
Distinct biological activities of C3 and ADP-ribosyltransferase-deficient C3-E174Q
15
Distinct biological activities of C3 and ADP-ribosyltransferase-deficient C3-E174Q Astrid Rohrbeck, Tanja Kolbe, Sandra Hagemann, Harald Genth and Ingo Just Institute of Toxicology, Medizinische Hochschule Hannover, Hanover, Germany Keywords ADP-ribosyltransferase; apoptosis; C3 exoenzyme; proliferation; RhoA Correspondence A. Rohrbeck, Department of Toxicology, Medizinische Hochschule Hannover, Carl-Neuberg-Straße 1, D-30625 Hanover, Germany Fax: +49 511 5322879 Tel: +49 511 5322807 E-mail: [email protected] (Received 25 October 2011, revised 15 May 2012, accepted 16 May 2012) doi:10.1111/j.1742-4658.2012.08645.x Low-molecular-weight GTP-binding proteins of the Rho family control the organization of the actin cytoskeleton in eukaryotic cells. Dramatic reorga- nization of the actin cytoskeleton is caused by the C3 exoenzyme derived from Clostridium botulinum (C3), based on ADP-ribosylation of RhoA ⁄ B ⁄ C. In addition, wild-type as well as ADP-ribosyltransferase-defi- cient C3-E174Q induce axonal outgrowth of primary murine hippocampal neurons and prevent growth cone collapse, indicating a non-enzymatic mode of action. In this study, we compared the effects of C3-E174Q and wild-type C3 in the murine hippocampal cell line HT22. Treatment of HT22 cells with C3 resulted in Rho ADP-ribosylation and cell rounding. The ADP-ribosyltransferase-deficient mutant C3-E174Q did not induce either Rho ADP-ribosylation or morphological changes. C3 as well as C3-E174Q treatment resulted in growth arrest, reduced expression of cyclin D levels, and increased expression of RhoB, a negative regulator of cell-cycle progression. Serum starvation induced apoptosis in HT22 cells, as determined on the basis of increased expression of caspase-9 and Bax. C3 but not C3-E174Q protected serum-starved HT22 cells from apoptosis. This is the first study separating ADP-ribosyltransferase-dependent from ADP-ribosyltransferase-independent effects of C3. While morphological changes and anti-apoptotic activity strictly depend on ADP-ribosyltransfer- ase activity, the anti-proliferative effects are independent of ADP-ribosyl- transferase activity. Structured digital abstract l Rhotekin physically interacts with RHOA by pull down ( View interaction) Introduction Low-molecular-weight GTP-binding proteins of the Rho family control the organization of the actin cyto- skeleton in eukaryotic cells and are involved in the reg- ulation of gene expression, cell proliferation, apoptosis and axonal growth [1,2]. The Rho family members RhoA, B and C exhibit significant amino acid sequence identity (approximately 85%) and are thought to inter- act with the same effectors. RhoB appears to possess unique functions compared to RhoA and RhoC. RhoB is exclusively localized to membranes, both plasma membrane and endosomes, even when inactive [3]. RhoA is cytosolic within the GDI complex, and only moves to the plasma membrane upon activation and dissociation of RhoGDI. Interestingly, RhoGDI-1 does not bind to RhoB, consistent with the finding that RhoB is always found in association with membranes [4]. RhoB has a specialized function in intracellular trafficking of cytokine receptors such as epidermal growth factor receptor [5]. Additionally, RhoB protein is relatively short-lived in cells, and its expression can Abbreviations ART, ADP-ribosyltransferase; C3, C3 exoenzyme derived from Clostridium botulinum; C3-E174Q, ADP-ribosyltransferase-deficient C3. FEBS Journal 279 (2012) 2657–2671 ª 2012 The Authors Journal compilation ª 2012 FEBS 2657
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Distinct biological activities of C3 and ADP-ribosyltransferase-deficient C3-E174Q

Distinct biological activities of C3 andADP-ribosyltransferase-deficient C3-E174QAstrid Rohrbeck, Tanja Kolbe, Sandra Hagemann, Harald Genth and Ingo Just
Institute of Toxicology, Medizinische Hochschule Hannover, Hanover, Germany
Keywords
ADP-ribosyltransferase; apoptosis;
C3 exoenzyme; proliferation; RhoA
Correspondence
A. Rohrbeck, Department of Toxicology,
Medizinische Hochschule Hannover,
Carl-Neuberg-Straße 1, D-30625 Hanover,
Germany
Fax: +49 511 5322879
Tel: +49 511 5322807
E-mail: [email protected]
(Received 25 October 2011, revised 15 May
2012, accepted 16 May 2012)
doi:10.1111/j.1742-4658.2012.08645.x
Low-molecular-weight GTP-binding proteins of the Rho family control the
organization of the actin cytoskeleton in eukaryotic cells. Dramatic reorga-
nization of the actin cytoskeleton is caused by the C3 exoenzyme derived
from Clostridium botulinum (C3), based on ADP-ribosylation of
RhoA ⁄B ⁄C. In addition, wild-type as well as ADP-ribosyltransferase-defi-
cient C3-E174Q induce axonal outgrowth of primary murine hippocampal
neurons and prevent growth cone collapse, indicating a non-enzymatic
mode of action. In this study, we compared the effects of C3-E174Q and
wild-type C3 in the murine hippocampal cell line HT22. Treatment of
HT22 cells with C3 resulted in Rho ADP-ribosylation and cell rounding.
The ADP-ribosyltransferase-deficient mutant C3-E174Q did not induce
either Rho ADP-ribosylation or morphological changes. C3 as well as
C3-E174Q treatment resulted in growth arrest, reduced expression of
cyclin D levels, and increased expression of RhoB, a negative regulator of
cell-cycle progression. Serum starvation induced apoptosis in HT22 cells, as
determined on the basis of increased expression of caspase-9 and Bax. C3
but not C3-E174Q protected serum-starved HT22 cells from apoptosis.
This is the first study separating ADP-ribosyltransferase-dependent from
ADP-ribosyltransferase-independent effects of C3. While morphological
changes and anti-apoptotic activity strictly depend on ADP-ribosyltransfer-
ase activity, the anti-proliferative effects are independent of ADP-ribosyl-
transferase activity.
Structured digital abstractl Rhotekin physically interacts with RHOA by pull down (View interaction)
Introduction
Low-molecular-weight GTP-binding proteins of the
Rho family control the organization of the actin cyto-
skeleton in eukaryotic cells and are involved in the reg-
ulation of gene expression, cell proliferation, apoptosis
and axonal growth [1,2]. The Rho family members
RhoA, B and C exhibit significant amino acid sequence
identity (approximately 85%) and are thought to inter-
act with the same effectors. RhoB appears to possess
unique functions compared to RhoA and RhoC. RhoB
is exclusively localized to membranes, both plasma
membrane and endosomes, even when inactive [3].
RhoA is cytosolic within the GDI complex, and only
moves to the plasma membrane upon activation and
dissociation of RhoGDI. Interestingly, RhoGDI-1 does
not bind to RhoB, consistent with the finding that
RhoB is always found in association with membranes
[4]. RhoB has a specialized function in intracellular
trafficking of cytokine receptors such as epidermal
growth factor receptor [5]. Additionally, RhoB protein
is relatively short-lived in cells, and its expression can
Abbreviations
ART, ADP-ribosyltransferase; C3, C3 exoenzyme derived from Clostridium botulinum; C3-E174Q, ADP-ribosyltransferase-deficient C3.
FEBS Journal 279 (2012) 2657–2671 ª 2012 The Authors Journal compilation ª 2012 FEBS 2657

be elevated by a number of stimuli, including epidermal
growth factor, transforming growth factor b and geno-
toxic stress [6,7]. Moreover, silencing of RhoA induced
strong up-regulation of both total and active RhoB
protein levels [8].
RhoA, RhoB and RhoC are substrates of bacterial
C3-like exoenzymes, which inactivate RhoA, B and C
by ADP-ribosylation at the amino acid asparagine at
position 41 [9]. In cultured cells, Rho ADP-ribosylation
causes dramatic changes in the actin cytoskeleton of
most cell types, resulting in rounding of cultured
fibroblasts or epithelial cells [10,11]. However, C3 from
Clostridium botulinum (C3bot) possesses an additional
axonotrophic activity independently of its inherent
ADP-ribosyltransferase (ART) activity, as C3 induces
axonal and dendritic growth, and, in addition, branch-
ing and synapse formation in cultured primary murine
hippocampal neurons [12]. This axonotrophic activity
is specific for C3bot, as C3 isoforms from other
microbes do not exhibit axon and dendrite growth-
promoting activity in hippocampal neurons [13]. More-
over, the axonotrophic activity of the ART-deficient
mutant C3bot-E174Q is as effective as the enzyme-
active C3bot. It is known that C3 stimulates growth
cone formation in neuronal cells N1E-115 by inhibition
of growth cone collapse through RhoA inactivation
[14]. On the other hand, C3 induces axonal outgrowth
from DRG (dorsal root ganglion) neurons, but forma-
tion of lamellipodia and filopodia in the growth cones
of DRG is not detectable under these conditions [15].
However, in addition to axon growth-promoting
activity, C3 causes inhibition of cell growth in cultured
rat pheochromocytoma PC-12 cells and formation of
neurites [16]. Growing evidence suggests that C3 influ-
ences cell proliferation by inactivation of RhoA. RhoA
is known to be instrumental in the kinetics of
cyclin D1 expression [17]. Moreover, others have
observed inhibition of cyclin D1 expression upon C3
treatment [18]. RhoA also suppresses p21 levels in
multiple normal and transformed cell lines [19]. Degra-
dation of G1 cyclin-dependent kinase inhibitor
p27(KIP1) is inhibited by C3, further supporting the
important role of RhoA in cell proliferation [20]. Inter-
estingly, RhoB negatively regulates cell proliferation
[21]. A negative role of RhoB in growth control would
contrast with the positive effects of RhoA and RhoC
[22].
Additionally, C3 also appears to play a role in sur-
vival signalling. It was shown that C3-catalyzed inacti-
vation of RhoA induces apoptosis in haematopoietic
cells [23] and causes adhesion-independent apoptosis
[24]. Recently, it has been reported that C3 results in
activation of caspase-3 and thus apoptosis of cardio-
myocytes [25]. By contrast, inactivation of RhoA by
C3 protects neurons from cell death [26]. In another
study, it was observed that, after contusion injury, cell-
permeable C3 decreased neuronal and glial apoptosis,
as detected by TUNEL [27].
In this study, the biological effects of C3 and C3-
E174Q were analyzed in the mouse hippocampal HT22
cells, a cell line that exhibits sensitivity to C3. C3 and
C3-E174Q both induced inhibition of cell proliferation
and cyclin D1 down-regulation as well as RhoB expres-
sion. These effects thus appear to be independent of the
ART activity of C3. In contrast, actin re-organization
and anti-apoptotic activity were specifically observed in
C3-treated cells, thus appeared to depend on the ART
activity. This report strongly indicates that C3 pos-
sesses ART-independent activities.
Results
ART dependency of the morphological changes
induced by C3
HT22 cells exhibited spindle-shaped morphology
(Fig. 1A). Upon C3 treatment for 72 h, a population
of HT22 cells became round and exhibited pronounced
Fig. 1. Morphology of HT22 cells after treatment with C3. HT22 cells were treated for 72 h with 500 nM of C3 or C3-E174Q. Untreated cells
served as a control. Phase-contrast microscopic images are shown. Scale bar = 50 lm.
Effect of C3 exoenzyme on hippocampal cells A. Rohrbeck et al.
2658 FEBS Journal 279 (2012) 2657–2671 ª 2012 The Authors Journal compilation ª 2012 FEBS

retraction fibers (Fig. 1). In contrast, treatment with
C3-E174Q did not cause any morphological changes
(Fig. 1A), suggesting that the morphological changes
are dependent on the ART activity of C3. C3bot
ADP-ribosylated RhoA from HT22 cells in a time-
and concentration-dependent manner, as evidenced by
the apparent shift of ADP-ribosylated RhoA to higher
molecular mass in SDS ⁄PAGE (Fig. 2A) and sequen-
tial [32P]ADP-ribosylation of non-ADP-ribosylated
Rho (Fig. 2B). In contrast, C3-E174Q did not ADP-ri-
bosylate cellular RhoA (Fig. 2A–C). Furthermore, C3-
E174Q did not possess any ART activity in a cell-free
system (Fig. S1). RhoA ADP-ribosylation by C3
resulted in a reduced level of RhoA-GTP, but treat-
ment with C3-E174Q did not affect the level of RhoA-
GTP (Fig. 3). These observations were consistent with
previous reports showing that C3-E174Q lacked ART
activity [13] and that ADP-ribosylation resulted in
RhoA inactivation [28,29].
Binding measurements for purified RhoA using
microscale thermophoresis
Next, we investigated binding of fluorescence-labelled
RhoA to C3 exoenzyme using microscale thermopho-
resis. Binding of C3 to RhoA was readily observed as
Fig. 2. Sensitivity of HT22 cells towards C3
exoenzyme. (A) Exposure of HT22 cells to
increasing concentrations of C3 causes a
molecular mass shift of RhoA in SDS ⁄ PAGE
and a decrease in RhoA level in a concentra-
tion- and time-dependent manner. Cells trea-
ted with the C3-E174Q did not show an
RhoA shift. (B) HT22 cells were exposed to
increasing concentrations of C3 for 8 or
24 h at 37 �C. Cells were lysed and sub-
jected to sequential [32P]ADP-ribosylation.
Phosphorimages from representative experi-
ments are shown. (C) Densitometric mea-
surements for a representative experiment.
A. Rohrbeck et al. Effect of C3 exoenzyme on hippocampal cells
FEBS Journal 279 (2012) 2657–2671 ª 2012 The Authors Journal compilation ª 2012 FEBS 2659

a change in the thermophoretic property of the
fluorescently labelled RhoA upon complex formation.
The C3–RhoA complex shows a stronger increase of
normalized fluorescence than the unbound protein.
The signal shown in Fig. 4A is a binding curve, and
starts at an Fnorm of approximately 830 units. When
the concentration of C3 is increased, the microscale
thermophoresis signal increases to approximately
920 units. Thus, a significant amount of the RhoA is
in complex with C3. From the data, we inferred a
dissociation coefficient of 32.18 lM (Fig. 4A). We next
assessed the interaction of RhoA with C3-E174Q.
Figure 4B shows the resulting sigmoidal binding curve
for the C3-E174Q–RhoA interaction, with significant
differences between low and high C3-E174Q concen-
trations and a calculated Kd of 7.23 lM. These findings
indicate a four- to fivefold higher affinity of RhoA for
C3-E174Q compared to C3.
Distinct kinetics of RhoB expression induced by
C3 and C3-E174Q
RhoA ADP-ribosylation by C3 has been reported to
result in RhoB expression [30]. Consistently, RhoA
ADP-ribosylation in C3-treated HT22 cells was
accompanied by pronounced RhoB expression
(Fig. 5A). Interestingly, RhoB expression was also
observed in HT22 cells upon prolonged treatment with
C3-E174Q for 2–6 days (Fig. 5B). Under these condi-
tions, neither RhoA nor RhoB was ADP-ribosylated,
as evidenced by the lack of shift of RhoA and RhoB
to apparent higher molecular masses in SDS ⁄PAGE
(Fig. 5B). However, RhoB expression in C3-E174Q-
treated cells correlated with a decreased cellular level
of RhoA. C3-E174Q thus appeared to induce RhoB
expression independently of RhoA ADP-ribosylation.
Reduced cell proliferation upon treatment with
both C3 and C3-E174Q
RhoB has been implicated in the regulation of cell
proliferation, as ectopic expression of constitutively
active RhoB inhibits proliferation [31]. Next, the effects
of C3 and C3-E174Q on HT22 proliferation was analy-
sed over a 7-day period (Fig. 6). Untreated cells
showed a reduced growth rate, probably due to density
inhibition. The cells reach confluence and form a
monolayer at day 5 (Fig. 6). Upon treatment with
either C3 or C3-E174Q, exponential growth was
observed up to 2 days, and then proliferation ceased.
Fig. 3. GTP-RhoA levels were determined in lysates of HT22 cells using pull-down assay. HT22 cells were treated with either C3 or
C3-E174Q for various times. (A) Cell lysates were subjected to Rho binding by pull-down assay followed by western blot analysis using anti-
bodies against RhoA or b-actin. Western blot analyses from representative experiments are presented (n = 3). GTPyS = positive control.
(B,C) Cellular levels of GTP-RhoA proteins at 24 h (B) and 72 h (C) were quantified using KODAK 1D image analysis software. All signal intensi-
ties were normalized to the intensity of the corresponding b-actin signal.
Effect of C3 exoenzyme on hippocampal cells A. Rohrbeck et al.
2660 FEBS Journal 279 (2012) 2657–2671 ª 2012 The Authors Journal compilation ª 2012 FEBS

In C3-treated cells, the number of viable cell decreased
by approximately 30%, probably due to C3-mediated
cytotoxicity. The number of C3-E174Q-treated cells
remained almost constant from 3 to 7 days, suggesting
that C3-E174Q inhibited cell proliferation (Fig. 6). To
determine whether the anti-proliferative activity of C3
and C3-E174Q was due to blocked G1–S transition, cel-
lular levels of cyclin D1 mRNA and cyclin D1 protein
were analysed by quantitative RT-PCR and western
blot analysis, respectively. C3 and C3-E174Q compara-
bly down-regulated both cyclin D1 mRNA (Fig. 7A)
and cyclin D1 protein (Fig. 7B) levels in time-series
experiments. As shown in Fig. 6B,C, untreated cells
show a time-dependent decrease in cyclin D1, probably
due to contact inhibition [32]. These observations
strongly suggest that C3-induced inhibition of cell pro-
liferation is independent of ART activity.
ART dependency of the anti-apoptotic
activity of C3
Depending on the cell line and the experimental condi-
tions, C3 has been reported to exhibit either pro-apop-
totic [33] or anti-apoptotic activity [34]. To determine
whether C3 promoted or inhibited apoptosis of HT22
cells, starvation-induced apoptosis of HT22 cells was
analysed in the presence and absence of C3. C3 treat-
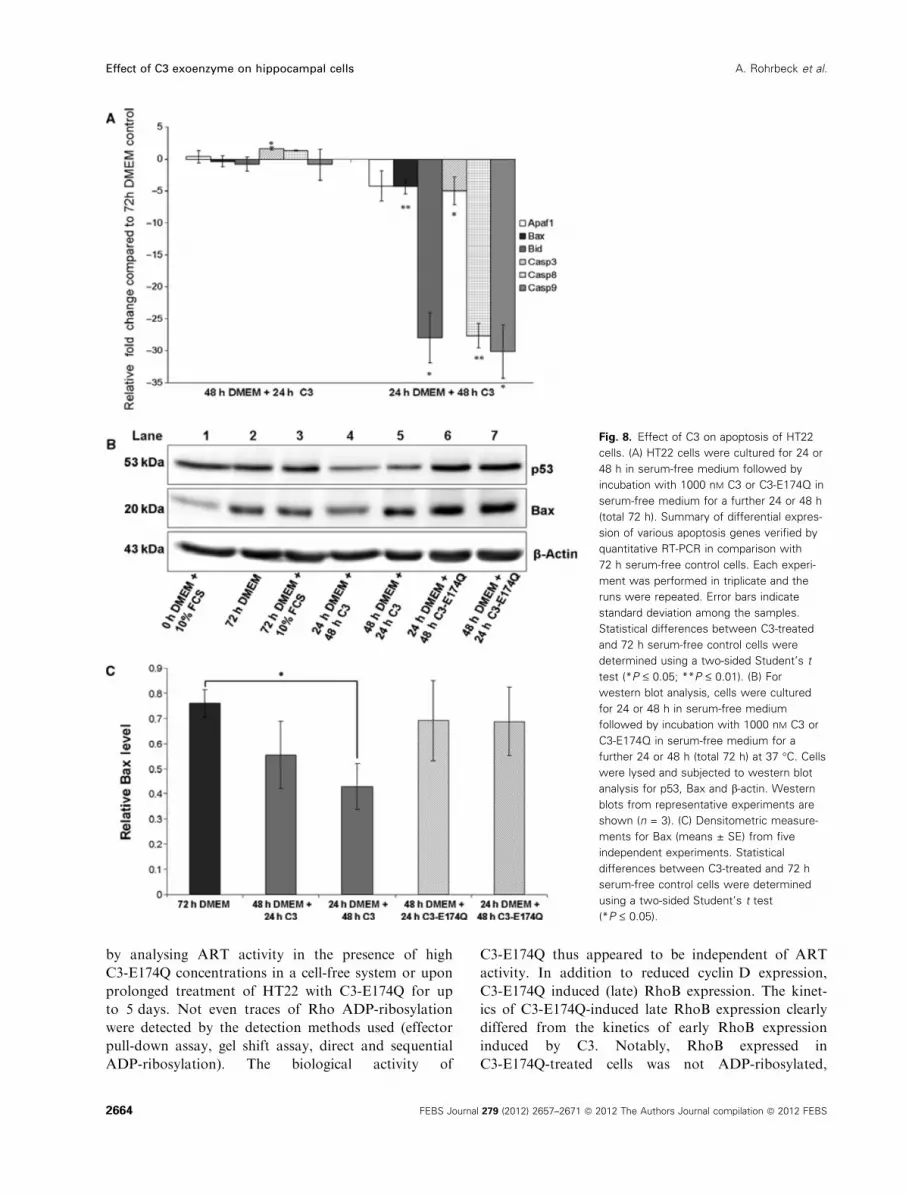
ment for 48 h resulted in a significant reduction of bax
mRNA and caspase-3 mRNA (Fig. 8A). To elucidate
the role of additional apoptotic factors, expression of
caspase-8 and caspase-9 mRNA was assessed. Both
caspases were strongly down-regulated by C3
(Fig. 8A). However, C3-E174Q did not influence
expression of the genes studied (data not shown).
As p53 is an important player in various signalling
pathways including apoptosis, the protein level of p53
was examined. Wild-type C3 induced a decrease in
p53, as shown by western blot analysis. In contrast,
C3-E174Q did not alter p53 protein level (Fig. 8B).
Furthermore, starvation-induced apoptosis of HT22
cells resulted in expression of Bax (Fig. 8B, lanes 1
and 2). To test for changes in the expression of Bax,
HT22 cells were cultured for 24 h in serum-free med-
ium to induced apoptosis, and then incubated with C3
or C3-E174Q in serum-free medium for further 24 or
48 h (Fig. 8B, lanes 4–7). We observed a reduced pro-
tein level of Bax 48 h after apoptosis induction if C3
was added after 24 h starvation-induced apoptosis
(Fig. 8, Lane 4). C3-E174Q did not reduced Bax 48 h
after apoptosis induction in comparison with 72 h
Fig. 4. Microscale thermophoresis binding
analysis. The concentration of the fluores-
cently labelled RhoA is kept constant and
the C3 exoenzyme is titrated. (A) Binding
curve for labelled RhoA–C3. (B) Binding
curve for labelled RhoA–C3-E174Q. The
binding curve is sigmoidal and reaches a pla-
teau. Binding curves from representative
experiments are shown (n = 2).
A. Rohrbeck et al. Effect of C3 exoenzyme on hippocampal cells
FEBS Journal 279 (2012) 2657–2671 ª 2012 The Authors Journal compilation ª 2012 FEBS 2661

serum-deprived control cells (Fig. 8, lanes 6 and 2).
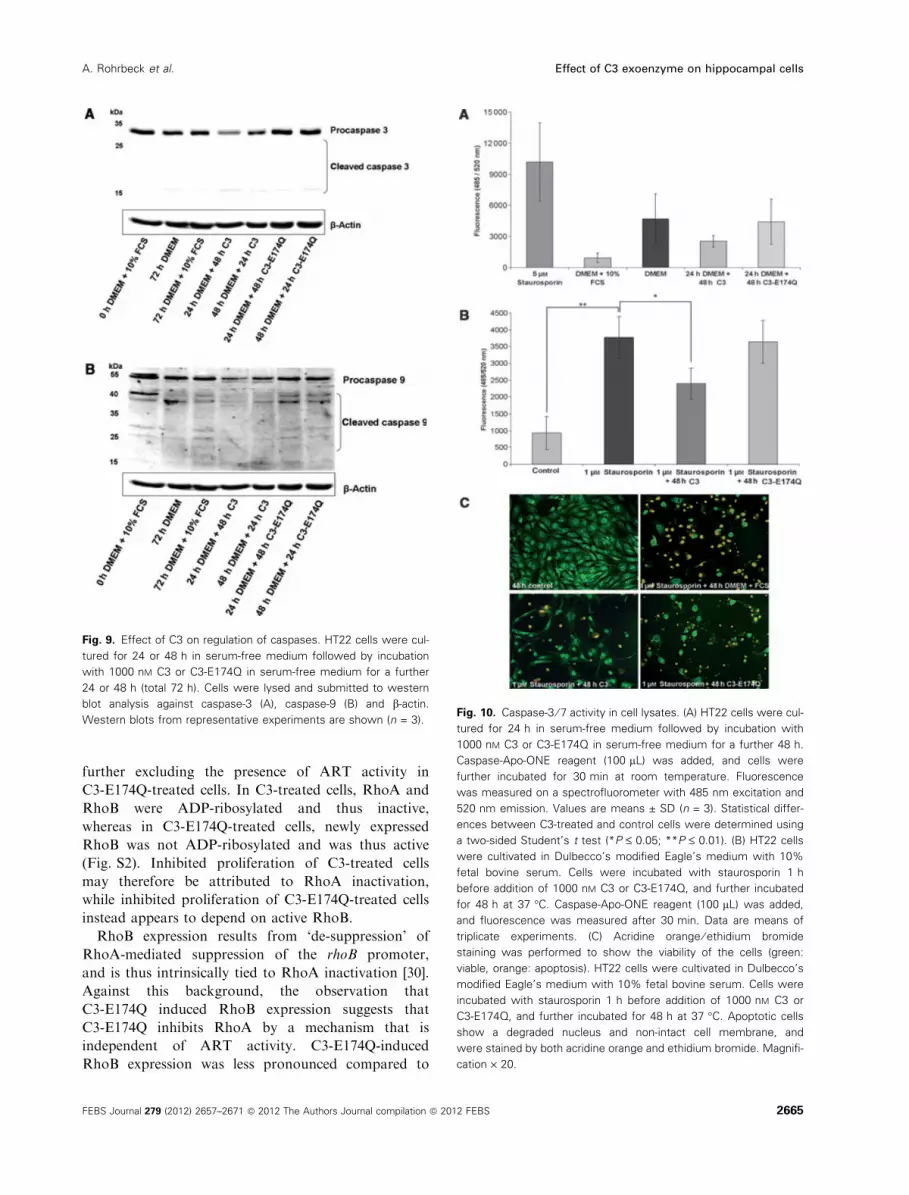
Similar results were found for the pro-forms of
caspase-3 and caspase-9 (Fig. 9).
Next, we determined the enzymatic activity of
caspase-3 using a caspase-3 ⁄ 7 ApoONE assay. Serum
starvation for 48 h induced was a threefold increase in
caspase-3 ⁄ 7 activity compared with control cells
(Fig. 10A). This finding strongly supports previous
reports showing that serum starvation results in induc-
tion of cell death [35,36]. Consistent with the results of
western blot analysis (Fig. 9), caspase-3 activity was
reduced in C3- but not in C3-E174Q-treated cells at
48 h after apoptosis induction.
To further provide evidence for the anti-apoptotic
activity of C3, staurosporin-induced apoptosis of
HT22 cells was analysed for its responsiveness to C3.
Staurosporin treatment strongly activated caspases in
HT22 cells. C3 (but not C3-E174Q) reduced staurospo-
rin-induced caspase activation (Fig. 10B).
Finally, staurosporine-induced loss of membrane
integrity was analysed using acridine orange ⁄ ethidiumbromide staining. Staurosporin-treated HT22 cells
lost their spindle-like morphology, shrank, detached
from the matrix and were stained orange, indicating
loss of membrane integrity. Loss of membrane integ-
rity was decreased in C3-treated cells but not in
C3-E174Q-treated cells, further indicating that the
anti-apoptotic activity of C3 depended on the ART
activity.
Discussion
Most cultured cell lines are almost insensitive to C3,
unless C3 has been fused to ectopic cell entry domains.
A previous study showing that C3 and C3-E174Q
induced biological effects in primary hippocampal neu-
rons [13] indicated that neuron-derived cells exhibit
remarkable sensitivity to C3bot independently of fused
cell entry domains. In this study, SV40-transformed
hippocampal neuronal HT22 cells were used as a cell
culture model to differentially study the biological
effects of C3 and C3-E174Q [37].
Fig. 5. RhoB up-regulation. (A) HT22 cells were exposed to 500 nM
C3-E174Q for up to 6 days or 500 nM C3 for 24 h at 37 �C. Cells
were lysed and submitted to western blot analysis for RhoA, RhoB
and b-actin (n = 3). (B) HT22 cells were exposed to 500 nM C3 or
C3-E174Q for up to 6 days. Cellular levels of RhoB proteins were
determined by western blot analysis and the signal intensity was
measured densiometrically (n = 3). The signal intensity was normal-
ized to the intensity of the corresponding b-actin signal. The differ-
ences in results were not statistically significant.
Fig. 6. Anti-proliferative effects of C3
exoenzyme. Cell numbers were counted in a
trypan blue exclusion assay using a
Neubauer counting chamber. Viable cells are
not detected by trypan blue but exclude the
dye. Incubation of HT22 cells with 500 nM of
wild-type C3 for 7 days caused a significant
inhibition in cell number 72 h after C3 appli-
cation compared to untreated cells. Results
were statistically significant relative to the
control at 72 h (with the exception of
C3-E174Q at 5 days) (P £ 0.05). Experiments
for each sample were performed in triplicate,
and three independent experiments were
performed (n = 9 for each value).
Effect of C3 exoenzyme on hippocampal cells A. Rohrbeck et al.
2662 FEBS Journal 279 (2012) 2657–2671 ª 2012 The Authors Journal compilation ª 2012 FEBS

C3 induced morphological changes in HT22 cells
(including cell rounding and pronounced formation of
retraction fibres), RhoA ADP-ribosylation and RhoA
degradation. Furthermore, C3 exhibited anti-prolifera-
tive and anti-apoptotic activity. These effects have
been reported for cell-permeable versions of C3 in var-
ious cellular systems including other neuronal cell lines
such as PC-12 cells. All these effects have been attri-
buted to C3-catalysed ADP-ribosylation. Consistent
with this view, we found that C3-induced cell rounding
as well as the anti-apoptotic activity strictly depend on
Rho ADP-ribosylation. In particular, apoptosis
induced by serum starvation or staurosporin is specifi-
cally inhibited by C3 (but not by C3-E174Q). Recently,
it was shown that inhibition of the Rho ⁄ROCK (Rho
associated kinase) pathway by C3 exoenzyme rescues
transplanted cells from apoptosis both in vitro and
in vivo [38]. Another study showed that inhibition of
RhoA activity by C3 attenuated thrombin-induced cell
death in cultured neurons and astrocytes [39]. In agree-
ment with these studies, we show down-regulation of
caspases and pro-apoptotic Bax in C3-treated HT22
cells in which apoptosis was induced by serum starva-
tion or staurosporin. In addition, we demonstrate inhi-
bition of caspase-3 activity by C3. All these findings
strongly suggest an anti-apoptotic effect of C3 in culti-
vated hippocampal HT22 cells. In animal models, ben-
eficial neuro-protective and neuro-regenerative effects
were observed with both C3 and C3-E174Q [40,41].
Possibly, anti-apoptotic or pro-apoptotic effects of C3
depends on tissue and cell types, but it appears that
C3 is predominantly anti-apoptotic in neuronal cells.
Whereas C3 specifically exhibited cell rounding and
anti-apoptotic activity, both C3 and C3-E174Q
reduced cell proliferation as well as cyclin D1 expres-
sion with comparable kinetics. Thus we conclude that
C3-E174Q enters HT22 cells comparably to C3.
C3-E174Q did not exhibit ART activity, as evidenced
Fig. 7. Effect of C3 on cell-cycle regulation
of HT22 cells. (A) Quantitative RT-PCR of
C3bot-treated cells. Summary of differential
expression of the cyclin D1 gene verified by
quantitative RT-PCR in comparison with
untreated control cells. Each experiment
was performed in triplicate and the runs
were repeated. Error bars indicate the
standard deviation among the samples.
Statistical differences between C3-treated
and control cells were determined using a
two-sided Student’s t test (*P £ 0.05;
**P £ 0.01; ***P £ 0.001). (B) For western
blot analysis, HT22 cells were exposed to
500 nM C3 or C3-E174Q for up to 6 days at
37 �C. Cells were lysed and subjected to
western blot analysis against cyclin D1
and b-actin. Western blots from representa-
tive experiments are shown (n = 3).
(C) Densitometric measurements for cyclin
D1 (means ± SE) from three independent
experiments. The results were not statisti-
cally significant.
A. Rohrbeck et al. Effect of C3 exoenzyme on hippocampal cells
FEBS Journal 279 (2012) 2657–2671 ª 2012 The Authors Journal compilation ª 2012 FEBS 2663
by analysing ART activity in the presence of high
C3-E174Q concentrations in a cell-free system or upon
prolonged treatment of HT22 with C3-E174Q for up
to 5 days. Not even traces of Rho ADP-ribosylation
were detected by the detection methods used (effector
pull-down assay, gel shift assay, direct and sequential
ADP-ribosylation). The biological activity of
C3-E174Q thus appeared to be independent of ART
activity. In addition to reduced cyclin D expression,
C3-E174Q induced (late) RhoB expression. The kinet-
ics of C3-E174Q-induced late RhoB expression clearly
differed from the kinetics of early RhoB expression
induced by C3. Notably, RhoB expressed in
C3-E174Q-treated cells was not ADP-ribosylated,
Fig. 8. Effect of C3 on apoptosis of HT22
cells. (A) HT22 cells were cultured for 24 or
48 h in serum-free medium followed by
incubation with 1000 nM C3 or C3-E174Q in
serum-free medium for a further 24 or 48 h
(total 72 h). Summary of differential expres-
sion of various apoptosis genes verified by
quantitative RT-PCR in comparison with
72 h serum-free control cells. Each experi-
ment was performed in triplicate and the
runs were repeated. Error bars indicate
standard deviation among the samples.
Statistical differences between C3-treated
and 72 h serum-free control cells were
determined using a two-sided Student’s t
test (*P £ 0.05; **P £ 0.01). (B) For
western blot analysis, cells were cultured
for 24 or 48 h in serum-free medium
followed by incubation with 1000 nM C3 or
C3-E174Q in serum-free medium for a
further 24 or 48 h (total 72 h) at 37 �C. Cells
were lysed and subjected to western blot
analysis for p53, Bax and b-actin. Western
blots from representative experiments are
shown (n = 3). (C) Densitometric measure-
ments for Bax (means ± SE) from five
independent experiments. Statistical
differences between C3-treated and 72 h
serum-free control cells were determined
using a two-sided Student’s t test
(*P £ 0.05).
Effect of C3 exoenzyme on hippocampal cells A. Rohrbeck et al.
2664 FEBS Journal 279 (2012) 2657–2671 ª 2012 The Authors Journal compilation ª 2012 FEBS
further excluding the presence of ART activity in
C3-E174Q-treated cells. In C3-treated cells, RhoA and
RhoB were ADP-ribosylated and thus inactive,
whereas in C3-E174Q-treated cells, newly expressed
RhoB was not ADP-ribosylated and was thus active
(Fig. S2). Inhibited proliferation of C3-treated cells
may therefore be attributed to RhoA inactivation,
while inhibited proliferation of C3-E174Q-treated cells
instead appears to depend on active RhoB.
RhoB expression results from ‘de-suppression’ of
RhoA-mediated suppression of the rhoB promoter,
and is thus intrinsically tied to RhoA inactivation [30].
Against this background, the observation that
C3-E174Q induced RhoB expression suggests that
C3-E174Q inhibits RhoA by a mechanism that is
independent of ART activity. C3-E174Q-induced
RhoB expression was less pronounced compared to
Fig. 9. Effect of C3 on regulation of caspases. HT22 cells were cul-
tured for 24 or 48 h in serum-free medium followed by incubation
with 1000 nM C3 or C3-E174Q in serum-free medium for a further
24 or 48 h (total 72 h). Cells were lysed and submitted to western
blot analysis against caspase-3 (A), caspase-9 (B) and b-actin.
Western blots from representative experiments are shown (n = 3).Fig. 10. Caspase-3 ⁄ 7 activity in cell lysates. (A) HT22 cells were cul-
tured for 24 h in serum-free medium followed by incubation with
1000 nM C3 or C3-E174Q in serum-free medium for a further 48 h.
Caspase-Apo-ONE reagent (100 lL) was added, and cells were
further incubated for 30 min at room temperature. Fluorescence
was measured on a spectrofluorometer with 485 nm excitation and
520 nm emission. Values are means ± SD (n = 3). Statistical differ-
ences between C3-treated and control cells were determined using
a two-sided Student’s t test (*P £ 0.05; **P £ 0.01). (B) HT22 cells
were cultivated in Dulbecco’s modified Eagle’s medium with 10%
fetal bovine serum. Cells were incubated with staurosporin 1 h
before addition of 1000 nM C3 or C3-E174Q, and further incubated
for 48 h at 37 �C. Caspase-Apo-ONE reagent (100 lL) was added,
and fluorescence was measured after 30 min. Data are means of
triplicate experiments. (C) Acridine orange ⁄ ethidium bromide
staining was performed to show the viability of the cells (green:
viable, orange: apoptosis). HT22 cells were cultivated in Dulbecco’s
modified Eagle’s medium with 10% fetal bovine serum. Cells were
incubated with staurosporin 1 h before addition of 1000 nM C3 or
C3-E174Q, and further incubated for 48 h at 37 �C. Apoptotic cells
show a degraded nucleus and non-intact cell membrane, and
were stained by both acridine orange and ethidium bromide. Magnifi-
cation · 20.
A. Rohrbeck et al. Effect of C3 exoenzyme on hippocampal cells
FEBS Journal 279 (2012) 2657–2671 ª 2012 The Authors Journal compilation ª 2012 FEBS 2665

C3-induced RhoB expression, allowing the conclusion
that C3-E174Q inhibited RhoA less efficiently com-
pared to C3. This view is consistent with the observa-
tion that C3 (but not C3-E174Q) reduced the cellular
level of RhoA-GTP in HT22 cells. The mechanism by
which C3-E174Q inhibits RhoA is most likely distinct
from a covalent modification and may include RhoA
sequestration. This is supported by the findings of
microscale thermophoresis. In these experiments, RhoA
binds with four- to fivefold higher affinity to C3-E174Q
than to C3. It is conceivable that RhoA is retained in
the cytosol through sequestration by C3-E174Q.
In summary, we demonstrate that murine hippocam-
pal HT22 cells are sensitive to both C3 and C3-E174Q.
C3 induced cell rounding, inhibition of cell prolifera-
tion and anti-apoptotic effects. C3-E174Q induced
RhoB expression and inhibition of cell proliferation
independently of ART activity, with RhoB probably
being involved in anti-proliferative effects.
Experimental procedures
Cell culture
Murine hippocampal HT22 cells were cultivated in Dul-
becco’s modified essential medium (Biochrom AG, Berlin,
Germany, +10% fetal bovine serum, 100 gÆmL)1 penicillin,
100 unitsÆmL)1 streptomycin and 1 mM sodium pyruvate)
at 37 �C and 5% CO2. Upon confluence, cells were pas-
saged.
For the growth kinetics experiments, cells were seeded
onto 3.5 cm plates at a concentration of 50 000 cellsÆmL)1.
Cells were grown for 24 h before treatment, and then C3
exoenzyme from Clostridium botulinum was added to the
medium at a concentration of 500 nM. After 12, 24 and
48 h, the medium was removed and the cells were washed
with NaCl ⁄Pi.
After the indicated time points, cells were detached with
0.25% trypsin solution. A 1 mL volume of culture medium
was added to neutralize the trypsin action. Cells were
counted every day by the trypan blue exclusion assay using
a Neubauer counting chamber (Carl Roth GmbH, Karls-
ruhe, Germany) to determine the number of viable cells.
For apoptosis experiments, cells were seeded onto 3.5 cm
plates at a concentration of 100 000 cellsÆmL)1. After 24 h,
the medium was changed to serum-free medium, and 48 h
later, cells were treated with 1000 nM C3 in serum-free
medium. The medium was replaced every 48 h by new
serum-free protein-containing medium.
Cell lysis for western blot analysis
After toxin treatment, cells were washed and scraped into
Laemmli sample buffer. The obtained suspension was sha-
ken at 37 �C for 10 min. Ultrasonic disruption was per-
formed using a cycle of 10 · 5 s, 5 · 10% sonic energy
with a sonotrode (Bandelin Electronic, Berlin, Germany).
The lysate was then incubated at 95 �C for 10 min and sub-
jected to SDS ⁄PAGE.
Western blot analysis
Complete lysate proteins were separated using SDS ⁄PAGE
and subsequently transferred onto nitrocellulose membranes
by a tank blot system. The membranes were blocked using
5% w ⁄ v non-fat dried milk for 60 min; incubation with pri-
mary antibody was performed overnight at 4 �C, and treat-
ment with the secondary antibody was performed at room
temperature for 1 h. For immunoblotting, the following
primary antibodies were used: RhoA was identified using a
mouse monoclonal IgG from Santa Cruz Biotechnologies
(Santa Cruz, CA, USA); antibodies against RhoB (rabbit
polyclonal IgG) were obtained from Bethyl Laboratories
(Montgomery, TX, USA); Bax (p-19) monoclonal antibody
was purchased from Santa Cruz Biotechnologies, pAkt
(Ser473), caspase-3, p53 and cyclin D1 antibodies were pur-
chased from Cell Signaling Technologies, and caspase-9
rabbit polyclonal antibody antibody was purchased from
Biomol GmbH (Hamburg, Germany); Identification of
C3bot was achieved using an affinity-purified rabbit poly-
clonal antibody that was raised against the full-length
exoenzyme C3bot (accession number CAA41767). Actin
(Sigma-Aldrich, Munich, Germany) was used as a loading
control.
For the chemiluminescence reaction, ECL Femto (Pierce ⁄Thermo Fisher Scientific Inc., Rockford, IL, USA) or
Immobilon (Millipore, Schwalbach, Germany) was used.
All signals were analysed densitometrically using KODAK
1D software (Kodak GmbH, Stuttgart, Germany) and
normalized to b-actin signals.
Expression and purification of recombinant
C3 proteins
C3 wild-type and C3-E174Q were expressed as recombinant
glutathione S-transferase (GST) fusion proteins in Escheri-
chia coli TG1 harbouring the respective DNA fragment in
plasmid pGEX-2T (Amersham Life Sciences, Arlington
Heights, IL, USA). Bacteria were grown at 37 �C in 1 L of
LB medium containing 100 lgÆmL)1 ampicillin to an atten-
uance at 595 nm of 0.7. Isopropyl-thio-b-D-galactoside was
added to a final concentration of 0.2 mM, and the cultures
were incubated at 37 �C for another 3 h. Bacteria were sed-
imented at 7700 g (15 min, 4 �C) and resuspended in
20 mL lysis buffer (50 mM Tris, pH 8, 150 mM NaCl, 2 mM
MgCl2, 2 mM dithiothreitol), complemented with EDTA-
free protease inhibitor cocktail (Roche Pharma AG, Gren-
zach-Wyhlen, Germany). Bacteria were lysed by means of a
French pressure cell press system (SIM Aminco Spectronic
Effect of C3 exoenzyme on hippocampal cells A. Rohrbeck et al.
2666 FEBS Journal 279 (2012) 2657–2671 ª 2012 The Authors Journal compilation ª 2012 FEBS

Instruments, New York, USA), and the bacterial debris
was sedimented at 12 000 g (15 min, 4 �C). The supernatant
was added to 4 mL of a 50% slurry of glutathione–
Sepharose 4B beads in lysis buffer, and incubated for 4 h at
4 �C while shaking. The slurry was poured into a dis-
posable Econo-Pac� chromatography column (Bio-Rad
Laboratories GmbH, Munchen, Germany) and washed five
times with 10 bed volumes of lysis buffer. For cleavage of
the fusion proteins from glutathione S-transferase, the
beads were incubated with five ‘‘NIH’’ units of thrombin
for 12 h at 4 �C. Thereafter, the protein was eluted with
12 mL lysis buffer. Thrombin was removed by precipitation
with benzamidine–Sepharose beads (AP Biosciences, New
York, USA). After centrifugation at 500 g (10 min, room
temperature), the supernatant was exchanged with 20 mM
HEPES pH 7.5 using PD-10 columns (GE healthcare,
Munchen, Germany), sterile-filtered (0.22 lm) and used for
cell culture experiments as indicated. ART activity was
measured by an in vitro ADP-ribosylation assay.
ADP-ribosylation of Rho in HT22 cells
To verify the effectiveness of ADP-ribosylation of Rho by
C. botulinum exoenzyme C3, the HT22 cells were either
left untreated or were incubated with increasing concentra-
tions of the exotoxin (10–1000 nM) for 8–24 h. The cells
were then washed with NaCl ⁄Pi and scraped into 100 lLof lysis buffer (20 mM Tris ⁄HCl (pH 7.4), 1% Triton
X-100, 10 mM NaCl, 5 mM MgCl2, 1 mM phen-
ylmethanesulfonyl fluoride, 5 mM dithiothreitol). The
obtained suspension was shaken at 37 �C for 10 min.
Ultrasonic disruption was performed using a cycle of
10 · 5 s, 5 · 10% sonic energy with a sonotrode (Bandelin
Electronic). Protein concentrations were measured by the
Bradford method. Cell lysates containing equal amounts
of protein were incubated with 1000 nM recombinant
C. botulinum exoenzyme C3 and 1 lCi [32P]NAD (Amer-
sham Life Sciences, Arlington Heights, IL, USA) in 20 lLof 4· buffer containing 50 mM HEPES (pH 7.3), 10 mM
MgCl2, 10 mM dithiothreitol, 10 mM thymidine and 10 lM
NAD at 37 �C for 20 min. The reaction was terminated
by addition of Laemmli sample buffer, and then incubated
at 95 �C for 10 min. Samples were resolved by
SDS ⁄PAGE on 15% gels, and the ADP-ribosylated Rho
was analyzed by phosphorimaging (Cyclone, Packard
American Instrument, MA, USA).
Pull-down assay
RhoA activity was measured by pull-down assay using a
GST fusion protein of the Rho binding domain. HT22 cells
were homogenized in lysis buffer [20 mM Tris ⁄HCl (to a
total of 25 lL) pH 7.4], 1% Triton X-100, 10 mM NaCl,
5 mM MgCl2, 1 mM phenylmethanesulfonyl fluoride, 5 mM
dithiothreitol). Cleared lysates were incubated for 60 min at
4 �C with GST–Rho binding domain-coupled beads (20–
30 lg per sample). Precipitates were washed three times
with the binding buffer and suspended in the SDS sample
buffer. Proteins were separated by 15% SDS ⁄PAGE and
transferred to a nitrocellulose membrane. GTP-bound
RhoA and total RhoA were detected by western blot analy-
sis using a monoclonal RhoA antibody (Santa Cruz Bio-
technology).
RNA isolation
RNA extraction was performed using an RNeasy Mini Kit
(Qiagen, Santa Clarita, CA, USA) according to the manu-
facturer’s instructions. The RNA concentration of the sam-
ples was then measured using a NanoDrop ND-1000
spectrophotometer (PeqLab Biotechnology GmbH, Erlan-
gen, Germany).
RT-PCR
RNA (2 lg) from each sample was mixed with an appro-
priate quantity of water (to a total of 25 lL) and dena-
tured for 5 min at 65 �C by using an Eppendorf
Thermomixer (Eppendorf AG, Hamburg, Germany). The
reaction mix for the reverse transcription comprised 2 lL10· reverse transcription buffer (Omniscript RT-Kit,
Qiagen, Hilden, Germany), 2 lL dNTPs, 2 lL random
hexamers (primers), 0.25 lL RNasin and 1 lL reverse
transcriptase (Omniscript RT Kit, Qiagen, Hilden, Ger-
many), and was added previously cooled samples. The
composition of the mix is appropriate for an RNA con-
centration between 50 ng and 2 lg. The samples were
subsequently incubated for 1 h at 37 �C. During this step,
the RNA is converted to cDNA. The enzymes were then
denaturated by incubation for 5 min at 95 �C in the Ep-
pendorf Thermomixer. Finally the samples were cooled
and frozen at )20 �C.Real-time RT-PCR measurements were performed using
an ABI PRISM 7500 sequence detection system instrument
(Applied Biosystems, Carlsbad, CA, USA). PCR reactions
were performed according to the manufacturer’s instruc-
tions using GoTaq� qPCR Master Mix (Promega GmbH,
Mannheim, Germany). Gene expression levels were normal-
ized to those of b2-microglobulin, which was found to be
stably expressed. The specificity of primers was confirmed
by agarose-gel electrophoresis of PCR products.
Each experiment was performed in triplicate and the runs
were repeated. Mean CT values were calculated using the
ABI PRISM software, and relative gene expression levels were
expressed as the difference between mean CT values for the
target gene and the control gene b2-microglobulin. A nega-
tive control was always included in RT-PCR experiments.
Additionally, a paired two-tailed Student’s t test was per-
formed, and the results were considered significant when
the P value was <0.05.
A. Rohrbeck et al. Effect of C3 exoenzyme on hippocampal cells
FEBS Journal 279 (2012) 2657–2671 ª 2012 The Authors Journal compilation ª 2012 FEBS 2667

The primers (Table 1) were designed in several steps as
described below. The mRNA sequences of the single genes
were found on http://www.ncbi.nlm.nih.gov/. The cDNA
sequences of the same genes were found on http ⁄ ⁄genome.ucsc.edu ⁄ cgi-bin ⁄ hgGateway. Primer design was
performed using web tool Primer3Plus (http://www.
bioinformatics.nl/cgi-bin/primer3plus/primer3plus.cgi). We
used the UCSC Genome Browser (http://genome.ucsc.edu)
to control the intron spanning of primers. PCR primers
were synthesized by Eurofins (Ebersberg, Germany).
Caspase activity assay
Caspase-3 ⁄ 7 activity was measured using the Apo-ONE�homogenous caspase-3 ⁄ 7 assay (Promega GmbH, Man-
nheim, Germany). Cells were seeded in 96-well plates, and
the next day, the medium was replaced with serum-free
medium for 48 h. After apoptosis induction, cells were
incubated for 24 and 48 h with 1000 nM C3 in serum-free
medium. Cells were lysed, and caspase-3 ⁄ 7 activity was
measured by cleavage of the caspase-3 ⁄ 7 substrate rhoda-
mine 110 [bis-(N-benzyloxycarbonyl-L-aspartyl-L-glutamyl-L-
valyl-L-aspartic acid amide)] (Z-DEVD-R110). Samples were
measured on a spectrofluorometer with 485 nm excitation
and 520 nm emission. Control cells were treated with 5 lM
staurosporin.
Staurosporin from Streptomyces spheroides (Sigma-
Aldrich, St. Louis, MO, USA) was kept in a stock solution
of 10 mM in 100% dimethyl sulfoxide and diluted to a final
concentration of 1 lM in serum-free medium before incuba-
tion. Controls were incubated with a corresponding concen-
tration of dimethyl sulfoxide. When the effect of
staurosporin concentration was investigated, final concen-
trations of 5000 and 1000 nM were used. Cells were incu-
bated with staurosporin 1 h before addition of C3.
Acridine orange ⁄ ethidium bromide staining of
cells
To determine cell viability, cells were subjected to acridine
orange ⁄ ethidium bromide staining. Cells were treated with
1 lM staurosporin for 1 h, then washed with NaCl ⁄Pi, and
1000 nM C3 or C3-E174Q was added. After 48 h, 10 lL of
a solution containing 5 mgÆmL)1 acridine orange and
10 mgÆmL)1 ethidium bromide was added. The cells were
then visualized by fluorescence microscopy (Zeiss Axiovert
200 M, Carl Zeis GmbH, Gottingen, Germany), using a
blue filter.
Microscale thermophoresis binding analysis
Thermophoresis was used to measure the binding interactions
between RhoA and C3 exoenzyme. For the experiments,
RhoA was labelled using Monolith NT� Protein Labeling
Kit Red-NHS (NanoTemper Technologies GmbH, Munich,
Germany) according to the manufacturer’s instructions. NT-
647-labelled RhoA was used at a concentration of 250 nM.
C3 or C3-E174Q was titrated in 1 : 1 dilutions starting at
30 lM (16 samples of a serial dilution). The experiments were
performed in HEPES buffer with 0.01% Tween in standard
treated capillaries (NanoTemper Technologies GmbH).
The measurements were performed using a NanoTemper
Monolith NT.015 instrument. All measurements were per-
formed at 20%, 40% and 80% IR-Laser (infra red-Laser),
with a laser-on time of 15 s, and a laser-off time of 5 s.
Table 1. Primer pairs used for quantitative RT-PCR (Ccnd1 ¼ cyclin D1; Bax ¼ BCL2-associated X protein; Casp9 ¼ caspase 9; Casp3 ¼caspase 3; Casp8 ¼ caspase 8; Apaf1 ¼ apoptotic peptidase activating factor 1; BID ¼ BH3 interacting domain death agonist; B2M ¼ beta-2
microglobulin).
Gene Reference sequence Primer Length (bp) Product (bp)
Proliferation Ccnd1 NM_007631.2 Forward: AGTGCGTGCAGAAGGAGATT
Reverse: CACAACTTCTCGGCAGTCAA
20 238
Apoptosis Bax NM_007527.3 Forward: CGAGCTGATCAGAACCATCA
Reverse: CTCAGCCCATCTTCTTCCAG
20 191
Casp9 NM_015733.4 Forward: ACGCTCTGCTGAGTCGAGA
Reverse: CCTTGGCCTGTGTCCTCTAA
19
20
169
Casp3 NM_009810.2 Forward: GGGCCTGTTGAACTGAAAAA
Reverse: CCGTCCTTTGAATTTCTCCA
20 242
Casp8 NM_009812.2 Forward: GCGTGGAACAGGAAGTGAGT
Reverse: TGGGATGTAGTCCAAGCACA
20 248
Apaf1 NM_009684.2 Foward: GTTCAAAGCCGAGACAGGAG
Reverse: ATTGACTTGCTCCGAGTGCT
20 187
BID NM_007544.3 Forward: TCACAGACCTGCTGGTGTTC
Reverse: GTCTGGCAATGTTGTGGATG
20 219
Housekeeping B2M NM_009735.3 Forward: ATTCACCCCCACTGAGACTG
Reverse: GCTATTTCTTTCTGCGTGCAT
20
21
192
Effect of C3 exoenzyme on hippocampal cells A. Rohrbeck et al.
2668 FEBS Journal 279 (2012) 2657–2671 ª 2012 The Authors Journal compilation ª 2012 FEBS

Sixteen subsequent thermophoresis measurements for each
IR-Laser option were performed to determine the binding
affinity. Plotting the normalized fluorescence Fnorm =
Fhot ⁄Fcold at given time t against the labelled RhoA concen-
tration produced a binding curve. The binding curves were
fitted, and Kd values were obtained.
Reproducibility of the experiments and statistics
All experiments were performed independently at least three
times. Results from representative experiments are shown in
the figures. Values (n ‡ 3) are means ± SEM. The two-
sided unpaired Student’s t test was used throughout the
study to assess the statistical significance of the differences
between two sets of data. Differences were considered to be
statistically significant at P £ 0.05.
Acknowledgements
We thank Moran Jerabek-Willemsen (NanoTem-
per Technologies GmbH, Munich, Germany) and
Professor Dietmar J. Manstein (Department of
Biophysical Chemistry, Medical School of Hanover,
Germany) for kind support with microscale thermo-
phoresis experiments. This work was supported by
funding from the Deutsche Forschungsgemeinschaft
(project JU231 ⁄ 5).
References
1 Hall A (1998) Rho GTPases and the actin cytoskeleton.
Science 279, 509–514.
2 To KC, Church J & O’Connor TP (2008) Growth cone
collapse stimulated by both calpain- and Rho-mediated
pathways. Neuroscience 153, 645–653.
3 Ellis S & Mellor H (2000) Regulation of endocytic
traffic by rho family GTPases. Trends Cell Biol 10,
85–88.
4 Michaelson D, Silletti J, Murphy G, D’Eustachio P,
Rush M & Philips MR (2001) Regulation by hypervari-
able regions and RhoGDI binding. J Cell Biol 152,
111–126.
5 Wallar BJ, Deward AD, Resau JH & Alberts AS (2007)
RhoB and the mammalian Diaphanous-related formin
mDia2 in endosome trafficking. Exp Cell Res 313, 560–
571.
6 Engel ME, Datta PK & Moses HL (1998) RhoB is
stabilized by transforming growth factor b and antago-
nizes transcriptional activation. J Biol Chem 273, 9921–
9926.
7 Fritz G, Kaina B & Aktories K (1995) The ras-related
small GTP-binding protein RhoB is immediate-early
inducible by DNA damaging treatments. J Biol Chem
270, 25172–25177.
8 Ho TT, Merajver SD, Lapiere CM, Nusgens BV &
Deroanne CF (2008) RhoA-GDP regulates RhoB
protein stability. Potential involvement of RhoGDIa.J Biol Chem 283, 21588–21598.
9 Just I, Huelsenbeck SC & Genth H (2010) Clostridium
botulinum C3 exoenzyme: Rho-inactivating tool in cell
biology and a neurotrophic agent. Open Toxinol J 3,
19–23.
10 Paterson HF, Self AJ, Garrett MD, Just I, Aktories K
& Hall A (1990) Microinjection of recombinant p21rho
induces rapid changes in cell morphology. J Cell Biol
111, 1001–1007.
11 Shiokawa S, Sakai K, Akimoto Y, Suzuki N,
Hanashi H, Nagamatsu S, Iwashita M, Nakamura Y,
Hirano H & Yoshimura Y (2000) Function of the
small guanosine triphosphate-binding protein RhoA
in the process of implantation. J Clin Endocrinol
Metab 85, 4742–4749.
12 Just I, Rohrbeck A, Huelsenbeck SC & Hoeltje M
(2011) Therapeutic effects of Clostridium botulinum C3
exoenzyme. Naunyn Schmiedebergs Arch Pharmacol 383,
247–252.
13 Ahnert-Hilger G, Holtje M, Große G, Pickert G,
Mucke C, Nixdorf-Bergweiler B, Boquet P, Hofmann F
& Just I (2004) Differential effects of Rho GTPases on
axonal and dendritic development in hippocampal neu-
rons. J Neurochem 90, 9–18.
14 Kozma R, Sarner S, Ahmed S & Lim L (1997) Rho
family GTPases and neuronal growth cone remodelling:
relationship between increased complexity induced by
Cdc42Hs, Rac1, and acetylcholine and collapse induced
by RhoA and lysophosphatidic acid. Mol Cell Biol 17,
1201–1211.
15 Jin Z & Strittmatter SM (1997) Rac1 mediates collaps-
ing-1-induced growth cone collapse. J Neurosci 17,
6256–6263.
16 Nishiki T, Narumiya S, Morii N, Yamamot M,
Fujiwara M, Kamata Y, Sakaguchi G & Kozaki S
(1990) ADP-ribosylation of the rho ⁄ rac proteins
induces growth inhibition, neurite outgrowth and
acetylcholine esterase in cultured PC-12 cells. Biochem
Biophys Res Commun 167, 265–272.
17 Watts KL, Cottrell E, Hoban PR & Spiteri MA (2006)
RhoA signaling modulates cyclin D1 expression in
human lung fibroblasts; implications for idiopathic
pulmonary fibrosis. Respir Res 7, 88.
18 Danen EHJ, Sonnenveld P, Sonnenberg A & Yamada
KM (2000) Dual stimulation of Ras ⁄mitogen-activated
protein kinase and RhoA by cell adhesion to
fibronectin supports growth factor stimulated
cell cycle progression. J Cell Biol 151,
1413–1422.
19 Adnane J, Bizouarn FA, Qian Y, Hamilton AD &
Sebti SM (1998) p21WAF1 ⁄CIP1 is upregulated by the
A. Rohrbeck et al. Effect of C3 exoenzyme on hippocampal cells
FEBS Journal 279 (2012) 2657–2671 ª 2012 The Authors Journal compilation ª 2012 FEBS 2669

geranylgeranyltransferase I inhibitor GGTI-298 through
a transforming growth factor b- and Sp1-responsive
element: involvement of the small GTPase RhoA.
Mol Cell Biol 18, 6962–6970.
20 Weber JD, Hu W, Jefcoat SC Jr, Raben DM & Baldas-
sare JJ (1997) Ras-stimulated extracellular signal-related
kinase 1 and RhoA activities coordinate platelet-derived
growth factor-induced G1 progression through the inde-
pendent regulation of cyclin D1 and p27. J Biol Chem
272, 32966–32971.
21 Du W & Prendergast GC (1999) Geranylgeranylated
RhoB mediates inhibition of human tumor cell
growth by farnesyltransferase inhibitors. Cancer Res 59,
5924–5928.
22 van Golen KL, Wu ZF, Qiao XT, Bao LW & Marajver
SD (2000) RhoC GTPase, a novel transforming onco-
gene for human mammary epithelial cells that partially
recapitulates the inflammatory breast cancer phenotype.
Cancer Res 60, 5832–5838.
23 Moorman JP, Bobak DA & Hahn CS (1996) Inactiva-
tion of the small GTP binding protein Rho induces
multinucleate cell formation and apoptosis in murine
T lymphoma EL4. J Immunol 156, 4146–4153.
24 Bobak D, Moorman J, Guanzon A, Gilmer L & Hahn
C (1997) Inactivation of the small GTPase Rho disrupts
cellular attachment and induces adhesion-dependent
and adhesion-independent apoptosis. Oncogene 15,
2179–2189.
25 Krijnen PA, Sipkens JA, Molling JW, Rauwerda JA,
Stehouwer CD, Muller A, Paulus WJ, van Nieuw
Amerongen GP, Hack CE, Verhoeven AJ et al. (2010)
Inhibition of Rho–ROCK signaling induces apoptotic
and non-apoptotic PS exposure in cardiomyocytes via
inhibition of flippase. J Mol Cell Cardiol 49, 781–790.
26 Bertrand J, Di Polo A & McKerracher L (2007)
Enhanced serviva and regenration of axotomized retinal
neurons by repeated delivery of cell-permeable C3-like
Rho antagonists. Neurobiol Dis 25, 65–72.
27 Dubreuil CI, Winton MJ & McKerracher L (2003) Rho
activation patterns after spinal cord injury and the role
of activated Rho in apoptosis in the central nervous
system. J Cell Biol 162, 233–243.
28 Genth H, Schmidt M, Gerhard R, Aktories K & Just I
(2003) Activation of phospholipase D1 by ADP-ribosy-
lated RhoA. Biochem Biophys Res Commun 302,
127–132.
29 Genth H, Gerhard R, Maeda A, Amano M, Kaibuchi
K, Aktories K & Just I (2003) Entrapment of Rho
ADP-ribosylated by Clostridium botulinum C3 exoen-
zyme in the Rho-guanine nucleotide dissociation inhibi-
tor-1 complex. J Biol Chem 278, 28523–28527.
30 Hulsenbeck J, Dreger SC, Gerhard R, Fritz G, Just I &
Genth H (2007) Upregulation of the immediate early
gene product RhoB by exoenzyme C3 from Clostridium
limosum and toxin B from Clostridium difficile.
Biochemistry 46, 4923–4931.
31 Wang X, Chen Y, Wang Y, Zhu X, Ma Y, Zhang S &
Lu J (2009) Role of RHOB in the antiproliferative
effect of glucocorticoid receptor on macrophage
RAW264.7 cells. J Endocrinol 200, 35–43.
32 Chassot AA, Lossaint G, Turchi L, Meneguzzi G,
Fisher D, Ponzio G & Dulic V (2008) Confluence-
induced cell cycle exit involves pre-mitotic CDK inhibi-
tion by p27(Kip1) and cyclin D1 downregulation. Cell
Cycle 7, 2038–2046.
33 Hippenstiel S, Schmeck B, N’Guessan PD, Seybold J,
Krull M, Preissner K, Eichel-Streiber CV & Suttorp N
(2002) Rho protein inactivation induced apoptosis of
cultured human endothelial cells. Am J Physiol Lung
Cell Mol Physiol 283, L830–L838.
34 Mills JC, Stone NL, Erhardt J & Pittman RN (1998)
Apoptotic membrane blebbing is regulated by myosin
light chain phosphorylation. J Cell Biol 140,
627–636.
35 Gerber HP, Dixit V & Ferrara N (1998) Vascular endo-
thelial growth factor induces expression of the antia-
poptotic proteins Bcl-2 and A1 in vascular endothelial
cells. J Biol Chem 273, 13313–13316.
36 Fujio Y, Guo K, Mano T, Mitsuuchi Y, Testa JR &
Walsh K (1999) Cell cycle withdrawal promotes myo-
genic induction of Akt, a positive modulator of myocyte
survival. Mol Cell Biol 19, 5073–5082.
37 Liu J, Li L & Suo WZ (2009) HT22 hippocampal
neuronal cell line possesses functional cholinergic
properties. Life Sci 84, 267–271.
38 Koyanagi M, Takahashi J, Arakawa Y, Doi D,
Fukuda H, Hayashi H, Narumiya S & Hashimoto N
(2008) Inhibition of the Rho ⁄ROCK pathway reduces
apoptosis during transplantation of embryonic stem
cell-derived neural precursors. J Neurosci Res 86, 270–
280.
39 Donovan FM, Pike CJ, Cotman CW & Cunningham
DD (1997) Thrombin induces apoptosis in cultured
neurons and astrocytes via a pathway requiring tyro-
sine kinase and RhoA activities. J Neurosci 17, 5316–
5326.
40 Boato F, Hendrix S, Huelsenbeck SC, Hofmann F,
Grosse G, Djalali S, Klimaschewski L, Auer M, Just I,
Ahnert-Hilger G et al. (2010) C3 peptide enhances
recovery from spinal cord injury by improved regenera-
tive growth of descending fiber tracts. J Cell Sci 123,
1652–1662.
41 Holtje M, Djalali S, Hofmann F, Munster-Wandowski
A, Hendrix S, Boato F, Dreger SC, Grosse G, Henne-
berger C, Grantyn R et al. (2009) A 29-amino acid frag-
ment of Clostridium botulinum C3 protein enhances
neuronal outgrowth, connectivity, and reinnervation.
FASEB J 23, 1115–1126.
Effect of C3 exoenzyme on hippocampal cells A. Rohrbeck et al.
2670 FEBS Journal 279 (2012) 2657–2671 ª 2012 The Authors Journal compilation ª 2012 FEBS

Supporting information
The following supplementary information is available:
Fig. S1. ADP-ribosylation of recombinant GST-RhoA
by C3.
Fig. S2. RhoB up-regulation in C3- and C3-E174Q-
treated HT22 cells.
This supplementary material can be found in the
online version of this article.
Please note: As a service to our authors and readers,
this journal provides supporting information supplied
by the authors. Such materials are peer-reviewed and
may be reorganized for online delivery, but are not
copy-edited or typeset. Technical support issues arising
from supporting information (other than missing files)
should be addressed to the authors.
A. Rohrbeck et al. Effect of C3 exoenzyme on hippocampal cells
FEBS Journal 279 (2012) 2657–2671 ª 2012 The Authors Journal compilation ª 2012 FEBS 2671




















