
A Novel ADP/ATP Transporter in the Mitosome of the Microaerophilic Human Parasite Entamoeba...
10
Current Biology, Vol. 15, 737–742, April 26, 2005, ©2005 Elsevier Ltd All rights reserved. DOI 10.1016/j.cub.2005.02.068 A Novel ADP/ATP Transporter in the Mitosome of the Microaerophilic Human Parasite Entamoeba histolytica Ka Wai Chan, 1,5 Dirk-Jan Slotboom, 1,5 Sian Cox, 3 ses confirm that the Entamoeba ADP/ATP carrier is distinct from archetypal mitochondrial ADP/ATP carri- T. Martin Embley, 2, * Olivier Fabre, 1 Mark van der Giezen, 3,6 Marilyn Harding, 1 ers, an observation that is supported by its different substrate and inhibitor specificity. Because many David S. Horner, 4 Edmund R.S. Kunji, 1, * Gloria León-Avila, 3,7 and Jorge Tovar 3 functions of yeast and human mitochondria rely on solutes transported by specialized members of this 1 Dunn Human Nutrition Unit Medical Research Council family, the Entamoeba mitosome must contain only a small subset of these processes requiring adenine Hills Road Cambridge CB2 2XY nucleotide exchange. United Kingdom 2 School of Biology The Devonshire Building Results and Discussion University of Newcastle upon Tyne Newcastle NE1 7RU Identification of an MCF Homolog on the Entamoeba histolytica Genome United Kingdom 3 School of Biological Sciences We identified a homolog of the mitochondrial carrier family (MCF) on the Entamoeba histolytica genome (TIGR) Royal Holloway University of London with BLAST searches. The translated protein contains a tripartite structure [ 8], signature motifs (IPR001993), and Egham, Surrey TW20 0EX United Kingdom other features typical of MCF members (Figure 1A). The Entamoeba carrier contains 276 amino acid residues 4 Dipartimento di Scienze Biomolecolare e Biotecnologie and is therefore the smallest MCF member so far iden- tified, and the loop regions linking the transmembrane University of Milan Via Celoria 26 a-helices are extremely short [9](Figure 1B). Most eu- karyotes have many mitochondrial carriers, typically 20133 Milan Italy between 30 and 60, that transport different substrates required or produced by the mitochondrion [7]. In con- trast, the genomes of the malaria parasite Plasmodium Summary falciparum [10] and the intestinal parasite Cryptospori- dum parvum [11] have only nine and five mitochondrial Recent data suggest that microaerophilic and para- carriers, respectively. This may reflect the propensity of sitic protozoa, which lack oxidative phosphorylation, parasites to use host metabolites rather than to make nevertheless contain mitochondrial homologs [1–6], their own, leading to the elimination of metabolic path- organelles that share common ancestry with mito- ways and transport steps surplus to requirements. The chondria. Such widespread retention suggests there reductionist tendency has been taken much further by may be a common function for mitochondrial homo- Entamoeba histolytica because we found only a single logs that makes them essential for eukaryotic cells. mitochondrial carrier on its genome. We determined the mitochondrial carrier family (MCF) complement of the Entamoeba histolytica mitochon- drial homolog, also known as a crypton [5] or more Localization of the Entamoeba MCF Protein commonly as a mitosome [3]. MCF proteins support The Entamoeba mitosome imports chaperonin 60 mitochondrial metabolic energy generation, DNA rep- (Cpn60) [3, 5], a protein that is of a-proteobacterial an- lication, and amino-acid metabolism by linking bio- cestry and is typically found in mitochondria, where it chemical pathways in the mitochondrial matrix with is involved in the ATP-dependent folding of organellar those in the cytosol [7]. MCF diversity thus closely proteins [12]. The Entamoeba Cpn60 clusters with the mirrors important facets of mitochondrial metabolic Dictyostelium mitochondrial protein in phylogenetic diversity. The Entamoeba histolytica mitosome has analyses [13], and it has an amino-acid extension sim- lost all but a single type of MCF protein, which trans- ilar to known mitochondrial targeting signals [3, 14]. ports ATP and ADP via a novel mechanism that is not Deletion of this extension prevents import of Cpn60 reliant on a membrane potential. Phylogenetic analy- into the mitosome, but import can be restored by addi- tion of a functional mitochondrial-targeting signal from Trypanosoma cruzi [3]. *Correspondence: [email protected] (T.M.E.); ek@mrc-dunn. Western blotting (Figure 2A) showed that the Enta- cam.ac.uk (E.R.S.K.) 5 These authors contributed equally to this work. moeba MCF protein occurred in the same Entamoeba 6 Present address: School of Biological Sciences, Queen Mary Uni- cell fractions as those containing Cpn60, strongly sug- versity of London, Mile End Road, London E1 4NS, United gesting that it is in the same compartment. Mitochon- Kingdom. drial carriers are targeted to, and inserted into, the 7 Present address: Genetics and Molecular Biology Department, mitochondrial inner membrane via a second import Centro de Investigación y de Estudios Avanzados del Instituto Poli- técnico Nacional, Zacatenco, 07360 Mexico City, Mexico. pathway that does not require an N-terminal leader se-
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of A Novel ADP/ATP Transporter in the Mitosome of the Microaerophilic Human Parasite Entamoeba...

Current Biology, Vol. 15, 737–742, April 26, 2005, ©2005 Elsevier Ltd All rights reserved. DOI 10.1016/j.cub.2005.02.068
A Novel ADP/ATP Transporterin the Mitosome of the MicroaerophilicHuman Parasite Entamoeba histolytica
Ka Wai Chan,1,5 Dirk-Jan Slotboom,1,5 Sian Cox,3
T. Martin Embley,2,* Olivier Fabre,1
Mark van der Giezen,3,6 Marilyn Harding,1
David S. Horner,4 Edmund R.S. Kunji,1,*Gloria León-Avila,3,7 and Jorge Tovar3
1Dunn Human Nutrition UnitMedical Research CouncilHills RoadCambridge CB2 2XYUnited Kingdom2School of BiologyThe Devonshire BuildingUniversity of Newcastle upon TyneNewcastle NE1 7RUUnited Kingdom3School of Biological SciencesRoyal HollowayUniversity of LondonEgham, Surrey TW20 0EXUnited Kingdom4Dipartimento di Scienze Biomolecolare e
BiotecnologieUniversity of MilanVia Celoria 2620133 MilanItaly
Summary
Recent data suggest that microaerophilic and para-sitic protozoa, which lack oxidative phosphorylation,nevertheless contain mitochondrial homologs [1–6],organelles that share common ancestry with mito-chondria. Such widespread retention suggests theremay be a common function for mitochondrial homo-logs that makes them essential for eukaryotic cells.We determined the mitochondrial carrier family (MCF)complement of the Entamoeba histolytica mitochon-drial homolog, also known as a crypton [5] or morecommonly as a mitosome [3]. MCF proteins supportmitochondrial metabolic energy generation, DNA rep-lication, and amino-acid metabolism by linking bio-chemical pathways in the mitochondrial matrix withthose in the cytosol [7]. MCF diversity thus closelymirrors important facets of mitochondrial metabolicdiversity. The Entamoeba histolytica mitosome haslost all but a single type of MCF protein, which trans-ports ATP and ADP via a novel mechanism that is notreliant on a membrane potential. Phylogenetic analy-
*Correspondence: [email protected] (T.M.E.); [email protected] (E.R.S.K.)
5 These authors contributed equally to this work. 6 Present address: School of Biological Sciences, Queen Mary Uni-versity of London, Mile End Road, London E1 4NS, UnitedKingdom. 7 Present address: Genetics and Molecular Biology Department,Centro de Investigación y de Estudios Avanzados del Instituto Poli-técnico Nacional, Zacatenco, 07360 Mexico City, Mexico.ses confirm that the Entamoeba ADP/ATP carrier isdistinct from archetypal mitochondrial ADP/ATP carri-ers, an observation that is supported by its differentsubstrate and inhibitor specificity. Because manyfunctions of yeast and human mitochondria rely onsolutes transported by specialized members of thisfamily, the Entamoeba mitosome must contain onlya small subset of these processes requiring adeninenucleotide exchange.
Results and Discussion
Identification of an MCF Homologon the Entamoeba histolytica GenomeWe identified a homolog of the mitochondrial carrierfamily (MCF) on the Entamoeba histolytica genome (TIGR)with BLAST searches. The translated protein contains atripartite structure [8], signature motifs (IPR001993), andother features typical of MCF members (Figure 1A). TheEntamoeba carrier contains 276 amino acid residuesand is therefore the smallest MCF member so far iden-tified, and the loop regions linking the transmembranea-helices are extremely short [9] (Figure 1B). Most eu-karyotes have many mitochondrial carriers, typicallybetween 30 and 60, that transport different substratesrequired or produced by the mitochondrion [7]. In con-trast, the genomes of the malaria parasite Plasmodiumfalciparum [10] and the intestinal parasite Cryptospori-dum parvum [11] have only nine and five mitochondrialcarriers, respectively. This may reflect the propensity ofparasites to use host metabolites rather than to maketheir own, leading to the elimination of metabolic path-ways and transport steps surplus to requirements. Thereductionist tendency has been taken much further byEntamoeba histolytica because we found only a singlemitochondrial carrier on its genome.
Localization of the Entamoeba MCF ProteinThe Entamoeba mitosome imports chaperonin 60(Cpn60) [3, 5], a protein that is of a-proteobacterial an-cestry and is typically found in mitochondria, where itis involved in the ATP-dependent folding of organellarproteins [12]. The Entamoeba Cpn60 clusters with theDictyostelium mitochondrial protein in phylogeneticanalyses [13], and it has an amino-acid extension sim-ilar to known mitochondrial targeting signals [3, 14].Deletion of this extension prevents import of Cpn60into the mitosome, but import can be restored by addi-tion of a functional mitochondrial-targeting signal fromTrypanosoma cruzi [3].
Western blotting (Figure 2A) showed that the Enta-moeba MCF protein occurred in the same Entamoebacell fractions as those containing Cpn60, strongly sug-gesting that it is in the same compartment. Mitochon-drial carriers are targeted to, and inserted into, themitochondrial inner membrane via a second importpathway that does not require an N-terminal leader se-

Current Biology738
Figure 1. Sequence Analysis of the Adenine Nucleotide Carrier from Entamoeba histolytica
(A) The putative membrane topology of the carrier; the schematic representation indicates the key residues of the signature motif (red circles)and the three amino-acid sequence repeats (bordered by blue dashed lines) that are typical for members of the mitochondrial carrier family.Colored circles represent the amino-acid residues of the Entamoeba carrier; these residues correspond to the bovine ADP/ATP carrierresidues that are involved in binding of carboxy-atractyloside. Shown in green are amino-acid residues that are involved in the binding of thesulfate groups of carboxy-atractyloside; in blue, residues that bind the carboxy group of the inhibitor; and in purple, residues that have vander Waals interactions [9].(B) Alignment of the amino sequence of the Entamoeba adenine nucleotide carrier (top), the bovine ADP/ATP carrier (middle), and the yeastADP/ATP carrier 3 (AAC3) (bottom). The black bars indicate the position of the transmembrane regions in the amino-acid sequence asdeduced from the structure of the bovine ADP/ATP carrier [9]. The red bars indicate key amino-acid residues of the signature motif. The blue,green, and purple bars indicate the position of the key residues involved in the binding of carboxy-atractyloside by the bovine ADP/ATPcarrier as in (A).
quence [15]. The Entamoeba carrier was expressed in FMyeast and correctly targeted to the inner membrane of
mitochondria (Figures 2B–2D). The possession of an TLMCF protein, complete with appropriate signals to
guide heterologous mitochondrial import [16], provides tcadditional strong evidence that the Entamoeba mito-
some is a mitochondrial homolog [3, 5]. w
unctional Characterization of the EntamoebaCF Proteinhe Entamoeba carrier was expressed in the bacteriumactococcus lactis [17]. Membrane vesicles containinghe Entamoeba carrier took up radio-labeled ATP in ex-hange for ADP (see the Supplemental Data availableith this article online). The uptake of radio-labeled
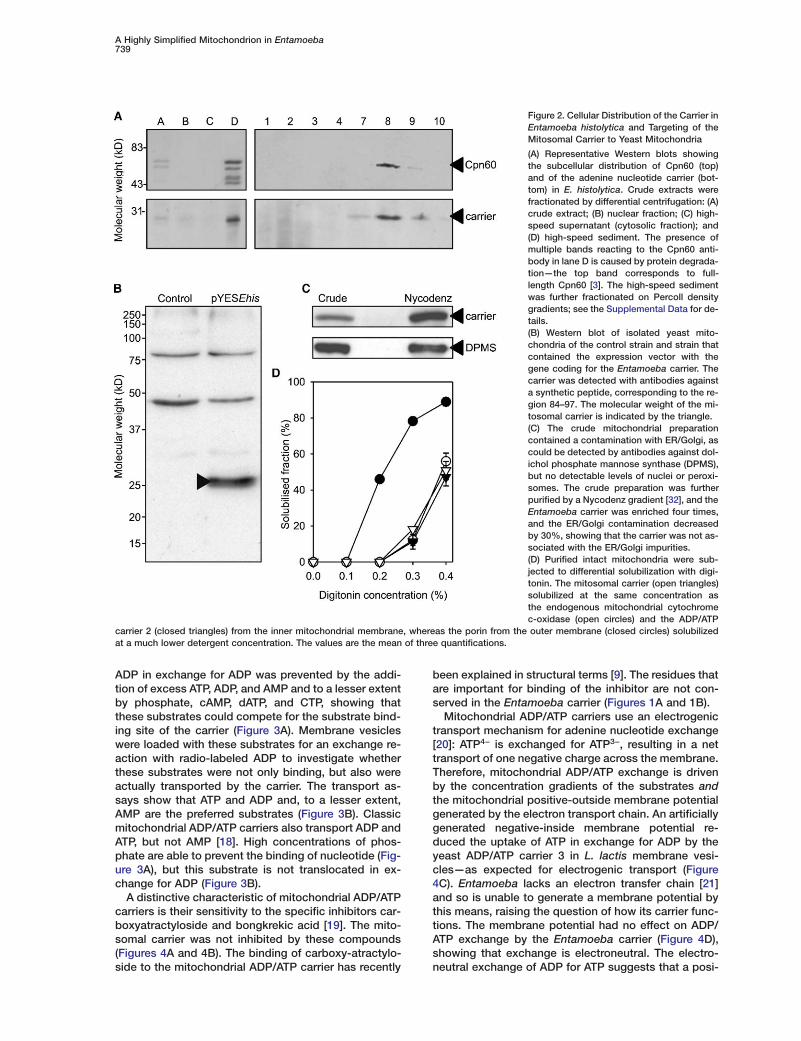
A Highly Simplified Mitochondrion in Entamoeba739
Figure 2. Cellular Distribution of the Carrier inEntamoeba histolytica and Targeting of theMitosomal Carrier to Yeast Mitochondria
(A) Representative Western blots showingthe subcellular distribution of Cpn60 (top)and of the adenine nucleotide carrier (bot-tom) in E. histolytica. Crude extracts werefractionated by differential centrifugation: (A)crude extract; (B) nuclear fraction; (C) high-speed supernatant (cytosolic fraction); and(D) high-speed sediment. The presence ofmultiple bands reacting to the Cpn60 anti-body in lane D is caused by protein degrada-tion—the top band corresponds to full-length Cpn60 [3]. The high-speed sedimentwas further fractionated on Percoll densitygradients; see the Supplemental Data for de-tails.(B) Western blot of isolated yeast mito-chondria of the control strain and strain thatcontained the expression vector with thegene coding for the Entamoeba carrier. Thecarrier was detected with antibodies againsta synthetic peptide, corresponding to the re-gion 84–97. The molecular weight of the mi-tosomal carrier is indicated by the triangle.(C) The crude mitochondrial preparationcontained a contamination with ER/Golgi, ascould be detected by antibodies against dol-ichol phosphate mannose synthase (DPMS),but no detectable levels of nuclei or peroxi-somes. The crude preparation was furtherpurified by a Nycodenz gradient [32], and theEntamoeba carrier was enriched four times,and the ER/Golgi contamination decreasedby 30%, showing that the carrier was not as-sociated with the ER/Golgi impurities.(D) Purified intact mitochondria were sub-jected to differential solubilization with digi-tonin. The mitosomal carrier (open triangles)solubilized at the same concentration asthe endogenous mitochondrial cytochromec-oxidase (open circles) and the ADP/ATP
carrier 2 (closed triangles) from the inner mitochondrial membrane, whereas the porin from the outer membrane (closed circles) solubilizedat a much lower detergent concentration. The values are the mean of three quantifications.
ADP in exchange for ADP was prevented by the addi-tion of excess ATP, ADP, and AMP and to a lesser extentby phosphate, cAMP, dATP, and CTP, showing thatthese substrates could compete for the substrate bind-ing site of the carrier (Figure 3A). Membrane vesicleswere loaded with these substrates for an exchange re-action with radio-labeled ADP to investigate whetherthese substrates were not only binding, but also wereactually transported by the carrier. The transport as-says show that ATP and ADP and, to a lesser extent,AMP are the preferred substrates (Figure 3B). Classicmitochondrial ADP/ATP carriers also transport ADP andATP, but not AMP [18]. High concentrations of phos-phate are able to prevent the binding of nucleotide (Fig-ure 3A), but this substrate is not translocated in ex-change for ADP (Figure 3B).
A distinctive characteristic of mitochondrial ADP/ATPcarriers is their sensitivity to the specific inhibitors car-boxyatractyloside and bongkrekic acid [19]. The mito-somal carrier was not inhibited by these compounds(Figures 4A and 4B). The binding of carboxy-atractylo-side to the mitochondrial ADP/ATP carrier has recently
been explained in structural terms [9]. The residues thatare important for binding of the inhibitor are not con-served in the Entamoeba carrier (Figures 1A and 1B).
Mitochondrial ADP/ATP carriers use an electrogenictransport mechanism for adenine nucleotide exchange[20]: ATP4− is exchanged for ATP3−, resulting in a nettransport of one negative charge across the membrane.Therefore, mitochondrial ADP/ATP exchange is drivenby the concentration gradients of the substrates andthe mitochondrial positive-outside membrane potentialgenerated by the electron transport chain. An artificiallygenerated negative-inside membrane potential re-duced the uptake of ATP in exchange for ADP by theyeast ADP/ATP carrier 3 in L. lactis membrane vesi-cles—as expected for electrogenic transport (Figure4C). Entamoeba lacks an electron transfer chain [21]and so is unable to generate a membrane potential bythis means, raising the question of how its carrier func-tions. The membrane potential had no effect on ADP/ATP exchange by the Entamoeba carrier (Figure 4D),showing that exchange is electroneutral. The electro-neutral exchange of ADP for ATP suggests that a posi-

Current Biology740
Figure 3. Substrate Specificity of the Mitoso-mal Carrier as Determined by Competition(A) and Active Transport (B)
(A) Initial uptake rate of radio-labeled ADP inthe presence or absence of a 3,333-foldhigher concentration of the indicated nonla-beled compounds.(B) Initial uptake rate of radio-labeled ADPinto fused membrane vesicles that were pre-loaded with different substrates. Only whenthe substrates are transported will exchangeoccur. High concentrations of phosphate areable to prevent the binding of nucleotide (A),but Pi is not translocated (B). In both typesof experiments (A and B), membrane vesi-cles were loaded with 2 mM substrate anddiluted 200-fold in a buffer containing 0.6 �M[14C]-ADP. The initial uptake rates were cal-culated from the accumulation of radio-labeled ADP after 20 s as determined byscintillation counting after removal of exter-nal radio-labeled substrate by filtration.
tive counter ion is transported to compensate for the “bcharge difference, but the type of ion has not been
identified. The peroxisomal AMP/ATP carrier [22] and aTthe mitochondrial GDP/GTP carrier [23] both use pro-
tons as counter ions for electron neutral exchange of rgnucleotides, and this may be the case for the mitoso-
mal carrier too. shm
Phylogenetic Analysis of the Entamoeba oMCF Protein aWe carried out a phylogenetic analysis of known MCFsubfamilies (Supplemental Data). The Entamoeba pro-tein formed a cluster together with uncharacterized ho- C
Tmologs from the aerobic slime mold Dictyostelium dis-coideum, a close relative of Entamoeba [24]. Classic c
dmitochondrial ADP/ATP carriers formed a separatecluster containing another homolog from Dictyostelium. i
yThe unfinished Dictyostelium genome contains 31 dif-ferent mitochondrial carriers, representing most of the b
krecognized MCF subfamilies [25, 26] from animals,plants, and protists. Recent phylogenetic analyses [27, l
628] suggest that eukaryotes can be divided into two
supergroups” comprising animals, fungi, and amoe-ae such as Entamoeba and Dictyostelium, or plants,lgae, and diverse protists including Plasmodium andrypanosoma. Gene fusion data [28] suggest that theoot of the eukaryotic tree is between these two super-roups. Because most MCF subfamilies occur on bothides of this rooted tree, the ancestral organelle mustave already contained a broad repertoire of MCFembers. Entamoeba has subsequently lost all but onef these carriers during its adaptation to a parasitic andnaerobic lifestyle.
oncluding Remarkshe Entamoeba mitosome has reduced its transportapacity mediated by MCF members to an unprece-ented degree and must therefore carry out only a lim-
ted subset of mitochondrial functions known fromeast mitochondria. Consistent with this hypothesis areiochemical data that suggest that Entamoeba lacksey mitochondrial pathways and that energy metabo-
ism is cytosolic [21, 29]. Mitochondrial chaperonin0—the only other protein currently known to localize
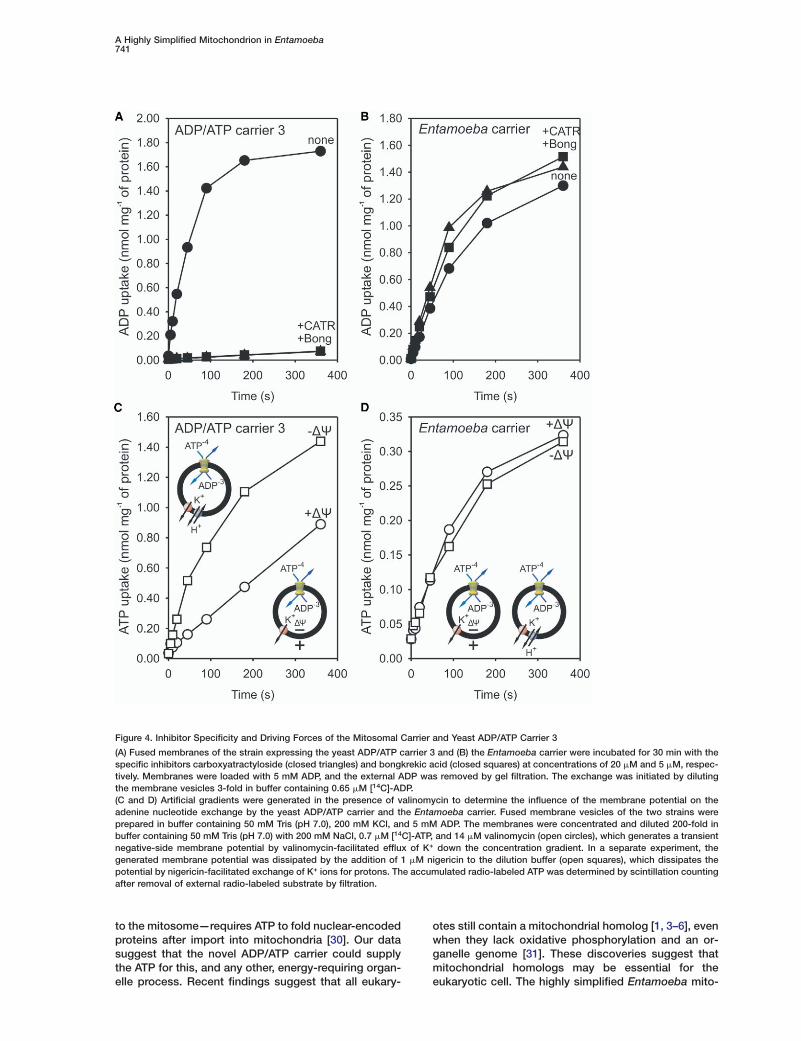
A Highly Simplified Mitochondrion in Entamoeba741
Figure 4. Inhibitor Specificity and Driving Forces of the Mitosomal Carrier and Yeast ADP/ATP Carrier 3
(A) Fused membranes of the strain expressing the yeast ADP/ATP carrier 3 and (B) the Entamoeba carrier were incubated for 30 min with thespecific inhibitors carboxyatractyloside (closed triangles) and bongkrekic acid (closed squares) at concentrations of 20 �M and 5 �M, respec-tively. Membranes were loaded with 5 mM ADP, and the external ADP was removed by gel filtration. The exchange was initiated by dilutingthe membrane vesicles 3-fold in buffer containing 0.65 �M [14C]-ADP.(C and D) Artificial gradients were generated in the presence of valinomycin to determine the influence of the membrane potential on theadenine nucleotide exchange by the yeast ADP/ATP carrier and the Entamoeba carrier. Fused membrane vesicles of the two strains wereprepared in buffer containing 50 mM Tris (pH 7.0), 200 mM KCl, and 5 mM ADP. The membranes were concentrated and diluted 200-fold inbuffer containing 50 mM Tris (pH 7.0) with 200 mM NaCl, 0.7 �M [14C]-ATP, and 14 �M valinomycin (open circles), which generates a transientnegative-side membrane potential by valinomycin-facilitated efflux of K+ down the concentration gradient. In a separate experiment, thegenerated membrane potential was dissipated by the addition of 1 �M nigericin to the dilution buffer (open squares), which dissipates thepotential by nigericin-facilitated exchange of K+ ions for protons. The accumulated radio-labeled ATP was determined by scintillation countingafter removal of external radio-labeled substrate by filtration.
to the mitosome—requires ATP to fold nuclear-encodedproteins after import into mitochondria [30]. Our datasuggest that the novel ADP/ATP carrier could supplythe ATP for this, and any other, energy-requiring organ-elle process. Recent findings suggest that all eukary-
otes still contain a mitochondrial homolog [1, 3–6], evenwhen they lack oxidative phosphorylation and an or-ganelle genome [31]. These discoveries suggest thatmitochondrial homologs may be essential for theeukaryotic cell. The highly simplified Entamoeba mito-

Current Biology742
some and the recently completed Entamoeba genomewill provide valuable tools toward identifying why thismight be so.
1
1Supplemental DataSupplemental Data including detailed Experimental Proceduresand four supplemental figures are available at http://www.current-biology.com/cgi/content/full/15/8/737/DC1/. 1
1Acknowledgments
Preliminary genomic sequence data for Entamoeba were obtainedfrom the Sanger Institute at ftp://ftp.sanger.ac.uk/pub/pathogens/
1E_histolytica and the Institute for Genomic Research at http://www.tigr.org. Sequencing of the TIGR Entamoeba histolytica Ge-nome Project was accomplished with support from the National
1Institute of Allergy and Infectious Diseases (NIAID). D.J.S. was sup-ported by a long-term fellowship of the International Human Fron-tier Science Program. K.W.C., M.H., and E.R.S.K. were supported
1by the Medical Research Council. E.R.S.K. acknowledges the sup-port of the European Molecular Biology Organisation (EMBO)Young Investigators Programme. M.v.d.G. was supported in part by
1the Leverhulme Trust. S.C. acknowledges support from the NuffieldFoundation. Research at Royal Holloway was supported by a grantfrom the Biotechnology and Biological Sciences Research Council(BBSRC) to J.T.
2Received: October 7, 2004Revised: February 21, 2005Accepted: February 21, 2005 2Published: April 26, 2005
2References
1. Tovar, J., Leon-Avila, G., Sanchez, L.B., Sutak, R., Tachezy, J.,Van Der Giezen, M., Hernandez, M., Muller, M., and Lucocq,J.M. (2003). Mitochondrial remnant organelles of Giardia func- 2tion in iron-sulphur protein maturation. Nature 426, 172–176.
2. Embley, T.M., van der Giezen, M., Horner, D.S., Dyal, P.L., Bell,S., and Foster, P.G. (2003). Hydrogenosomes, mitochondria and 2early eukaryotic evolution. IUBMB Life 55, 387–395.
3. Tovar, J., Fischer, A., and Clark, C.G. (1999). The mitosome, anovel organelle related to mitochondria in the amitochondrialparasite Entamoeba histolytica. Mol. Microbiol. 32, 1013–1021.
4. Williams, B.A., Hirt, R.P., Lucocq, J.M., and Embley, T.M. (2002).A mitochondrial remnant in the microsporidian Trachipleisto- 2phora hominis. Nature 418, 865–869.
5. Mai, Z., Ghosh, S., Frisardi, M., Rosenthal, B., Rogers, R., andSamuelson, J. (1999). Hsp60 is targeted to a cryptic mitochon-drion-derived organelle (“crypton”) in the microaerophilic pro- 2tozoan parasite Entamoeba histolytica. Mol. Cell. Biol. 19,2198–2205.
6. Hrdy, I., Hirt, R.P., Dolezal, P., Bardonova, L., Foster, P.G., 2Tachezy, J., and Embley, T.M. (2004). Trichomonas hydrogeno-somes contain the NADH dehydrogenase module of mitochon-drial complex I. Nature 432, 618–622. 2
7. Kunji, E.R. (2004). The role and structure of mitochondrial carri-ers. FEBS Lett. 564, 239–244. 2
8. Saraste, M., and Walker, J.E. (1982). Internal sequence repeatsand the path of polypeptide in mitochondrial ADP/ATP translo-case. FEBS Lett. 144, 250–254. 3
9. Pebay-Peyroula, E., Dahout-Gonzalez, C., Kahn, R., Trezeguet,V., Lauquin, G.J., and Brandolin, G. (2003). Structure of mito- 3chondrial ADP/ATP carrier in complex with carboxyatractylo-side. Nature 426, 39–44.
10. Gardner, M.J., Hall, N., Fung, E., White, O., Berriman, M., Hy-man, R.W., Carlton, J.M., Pain, A., Nelson, K.E., Bowman, S., 3et al. (2002). Genome sequence of the human malaria parasitePlasmodium falciparum. Nature 419, 498–511.
11. Abrahamsen, M.S., Templeton, T.J., Enomoto, S., Abrahante,
J.E., Zhu, G., Lancto, C.A., Deng, M., Liu, C., Widmer, G., Tzi-pori, S., et al. (2004). Complete genome sequence of the api-complexan, Cryptosporidium parvum. Science 304, 441–445.
2. Martin, J. (1997). Molecular chaperones and mitochondrial pro-tein folding. J. Bioenerg. Biomembr. 29, 35–43.
3. Horner, D.S., and Embley, T.M. (2001). Chaperonin 60 phylog-eny provides further evidence for secondary loss of mito-chondria among putative early-branching eukaryotes. Mol.Biol. Evol. 18, 1970–1975.
4. Clark, C.G., and Roger, A.J. (1995). Direct evidence for second-ary loss of mitochondria in Entamoeba histolytica. Proc. Natl.Acad. Sci. USA 92, 6518–6521.
5. Sirrenberg, C., Endres, M., Folsch, H., Stuart, R.A., Neupert,W., and Brunner, M. (1998). Carrier protein import into mito-chondria mediated by the intermembrane proteins Tim10/Mrs11 and Tim12/Mrs5. Nature 391, 912–915.
6. Pfanner, N., and Geissler, A. (2001). Versatility of the mitochon-drial protein import machinery. Nat. Rev. Mol. Cell Biol. 2,339–349.
7. Kunji, E.R., Slotboom, D.J., and Poolman, B. (2003). Lactococ-cus lactis as host for overproduction of functional membraneproteins. Biochim. Biophys. Acta 1610, 97–108.
8. Pfaff, E., and Klingenberg, M. (1968). Adenine nucleotide trans-location of mitochondria. 1. Specificity and control. Eur. J. Bio-chem. 6, 66–79.
9. Brandolin, G., Le Saux, A., Trezeguet, V., Lauquin, G.J., andVignais, P.V. (1993). Chemical, immunological, enzymatic, andgenetic approaches to studying the arrangement of the pep-tide chain of the ADP/ATP carrier in the mitochondrial mem-brane. J. Bioenerg. Biomembr. 25, 459–472.
0. Klingenberg, M. (1985). The ADP/ATP carrier in mitochondrialmembranes. In The Enzymes of Biological Membranes, A.N.Martonosi, ed. (New York: Plenum Press), pp. 511–553.
1. Reeves, R.E. (1984). Metabolism of Entamoeba histolyticaSchaudinn, 1903. Adv. Parasitol. 23, 105–142.
2. Lasorsa, F.M., Scarcia, P., Erdmann, R., Palmieri, F., Rotten-steiner, H., and Palmieri, L. (2004). The yeast peroxisomal ade-nine nucleotide transporter: Characterization of two transportmodes and involvement in delta pH formation across peroxi-somal membranes. Biochem. J. 381, 581–585.
3. Vozza, A., Blanco, E., Palmieri, L., and Palmieri, F. (2004). Iden-tification of the mitochondrial GTP/GDP Transporter inSaccharomyces cerevisiae. J. Biol. Chem. 279, 20850–20857.
4. Bapteste, E., Brinkmann, H., Lee, J.A., Moore, D.V., Sensen,C.W., Gordon, P., Durufle, L., Gaasterland, T., Lopez, P., Muller,M., et al. (2002). The analysis of 100 genes supports the group-ing of three highly divergent amoebae: Dictyostelium, Enta-moeba, and Mastigamoeba. Proc. Natl. Acad. Sci. USA 99,1414–1419.
5. Kuan, J., and Saier, M.H., Jr. (1993). The mitochondrial carrierfamily of transport proteins: Structural, functional, and evolu-tionary relationships. Crit. Rev. Biochem. Mol. Biol. 28, 209–233.
6. El Moualij, B., Duyckaerts, C., Lamotte-Brasseur, J., and Sluse,F.E. (1997). Phylogenetic classification of the mitochondrialcarrier family of Saccharomyces cerevisiae. Yeast 13, 573–581.
7. Baldauf, S., Roger, A., Wenk-Siefert, I., and Doolittle, W.F.(2000). A kingdom-level phylogeny of eukaryotes based oncombined protein data. Science 290, 972–977.
8. Stechmann, A., and Cavalier-Smith, T. (2003). The root of theeukaryote tree pinpointed. Curr. Biol. 13, R665–R666.
9. Muller, M. (2003). Energy metabolism. Part I. Anaerobic proto-zoa. In Molecular Medical Parasitology, J. Marr, T. Nielson, andR. Komuniecki, eds. (London: Academic Press), pp. 125–139.
0. Gething, M.J., and Sambrook, J. (1992). Protein folding in thecell. Nature 355, 33–45.
1. Leon-Avila, G., and Tovar, J. (2004). Mitosomes of Entamoebahistolytica are abundant mitochondrion-related remnant organ-elles that lack a detectable organellar genome. Microbiol. 150,1245–1250.
2. Diekert, K., de Kroon, A.I., Kispal, G., and Lill, R. (2001). Isola-tion and subfractionation of mitochondria from the yeastSaccharomyces cerevisiae. Methods Cell. Biol. 65, 37–51.

S1Supplemental Data
A Novel ADP/ATP Transporterin the Mitosome of the MicroaerophilicHuman Parasite Entamoeba histolytica
Ka Wai Chan, Dirk-Jan Slotboom, Sian Cox,T. Martin Embley, Olivier Fabre,Mark van der Giezen, Marilyn Harding,David S. Horner, Edmund R.S. Kunji,Gloria Leon-Avila, and Jorge Tovar
Supplemental Experimental Procedures freeze-thawing. Crude extracts were fractionated by differentialcentrifugation as described previously [S6]. The postnuclear super-natant was centrifuged at 100,000 � g for 30 min to generate aCloning of the Mitosomal Carrier
The mitosomal adenine nucleotide carrier was identified with BlastP high-speed supernatant (the cytosolic fraction) and a high-speedsediment. The latter was resuspended in SMDI (0.25 M sucrose, 10on data generated by the Entamoeba histolytica Genome Project
and amplified by PCR from E. histolytica genomic DNA. Rapid ampli- mM MOPS, and 10 mM DTT at pH 7.2) containing Complete ProteaseInhibitors (Roche) and loaded on a 45% Percoll density gradient infication of cDNA ends (RACE) was used to amplify the full open
reading frame (ORF) from a cDNA library. The gene was modified SMDI [S7]. Fractions were collected after centrifugation at 68,000 � gfor 30 min at 4�C.by PCR to introduce a BspHI site coinciding with the start codon
and an NheI site immediately after the stop codon. The product wasligated into vector pNZ8048 [S1]. The resulting plasmid pNZEhis Enzyme Assays for Acid Phosphatase
Acid phosphatase activity was determined with p-nitrophenyl phos-was transformed into Lactococcus lactis NZ9000.phate (p-NPP) as substrate [S8]. Fractions (0.1 ml) were incubatedwith 0.9 ml of substrate solution (Sigma) for 30 min at 37�C. Reac-Expression and Localization Studies of the Entamoeba
Carrier in Yeast tions were terminated by addition of 0.2 ml 2N NaOH, and the con-centration of p-nitophenol was determined with an extinction coeffi-The E. histolytica gene was amplified by PCR with primers to intro-
duce an in-frame NcoI site, incorporating the start codon, as well cient of 18.5 mM cm�1. One unit of activity is defined as the amountof enzyme hydrolizing 1 nmol of substrate per min at 37�C.as an NheI site after the 3� stop codon. The product was cloned
into pYES-Pgal-aac2 [S2], resulting in the expression vector pYES-Ehis, which was transformed into S. cerevisiae INVSc1. Yeast mito- Isolation of Membrane Vesicles of Lactococcus lactis
Lactococcus lactis strain NZ9000/pNZEhis was grown at 30�C inchondria were prepared [S3], and soluble and peripheral proteinswere removed by rupture of the outer mitochondrial membrane to M17 broth supplemented with 1% glucose and 5 �g/ml chloram-
phenicol. Expression was induced at A600 of 0.6 by the addition ofproduce mitoplasts and then by sodium carbonate extraction [S4].For differential solubilization, mitochondria were extracted with dig- a dilution of 1:1000 of Nisin A stock [S1]. Cultures were incubated
for 3 hr at 30�C and left at 20�C for an additional 16 hr. Cells wereitonin [S5], and proteins in the soluble fraction were precipitatedwith 10% trichloroacetic acid. collected by centrifugation for 10 min at 6,000 � g and 4�C and
were washed once in ice-cold Tris-buffered saline (TBS; 20 mM Tris[pH7.4] and 150 mM NaCl). The final pellet was resuspended in 50Preparation of E. histolytica Cellular Fractions
E. histolytica HM-1:IMSS clone 9 was grown in YI-S medium with ml TBS, and the cells were lysed with a Constant Systems 2.2 kVdisrupter at 30,000 psi and 0.5�C. Lysed cells were left for 10 min on15% adult bovine serum. E. histolytica trophozoites were lysed by
Figure S1. Protein Content of Entamoeba Cell Fractions
(A) Representative Coomassie-blue-stained gel of crude cell extracts fractionated by differential centrifugation: (A) crude extract; (B) nuclearfraction; (C) high-speed supernatant; and (D) high-speed sediment. The gel corresponds to the Western blot shown in Figure 2A.(B) The high-speed sediment was further fractionated on a 45% Percoll density gradient as described in the Supplemental ExperimentalProcedures. (B) shows a corresponding Coomassie-stained gel showing protein distribution in all fractions; these fractions correspond tolanes 1–10 in Figure 2A.

S2
Figure S2. Distribution of the LysosomalMarker Enzyme Acid Phosphatase Activity inPercoll Gradient Fractions
Acid phosphatase (AP) activity was assayedin fractions 1–10 of the Percoll gradientshown in Figure 2A, as described in the Sup-plemental Experimental Procedures. The ex-tent of mitosome enrichment in organellarpreparations is illustrated by comparing thedistribution of the mitosomal protein Cpn60(Figure 2), the pattern of Coomassie staining(Figure S1B), and the distribution of contami-nating lysosomal vesicles in the Percoll gradi-ent, as indicated by the marker enzyme acidphosphatase.
ice, and whole cells were subsequently removed by centrifugation at in buffer containing radio-labeled substrate, competitors, inhibitors,or ionophores as indicated. In the second method, the substrate-11,000 � g for 10 min at 4�C. The membrane vesicles were collected
by ultracentrifugation at 200,000 � g for 30 min at 4�C after one loaded fused membranes were concentrated in 50 �l buffer anddirectly diluted 200-fold in buffer containing radio-labeled substrate,wash step with TBS.competitors, inhibitors, or ionophores, depending on the experi-ment. All uptake experiments were performed at 25�C with constantProduction and Purification of Antibodiesstirring. Uptake was stopped by addition of 4 ml of ice-cold TBSAntibodies against a synthetic peptide, corresponding to the regionand then by rapid filtration over cellulose nitrate filters (0.45 �m84–97 of the mitosomal carrier, were produced in hen (Agrisera).pore size). The filters were washed once with 2 ml ice-cold TBS andThe antibodies were affinity purified with mitosomal carrier protein.transferred to a vial for scintillation counting.
SDS-Polyacrylamide Gel Electrophoresisand Western Blotting Phylogenetic AnalysesSamples were loaded onto 15% SDS-PAGE gel and separated at Alignment of conceptually translated protein sequences of all avail-30 mA. For Western analysis, proteins were transferred onto a poly- able MCF sequences was performed with ClustalW [S9], with manualvinylidene fluoride (PVDF) membrane at 120 mA for 1 hr. The mem- adjustment of the resulting alignment. Phylogenetic analysis of thisbrane was blocked with 5% (w/v) skimmed milk in TBS buffer con- data set (199 sequences, 232 positions) was performed withtaining 0.1% (v/v) Tween overnight at room temperature with gentle MrBayes [S10], with 200,001 generations under the Whelan andagitation. The purified primary antibody and secondary antibody Goldman (WAG) amino-acid substitution model, with one invariable(anti-chicken HRP; Sigma) were used at titers of 1:250 and 1:20,000, and eight invariable substitution-rate categories. 501 trees wererespectively. Antibody-stained protein bands were visualized with discarded as “burn-in.”ECL (Amersham). On the basis of this analysis, a second data set of 39 sequences
most closely related to the Entamoeba histolytica carrier was ana-Preparation of Liposomes and Fused Membranes lyzed further. Bootstrap replicate data sets (100) were each analyzedEscherichia coli total lipid extract and egg yolk phosphatidylcholine with MrBayes as before. Branch lengths for the bootstrap consensus(Avanti Polar Lipids) were mixed in a 3:1 w/w ratio in buffer to a tree (Figure S4) were estimated with Tree-Puzzle version 5. Con-final lipid concentration of 20 mg/ml. Membrane vesicles were mixed straint trees, consistent with different hypotheses of relationship forwith liposomes in a 1:5 (w/w) protein to lipid ratio in 800 �l of the Entamoeba carrier, were constructed with MrBayes and evalu-buffer in the presence of substrates, ionophores, and inhibitors. ated against the best unconstrained tree we found with the approxi-Membranes and liposomes were fused by freezing in liquid nitrogen mately unbiased (AU) test [S11]. Using this test, we were able toand thawing at room temperature seven times. reject at p � 0.05 the hypothesis that the Entamoeba MCF was a
member of the classical ADP/ATP carrier clade.Transport AssaysThe fused membranes were extruded nine times through 400 nmpolycarbonate filters and collected by centrifugation for 30 min at Supplemental References390,000 � g and 4�C. The pellet was resuspended in the appropriatebuffer, containing the substrate, inhibitors, or ionophores. Two dif- S1. Kunji, E.R., Slotboom, D.J., and Poolman, B. (2003). Lactococ-
cus lactis as host for overproduction of functional membraneferent transport methods were applied. In one method, the externalsubstrate was removed by passing the fused membrane suspension proteins. Biochim. Biophys. Acta 1610, 97–108.
S2. van der Giezen, M., Slotboom, D.J., Horner, D.S., Dyal, P.L.,over a 3.5 ml bed volume Sephadex G-75 gel filtration column equili-brated with buffer without substrate. The fused membranes were Harding, M., Xue, G.P., Embley, T.M., and Kunji, E.R. (2002).
Conserved properties of hydrogenosomal and mitochondrialeluted in 1 ml buffer, and transport was initiated by 4-fold dilution

S3
S6. Tovar, J., Fischer, A., and Clark, C.G. (1999). The mitosome,a novel organelle related to mitochondria in the amitochondrialparasite Entamoeba histolytica. Mol. Microbiol. 32, 1013–1021.
S7. Bradley, P.J., Lahti, C.J., Plumper, E., and Johnson, P.L. (1997).Targeting and translocation of proteins into the hydrogeno-some of the protist Trichomonas: Similarities with mitochon-drial protein import. EMBO J. 16, 3484–3493.
S8. Aguirre-Garcia, M.M., Cerbon, J., and Talamas-Rohana, P.(2000). Purification and properties of an acid phosphatasefrom Entamoeba histolytica HM-1:IMSS. Int. J. Parasitol. 30,585–591.
S9. Thompson, J.D., Higgins, D.G., and Gibson, T.J. (1994). ClustalW: Improving the sensitivity of progressive multiple sequencealignment through sequence weighting position specific gappenalties and weight matrix choice. Nucleic Acids Res. 22,4673–4680.
S10. Huelsenbeck, J.P., and Ronquist, F. (2001). MRBAYES: Bayes-ian inference of phylogenetic trees. Bioinformatics 17,754–755.
S11. Shimodaira, H. (2002). An approximately unbiased test of phy-logenetic tree selection. Syst. Biol. 51, 492–508.
Figure S3. Expression and Functional Analysis of the MitosomalCarrier in Lactococcal Membranes
(A) Coomassie-stained SDS-polyacrylamide gel and Western-blotanalysis of membrane vesicles of the L. lactis control (lane 1) andexpression strain NZ9000/pNZEhis (lane 2), with purified antibodiesraised against peptide 84–97 of the mitosomal carrier. Approxi-mately 15 �g total membrane protein was loaded per lane. Thetriangle indicates the band of the expressed mitosomal carrier.(B) [14C]-ATP uptake by membrane vesicles of the expressionNZ9000/pNZEhis (closed triangles) and control strain (closed circles)preloaded with 5 mM ADP. Membranes of a strain expressing theyeast ADP/ATP carrier 3 were used as a positive control (opencircles). After removal of external ADP by gel filtration, the exchangewas initiated by diluting the membrane vesicles 3-fold in buffercontaining 1 �M valinomycin, 1 �M nigericin, and 0.7 �M [14C]-ATP.The accumulated radiolabeled ATP was determined by scintillationcounting after removal of external radio-labeled substrate by fil-tration.
ADP/ATP carriers: A common origin for both organelles. EMBOJ. 21, 572–579.
S3. Stuart, J.A., Harper, J.A., Brindle, K.M., Jekabsons, M.B., andBrand, M.D. (2001). A mitochondrial uncoupling artifact canbe caused by expression of uncoupling protein 1 in yeast.Biochem. J. 356, 779–789.
S4. Diekert, K., de Kroon, A.I., Kispal, G., and Lill, R. (2001). Isola-tion and subfractionation of mitochondria from the yeast Sac-charomyces cerevisiae. Methods Cell Biol. 65, 37–51.
S5. Palmieri, L., Vozza, A., Agrimi, G., De Marco, V., Runswick,M.J., Palmieri, F., and Walker, J.E. (1999). Identification of theyeast mitochondrial transporter for oxaloacetate and sulfate.J. Biol. Chem. 274, 22184–22190.

S4
Figure S4. Phylogenetic Analysis of the Mitosomal ADP/ATP Carrier
The tree shown is a Bayesian consensus tree from 100 bootstrap replicate data sets. Bootstrap values over 50% are given at relevant nodes.The scale bar represents 0.1 substitutions per site.




















