
Bone phenotype of the aromatase deficient mouse
11
Journal of Steroid Biochemistry & Molecular Biology 79 (2001) 49–59 Bone phenotype of the aromatase deficient mouse Orhan K. Öz a,∗ , Gen Hirasawa a , Jonathan Lawson a , Lydia Nanu a , Anca Constantinescu a , Peter P. Antich a , Ralph P. Mason a , Edward Tsyganov a , Robert W. Parkey a , Joseph E. Zerwekh b , Evan R. Simpson c a Department of Radiology, UT Southwestern Medical Center at Dallas, Dallas, TX 75390, USA b Department of Internal Medicine, UT Southwestern Medical Center at Dallas, Dallas, TX 75390, USA c Victorian Breast Cancer Consortium, Prince Henry’s Institute of Medical Research, P.O. Box 5152, Clayton, Vic., Australia Abstract Estrogens are important for normal bone growth and metabolism. The mechanisms are incompletely understood. Thus, we have un- dertaken characterization of the skeletal phenotype of aromatase (ArKO) deficient mice. No abnormalities have been noted in skeletal patterning in newborns. Adult ArKO mice show decreased femur length and decreased peak Bone Mineral Density (BMD) with acceler- ated bone loss by 7 months of age in females. Magnetic resonance microscopy (MR) and microCT (CT) imaging disclosed decreased cancellous connectivity and reduced cancellous bone volume in ArKO females. Bone formation rate (BFR) is increased in ArKO females and decreased in ArKO males. Estradiol therapy reverses these changes. This anabolic effect of estradiol in the male skeleton is supported by 18-F − Positron Emission Tomography (PET) imaging, which clearly demonstrates decreased spinal uptake, but marked increase after estradiol therapy. Serum IGF-1 levels are high in young female ArKO mice but low in young ArKO males. The reduced BMD in ArKO females, despite the presence of elevated serum IGF 1, suggests that other mechanism(s) are operative. There is increased B-cell lym- phopoiesis in adult female ArKO bone marrow cells. These results show that ArKO mice show the effects of estrogen deficiency on bone growth, mass, metabolism, microarchitecture and the hematopoietic microenvironment. © 2002 Elsevier Science Ltd. All rights reserved. Keywords: Aromatase; Bone; Dual energy X-ray absorptiometry; Estrogen; Histomorphometry; Insulin-like growth factor 1; Major urinary protein 1. Introduction Estrogens are known to be important in the establishment and maintenance of normal bone mass in women and men. In particular, loss of estrogen at the menopause leads to ac- celerated bone loss in women [1] and is also contributory to the slow bone loss seen with aging in men [1,2]. Though the mechanisms of estrogens’ actions in maintenance of bone are incompletely understood, it is clear that they act ultimately by controlling the balance between resorption or osteoclastic activity and bone formation or osteoblastic activity. While most research has focused on estrogens’ effects on osteoclas- togenesis, recent evidence suggests that estrogen also stim- ulates osteoblast activity [3]. Cytokines and growth factors are known mediators of these effects on bone [4]. Moreover estrogens are necessary for acquisition of peak Bone Min- eral Density (BMD) in the maturing skeletons of both sexes (reviewed in [5]). In terms of the accrual of bone mass to Proceedings of the Symposium: ‘Aromatase 2000 and the Third Generation’ (Port Douglas, Australia, 3–7 November 2000). ∗ Corresponding author. Fax: +1-214-648-2727. E-mail address: [email protected] (O.K. Öz). peak BMD, the GH–IGF-1 axis is known to be important and in at least one case of estrogen deficiency, Turner’s syn- drome, there are abnormalities in GH secretion. This raises the possibility of a causal relationship. Estrogens also regulate the hematopoietic component of bone. Ovariectomy, though not a pure estrogen defi- cient state, results in increased B-cell lymphopoiesis in the mouse bone marrow. This and the associated bone loss can be prevented by estrogen administration [6]. On the other hand, the osteosclerosis induced by high doses of estrogen [7] is preceded by suppression of hematopoiesis including B-cell lymphopoiesis [8]. The osteogenic precursor popu- lation increases, but after the effects on the hematopoietic component. Thus, estrogens’ anti-resorptive and poten- tial pro-formation effects may involve regulation of the hematopoietic component of the marrow. Estrogens are synthesized from androgen precursors by the enzyme aromatase. In order to unravel the mechanisms underlying the multiple effects of estrogens animal mod- els are necessary. The aromatase deficient mouse or ArKO mouse [9] is such a model. We have previously reported that ArKO have reduced cancellous bone volume and a sexually dimorphic pattern of remodeling, high turnover in females 0960-0760/02/$ – see front matter © 2002 Elsevier Science Ltd. All rights reserved. PII:S0960-0760(01)00130-3
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Bone phenotype of the aromatase deficient mouse

Journal of Steroid Biochemistry & Molecular Biology 79 (2001) 49–59
Bone phenotype of the aromatase deficient mouse�
Orhan K. Öza,∗, Gen Hirasawaa, Jonathan Lawsona, Lydia Nanua, Anca Constantinescua,Peter P. Anticha, Ralph P. Masona, Edward Tsyganova, Robert W. Parkeya,
Joseph E. Zerwekhb, Evan R. Simpsonca Department of Radiology, UT Southwestern Medical Center at Dallas, Dallas, TX 75390, USA
b Department of Internal Medicine, UT Southwestern Medical Center at Dallas, Dallas, TX 75390, USAc Victorian Breast Cancer Consortium, Prince Henry’s Institute of Medical Research, P.O. Box 5152, Clayton, Vic., Australia
Abstract
Estrogens are important for normal bone growth and metabolism. The mechanisms are incompletely understood. Thus, we have un-dertaken characterization of the skeletal phenotype of aromatase (ArKO) deficient mice. No abnormalities have been noted in skeletalpatterning in newborns. Adult ArKO mice show decreased femur length and decreased peak Bone Mineral Density (BMD) with acceler-ated bone loss by 7 months of age in females. Magnetic resonance microscopy (MR) and microCT (�CT) imaging disclosed decreasedcancellous connectivity and reduced cancellous bone volume in ArKO females. Bone formation rate (BFR) is increased in ArKO femalesand decreased in ArKO males. Estradiol therapy reverses these changes. This anabolic effect of estradiol in the male skeleton is supportedby 18-F− Positron Emission Tomography (PET) imaging, which clearly demonstrates decreased spinal uptake, but marked increase afterestradiol therapy. Serum IGF-1 levels are high in young female ArKO mice but low in young ArKO males. The reduced BMD in ArKOfemales, despite the presence of elevated serum IGF 1, suggests that other mechanism(s) are operative. There is increased B-cell lym-phopoiesis in adult female ArKO bone marrow cells. These results show that ArKO mice show the effects of estrogen deficiency on bonegrowth, mass, metabolism, microarchitecture and the hematopoietic microenvironment. © 2002 Elsevier Science Ltd. All rights reserved.
Keywords: Aromatase; Bone; Dual energy X-ray absorptiometry; Estrogen; Histomorphometry; Insulin-like growth factor 1; Major urinary protein
1. Introduction
Estrogens are known to be important in the establishmentand maintenance of normal bone mass in women and men.In particular, loss of estrogen at the menopause leads to ac-celerated bone loss in women [1] and is also contributory tothe slow bone loss seen with aging in men [1,2]. Though themechanisms of estrogens’ actions in maintenance of bone areincompletely understood, it is clear that they act ultimatelyby controlling the balance between resorption or osteoclasticactivity and bone formation or osteoblastic activity. Whilemost research has focused on estrogens’ effects on osteoclas-togenesis, recent evidence suggests that estrogen also stim-ulates osteoblast activity [3]. Cytokines and growth factorsare known mediators of these effects on bone [4]. Moreoverestrogens are necessary for acquisition of peak Bone Min-eral Density (BMD) in the maturing skeletons of both sexes(reviewed in [5]). In terms of the accrual of bone mass to
� Proceedings of the Symposium: ‘Aromatase 2000 and the ThirdGeneration’ (Port Douglas, Australia, 3–7 November 2000).
∗ Corresponding author. Fax:+1-214-648-2727.E-mail address: [email protected] (O.K. Öz).
peak BMD, the GH–IGF-1 axis is known to be importantand in at least one case of estrogen deficiency, Turner’s syn-drome, there are abnormalities in GH secretion. This raisesthe possibility of a causal relationship.
Estrogens also regulate the hematopoietic componentof bone. Ovariectomy, though not a pure estrogen defi-cient state, results in increased B-cell lymphopoiesis in themouse bone marrow. This and the associated bone loss canbe prevented by estrogen administration [6]. On the otherhand, the osteosclerosis induced by high doses of estrogen[7] is preceded by suppression of hematopoiesis includingB-cell lymphopoiesis [8]. The osteogenic precursor popu-lation increases, but after the effects on the hematopoieticcomponent. Thus, estrogens’ anti-resorptive and poten-tial pro-formation effects may involve regulation of thehematopoietic component of the marrow.
Estrogens are synthesized from androgen precursors bythe enzyme aromatase. In order to unravel the mechanismsunderlying the multiple effects of estrogens animal mod-els are necessary. The aromatase deficient mouse or ArKOmouse [9] is such a model. We have previously reported thatArKO have reduced cancellous bone volume and a sexuallydimorphic pattern of remodeling, high turnover in females
0960-0760/02/$ – see front matter © 2002 Elsevier Science Ltd. All rights reserved.PII: S0960-0760(01)00130-3

50 O.K. Öz et al. / Journal of Steroid Biochemistry & Molecular Biology 79 (2001) 49–59
and low turnover in males [10]. We also reported male micehave significantly reduced femur length. Since that origi-nal study we have continued our analysis of the bone phe-notype by studying longitudinal growth dynamically, bonemineral accretion, remodeling, and B-cell lymphopoiesis inadditional animals. The results are reported below.
2. Materials and methods
2.1. Animals
ArKO mice and their wt littermates were produced bynonbrother-sister breeding of F6 heterozygous animals. Allanimals were from the hybrid 129S6/SvEv Taconic/C57Bl/6background. Animals had unlimited access to drinkingwater and were maintained on standard chow. For estradiolreplacement experiments, animals were injected subcuta-neously with 10�g/kg of estradiol in corn oil three timesper week. All protocols were approved by the IACRAC ofUT Southwestern Medical Center.
2.2. Skeletal staining
Skin and viscera were removed from newborn mice. Thecarcasses were fixed overnight in 95% ethanol. The fixedskeletons were stained for 24 h in a 0.3% w/v alcian bluesolution prepared in 70% ethanol. After clearing the resid-ual stain by soaking in 95% EtOH for several hours, theskeletons were transferred to 2% KOH for 24 h or longerdepending upon the size of the animal to clear any re-maining flesh. The cleared skeletons were transferred to 1%KOH/0.05 Alizarin red S for overnight staining. The skele-tons were then cleared for a least 2 days in 1% KOH/20%glycerol. Stained cleared skeletons were stored in glycerol:95% EtOH (1:1).
2.3. Static and dynamic histomorphometry
Static and dynamic histomorphometry were performed aspreviously described [10]. Bone Formation Rate (BFR) wascalculated as the product of the mineral apposition rate timesthe fraction of osteoid surface covered by a double label. Itrepresents the volume of mineralized new bone formed perunit area of osteoid surface per unit time.
2.4. Flow cytometry
Bone marrow cells were prepared from lumbar spine andfemurs of 8-month-old wt and ArKO females by crushingthe bones in PBS containing 2% fetal bovine serum (FBS)and 1 mM EDTA. A single cell suspension was preparedby first filtering the resultant supernatant through a nylonmesh followed by repeated dispersion through a 22-gaugeneedle. The cells (2.5× 105) were then incubated with
FITC-conjugated anti-B220(CD45R) (Pharmingen, SanDiego, CA USA) for 15 min on ice, washed once, and fixedfor 10 min with formalin. Stained cells were analyzed on aBecton Dickinson FACS system. Unstained cells were usedas negative controls.
2.5. DEXA
Whole body DEXA was performed using the PIXImusdensitometer (Lunar Corporation, Madison, WI, USA)equipped with software version 1.43.
2.6. Urine and serum analysis
Urine was collected from mice as a spontaneous voidgenerated at the time animals were handled between 1500 hand 1730 h. The urine was diluted 1:8 and analyzed bySDS-PAGE. For plasma IGF-1 analysis, blood was col-lected from the retro-orbital venous plexus into heparinizedpipettes. Blood was always collected at the same time eachday, about 1530. Plasma was separated from clotted bloodby centrifugation. IGF-1 concentration was determined byRIA (DSL, Inc. Webster, TX, USA).
2.7. Magnetic resonance and microcomputedtomography imaging
MR imaging was performed using an Omega 9.4T MRIsystem (GE/Bruker, Fremont, CA) and a home built slottedtube resonator coil (10 mm i.d.). Shimming on the water sig-nal gave a typical linewidth of 70 Hz. Data were acquiredusing a simple partial saturation 3D spin-echo sequencewith hard pulses (π /2 = 55 �s), a repetition time (TR) of150 ms, an echo time (TE) of 16 ms and 64× 64 phaseencode steps over the small dimensions. One hundred andtwenty eight read-out points for the long axis with a typicalFOV 4× 4× 16 mm gave a 62.5× 62.5× 125 �m resolu-tion. Sine-bell apodization was applied prior to 3D Fouriertransformation. Images were interpolated (up to 8 fold) andedited using NIH Image shareware (NIMH, Bethesda, MD)and analyzed usingc language software developed in house[11]. Connectivity density was determined for regions ofthe metaphysis expected to be rich in trabecular structureusing algorithms described previously [11]. The vertebralbodies were scanned on a�CT 20 system (Scanco USA,Inc. Wayne, PA) at a resolution of 18�m3. A set of imageswas obtained from each sample. Three-dimensional analy-sis was conducted on manually selected volume of interestto calculate trabecular bone morphometric parameters.
2.8. Positron emission tomography imaging
Following intravenous administration of 500�Ci of car-rier free 18-F− , 1 h delayed whole body images were ac-quired on an in-house designed PET imaging system [12].

O.K. Öz et al. / Journal of Steroid Biochemistry & Molecular Biology 79 (2001) 49–59 51
Following reconstruction, the images were displayed astwo-dimensional projections.
2.9. Statistical analysis
Data are expressed as the mean± S.D. or standard error(S.E.). In all instancesP<0.05 was taken to be significant.For normally distributed data, analysis was done using thet-test. Otherwise, the Mann–Whitney test was utilized.
3. Results
3.1. Bone patterning
To determine whether aromatase deficiency results in de-velopmental abnormalities in skeletal patterning, we haveexamined the newborn skeleton after staining for bone andmineralized cartilage (Fig. 1). After careful study of three
Fig. 1. Alizarin red and Alcian blue staining of newborn skeletons. A, Wt female; B, ArKO female; C, Wt male; D, ArKO male. Skeletons were clearedand stained as described in Section 2. In Panel A one femur of the newborn has been disarticulated during processing and the offset of the bones ofposterior skull in the males is a processing artifact. No gross abnormalities, such as missing bones and large areas of undermineralization are present.
litters we have not found any consistent differences betweenArKO animals and their wt littermates. In this particular ex-ample, the cartilaginous portions of the wild-type femaleskull show more mineralization (blue) than the ArKO fe-male. On the other hand, it appears that these areas are moremineralized in the ArKO male relative to wildtype control.The ArKO male also shows greater mineralization of thetibial epiphysis compared with its wt littermate. However,in other litters we have seen reversal of these relationshipswith no consistent trend from litter to litter. In the currentexample, the ArKO male demonstrates shortened femurs.Again, this pattern was not consistent in the three litters wehave examined to date.
3.2. Longitudinal bone growth
Previously, we have shown that adult male ArKO micehave decreased mean femur length as compared with wt lit-termates. Study of the dynamics of femur growth show this

52 O.K. Öz et al. / Journal of Steroid Biochemistry & Molecular Biology 79 (2001) 49–59
Fig. 2. Longitudinal growth of the femur with age. In a cross-sectional study design femur length was followed from the time of weaning to 7 months.There is significant difference in femur length between Wt and ArKO females only at the 46–65 day range. Note male ArKO mice show an absence ofthe accelerated growth during puberty as compared with Wt. For time intervals after 46–65, the difference is statistically significant.
to be a consequence of the absence of rapid growth seen inmale wt mice during the postpubertal period (Fig. 2). Onthe other hand, the female ArKO animals demonstrate sig-nificantly longer femurs (ArKO 14.8 mm vs. wt 14.5,P =0.04) at the 46–65 day range (Fig. 2), though at the 6–7month range the relationship between the means was re-versed (wt 16.4 vs. 16.1 mm ArKO) and nearly significantlydifferent (P = 0.06). This contrasts with the accumulation
of body weight and nasal–anal length which is significantlyincreased by 5 weeks of age in ArKO females as comparedwith wt littermates (Hirasawa and Öz, unpublished data)but not in males. We have previously reported this sexu-ally dimorphic pattern in body size of 3–4-month-old ArKOanimals [13].
IGF-1 has been shown to be an important regulator ofbody mass and bone growth. Therefore, we investigated

O.K. Öz et al. / Journal of Steroid Biochemistry & Molecular Biology 79 (2001) 49–59 53
Table 1Serum IGF-1 levels
Age IGF-1 (ng/ml)
Males Females
Wt ArKO Wt ArKO
1–2 months 675± 132.8 (n = 4) 474.4± 160.1 (n = 5) 526.7± 90.7 (n = 3) 733± 79.6 (n = 4)
ArKO significantly different from Wt,P<0.05.
IGF-1 serum levels in ArKO and wt type mice. Serum IGF-1levels in young male ArKO mice are significantly lowerthan controls. On the other hand, ArKO females have sig-nificantly higher levels than wt female controls (Table 1).This correlates well with body weight results. Serum IGF-1is mainly produced by the liver and that production is regu-lated by growth hormone [14]. Since growth hormone levelsare difficult to measure because of rhythmic variations or
Fig. 3. Major Urinary Protein levels in ArKO and Wt littermates. Equal volumes of diluted urine were analyzed by SDS-PAGE under reducing conditions.Lanes A–G, male animals. Lane H–M female animals. The animals are grouped by litters. Notice that ArKO males have lower levels of MUP relativeto littermates, whereas female ArKO animals have higher levels than Wt littermates.
pulsatility we measured levels of the major urinary protein(MUP) as a surrogate [15]. Consistent with the low IGF-1levels ArKO males show decreased MUP in their urine andArKO females, which have higher serum IGF-1, show in-creased MUP levels relative to their controls (Fig. 3). Notethere is variability in MUP levels between ArKO males ofdifferent litters but ArKO males show lower levels than wtmales of the same litter.

54 O.K. Öz et al. / Journal of Steroid Biochemistry & Molecular Biology 79 (2001) 49–59
Table 2Lumbar spine BMD (mg/cm2) of female mice as a function of age
Age Wt ArKO P
Genotype3 weeks 28.8± 2.4 30.6± 2.2 NS2 months 53.6± 2.9 51.5± 3.5 NS3 months 61.3± 5.4 51.0± 2.9 <0.014 months 66.8± 5.4 52.1± 2.9 <0.017 months 66.4± 4.3 47.6± 4.4 <0.0110 months 65.6± 5.5 46.2± 5.8 <0.0112 months 67.2± 5.5 45.9± 5.3 <0.01
Table 3Lumbar spine BMD of male mice as a function of age
Age Wt ArKO P
Genotype3 weeks 29.1± 1.6 27.3± 1.4 <0.052 months 49.2± 3.4 47.8± 4.1 NS3 months 55.2± 4.1 53.1± 3.8 <0.014 months 58.5± 5.1 54.3± 4.4 <0.017 months 58.2± 5.1 53.4± 5.3 <0.0112 months 58.2± 3.8 53.6± 5.3 <0.01
n≥4 in each group.
3.3. Bone mass accretion and change with aging orestradiol therapy
Estrogens are now known to influence bone mass accrualnot only in human females but males as well. Therefore,we have used DEXA to examine the acquisition of peakBMD and maintenance with age. Irrespective of sex, as apopulation, ArKO animals have decreased peak BMD com-pared with wt controls (Tables 2 and 3). Furthermore, asthe animals become older they show an accelerated loss ofspinal BMD, females much more dramatically than males.The ArKO skeleton responds appropriately to estradiol as1 month of estradiol therapy increased BMC in male andfemale ArKO animals relative to placebo controls (Table 4).
3.4. Bone remodeling
We have previously reported sexual dimorphism in theremodeling patterns of 5–7-month-old ArKO animals [10].These results are summarized in Table 5. Adult males showa low turnover pattern while adult females show a highturnover pattern. This is confirmed by tetracycline doublelabel patterns in which ArKO females show increasedtetracycline labeled surfaces (both single and double) as
Table 4Effect of short term estradiol therapy on ex-vivo spinal BMC
ArKO males ArKO females
Baseline (g/cm× 10− 3) 20.3 (n = 3) 17 (n = 3)Estradiol therapy 21.6 (n = 3) 20.8 (n = 3)Change (%) 6 18
Table 5Summary of static and dynamic histomorphometry analysis of the lumbarspine
Compared with wt littermate
ArKO female ArKO male
Structural parameters
BV/TV (%)
C.Th (�m) –
Tb.Th (�m)
Surface parametersFormationOV/BV (%) ↑ –
OS/BS (%) ↑Ob.S/BS (%) –
Resorption
Oc.S/BS (%) –
ES/BS (%) –
Dynamic parameters
MS/BS′ (%) ↑MS/BS (2X%)
Modified from [10]. Bold arrows indicate significant differences. Thinarrows indicate direction of trends though the differences were not sig-nificant. Horizontal lines indicate there is no trend to suggest a change.
compared with wt female or male animals and ArKO males.ArKO males show the lowest double labeled surface ofany of the animal groups (Fig. 4). This is shown by dra-matic elevations of BFR in ArKO females and decreasedBFR in ArKO males (Table 6). Estradiol therapy, as inovariectomized mice, decreases BFR in ArKO females.However, the same therapeutic regimen stimulated BFR inArKO males (Table 6) as well as other histomorphometricmeasures of bone remodeling (data not shown).
Fluorine-18 positron emission tomography (PET) is animaging method in which F-18 is retained in the skeleton inproportion to the degree of remodeling that is occurring at
Table 6Bone formation rates
Genotype BFR (�m3/mm2 per year)
Female Male
Mean± S.E.M. P Mean± S.E.M. P
WT 374.5± 58.4 – 384.5± 6.8 –ArKO 401.9 ± 45.3 NSa 81.3 ± 30.4a <0.0001ArKO+E2 130.3± 22.4 <0.01b 192 ± 45.9b 0.006
NS, not significant.aArKO vs. WT.bArKO + E2 vs. ArKO.

O.K. Öz et al. / Journal of Steroid Biochemistry & Molecular Biology 79 (2001) 49–59 55
Fig. 4. Double labeled surfaces in spinal cancellous bone of Wt and ArKO animals. Panel A, Wt female. Panel B, ArKO female. Panel C, Wt male.Panel D, ArKO male. Notice that female mice have more label incorporation compared with males and that there are more double labeled surfaces inArKO females compared with other groups.
the time of imaging [16]. Therefore, we tested whether maleArKO animals would show decreased F-18 retention com-pared with wt males and whether estradiol therapy wouldcorrect this decrease. Fig. 5 shows decreased activity in maleArKO lumbar spine (yellow vs. red) but a more normal pat-tern (more red) in the lumbar spine after therapy.
3.5. Bone hematopoietic microenvironment
Ovariectomy has been reported to increase B-cell lym-phopoiesis in bone marrow cells. Therefore, we comparedthe bone marrow derived from wt and ArKO mice for cellsstaining positive for the B-cell marker B220. A representa-tive experiment is shown in Fig. 6. The ArKO female clearlyshows higher levels of the two populations of B220 positivecells. This same pattern has been seen in three independentexperiments.
3.6. Bone 3D microarchitecture
To demonstrate whether there is an alteration in the3D structure of cancellous bone we have employed MRmicroscopy and�CT on femurs and vertebral bodies,
respectively, of female wt and ArKO littermates at 4 monthsof age. As expected the ArKO animals have decreased bonevolume in the spine (Fig. 7A and B) and femur (Fig. 7Cand D). Furthermore, the connectivity is also decreased.For example, in the vertebral bodies examined by�CT themean connectivity density is 115.5 mm− 3 for Wt femaleand 44.4 mm− 3 for the ArKO female. Femurs examinedby MR microscopy show similar decreased connectivitydensity, 119 vs. 44.
4. Discussion and conclusion
Bone is a multi-compartment dynamic organ composedof mineralized tissue, stromal, and hematopoietic marrowcompartments. The present report demonstrates that estrogeninsufficiency caused by aromatase deficiency in the ArKOmice results in abnormalities in bone growth, mass, remodel-ing, microarchitecture and the hematopoietic microenviron-ment. It extends our previous report [10] by showing whenthe abnormalities in longitudinal growth arise. Furthermore,the abnormalities are sexually dimorphic in that in the earlypost-pubertal period the ArKO females have longer bones
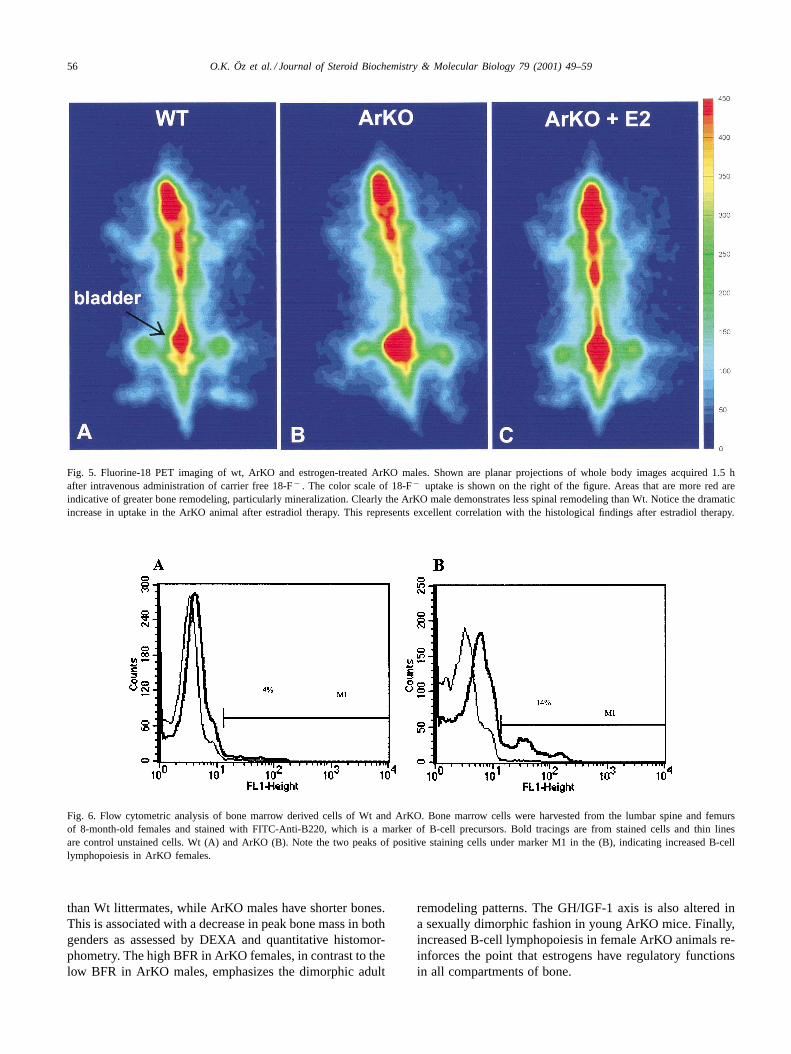
56 O.K. Öz et al. / Journal of Steroid Biochemistry & Molecular Biology 79 (2001) 49–59
Fig. 5. Fluorine-18 PET imaging of wt, ArKO and estrogen-treated ArKO males. Shown are planar projections of whole body images acquired 1.5 hafter intravenous administration of carrier free 18-F− . The color scale of 18-F− uptake is shown on the right of the figure. Areas that are more red areindicative of greater bone remodeling, particularly mineralization. Clearly the ArKO male demonstrates less spinal remodeling than Wt. Notice thedramaticincrease in uptake in the ArKO animal after estradiol therapy. This represents excellent correlation with the histological findings after estradioltherapy.
Fig. 6. Flow cytometric analysis of bone marrow derived cells of Wt and ArKO. Bone marrow cells were harvested from the lumbar spine and femursof 8-month-old females and stained with FITC-Anti-B220, which is a marker of B-cell precursors. Bold tracings are from stained cells and thin linesare control unstained cells. Wt (A) and ArKO (B). Note the two peaks of positive staining cells under marker M1 in the (B), indicating increased B-celllymphopoiesis in ArKO females.
than Wt littermates, while ArKO males have shorter bones.This is associated with a decrease in peak bone mass in bothgenders as assessed by DEXA and quantitative histomor-phometry. The high BFR in ArKO females, in contrast to thelow BFR in ArKO males, emphasizes the dimorphic adult
remodeling patterns. The GH/IGF-1 axis is also altered ina sexually dimorphic fashion in young ArKO mice. Finally,increased B-cell lymphopoiesis in female ArKO animals re-inforces the point that estrogens have regulatory functionsin all compartments of bone.

O.K. Öz et al. / Journal of Steroid Biochemistry & Molecular Biology 79 (2001) 49–59 57
Fig. 7. MR microscopy and�CT imaging of femurs or vertebrae from Wt and ArKO females.�CT 3D renderings (A, B). MC microscopy (C, D). A,C Wt female. B, D ArKO female. In C and D the gray scale has been inverted so that bone appears white and a part of the cortical bone has been ‘cutaway’ to reveal the difference in trabecular structure. Notice the decreased trabecular bone in the femoral metaphysis of the ArKO animal.
The longitudinal bone phenotype in ArKO mice is ageand sex dependent. By 7 months of age the ArKO femalefemurs are nearly significantly shorter (P = 0.06) while fe-murs from ArKO males show much more significant short-ening (P = 0.0001). Both sexes of ArKO animals experi-ence a reduced longitudinal femur growth rate after the earlypost-pubertal period (i.e. 46–65 days old). From this time to7 months of age the mean femur length increased by 12.4%in wt females versus 9% in ArKO females; however, the maleArKO mice show a more dramatic difference with only a2% increase compared with a 10.1% increase in males. Thelimiting step in longitudinal growth rate regulation withinlong bones occurs at the level of the growth plate. There-fore, these changes may be secondary to a problem(s) in therequisite osteoconduction by growth plate septae. These sex-ually dimorphic differences in longitudinal bone growth ob-served in the early post-pubertal ArKO animals could resultfrom a number of paracrine/autocrine and/or endocrine ab-normalities, including GH acting through IGF-1, Vitamin D,and thyroid hormone. For example, the altered serum IGF-1levels observed in ArKO mice during puberty (1 month),low in males but high in females, appear to be consistent
with the observed decreased and increased bone lengths, re-spectively. However, recent data suggest autocrine/paracrineeffects of IGF-1 in the local skeletal environment regulategrowth [17] as opposed to an endocrine mode of regulationby hepatic IGF-1 [14]. It remains to be tested whether theremight be local changes of IGF-1 in ArKO bone. Several otherparacrine/autocrine factors are known to regulate growthplate chondrocyte proliferation and differentiation includ-ing TGF-�, FGF-2, PTHrP, BMPs 6 and 7, and VitaminD [18]. Of particular interest is the PTHrP/IHH regulatoryfeedback system. Originally identified as a critical feedbacksystem regulating embryonic skeletal development, this sys-tem has recently been shown to be expressed in epiphy-seal cartilage during the early post-pubertal period in mice[19], and the pubertal period in rats [20]. To our knowl-edge there are no published studies on the regulation ofthis feedback system by estrogens; however, estrogens havebeen shown to regulate PTHrP expression in breast cancercells though the results are conflicting [21,22]. Neverthe-less, these results raise the question of dysregulation of thissystem as the cause of the altered longitudinal growth inArKO mice.

58 O.K. Öz et al. / Journal of Steroid Biochemistry & Molecular Biology 79 (2001) 49–59
A defect in growth plate function might also be the basisfor the decreased peak bone mass in the lumbar spine, if tra-becular number is decreased early in life. Indeed at 3 weeksof age ArKO males have lower spinal BMDs but not ArKOfemales. However, both sexes show reduced spinal BMDat 3 months. If local IGF-1 changes are represented by thechanges in circulating levels then the differences may besecondary to changes in the GH/IGF-1 axis. The reducedMUP levels, a family of proteins produced in the liver, ofArKO males is more characteristic of the low continuousGH release seen in female mice as opposed the high MUPlevels in normal male mice which release high levels of GHin a pulsatile fashion [15]. On the other hand, the high MUPlevels in ArKO females is more characteristic of normalmale levels. Altered MUP levels in ArKO animals suggestthat estrogens or possibly the estrogen to testosterone ratioregulates the pattern of GH secretion in both sexes. What-ever the case with IGF-1, reduced peak BMD in both sexesindicates normal levels of estrogens are required to attainpeak BMD.
Maintenance of skeletal mass after establishment of peakBMD requires normal coupling of bone formation and re-sorption. While a growth plate malfunction might explainthe low peak BMD it cannot explain the abnormal patternsof turnover demonstrated by tetracycline labeling and PETimaging. The loss of bone mass in aging ArKO femalesis consistent with increased osteoclastic activity. The lowturnover pattern with little change in BMD demonstratedin ArKO males might be the result of a defect(s) in os-teoblast differentiation or function. Since osteoclastogenesisis coupled to osteoblasts through the OPG/RANK/RANKLsystem the low osteoclastic surface may also be explainedby a problem with osteoblasts/stromal cells in ArKO mice.We are currently investigating whether male ArKO animalshave fewer osteoblast progenitor cells or faulty differentia-tion in in vitro studies. The dramatic anabolic response (highBFR and increased osteoblastic surface (data not shown)) ofArKO males to estradiol indicates the problem is a conse-quence of estrogen deficiency and suggests the defect is inprogenitor differentiation. Why there is sexual dimorphismin the remodeling pattern remains unclear.
The increased B cell lymphopoiesis demonstrated inArKO females is similar to the results observed in ovariec-tomized mice [6]. However, ovariectomy does not resultin estrogen deficiency alone. Our results show unequivo-cally that estrogen deficiency is sufficient for increased Bcell lymphopoiesis in bone marrow cells. Previous studieshave shown that cell-to-cell interaction between B lympho-cytes and human bone marrow stromal cells triggers theproduction of the bone resorbing cytokines IL-1 and IL-6[23]. Thus, the increased bone loss seen in adult ArKO fe-males may be related to this increase in B cells in the bonemicroenvironment.
In summary, the bone phenotype of the ArKO mouseshows many of the phenotypes associated with estrogendeficiency, decreased peak BMD in both sexes; abnormal
longitudinal growth; alterations in bone remodeling; in-creased B-cell lymphopoiesis; and an altered GH/IGF-1axis. Understanding the underlying mechanisms shouldimpact our understanding of growth plate function and reg-ulation, bone remodeling in adults, and the health care ofpatients with osteopenia and osteoporosis.
Acknowledgements
The authors wish to thank Dr Rasesh Kapadia and DrDouglass Adams for�CT scanning and analysis, Dr MarkWessels for MR microscopy, Dr James Richardson and JohnShelton for photography of stained skeletons and VirginiaBarnett for assistance with graphics. This work was sup-ported by grants 1-K08-HDO1463-01 from the NICHD ofthe NIH, and in part by funds from the Effie and WoffardCain Chair endowment, institutional funds from the Centerfor Mineral Metabolism and Clinical Research and a grantfrom the Victorian Breast Cancer Research Consortium, Inc.
References
[1] B.L. Riggs, S. Khosla, L.J. Melton III, A unitary model forinvolutional osteoporosis: estrogen deficiency causes both type I andtype II osteoporosis in postmenopausal women and contributes tobone loss in aging men, J. Bone Miner. Res. 13 (5) (1998) 763–773.
[2] S. Khosla, L.J. Melton III, E.J. Atkinson, W.M. O′Fallon, G.G.Klee, B.L. Riggs, Relationship of serum sex steroid levels and boneturnover markers with bone mineral density in men and women: akey role for bioavailable estrogen, J. Clin. Endocrinol. Metab. 83 (7)(1998) 2266–2274.
[3] J.H. Tobias, J.E. Compston, Does estrogen stimulate osteoblastfunction in postmenopausal women? Bone 24 (2) (1999) 121–124.
[4] S.C. Manolagas, Birth and death of bone cells: basic regulatorymechanisms and implications for the pathogenesis and treatment ofosteoporosis, Endocr. Rev. 21 (2) (2000) 115–137.
[5] L.A. Soyka, W.P. Fairfield, A. Klibanski, Hormonal determinants anddisorders of peak bone mass in children, J. Clin. Endocrinol. Metab.85 (11) (2000) 3951–3963.
[6] Y. Onoe, C. Miyaura, M. Ito, H. Ohta, S. Nozawa S, T. Suda,Comparative effects of estrogen and raloxifene on B lymphopoiesisand bone loss induced by sex steroid deficiency in mice, J. BoneMiner. Res. 15 (3) (2000) 541–549.
[7] M.R. Urist, A.M. Budy, F.C. Mclean, Endosteal bone formation inestrogen-treated mice, J. Bone Joint Surg. 32A (1950) 143–162.
[8] M.J. Perry, A. Samuels, D. Bird, J. Tobias, Effects of high-doseestrogen on murine hematopoietic bone marrow precede those onosteogenesis, Am. J. Physiol. Endocrinol. Metab. 279 (2000) E1159–E1165.
[9] C.R. Fisher, K.H. Graves, A.F. Parlow, E.R. Simpson,Characterization of mice deficient in aromatase (ArKO) because oftargeted disruption of the cyp19 gene, Proc. Natl. Acad. Sci. USA95 (12) (1998) 6965–6970.
[10] O.K. Öz, J.E. Zerwekh, C. Fisher, K. Graves, L. Nanu, R. Millsaps,E.R. Simpson, Bone has a sexually dimorphic response to aromatasedeficiency, J. Bone Miner. Res. 15 (3) (2000) 507–514.
[11] M. Wessels, R.P. Mason, P.P. Antich, J.E. Zerwekh, C.Y. Pak,Connectivity in human cancellous bone by three-dimensionalmagnetic resonance imaging, Med. Phy. 24 (9) (1997) 1409–1420.

O.K. Öz et al. / Journal of Steroid Biochemistry & Molecular Biology 79 (2001) 49–59 59
[12] P. Antich, R. Parkey, N. Slavin, E. Tsyganov, A. Zinchenko, CompactCompton Camera Design: Parameters and Imaging Algorithm,Nuclear Science Symposium and Medical Imaging Conference, Lyon,France, 15–20 October 2000, Abstract 225.
[13] M.E.E. Jones, A.W. Thorburn, K.L. Britt, K.N. Hewitt, N.G. Wreford,J. Proietto, O.K. Oz, B.J. Leury, K.M. Robertson, S. Yao, E.R.Simpson, Aromatase-deficient (ArKO) mice have a phenotype ofincreased adiposity, Proc. Natl. Acad. Sci. USA 97 (23) (2000)12735–12740.
[14] W.H. Daughaday, P. Rotwein, Insulin-like growth factors I and II.Peptide, messenger ribonucleic acid and gene structures, serum, andtissue concentrations, Endocr. Rev. 10 (1) (1989) 68–91.
[15] G. Norstedt, R. Palmiter, Secretory rhythm of growth hormoneregulates sexual differentiation of mouse liver, Cell 36 (4) (1984)805–812.
[16] C. Schiepers, J. Nuyts, G. Bormans, J. Dequeker, R. Bouillon, L.Mortelmans, A. Verbruggen, M. De Roo, Fluoride kinetics of theaxial skeleton measured in vivo with fluorine-18-fluoride PET, J.Nucl. Med. 38 (12) (1997) 1970–1976.
[17] K. Sjogren, J.L. Liu, K. Blad, S. Skrtic, O. Vidal, V. Wallenius,D. LeRoith, J. Tornell, O.G. Isaksson, J.O. Jansson, C. Ohlsson,Liver-derived insulin-like growth factor I (IGF-I) is the principalsource of IGF-I in blood but is not required for postnatal body growthin mice, Proc. Natl. Acad. Sci. USA 96 (12) (1999) 7088–7092.
[18] E.B. Hunziker, Growth plate formation, structure and regulation,J.A. Buckwalter M.G. Ehrlich L.J. Sandell S.B. Trippel (Eds.),Skeletal Growth and Development Clinical Issues and Basic ScienceAdvances (1998) 187–202American Academy of OrthopaedicSurgeonsRosemont, IL.
[19] V. Kartsogiannis, J. Moseley, B. McKelvie, S.T. Chou, D.K. Hards,K.W. Ng, T.J. Martin, H. Zhou, Temporal expression of PTHrP duringendochondral bone formation in mouse and intramembranous boneformation in an in vivo rabbit model, Bone 21 (5) (1997) 385–392.
[20] B.C. van der Eerden, M. Karperien, E.F. Gevers, C.W. Lowik, J.M.Wit, Expression of Indian hedgehog, parathyroid hormone-relatedprotein, and their receptors in the postnatal growth plate of the rat:evidence for a locally acting growth restraining feedback loop afterbirth, J. Bone Miner. Res. 15 (6) (2000) 1045–1055.
[21] J.L. Funk, H. Wei, Regulation of parathyroid hormone-relatedprotein expression in MCF-7 breast carcinoma cells by estrogenand antiestrogens, Biochem. Biophys. Res. Commun. 251 (3) (1998)849–854.
[22] J. Kurebayashi, H. Sonoo, Parathyroid hormone-related proteinsecretion is inhibited by oestradiol and stimulated by antioestrogensin KPL-3C human breast cancer cells, Br. J. Cancer 75 (1997) 1912.
[23] L.J. Jarvis, T.W. LeBien, Stimulation of human bone marrowstromal cell tyrosine kinases and IL-6 production by contact with Blymphocytes, J. Immunol. 155 (5) (1995) 2359–2368.




















