
Doxycycline is Neuroprotective Against Nigral Dopaminergic Degeneration by a Dual Mechanism...
11
Doxycycline is Neuroprotective Against Nigral Dopaminergic Degeneration by a Dual Mechanism Involving MMP-3 Yuri Cho Hyo Jin Son Eun-Mee Kim Ji Hyun Choi Sung Tae Kim In Jung Ji Dong Hee Choi Tong H. Joh Yoon Seong Kim Onyou Hwang Received: 20 October 2008 / Revised: 15 May 2009 / Accepted: 18 June 2009 / Published online: 7 July 2009 Ó Springer Science+Business Media, LLC 2009 Abstract In Parkinson disease (PD), the dopaminergic (DAergic) neurons in the substantia nigra undergo degen- eration. While the exact mechanism for the degeneration is still not completely understood, neuronal apoptosis and inflammation are thought to play roles. We have recently obtained evidence that matrix metalloproteinase (MMP)-3 plays a crucial role in the apoptotic signal in DAergic cells as well as activation of microglia. The present study tested whether doxycycline might modulate MMP-3 and provide neuroprotection of DAergic neurons. Doxycycline effec- tively suppressed the expression of MMP-3 induced in response to cellular stress in the DAergic CATH.a cells. This was accompanied by protection of CATH.a cells as well as primary cultured mesencephalic DAergic neurons via attenuation of apoptosis. The active form of MMP-3, released under the cell stress condition, was also decreased in the presence of doxycycline. In addition, doxycycline led to downregulation of MMP-3 in microglial BV-2 cells exposed to lipopolysaccharide (LPS). This was accompa- nied by suppression of production of nitric oxide and TNF- a, as well as gene expression of iNOS, TNF-a, IL-1b, and COX-2. In vivo, doxycycline provided neuroprotection of the nigral DAergic neurons following MPTP treatment, as assessed by tyrosine hydroxylase immunocytochemistry and silver staining, and suppressed microglial activation and astrogliosis as assessed by Iba-1 and GFAP immuno- chemistry, respectively. Taken together, doxycycline showed neuroprotective effect on DAergic system both in vitro and in vivo and this appeared to derive from anti- apoptotic and anti-inflammatory mechanisms involving downregulation of MMP-3. Keywords Doxycycline Á Matrix metalloproteinase-3 Á Parkinson’s disease Á Microglia Á Dopaminergic neurons Introduction In Parkinson’s disease (PD), the dopaminergic (DAergic) neurons in the substantia nigra (SN) undergo degeneration. These DAergic neurons are particularly vulnerable due to the presence of reactive oxygen species-generating mole- cules, including dopamine (DA) itself and iron, as well as low antioxidants. Various cellular stresses, therefore, can cause activation of cascade of events leading to apoptotic death of the DAergic neurons. In addition, SN contains a relatively high number of microglia (Kim et al. 2000) and neuroinflammation involving activated microglia plays an active role in the pathogenesis of PD (Sanchez-Pernaute et al. 2004; Walker and Lue 2005). Matrix metalloproteinase (MMP)-3 is a member of the MMP family, which has been widely recognized to be involved in degradation and remodeling of the extracellular matrix (Visse and Nagase 2003). More recently, evidence that MMP-3 plays a critical role in DAergic neurodegen- eration has been demonstrated. We have observed that MMP-3 participates in the apoptotic signaling in DAergic cells, and its pharmacological inhibition, suppression of Yuri Cho and Hyo Jin Son contributed equally. Y. Cho Á H. J. Son Á E.-M. Kim Á J. H. Choi Á S. T. Kim Á I. J. Ji Á D. H. Choi Á O. Hwang (&) Department of Biochemistry and Molecular Biology, University of Ulsan College of Medicine, 388-1 Pungnap-dong, Songpa-ku, Seoul 138-736, Korea e-mail: [email protected] T. H. Joh Á Y. S. Kim Department of Neurology and Neuroscience, Weill Medical College of Cornell University, New York, NY 10021, USA 123 Neurotox Res (2009) 16:361–371 DOI 10.1007/s12640-009-9078-1
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Doxycycline is Neuroprotective Against Nigral Dopaminergic Degeneration by a Dual Mechanism...

Doxycycline is Neuroprotective Against Nigral DopaminergicDegeneration by a Dual Mechanism Involving MMP-3
Yuri Cho Æ Hyo Jin Son Æ Eun-Mee Kim Æ Ji Hyun Choi ÆSung Tae Kim Æ In Jung Ji Æ Dong Hee Choi ÆTong H. Joh Æ Yoon Seong Kim Æ Onyou Hwang
Received: 20 October 2008 / Revised: 15 May 2009 / Accepted: 18 June 2009 / Published online: 7 July 2009
� Springer Science+Business Media, LLC 2009
Abstract In Parkinson disease (PD), the dopaminergic
(DAergic) neurons in the substantia nigra undergo degen-
eration. While the exact mechanism for the degeneration is
still not completely understood, neuronal apoptosis and
inflammation are thought to play roles. We have recently
obtained evidence that matrix metalloproteinase (MMP)-3
plays a crucial role in the apoptotic signal in DAergic cells
as well as activation of microglia. The present study tested
whether doxycycline might modulate MMP-3 and provide
neuroprotection of DAergic neurons. Doxycycline effec-
tively suppressed the expression of MMP-3 induced in
response to cellular stress in the DAergic CATH.a cells.
This was accompanied by protection of CATH.a cells as
well as primary cultured mesencephalic DAergic neurons
via attenuation of apoptosis. The active form of MMP-3,
released under the cell stress condition, was also decreased
in the presence of doxycycline. In addition, doxycycline
led to downregulation of MMP-3 in microglial BV-2 cells
exposed to lipopolysaccharide (LPS). This was accompa-
nied by suppression of production of nitric oxide and TNF-
a, as well as gene expression of iNOS, TNF-a, IL-1b, and
COX-2. In vivo, doxycycline provided neuroprotection of
the nigral DAergic neurons following MPTP treatment, as
assessed by tyrosine hydroxylase immunocytochemistry
and silver staining, and suppressed microglial activation
and astrogliosis as assessed by Iba-1 and GFAP immuno-
chemistry, respectively. Taken together, doxycycline
showed neuroprotective effect on DAergic system both in
vitro and in vivo and this appeared to derive from anti-
apoptotic and anti-inflammatory mechanisms involving
downregulation of MMP-3.
Keywords Doxycycline � Matrix metalloproteinase-3 �Parkinson’s disease � Microglia � Dopaminergic neurons
Introduction
In Parkinson’s disease (PD), the dopaminergic (DAergic)
neurons in the substantia nigra (SN) undergo degeneration.
These DAergic neurons are particularly vulnerable due to
the presence of reactive oxygen species-generating mole-
cules, including dopamine (DA) itself and iron, as well as
low antioxidants. Various cellular stresses, therefore, can
cause activation of cascade of events leading to apoptotic
death of the DAergic neurons. In addition, SN contains a
relatively high number of microglia (Kim et al. 2000) and
neuroinflammation involving activated microglia plays an
active role in the pathogenesis of PD (Sanchez-Pernaute
et al. 2004; Walker and Lue 2005).
Matrix metalloproteinase (MMP)-3 is a member of the
MMP family, which has been widely recognized to be
involved in degradation and remodeling of the extracellular
matrix (Visse and Nagase 2003). More recently, evidence
that MMP-3 plays a critical role in DAergic neurodegen-
eration has been demonstrated. We have observed that
MMP-3 participates in the apoptotic signaling in DAergic
cells, and its pharmacological inhibition, suppression of
Yuri Cho and Hyo Jin Son contributed equally.
Y. Cho � H. J. Son � E.-M. Kim � J. H. Choi �S. T. Kim � I. J. Ji � D. H. Choi � O. Hwang (&)
Department of Biochemistry and Molecular Biology, University
of Ulsan College of Medicine, 388-1 Pungnap-dong, Songpa-ku,
Seoul 138-736, Korea
e-mail: [email protected]
T. H. Joh � Y. S. Kim
Department of Neurology and Neuroscience, Weill Medical
College of Cornell University, New York, NY 10021, USA
123
Neurotox Res (2009) 16:361–371
DOI 10.1007/s12640-009-9078-1

expression by siRNA, and gene deletion all lead to pro-
tection of the cells (Choi et al. 2008). In addition, MMP-3
is released from apoptotic DAergic cells (Kim et al. 2005b;
Choi et al. 2008), and this in turn can cause activation of
microglia and production of proinflammatory cytokines
and superoxide (Kim et al. 2005b; Kim et al. 2007).
Furthermore, MMP-3 is produced inside the activated
microglia as well, and mediates the generation of proin-
flammatory molecules (Mun-Bryce et al. 2002; Nut-
tall et al. 2007; Woo et al. 2008). Taken together,
modulation of MMP-3 may be used to protect against
neurodegeneration via direct neuroprotection, downregu-
lation of its release, and/or suppression of neuroinflam-
matory responses.
Currently no therapy has been definitively shown to
provide neuroprotection for PD and efforts are being made
to find ways to delay or halter the DAergic neurodegen-
eration. Doxycycline, a semi-synthetic tetracycline deriv-
ative, has been reported to downregulate MMPs (Boyle
et al. 1998; Brown et al. 2004; Burggraf et al. 2007) via a
mechanism that is distinct from its antibiotic activity
(Golub et al. 1994; Sapadin and Fleischmajer 2006; Soory
2008). Therefore, it was possible that doxycycline might
suppress MMP-3 induction and thus provide neuroprotec-
tion via both anti-apoptotic and anti-inflammatory mecha-
nisms. Doxycycline has a high penetration rate into the
brain and cerebrospinal fluid (Andersson and Alestig
1976), a prerequisite for a drug for the central nervous
system.
The present study shows the evidence that doxycycline
indeed downregulates the cellular stress-induced MMP-3
expression and suppresses apoptosis in DAergic cells. It
also downregulates MMP-3 release from these cells and
induction of MMP-3 in microglia in vitro. In addition,
doxycycline provides protection of the nigral DAergic
neurons in a 1-methyl-4-phenyl-1,2,3,6-tetrahydropyrimi-
dine (MPTP)-induced mouse PD model.
Materials and Methods
Chemicals
Culture media, fetal bovine serum (FBS), L-glutamine,
trypsin/EDTA, and penicillin–streptomycin were pur-
chased from GibcoBRL (Gaithersburg, MD). Lipopoly-
saccharide (LPS), tetrahydrobiopterin (BH4), doxycycline,
and MPTP were purchased from Sigma Chemical (St.
Louis, MO). First strand cDNA synthesis kit for RT-PCR
was purchased from MBI Fermentas (Ontario, Canada).
Protein concentration of samples was determined by
Bradford assay (BioRad, Hempstead, UK). Antibodies used
were as follows: polyclonal anti-MMP-3 (R&D System,
Minneapolis, MN), anti-tyrosine hydroxylase (TH) (Protos,
New York, NY), anti-Iba-1 (Wako Chemicals, Osaka,
Japan) and anti-activated caspase-3 (Cell Signaling Tech-
nology, Beverly, MA) and monoclonal anti-TH (R&D
system), and anti-glial fibrillary acidic protein (GFAP; Cell
Signaling Technology, Beverly, MA) and anti-b-actin
(Santa Cruz Biotechnology, Santa Cruz, CA). Alexa Fluor
488 goat anti-rabbit IgG and Alexa Fluor 546 donkey anti-
mouse IgG were from Molecular Probes (Eugene, OR). In
situ cell death detection kit for TUNEL staining was from
Roche Diagnostics GmbH (Penzberg, Germany). Vecta-
stain ABC kit and biotinylated secondary antibodies were
from Vector Laboratories (Burlingame, CA). Ready-Set-
Go mouse TNF-a ELISA kit was from eBioscience (San
Diego, CA). Chemiluminescence detection system was
obtained from Pierce Chemical (Rockford, IL). All other
chemicals were reagent grade and were purchased from
Sigma Chemical or Merck (Rahway, NJ). [3H]DA was
from Amersham Biosciences (Piscataway, NJ) and Taq
polymerase was from Roche Applied Science (Indianapo-
lis, IN). Laminin, Trizol reagent, and superscript II reverse
transcriptase were obtained from Invitrogen (Carlsbad,
CA).
Animals
All procedures were performed in compliance with the
Laboratory Animal Manual of the University of Ulsan
Asan Institute for Life Sciences, which follow the guide-
lines set forth by the Society for Neuroscience. Male
C57Bl/6 mice and pregnant Sprague-Dawley rats were
obtained from Orient Charles River Technology (Seoul,
Korea).
Cell Cultures
The BV-2 mouse microglial cells, originally produced by
Dr. V. Bocchini (Blasi et al. 1990) were grown and
maintained in Dulbecco’s modified Eagle’s medium with
10% FBS, 100 IU/l penicillin, and 10 lg/ml streptomycin
at 37�C in a humidified incubator under 5% CO2. For
experiments, BV-2 cells were plated at a density of
2.5 9 105 cells/well in 24-well polystyrene plates or
2.6 9 106 cells/60 mm plate. CATH.a cells (Suri et al.
1993) were grown in RPMI 1640 containing 8% horse
serum, 4% FBS, and penicillin–streptomycin. They were
plated at a density of 1.5 9 105 cells/well in 24-well
polystyrene plates or 1.5 9 106 cells/60 mm plate. After
24 h, the cells were fed with fresh medium and treated. For
primary culture of mesencephalic neurons, the ventral
mesencephalon was removed from 14-day gestation rat
embryo and incubated with 0.01% trypsin in HBSS for
15 min at 37�C. After trituration, 3 9 105 cells were plated
362 Neurotox Res (2009) 16:361–371
123

on each polystyrene cover slide that had been precoated
with 100 lg/ml poly-L-lysine and 4 lg/ml laminin and
placed in a 24-well culture plate. The cells were maintained
at 37�C in a humidified atmosphere with 5% CO2 in neu-
robasal medium supplemented with B-27, 2 mM gluta-
mine, 100 IU/l penicillin, and 10 lg/ml streptomycin. On
day 5 or 6 in vitro, the neurons were fed with fresh medium
and treated.
Cell Staining
For immunofluorescence staining, cells were fixed in cold
4% paraformaldehyde in 0.1 M phosphate buffered saline
(PBS), pH 7.4 for 30 min at room temperature. After
washing twice in PBS, the cells were incubated for 1 h in
blocking solution (5% FBS and 0.3% Triton X-100 in 0.1
M PBS), followed by overnight incubation with appropri-
ate primary antibody diluted in incubation solution (5%
FBS and 0.2% Triton X-100 in 0.1 M PBS) at 4�C. After
two washes in PBS, the samples were incubated at room
temperature for 1 h with appropriate fluorescence-labeled
secondary antibody in the incubation solution. For TH
immunostaining, mouse monoclonal anti-TH antibody
(1:5009) and Alexa Fluor 546 goat anti-mouse IgG
(1:2009) were used. For caspase-3, rabbit polyclonal anti-
caspase-3 antibody (1:1009) and Alexa Fluor 488 goat
anti-rabbit IgG (1:2009) were used. As a negative control,
the samples were incubated with the secondary antibody
only. The cells were washed in PBS, mounted on a glass
slide, and viewed by confocal microscopy (TCS-ST2;
Leica, Wetzlar, Germany). TUNEL staining was performed
according to the manufacturer’s protocol using fluorescein
and analyzed by confocal microscopy.
DA Uptake Assay
The primary cultured neurons were rinsed with Krebs–
Ringer solution and incubated in Krebs–Ringer solution
containing 1 mM ascorbate, 2 mM b-alanine, and 100 lM
pargyline at 37�C for 5 min in a humidified 5% CO2
atmosphere. The neurons were further incubated in the
presence of 25 nM [3H]DA (45 Ci/mmol) for 15 min.
Control neurons were treated in the same solution but
containing the DA uptake blocker bupropion (10 lM). The
cells were then washed with ice-cold Krebs–Ringer solu-
tion and lysed with 0.5 M NaOH. Radioactivity was
measured using a liquid scintillation counter (Beckman,
Fullerton, CA).
RT-PCR
Reverse transcription (RT) reaction was performed using
5 lg total RNA isolated from BV-2 or CATH.a cells.
Polymerase chain reaction (PCR) was performed, using the
cDNA product as a template, at 94�C for 1 min, 60�C for
1 min, and 72�C for 1 min for MMP-3 and at 94�C for
30 s, 60�C for 30 s, and 72�C for 30 s for the others. After
30 cycles of amplification, the PCR product was subjected
to agarose gel electrophoresis. RT-PCR against b2M was
performed as an internal control. Analysis of PCR products
on 1.5% agarose gels showed single-band amplification
products with respective expected sizes. The following
primers were used for PCR: MMP-3 (forward, CTGG
TACCAACCTATTCCTGGTTGC; reverse, TCTCTTTAT
TAGAAATGGCAGCATCG), iNOS (forward, ATGTCC
GAAGCAAACATCAC; reverse, TAATGTCCAGGAAG
TAGGTG), TNF-a (forward, CAGACCCTCACACTCA-
GATCATCTT; reverse, CAGAGCAATGACTCCAAAG-
TAGACCT), IL-1b (forward, ATGGCAACTGTTCCTG
AACTCAACT; reverse, CAGGACAGGTATAGATTCT
TTCCTTT), COX-2 (forward, CAGCAAATCCTTGCT
GTTCC; reverse, TGGGCAAAGAATGCAAACATC), and
b2M (forward, GCTATCCAGAAAACCCCTCAA; reverse,
CATGTCTCGATCCCAGTAGACGGT).
Western Blot
Equal amounts (20 lg) of protein were separated on 10%
SDS polyacrylamide gel and transferred onto polyvinyli-
dene difluoride-nitrocellulose filters membrane. Mem-
branes were blocked for 1 h in 20 mM Tris-HCl containing
137 mM NaCl and 0.05% Tween 20 (TBST) containing
5% skim milk at room temperature, incubated overnight
with primary antibody against MMP-3 (1:1000) or b-actin
(1:1000) at 4�C followed by horseradish peroxidase-con-
jugated secondary antibodies for 1 h at room temperature.
Protein bands were detected by chemiluminescence.
Lactate Dehydrogenase (LDH) Assay
Degrees of cell death were assessed by activity of LDH
released into the culture medium using the method previ-
ously described (Choi et al. 2004). Aliquots (50 ll) of cell
culture medium were incubated at room temperature in the
presence of 0.26 mM NADH, 2.87 mM sodium pyruvate,
and 100 mM potassium phosphate buffer (pH 7.4) in a total
volume of 200 ll. The rate of NAD? formation was
monitored for 5 min at 2-s intervals at 340 nm using a
microplate spectrophotometer (SPECTRA MAX 340 pc;
Molecular Devices, Menlo Park, CA).
Griess Reaction
Accumulated nitrite, an oxidative metabolite of NO, was
measured in the cell supernatant by the Griess reaction as
Neurotox Res (2009) 16:361–371 363
123

previously described (Seo et al. 2005). Briefly, 200 ll
aliquots of cell medium was mixed with 100 ll of Griess
reagent (1% sulfanilamide and 0.1% naphthylethylenedi-
amine dihydrochloride in 2.5% H3PO4) in a 96-well
microtiter plate, and the absorbance was read at 540 nm
using the plate reader. The nitrite concentration was
determined using a curve calibrated with sodium nitrite
standards.
Measurement of TNF-a Levels
TNF-a was measured by ELISA according to the protocol
provided by the manufacturer. The concentration of TNF-awas calculated according to a standard curve.
MPTP Mice
Male C57Bl/6 mice weighing 23–25 g were maintained in
a temperature- and humidity-controlled room with a 12-h
light–dark cycle and food and water available ad libitum.
The animals were divided into four groups: saline treated
control, MPTP treated, MPTP plus doxycycline treated,
and doxycycline treated groups (n = 10). Doxycycline was
administered intraperitoneally at 45 mg/kg twice at 12-h
interval. The first doxycycline administration was done
30 min before the first MPTP injection. MPTP was injected
intraperitoneally 4 times in a single day, 2 h apart at
20 mg/kg dissolved in saline. The animals were killed
7 days after the first injection; they were deeply anesthe-
tized (80 mg/kg ketamine and 20 mg/kg xylazine, injected
intraperitoneally) and transcardially fixed in 4% parafor-
maldehyde. Brains were promptly removed and postfixed
in 4% paraformaldehyde. After cryoprotection, the tissue
was cut into 20-lm sections on a vibrating-blade micro-
tome (VT 1000S; Leica, Nussloch, Germany), which were
stored in 0.08% sodium azide in PBS at 4�C until analysis.
Immunohistochemistry
Immunohistochemistry of the nigral and striatal brain tis-
sue sections was performed as described previously
(Hwang et al. 1998), using antisera against TH (1:1,0009),
Iba-1 (1:3009), or GFAP (1:1009), followed by Vecta-
stain ABC kit and biotinylated secondary antibody.
Quantitation of the TH-positive neurons was done by the
method described previously (Kim et al. 2006).
Amino-Cupric-Silver Staining
Amino-cupric-silver degeneration staining was carried out
by the method as described previously (de Olmos et al.
1994; Kim et al. 2005a). For counterstaining, the sections
previously stained and mounted on gelatin-coated slides
were immersed in 0.5% neutral red for 15 min, rinsed
twice in distilled H2O, dehydrated, cleared in pure xylene,
and coverslipped using DPX as a mounting medium. For
quantitation, we randomly selected a total of 2500 small
circles (10 lm in diameter) from microscopic fields of SN
and striatum (1009 magnification) sections. Their optical
density was measured using Science Lab 2001 Image
Gauge Version 4.0 software (Fuji Photo, Tokyo, Japan). To
account for differences in background staining intensity,
the background optical density was represented by the
average of the smallest 25 values in 2500 circles, the mean
of which was subtracted from the optical density of silver
stain-positive area to provide the final optical density
value.
Data Analyses
Data are expressed as mean ± SEM of independent
experiments. Comparisons of three of more groups were
analyzed by one-way ANOVA (analysis of variance) and
Student-Newman-Keuls comparison test. Statistical tests
were carried out using PRISM (GraphPad Software, San
Diego, CA). A value of P \ 0.05 was considered statisti-
cally significant.
Results
Doxycycline Downregulates MMP-3 Gene Expression
in DAergic Cells
We have previously established that exposure to BH4, an
endogenous molecule present in DAergic cells serving as a
cofactor for DA synthesis, causes oxidative stress (Choi
et al. 2000; Choi et al. 2003; Lee et al. 2007) and upreg-
ulation of MMP-3 (Choi et al. 2008) in cultured DAergic
CATH.a cells as well as primary cultured mesencephalic
DAergic neurons. Using this system, we first tested whe-
ther doxycycline might suppress the MMP-3 induction. As
shown by the RT-PCR data in Fig. 1a, the dramatic
upregulation of MMP-3 mRNA by the exposure to BH4
was suppressed by the co-treatment with doxycycline. At
1 lM doxycycline, the MMP-3 induction was completely
blocked. Accurate quantitation could not be done because
the bands corresponding to MMP-3 for untreated and
doxycycline-treated cells were barely detectable.
The suppression of MMP-3 induction by doxycycline
was also apparent at the protein level, as determined by
western blot analysis (Fig. 1b). We have previously
observed that under this cellular stress condition, most
MMP-3 is cleaved to the catalytically active 48 kD protein
(actMMP-3) (Choi et al. 2008). Whereas BH4 caused a
dramatic increase in actMMP-3 (6.86 ± 0.72 fold), this
364 Neurotox Res (2009) 16:361–371
123
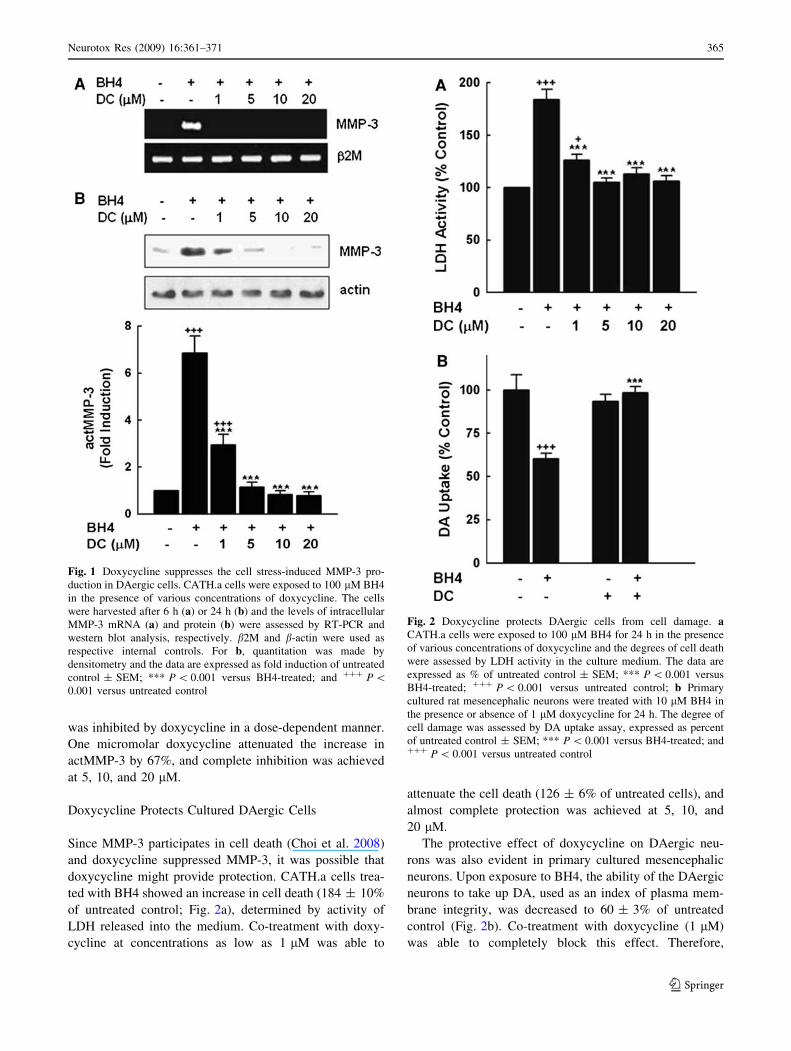
was inhibited by doxycycline in a dose-dependent manner.
One micromolar doxycycline attenuated the increase in
actMMP-3 by 67%, and complete inhibition was achieved
at 5, 10, and 20 lM.
Doxycycline Protects Cultured DAergic Cells
Since MMP-3 participates in cell death (Choi et al. 2008)
and doxycycline suppressed MMP-3, it was possible that
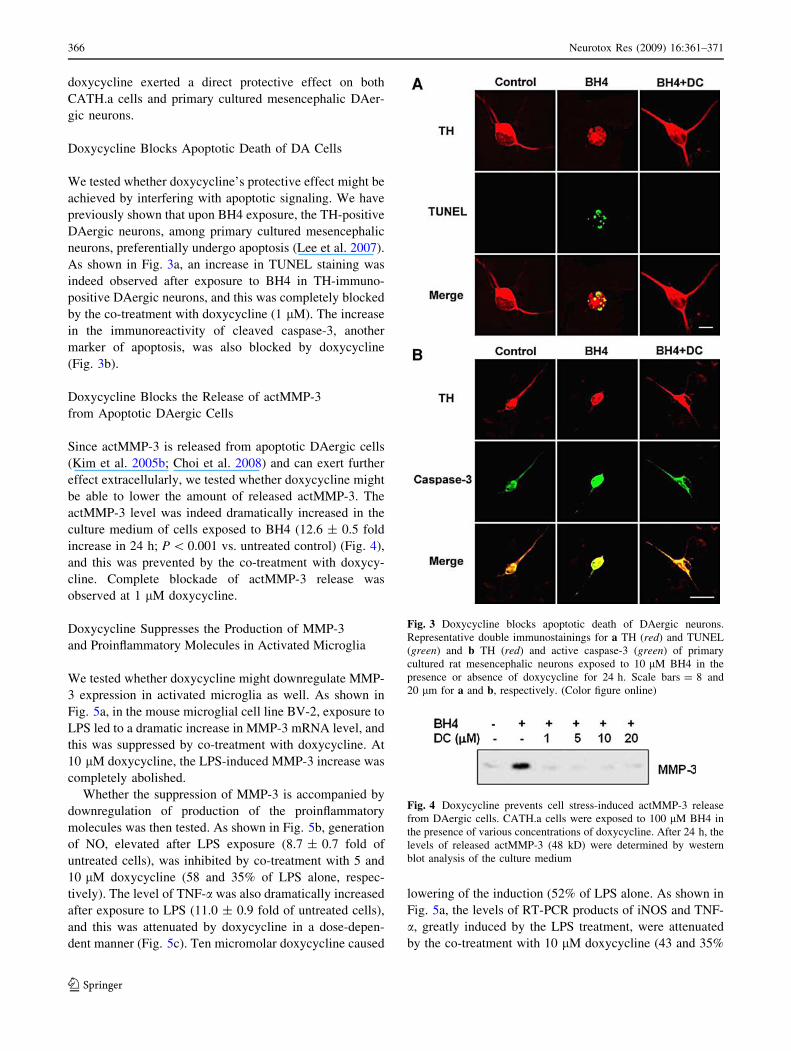
doxycycline might provide protection. CATH.a cells trea-
ted with BH4 showed an increase in cell death (184 ± 10%
of untreated control; Fig. 2a), determined by activity of
LDH released into the medium. Co-treatment with doxy-
cycline at concentrations as low as 1 lM was able to
attenuate the cell death (126 ± 6% of untreated cells), and
almost complete protection was achieved at 5, 10, and
20 lM.
The protective effect of doxycycline on DAergic neu-
rons was also evident in primary cultured mesencephalic
neurons. Upon exposure to BH4, the ability of the DAergic
neurons to take up DA, used as an index of plasma mem-
brane integrity, was decreased to 60 ± 3% of untreated
control (Fig. 2b). Co-treatment with doxycycline (1 lM)
was able to completely block this effect. Therefore,
Fig. 1 Doxycycline suppresses the cell stress-induced MMP-3 pro-
duction in DAergic cells. CATH.a cells were exposed to 100 lM BH4
in the presence of various concentrations of doxycycline. The cells
were harvested after 6 h (a) or 24 h (b) and the levels of intracellular
MMP-3 mRNA (a) and protein (b) were assessed by RT-PCR and
western blot analysis, respectively. b2M and b-actin were used as
respective internal controls. For b, quantitation was made by
densitometry and the data are expressed as fold induction of untreated
control ± SEM; *** P \ 0.001 versus BH4-treated; and ??? P \0.001 versus untreated control
Fig. 2 Doxycycline protects DAergic cells from cell damage. aCATH.a cells were exposed to 100 lM BH4 for 24 h in the presence
of various concentrations of doxycycline and the degrees of cell death
were assessed by LDH activity in the culture medium. The data are
expressed as % of untreated control ± SEM; *** P \ 0.001 versus
BH4-treated; ??? P \ 0.001 versus untreated control; b Primary
cultured rat mesencephalic neurons were treated with 10 lM BH4 in
the presence or absence of 1 lM doxycycline for 24 h. The degree of
cell damage was assessed by DA uptake assay, expressed as percent
of untreated control ± SEM; *** P \ 0.001 versus BH4-treated; and??? P \ 0.001 versus untreated control
Neurotox Res (2009) 16:361–371 365
123
doxycycline exerted a direct protective effect on both
CATH.a cells and primary cultured mesencephalic DAer-
gic neurons.
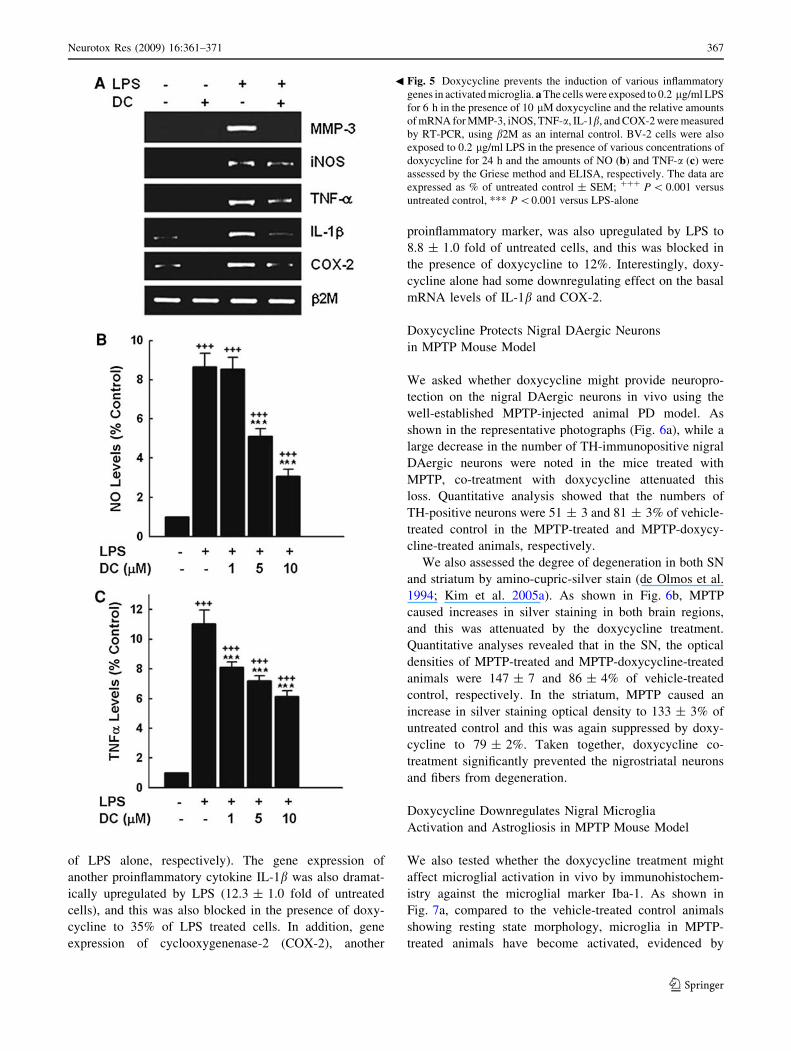
Doxycycline Blocks Apoptotic Death of DA Cells
We tested whether doxycycline’s protective effect might be
achieved by interfering with apoptotic signaling. We have
previously shown that upon BH4 exposure, the TH-positive
DAergic neurons, among primary cultured mesencephalic
neurons, preferentially undergo apoptosis (Lee et al. 2007).
As shown in Fig. 3a, an increase in TUNEL staining was
indeed observed after exposure to BH4 in TH-immuno-
positive DAergic neurons, and this was completely blocked
by the co-treatment with doxycycline (1 lM). The increase
in the immunoreactivity of cleaved caspase-3, another
marker of apoptosis, was also blocked by doxycycline
(Fig. 3b).
Doxycycline Blocks the Release of actMMP-3
from Apoptotic DAergic Cells
Since actMMP-3 is released from apoptotic DAergic cells
(Kim et al. 2005b; Choi et al. 2008) and can exert further
effect extracellularly, we tested whether doxycycline might
be able to lower the amount of released actMMP-3. The
actMMP-3 level was indeed dramatically increased in the
culture medium of cells exposed to BH4 (12.6 ± 0.5 fold
increase in 24 h; P \ 0.001 vs. untreated control) (Fig. 4),
and this was prevented by the co-treatment with doxycy-
cline. Complete blockade of actMMP-3 release was
observed at 1 lM doxycycline.
Doxycycline Suppresses the Production of MMP-3
and Proinflammatory Molecules in Activated Microglia
We tested whether doxycycline might downregulate MMP-
3 expression in activated microglia as well. As shown in
Fig. 5a, in the mouse microglial cell line BV-2, exposure to
LPS led to a dramatic increase in MMP-3 mRNA level, and
this was suppressed by co-treatment with doxycycline. At
10 lM doxycycline, the LPS-induced MMP-3 increase was
completely abolished.
Whether the suppression of MMP-3 is accompanied by
downregulation of production of the proinflammatory
molecules was then tested. As shown in Fig. 5b, generation
of NO, elevated after LPS exposure (8.7 ± 0.7 fold of
untreated cells), was inhibited by co-treatment with 5 and
10 lM doxycycline (58 and 35% of LPS alone, respec-
tively). The level of TNF-a was also dramatically increased
after exposure to LPS (11.0 ± 0.9 fold of untreated cells),
and this was attenuated by doxycycline in a dose-depen-
dent manner (Fig. 5c). Ten micromolar doxycycline caused
lowering of the induction (52% of LPS alone. As shown in
Fig. 5a, the levels of RT-PCR products of iNOS and TNF-
a, greatly induced by the LPS treatment, were attenuated
by the co-treatment with 10 lM doxycycline (43 and 35%
Fig. 3 Doxycycline blocks apoptotic death of DAergic neurons.
Representative double immunostainings for a TH (red) and TUNEL
(green) and b TH (red) and active caspase-3 (green) of primary
cultured rat mesencephalic neurons exposed to 10 lM BH4 in the
presence or absence of doxycycline for 24 h. Scale bars = 8 and
20 lm for a and b, respectively. (Color figure online)
Fig. 4 Doxycycline prevents cell stress-induced actMMP-3 release
from DAergic cells. CATH.a cells were exposed to 100 lM BH4 in
the presence of various concentrations of doxycycline. After 24 h, the
levels of released actMMP-3 (48 kD) were determined by western
blot analysis of the culture medium
366 Neurotox Res (2009) 16:361–371
123
of LPS alone, respectively). The gene expression of
another proinflammatory cytokine IL-1b was also dramat-
ically upregulated by LPS (12.3 ± 1.0 fold of untreated
cells), and this was also blocked in the presence of doxy-
cycline to 35% of LPS treated cells. In addition, gene
expression of cyclooxygenenase-2 (COX-2), another
proinflammatory marker, was also upregulated by LPS to
8.8 ± 1.0 fold of untreated cells, and this was blocked in
the presence of doxycycline to 12%. Interestingly, doxy-
cycline alone had some downregulating effect on the basal
mRNA levels of IL-1b and COX-2.
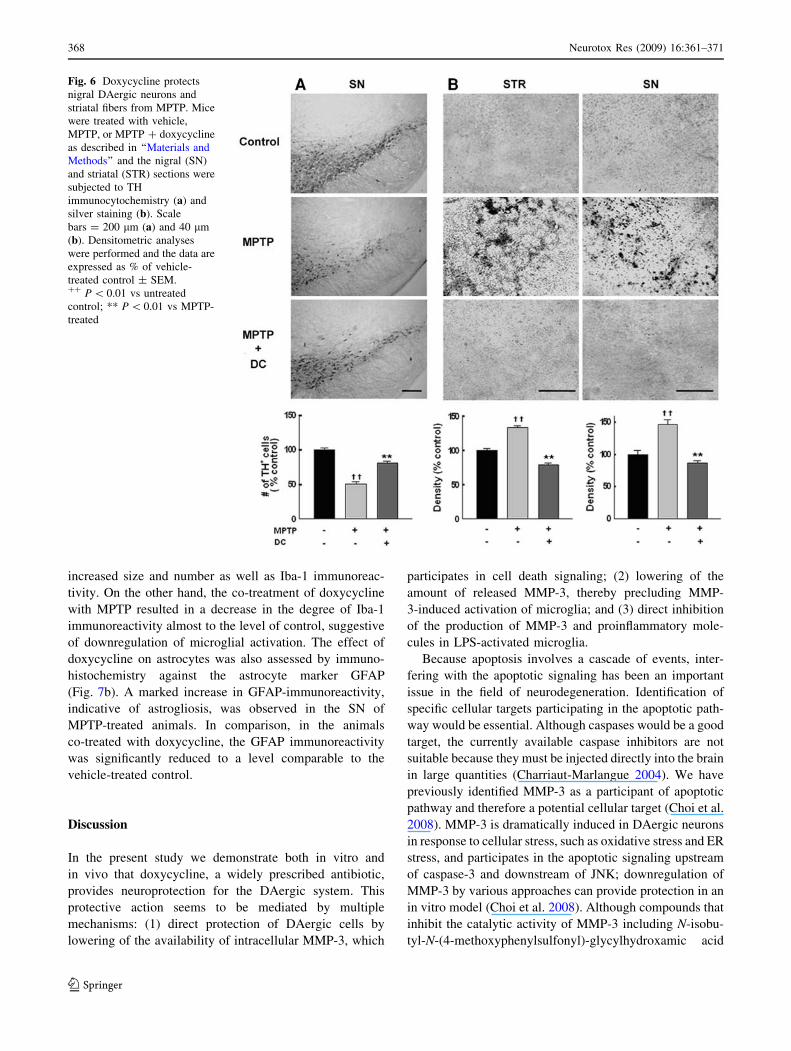
Doxycycline Protects Nigral DAergic Neurons
in MPTP Mouse Model
We asked whether doxycycline might provide neuropro-
tection on the nigral DAergic neurons in vivo using the
well-established MPTP-injected animal PD model. As
shown in the representative photographs (Fig. 6a), while a
large decrease in the number of TH-immunopositive nigral
DAergic neurons were noted in the mice treated with
MPTP, co-treatment with doxycycline attenuated this
loss. Quantitative analysis showed that the numbers of
TH-positive neurons were 51 ± 3 and 81 ± 3% of vehicle-
treated control in the MPTP-treated and MPTP-doxycy-
cline-treated animals, respectively.
We also assessed the degree of degeneration in both SN
and striatum by amino-cupric-silver stain (de Olmos et al.
1994; Kim et al. 2005a). As shown in Fig. 6b, MPTP
caused increases in silver staining in both brain regions,
and this was attenuated by the doxycycline treatment.
Quantitative analyses revealed that in the SN, the optical
densities of MPTP-treated and MPTP-doxycycline-treated
animals were 147 ± 7 and 86 ± 4% of vehicle-treated
control, respectively. In the striatum, MPTP caused an
increase in silver staining optical density to 133 ± 3% of
untreated control and this was again suppressed by doxy-
cycline to 79 ± 2%. Taken together, doxycycline co-
treatment significantly prevented the nigrostriatal neurons
and fibers from degeneration.
Doxycycline Downregulates Nigral Microglia
Activation and Astrogliosis in MPTP Mouse Model
We also tested whether the doxycycline treatment might
affect microglial activation in vivo by immunohistochem-
istry against the microglial marker Iba-1. As shown in
Fig. 7a, compared to the vehicle-treated control animals
showing resting state morphology, microglia in MPTP-
treated animals have become activated, evidenced by
Fig. 5 Doxycycline prevents the induction of various inflammatory
genes in activated microglia. a The cells were exposed to 0.2 lg/ml LPS
for 6 h in the presence of 10 lM doxycycline and the relative amounts
of mRNA for MMP-3, iNOS, TNF-a, IL-1b, and COX-2 were measured
by RT-PCR, using b2M as an internal control. BV-2 cells were also
exposed to 0.2 lg/ml LPS in the presence of various concentrations of
doxycycline for 24 h and the amounts of NO (b) and TNF-a (c) were
assessed by the Griese method and ELISA, respectively. The data are
expressed as % of untreated control ± SEM; ??? P \ 0.001 versus
untreated control, *** P \0.001 versus LPS-alone
b
Neurotox Res (2009) 16:361–371 367
123
increased size and number as well as Iba-1 immunoreac-
tivity. On the other hand, the co-treatment of doxycycline
with MPTP resulted in a decrease in the degree of Iba-1
immunoreactivity almost to the level of control, suggestive
of downregulation of microglial activation. The effect of
doxycycline on astrocytes was also assessed by immuno-
histochemistry against the astrocyte marker GFAP
(Fig. 7b). A marked increase in GFAP-immunoreactivity,
indicative of astrogliosis, was observed in the SN of
MPTP-treated animals. In comparison, in the animals
co-treated with doxycycline, the GFAP immunoreactivity
was significantly reduced to a level comparable to the
vehicle-treated control.
Discussion
In the present study we demonstrate both in vitro and
in vivo that doxycycline, a widely prescribed antibiotic,
provides neuroprotection for the DAergic system. This
protective action seems to be mediated by multiple
mechanisms: (1) direct protection of DAergic cells by
lowering of the availability of intracellular MMP-3, which
participates in cell death signaling; (2) lowering of the
amount of released MMP-3, thereby precluding MMP-
3-induced activation of microglia; and (3) direct inhibition
of the production of MMP-3 and proinflammatory mole-
cules in LPS-activated microglia.
Because apoptosis involves a cascade of events, inter-
fering with the apoptotic signaling has been an important
issue in the field of neurodegeneration. Identification of
specific cellular targets participating in the apoptotic path-
way would be essential. Although caspases would be a good
target, the currently available caspase inhibitors are not
suitable because they must be injected directly into the brain
in large quantities (Charriaut-Marlangue 2004). We have
previously identified MMP-3 as a participant of apoptotic
pathway and therefore a potential cellular target (Choi et al.
2008). MMP-3 is dramatically induced in DAergic neurons
in response to cellular stress, such as oxidative stress and ER
stress, and participates in the apoptotic signaling upstream
of caspase-3 and downstream of JNK; downregulation of
MMP-3 by various approaches can provide protection in an
in vitro model (Choi et al. 2008). Although compounds that
inhibit the catalytic activity of MMP-3 including N-isobu-
tyl-N-(4-methoxyphenylsulfonyl)-glycylhydroxamic acid
Fig. 6 Doxycycline protects
nigral DAergic neurons and
striatal fibers from MPTP. Mice
were treated with vehicle,
MPTP, or MPTP ? doxycycline
as described in ‘‘Materials and
Methods’’ and the nigral (SN)
and striatal (STR) sections were
subjected to TH
immunocytochemistry (a) and
silver staining (b). Scale
bars = 200 lm (a) and 40 lm
(b). Densitometric analyses
were performed and the data are
expressed as % of vehicle-
treated control ± SEM.?? P \ 0.01 vs untreated
control; ** P \ 0.01 vs MPTP-
treated
368 Neurotox Res (2009) 16:361–371
123

(NNGH) are available and would be effective in vitro, their
high cost and low penetration into the brain make them
unsuitable as a neuroprotective drug. In addition, adminis-
tration of MMP-3-specific siRNA would involve a rather
invasive method. We report in the present study that the drug
doxycycline, which is known to enter the brain efficiently
(Andersson and Alestig 1976), can downregulate MMP-3
and provide DAergic neuroprotection.
We show the evidence that doxycycline provides direct
protection on DAergic neurons and that this is via inhib-
iting apoptosis, as demonstrated by the decreases in
TUNEL staining and active caspase-3. Doxycycline’s
protective effect has been observed in stroke models pre-
viously, and this has been thought to occur mainly by
inhibiting microglial activation. Lai and Todd (2006)
reported, in hypoxia-activated microglia model, that
doxycycline confers neuroprotection by regulating mi-
croglial, not neuronal, factors. We show for the first time
using isolated and glia-free cell culture systems that
doxycycline has a direct protective effect on DAergic cells
as well.
That actMMP-3 is released from apoptotic DAergic
cells (Kim et al. 2005b; Choi et al. 2008) and this in turn
causes microglial activation (Kim et al. 2005b) has been
shown. We observe in the present study that the release of
actMMP-3 is decreased in the presence of doxycycline.
The decrease in the amount of released actMMP may be
due to the lowered intracellular availability. However, one
cannot completely eliminate the possibility that a mecha-
nism for actMMP-3 release that is separate from but
coordinated with the intracellular apoptotic signaling may
exist. In fact, actMMP-3 release was observed to be com-
pletely abolished at 1 lM doxycycline (Fig. 4), but this
attenuated the intracellular actMMP-3 accumulation by
only 50% (Fig. 1b).
The proinflammatory and cytotoxic substances released
from activated microglia is thought to exert secondary
damage to nearby neurons that are otherwise healthy.
Because MMP-3 seems to play an important role in neur-
oinflammatory responses by eliciting the production of
TNF-a, IL-1b, and NO (Mun-Bryce et al. 2002; Kim et al.
2005b; Cauwe et al. 2007; Woo et al. 2008), the lowering
of its availability may preclude the secondary and perpetual
damage caused by neuroinflammation. The source of
MMP-3 activity can be activated microglia themselves as
well as the apoptotic DAergic neurons. MMP-3 expression
is increased in activated microglia (Rosenberg 2002; Woo
et al. 2008; the present data), and both actMMP-3 protein
and MMP-3 catalytic activity have been observed in the
medium of LPS-activated microglia (Woo et al. 2008). We
show in the present study that doxycycline suppresses the
LPS-induced increase in MMP-3 gene expression and that
this is accompanied by the downregulation of production of
NO and TNF-a, and the expression of iNOS, TNF-a, and
IL-1b.
Neuroprotective effect of doxycycline on DAergic
neurons was confirmed in vivo, in the MPTP-treated ani-
mal model of PD. The MPTP-induced degeneration of the
nigral DAergic neurons was attenuated by the co-treatment
with doxycycline. We have previously shown that MPP?,
the metabolite of MPTP, induced expression of MMP-3 in
primary cultured mesencephalic DAergic neurons and that
the nigral DAergic neurons of MMP-3 knockout animals
were relatively resistant to MPTP (Kim et al. 2007). Thus,
the protective effect of doxycycline shown in the MPTP-
treated animals is likely due to combination of both sup-
pression of apoptotic signal in the neurons and suppression
of microglial production of proinflammatory molecules,
both involving MMP-3 downregulation. Astrogliosis in the
MPTP-treated animals was also found to be suppressed by
the doxycycline co-treatment. It is possible that the soluble
factors that are released from activated microglia and
stimulate astrogliosis (Mohri et al. 2006; Rohl et al. 2007;
Zhang et al. 2009) are reduced under the condition that
microglia activation is suppressed by the doxycycline
administration.
Because doxycycline also suppresses other MMPs, it is
possible that inhibition of their activities also contributes to
the neuroprotective effect of doxycycline. However, we
have observed that in our BH4-induced CATH.a cell death
Fig. 7 Doxycycline suppresses microglial activation and astrogliosis
in SN of MPTP treated mice. Mice were treated with vehicle, MPTP,
or MPTP ? doxycycline as described in ‘‘Materials and Methods’’
and the nigral sections were subjected to Iba-1 (a) and GFAP (b)
immunohistochemistry. Scale bars = 50 lm
Neurotox Res (2009) 16:361–371 369
123

model (Choi et al. 2008) as well as ER stress model (Kim
et al. unpublished results), no changes were observed in the
mRNA levels of MMP-1, MMP-7, and MMP-9, belonging
to the collagenase, matrilysin, and gelatinase subtypes,
respectively, as well as MMP-2, which is usually present
ubiquitously. Therefore, at least in our cell system, the
levels of other MMP subtypes do not seem to be different
between healthy and apoptotic conditions and therefore
may not play an important role in the cell death
mechanism.
It is well known that another tetracycline derivation,
minocycline, also possesses the neuroprotective effect as
doxycycline (Stirling et al. 2005; Domercq and Matute
2004; Thomas et al. 2003). However, the drug is also
known to cause side effects such as vestibular toxicity
(Cunha 2006) and effects on bone and teeth formation.
During the on-going clinical trial for PD, the issue of
decreased tolerability of minocycline has recently been
raised to be a concern (NINDS NET-PD Investigators
2008). That is, due to dizziness, nausea, headache, joint
pain, and tooth discoloration, 23% of subjects prematurely
discontinued the minocycline treatment during the study
period. On the other hand, doxycycline has the least toxic
side effects of the tetracyclines (Toth et al. 1988; Smith and
Leyden 2005). Although doxycycline and minocycline
have similar chemical structures, they have been reported
to exhibit differences in patterns of neuroprotection in
some models of cerebral ischemia (Yrjanheikki et al. 1998;
Fox et al. 2005; Jantzie et al. 2005) and in antioxidant
activity (Kraus et al. 2005). In addition, doxycycline is a
more potent inhibitor of MMPs (Gilbertson-Beadling et al.
1995; Yao et al. 2007).
The effective concentration of doxycycline at which the
protective effect is observed in our hands is in the range of
concentration of plasma doxycycline in human [(3.75–
19.62 lM (Prall et al. 2002); 1.83–13.18 lM (Smith et al.
2008)]. Therefore, doxycycline, which is already available
in clinical situations and has protective action with minor
side effects, good tolerance, and excellent blood brain
barrier penetration, may be considered as a candidate
toward the therapeutic strategy for PD.
Acknowledgment This work was supported by Brain Research
Center of the 21st Century Frontier Research Program of the Ministry
of Science & Technology (2009K0012510) to O. Hwang.
References
Andersson H, Alestig K (1976) The penetration of doxycycline into
CSF. Scand J Infect Dis Suppl 9:17–19
Blasi E, Barluzzi R, Bocchini V, Mazzolla R, Bistoni F (1990)
Immortalization of murine microglial cells by a v-raf/v-myc
carrying retrovirus. J Neuroimmunol 27:229–237
Boyle JR, McDermott E, Crowther M, Willis AD, Bell PR, Thompson
MM (1998) Doxycycline inhibits elastin degradation and reduces
metalloproteinase activity in a model of aneurysmal disease.
J Vasc Surg 27:354–361
Brown DL, Desai KK, Vakili BA, Nouneh C, Lee HM, Golub LM
(2004) Clinical and biochemical results of the metalloproteinase
inhibition with subantimicrobial doses of doxycycline to prevent
acute coronary syndromes (MIDAS) pilot trial. Arterioscler
Thromb Vasc Biol 24:733–738
Burggraf D, Trinkl A, Dichgans M, Hamann GF (2007) Doxycycline
inhibits MMPs via modulation of plasminogen activators in focal
cerebral ischemia. Neurobiol Dis 25:506–513
Cauwe B, Van den Steen PE, Opdenakker G (2007) The biochemical,
biological, and pathological kaleidoscope of cell surface sub-
strates processed by matrix metalloproteinases. Crit Rev Bio-
chem Mol Biol 42:113–185
Charriaut-Marlangue C (2004) Apoptosis: a target for neuroprotec-
tion. Therapie 59:185–190
Choi HJ, Jang YJ, Kim HJ, Hwang O (2000) Tetrahydrobiopterin is
released from and causes preferential death of catecholaminergic
cells by oxidative stress. Mol Pharmacol 58:633–640
Choi HJ, Kim SW, Lee SY, Hwang O (2003) Dopamine-dependent
cytotoxicity of tetrahydrobiopterin: a possible mechanism for
selective neurodegeneration in Parkinson’s disease. J Neurochem
86:143–152
Choi HJ, Lee SY, Cho Y, Hwang O (2004) JNK activation by
tetrahydrobiopterin: implication for Parkinson’s disease. J Neu-
rosci Res 75:715–721
Choi DH, Kim EM, Son HJ, Joh TH, Kim YS, Kim D, Flint Beal M,
Hwang O (2008) A novel intracellular role of matrix metallo-
proteinase-3 during apoptosis of dopaminergic cells. J Neuro-
chem 106:405–415
Cunha BA (2006) New uses for older antibiotics: nitrofurantoin,
amikacin, colistin, polymyxin B, doxycycline, and minocycline
revisited. Med Clin North Am 90:1089–1107
de Olmos JS, Beltramino CA, de Lorenzo S (1994) Use of an amino-
cupric-silver technique for the detection of early and semiacute
neuronal degeneration caused by neurotoxicants, hypoxia, and
physical trauma. Neurotoxicol Teratol 16:545–561
Domercq M, Matute C (2004) Neuroprotection by tetracyclines.
Trends Pharmacol Sci 25:609–612
Fox C, Dingman A, Derugin N, Wendland MF, Manabat C, Ji S,
Ferriero DM, Vexler ZS (2005) Minocycline confers early but
transient protection in the immature brain following focal
cerebral ischemia-reperfusion. J Cereb Blood Flow Metab 25:
1138–1149
Gilbertson-Beadling S, Powers EA, Stamp-Cole M, Scott PS, Wallace
TL, Copeland J, Petzold G, Mitchell M, Ledbetter S, Poorman R
(1995) The tetracycline analogs minocycline and doxycycline
inhibit angiogenesis in vitro by a non-metalloproteinase-
dependent mechanism. Cancer Chemother Pharmacol 36:
418–424
Golub LM, Evans RT, McNamara TF, Lee HM, Ramamurthy NS
(1994) Non-antimicrobial tetracycline inhibits gingival matrix
metalloproteinases in Porphyromonas gingivalis-induced peri-
odontitis in rats. Ann NY Acad Sci 732:96–111
Hwang O, Baker H, Gross S, Joh TH (1998) Localization of GTP
cyclohydrolase in monoaminergic but not nitric oxide-producing
cells. Synapse 28:140–153
Jantzie LL, Cheung PY, Todd KG (2005) Doxycycline reduces
cleaved caspase-3 and microglial activation in an animal model
of neonatal hypoxia-ischemia. J Cereb Blood Flow Metab 25:
314–324
Kim WG, Mohney RP, Wilson B, Jeohn GH, Liu B, Hong JS (2000)
Regional difference in susceptibility to lipopolysaccharide-
370 Neurotox Res (2009) 16:361–371
123

induced neurotoxicity in the rat brain: role of microglia.
J Neurosci 20:6309–6316
Kim ST, Choi JH, Chang JW, Kim SW, Hwang O (2005a)
Immobilization stress causes increases in tetrahydrobiopterin,
dopamine, and neuromelanin and oxidative damage in the
nigrostriatal system. J Neurochem 95:89–98
Kim YS, Kim SS, Cho JJ, Choi DH, Hwang O, Shin DH, Chun HS,
Beal MF, Joh TH (2005b) Matrix metalloproteinase-3: a novel
signaling proteinase from apoptotic neuronal cells that activates
microglia. J Neurosci 25:3701–3711
Kim ST, Choi JH, Kim D, Hwang O (2006) Increased tyrosine
hydroxylase and neuromelanin in the SN of middle aged mice.
Neurosci Lett 396:263–268
Kim YS, Choi DH, Block ML, Lorenzl S, Yang L, Kim YJ, Sugama
S, Cho BP, Hwang O, Browne SE, Kim SY, Hong JS, Beal MF,
Joh TH (2007) A pivotal role of matrix metalloproteinase-3
activity in dopaminergic neuronal degeneration via microglial
activation. FASEB J 21:179–187
Kraus RL, Pasieczny R, Lariosa-Willingham K, Turner MS, Jiang A,
Trauger JW (2005) Antioxidant properties of minocycline:
neuroprotection in an oxidative stress assay and direct radical-
scavenging activity. J Neurochem 94:819–827
Lai AY, Todd KG (2006) Hypoxia-activated microglial mediators of
neuronal survival are differentially regulated by tetracyclines.
Glia 53:809–816
Lee SY, Moon Y, Hee Choi D, Jin Choi H, Hwang O (2007)
Particular vulnerability of rat mesencephalic dopaminergic
neurons to tetrahydrobiopterin: Relevance to Parkinson’s dis-
ease. Neurobiol Dis 25:112–120
Mohri I, Taniike M, Taniguchi H, Kanekiyo T, Aritake K, Inui T,
Fukumoto N, Eguchi N, Kushi A, Sasai H, Kanaoka Y, Ozono K,
Narumiya S, Suzuki K, Urade Y (2006) Prostaglandin D2-
mediated microglia/astrocyte interaction enhances astrogliosis
and demyelination in twitcher. J Neurosci 9:4383–4393
Mun-Bryce S, Lukes A, Wallace J, Lukes-Marx M, Rosenberg GA
(2002) Stromelysin-1 and gelatinase A are upregulated before
TNF-alpha in LPS-stimulated neuroinflammation. Brain Res
933:42–49
NINDS NET-PD Investigators (2008) A pilot clinical trial of creatine
and minocycline in early Parkinson disease: 18-month results.
Clin Neuropharmacol 31:141–150
Nuttall RK, Silva C, Hader W, Bar-Or A, Patel KD, Edwards DR,
Yong VW (2007) Metalloproteinases are enriched in microglia
compared with leukocytes and they regulate cytokine levels in
activated microglia. Glia 55:516–526
Prall AK, Longo GM, Mayhan WG, Waltke EA, Fleckten B, Thompson
RW, Baxter BT (2002) Doxycycline in patients with abdominal
aortic aneurysms and in mice: Comparison of serum levels and
effect on aneurysm growth in mice. J Vasc Surgery 35:923–929
Rohl C, Lucius R, Sievers J (2007) The effect of activated microglia
on astrogliosis parameters in astrocyte cultures. Brain Res
1129:43–52
Rosenberg GA (2002) Matrix metalloproteinases in neuroinflamma-
tion. Glia 39:279–291
Sanchez-Pernaute R, Ferree A, Cooper O, Yu M, Brownell AL,
Isacson O (2004) Selective COX-2 inhibition prevents progres-
sive dopamine neuron degeneration in a rat model of Parkinson’s
disease. J Neuroinflammation 1:6
Sapadin AN, Fleischmajer R (2006) Tetracyclines: nonantibiotic
properties and their clinical implications. Am Acad Dermatol
54:258–265
Seo JW, Srisook E, Son HJ, Hwang O, Cha YN, Chi DY (2005)
Syntheses of NAMDA derivatives inhibiting NO production in
BV-2 cells stimulated with lipopolysaccharide. Bioorg Med
Chem Lett 15:3369–3373
Smith K, Leyden JJ (2005) Safety of doxycycline and minocycline: a
systematic review. Clin Ther 27:1329–1342
Smith VA, Khan-Lim D, Anderson L, Cook SD, Dick AD (2008)
Does orally administered doxycycline reach the tear film? Br J
Ophthalmol 92:856–859
Soory M (2008) A role for non-antimicrobial actions of tetracyclines
in combating oxidative stress in periodontal and metabolic
diseases: a literature review. Open Dent J 2:5–12
Stirling DP, Koochesfahani KM, Steeves JD, Tetzlaff W (2005)
Minocycline as a neuroprotective agent. Neuroscientist 11:308–
322
Suri C, Fung BP, Tischler AS, Chikaraishi DM (1993) Catecholam-
inergic cell lines from the brain and adrenal glands of tyrosine
hydroxylase-SV40 T antigen transgenic mice. J Neurosci 13:
1280–1291
Thomas M, Le WD, Jankovic J (2003) Minocycline and other
tetracycline derivatives: a neuroprotective strategy in Parkin-
son’s disease and Huntington’s disease. Clin Neuropharmacol
26:18–23
Toth A, Lesser ML, Naus G, Brooks C, Adams D (1988) Effect of
doxycycline on pre-menstrual syndrome: a double-blind random-
ized clinical trial. J Int Med Res 16:270–279
Visse R, Nagase H (2003) Matrix metalloproteinases and tissue
inhibitors of metalloproteinases: structure, function, and bio-
chemistry. Circ Res 92:827–839
Walker DG, Lue LF (2005) Investigations with cultured human
microglia on pathogenic mechanisms of Alzheimer’s disease and
other neurodegenerative diseases. J Neurosci Res 81:412–425
Woo MS, Park JS, Choi IY, Kim WK, Kim HS (2008) Inhibition of
MMP-3 or -9 suppresses lipopolysaccharide-induced expression
of proinflammatory cytokines and iNOS in microglia. J Neuro-
chem 106:770–780
Yao JS, Shen F, Young WL, Yang GY (2007) Comparison of
doxycycline and minocycline in the inhibition of VEGF-induced
smooth muscle cell migration. Neurochem Int 50:524–530
Yrjanheikki J, Keinanen R, Pellikka M, Hokfelt T, Koistinaho J (1998)
Tetracyclines inhibit microglial activation and are neuroprotective
in global brain ischemia. Proc Natl Acad Sci USA 95:15769–
15774
Zhang D, Hu X, Qian L, Wilson B, Lee C, Flood P, Langenbach R,
Hong JS (2009) Prostaglandin E2 released from activated
microglia enhances astrocyte proliferation in vitro. Toxicol Appl
Pharmacol 238(1):64–70
Neurotox Res (2009) 16:361–371 371
123




















