
Telocytes in Human Term Placenta - Morphology and Phenotype
15
Fax +41 61 306 12 34 E-Mail [email protected] www.karger.com Original Paper Cells Tissues Organs DOI: 10.1159/000319467 Telocytes in Human Term Placenta: Morphology and Phenotype Laura Suciu a, b Laurenţiu M. Popescu a, b Mihaela Gherghiceanu b Teodor Regalia a, b Mihnea I. Nicolescu a, b Mihail E. Hinescu a, b Maria-Simonetta Faussone-Pellegrini c a Department of Cellular and Molecular Medicine, ‘Carol Davila’ University of Medicine and Pharmacy, and b ‘Victor Babeş’ National Institute of Pathology, Bucharest, Romania; c Department of Anatomy, Histology and Forensic Medicine, University of Florence, Florence, Italy 2–3 very thin, long processes (tens to hundreds of microm- eters), with an uneven calibre ( ^0.5 m thick) and typical branching pattern . The dilations of processes accommodate caveolae, endoplasmic reticulum cisternae, and mitochon- dria. These TC have close contacts with perivascular SMC in stem villi. In situ, similar cells are positive for c-kit, CD34, vimentin, caveolin-1, vascular endothelial growth factor Key Words CD117/c-kit Human placenta Interstitial Cajal-like cells Telocytes Telopodes Abstract In the last few years, a new cell type – interstitial Cajal-like cell (ICLC) – has been described in digestive and extra-diges- tive organs. The name has recently been changed to telo- cytes (TC) and their typical thin, long processes have been named telopodes (TP). To support the hypothesis that TC may also be present in human placenta and add to the infor- mation already available, we provide evidence on the ultra- structure, immunophenotype, distribution, and interactions with the surrounding stromal cells of TC in the villous core of human term placenta. We used phase-contrast microscopy, light microscopy of semithin sections, transmission electron microscopy, immunohistochemistry, and immunofluores- cence of tissue sections or cell cultures, following a pre-es- tablished diagnostic algorithm. Transmission electron mi- croscopy showed cells resembling TC, most ( 76%) having Accepted after revision: April 8, 2010 Published online: July 27, 2010 Prof. Laurenţiu M. Popescu ‘Victor Babeş’ National Institute of Pathology Splaiul Independenţei 99–101 RO–050096 Bucharest (Romania) Tel. +40 21 319 4530, Fax +40 21 312 4885, E-Mail LMP @ jcmm.org © 2010 S. Karger AG, Basel 1422–6405/10/0000–0000$26.00/0 Accessible online at: www.karger.com/cto Abbreviations used in this paper BSA bovine serum albumin DMEM Dulbecco’s modified Eagle’s medium EDTA ethylenediaminetetraacetic acid ICC interstitial cells of Cajal ICLC interstitial Cajal-like cells IF immunofluorescence IHC immunohistochemistry iNOS inducible nitric oxide synthase PBS phosphate buffered saline SMC smooth muscle cells SMM smooth muscle myosin TC telocytes TEM transmission electron microscopy TP telopodes VEGF vascular endothelial growth factor SMA -smooth muscle actin The terminology used to describe some of the cells in this article has recently been changed. Please see the important note at the end of the paper.
-
Upload
univermed-cdgm -
Category
Documents
-
view
3 -
download
0
Transcript of Telocytes in Human Term Placenta - Morphology and Phenotype

Fax +41 61 306 12 34E-Mail [email protected]
Original Paper
Cells Tissues Organs DOI: 10.1159/000319467
Telocytes in Human Term Placenta: Morphology and Phenotype
Laura Suciu a, b Laurenţiu M. Popescu a, b Mihaela Gherghiceanu b Teodor Regalia a, b Mihnea I. Nicolescu a, b Mihail E. Hinescu a, b Maria-Simonetta Faussone-Pellegrini c
a Department of Cellular and Molecular Medicine, ‘Carol Davila’ University of Medicine and Pharmacy, and b ‘Victor Babeş’ National Institute of Pathology, Bucharest , Romania; c Department of Anatomy, Histology and Forensic Medicine, University of Florence, Florence , Italy
2–3 very thin, long processes (tens to hundreds of microm-eters), with an uneven calibre ( ̂ 0.5 � m thick) and typical branching pattern . The dilations of processes accommodate caveolae, endoplasmic reticulum cisternae, and mitochon-dria. These TC have close contacts with perivascular SMCin stem villi. In situ, similar cells are positive for c-kit, CD34, vimentin, caveolin-1, vascular endothelial growth factor
Key Words
CD117/c-kit � Human placenta � Interstitial Cajal-like cells � Telocytes � Telopodes
Abstract
In the last few years, a new cell type – interstitial Cajal-like cell (ICLC) – has been described in digestive and extra-diges-tive organs. The name has recently been changed to telo-cytes (TC) and their typical thin, long processes have been named telopodes (TP). To support the hypothesis that TC may also be present in human placenta and add to the infor-mation already available, we provide evidence on the ultra-structure, immunophenotype, distribution, and interactions with the surrounding stromal cells of TC in the villous core of human term placenta. We used phase-contrast microscopy, light microscopy of semithin sections, transmission electron microscopy, immunohistochemistry, and immunofluores-cence of tissue sections or cell cultures, following a pre-es-tablished diagnostic algorithm. Transmission electron mi-croscopy showed cells resembling TC, most ( � 76%) having
Accepted after revision: April 8, 2010 Published online: July 27, 2010
Prof. Laurenţiu M. Popescu ‘Victor Babeş’ National Institute of Pathology Splaiul Independenţei 99–101 RO–050096 Bucharest (Romania) Tel. +40 21 319 4530, Fax +40 21 312 4885, E-Mail LMP @ jcmm.org
© 2010 S. Karger AG, Basel1422–6405/10/0000–0000$26.00/0
Accessible online at:www.karger.com/cto
Abbreviations used in this paper
BSA bovine serum albuminDMEM Dulbecco’s modified Eagle’s mediumEDTA ethylenediaminetetraacetic acidICC interstitial cells of Cajal ICLC interstitial Cajal-like cells IF immunofluorescence IHC immunohistochemistry iNOS inducible nitric oxide synthasePBS phosphate buffered salineSMC smooth muscle cells SMM smooth muscle myosin TC telocytesTEM transmission electron microscopy TP telopodesVEGF vascular endothelial growth factor�SMA �-smooth muscle actin
The terminology used to describe some of the cells in this article has recently been changed. Please see the important note at the end of the paper.
steimlej
CopyBAhoch

Suciu /Popescu /Gherghiceanu /Regalia /Nicolescu /Hinescu /Faussone-Pellegrini
Cells Tissues Organs 2
(VEGF), and inducible nitric oxide synathase (iNOS). The c-kit-positive cells inconsistently co-express CD34, CD44, � SMA, S100, neuron-specific enolase, and nestin. Among cells with a morphologic TC profile in cell cultures, about 13% co-ex-press c-kit, vimentin, and caveolin-1; 70% of the c-kit-positive cells co-express CD34 and 12% co-express iNOS or VEGF. In conclusion, this study confirms the presence of TC in human term placenta and provides their ultrastructural and immu-nophenotypic characterization.
Copyright © 2010 S. Karger AG, Basel
Introduction
Recently, a new cell type has been identified in the in-terstitium of several cavitary or parenchymatous organs, including the placenta [McCloskey and Gurney, 2002; Povstyan et al., 2003; Popescu et al., 2005b; Hinescu et al., 2006; Suciu et al., 2007; Pieri et al., 2008; Hutchings et al., 2009]. To name these cells, the term interstitial Cajal-like cells (ICLC) was used because they share some morpho-logical features with a well-defined cell type present in the digestive tract, the interstitial cells of Cajal [Faussone-Pel-legrini and Thuneberg, 1999; Bussolati, 2005; Sanders et al., 2006; Farrugia, 2008; Garcia-Lopez et al., 2009]. After thorough investigation, ICLC appeared to be completely different from interstitial cells of Cajal (ICC), so the name has been changed to telocytes (TC) and their processes are called telopodes (TP) (see Note Added in Proof).
In a previous short note, we hypothesized that TC may also be present in human placenta [Suciu et al., 2007], a non-innervated organ [Reilly and Russell, 1977], but de-tailed information on their morphology and immuno-phenotype was still lacking [Suciu et al., 2007].
In the present study, we took advantage of (1) available ultrastructural criteria [Faussone-Pellegrini and Thune-berg, 1999; Popescu et al., 2005a; Pieri et al., 2008]; (2) a suggested phenotypic profile for TC, both in situ and in culture, and (3) a previous experience in identifying TC in other organs [McCloskey and Gurney, 2002; Povstyan et al., 2003; Popescu et al., 2005b; Hinescu et al., 2006; Suciu et al., 2007; Hutchings et al., 2009]. To ascertain a possible role(s) for TC in the physiology of the placenta, we focused on the distribution of presumed TC in the vil-lous core and on their interaction with surrounding stro-mal cells and the perivascular contractile sheath [Graf et al., 1995; Farley et al., 2004], a structure involved in fetal blood flow regulation.
Current opinion is that connective tissue in the hu-man placenta consists of a heterogeneous population of
cells – identified based on differential expression of cyto-skeletal proteins and some ultrastructural characteris-tics – that include fibroblasts, myofibroblasts, smooth muscle cells, Hofbauer, mesenchymal, vacuolated, fila-mented, and reticulum cells [Castellucci and Kaufmann, 1982; Martinoli et al., 1984; Kohnen et al., 1996; Demir et al., 1997; Benirschke et al., 2006; Sati et al., 2007]. A dif-ferentiation gradient starting with mesenchymal cells and ending with highly differentiated myofibroblasts or even smooth muscle cells (SMC) was suggested [Kohnen et al., 1996; Benirschke et al., 2006]. However, there is a stromal cell population that is not yet characterized.
Here we show that TC is one of the morphological and immmunohistochemical profiles that populate the ‘gray zone’ of the placenta stroma.
Materials and Methods
Fifteen human placentas were obtained from women aged 23–30 years following informed consent. The pregnancies were un-complicated (no pre-eclampsia, intrauterine growth restriction or pre-labor rupture of the membranes), and the babies were deliv-ered by elective pre-labor caesarean section at 38–40 weeks’ ges-tational age.
Cell Cultures from Human Placental Connective Tissue Tissue samples from 10 term placentas were processed to obtain
cell cultures from chorionic villi; one cotyledon was removed and washed in PBS, then chorionic villi were dissected under a stereo-microscope and minced into small pieces of about 1 mm 3 . Tissue fragments were incubated for 60 min on a rocking table in 0.05% trypsin/0.02% EDTA (Biochrom, Berlin, Germany) at 37 ° C then collected by gravity sedimentation. The supernatant, which con-tained trophoblast cells and contaminating decidua, was discard-ed. The remaining villous core was treated with 1 mg/ml collage-nase IV (Sigma Chemical, St. Louis, Mo., USA) for 30 min at 37 ° C. The supernatant was discarded and the remaining tissue frag-ments were dissociated by repeated pipetting. The resulting single-cell suspension and small cell aggregates were plated in 25 cm 2 NUNC Flasks in DMEM (Biochrom) supplemented with 10% fetal calf serum and 100 U/ml penicillin/100 � g/ml streptomycin (both from Sigma) at 37 ° C in a humidified atmosphere with 5% CO 2 . After 5 days, colonies of fibroblast-like cells began to form around the small explants and in 2 weeks they covered more than 80% of the culture dish. When used in immunofluorescence (IF) experi-ments, the cells were detached and replated on glass coverslips.
Vital Staining of Cultured Cells Cells grown on cover slips were labeled with MitoTracker
Green FM (Molecular Probes, Eugene, Oreg., USA). Cells were incubated with 80 n M MitoTracker Green FM in phenol red-free DMEM supplemented with 10% fetal calf serum and 1% antibi-otic/antimycotic solution (Sigma) for 30 min at 37 ° C. Cells were washed and examined by fluorescence microscopy (450–490 nm excitation light, 520 nm barrier filter; Nikon TE300 microscope).
Telocytes in Human Term Placenta Cells Tissues Organs 3
Immunofluorescence of Cultured Cells Immunofluorescent labeling (single or double) was performed
on cells grown on cover slips. Samples were fixed in 2% parafor-maldehyde for 10 min, washed in PBS, then incubated in PBS con-taining 2% BSA for another 10 min. Afterwards, the cells were permeabilized with 0.075% saponin in the BSA solution for 10 min (all reagents were from Sigma). Incubation with the primary antibodies was performed at room temperature for 1 h using the anti-human antibodies listed in table 1 , at the specificities and working dilutions shown.
After three serial rinses, the primary antibodies were detected with secondary anti-mouse antibodies (of the appropriate iso-type), or anti-rabbit antibody conjugated to AlexaFluor 546 or 488, all from Molecular Probes. Finally, the nuclei were counter-stained with 1 � g/ml Hoechst 33342 (Sigma).
Samples were examined under a Nikon TE300 microscope equipped with a Nikon DX1 camera, Nikon PlanApo 40 ! and 60 ! objectives, and the appropriate fluorescence filters.
Immunolabelling of Tissue Sections Human term placenta samples were fixed in 4% formalin and
embedded in paraffin according to routine histology protocols, then tested for antigen expression by immunohistochemistry (IHC) or colocalization by IF, using the anti-human antibodies listed in table 1 .
The 4- � m thick sections were deparaffinized in xylene, hy-drated in an alcohol series, and washed in distilled water. Antigen retrieval was performed enzymatically for cytokeratin 7 and
smooth muscle myosin (SMM; 15 min in 0.5% trypsin/1% CaCl 2 , at 37 ° C) and pressure cooking in TRIS-citrate buffer for all other antibodies. The sections were set aside to reach room tempera-ture, washed in PBS, and incubated in 2% BSA for 20 min.
For IHC, the incubation with primary antibody was per-formed at room temperature for 1 h, followed by standard strep-tavidin-biotin labeling and development in 3,3 � -diaminobenzidine. The sections were then counterstained with Mayer’s haematoxy-lin and photographs were taken with a Nikon Eclipse 600 micro-scope.
For double-labeling experiments, the sections were treated with fluorescent-tagged secondary antibodies after incubation with primary antibodies. Nuclei were counterstained with 1 � g/ml Hoechst 33342 (Sigma). Sections were viewed under a Nikon TE300 microscope equipped with suitable fluorescence filters.
Negative and Positive Controls For both IF and IHC, negative controls were obtained follow-
ing the same protocols, but with the primary antibodies omitted. The sections were incubated either with PBS, the appropriate mouse IgG isotype controls (IgG 1 clone DAK-GO1, IgG 2a clone DAK-GO5, or IgG 2b clone DAK-GO9, all from Dako) or negative control rabbit immunoglobulin fraction (normal) (Dako), de-pending on the primary antibody used and at the same final dilu-tion. Human ileum and colon were chosen as IHC/IF-positive controls for archetypal ICC-specific markers [Faussone-Pellegri-ni and Thuneberg, 1999]. Human samples, that contained no per-sonal identifiers to link the sample with the person from whom it
Table 1. I mmunophenotype synopsis
Antibody Source Cataloguenumber
Clone Dilution TC positivityin cell culture
TC positivity in tissue sections
CD117/c-kit1 Dako A4502 polyclonal 1:100 ++CD117/c-kit1 Santa Cruz Sc-13508 Ab81 1:100 ++CD341 Dako M7165 QBEnd-10 1:25 +/–CD341 BD Biosciences 550760 563 1:50 +Vimentin Dako M7020 V9 1:100 ++ ++Caveolin-1 Santa Cruz Sc-894 polyclonal 1:50 ++ ++CD44 Dako M7082 DF 1485 1:25 + +CD45 BD Biosciences 347460 2D1 1:100 – NTCD90 BD Biosciences 555593 5E10 1:100 – NTCytokeratin 7 Santa Cruz Sc-52322 OVTL 12-30 1:10 – –Desmin Dako M0760 D33 1:50 – +/–Nestin Santa Cruz Sc-23927 10c2 1:200 + +/–NOS2 Santa Cruz Sc-651 polyclonal 1:50 + +/–NSE Dako M0873 BBS/NC/VI-H14 1:75 + +/–S-100 Dako Z 0311 polyclonal 1:100 + +SMM Dako M3558 SMMS-1 1:100 – –VEGF Santa Cruz Sc-152 polyclonal 1:100 + +�SMA NeoMarkers MS-113-P1 Ab1 1:100 + +
T he intensity of TC reactivity was assessed semiquantitatively: – negative; +/– occasionally/weakly positive; + positive; ++ strong-ly positive; NT = not tested.
1 Different antibodies against the same antigen were used for different applications (IF/IHC).

Suciu /Popescu /Gherghiceanu /Regalia /Nicolescu /Hinescu /Faussone-Pellegrini
Cells Tissues Organs 4
came, were obtained from the collection of paraffin-embedded archived tissue samples of Victor Babeş Institute of Pathology.
Transmission Electron Microscopy Small tissue samples from 6 human term placentas were pro-
cessed and Epon embedded for transmission electron microscopy (TEM) as previously described [Hinescu et al., 2006].
Semi-thin sections ( ̂ 1 � m thick) were stained with toluidine blue and examined by light microscopy (Nikon Eclipse E600). Different investigators, who were blinded to the experimental de-sign, counted the number of cells with suggestive TC morphology within randomly selected images (n = 40) of semi-thin sections to determine the relative proportion of TC in the stem villi of differ-ent diameters.
Digital electron micrographs of 60-nm ultrathin sections were taken with MegaView III CCD using iTEM-SIS software (Olym-pus, Soft Imaging System GmbH, Germany) on a Morgagni 286 TEM (FEI Company, Eindhoven, The Netherlands) at 60 kV.
Quantitative Analysis Organelle relative volumes, in cells with ultrastructural fea-
tures of TC, were obtained using a point-counting morphometric approach [Weibel, 1979], on 100 randomly chosen TC electronmi-crographs. Low magnifications (9,100 ! ) were used for eval uation of the cell, extracellular space and mitochondria and higher mag-nifications (38,000 ! ) for caveolae, and endoplasmic reticulum. The volume fractions of the cell organelles were calculated after subtraction of the extracellular space [Weibel, 1979].
This study was approved by the Bioethics Committee of the ‘Victor Babeş’ National Institute of Pathology, Bucharest, and re-spects the principles governing clinical research as set out in the Declaration of Helsinki (1989) and Good Clinical Practice guide-lines.
Results
Cell Culture In primary cultures, cells resembling TC were success-
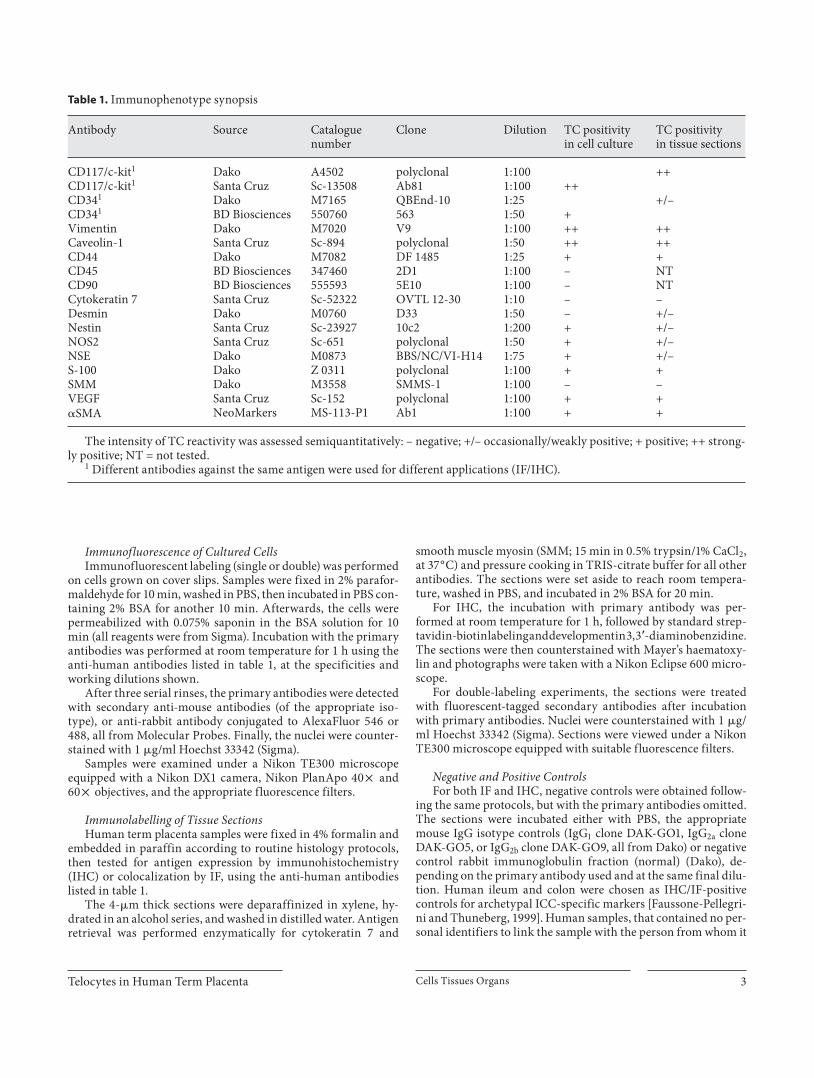
fully maintained and could be easily distinguished from SMC by phase-contrast microscopy before the culture reached confluence. After 4 days in culture, these cells displayed a typical TC morphology [Popescu et al., 2005a], with long, thin, moniliform processes extending from the small cell body ( fig. 1 a). Based on the morphology, an es-timate of the in vitro stromal cell population (n = 500) showed 13.75 8 1.1% TC, 25 8 1.5% SMC, and 51.25 8 2.7% fibroblast-like cells. The rest of the cell popula-tion was represented by a small population of epithelial-like cells that persisted even after multiple passages.
Vital staining using MitoTracker Green FM, a lipo-philic selective dye that becomes fluorescent once it ac-cumulates in the lipid environment of mitochondria, proved that these organelles were also present at the dila-tion process level, as shown in figure 1 b.
IF Staining of Cell Cultures IF for vimentin, cytokeratin 7 and CD45 antigens
along with a morphologic assessment confirmed that contamination with cytokeratin-positive trophoblast cells and CD45-positive blood cells was negligible, and more than 90% of cells were of mesenchymal origin (in-cluding endothelial cells and macrophages). Cells with suggestive TC morphology were c-kit positive. In order to analyze TC survival and the possibility of maintaining them in cell culture, we determined the proportion rep-resented by this population in different passages in 6 sep-arate cultures. The results correlate with the ‘morpho-logical assessment’ showing that they represent 12.66 8 1.2% of cells in primary culture and stabilize at around 15% after 5 passages.
Seventy percent of the presumptive TC were also pos-itive for CD34 ( fig. 1 d) and all expressed vimentin and caveolin-1 ( fig. 1 c, e). About 20% of the vimentin-positive cell population had a TC morphology. Conversely, these cells were negative for SMM, desmin and CD90 ( fig. 1 c). About 12% of cultured cells were double positive for c-kit and inducible nitric oxide synthase (iNOS) or c-kit and vascular endothelial growth factor (VEGF), and also shared the TC morphologic features.
In situ Immunophenotyping A set of antibodies was tested by IHC and IF. The most
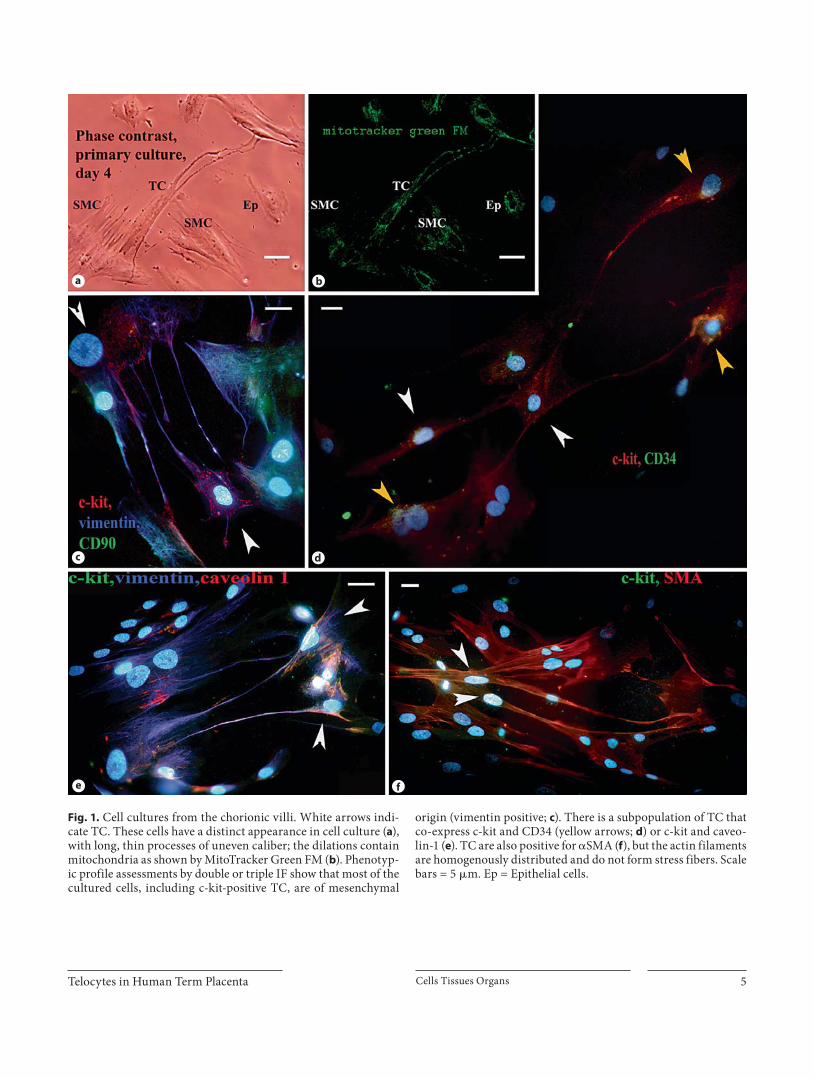
reliable TC markers were identified on paraffin-embed-ded samples by IHC. c-kit was mostly expressed in the trophoblast cells, but also in some cells with long, slender processes located just beneath the trophoblast or sur-rounding blood vessels, having either a circular or a lon-gitudinal arrangement ( fig. 2 a, b). Human ileum muscle coat served as positive control for the c-kit staining.
Vimentin reactivity was also tested because it is known to mark cells of mesenchymal origin, including TC, and all stromal cells from the villous core were indeed vimen-tin positive. The cells with suggestive TC morphology, similar to stromal c-kit-positive cells, were concentrated beneath the trophoblast and their processes ran parallel to the basement membrane and in a circular arrangement around blood vessels ( fig. 2 c). The same type of cells test-ed positive for caveolin-1, and some were positive for iNOS ( fig. 2 d).
Very few cells surrounding blood vessels in large stem villi or just beneath the trophoblast in small villi display-ing TC morphology were positive for CD34 ( fig. 2 e). En-dothelial cells were also specifically labeled by CD34 monoclonal antibodies, so we used them as positive con-trols.
Telocytes in Human Term Placenta Cells Tissues Organs 5
a b
c d
e f
Fig. 1. Cell cultures from the chorionic villi. White arrows indi-cate TC. These cells have a distinct appearance in cell culture ( a ), with long, thin processes of uneven caliber; the dilations contain mitochondria as shown by MitoTracker Green FM ( b ). Phenotyp-ic profile assessments by double or triple IF show that most of the cultured cells, including c-kit-positive TC, are of mesenchymal
origin (vimentin positive; c ). There is a subpopulation of TC that co-express c-kit and CD34 (yellow arrows; d ) or c-kit and caveo-lin-1 ( e ). TC are also positive for � SMA ( f ), but the actin filaments are homogenously distributed and do not form stress fibers. Scale bars = 5 � m. Ep = Epithelial cells.
Suciu /Popescu /Gherghiceanu /Regalia /Nicolescu /Hinescu /Faussone-Pellegrini
Cells Tissues Organs 6
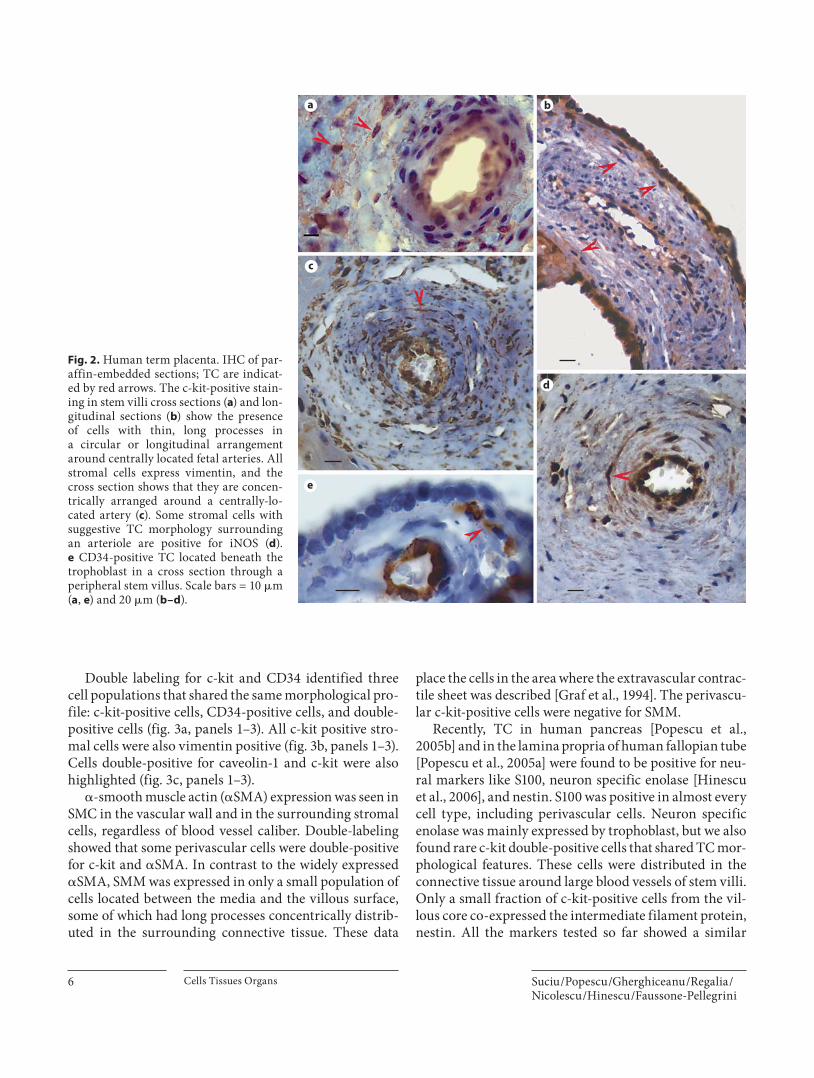
Double labeling for c-kit and CD34 identified three cell populations that shared the same morphological pro-file: c-kit-positive cells, CD34-positive cells, and double-positive cells ( fig. 3 a, panels 1–3). All c-kit positive stro-mal cells were also vimentin positive ( fig. 3 b, panels 1–3). Cells double-positive for caveolin-1 and c-kit were also highlighted ( fig. 3 c, panels 1–3).
� -smooth muscle actin ( � SMA) expression was seen in SMC in the vascular wall and in the surrounding stromal cells, regardless of blood vessel caliber. Double-labeling showed that some perivascular cells were double- positive for c-kit and � SMA. In contrast to the widely expressed � SMA, SMM was expressed in only a small population of cells located between the media and the villous surface, some of which had long processes concentrically distrib-uted in the surrounding connective tissue. These data
place the cells in the area where the extravascular contrac-tile sheet was described [Graf et al., 1994]. The perivascu-lar c-kit-positive cells were negative for SMM.
Recently, TC in human pancreas [Popescu et al., 2005b] and in the lamina propria of human fallopian tube [Popescu et al., 2005a] were found to be positive for neu-ral markers like S100, neuron specific enolase [Hinescu et al., 2006], and nestin. S100 was positive in almost every cell type, including perivascular cells. Neuron specific enolase was mainly expressed by trophoblast, but we also found rare c-kit double-positive cells that shared TC mor-phological features. These cells were distributed in the connective tissue around large blood vessels of stem villi. Only a small fraction of c-kit-positive cells from the vil-lous core co-expressed the intermediate filament protein, nestin. All the markers tested so far showed a similar
a b
c
d
e
Fig. 2. Human term placenta. IHC of par-affin-embedded sections; TC are indicat - ed by red arrows. The c-kit-positive stain-ing in stem villi cross sections ( a ) and lon-gitudinal sections ( b ) show the presenceof cells with thin, long processes ina circular or longitudinal arrangement around centrally located fetal arteries. All stromal cells express vimentin, and the cross section shows that they are concen-trically arranged around a centrally-lo-cated artery ( c ). Some stromal cells with suggestive TC morphology surroundingan arteriole are positive for iNOS ( d ). e CD34-positive TC located beneath the trophoblast in a cross section through a peripheral stem villus. Scale bars = 10 � m ( a , e ) and 20 � m ( b–d ).
Telocytes in Human Term Placenta Cells Tissues Organs 7
a1
b1 b2 b3
c1 c2 c3
d1 d2 d3
a2 a3
Fig. 3. Double IF staining on paraffin-embedded sections of human term placenta. Panels a1–d1 show perivas-cular c-kit-positive cells in stem villi. Panels a2–d2 show positive staining for CD34, vimentin, caveolin-1, and VEGF. Panels a3–d3 are merged images of the two previous slides and show double-positive cells. TC express-ing the tested markers are indicated by white arrows. Scale bars = 20 � m ( a , b , d ) and 10 � m ( c ). L = Lumen;M = media; PVS = perivascular space.
Suciu /Popescu /Gherghiceanu /Regalia /Nicolescu /Hinescu /Faussone-Pellegrini
Cells Tissues Organs 8
a b
c d
e f
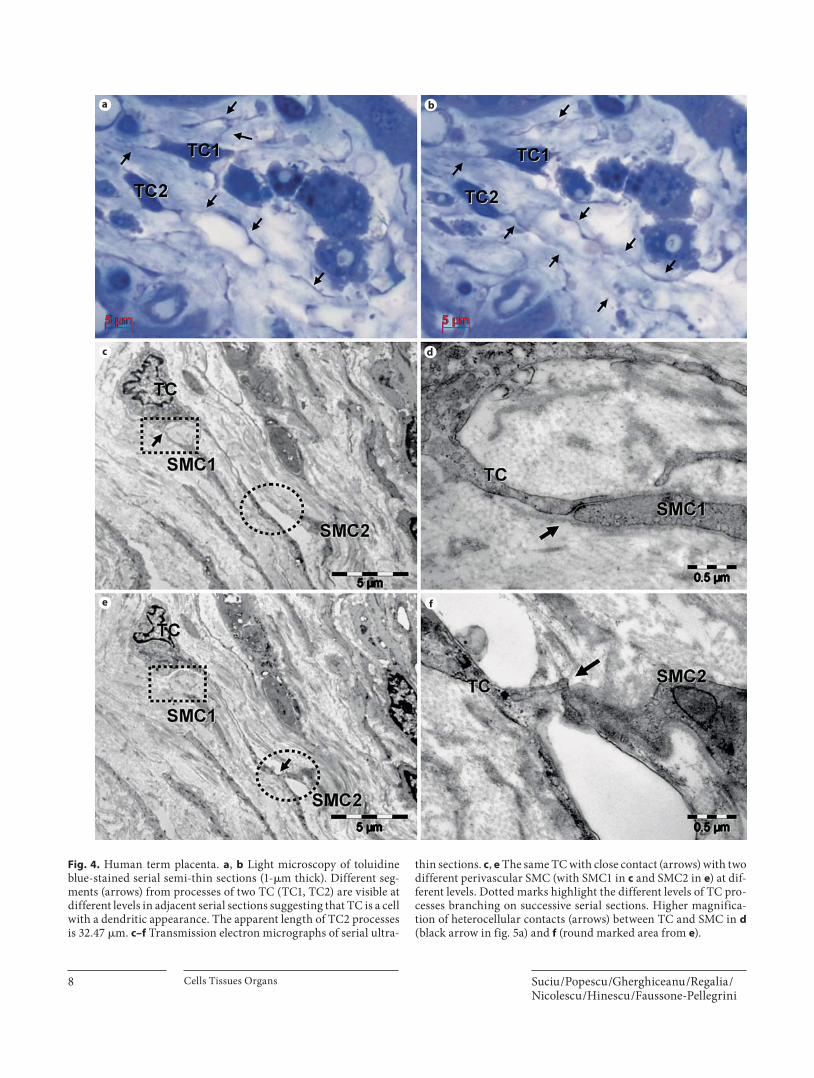
Fig. 4. Human term placenta. a , b Light microscopy of toluidine blue-stained serial semi-thin sections (1- � m thick). Different seg-ments (arrows) from processes of two TC (TC1, TC2) are visible at different levels in adjacent serial sections suggesting that TC is a cell with a dendritic appearance. The apparent length of TC2 processes is 32.47 � m. c–f Transmission electron micrographs of serial ultra-
thin sections. c , e The same TC with close contact (arrows) with two different perivascular SMC (with SMC1 in c and SMC2 in e ) at dif-ferent levels. Dotted marks highlight the different levels of TC pro-cesses branching on successive serial sections. Higher magnifica-tion of heterocellular contacts (arrows) between TC and SMC in d (black arrow in fig. 5a) and f (round marked area from e ).
Telocytes in Human Term Placenta Cells Tissues Organs 9
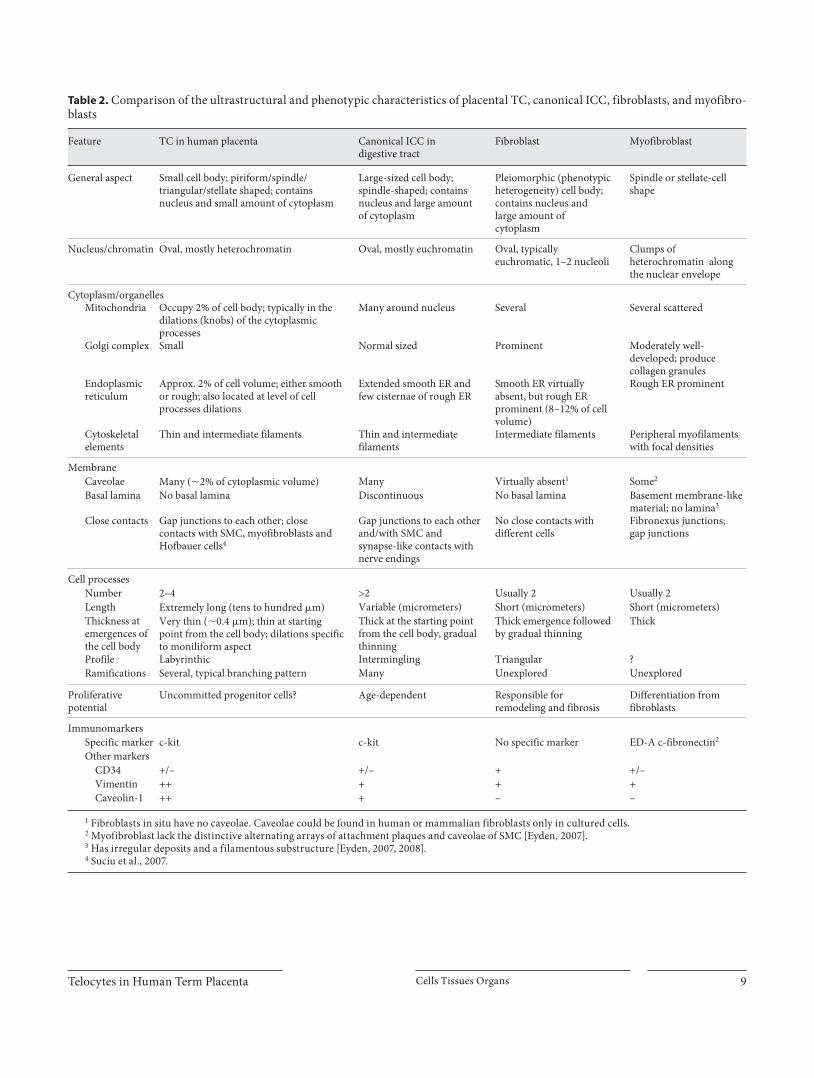
Table 2. Comparison of the ultrastructural and phenotypic characteristics of placental TC, canonical ICC, fibroblasts, and myofibro-blasts
Feature TC in human placenta Canonical ICC in digestive tract
Fibroblast Myofibroblast
General aspect Small cell body; piriform/spindle/triangular/stellate shaped; contains nucleus and small amount of cytoplasm
Large-sized cell body; spindle-shaped; contains nucleus and large amount of cytoplasm
Pleiomorphic (phenotypic heterogeneity) cell body; contains nucleus and large amount of cytoplasm
Spindle or stellate-cell shape
Nucleus/chromatin Oval, mostly heterochromatin Oval, mostly euchromatin Oval, typically euchromatic, 1–2 nucleoli
Clumps of heterochromatin along the nuclear envelope
Cytoplasm/organellesMitochondria Occupy 2% of cell body; typically in the
dilations (knobs) of the cytoplasmic processes
Many around nucleus Several Several scattered
Golgi complex Small Normal sized Prominent Moderately well-developed; produce collagen granules
Endoplasmicreticulum
Approx. 2% of cell volume; either smooth or rough; also located at level of cell processes dilations
Extended smooth ER and few cisternae of rough ER
Smooth ER virtually absent, but rough ER prominent (8–12% of cell volume)
Rough ER prominent
Cytoskeletalelements
Thin and intermediate filaments Thin and intermediate filaments
Intermediate filaments Peripheral myofilaments with focal densities
MembraneCaveolae Many (�2% of cytoplasmic volume) Many Virtually absent1 Some2
Basal lamina No basal lamina Discontinuous No basal lamina Basement membrane-like material; no lamina3
Close contacts Gap junctions to each other; close contacts with SMC, myofibroblasts and Hofbauer cells4
Gap junctions to each other and/with SMC and synapse-like contacts with nerve endings
No close contacts with different cells
Fibronexus junctions;gap junctions
Cell processesNumber 2–4 >2 Usually 2 Usually 2Length Extremely long (tens to hundred �m) Variable (micrometers) Short (micrometers) Short (micrometers)Thickness atemergences ofthe cell body
Very thin (�0.4 �m); thin at starting point from the cell body; dilations specific to moniliform aspect
Thick at the starting point from the cell body, gradual thinning
Thick emergence followed by gradual thinning
Thick
Profile Labyrinthic Intermingling Triangular ?Ramifications Several, typical branching pattern Many Unexplored Unexplored
Proliferativepotential
Uncommitted progenitor cells? Age-dependent Responsible for remodeling and fibrosis
Differentiation from fibroblasts
ImmunomarkersSpecific marker c-kit c-kit No specific marker ED-A c-fibronectin2
Other markersCD34 +/– +/– + +/–Vimentin ++ + + +Caveolin-1 ++ + – –
1 Fibroblasts in situ have no caveolae. Caveolae could be found in human or mammalian fibroblasts only in cultured cells.2 Myofibroblast lack the distinctive alternating arrays of attachment plaques and caveolae of SMC [Eyden, 2007].3 Has irregular deposits and a filamentous substructure [Eyden, 2007, 2008]. 4 Suciu et al., 2007.

Suciu /Popescu /Gherghiceanu /Regalia /Nicolescu /Hinescu /Faussone-Pellegrini
Cells Tissues Organs 10
tissue distribution of the presumed TC in close proxim-ity to fetal blood vessels from stem villi, regardless of their calibre.
We also tested the expression pattern of iNOS. A pop-ulation of extravascular stromal cells with suggestive TC morphology were positive for this marker, and only some of them were also c-kit positive. In term placenta, such cells were located around blood vessels in all types of stem villi. The rest of the iNOS-positive cells presumably belong to the monocyte/macrophage lineage. Some of the c-kit-positive cells were also positive for VEGF, another potentially ‘function-related’ marker. The processes of these cells extended into the axis of the stem villi ( fig. 3 d, panels 1–3). VEGF-positive cells could be seen in the ter-minal villi as well, but their morphology corresponded to Hofbauer cells.
Light Microscopy on Semithin Sections TC were easily identifiable as cells with long cellular
processes that surround blood vessels or extend into the connective tissue beneath the trophoblast in toluidine-blue semi-thin sections of both large and peripheral stem villi. The processes of these cells appears as segments in adjacent serial sections ( fig. 4 a, b). Because of their wavy course, the TC processes (TP) leave the section planes, suggesting that these cells have a dendritic rather than a sail-like appearance.
The relative proportion of these cells on semi-thin sec-tions was � 8% in large stem villi, and � 3% in smaller ones.
Transmission Electron Microscopy TEM examination was fundamental in identifying the
TC ( fig. 4 c, e, 5 a) as a distinct stromal cell type in human term placenta in addition to fibroblasts, myofibroblasts, SMC, Hofbauer cells, and mast cells.
The TC showed typical features [Ciontea et al., 2005; Popescu et al., 2005a]: a dendritic appearance, a small round or triangular body, and two to three processes which branch dichotomously ( fig. 5 a). These processes are key features of the TC, being long and moniliform, thin segments (0.1–0.5 � m) that alternate with dilations accommodating mitochondria and endoplasmic reticu-lum. Often, segments of the TP were observed without connection to the cellular body, and this aspect made it difficult to evaluate their length properly.
A rather heterochromatic nucleus, the presence of ca-veolae on the plasma membrane (over 2% of the cytoplas-mic volume), and an endoplasmic reticulum ( � 2%, rough and/or smooth) that was less well represented favor TC
in the differential comparison with fibroblasts ( fig. 5 b). None of the TC had the typical ultrastructural features of myofibroblasts ( fig. 5 c). The myofibroblasts were large spindle-shaped cells with myofilaments and focal densi-ties in the periphery, fibronectin fibrils adjacent to cell membrane, and distinctive fibronexus junctions ( fig. 5 d, e). Often, lipid droplets were seen in the cytoplasm of fi-broblasts and myofibroblasts but not in the cytoplasm of TC. Comparative features of TC from human placenta, ICC from digestive tract [Huizinga et al., 1997; Junquera et al., 2007; Pieri et al., 2008; Kostin and Popescu, 2009], fibroblast [Goldsmith et al., 2004; Camelliti et al., 2005] and myofibroblast [Eyden, 2007, 2008] are presented in table 2 .
TC were found in the large stem villi, with their long, slender processes surrounding the blood vessel wall or interposed between arterioles and the trophoblast base-ment membrane in small stem villi ( fig. 4 c, e). The aver-age distance between first layer of perivascular TC and the blood vessel wall ranged from 190.87 8 16.8 nm in the small stem villi to 507.84 8 15.1 nm in large ones. The TC were connected by gap junctions ( fig. 6 ) in a network extending into the placental stroma between concentric layers formed by SMC and myofibroblasts around large blood vessels.
The TC had close contacts with mast cells, Hofbauer cells, myofibroblasts, and SMC. An interesting aspect was seen in serial ultrathin sections, which showed that one TC had close contacts with different perivascular SMC ( fig. 4 c–f, 5 a) through different branches.
Fig. 5. Electron micrographs show three different types of fibro-blast-like stromal cells in human term placenta. a Another section showing the same TC from figure 4c, e. The TC has few organelles in the perinuclear area and three emerging thin processes (red ar-rows); black arrowheads mark the branching points (high magni-fication in fig. 4d). One TC process is in close apposition with a SMC (black arrow). Digitally colored images emphasize TC (blue) and SMC (brown). b A placental fibroblast (Fb) has abundant rough endoplasmic reticulum with dilated cisternae. c A placental myofibroblast (MyFb) has cortical myofilaments (arrows) and abundant rough endoplasmic reticulum, lipid droplets (l), corti-cal myofilaments, and dense bodies. d , e Electron micrographs show small fibronexus junctions as diagnostic features of myofi-broblasts. Fibronectin fibrils (arrows) are co-linear with intracel-lular myofilaments (asterisks).

Telocytes in Human Term Placenta Cells Tissues Organs 11
a b
c
d
e
l

Suciu /Popescu /Gherghiceanu /Regalia /Nicolescu /Hinescu /Faussone-Pellegrini
Cells Tissues Organs 12
Discussion
We characterize a cell population in the human term placenta that was previously included in the vast popula-tion of placental stromal cells. The results we obtained so far prove that it has many phenotypic and morphologic features related to the archetypal TC.
Telocytes: Distribution, Morphology and Immunophenotype The cells we identified as TC in the human term pla-
centa are located in the interstitium and their typical, long, thin, moniliform processes could be detected both by light and electron microscopy. Through these process-es, these cells are likely to form a 3-dimensional network on the axis of the villi. The morphology of TC and their intravillous organization is markedly similar to that of cells previously observed under TEM and scanning elec-tron microscopy [Castellucci and Kaufmann, 1982; Mar-
tinoli et al., 1984; Sati et al., 2007] and included in the mesenchymal or stromal population.
In addition to confirming these previous studies, our present data describe the immunophenotype of TC, which may be of value when considering their possible function.
We did not observe significant dissimilarities between the phenotypic profile of in situ placental TC and that of cultured cells ( table 1 ). Indeed, either in situ or in culture, these cells were consistently found to be positive for c-kit. Some previous studies did not demonstrate any c-kit pos-itivity in human placenta or endometrium [Lammie et al., 1994], while some others reported c-kit expression in trophoblast and, to a lesser extent, in scattered cells of the placental villous core [Kauma et al., 1996]. These con-trasting results could be explained by either a ‘switching’ phenotype behavior in similar cell populations, by ex-amination of dissimilar cell (sub)populations, or by var-ied technical procedures in different studies. A similar
Fig. 6. Electron micrograph shows a gap junction (inset, arrows) connecting the processes of two TC (TC1, TC2) in the vicinity of a blood vessel from the placental villous core.

Telocytes in Human Term Placenta Cells Tissues Organs 13
controversy regarding the presence or absence of c-kit ex-pression exists for myometrial TC [Hutchings et al., 2009].
We also found that CD34 was expressed by placental TC similar to those of the gut [Pieri et al., 2008] and oth-er organs – pancreas [Popescu et al., 2005b], atrial myo-cardium [Hinescu et al., 2006], fallopian tube [Popescu et al., 2005a], myometrium [Ciontea et al., 2005] and mes-entery [Hinescu et al., 2008]. The other consistently-ex-pressed marker was vimentin, confirming that TC are of mesenchymal origin.
Placental TC were negative for SMM and desmin staining. Together with the less organized distribution of � SMA observed in cultured cells ( fig. 1 f), these data may represent additional arguments for a differential compar-ison with SMC [Leik et al., 2004] and myofibroblasts, the cell types that many immunohistochemical [Feller et al., 1985; Kohnen et al., 1996] and some electron microscop-ic studies [Demir et al., 1992] have estimated as being the most abundant in the villi stroma. Finally, cultured TC do not express the cell adhesion molecule CD90 ( fig. 1 c), which was previously identified on the surface of fibro-blasts [Sorrell and Caplan, 2009].
Telocytes: A Subpopulation of Progenitor Cells in Human Placenta Currently, it is generally accepted that the young con-
nective tissue in the placenta is composed of mesenchy-mal cells that will eventually differentiate into all stromal cells. This might be the missing link regarding the origin of TC, which could be considered an early step in the dif-ferentiation process of the mesenchymal cells that ac-quire different functions depending on location. Besides being c-kit-positive, other stem-cell markers were found to be co-expressed on TC including CD34, CD44 [Blond-heim et al., 2006], and nestin [Lendahl et al., 1990]. This phenotypic profile implies that it is reasonable to con-sider placental TC, in addition to having specific func-tional roles in their own right, as a subpopulation of pro-genitor cells among the stromal cells of human placenta.
Telocytes and Blood Flow Regulation The structural properties of the placenta exchange
barrier need to take into account all factors potentially affecting placental supply capacity [Desforges and Sibley, 2009]. However, current conceptual models disregard the possible interposition of TC into the blood flow regula-tion.
Examined from this perspective, placental TC have multiple projections connected by gap junctions with neighboring TC in a network-like distribution around
blood vessels, and are in close apposition with the SMC and the surrounding myofibroblasts. SMC or myofibro-blasts form a clearly-defined, perivascular contractile sheath previously demonstrated in large stem villi [Graf et al., 1994] and anchoring villi [Farley et al., 2004], and it was suggested that the contraction of these cells adjusts the blood flow in fetal vessels and increases the turgor, imparting mechanical stability to the villous tree in the maternal blood stream. Moreover, contraction of longi-tudinally-arranged myofibroblasts within anchoring vil-li may influence the length and width of the intervillous space, thus regulating maternal intervillous blood pres-sure [Graf et al., 1994; Farley et al., 2004]. The stimulus for this type of contraction is not yet established because the placenta is a non-innervated organ. Even if no distinct junctions between TC and these cell types were apparent, we found many areas of close contact that suggest some sort of communication or coupling. Another possible role for TC is intervention in blood flow regulation through an inhibitory signaling pathway that involves NO. It is well known that NO acts as a vasodilator [Moncada et al., 1991]. Previous studies showed that iNOS may be the ma-jor producer of NO in the rat placenta and placental iNOS mRNA expression increases in the final gestational stag-es and is likely to generate sufficient NO to maintain local placental blood flow [Takizawa et al., 2002]. Our study, however, showed that only a fraction of TC expressed iNOS at a level detectable by immuno staining techniques, possibly because all samples were harvested from women who had pre-labor caesarean sections after normal preg-nancies. NOS activity and NO production are also sig-nificantly increased in placentas affected by pre-eclamp-sia and eclampsia. Their levels are related to the severity of the disorder; however, their presence may represent a physiological adaptive response to overcome vascular re-sistance and prevent platelet and leukocyte adhesion to the surface of placental villi or within the intervillous spaces [Shaamash et al., 2001].
Further exploration of the potential role of placental TC in the regulation of fetal placental blood flow could be of great interest in developing our understanding of the pathological processes involved in pre-eclampsia and intrauterine growth restriction.
Telocytes and Tissue Remodeling Remodeling is a process that takes place throughout
the entire life of the placenta along with maturation of the intermediate villi. The presence of a subtrophoblastic macrophage population may be important for the initia-tion of stromal fibrosis, as well as for angiogenesis, by

Suciu /Popescu /Gherghiceanu /Regalia /Nicolescu /Hinescu /Faussone-Pellegrini
Cells Tissues Organs 14
modulating endothelial cell proliferation through soluble angiogenic factors [Seval et al., 2007]. However, as dem-onstrated by our results, TC themselves could contribute to this process, since these cells express CD44 and VEGF. CD44 expression supports a role for this cell in stabilizing and trimming the hyaluronan network that is important in the maintenance of the structural integrity of the pla-centa and intervening in tissue remodeling later in gesta-tion [Marzioni et al., 2001]. The expression of VEGF is relevant because this molecule acts as a potent angiogen-ic agent [Reynolds et al., 2009] in promoting endothelial cell proliferation and vascular permeability [Keck et al., 1989], and is a maintenance factor for newly-formed blood vessels [Alon et al., 1995].
In conclusion, this study shows that TC exist in human term placenta. On the basis of their distribution and mor-phology, we suggest that these cells may be involved in tissue remodeling and, since the placenta is a non-inner-vated organ, they may also intervene in blood flow reg-ulation. Considering their immunophenotype, TC may also be a subpopulation of progenitor cells. Currently,
however, the presumed role(s) of placental TC is the sub-ject of speculation, and further studies are required, in-cluding studies in preterm placental tissue.
Note Added in Proof
Rationale for the Terms ‘Telocytes’ and ‘Telopodes’ In 2005, we described a new type of cell that has be-
come known as the ‘interstitial Cajal-like cell’ (ICLC). We named these cells ICLC because they were apparently similar to the well-known gastrointestinal interstitial cells of Cajal (ICC). However, electron microscopy and immu-nohistochemistry have now shown them to be completely different from ICC. In a recently published review from our laboratory [Popescu and Faussone-Pellegrini, 2010], we coined the term ‘telocytes’ as a new name for ICLC and ‘telopodes’ for their prolongations. This terminology should prevent further confusion with other interstitial cell types (e.g. fibroblasts, fibrocytes, fibroblast-like cells, mesenchymal cells and interstitial Cajal cells).
References
Alon, T., I. Hemo, A. Itin, J. Pe’er, J. Stone, E. Kes-het (1995) Vascular endothelial growth factor acts as a survival factor for newly formed ret-inal vessels and has implications for retinop-athy of prematurity. Nat Med 1: 1024–1028.
Benirschke, K., P. Kaufmann, R. Baergen (2006) Pathology of the Human Placenta. New York, Springer.
Blondheim, N.R., Y.S. Levy, T. Ben-Zur, A. Burshtein, T. Cherlow, I. Kan, R. Barzilai, M. Bahat-Stromza, Y. Barhum, S. Bulvik, E. Melamed, D. Offen (2006) Human mesen-chymal stem cells express neural genes, sug-gesting a neural predisposition. Stem Cells Dev 15: 141–164.
Bussolati, G. (2005) Of GISTs and EGISTs, ICCs and ICs. Virchows Arch 447: 907–908.
Camelliti, P., T.K. Borg, P. Kohl (2005) Structur-al and functional characterisation of cardiac fibroblasts. Cardiovasc Res 65: 40–51.
Castellucci, M., P. Kaufmann (1982) A three-di-mensional study of the normal human pla-cental villous core. II. Stromal architecture. Placenta 3: 269–285.
Ciontea, S.M., E. Radu, T. Regalia, L. Ceafalan, D. Cretoiu, M. Gherghiceanu, R.I. Braga, M. Malincenco, L. Zagrean, M.E. Hinescu, L.M. Popescu (2005) C-kit immunopositive inter-stitial cells (Cajal-type) in human myome-trium. J Cell Mol Med 9: 407–420.
Demir, R., N. Demir, G. Kohnen, G. Kosanke, A. Mironov, I. Uestuenel, E. Kocamaz (1992) Ultrastructure and distribution of myofibro-blast-like cells in human placental stem villi. Electron Microscopy 3: 509–510.
Demir, R., G. Kosanke, G. Kohnen, S. Kertschan-ska, P. Kaufmann (1997) Classification of human placental stem villi: review of struc-tural and functional aspects. Microsc Res Tech 38: 29–41.
Desforges, M., C.P. Sibley (2010) Placental nutri-ent supply and fetal growth. Int J Dev Biol 54: 377–390.
Eyden, B. (2007) The myofibroblast: a study of normal, reactive and neoplastic tissues, with an emphasis on ultrastructure. J Submicrosc Cytol Pathol 7–166.
Eyden, B. (2008) The myofibroblast: phenotypic characterization as a prerequisite to under-standing its functions in translational medi-cine. J Cell Mol Med 12: 22–37.
Farley, A.E., C.H. Graham, G.N. Smith (2004) Contractile properties of human placental anchoring villi. Am J Physiol Regul Integr Comp Physiol 287: R680–R685.
Farrugia, G. (2008) Interstitial cells of Cajal in health and disease. Neurogastroenterol Mo-til 20(suppl 1): 54–63.
Faussone-Pellegrini, M.S., L. Thuneberg (1999) Guide to the identification of interstitial cells of Cajal. Microsc Res Tech 47: 248–266.
Feller, A.C., H. Schneider, D. Schmidt, M.R. Par-waresch (1985) Myofibroblast as a major cel-lular constituent of villous stroma in human placenta. Placenta 6: 405–415.
Garcia-Lopez, P., V. Garcia-Marin, R. Martinez-Murillo, M. Freire (2009) Updating old ideas and recent advances regarding the Intersti-tial Cells of Cajal. Brain Res Rev 61: 154–169.
Goldsmith, E.C., A. Hoffman, M.O. Morales, J.D. Potts, R.L. Price, A. McFadden, M. Rice, T.K. Borg (2004) Organization of fibroblasts in the heart. Dev Dyn 230: 787–794.
Graf, R., J.U. Langer, G. Schonfelder, T. Oney,S. Hartel-Schenk, W. Reutter, H.H. Schmidt (1994) The extravascular contractile system in the human placenta: morphological and immunocytochemical investigations. Anat Embryol (Berl) 190: 541–548.
Graf, R., G. Schonfelder, M. Muhlberger, M. Guts-mann (1995) The perivascular contractile sheath of human placental stem villi: its isola-tion and characterization. Placenta 16: 57–66.
Hinescu, M.E., M. Gherghiceanu, E. Mandache, S.M. Ciontea, L.M. Popescu (2006) Intersti-tial Cajal-like cells (ICLC) in atrial myocar-dium: ultrastructural and immunohisto-chemical characterization. J Cell Mol Med 10: 243–257.

Telocytes in Human Term Placenta Cells Tissues Organs 15
Hinescu, M.E., L.M. Popescu, M. Gherghiceanu, M.S. Faussone-Pellegrini (2008) Interstitial Cajal-like cells in rat mesentery: an ultra-structural and immunohistochemical ap-proach. J Cell Mol Med 12: 260–270.
Huizinga, J.D., L. Thuneberg, J.M. Vanderwin-den, J.J. Rumessen (1997) Interstitial cells of Cajal as targets for pharmacological inter-vention in gastrointestinal motor disorders. Trends Pharmacol Sci 18: 393–403.
Hutchings, G., O. Williams, D. Cretoiu, S.M. Ciontea (2009) Myometrial interstitialcells and the coordination of myometrial contractility. J Cell Mol Med 13: 4268–4282.
Junquera, C., C. Martinez-Ciriano, T. Castiella, P. Serrano, M.J. Azanza, S.R. Junquera (2007) Immunohistochemical and ultra-structural characteristics of interstitial cells of Cajal in the rabbit duodenum: presence of a single cilium. J Cell Mol Med 11: 776–787.
Kauma, S., T. Huff, G. Krystal, J. Ryan, P. Takacs, T. Turner (1996) The expression of stem cell factor and its receptor, c-kit in human endo-metrium and placental tissues during preg-nancy. J Clin Endocrinol Metab 81: 1261–1266.
Keck, P.J., S.D. Hauser, G. Krivi, K. Sanzo, T. War-ren, J. Feder, D.T. Connolly (1989) Vascular permeability factor, an endothelial cell mito-gen related to PDGF. Science 246: 1309–1312.
Kohnen, G., S. Kertschanska, R. Demir, P. Kaufmann (1996) Placental villous stroma as a model system for myofibroblast differen-tiation. Histochem Cell Biol 105: 415–429.
Kostin, S., L.M. Popescu (2009) A distinct type of cell in myocardium: interstitial Cajal-like cells (ICLCs). J Cell Mol Med 13: 295–308.
Lammie, A., M. Drobnjak, W. Gerald, A. Saad, R. Cote, C. Cordon-Cardo (1994) Expression of c-kit and kit ligand proteins in normal human tissues. J Histochem Cytochem 42: 1417–1425.
Leik, C.E., A. Willey, M.F. Graham, S.W. Walsh (2004) Isolation and culture of arterial smooth muscle cells from human placenta. Hypertension 43: 837–840.
Lendahl, U., L.B. Zimmerman, R.D. McKay (1990) CNS stem cells express a new class of intermediate filament protein. Cell 60: 585–595.
Martinoli, C., M. Castellucci, D. Zaccheo, P. Kaufmann (1984) Scanning electron micros-copy of stromal cells of human placental vil-li throughout pregnancy. Cell Tissue Res 235: 647–655.
Marzioni, D., C. Crescimanno, D. Zaccheo, R. Coppari, C.B. Underhill, M. Castellucci (2001) Hyaluronate and CD44 expression patterns in the human placenta throughout pregnancy. Eur J Histochem 45: 131–140.
McCloskey, K.D., A.M. Gurney (2002) Kit posi-tive cells in the guinea pig bladder. J Urol 168: 832–836.
Moncada, S., R.M. Palmer, E.A. Higgs (1991) Nitric oxide: physiology, pathophysiology, and pharmacology. Pharmacol Rev 43: 109–142.
Pieri, L., M.G. Vannucchi, M.S. Faussone-Pel-legrini (2008) Histochemical and ultrastruc-tural characteristics of an interstitial cell type different from ICC and resident in the muscle coat of human gut. J Cell Mol Med 12: 1944–1955.
Popescu, L.M., S.M. Ciontea, D. Cretoiu, M.E. Hinescu, E. Radu, N. Ionescu, M. Ceausu, M. Gherghiceanu, R.I. Braga, F. Vasilescu, L. Zagrean, C. Ardeleanu (2005a) Novel type of interstitial cell (Cajal-like) in human fallo-pian tube. J Cell Mol Med 9: 479–523.
Popescu, L.M, M.S. Faussone-Pellegrini (2010) Telocytes – A case of serendipity: the wind-ing way from Interstitial Cells of Cajal (ICC), via Interstitial Cajal-Like Cells (ICLC) to Telocytes. J Cell Mol Med 14: 729–740.
Popescu, L.M., M.E. Hinescu, N. Ionescu, S.M. Ciontea, D. Cretoiu, C. Ardelean (2005b) In-terstitial cells of Cajal in pancreas. J Cell Mol Med 9: 169–190.
Povstyan, O.V., D.V. Gordienko, M.I. Harhun, T.B. Bolton (2003) Identification of intersti-tial cells of Cajal in the rabbit portal vein. Cell Calcium 33: 223–239.
Reilly, R.D., P.T. Russell (1977) Neurohisto-chemical evidence supporting an absence of adrenergic and cholinergic innervation in the human placenta and umbilical cord. Anat Rec 188: 277–286.
Reynolds, L.P., P.P. Borowicz, J.S. Caton, K.A. Vonnahme, J.S. Luther, D.S. Buchanan, S.A. Hafez, A.T. Grazul-Bilska, D.A. Redmer (2010) Uteroplacental vascular development and placental function: an update. Int J Dev Biol 54: 355–366.
Sanders, K.M., S.D. Koh, S.M. Ward (2006) In-terstitial cells of cajal as pacemakers in the gastrointestinal tract. Annu Rev Physiol 68 : 307–343.
Sati, L., Y. Seval, A. Yasemin Demir, G. Kosanke, G. Kohnen, R. Demir (2007) Cellular diver-sity of human placental stem villi: an ultra-structural and immunohistochemical study. Acta Histochem 109: 468–479.
Seval, Y., E.T. Korgun, R. Demir (2007) Hofbau-er cells in early human placenta: possible im-plications in vasculogenesis and angiogene-sis. Placenta 28 : 841–845.
Shaamash, A.H., E.D. Elsonosy, M.M. Zakhari, S.H. Radwan, H.M. El-Dien (2001) Placental nitric oxide synthase (NOS) activity and ni-tric oxide (NO) production in normal preg-nancy, pre-eclampsia and eclampsia. Int J Gynaecol Obstet 72: 127–133.
Sorrell, J.M., A.I. Caplan (2009) Fibroblasts-a diverse population at the center of it all. Int Rev Cell Mol Biol 276: 161–214.
Suciu, L., L.M. Popescu, M. Gherghiceanu (2007) Human placenta: de visu demonstra-tion of interstitial Cajal-like cells. J Cell Mol Med 11: 590–597.
Takizawa, T., H. Yoshikawa, M. Yamada, H. Morita (2002) Expression of nitric oxide syn-thase isoforms and detection of nitric oxide in rat placenta. Am J Physiol Cell Physiol 282: C762–C767.
Weibel, E. (1979) Stereological Methods: Practi-cal Methods for Biological Morphometry. New York, Academic Press.
steimlej
CopyBAhoch




















