation after sectioning one optic nerve
Successful stimulation experiments were done in 9 rabbits, 8 30 days after cutting the right optic nerve intracranially. Of these animals, the right eye was blind, according to the criteria of pupillary reflexes, flash- evoked cortical and collicular potentials, op tokinetic stimulation, and be- havioral responses. Optokinetic stimulation of the left eye caused con- jugate optokinetic responses of both eyes. Apart from this, the animals behaved normally. Circling or spontaneous nystagmus was at no time observed.
Subsequent histological inspection showed massive desintegration of the fibers of the left optic tract. (About 90% of the optic nerve fibers crosses in the chiasma of pigmented rabbit; Giolli and Guthrie, (1969).)
Responses to superior collicular stimulation in these animals proved to be altered profoundly. Whereas from the right (normal) colliculus the usual response pattern was obtained, stimulation of the left (de-affer- ented) colliculus was quite ineffective. Responses were very small, badly reproducible or entirely absent (Fig. 6). The only exception was a ct .... a
bri 1. 2.
......... ~ . . ~
L4
1 8 * , i i i '
-\------- ~ \ \ ' .-- i-
.dL -__ at lDses
Fig. 6 . Eye movements elicited from the left (de-afferented) and right (normal) superior colliculus, 30 days after section of the right optic nerve. The insets show loca- tions of the stimuli in the colliculus seen from above (right) and in a transversal section. Position 9 is shown to he inside the left optic .tract. (See Fig. 1 for abbreviation key.)

R A B B I T OClJLOMOTOR A R E A S 13
Fig. 7. Transversal sections through left half of the brainstem, 5 10 mm behind the Pooled results of stimulation in 9 rabbits, 8-30 days after transection
(0 ) none, (+) weak (up to 5',/sec), (See Fig. 1 for abbreviation key.)
coronal suture. of the right optic nerve. (0) intermediate (5-2O0/sec), (@) strong (20°/sec.) .
Coding of the effects:
small area near the rostrolateral collicular edge, which remained always responsive after de-afferentiation.
A systematic exploration of the left (de-afferented) meso-diencephalic area gave the following results (Fig. 7). No systematic oculomotor re- sponses were elicited from the lateral and medial geniculate nuclei, the lateral posterior nucleus, the medial pretectal area, the olivary, anterior- and posterior pretectal nuclei, the ventrobasal complex and the superior colliculus, except its rostra1 edge. Very intense nystagmus (slow phase in steady state faster than 20"/sec) was elicited from two areas:
(1) the optic tract dorsal to the anterior pretectal nucleus, (2) the midbrain tegmentum.
The sensitive site in the optic tract corresponds to the location of the nucleus of the optic tract, which receives a dense terminal projection of optic tract fibers (see Scalia, 1972). Since all crossed primary optic fibers in these optic tracts were presumably degenerated, these results may be explained by stimulation of the cell bodies of the nucleus of the optic tract (which were still clearly recognizable amid the degenerating fibers) or their efferent fibers. The nucleus of the optic tract is situated

14 COLL E WIJN
Fig. 5. Effect of stimulation of the superior colliculus, 1 2 days after transection of From the colliculus proper (sites 3- 61 weak effects are obtained,
From the These
(See Fig. 1 for
both optic nerves. which in this case are inverted in polariiy in comparison to normal animals. rostrolateral edges (sites 1 and 2) the normal, vigorous responses are obtained. locations were inside or very close to the optic tract (inset left above). ahbreviation key.)
very close to the rostrolateral edge of the superior colliculus. This rela- tionship accoun I s for the apparent high sensitivity of this collicular area.
The midbrain tegmentum does not, receive primary optic fibers but has been often considered a premotor area for eye movements (Komatsuzaki, Alpert, Harris and Cohen, 1972).
The change in effects when Ihe stimulus electrode was moved, indi- cated that the error due to current spread (at 0.1 mA) was maximally 1 mm. I n view of the coiisistency of the location in 9 rabbits (Fig. 7) , the reconstruction of the stimulus sites apparently introduced no wider spread.
I n regions close to the two maximally effective areas, weak or interme- diate responses were often obtained. All responses were nystagmic in nature, with the slow phase directed ipsilaterally.

RABBIT OCULOMOTOR AREAS 15
Stimulation after sectioning both optic nerves Similar stimulation experiments were performed in a number of bi-
laterally blinded animals. These rabbits were blind according to all criteria. They remained in good coiidition, but had to be handled care- fully since they panicked easily when they were touched.
Responses elicited by collicular stimulation were weak, absent or ab- normal in direction (Fig. 8). Again, strongly positive responses were ob- tained in a small area immediately rostrolateral to the colliculus.
Systematic exploration in depth in 3 animals showed that the position with the lowest threshold and the highest effect was again located inside the optic tract, dorsal to the anterior pretectal nucleus. Also from the midbrain tegmentum strong responses were obtained. All other struc- tures named so far were unresponsive. These results are in complete agreement with those after unilateral optic nerve section.
Eliciting of pure saccades None of the areas mentioned so far in the normal or optically de-
affereiited brain was effective in eliciting pure saccades, other than as fast phases of a nystagmus. However, i t was found that saccadic reac- tions could be easily and consistently obtained by stimulation close to the midline. As shown in Figure 9, saccades were elicited from this area by brief (e.g., 200 msec) stimulus trains. The latency was about 200 msec. A rebound saccade usually followed termination of the stimulus. Longer stimulus trains resulted in a sequence of saccades in the same direction with an interval of about 0.5 1 sec. These reactions could be obtained as easily with the eyes open (Fig. 9C,E,F) as with eyes closed (Fig. 9A,B,D). The direction of the saccades was always ipsilateral to the stimulus.
Histological reconstruct ion of stimulus positions that elicited saccades (3 animals pooled) showed that all these positions were localized within or very close to the central mesencephalic grey (Fig. 10).
Fig. 9. Saccades elicited by stimulation of the midbrain close to the midline (see Fig. 10 for histology). Stimulus iiidicated by horizontal bars: A, R, C, 40 'sec, 0.1 mA; D, E, F 20 sec, 0.1 mA. A, R, D eyes closed; C, E, F eyes open. A-E were elicited from the same stimulus position, F from a different one.

16 COLL E WIJN
Fig. 10. were elicited. saccadic responses.
Histological reconstruction of stimulus positions ( + ) from where saccades (0 ) no They are all localized within or very close to the central grey.
Pooled results of 3 rabbits.
A systematic investigation of the influence of stimulus parameters and position within the central grey upon size and direction of the saccades has yet to be completed.
DISCUSSION
Role of superior colliculus in eye movements
The present findings indicate that the function of the superior col- liculus in the rabbit’s eye movements is quite limited. It has been shown that normal optokinetic reactions can still be obtained after complete re- moval of the colliculi. Earlier investigators have reached similar con- clusions, but usually on the basis of incomplete lesions. Proctor (1962) mentions positive optokinetic reactions after a 25 % unilateral removal of the superior colliculus in one rabbit, and 85% unilateral removal in one cat. Also Hobbelen and Collewijn (1971) found that partial lesions of the superior colliculi in the rabbit left optokinetic reactions largely intact.

RABBIT 0C:JLOMOTOR AREAS 17
Pasik, Pasik and Bender (1966) reviewed the then existing evidence and concluded that it was not conclusive for considering the superior col- liculus a center for optokinetic or other eye movements. The present re- sults confirm that the colliculus is not required €or optokinetic reactions in the rabbit.
However, a n area very close to the rostrolateral margin of the col- liculus appears to be essential in optokinetic reactions. This area was un- doubtedly damaged in the experiments of Hobbelen and Collewijn (1971). In the present experiments it was invariably found that even slight damage in this zone abolishes optokinetic reactions with the slow phase to the ipsilateral side and leads to a spontaneous iiystagmus with the slow phase to the contralateral side. However, such lesions inevitably damage many fibers in the optic tract and the brachium of the superior colliculus as well as pretectal structures and therefore do not lead to very specific conclusions.
Electrical stimulation of the superior colliculus did elicit nystagmic eye movements, in agreement with earlier findings in the literature. Especially the report of Blohmke (1929) proved to be remarkably de- tailed; this author described the same horizontal nystagmus and the tonic vertical reactions. He also found that the tecto-thalamic transi- tion zone was the most effective stimulus site. Arimoto (1958) also re- ported that “tectal nystagmus” was elicited most easily from a location rostrolateral to the superior colliculus. Bergmann, Costin, Gutman and Chaimovitz (1964) emphasize that they could obtain nystagmus (fast phase contralateral) by collicular stimulation only in darkness; as should be expected in view of our present understanding of optokinetic reflexes as a stabilizing visual feedback system.
The present findings that optic de-afferen tiation severely diminishes or abolishes the oculomot or effects of collicular stimulation can be inter- preted in tvio ways.
The first possibility is that the colliculus contains no true efferent oculomotor fibers and that by collicular stimulation other centers are ex- cited by antidromic collateral stimulation of primary optic fibers. The concept that optic fibers may bifurcate and terminate in several end stations has received considerable support recently, both anatomically (Giolli and Guthrie, 1969) and electrophysiologically (Hoffmann, 1973). This means that fibers from direction selective retinal ganglion cells (as- suming that these are involved in optokinetic reactions; see Oyster et al. 1972) could conceivably terminate as well in the colliculus, in certain pre- tectal nuclei and in the lateral geniculate nucleus. Optic nerve section would abolish these collateral reactions. Indeed it does abolish re- sponses from both colliculus and lateral geniculate nucleus, but apparently not from the nucleus of the optic tract. The detrimental effect of optic nerve section upon collicular excitability was noticed for horizontal nys- tagmus as well as the vertical tonic deviations. The few remaining

18 COLLE W I J N
reactions were disorganized and stereotyped; often a similar reaction was obtained from any place in the right as well as the left colliculue. De- afferentiation certainly did not uncover any subtle collicular fixation mechanism, which in the normal animal might have been masked by nystagmic reaction to stimulation.
A second possibility is that the colliculus normally does process visual information into oculomotor commands, but that this system is dis- organized by de-afferentiation in such a way that it can no longer be activated by electrical stimulation. Since the oculomotor effects of collicular stimulation in the monkey are so highly specific, it would be of great interest if the effect of optic de-afferentiation were tested in that animal.
Nystagmogen ic zones
The concept of collateral stimulation fits well with earlier findings that horizontal nystagmus can be obtained by stimulating anywhere along the rabbit’s visual pathway, a finding formulated by Gutman, Bergmann, Chaimovitz and Costin (1963). These authors found that stimulation of optic fibers from the right eye led to nystagmus with the slow phase to the left, and vice versa. This is in accordance with the directional preference for optokinetic stimulation in the temporo-nasal direction of each eye (Collewijn, 1969). This generalization seems to deprive the original finding of a “nystagmogenic area’’ in the meso-diencephalon by Lach- mann, Bergmann and Monnier (1958) of its specificity. The localization of this nystagmogenic area, in subsequent publications of these investi- gators, has remained somewhat problematic. Bergmann, Chaimovitz, Gutman and Zelig (1963) stated that the region of greatest sensitivity was located a t a distance of 7.5 mm caudal to the coronal suture and 4.5 mm from the midline. Although the depth was not mentioned, these hori- zontal coordinates exactly agree with the present findings for the nucleus of the optic tract. Costin, Bergmann, Chaimovitz and Ginath (1966) recognized that the lateral geniculate nucleus and optic radiation were the most sensitive structures. On the other hand, Monnier (1967) has claimed a localization in the intralaminary system, medial to the lateral geniculate nucleus. Many of these controversial findings are, apart from collateral effects, probably due to current spread, since current intensity was not controlled in those experiments.
In the light of the present evidence it seems likely that a genuine nystagmogenic area may be present after all in the nucleus of the optic tracl, since in generating nystagmus it is clearly the most effective site and its responsiveness survives optic de-afferentiation. In a few preliminary experiments, I found that horizontal nystagmus from this site can still be elicited several months after bilateral collicular ablation (90 % complete) combined with bilateral optic nerve section, which would indi- cate that collicular efferents are not involved in this effect.

RABBIT OCULOMOTOR AREAS 19
The optokinetic pathway
The sum of the present evidence would then suggest the following path- way for horizontal optokinetic reactions. Primary optic nerve fibers (possibly specifically direction selective fibers) terminate in the nucleus of the optic tract. Collaterals of these fibers may end in the colliculus and other stations. From the nucleus of the optic tract, secondary (truly oculomotor) fibers may pass to the next station. The efferent connections of the nucleus of the optic tract are not yet known, but with some bold speculation one might conceive that the midbrain tegmentum might be among them. The present findings indicate that this structure can gen- erate nystagmus of the same type as the nucleus of the optic tract. Komatsuzaki et al. (1972) found that lesions in the mesencephalic reticu- lar formation strongly interfered with optokinetic reactions in the mon- key. They suppose that this area is especially involved in generating the slow phase to the ipsilateral side; this suggestion is in agreement with the present results. It should be emphasized that none of the stimulation sites used in this study and others have elicited a vertical or rotatory nystagmus, although optokinetic reactions in these directions are well developed (Collewijn and Noorduin, 1972). It may be that these are mediated through other pathways. Another possibility is that elements involved in upward and downward eye movements are thoroughly mixed and balanced so that they are not revealed by the present (rather crude) stimulation. The vertical movements that were obtained were mostly tonic and not nystagmic.
Not only the anatomical connections, but also the neurophysiological properties of units in the nucleus of the optic tract are still unknown. Recently Stewart, Towns and Birt (1973) reported on the visual receptive field characteristics of the other pretectal fiuclei which are known to receive optic afferents: the anterior and posterior pretectal nuclei. It may be of some significance that the only place where they found direc- tion selective units in abundance was the dorsal part of the posterior pretectal nucleus (named by them pretectal nucleus). This location is very close to the lowest threshold site for eye movements, the nucleus of the optic tract, which was not specifically included in their investi- gation. A finding of a large concentration of direction selective units in the nucleus of the optic tract would obviously strengthen the present hypothesis for the optokinetic pathway very strongly.
Possible role of accessory optic tract
A role of the accessory optic fiber system in eye movements has been repeatedly suggested. I n the rabbit, the transpeduncular tract emerges from the brachium of the superior colliculus and courses ventrally over the surface of the cerebral peduncle to terminate in the nucleus of the peduncular tract (Giolli, 1961). It runs close to the best trigger zone for nystagmus. However, Marg (1964) states that stimulation of the nu-

20 COLLEWIJN
cleus of the peduncular tract in undrugged animals did not produce specific eye movements. Recently, LAzBr (1973) reported that lesions of the basal optic root, the homologue of this tract in the frog, specifically abolished optokinetic reactions. The present findings of a trigger zone in the optic tract as well as the vulnerability of optokinetic reactions to lesions a t this site are not incompatable with a mediation of optokinetic reflexes through the transpeduncular tract. However, preliminary stimulation experiments a t the ventrolateral side of the meso-diencephalic area, where the transpeduncular tract is running, never elicited any eye movement. More significant is the finding that stimulation a t the site of the optic tract nucleus was still equally effective after sectioning of the optic nerves. Supposedly, the transpeduncular tract was totally de- generated in these animals. Selective destruction of the accessory optic system is hard to achieve, and IAzAr’s report suggests that other parts of the visual pathway were destroyed as well in his animals following basal root lesions, since they seemed to be completely blind and the pupils were unresponsive to light. In other experiments, LBzBr destroyed the nucleus of the accessory optic tract and found that optokinelic reactions were abolished by this procedure. However, it is not clear how much damage was done to the midbrain tegmentum by these lesions.
It must be concluded that there is no firm evidence for a role of the accessory optic system in optomotor reactions.
Generation of pure saccades
A quite unexpected finding was a trigger zone for saccadic (ipsilateral) eye movements in the mesencephalic grey. This area has not been specifically implicated before in eye movements, although Render and Shanzer (1964) include i t among the many structures that yielded con- jugate eye movements upon stimulation in alert monkeys. The present results are preliminary and do not permit conclusioiis about the nature of these fast movements. A role in a fixation mechanism appears unlikely since the central grey is not a visual projection center. Possibly, this area is involved in the generation of the fast phase of nystagmus, about which we know very little. The finding that they were usually followed by a re- bound saccade after termination of the stimulus (Fig. 9) is in contrast with the results in the monkey’s colliculus (Robinson, 1972) and may also argue against a visually guided system. The only other known place in the rabbit from where pure saccades can be elicited is an area in the cerebellum (Collewijn, unpublished observations). The finding that these saccades could be elicited as easily with open as with covered eyes is explained by the earlier findings that the optokinetic stabilization system does not respond to stepwise displacements (Collewijn, 1972a).
Thanks are due to Miss W. Zuidam for excellent histological and secretarial assis- tance.

RABBIT OCULOMOTOR AREAS 21
REFERENCES
ARIMOTO, K. (1958). Superior colliculus as center of nystagmus.
BENDER, B. and QHANZER, S. (1964).
Wahayamu Med.
Oculomotor pathways defined by electric stimu- In: The Oculomotor System,
BERGMANN, F., CHAIMOVITZ, M., GUTMAN, J. and ZELIG, S. (1963). Optokinetic J . Physiol. (London) 168:
Nystagmus
Uber den durch elektrische Reizung des Hirnstammes auslijs-
Input-output rela-
Latency and gain of the rabbit's optokinetic reactions to
Brain
Vertical and torsional optokinetic eye movements in the rabbit.
Nystagmus evoked in the rabbit by electrical stimulation of the lateral geniculate body. Ex- perientia 22: 102-103.
GERHARD, L. (1968). Springer, Berlin.
G I O I ~ , R. A. (1961). An experimental study of the accessory optic tracts (trans- peduncular tracts and anterior accessory optic tracts) in the rabbit. J. Comp. Neurol. 117: 77-95.
GIOLLI, R. A. and GUTHRIE, M. D. (1969). The primary optic projections in the rabbit. An experimental degeneration study.
GUTMAN, J., BERGMANN, F., CHAIMOVITZ, M. and COSTIN, A. (1963). Nystagmus evoked by stimulation of the optic pathways in the rabbit. Exp. Neurol. 8: 132-142.
HORRELEN, J. F. and COLLEWIJN, H. (1971). Effect of cerebro-cortical and collicular ablations upon the optokinetic reactions in the rabbit. Doc. Ophthalmol. 30: 227-236.
HOFFMANN, K. P. (1973). Conduction velocity in pathways from retina to superior colliculus in the cat: J . Neurophysiol. 36: 409-424.
HUGHES, A. (1971). Topographical relationships between the anatomy and physiology of the rabbit visual system.
KOMATSUZAKI, A,, ALPERT, J., HARRIS, H. E. and COHEN B. (1972). Effects of mesen- cephalic reticular formation lesions on optokinetic nystagmus. Exp. Neurol. 34:
Central nystagmus elicited Amer. J . Physiol. 193: 328-
Role of the accessory optic system in the optokinetic nystagmus of
The accessory optic system.
Rep. 4: 1-35.
lation and lesions in the brain stem of the monkey. Bender, M. B. (Ed.).
nystagmus and its interaction with central nystagmus.
Hoeber-Harper and Row, New York.
318-331. BERGMANN, F., COSTIN, A,, GUTMAN, J. and CHAIMOVITZ, M. (1964).
BLOHMKE, A. (1929).
CoImcwIm, H. (1969). Optokinetic eye movements in the rabbit:
COmEwIJN, H. (1972a).
COLLEWIJN, H. (1972b).
COLLEWIJN, H. and NOORDUIN, H. (1972).
COSTIN, A., BERGMANN, F., CHAIMOVITZ, M. and GINATH, Y. (1966).
evoked from the superior colliculus of the rabbit. Exp. Neurol. 9: 386 -399.
baren Nystagmus heim Kaninchen.
tions. Vision Res. 9: 117-132.
small movements.
2. Hals, Nasen Ohrenheillz. 23: 213-241.
Brain Res. 36: 59-70. An analog model of the rabbit's optokinetic system.
R ~ S . 36: 71-88.
Pfliigers Arch. 332: 87-95.
Atlas des Mittel- und Zwischenhirns cles Kaninchens.
J. Comp. Neurol. 136: 99-126.
A correlation with receptive field properties.
Doc. Ophthalmol. 30: 33-159.
522-534. LACHMANN, J., BERGMANN, F. and MONNIER, M. (1958).
by stimulation of the meso-diencephalon in the rabbit. 334.
the frog. LLzAR, G. (1973).
MARG, E. (1964). MONNIER, M. (1967). Central mechanisms of vestibular and optokinetic nystagmus.
In: Myotatic, Kinesthetic and Vestibular Mechanisms, De Reuck, A. V. S. and Knight, J. (Eds.).
OYSTER, C. W., TAKAHASHI, E. and COLLEWIJN, H. (1972). Direction-selective retinal ganglion cells and control of optokinetic nystagmus in the rabbit. Vision Res. 12:
Brain, Behau. Euol. 5: 443-460. Ann. N.Y. Acad. Sci. 117: 35-52.
Ciba Foundation Symposium, Churchill, London.
183-193.

22 COLLEWIJN
PASIK, T., PASIK, P. and BENDER, M. B. (1966). The superior colliculi and eye move- ments.
PROCTOR, L. R. (1962). Experimental observations on postural nystagmus. 111. Lesions of the colliculi.
ROBINSON, D. A. (1972). Eye movements evoked by collicular stimulation in the alert monkey.
SCALIA, F. (1972). The termination of retinal axons in the pretectal region of mammals. J . Comp. Neurol. 145: 223-258.
SCHILLER, P. H. and STRYKER, M. (1972). Single-unit recording and stimulation in superior colliculus of the alert rhesus monkey.
STEWART, D. L., TOWNS, L. C. and BIRT, D. (1973). Visual receptive-field characteris- tics of posterior thalamic and pretectal neurons in the rabbit. Bruin Res. 57: 43-57.
STRASCHILL, M. and RIEGER, P. (1973). Eye movements evoked by focal stimulation of the cat's superior colliculus. Bruin Res. 59: 211-227.
WURTZ, R. H. and GOLDBERG, M. E. (1972). Activity of superior colliculus in behaving monkey. 111. Cells discharging before eye movements. J . Neurophysiol. 35: 575- 586.
Arch. Neurol. (Chicago) 15: 420-436.
Ann. Otol. Rhino/. Laryngol. 71: 891-912.
Vision Res. 12: 1795-1808.
J . Neurophysiol. 35: 915--924.
Accepted for publication September 3, 1974
Copyright © 2022 FDOKUMEN




















