
Amphetamine triggers an increase in met-enkephalin simultaneously in brain areas and immune cells
14
Amphetamine triggers an increase in met-enkephalin simultaneously in brain areas and immune cells María A. Assis a,1 , César Collino b,c , María de L. Figuerola d , Claudia Sotomayor b , Liliana M. Cancela a, ⁎ a Departamento de Farmacología, Facultad de Ciencias Químicas, Universidad Nacional de Córdoba, Ciudad Universitaria, X5000HUA, Córdoba, Argentina b Departamento de Bioquímica Clínica, CIBIC-CONICET, Facultad de Ciencias Químicas, Universidad Nacional de Córdoba, Ciudad Universitaria, X5000HUA, Córdoba, Argentina c Centro de Química Aplicada, CEQUIMAP, Facultad de Ciencias Químicas, Universidad Nacional de Córdoba, Ciudad Universitaria, X5000HUA, Córdoba, Argentina d Centro de Investigaciones Endocrinólogicas (CEDIE), CONICET, Hospital de Niños Ricardo Gutiérrez, Gallo 1360, 1425, Buenos Aires, Argentina Received 21 October 2005; received in revised form 16 May 2006; accepted 17 May 2006 Abstract We analyzed effects of amphetamine on proenkephalin-derived peptides in brain areas and immune cells in rats. Acute, as well as a repeated amphetamine treatment, decreased the concanavalin-A-induced lymphocyte proliferation, concomitantly with an increase of free met-enkephalin in nucleus accumbens, prefrontal cortex, spleen, thymus and splenic macrophages. Proenkephalin protein increased in prefrontal cortex, thymus (32 kDa isoform), nucleus accumbens and spleen (44 kDa isoform), while proenkephalin mRNA levels decreased in brain stem. The influence of met-ENK in key brain areas for sensitization and in immune organs is consistent with the idea that changes on met-ENK could underlie amphetamine's effects on brain and IS. © 2006 Elsevier B.V. All rights reserved. Keywords: Psychostimulants; Proenkephaline; Sensitization; T-cell proliferation; Rat 1. Introduction Drug addiction is a devastating illness with deleterious consequences not only on behavior, but also on immune system (IS) functions, rendering the individuals more vulnerable to infectious diseases. It is currently known that drugs of abuse clearly perturb immune functions as do stress, mood and emotion (Woiciechowsky et al., 1999; Pruett, 2001; Galinowski et al., 1992; Baldwing et al., 1998; Padgett and Glaser, 2003). Furthermore, the interchangeability between psychostimulant drugs and stress observed at behavioral level, showing long-lasting changes in dopami- nergic neurons and in the pituitary functions (Diaz-Otañez et al., 1997; Knych and Eisenberg, 1979; Saal et al., 2003), was also found in the IS (Basso et al., 1999). Sensitization is a well known adaptive process which has been associated to the long-lasting behavioral consequences following exposure to psychostimulants (Robinson and Berridge, 2000; Vezina et al., 2002). Several of the changes Journal of Neuroimmunology 178 (2006) 62 – 75 www.elsevier.com/locate/jneuroim Abbreviations: IS, immune system; DA, dopamine; NAc, nucleus accumbens; PfC, prefrontal cortex; ENK, enkephalin; AMPH, amphet- amine; Con A, concanavalin-A; VEH, vehicle; ZT, zeitgeber; RIA, radioimmunoassay; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; FITC, fluorescein isothiocyanate; MoAb, monoclonal antibody; PE, R- phycoerythrin; CREB, cAMP-response element binding protein; AP-1, activator protein-1. ⁎ Corresponding author. Tel.: +54 351 4344974x161; fax: +54 351 4334420. E-mail address: [email protected] (L.M. Cancela). 1 Fellowship from CONICET. 0165-5728/$ - see front matter © 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.jneuroim.2006.05.009
Transcript of Amphetamine triggers an increase in met-enkephalin simultaneously in brain areas and immune cells

gy 178 (2006) 62–75www.elsevier.com/locate/jneuroim
Journal of Neuroimmunolo
Amphetamine triggers an increase in met-enkephalin simultaneouslyin brain areas and immune cells
María A. Assis a,1, César Collino b,c, María de L. Figuerola d,Claudia Sotomayor b, Liliana M. Cancela a,⁎
a Departamento de Farmacología, Facultad de Ciencias Químicas, Universidad Nacional de Córdoba,Ciudad Universitaria, X5000HUA, Córdoba, Argentina
b Departamento de Bioquímica Clínica, CIBIC-CONICET, Facultad de Ciencias Químicas, Universidad Nacional de Córdoba,Ciudad Universitaria, X5000HUA, Córdoba, Argentina
c Centro de Química Aplicada, CEQUIMAP, Facultad de Ciencias Químicas, Universidad Nacional de Córdoba,Ciudad Universitaria, X5000HUA, Córdoba, Argentina
d Centro de Investigaciones Endocrinólogicas (CEDIE), CONICET, Hospital de Niños Ricardo Gutiérrez,Gallo 1360, 1425, Buenos Aires, Argentina
Received 21 October 2005; received in revised form 16 May 2006; accepted 17 May 2006
Abstract
We analyzed effects of amphetamine on proenkephalin-derived peptides in brain areas and immune cells in rats. Acute, as well as arepeated amphetamine treatment, decreased the concanavalin-A-induced lymphocyte proliferation, concomitantly with an increase of freemet-enkephalin in nucleus accumbens, prefrontal cortex, spleen, thymus and splenic macrophages. Proenkephalin protein increased inprefrontal cortex, thymus (32 kDa isoform), nucleus accumbens and spleen (44 kDa isoform), while proenkephalin mRNA levels decreasedin brain stem. The influence of met-ENK in key brain areas for sensitization and in immune organs is consistent with the idea that changes onmet-ENK could underlie amphetamine's effects on brain and IS.© 2006 Elsevier B.V. All rights reserved.
Keywords: Psychostimulants; Proenkephaline; Sensitization; T-cell proliferation; Rat
1. Introduction
Drug addiction is a devastating illness with deleteriousconsequences not only on behavior, but also on immune
Abbreviations: IS, immune system; DA, dopamine; NAc, nucleusaccumbens; PfC, prefrontal cortex; ENK, enkephalin; AMPH, amphet-amine; Con A, concanavalin-A; VEH, vehicle; ZT, zeitgeber; RIA,radioimmunoassay; GAPDH, glyceraldehyde-3-phosphate dehydrogenase;FITC, fluorescein isothiocyanate; MoAb, monoclonal antibody; PE, R-phycoerythrin; CREB, cAMP-response element binding protein; AP-1,activator protein-1.⁎ Corresponding author. Tel.: +54 351 4344974x161; fax: +54 351
4334420.E-mail address: [email protected] (L.M. Cancela).
1 Fellowship from CONICET.
0165-5728/$ - see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.jneuroim.2006.05.009
system (IS) functions, rendering the individuals morevulnerable to infectious diseases. It is currently known thatdrugs of abuse clearly perturb immune functions as do stress,mood and emotion (Woiciechowsky et al., 1999; Pruett,2001; Galinowski et al., 1992; Baldwing et al., 1998; Padgettand Glaser, 2003). Furthermore, the interchangeabilitybetween psychostimulant drugs and stress observed atbehavioral level, showing long-lasting changes in dopami-nergic neurons and in the pituitary functions (Diaz-Otañez etal., 1997; Knych and Eisenberg, 1979; Saal et al., 2003), wasalso found in the IS (Basso et al., 1999).
Sensitization is a well known adaptive process which hasbeen associated to the long-lasting behavioral consequencesfollowing exposure to psychostimulants (Robinson andBerridge, 2000; Vezina et al., 2002). Several of the changes

63M.A. Assis et al. / Journal of Neuroimmunology 178 (2006) 62–75
that underlie this process occur in the mesocorticolimbicdopamine (DA) systems, of which the ventral tegmentalarea, nucleus accumbens (NAc) and prefrontal cortex (PfC)are the main components. However, enhanced dopaminer-gic transmission is only one component of the motivecircuit, which mediates behavioral sensitization to psychos-timulants. Other neurotransmitter systems, such as enke-phalins (ENKs) and glutamate, are also among the mostimportant neurotransmitters in the mesocorticolimbic sys-tems associated with the development and/or expression ofsensitization (Kalivas and Stewart, 1991; Pierce andKalivas, 1997a). Notwithstanding, no evidence has so farbeen provided regarding possible psychostimulant-inducedlong-lasting changes in the ENK levels in these brain areas.
It has been shown that the administration of centralnervous system (CNS) acting drugs into the lateralventricle of the brain, induced a similar effect on the ISas that observed following the systemic infusion of thedrug (Pellegrino and Bayer, 1998a, 2000) suggesting that acentral component could be involved. Related to this, itwas found that brain DA and opioid systems have beeninvolved in the impairment of IS functions followingpsychostimulants and/or stress (Basso et al., 1999; Saraviaet al., 1998; Kubera et al., 2002; Jankovic and Radulovic,1992). However, it is also possible that autocrine andparacrine actions of opioid peptides and DA produced byimmune cells may be responsible for the effects of drugabuse on IS (Gordon and Barnes, 2003; Pellegrino andBayer, 1998b).
ProENK mRNA and proENK derived peptides weredetected in immune cells of bone marrow, thymus, spleenand alveolar macrophages (Vindrola et al., 1990; Saravia etal., 1993; Linner et al., 1995, 1991; Padrós et al., 1989;Kuis et al., 1991; Roth et al., 1989; Rittner et al., 2001).However, the myeloid lineage cells are the only immunecells that contain the prohormone converting enzymes PC1and/or PC2, which are involved in the post-translationalprocessing of proENK to produce the opioid peptides (Rothet al., 1989; Vindrola and Lindberg, 1992; Mathis andLindberg, 1992). Because of this, only macrophages andneutrophils are able to synthesize opioid peptides derivedfrom proENK (i.e. met-ENK) (Saravia et al., 1998; Kuis etal., 1991). In agreement with this, it was shown that T-lymphocytes release cryptic met-ENK-containing peptidesbut they do not have opioid activity (Roth et al., 1989;Padrós et al., 1995). On the other hand, several physiolog-ical studies have demonstrated the immunosuppressiveeffect of opioid peptides (Jankovic and Radulovic, 1992).Interestingly, a stressful stimulus inducing immunosuppres-sion modified proENK-derived peptide levels and theirrelease from cells of peripheral immune tissues (Saravia etal., 1998) as it did in CNS (Borsook et al., 1994; Lightmanand Young, 1987). In addition to these neuropeptides, thecatecholamines, including DA, are also present in lympho-cytes, macrophages and neutrophils, which also express thetransporters, receptors and synthesis enzymes for these
neurotransmitters (Bergquist et al., 1994; Amenta et al.,2001; McKenna et al., 2002; Cosentino et al., 2002).
It has been shown that long-lasting behavioral sensitiza-tion is induced following either a single dose or a chronicamphetamine (AMPH) treatment (Vanderschuren and Kali-vas, 2000). In this work, we showed that a single dose ofAMPH is also able to induce effects on IS, which wereobservable after 4 days, as in the case of a repeated regimewith the drug. At the same time of observation, it could beseen that both acute and repeated AMPH treatments hadinduced a simultaneous increase of met-ENK in IS, as well asin CNS. We also found that AMPH was a sufficient stimulusfor splenic T-lymphocytes to produce almost as many crypticproENK-derived peptides as concanavalin-A (Con A).Interestingly, it was shown that splenic macrophages werethe cells that synthesized elevated amounts of met-ENKfollowing the drug. All these data suggest that the activationof enkephalinergic systems in CNS and IS may be involvedin the AMPH's effects on both systems. Furthermore,changes in the proENK-derived peptide levels fromperipheral immune cells might be an index reflecting anopioid central effect of AMPH.
2. Materials and methods
2.1. Animals
Adult male Wistar rats (250–330 g) from the Facultad deCiencias Veterinarias of the Universidad Nacional de LaPlata (Buenos Aires, Argentina) were maintained at 20–24 °C under a 12 h light–dark cycle (lights on at 07:00 a.m.)with free access to food and water. Rats were collectivelyhoused in cages in the experimental room for at least 7 daysbefore starting the experiments. All procedures wereconducted in accordance with the NIH Guide for the Careand Use of Laboratory Animals as approved by Animal Careand Use Committee of the Facultad de Ciencias Químicas,Universidad Nacional de Córdoba.
2.2. Drugs
For all experiments D-AMPH sulfate (Sigma Co, St.Louis, MO) was dissolved in an isotonic saline solution(0.9% NaCl), which was also used for vehicle (VEH) controlinjections. All injections were administered intraperitoneallyin a volume of 1 ml/kg and the treatments were made at 11 a.m. (ZT 4) to avoid the influence of the circadian rhythm onthe immune response (Haus and Smolensky, 1999) and onthe behavioral sensitization to psychostimulants (Abarca etal., 2002).
2.3. Experimental procedures
2.3.1. Repeated AMPH treatmentRats were randomly assigned to one of three treatments:
VEH group, AMPH 5×1 (1 mg/kg/day i.p.) and AMPH 5×2

Table 1Amplification parameters of proENK, GAPDH and c-fos mRNA
ProENK: exons I–III GAPDH and c-fos
94 °C–5 min 94 °C–5 min60 °C–2 min 60 °C–2 min72 °C–3 min 72 °C–3 min1 cycle 1 cycle94 °C–1 min 94 °C–1 min60 °C–2 min 60 °C–1 min72 °C–3 min 72 °C–1.5 min30 cycles 30 cycles94 °C–1 min 94 °C–1 min60 °C–2 min 60 °C–2 min72 °C–10 min 72 °C–10 min1 cycle 1 cycle
64 M.A. Assis et al. / Journal of Neuroimmunology 178 (2006) 62–75
(2 mg/kg/day i.p.). The AMPH or VEH treatment wasadministered during days 1–5, and on day 9 animals werekilled by decapitation (4 days following the last druginjection) (Fig. 2A). Spleens were removed and mononu-clear cells were isolated as mentioned below. Since weobtained the same proliferative response in untreated controlanimals as in the VEH group in previous experiments (datanot shown), this indicated that there was no influence ofinjection on this parameter, therefore we decided to use VEHas the control group.
2.3.2. Acute AMPH treatmentRats were randomly assigned to one of five treatments:
VEH group, AMPH 1×0.5 (0.5 mg/kg i.p.), AMPH 1×1(1 mg/kg i.p.), AMPH 1×2.5 (2.5 mg/kg i.p.) and AMPH1×5 (5 mg/kg i.p.). The AMPH or VEH treatment wasadministered during day 1, and on day 5 animals were killedby decapitation (4 days following drug injection) (Fig. 2B).Spleens were removed and mononuclear cells were isolated asmentioned below.
As similar dose–response effects were obtained followingacute and repeated AMPH treatment, a single dose of AMPH(5mg/kg i.p.) was chosen to asses the possible effect on spleencellularity and to observe the long-lasting effects of proENKmRNA, proteins and derived peptides on IS and CNS.
Once rats were decapitated, the brain, spleen, thymus andbone marrow were removed. The following brain areas ofboth hemispheres were dissected: cerebellum, caudateputamen, PfC, hippocampus, NAc, septum and brainstem.
2.3.3. Isolation of mononuclear cellsSpleen cell suspensions were obtained by gently grinding
tissue into RPMI 1640 culture medium (Sigma-Aldrich,Steinheim, Germany) under sterile conditions. Mononuclearcells were separated by Ficoll–Hypaque density gradient(1.083 g/ml) centrifugation. The mononuclear cells concen-trated at the surface were collected and washed twice inRPMI 1640 medium. After cell counting, splenocytes weresuspended at a final concentration of 2×106 cells/ml inRPMI 1640 medium supplemented with 10% of inactivatedfetal bovine serum, 2 mM glutamine, 10 mM sodiumbicarbonate, 100 UI/ml penicillin and 100 μg/ml streptomy-cin (denominated complete RPMI 1640 medium).
To evaluate the proENK-derived peptides released fromcultured spleen cells, part of the splenic mononuclear cells
Fig. 1. The specificity of met-enkephalin-Arg-Phe (ME-Arg-Phe)-directedantiserum (dilution 1 :1000) was determined incubating it (previous to theWestern blot) with different concentrations of the synthetic peptide used toproduce the conjugate injected to the rabbits. 100 μg of proteins from ratthymus acidic extracts were electrophoresed and analyzed by Western blot.
were cultured in flasks (TPP, Switzerland) for 48 h withcomplete RPMI 1640 medium with or without Con A, asdescribed below. Then, the supernatants were separated fromthe non-adherent cells by centrifugation, and used forradioimmunoassay (RIA) (Saravia et al., 1998). The adherentcells (macrophages) from culture without Con A wereremoved with 2 M acetic acid after two washes with salinesolution, boiled for 15 min, homogenized by sonication,centrifuged at 50,000×g for 1 h, and used for RIA.
2.3.4. Mitogenic assayThe mitogen Con A (Sigma-Aldrich, Steinheim, Germany)
was used to evaluate the splenic T-cell response. Splenocyteswere added in quadruplicate to each well of a 96-well flat-bottom tissue culture plates (TPP, Switzerland) in the presenceof 5 μg/ml of Con A. The plates were incubated at 37 °C in ahumidified atmosphere with 5% CO2 for 48 h. 18 h beforeharvesting, cells were pulsed with 1 μCi [3H]-thymidine(PerkinElmer Life Sciences, Wellesley, MA). Finally, quad-ruplicates were collected onto glass fiber filter paper (Watt-man, UK) using a Scatron micro cell harvester (SIEM,Córdoba, Arg.). A 1205 Betaplate liquid scintillation counter(PerkinElmer Life Sciences, Wellesley, MA) was used todetect the incorporation of radioactive thymidine in a Packard,Tri-Carb Liquid Scintillation Analyzer. The radioactivity wasexpressed as cpm and the percentage of responses wascalculated relative to VEH-treated animals.
2.3.5. Flow cytometry studiesIn order to evaluate if the differences in the lymphopro-
liferative response could be produced by a direct AMPH effecton lymphocyte subpopulations, the spleens were weighed andtheir cellularity were assessed by flow cytometry. Cellsuspensions were obtained by gently grinding tissue intoRPMI 1640 culture medium without phenol red (Sigma-Aldrich, Steinheim, Germany), counted and suspended at afinal concentration of 2×106 cells/ml. To analyse the totalcount of T- and B- and NK-cells in these tissues, three-colourimmunofluorescence staining was performed and the intensityof stained cells was analysed by flow cytometry (CytoronAbsolute flow cytometer; Ortho Diagnostic System, Raritan,

Fig. 2. Schematic diagrams of repeated (A) and acute (B) AMPH treatments. C: Dose-dependent effects of repeated and acute AMPH treatment on theimmunoproliferative response Percentage of proliferative responses related to the control groups (VEH). Data show the mean±S.D. of five rats per group.⁎p<0.05 and ⁎⁎p<0.01. These data are representative of at least two independent experiments.
65M.A. Assis et al. / Journal of Neuroimmunology 178 (2006) 62–75
NJ, USA). 100 μL of cells suspension were incubated withfluorescein isothiocyanate-conjugated mouse anti-rat CD161amonoclonal antibody (MoAb), R-phycoerythrin-conjugatedmouse anti-rat CD3 MoAb and Cy-Chrome™-conjugatedmouse anti-rat CD45RAMoAb (BD Bioscience, NJ, USA) inorder to determine NK-, T- and B-cells, respectively. Theincubation was for 30 min in the dark, followed by 15 minwith lysis buffer and three washes in PBS (pH 7.2). After that,the cells were fixed with 2% formaldehyde, washed tree timesin PBS and finally resuspended in ISOTON prior to analyse inthe cytometer.
Lymphocytes were gated based on their characteristic light-scatter and corroborated by a morphologic analysis by May–Grünwald–Giemsa. Fluorescence intensity was depicted on athree-decade logarithmic scale and in single-parameteranalysis as histograms. Absolute numbers of lymphocyteswere calculated according to lymphocyte subset percentages
and the absolute value of leukocytes (analyzed in a Coulter T-540 hematology analyzer).
2.3.6. Total and free met-ENK radioimmunoassayFrozen immune tissues and brain areas were suspended in
1 M acetic acid containing 50 mM HCl, boiled for 15 min,homogenized with a Polytron, and centrifuged at 50,000×gfor 1 h. An aliquot of the supernatant was lyophilized andreconstituted in 50 mM Tris–HCl buffer, pH 8.4, and 2 mMCaCl2. Free and total immunoreactive met-ENK, thosebeing, previous to and after sequential enzymatic digestionwith trypsin and carboxypeptidase B, were determined byRIA (Vindrola et al., 1990; Padrós et al., 1989).
Culture medium was diluted in Tris buffer forenzymatic digestion and peptides assayed as describedbefore (Saravia et al., 1998; Padrós et al., 1989). Specificmet-ENK antiserum was obtained and characterized as

Table 2Effect of the acute AMPH treatment (5 mg/kg i.p.) on spleen weight(mg spleen/g body weight) and cellularity (cell number×106/mg spleen)
Spleens VEH treatment AMPH treatment
Weight 2.08±0.19 2.03±0.13Leucocytes 566.5±28.1 549.9±27.9Lymphocytes 447.3±24.9 437.2±21.3T-cells 141.5±23.4 135.4±9.6B-cells 247.7±8.0 258.0±16.0NK cells 1.5±0.5 1.7±0.2
The results are the means±S.D. of four rats.
66 M.A. Assis et al. / Journal of Neuroimmunology 178 (2006) 62–75
previously described (Vindrola et al., 1990; Padrós et al.,1989), and used to identify the carboxy-terminal portion ofmet-ENK.
2.3.7. ImmunoblottingAliquots of acidic tissue extracts were dried, suspended
in 2× Laemmli buffer and boiled for 5 min. 50 μg ofproteins were applied to 15% (proENK-derived proteins) or8.8% (PC1 and Fos) acrylamide gel using the mini-proteangel system (BioRad). Electrophoresis and immunoblottingwere carried out as described previously (Saravia et al.,1998). The second antibody was conjugated with alkalinephosphatase (BioRad Laboratories, Hercules, CA) anddeveloped with the respective color reagent (BioRad kit).Antibodies against met-ENK-Arg-Phe (final dilution1 : 1000) (Fig. 1), N-terminal PC1 (DAM3-B6) (finaldilution 1 :1000), Fos (final dilution 1 :500) and actin(1 :500) (Santa Cruz, Inc. CA) were used in these studies.Control experiments were run simultaneously, omitting thefirst antibody. PC1 enzyme was mostly found as a long andstable pool in neuroendocrine cells (Rittner et al., 2001),while Fos mainly appeared as a consequence of tissue or cellactivation. The measurement of these two proteins with aknown pattern of expression in rat brain served as areference of the specific response of proENK proteins. Sincean increase in proENK and met-ENK after AMPH treatmentwas only obtained in PfC, we also measured PC1 and Fos inthis structure.
2.3.8. RNA isolationTotal cellular RNA was extracted from the immune
tissues and brain areas with TRIZOL isolation solvent asdescribed by the manufacturer (Invitrogen, Inc.). RNA wasprecipitated and suspended in 50 μl diethylpyrocarbonate-treated-dH2O and the concentration was estimated after opticdensity lecture at 260 nm using a GeneQuantPro spectro-photometer (Amersham-Pharmacia, Buckinghamshire, UK).The integrity of RNA was determined by applying 1 μg in1.5% agarose gel containing ethidium bromide followed byUV visualization in a transiluminator (Stratagene, La Jolla,CA). mRNAwas purified by mini kit Oligotex, following thespecification described by the manufacturer (Qiagen Inc.,Hilden, Germany).
2.3.9. RT–PCR analysis5 μg of total RNAwere incubated for 5 min at 90 °C, and
reverse transcription was carried out in a final volume of20 μl, adding 500 ng of random hexamers, 100 units ofMoloney murine leukemia virus reverse transcriptase(Promega, Madison, WI), 0.5 mM dNTPs and 20 units ofRNasin (Promega, Madison, WI). The samples wereincubated for 60 min at room temperature, followed by45 min at 37 °C. Finally, tubes were heated to 95 °C for5 min. Samples containing 0.5 μg of RT were amplified inthe presence of 200 μM of dNTPs (Roche Diagnostics,Mannheim, Germany), 1 μM of sense and antisense primers
(Life Technology, USA), 1.5 mM MgCl2 and 1.25 units TaqDNA polymerase (Roche Diagnostics, Mannheim, Ger-many) and were placed in a thermal cycler (Robocycler 40,Stratagene, La Jolla, CA). The parameters of the amplifica-tion are specified in Table 1. A proENK-specific primers pairwas designed and assessed for specificity by a computer-assisted search of up-dated versions of Oligo-4 program. ThePCR primer sequences for c-fos and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were obtained from aprevious publication (Saravia et al., 1998). All primers usedwere synthesized by Life Technologies.
ProENK exon I–exon III: 5′–CCAGCTACCTGCGC-CATCTGAACAA–3′ and 5′–AGAAGCTTTTCC-TGTTGGTGGCTATCTTT–3′ (545 bp).c- fos : 5′–TGCATGAATTCCCCAGCCGACTC-CTTCTCCA– 3 ′ a n d 5 ′ –TGCATAAGCTT-CAGCTCCCTCCTCCGATTCC–3′ (388 bp).GAPDH: 5′–TGCATGAATTCTGATTCTACCCACGG-CAAGT–3 ′ and 5 ′–TGCATAAGCTTGTCAT-GAGCCCTTCCACGAT–3′ (380 bp).
The PCR products were analyzed on ethidium bromide3% agarose gels followed by UV visualization in atransiluminator (Stratagene).
2.4. Statistical analysis
Cell proliferation was measured by [3H]-thymidineincorporation assay and results were plotted as thepercentage of changes between AMPH- and VEH-treatedrats. Data represent means±S.D., and correspond toquadruplicate values of five different rats. Student's t-test(the level of significance was set at p<0.05) was used toevaluate the statistical significance.
3. Results
3.1. Dose-dependent effects of repeated and acute AMPHtreatments on the immunoproliferative response
Exposure of rats to repeated AMPH treatment resulted ina dose-dependent decrease in the in vitro splenic proliferativeresponse to Con A, relative to VEH-treated controls. This

67M.A. Assis et al. / Journal of Neuroimmunology 178 (2006) 62–75
effect was seen by a significant decrease in 3H-thymidineincorporation, by 45% (p<0.05) and 65% (p<0.01), in ConA-stimulated splenocytes, for rats treated with 1 or 2 mg/kg/day of AMPH, respectively (Fig. 2C).
Similar results were obtained after exposure to acute AMPHtreatment. 3H-thymidine incorporation was significantly re-duced in a dose-dependentmanner, by 37% (p<0.05), 50% and65%(p<0.01) in rats treatedwith 1, 2.5 or 5mg/kg, respectively(Fig. 2C). No significant differences were detected in ratstreated with 0.5 mg/kg relative to the control group.
As similar results were obtained after the repeated andacute VEH treatments in the proliferative response to ConA (p=NS), the data were pooled (5.6±0.5×104 cpm) andrepresented the maximum percentage of proliferation.
3.2. Effect of acute AMPH on the spleen weight andcellularity
No significant differences on spleen weights wereobserved after the acute AMPH (5 mg/kg) treatment, relativeto their controls (Table 2). In the same line, the lymphocytesubpopulations of spleens in rats treated with AMPH did notshow any difference, in absolute values as well as inpercentages, relative to their VEH-treated controls (Table 2).
3.3. Effect of AMPH on the proENK-derived peptide levelsin rat mesocorticolimbic areas
3.3.1. Radioimmunoassay proceduresThe acute AMPH treatment significantly increased the
free met-ENK levels in NAc (80%, p<0.05) and PfC (45%,p<0.05), compared with VEH-treated controls. This effectwas not seen in the other brain regions tested (Table 3,column 1 vs. 4).
No significant changes in total met-ENK levels weredetected in NAc and PfC after AMPH treatment (Table 3).However, free to total met-ENK ratios increased significant-ly in both regions (p<0.05) (Table 3, column 3 vs. 6).
To confirm if the changes after the acute AMPH treatmentin the met-ENK content are also observed after a repeatedAMPH treatment (AMPH 5×2), we studied the same brain
Table 3Effect of the acute (5 mg/kg i.p.) and repeated (2 mg/kg/day i.p.) AMPH treatmen
Brain areas IR-met-ENK levels (pg/mg protein)
Ac VEH Ac A
Free Total Ratio Free
Brainstem 3014±305 n.m. 3370±Caudate Putamen 1680±185 n.m. 1834±Cerebelum 1615±185 n.m. 1309±Hippocampus 1862±427 n.m. 1361±Nucleus accumbens 997±75 7663±96 0.13±0.02 1795±Prefrontal cortex 593±93 3436±295 0.17±0.03 861±Septum 3231±649 n.m. 2684±
The results are the means±S.D. of five rats. ap<0.01 and bp<0.05 compared to tAc=acute; Rep=repeated; IR= immunoreactive; n.m.=not measured; n.d.=not de
structures and immune tissues that showed a variation after theacute treatment. This AMPH repeated treatment was chosenbecause it produced a similar decrease of proliferativeresponse to that observed after acute AMPH (5 mg/kg i.p.).
The repeated AMPH treatment significantly increased thefree met-ENK levels in NAc (72%, p<0.01) and PfC (39%,p<0.01), compared with VEH-treated controls. (Table 3,column 7 vs. 8).
3.3.2. Western blotting studiesIn order to characterize high molecular weight proENK
molecules in NAc and PfC and their probable contentchanges induced by the acute AMPH treatment, weperformed immunoblotting analysis, using an antibodyagainst the C-terminal portion of proENK, which distin-guished three proteins of 32, 44 and 64 kDa in rat brain (Fig.3A). The 32 kDa molecule corresponded to the authenticproENK and the 64 kDa protein corresponds to therespective dimer (Lindberg and Shaw, 1992). The 44 kDamolecule corresponds to a novel isoform previouslydescribed in tumoral tissues and cells (Vindrola et al.,1998; Brar and Lowry, 1999). AMPH-treated rats showed asignificant increase in the 32 kDa proENK content in the PfC(p<0.05), while the other two proteins levels remainedpractically unchanged. On the other hand, the AMPHtreatment did not modify 32 kDa protein levels in NAc,where only a significant decrease (p<0.05) of the 44 kDamolecule was detected (Fig. 3).
PC1 enzyme appeared as the 68 kDa molecule and itshigh levels, as well as actin levels, were not affected inPfC and NAc of AMPH-treated rats, respectively.However, blotting results for Fos proteins in PfC ofAMPH-treated rats showed a significant increase in the35–37 kDa band, corresponding to highly stable isoformsof ΔFosB (Fig. 4).
3.4. Effect of acute AMPH on the proENK mRNA levels inrat mesocorticolimbic areas
The proENK mRNA content did not change in PfC andNAc, but showed a significant decrease in brainstem (p<0.05)
ts on brain IR-met-ENK levels (pg/mg protein)
MPH Rep VEH Rep AMPH
Total Ratio Free Free
640 n.m. n.m. n.m.160 n.m. n.m. n.m.249 n.m. n.m. n.m.334 n.m. n.m. n.m.226b 6952±1093 0.26±0.06a 1133±72 1952±177a
72b 3005±152 0.29±0.04a 724±67c 1008±55a
396 n.m. n.m. n.m.
heir controls (VEH-treated rats); cp<0.05 compared to the acute treatment;tectable levels.

Fig. 3. Regulation of proENK by AMPH in PfC and NAc. (A): Immunoblotting of proENK peptides in control rats (VEH) and in rats that received an acuteAMPH treatment (5 mg/kg i.p.) 4 days before. (B) Relative units of absorbance of 32, 44 and 64 kDa proENK isoforms. Data show the mean±S.D. of three ratsper group. ⁎p<0.05.
68 M.A. Assis et al. / Journal of Neuroimmunology 178 (2006) 62–75
(Fig. 5) after the acute AMPH treatment. The c-fos mRNAlevels were not changed in any of brain regions tested (Fig. 5).
In conclusion, the acute AMPH treatment increased freemet-ENK levels in PfC and NAc, as well as proENK content inPfC, and also decreased proENK mRNA levels in thebrainstem.
3.5. Effect of AMPH on proENK-derived peptide levels inrat immune organs
3.5.1. Radioimmunoassay proceduresThe acute AMPH treatment enhanced free met-ENK
levels in rat spleen (35%, p<0.05) and thymus (31%,p<0.05), relative to VEH-treated controls. This effect wasnot seen in the bone marrow (Table 4, column 1 vs. 4).However, total met-ENK (met-ENK plus cryptic met-ENK-containing peptides) did not change significantly in anytested organs. Free to total met-ENK ratios increasedsignificantly in the spleen and thymus, but not in the bonemarrow of the animals treated with AMPH (Table 4, column3 vs. 6). These results suggest an increased proENK post-translational processing towards low molecular weightpeptides in spleen and thymus.
Free met-ENK concentration was not detected in themedium of cultured splenocytes from all-treated animals(Table 4, column 1 vs. 4). However, total met-ENK levelsincreased significantly (65%, p<0.01) (Table 4, column 5 vs.2) in the medium of AMPH-treated rats splenocytes, culturedfor 48 h. This effect was observed in the absence of any kind ofmitogen, that is, with complete RPMI 1640 medium alone.Although the addition of T-lymphocyte mitogen Con A to theculture medium increased significantly the total met-ENKlevels in VEH-treated rats (66%, p<0.05, Table 4, column 2),it produced aminor non-significant effect (14%) on the already
elevated levels following AMPH. Free met-ENK concentra-tion was elevated in the macrophages (38%, p<0.01) fromacute AMPH-treated animals (Table 4, column 1 vs. 4).
The repeated AMPH-treated animals showed a similarpattern in met-ENK content. The repeated treatmentsignificantly increased the free met-ENK levels in spleen(35%, p<0.01), thymus (29%, p<0.01) and macrophages(45%, p<0.01) (Table 4, column 7 vs. 9), and also raisedthe total met-ENK levels in the medium of AMPH-treatedrats splenocytes (Table 4, column 8 vs. 10), cultured for48 h, compared with VEH-treated controls.
3.5.2. Western blotting studiesImmunoblotting studies of proENK proteins showed that
the spleen contained high levels of a 34–32 kDa doublet,together with 64 kDa (see before), 44 kDa (see before),27 kDa and 25 kDa proteins. Spleens of AMPH-treated ratshad a significant increase in the 44 kDa protein content,while the levels of the other proteins remained unchanged(Fig. 6). The thymus contained the same proteins asspleens, except for the 44 kDa molecule. Thymus ofAMPH-treated rats showed a significant increase in 34–32 kDa doublet content with no changes in the otherproteins (Fig. 6). Although both immune organs of AMPH-treated rats showed increased levels of proENK, differentisomers of this molecule were responsible for this change.Actin levels were not modified after AMPH treatment inany tissue assayed.
3.6. Effect of acute AMPH on the proENK mRNA levels inrat immune organs
RT-PCR for proENK, using exon I–exon III primers,amplified the expected fragment of 545 bp (Fig. 7A). The

Fig. 4. Regulation of PC1 and Fos-related proteins by AMPH in PfC. (A) Immunoblotting of PC1; (B) Fos-related proteins in control rats (VEH) and in rats thatreceived an acute AMPH treatment (5 mg/kg i.p.) 4 days before. (C) Relative units of absorbance of PC1 and 35–37 kDa isoform of ΔFosB. Data show themean±S.D. of four rats per group. ⁎p<0.05. MW: molecular weight marker.
69M.A. Assis et al. / Journal of Neuroimmunology 178 (2006) 62–75
proENK mRNA was detected in the thymus but not in thespleen. In this last organ, proENK was not even observedafter RT–PCR of purified mRNA (data not shown). Theseresults verified previous publications, indicating thatproENK mRNA expression in splenocytes may dependon cell activation (Linner et al., 1995). ProENK mRNAcontent did not change in immune organs 4 days after onedose of AMPH (Fig. 7), while this drug only induced asignificant decrease of c-fos mRNA levels in the thymus(p<0.05) (Fig. 7).
4. Discussion
This work has demonstrated that a single dose, as well asa repeated treatment with AMPH, can induce a decrease inthe splenocyte proliferation, as well as a simultaneousincrease in met-ENK levels in immune organs and in brainareas. In addition, the non-activated cultured splenocytesfrom AMPH-treated rats released nearly the same amount ofcryptic met-ENK-containing products as Con A-stimulated
control splenocytes. Within the cultured splenocytes, themacrophages were the cells where the increase in met-ENKwas observed. Since met-ENK has been consistently relatedto a decrease in immune function, our current results stronglysuggest that an increased in vivo release of met-ENK byperipheral monocytes/macrophages may be the primer forthe inhibitory proliferative response of T-lymphocyte. Theinfluence of the met-ENK system in key brain areas forsensitization, and in immune organs, is consistent with theidea that similar biological changes could underlie theAMPH's effects on the CNS and IS.
It has been shown that a single, as well as a repeatedtreatment with AMPH, induces a phenomenon of behavioralsensitization, which has been proposed as a key process inthe addiction to psychostimulants and other abuse drugs(Vanderschuren et al., 1999b; Vanderschuren and Kalivas,2000; Robinson and Berridge, 2000). Several alterations inthe mesocorticolimbic dopaminergic nuclei have beenidentified in the sensitization induced by psychostimulants.Within these nuclei, the ventral tegmental area, NAc, and
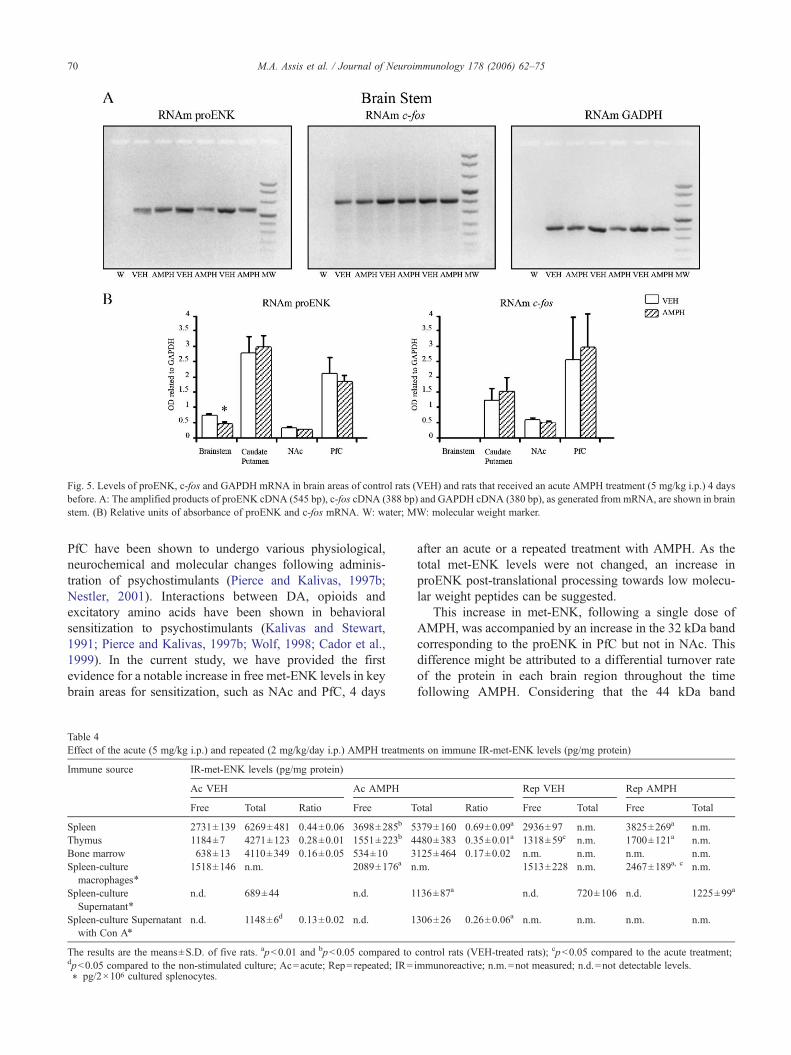
Fig. 5. Levels of proENK, c-fos and GAPDH mRNA in brain areas of control rats (VEH) and rats that received an acute AMPH treatment (5 mg/kg i.p.) 4 daysbefore. A: The amplified products of proENK cDNA (545 bp), c-fos cDNA (388 bp) and GAPDH cDNA (380 bp), as generated from mRNA, are shown in brainstem. (B) Relative units of absorbance of proENK and c-fos mRNA. W: water; MW: molecular weight marker.
70 M.A. Assis et al. / Journal of Neuroimmunology 178 (2006) 62–75
PfC have been shown to undergo various physiological,neurochemical and molecular changes following adminis-tration of psychostimulants (Pierce and Kalivas, 1997b;Nestler, 2001). Interactions between DA, opioids andexcitatory amino acids have been shown in behavioralsensitization to psychostimulants (Kalivas and Stewart,1991; Pierce and Kalivas, 1997b; Wolf, 1998; Cador et al.,1999). In the current study, we have provided the firstevidence for a notable increase in free met-ENK levels in keybrain areas for sensitization, such as NAc and PfC, 4 days
Table 4Effect of the acute (5 mg/kg i.p.) and repeated (2 mg/kg/day i.p.) AMPH treatmen
Immune source IR-met-ENK levels (pg/mg protein)
Ac VEH Ac AMPH
Free Total Ratio Free T
Spleen 2731±139 6269±481 0.44±0.06 3698±285b 5Thymus 1184±7 4271±123 0.28±0.01 1551±223b 4Bone marrow 638±13 4110±349 0.16±0.05 534±10 3Spleen-culture
macrophages⁎1518±146 n.m. 2089±176a n
Spleen-cultureSupernatant⁎
n.d. 689±44 n.d. 1
Spleen-culture Supernatantwith Con A⁎
n.d. 1148±6d 0.13±0.02 n.d. 1
The results are the means±S.D. of five rats. ap<0.01 and bp<0.05 compared todp<0.05 compared to the non-stimulated culture; Ac=acute; Rep=repeated; IR=⁎ pg/2×106 cultured splenocytes.
after an acute or a repeated treatment with AMPH. As thetotal met-ENK levels were not changed, an increase inproENK post-translational processing towards low molecu-lar weight peptides can be suggested.
This increase in met-ENK, following a single dose ofAMPH, was accompanied by an increase in the 32 kDa bandcorresponding to the proENK in PfC but not in NAc. Thisdifference might be attributed to a differential turnover rateof the protein in each brain region throughout the timefollowing AMPH. Considering that the 44 kDa band
ts on immune IR-met-ENK levels (pg/mg protein)
Rep VEH Rep AMPH
otal Ratio Free Total Free Total
379±160 0.69±0.09a 2936±97 n.m. 3825±269a n.m.480±383 0.35±0.01a 1318±59c n.m. 1700±121a n.m.125±464 0.17±0.02 n.m. n.m. n.m. n.m..m. 1513±228 n.m. 2467±189a, c n.m.
136±87a n.d. 720±106 n.d. 1225±99a
306±26 0.26±0.06a n.m. n.m. n.m. n.m.
control rats (VEH-treated rats); cp<0.05 compared to the acute treatment;immunoreactive; n.m.=not measured; n.d.=not detectable levels.

Fig. 6. Regulation of proENK by AMPH in thymus and spleen. (A) Immunoblotting of proENK isoforms in control rats (VEH) and in rats that received an acuteAMPH treatment (5 mg/kg i.p.) 4 days before. (B) Relative units of absorbance of 32, 4nd 64 kDa proENK isoforms. Data show the mean±S.D. of three rats pergroup. ⁎p<0.05.
71M.A. Assis et al. / Journal of Neuroimmunology 178 (2006) 62–75
corresponds to a novel isoform whose function is not totallyunderstood, further studies are necessary to interpret thedecrease of this band in NAc after AMPH treatment.
The increase in proENK in PfC 4 days after AMPHtreatment is in line with that observed in proENK RNAm instriatum following 3 h of acute or repeated AMPH treatment(Wang and McGinty, 1995, 1996; Mao and Wang, 2003).However, a decrease in proENK mRNA levels was observedin the central nucleus of the amygdala, 3 h after the lastinjection following acute and chronic treatment with AMPH(Turchan et al., 2002). It has been suggested that the increasein proENK RNAm by AMPH could be through increasedcytosolic Ca2+ levels that activate the phosphorylation of thetranscription factor CREB (Konradi et al., 1994), whichmight result in an enhanced proENK gene transcription(Rajadhyaksha et al., 1999). However, since the proENKgene promoter also has an AP-1 site that can be activated byFos–Jun proteins (Monnier and Loeffler, 1998), thesetranscription factors might also be involved in the proENKgene regulation following AMPH. A recent study hasprovided evidence, that following AMPH, the DNA-bindingactivity of AP-1 was increased in the human astrocyte-likeU373 MG cells, concomitantly to the psychostimulant'seffects on the proENK gene regulation (Malaplate-Armandet al., 2005). In this work, the brain proENK mRNA levelswere not modified 4 days after an acute AMPH treatment,with the exception of the brainstem where a decrease wasobserved. The lack of effect of AMPH on proENK mRNA inmost of the CNS areas could be related to the long intervalbetween AMPH injection and the assay (i.e. 4 days in ourstudy vs. 3 h in the previous study mentioned above). Other
authors also failed to show changes in brain proENK mRNAfollowing morphine (Tjon et al., 1997) and cocaine (Daunaisand McGinty, 1994; Álvarez Fischer et al., 2001). Our dataalso indicate that c-fos mRNA levels were unchanged in anyregion tested after AMPH treatment. These results supportprevious data indicating that proENK mRNA may behavelike early gene mRNAs (Weisinger, 1995).
The measurement of other inducible proteins, such asFos-related proteins, showed a similar pattern of change tothe 32 kDa proENK band following AMPH. The highlystable isoform of ΔFosB was the only one that remainedelevated in PfC of AMPH-treated rats, in agreement withprevious data showing that after chronic exposure to AMPHor cocaine, a sustained increase in these protein levels wasobserved in specific brain areas for several weeks (Chen etal., 1997; Ehrlich et al., 2002; McClung et al., 2004). Ourresults suggest that although the other isoforms (c-Fos, Fra-1and -2, FosB) are soon activated by an AMPH treatment,only ΔFosB content may remain elevated after 4 days inPfC. It is important to address that this is the first evidence ofan increase of this protein after a single dose of AMPH. Inaddition, since a related constitutive protein such as PC1 didnot show any change in PfC after AMPH administration, it islikely that this drug exerts a specific effect on some proteinsrather than having a general influence on protein synthesis.
Interestingly, in this study we also showed that AMPHmodified the met-ENK in IS in a similar way to that observedin CNS. Our findings indicate an increase in free met-ENKlevels in immune organs such as the spleen and the thymusobservable 4 days after acute or repeated AMPH. Inagreement with this, an increased free met-ENK and free

Fig. 7. Levels of proENK, c-fos and GAPDH mRNA in immune organs of control rats (VEH) and rats that received an acute AMPH treatment (5 mg/kg i.p.)4 days before. (A) The amplified products of proENK cDNA (545 bp), c-fos cDNA (388 bp) and GAPDH cDNA (380 bp), as generated from mRNA, are shownin thymus. (B) Relative units of absorbance of proENK and c-fos mRNA. W: water; MW: molecular weight marker.
72 M.A. Assis et al. / Journal of Neuroimmunology 178 (2006) 62–75
syn-ENK level was observed in cultured splenocytes ofacutely and chronically stressed rats, suggesting that a stressstimulus also enhanced proENK-derived peptide releasefrom thymus, spleen and bone marrow macrophages(Saravia et al., 1998). Moreover, macrophages of chronicallystressed rats have been suggested to have a tonic increasedrelease of proENK products (Saravia et al., 1998). Thus,similar mechanisms may be triggered by stress and drugs onIS, as was previously described for behavioral andneurochemical effects (Kalivas and Stewart, 1991; Antelmanet al., 1980; Pacchioni et al., 2002, in press).
In the current study, the medium of cultured ratsplenocytes did not have detectable levels of free met-ENK, although it did contain proENK-derived peptides.These peptides increased following AMPH to a similarextent to that induced by the T-cell mitogen in control rats.Moreover, the addition of Con A in the splenocytes ofAMPH-treated rats only induced a low additional increase inproENK-derived peptides, indicating that the other spleencells (like macrophages and B-cells) had a minimalinfluence. This finding suggests that one dose of AMPHby itself was sufficient stimulus for splenic T-lymphocytes torelease proENK-derived peptides, as in the case of mitogenicactivation. Spleen and alveolar macrophages contained PC1(Vindrola et al., 1994), which is involved in proENKprocessing in endocrine cells (Rittner et al., 2001; Vindrolaand Lindberg, 1992). Moreover, a previous report indicatedthat peripheral blood monocytes, but not T-cells, produced
free met-ENK (Kuis et al., 1991; Roth et al., 1989). Sinceonly macrophages have the machinery (PC1) to produce lowmolecular weight biologically active opioid peptides, in thepresent study cryptic proENK-derived peptides may havebeen released by lymphocytes. Related to this, we demon-strated that the macrophages are specifically the cells thatshow an increase in met-ENK following acute or repeatedAMPH in the spleen culture.
Spleens of acute AMPH-treated rats had an increase in the44 kDa protein content, while the thymus showed anincrease in the 34–32 kDa doublet corresponding to proENKcontent. It could be that different turnover rates of proENKthroughout the immune organs are responsible for thesechanges.
ProENK mRNA has been described in uncultured ratbone marrow and cultured mouse bone marrow-derivedmacrophages (Saravia et al., 1993), in thymic CD4+ cellsafter 15-day activation with Con A (Linner et al., 1991), andin splenic monocyte–macrophage and B-cells (Saravia et al.,1993; Behar et al., 1991). A single dose of AMPH did notmodify the proENK or c-fos mRNA levels in IS which couldbe also attributed to the fact that the assays were performed4 days after the drug. The only exception was that AMPHinduced a decrease in c-fos mRNA levels in the thymuswhich could be related, at least in part, to the decreased T-proliferative response after the drug. Further studies shouldconfirm if the repeated AMPH treatment exerts any influenceon pro-ENK mRNA and protein content in IS and CNS.

73M.A. Assis et al. / Journal of Neuroimmunology 178 (2006) 62–75
This study demonstrates that a single dose, as well as arepeated treatment with AMPH, induces a dose-dependentdecreasing effect in the Con A-proliferative response 4 daysfollowing the AMPH injection. Since the spleen weight andcellularity were not modified following acute AMPH, it islikely that the drug is exerting a specific influence onlymphocyte responsivenessmore than an effect on the absolutenumber of cells. This is the first result showing a influence ofAMPH on IS observable 4 days later, which is consistent withprevious reports which indicated a decreased splenocyteproliferation immediately after a challenge with this drug(Kubera et al., 2002; Freire-Garabal et al., 1991). Previousfindings from our laboratory also showed that chronic AMPHtreatment induced a persistent sensitization to the stress-induced immunosuppressive effects (Basso et al., 1999).
Met-ENK synthesized by endocrine and/or nervous tissuehas been involved in stress-induced immunosuppression(Saravia et al., 1998; Roda et al., 1996; Roy and Loh, 1996).In rat spleen, only macrophages produced free met-ENK,which had led to suggest that these cells may be the source ofmet-ENK, and could participate in the modulation of theimmune response (Saravia et al., 1998). ProENK is notprocessed to met-ENK in bone marrow–mononuclear cells,while in thymus– and spleen–mononuclear cells met-ENKrepresented 10% and 26% of total met-ENK-containingpeptides, respectively (Saravia et al., 1998). In the currentstudy, it is possible that an enhanced met-ENK release fromperipheral monocyte/macrophages is actually the primer forthe inhibitory proliferative response of T-lymphocytesfollowing AMPH. In agreement with our present findings,antisense oligonucleotides against proENK reduced theopioid expression and enhanced the proliferation of ConA-stimulated splenocytes (Fulford et al., 2000). Furtherstudies are necessary to clarify the role played by central vs.peripheral met-ENK in the AMPH's effects on IS.
Besides met-ENK, other systems affected by AMPH couldalso be exerting an influence on IS. DA is the neurotransmitterprimarily involved in the long-term expression of behavioralsensitization to AMPH (Vanderschuren et al., 1999a) and thereis evidence of an immunomodulatory role for it (Basu andDasgupta, 2000). It is highly probable that the dose-dependentdecreasing effect on proliferative response showed in thisstudy may be also linked to this neurotransmitter. In vivostudies fromour lab have provided evidence of an involvementof DA in the chronic AMPH-induced enhancement of thestress-induced immunosupression (Basso et al., 1999).However, in the most of in vivo studies so far is has beenimpossible to separate the effects of the drug on DA of CNSfrom any that might target immune cells directly. Whendiscussing the possible contribution of DA, we should alsotake into account the peripheral role of this catecholamine inthe neuroimmune network. Thus, DA may act as a transmitterbetween splenic sympathetic nerve terminals and immuno-cytes (Bencsics et al., 1997), and it has been hypothesized thatendogenous DA detected in lymphocytes could underlieseveral functions (Cosentino et al., 2002, 2003).
Summing up, we conclude that similar biologicalmechanisms could be triggered by AMPH to underlie itseffects on brain and immune organs. We propose thatchanges in proENK-derived peptides in peripheral T-lymphocytes may be reflecting an opioid central effect ofAMPH. Furthermore, proENK-derived peptides could beconsidered as one of the main targets for changes in the ISfunctions induced by a psychostimulant drug. Since drugs ofabuse not only modify the behavior of the individuals, butalso compromise their immune functions, increasing thevulnerability to numerous infectious diseases (Baldwing etal., 1998; Friedman and Eisenstein, 2004), it is highlyrelevant to find similar biological changes in immune organsand brain and follow them up at peripheral level.
Acknowledgments
This work was supported by grants from Ministerio deSalud de la Nación, Agencia Córdoba Ciencia, CONICETand SeCyT (Argentina), and IBRO Latin-American Com-mittee. Dr Cancela, Dr Sotomayor and Dr Figuerola areSenior Researchers of CONICET (Argentina). The authorswish to express their sincere gratitude to Mrs. Estela Salde,Mrs. Paula Icely and Mrs. Elsa R. Pereyra for their excellenttechnical assistance, to Dr. German Roth for his collabora-tion with the micro cell harvester and to Dr. JacquelineMcGinty for kind suggestions to the manuscript.
References
Abarca, C., Albrecht, U., Spanagel, R., 2002. Cocaine sensitization andreward are under the influence of circadian genes and rhythm. Proc. Natl.Acad. Sci. U. S. A. 99 (13), 9026–9030.
Álvarez Fischer, D., Schafer, M.K., Ferger, B., Gross, S., Westermann, R.,Weihe, E., Kuschinsky, K., 2001. Sensitization to the behavioural effectsof cocaine: alterations in tyrosine hydroxylase or endogenous opioidmRNAs are not necessarily involved. Naunyn-Schmiedeberg's Arch.Pharmacol. 363 (3), 288–294.
Amenta, F., Bronzetti, E., Cantalamessa, F., El-Assouad, D., Felici, L.,Ricci, A., Tayebati, S.K., 2001. Identification of dopamine plasmamembrane and vesicular transporters in human peripheral bloodlymphocytes. J. Neuroimmunol. 177, 133–142.
Antelman, S.M., Eichler, A.J., Black, C.A., Kocan, D., 1980. Interchange-ability of stress and amphetamine in sensitization. Science 207,329–331.
Baldwing, G.C., Roth, M.D., Tashkin, D.P., 1998. Acute and chroniceffects of cocaine on the immune system and the possible link to AIDS.J. Neuroimmunol. 83, 133–138.
Basso, A.M., Gioino, G., Molina, V.A., Cancela, L.M., 1999. Chronicamphetamine facilitates immunosuppression in response to a novelaversive stimulus: Reversal by Haloperidol pretreatment. Pharmacol.Biochem. Behav. 62, 307–314.
Basu, S., Dasgupta, S., 2000. Dopamine, a neurotransmitter, influences theimmune system. J. Neuroimmunol. 102, 113–124.
Behar, O.Z., Ovadia, H., Polakiewicz, R.D., Abramsky, O., Rosen, H., 1991.Regulation of proenkephalin A messenger ribonucleic acid levels innormal B lymphocytes: specific inhibition by glucocorticoid hormonesand superinduction by cycloheximide. Endocrinology 129, 649–655.
Bencsics, A., Sershen, H., Baranyi, M., Hashim, A., Lajtha, A., Vizi, E.S.,1997. Dopamine, as well as norepinephrine, is a link betweennoradrenergic nerve terminals and splenocytes. Brain Res. 761, 236–343.

74 M.A. Assis et al. / Journal of Neuroimmunology 178 (2006) 62–75
Bergquist, J., Tarkowski, A., Ekman, R., Ewing, A., 1994. Discovery ofendogenous catecholamines in lymphocytes and evidence for catechol-amine regulation of lymphocyte function via an autocrine loop. Proc.Natl. Acad. Sci. U. S. A. 91, 12912–12916.
Borsook, D., Konradi, C., Falkowski, O., Comb, M., Hyman, S.E., 1994.Molecular mechanisms of stress-induced proenkephalin gene regulation:CREB interacts with the proenkephalin gene in the mouse hypothalamusand is phosphorylated in response to hyperosmolar stress. Mol.Endocrinol. 8, 240–248.
Brar, B.K., Lowry, P.J., 1999. The differential processing of proenkephalinA in mouse and human breast tumour cell lines. J. Endocrinol. 161,475–484.
Cador, M., Bjijou, Y., Cailhol, S., Stinus, L., 1999. D-Amphetamine-induced behavioral sensitization: implication of a glutamatergic medialprefrontal cortex–ventral tegmental area innervation. Neuroscience 94,705–721.
Chen, J., Kelz, M.B., Hope, B.T., Nakabeppu, Y., Nestler, E.J., 1997.Chronic Fos-related antigens: stable variants of ΔFosB induced in brainby chronic treatments. J. Neurosci. 17, 4933–4941.
Cosentino, M., Zaffaroni, M., Marino, F., Bombelli, R., Ferrari, M., Rasini,E., Lecchini, S., Ghezzi, A., Frigo, G., 2002. Catecholamine productionand tyrosine hydroxylase expression in peripheral blood mononuclearcells from multiple sclerosis patients: effects of cell stimulation andpossible relevance for activation-induced apoptosis. J. Neuroimmunol.133 (1–2), 233–240.
Cosentino, M., Marino, F., Bombelli, R., Ferrari, M., Lecchini, S., Frigo, G.,2003. Unravelling dopamine (and catecholamine) physiopharmacologyin lymphocytes: open questions. Trends Immunol. 24 (11), 581–582.
Daunais, J.B., McGinty, J.F., 1994. Acute and chronic cocaine administra-tion differentially alters striatal opioid and nuclear transcription factormRNAs. Synapse 18 (1), 35–45.
Diaz-Otañez, C.S., Capriles, N., Cancela, L.M., 1997. D1 and D2 dopaminereceptors are involved in the restraint stress induced sensitization to thepsychostimulant effects of amphetamine. Pharmacol. Biochem. Behav.58 (1), 9–14.
Ehrlich, M.E., Sommer, J., Canas, E., Unterwald, E.M., 2002. Periadoles-cent mice show enhanced ΔFosB upregulation in response to cocaineand amphetamine. J. Neurosci. 22 (21), 9155–9159.
Freire-Garabal, M., Balboa, J.L., Núñez, M.J., Castaño, M.T., Llovo, J.B.,Fernández-Rial, J.C., Belmonte, A., 1991. Effects of amphetamine on T-cell immune response in mice. Life Sci. 49 (16), PL107–PL112.
Friedman, H., Eisenstein, T.K., 2004. Neurological basis of drugdependence and its effects on the immune system. J. Neuroimmunol.147 (1–2), 106–108.
Fulford, A.J., Harbuz, M.S., Jessop, D.S., 2000. Antisense inhibition of pro-opiomelanocortin and proenkephalin A messenger RNA translationalters rat immune cell function in vitro. J. Neuroimmunol. 106 (1–2),6–13.
Galinowski, A., Levy-Soussan, P., Loo, H., 1992. Schizophrenia andimmunity. Ann. Med. Psychol. 150, 138–142.
Gordon, J., Barnes, N.M., 2003. Lymphocytes transport serotonin anddopamine: agony or ecstasy? Trends Immunol. 24 (8), 438–443.
Haus, E., Smolensky, M.H., 1999. Biologic rhythms in the immune system.Chronobiol. Int. 16 (5), 581–622.
Jankovic, B.D., Radulovic, J., 1992. Enkephalins, brain and immunity:modulation of immune responses by methionine-enkephalin injectedinto the cerebral cavity. Int. J. Neurosci. 67 (1–4), 241–270.
Kalivas, P.W., Stewart, J., 1991. Dopamine transmission in the initiation andexpression of drug- and stress-induced sensitization of motor activity.Brain Res. Rev. 16, 223–244.
Knych, E.T., Eisenberg, R.M., 1979. Effect of amphetamine on plasmacorticosterone in the conscious rat. Neuroendocrinology 29, 110–118.
Konradi, C., Cole, R.L., Heckers, S., Hyman, S.E., 1994. Amphetamineregulates gene expression in rat striatum via transcription factor CREB.J. Neurosci. 14 (9), 5623–5634.
Kubera, M., Filip, M., Basta-Kaim, A., Nowak, E., Budziszewska, B.,Tetich, M., Holan, V., Korzeniak, B., 2002. The effect of amphetamine
sensitization on mouse immunoreactivity. J. Physiol. Pharmacol. 53 (2),233–252.
Kuis, W., Villiger, P.M., Leser, H.G., Lotz, M., 1991. Eifferential processingof proenkephalin-A by human peripheral blood monocytes and Tlymphocytes. J. Clin. Invest. 88 (3), 817–824.
Lightman, S.L., Young, W.S., 1987. Changes in hypothalamic preproenke-phalin A mRNA following stress and opiate withdrawal. Nature 328,643–645.
Lindberg, I., Shaw, E., 1992. Posttranslational processing of proenkephalinin SK-N-MC cells: evidence for phosphorylation. J. Neurochem. 58,448–453.
Linner, K.M., Beyer, H.S., Sharp, B.M., 1991. Induction of the messengerribonucleic acid for proenkephalin A in cultured murine CD4-positivethymocytes. Endocrinology 128 (2), 717–724.
Linner, K.M., Quist, H.E., Sharp, B.M., 1995. Met-enkephalin-containingpeptides encoded by proenkephalin A mRNA expressed in activatedmurine thymocytes inhibit thymocyte proliferation. J. Immunol. 154(10), 5049–5060.
Malaplate-Armand, C., Becuwe, P., Ferrari, L., Masson, C., Dauca, M.,Visvikis, S., Lambert, H., Batt, A.M., 2005. Effect of acute and chronicpsychostimulant drugs on redox status, AP-1 activation and pro-enkephalin mRNA in the human astrocyte-like U373 MG cells.Neuropharmacology 48 (5), 673–684.
Mao, L., Wang, J.Q., 2003. Contribution of ionotropic glutamate receptorsto acute amphetamine-stimulated preproenkephalin mRNA expressionin the rat striatum in vivo. Neurosci. Lett. 31; 346 (1–2), 17–20.
Mathis, J.P., Lindberg, I., 1992. Posttranslational processing of proenke-phalin in AtT-20 cells: evidence for cleavage at a Lys-Lys site.Endocrinology 131 (5), 2287–2296.
McClung, C.A., Ulery, P.G., Perrotti, L.I., Zachariou, V., Berton, O., Nestler,E.J., 2004. ΔFosB: a molecular switch for long-term adaptation in thebrain. Brain Res. Mol. Brain Res. 20; 132 (2), 146–154.
McKenna, F., McLaughlin, P.J., Lewis, B.J., Sibbring, G.C., Cummerson, J.A., Bowen-Jones, D., Moots, R.J., 2002. Dopamine receptor expressionon human T- and B-lymphocytes, monocytes, neutrophils, eosinophilsand NK cells: a flow cytometric study. J. Neuroimmunol. 132, 34–40.
Monnier, D., Loeffler, J.P., 1998. Pituitary adenylate cyclase-activatingpolypeptide stimulates proenkephalin gene transcription through AP1-and CREB-dependent mechanisms. DNA Cell Biol. 17 (2), 151–159.
Nestler, E.J., 2001. Molecular neurobiology of addiction. Am. J. Addict. 10(3), 201–217.
Pacchioni, A.M., Gioino, G., Assis, M.A., Cancela, L.M., 2002. A singleexposure to restraint stress induces behavioral and neurochemicalsensitization to stimulating effects of amphetamine: involvement ofNMDA receptors. Ann. N. Y. Acad. Sci. 965, 233–246.
Pacchioni, A.M., Cador, M., Bregonzio, C., Cancela, L.M., in press. Aglutamate–dopamine interaction in the persistent enhanced response toamphetamine in nucleus accumbens core but not shell following a singlerestraint stress. Neuropsychopharmacology (Advanced online publica-tion 19 April 2006).
Padgett, D.A., Glaser, R., 2003. How stress influences the immune response.Trends Immunol. 24 (8), 444–448.
Padrós, M.R., Vindrola, O., Zunszain, P., Fainboin, L., Finkielman, S.,Nahmod, V.E., 1989. Mitogenic activation of human lymphocytesinduce the release of proENK derived peptides. Life Sci. 242,229-223.
Padrós, M.R., Saravia, F., Vindrola, O., 1995. Antibodies against the amino-terminal portion of pro-enkephalin inhibit DNA synthesis in humanperipheral mononuclear cells. J. Neuroimmunol. 61 (1), 79–83.
Pellegrino, T., Bayer, B.M., 1998a. Modulation of immune cell functionfollowing fluoxetine administration in rats. Pharmacol. Biochem. Behav.59 (1), 151–157.
Pellegrino, T., Bayer, B.M., 1998b. In vivo effects of cocaine on immunecell function. J. Neuroimmunol. 83, 139–147.
Pellegrino, T.C., Bayer, B.M., 2000. Specific serotonin reuptake inhibitor-induced decrease in lymphocyte activity requires endogenous serotoninrelease. Neuroimmunomodulation 8, 179–187.

75M.A. Assis et al. / Journal of Neuroimmunology 178 (2006) 62–75
Pierce, R.C., Kalivas, P.W., 1997a. A circuitry model of the expression ofbehavioral sensitization to amphetamine-like psychostimulants. BrainRes. Rev. 25, 192–216.
Pierce, R.C., Kalivas, P.W., 1997b. A circuitry model of the expression ofbehavioral sensitization to amphetamine-like psychostimulants. BrainRes. Rev. 25, 192–216.
Pruett, S.B., 2001. Quantitative aspects of stress-induced immunomodula-tion. Int. Immunopharmacol. 1, 507–520.
Rajadhyaksha, A., Barczak, A., Macias, W., Leveque, J.C., Lewis, S.E.,Konradi, C., 1999. L-Type Ca(2+) channels are essential for glutamate-mediated CREB phosphorylation and c-fos gene expression in striatalneurons. J. Neurosci. 1; 19 (15), 6348–6359.
Rittner, H.L., Brack, A., Machelska, H.S., Mousa, A., Bauer, M., Schäfer,M., Stein, C., 2001. Opioid peptide-expressing leukocytes: identifica-tion, recruitment, and simultaneously increasing inhibition of inflam-matory pain. Anesthesiology 95 (2), 500–508.
Robinson, T.E., Berridge, K.C., 2000. The psychology and neurobiology ofaddiction: an incentive-sensitization view. Addiction 95 (Supplement 2),S91–S117.
Roda, L.G., Bongiorno, L., Trani, E., Urbani, A., Marini, M., 1996. Positiveand negative immunomodulation by opioid peptides. Int. J. Immuno-pharmacol. 18, 1–16.
Roth, K.A., Lorenz, R.G., Unanue, R.A., Weaver, C.T., 1989. Non-opiateactive proenkephalin-derived peptides are secreted by T helper cells.FASEB J. 3 (12), 2401–2407.
Roy, S., Loh, H.H., 1996. Effects of opioids on the immune system.Neurochem. Res. 21, 1375–1386.
Saal, D., Dong, Y., Bonci, A., Malenka, R.C., 2003. Drugs of abuse andstress trigger a common report synaptic adaptation in dopamine neurons.Neuron 37, 1–20.
Saravia, F.A., Ase, A., Aloyz, R., Kleid, M.C., Ines, M., Vida, R., Nahmod,V.E., Vindrola, O., 1993. Differential post-translational processing ofproENK in rat bone marrow and spleen mononuclear cells: evidence forsynenkephalin cleavage. Endocrinolgy 132, 1431–1437.
Saravia, F., Padros, M.R., Ase, A., Aloyz, R., Duran, S., Vindrola, O., 1998.Differential response to a stress stimulus of proENK peptide content inimmune cells of naive and chronically stressed rats. Neuropeptides 32(4), 351–359.
Tjon, G.H., Voorn, P., Vanderschuren, L.J., de Vries, T.J., Michiels, N.H.,Jonker, A.J., Klop, H., Nestby, P., Mulder, A.H., Schoffelmeer, A.N.,1997. Delayed occurrence of enhanced striatal preprodynorphin geneexpression in behaviorally sensitized rats: differential long-term effectsof intermittent and chronic morphine administration. Neuroscience 76(1), 167–176.
Turchan, J., Maj, M., Przewlocka, B., Przewlocki, R., 2002. Effect ofcocaine and amphetamine on biosynthesis of proenkephalin and
prodynorphin in some regions of the rat limbic system. Pol. J.Pharmacol. 54 (4), 367–372.
Vanderschuren, L.J.M.J., Kalivas, P.W., 2000. Alterations in dopaminergicand glutamatergic transmission in the induction and expression ofbehavioral sensitization: a critical review of preclinical studies.Psychopharmacology (Berl) 151 (2–3), 99–120.
Vanderschuren, L.J.M.J., Schoffelmeer, A.N.M., Mulder, A.H., De Vries,T.J., 1999a. Dopaminergic mechanisms mediating the long-termexpression of locomotor sensitization following pre-exposure tomorphine or amphetamine. Psychopharmacology 143, 244–253.
Vanderschuren, L.J.M.J., Schmidt, E.D., De Vries, T.J., Van Moorsel, C.A.,Tilders, F.J., Schoffelmeer, A.N., 1999b. A single exposure toamphetamine is sufficient to induce long-term behavioral, neuroendo-crine, and neurochemical sensitization in rats. J. Neurosci. 19 (21),9579–9586.
Vezina, P., Lorrain, D.S., Arnold, G.M., Austin, D., Suto, N., 2002.Sensitization of midbrain dopamine neuron reactivity promotes thepursuit of amphetamine. J. Neurosci. 22 (11), 4654–4662.
Vindrola, O., Lindberg, I., 1992. Biosynthesis of the prohormone convertasemPC1 in AtT-20 cells. Mol. Endocrinol. 6 (7), 1088–1094.
Vindrola, O., Padrós, M.R., Sterin-Prync, A., Ase, A., Finkielman, S.,Nahmod, V.E., 1990. Proenkephalin system in human polymorphonu-clear cells. J. Clin. Invest. 86, 531–537.
Vindrola, O., Mayer, A.M., Citera, G., Spitzer, J.A., Espinoza, L.R., 1994.Prohormone convertases PC2 and PC3 in rat neutrophils and macro-phages. Neuropeptides 27, 235–244.
Vindrola, O., Chervin, A., Vitale, M., Mella, A.N., Aloyz, R., Basso, A.,1998. Elevated proenkephalin-derived peptide levels in ACTH-produc-ing adenomas: nucleus and cytoplasm localization. Endocrine 8 (3),231–240.
Wang, J.Q., McGinty, J.F., 1995. Alterations in striatal zif/268,preprodynorphin and preproenkephalin mRNA expression induced byrepeated amphetamine administration in rats. Brain Res. 6; 673 (2),262–274.
Wang, J.Q., McGinty, J.F., 1996. D1 and D2 receptor regulation ofpreproenkephalin and preprodynorphin mRNA in rat striatum followingacute injection of amphetamine or methamphetamine. Synapse 22 (2),114–122.
Weisinger, G., 1995. The transcriptional regulation of the preproenkephalingene. Biochem. J. 307, 617–629.
Woiciechowsky, C., Schöning, B., Lanksch, W.R., Volk, H.D., Döcke, W.D.,1999. Mechanisms of brain-mediated systemic anti-inflammatorysyndrome causing immunodepression. J. Mol. Med. 77, 769–780.
Wolf, M.E., 1998. The role of excitatory amino acids in behavioralsensitization to psychomotor stimulants. Prog. Neurobiol. 54,679–720.



![Antiapoptotic effects of delta opioid peptide [D-Ala2, D-Leu5]enkephalin in brain slices induced by oxygen-glucose deprivation](https://static.fdokumen.com/doc/165x107/631998d7e9c87e0c091032dc/antiapoptotic-effects-of-delta-opioid-peptide-d-ala2-d-leu5enkephalin-in-brain.jpg)
















