
Brain aromatase and the control of male sexual behavior
20
J. Steroid Biochem. Molec. Biol. Vol. 44, No. 4-6, pp. 521-540, 1993 0960-0760/93$6.00 + 0.00 Printed in Great Britain. All rights reserved Copyright© 1993PergamonPress Ltd BRAIN AROMATASE AND THE CONTROL OF MALE SEXUAL BEHAVIOR J. BALTHAZART* and A. FOIDART Laboratory of General and Comparative Biochemistry, University of Li6ge (Bat. LI), 17 place Delcour, 4020 Li6ge, Belgium Summary--The activational effects of testosterone (T) on male copulatory behavior are mediated by its aromatization into estradiol. In quail, we have shown by stereotaxic implantation of steroids and metabolism inhibitors and by electrolytic lesions that the action of T and its aromatization take place in the sexually dimorphic medial preoptic nucleus (POM). The distribution and regulation of brain aromatase was studied in this species by product-formation assays measuring aromatase activity (AA) in microdissected brain regions and by immunocytochemistry (ICC). Aromatase-immunoreactive (ARO-ir) neurons were found in four brain regions: the POM, the septal region, the bed nucleus of the stria terminals (BNST) and the tuberal hypothalamus. ARO-ir cells actually outline the POM boundaries. ARO-ir material is found not only in the perikarya of neurons but also in the full extension of their cellular processes including the axons and the presynaptic boutons. This is confirmed at the light level by the demonstration of immunoreactive fibers and punctate structures in brain regions that are sometimes fairly distant from the closest ARO-ir cells. A lot of ARO-ir cells in the POM and BNST do not contain immunoreactive estrogen receptors (ER-ir) as demonstrated by double label ICC. These morphological data suggest an unorthodox role for the enzyme or the locally formed estrogens. In parallel with copulatory behavior, the preoptic AA decreases after castration and is restored by T to levels seen in sexually mature males. This probably reflects a change in enzyme concentration rather than a modulation of the activity in a constant number of molecules since the maximum enzymatic velocity (Vmax) only is affected while the affinity (Kin) remains unchanged. In addition, T increases the number of ARO-ir neurons in POM and other brain areas suggesting that the concentration of the antigen is actually increased. This probably involves the direct activation of aromatase transcription as demonstrated by RT-PCR studies showing that aromatase mRNA is increased following T treatment of castrates. These activating effects of T seem to result from a synergistic action of androgenic and estrogenic metabolites of the steroid. The anatomical substrate for these regulations remains unclear at present especially in POM where ARO-ir cells do not in general contain ER-ir while androgen receptors appear to be rare based on both [3H] dihydrotestosterone autoradiography and ICC. Transynaptic mechanisms of control may be considered. A modulation of brain aromatase by catecholamines is also suggested by a few pharmacological studies. This notion is further supported by anatomical data demonstrating dense projections of dopamine fl-hydroxylase and tyrosine hydroxylase-immunoreactive fibers around ARO-ir cells. INTRODUCTION Aromatase also called estrogen synthetase, an enzyme of the cytochrome P450 family (P- 450 .... ; EC 1.14.14.1), irreversibly transforms testosterone (T) into estradiol-17fl (E2) or an- drostenedione (A4) into estrone (El). This en- zyme was originally found in the ovary and placenta but in the early seventies, Naftolin and Proceedings of the Third International Aromatase Confer- ence. Basic and Clinical Aspects of Arornatase, Bologna, Italy, 14 17 June 1992. *To whom correspondence should be addressed coworkers [1, 2] were able to identify a low aromatase activity (AA) in the brain of several mammalian species. This enzymatic activity has since been identified in the brain of many species belonging to all vertebrate classes from jawed fishes to mammals. The presence of this enzyme in the brain of cyclosomes is questionable and it has not been detected in invertebrates, such as the lobster and the octopus [3-5]. Brain AA is usually low in higher vertebrates (orders of magnitude below activities normally observed in peripheral structures) but the en- zyme has a critical importance in the control of reproductive processes. There is a wealth of 521
Transcript of Brain aromatase and the control of male sexual behavior

J. Steroid Biochem. Molec. Biol. Vol. 44, No. 4-6, pp. 521-540, 1993 0960-0760/93 $6.00 + 0.00 Printed in Great Britain. All rights reserved Copyright © 1993 Pergamon Press Ltd
BRAIN AROMATASE AND THE CONTROL OF MALE SEXUAL BEHAVIOR
J. BALTHAZART* and A. FOIDART
Laboratory of General and Comparative Biochemistry, University of Li6ge (Bat. LI), 17 place Delcour, 4020 Li6ge, Belgium
Summary--The activational effects of testosterone (T) on male copulatory behavior are mediated by its aromatization into estradiol. In quail, we have shown by stereotaxic implantation of steroids and metabolism inhibitors and by electrolytic lesions that the action of T and its aromatization take place in the sexually dimorphic medial preoptic nucleus (POM). The distribution and regulation of brain aromatase was studied in this species by product-formation assays measuring aromatase activity (AA) in microdissected brain regions and by immunocytochemistry (ICC). Aromatase-immunoreactive (ARO-ir) neurons were found in four brain regions: the POM, the septal region, the bed nucleus of the stria terminals (BNST) and the tuberal hypothalamus. ARO-ir cells actually outline the POM boundaries. ARO-ir material is found not only in the perikarya of neurons but also in the full extension of their cellular processes including the axons and the presynaptic boutons. This is confirmed at the light level by the demonstration of immunoreactive fibers and punctate structures in brain regions that are sometimes fairly distant from the closest ARO-ir cells. A lot of ARO-ir cells in the POM and BNST do not contain immunoreactive estrogen receptors (ER-ir) as demonstrated by double label ICC. These morphological data suggest an unorthodox role for the enzyme or the locally formed estrogens. In parallel with copulatory behavior, the preoptic AA decreases after castration and is restored by T to levels seen in sexually mature males. This probably reflects a change in enzyme concentration rather than a modulation of the activity in a constant number of molecules since the maximum enzymatic velocity (Vmax) only is affected while the affinity (Kin) remains unchanged. In addition, T increases the number of ARO-ir neurons in POM and other brain areas suggesting that the concentration of the antigen is actually increased. This probably involves the direct activation of aromatase transcription as demonstrated by RT-PCR studies showing that aromatase mRNA is increased following T treatment of castrates. These activating effects of T seem to result from a synergistic action of androgenic and estrogenic metabolites of the steroid. The anatomical substrate for these regulations remains unclear at present especially in POM where ARO-ir cells do not in general contain ER-ir while androgen receptors appear to be rare based on both [3H] dihydrotestosterone autoradiography and ICC. Transynaptic mechanisms of control may be considered. A modulation of brain aromatase by catecholamines is also suggested by a few pharmacological studies. This notion is further supported by anatomical data demonstrating dense projections of dopamine fl-hydroxylase and tyrosine hydroxylase-immunoreactive fibers around ARO-ir cells.
INTRODUCTION
Aromatase also called estrogen synthetase, an enzyme of the cy tochrome P450 family (P- 450 . . . . ; EC 1.14.14.1), irreversibly t ransforms testosterone (T) into estradiol-17fl (E2) or an- drostenedione (A4) into estrone (El). This en- zyme was originally found in the ovary and placenta but in the early seventies, Naftol in and
Proceedings of the Third International Aromatase Confer- ence. Basic and Clinical Aspects of Arornatase, Bologna, Italy, 14 17 June 1992.
*To whom correspondence should be addressed
coworkers [1, 2] were able to identify a low aromatase activity (AA) in the brain o f several mammal ian species. This enzymatic activity has since been identified in the brain o f many species belonging to all vertebrate classes f rom jawed fishes to mammals . The presence o f this enzyme in the brain o f cyclosomes is questionable and it has not been detected in invertebrates, such as the lobster and the octopus [3-5].
Brain A A is usually low in higher vertebrates (orders o f magni tude below activities normally observed in peripheral structures) but the en- zyme has a critical importance in the control o f reproductive processes. There is a wealth o f
521

522 J. BALTFIAZART and A. FOIDART
information coming from mammalian and non- mammalian studies which supports the idea that many effects of T in the brain, in fact, require the transformation of this androgen into E2 (for review see [5,6]). This represents the "aroma- tization hypothesis" which states that, ulti- mately, the effect of T in the brain requires its aromatization into estrogen. There is also a stronger version of this hypothesis that has been called the "estrogen-receptor hypothesis" which proposes that the action of T depends on bind- ing of E2 (derived from T aromatization) to the estrogen receptors [6, 7].
In particular, the activation of male copula- tory behavior seems to depend on the central aromatization of T in many species (for review see [5, 6, 8]). This notion is supported by in vivo studies analyzing the behavioral effects of aromatizable vs non-aromatizable androgens, of estrogens vs androgens and of aromatase inhibitors or antiestrogens. In addition, a lot of studies have shown that AA is present in the brain areas that control male sex behavior and that the enzymatic activity is regulated by T in a manner consistent with the notion that the enzyme is a limiting step for the action of T on behavior.
In the following, I would like to review recent studies coming from my and from collaborating laboratories carried out on the Japanese quail (Coturnix japonica) , a species that has turned out to be an excellent model for the study of brain aromatase and of its implication in the control of male sex behavior. I will first briefly summarize the experimental data demonstrating that aromatase plays a key role in the activation by T of male sex behavior. The regulations of AA as studied by product-formation assays will later be considered. I will then describe more recent studies in which the distribution of aromatase could be studied in the brain of the quail at a cellular level of resolution with the help of an immunocytochemical procedure using a purified polyclonal antibody raised in rabbit against human placental aromatase. The mechanisms that control brain aromatase will finally be considered and I will describe separ- ately experiments analyzing the control of AA as studied by product-formation assays, the control of aromatase-immunoreactive (ARO-ir) cells as analyzed by immunocytochemistry (ICC) and finally the control of the concen- tration of aromatase mRNA as analyzed by a reverse transcriptase-polymerase chain reaction (PCR) technique.
TESTOSTERONE ACTIVATES MALE COPULATORY BEHAVIOR AND THIS EFFECT REQUIRES ITS
AROMATIZATION INTO ESTROGENS
In quail, like in most mammals and birds that have been studied, the activation of male copu- latory behavior by T requires its aromatization into estrogens. This notion is supported by several types of experimental data:
-estrogens per se can restore male behavior in castrated males [9-12];
.... aromatizable androgens, such as T or A4 activate male copulatory behavior while non-aromatizable androgens, such as 5~- dihydrotestosterone (5e-DHT) and the de- rived diols have no or only weak effects [9-12]
- aromatase inhibitors such as androstatri- enedione (ATD) or R76713 (6-[(4-chloro- phenyl)(l H - 1,2,4- triazol - 1 - yl)methyl] - 1- methyl-lH-benzotriazole) inhibit or com- pletely block the behavioral effects of T[lO, 13, 141 antiestrogens such as tamoxifen or nitro- mifene citrate (CI-628) block the T-induced male sexual behavior [15, 16].
These data clearly support the "aromatiza- tion hypothesis" and even the "estrogen- receptor hypothesis" as far as the activation of quail sexual behavior is concerned. They should however not lead to the conclusion that andro- gens per se are inactive at this level. It is indeed clear that 5c~-DHT has a weak but never- theless significant effect on copulatory behav- ior[17, 18]. In addition the non-aromatizable androgen, methyltrienolone or R1881 [19---22] restores a weak male sex behavior m castrated quail [12, 23]. Finally, antiandrogens, although less effective than the antiestrogens, are never° theless able to significantly diminish the T- induced male behavior[16]. It is also well established that non-aromatizable androgens synergize with low doses of estrogen to fully restore male behavior in quail [9, 10, 12], even if the precise mechanism underlying this syner- gism has not been identified so far. Therefore in vivo studies demonstrate that T aromatization plays a critical role for the activation of copw lation in castrated quail but they do not rule out a specific contribution of androgens. These data demonstrate that the hormonal specificity of the behavioral activation in quail is very similar to the specificity observed in the rat (see [6] for a more detailed discussion).

Aromatase and sexual behavior 523
THE MEDIAL PREOPTIC NUCLEUS IPOMI IS A SUFFICIENT SITE FOR T ACTION ON BEHAVIOR
Special efforts were devoted to the identifi- cation in quail of the brain site(s) implicated in the action of T on male sexual behavior. Atten- tion was centered on the preoptic area (POA) because, in both birds and mammals, this region binds sex steroids [24-27]. Studies based on the stereotaxic implantation of steroids and on elec- trolytic lesions also indicated that T action in the POA is, in many species, sufficient to acti- vate sex behavior in castrated males [28].
Morphometric studies of Nissl-stained sec- tions in the POA of quail were undertaken in collaboration with the group of Drs G. C. Panzica and C. Viglietti-Panzica (University of Torino, Italy). This led to the identification of a sexually dimorphic nucleus (SDN) in the quail POA. The POM is indeed significantly larger in male than in female quail [29, 30]. It is also T-sensitive: its volume regresses after castration and increases following T treatment [31, 32]. POM therefore provides a morphological signa- ture of T action in the POA.
These data indicating that POM, a structure located medially in the POA, was T-sensitive suggested that this nucleus might be implicated in the control of male copulatory behavior. This idea was reinforced by the identification of two neurochemical characteristics of this nucleus: the presence of estrogen receptors identified both by [3H]E2 autoradiography [33] and by immunocytochemistry [34] and the presence of a very high AA as measured by in vitro product- formation assays performed on microsamples that had been dissected by the Palkovits punch technique [35]. Biochemical studies also indi- cated the presence of estrogen receptors (ER) in the quail POA-anterior hypothalamus and these were showing, in addition, that ER occupancy was correlated with the expression of aggressive behavior [36].
Electrolytic lesions and stereotaxic implan- tations of steroids, aromatase inhibitors and steroid receptor antagonists were undertaken to test the role of POM in the activation of male behavior.
Previously published studies indicated that stereotaxic implants of T in the POA located in or near POM were able to activate sexual behavior in castrated male quail [37, 38] but these did not permit a precise determination of the active area since fairly large amounts of steroid had been implanted and the diffusion area was unknown. Detailed studies using
smaller amounts of hormone were performed and they confirmed that T implants must be within the boundaries of the POM as defined in Nissl-stained material in order to activate be- havior [39, 40]. In addition, electrolytic lesions produced deficits in the T-induced copulatory behavior of castrated males that were pro- portional to the amount of the nucleus that was destroyed [39]. A series of studies in which syn- thetic androgens or estrogens, antiandrogens, antiestrogens and aromatase inhibitors were stereotaxically implanted in the POM of cas- trated male quail demonstrated that T must be aromatized and the synthesized estrogens must interact locally in the nucleus with ER in order to produce a significant behavior acti- vation [14, 41]. The quail POM is therefore a unique model structure for the study of the activation of masculine sexual behavior by ster- oids in that it is a T-sensitive sexually differenti- ated nucleus which is at the same time a sufficient site for the induction of behavior by T. Effects of T in this nucleus are also mediated through local aromatization.
AA IS REGULATED BY T IN THE QUAIL POA AND APPEARS TO BE A LIMITING STEP IN THE
ACTIVATION OF BEHAVIOR
Studies in another avian species, the ring dove ( Streptophelia risoria ) originally demonstrated that T markedly increases AA in the brain and especially in the POA [42]. A similar control mechanism was later identified in the rat [43, 44] and in the Japanese quail [45]. Detailed para- metrical studies were carried out to analyze the relationships between the induction by T of AA in the POA and the activation of male copula- tory behavior in castrated male quail [13]. This demonstrated that the induction of AA by T in the POA is dose- and time-dependent. Levels of AA seen in sexually mature males were restored in castrated birds by a treatment with 20 to 40 mm silastic T capsules which is known to produce physiological levels of steroid in the plasma. The minimal dose of T (10 mm implant) which reliably restored copulatory behavior ap- proximately doubled the AA in the POA. A significant increase in AA was already observed 16 h after the start of the treatment with T and the induction was maximal after 48 h. Acti- vation of copulatory behavior followed a similar time-course but occurred with a delay of 24 to 48h. Direct manipulation of AA using an aromatase inhibitor, ATD confirmed that the

524 J. BALTHAZART and A. FOIDART
enzymatic activity is a limiting factor in the activation of copulatory behavior. If T-treated birds received ATD at the same time, the acti- vation of behavior was suppressed for at least 1 week and in parallel there was an inhibition of the AA in the POA i13]. It has been shown in addition that individual levels of AA in the POA-anterior hypothalamus are significantly correlated in quail with individual levels of aggressive behavior [46].
The control by T of AA was also studied specifically in POM by analyzing the production of E2 in gonadally intact, castrated or T-treated male quail. POM was dissected by the Palkovits "punch" technique [47], incubated in the pres- ence of tritiated T, and the radioactive E2 produced was quantified by scintillation count- ing after a purification by phenolic partition and thin layer chromatography. This confirmed that within POM, AA is strongly influenced by circulating T levels. Castration produced a 4- to 5-fold decrease in enzyme activity and treatment with T restored the activity to levels similar to those seen in intact birds [48].
AROMATASE-ICC IN THE QUAIL BRAIN
These studies showed that aromatization of T was required for the steroid to exert its behav- ioral effects (in vivo studies) and in addition, that AA in the brain and specifically in the POA/POM was modulated by T in a manner consistent with the notion that this enzyme was a limiting factor in the action of T on behavior.
The measure of AA by product-formation assay, however, forces us to ignore erroneously the potential heterogeneity of the aromatase distribution in the brain. By combining this assay with the dissection of very small samples by the Palkovits "punch" technique, we had been able to show that AA activity is heteroge- neously distributed in the quail brain (higher in the iimbic than in non-limbic areas, higher in the POA than in the hypothalamus). The presence of heterogeneity within the POA was also demonstrated by the finding that AA in POM was 3-4 times higher than in the rest of the area [36]. This however did not provide any information on the distribution of aromatase at the cellular level.
Cellular heterogeneity cannot be ignored when studying the brain and the mechanism of behavior control because not all neurons in an area are involved in a particular behavior. If aromatase is present in a region, it is therefore
critical to know whether it is found in the neurons that are anatomically linked to the behavior or in other unrelated cells (see [49] for a more detailed discussion of this issue). The limitations of biochemical assays in tube do not permit to obtain this type of information.
A significant progress in the study of brain aromatase was therefore made with the discov- ery of an ICC procedure that permits the visual- ization of ARO-ir cells in the brain of the quail and of a few other avian species [50, 51]. This technique is based on a primary polyclonat antibody raised in rabbit against human placet~+ tal aromatase that had been purified based on the detection of the catalytic activity m the eluates from chromatography columns[52]. The visualization of the bound antibody is then performed by a classical peroxidase antiperoxidase (PAP) system.
A variety of biochemical, immunological+ neuroanatomical and physiological criteria strongly support the notion that this ICC pro- cedure detects the enzyme responsible for E2 formation in the quail brain. The antibody which has been purified by affinity chromatog- raphy appears to be monospecific and recog- nizes human aromatase as judged by classical biochemical and immunological criteria (inhi- bition of product formation in assays using human placental microsomes, Western blotting, Ouchterlony double diffusion analysis)[52] Classical ICC validations (ommission of pri- mary or secondary antibody, preabsorbtion ol the primary antibody with an excess of antigen) suggest that the immunoreactive rnaterial ob- served on the quail brain sections corresponds to aromatase [50]. The antiaromatase antibody specifically inhibits in a dose-dependent manner estrogen formation in the quail brain during in vitro product-formation assays but does not affect other T metabolizing enzymes [50j. Finally. ARO-ir cells are detected only in brain regions where AA had been previously measured by product-formation assay[35, 501 and the number of these ARO-ir cells varies m parallel with AA in specific brain regions as ~ function of the sex or hormonal condition of the birds [13, 50, 53, 54] (also see below).
NEUROANATOMICAL DISTRIBUTION OF ARO-ir MATERIAL IN THE QUAIL BRAIN
ARO-ir cells were detected by this method in four major areas of the quail brain [50, 511 (see Fig. 1): (a) in the POA, the5' were located

Aromatase and sexual behavior 525
D
Fig. 1. Neuronanatomical distribution of ARO-ir cells in the quail brain. (A) Rostral part of the POA. (B) Caudal part of the POA at the level of the anterior commissure. (C) Full extension of the bed nucleus striae terminalis. (D) Tuberal hypothalamus. Notice that in A and B, the ARO-ir cells precisely outline the cytoarchitectonic boundaries of the POM. These sections were taken from a male that had been treated for 4 days with the aromatase inhibitor, R76713 which provides a better visualization of immunoreactive
cells (see text). Magnification bar = 500 #m.
exclusively in the P O M and they specifically out l ined the edges o f this s t ructure t h roughou t its ros t ra l to cauda l extent [Fig. 1 (A and B)]. A R O - i r cells const i tu te an excellent neuro- chemical m a r k e r o f P O M ; (b) in the septal region under the la teral ventricles in an area
extending f rom the level o f the nucleus accum- bens at the ros t ra l end to the level o f the nucleus str iae te rminals a t the cauda l end [the Ac -nST region; Fig. 1 (B)]; (c) in a V-shaped a rea extending f rom the dorsa l aspect o f the nucleus paravent r icu la r i s to the cauda l aspects o f the

526 J. BALTHAZART and A, FOIDART
nucleus striae terminals [Fig. 1 (C)]. This region is actually an extension in the caudal direction of the POM and Ac-nST groups which merged just after the disappearance of the anterior commissure. This group seems to correspond to the full extent of the nucleus striae terminals and (d) in the tuberal region, where they form a diagonal band of labeled cells running from a dorso-medial to a ventro-lateral point at the level of the nucleus inferiors hypothalami [Fig. 1 (D)].
During the original experiments, ARO-ir ma- terial was essentially detected in the perikarya of cells and in their proximal dendrites. The cell nuclei were always devoid of immunoreactivity and they appeared completely clear on the sections [50, 51]. This distribution was fully con- sistent with the accepted subceilular distribution of AA established by product-formation assays performed on subcellular fractions prepared by differential centrifugation. In steroidogenic structures, aromatase was known as a micro- somal enzyme [55]. In addition, previous ICC studies of the placenta employing another poly- clonal aromatase antibody had shown that immunopositivity was found in the rough endo- plasmic reticulum throughout the cytoplasm but not in the nucleus [56].
The immunocytochemical studies on the quail brain aromatase were extended at the electron microscope level in collaboration with Drs F. Naftolin and C. Leranth (Department of Ob- stetrics and Gynecology, Yale University). A pre-embedding PAP staining technique was used to analyze the subcellular distribution of ARO-ir material in the quail POA [57, 58]. Immunopositivity was detected throughout the perikarya of neurons including the full extent of their cellular processes. Intranuclear immunore- activity was never seen. There were numerous pre-synaptic boutons that contained immuno- positive small clear synaptic vesicles. Many of these immunoreactive axon terminals formed synapses which were either axo-somatic or den- dritic. In most cases, these synapses contacted immunonegative profiles but occasionally im- munoreactive boutons formed synaptic contacts with immunostained dendrites or cell bodies. This observation of ARO-ir material in pre- synaptic boutons is consistent with recent bio- chemical studies on the rat and quail brain demonstrating the presence of AA in synaptoso- mal preparations [59, 60]. This suggests that es- trogens may be produced in the vicinity of the pre- and post-synaptic membranes and that they
could have local effects that would not require interaction with the classical estrogen receptors (see [61] for further discussion of this problem). This suspicion is further supported by our double label immunocytochemical studies on the anatomical relationships between ER and aromatase (see below).
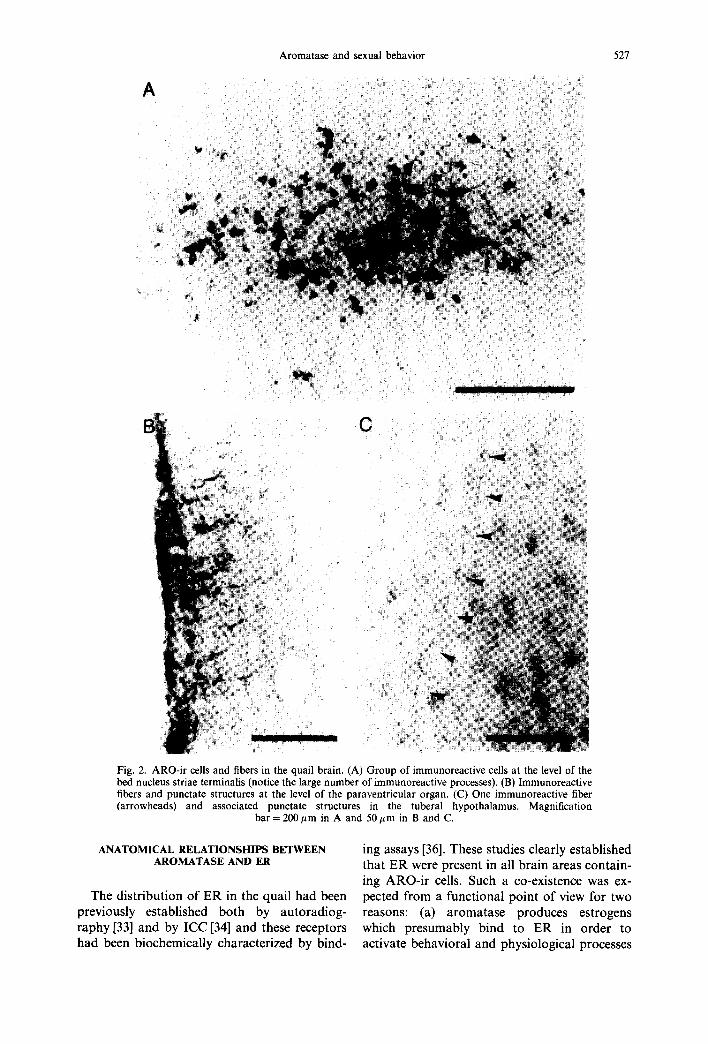
As mentioned above, ARO-ir material as observed in light microscopy was originally limited to the neuronal perikarya and their proximal dendrites. In the course of experiments designed to test the behavioral effects of aroma- tase inhibition, we observed however that the new non-steroidal aromatase inhibitor. R76713 [62, 63] strongly increases the aroma- tase-immunoreactivity in the quail brain [54]. The distribution of ARO-ir material was there- fore re-analyzed in the brain of quail that had been pretreated with R76713 and perfused with a mixture of paraformaldehyde (4%) and glu- taraldehyde (0.1%) in order to maintain an optimum morphology of the structures, lm- munoreactive cells surrounded by a dense plexus of immunoreactive fibers were observed in this way [see Fig. 2(A)]. Immunoreactive material was similarly detected in fiber-like structures associated with the third ventricle wall, especially at the level of the hypothalamic paraventricular organ. These possibly originate from tanycytes [Fig. 2(B)] but their exact ana- tomical nature remains to be elucidated. In addition, immunoreactive fibers associated with immunoreactive varicosities and punctuate structures were detected in brain regions that were often fairly distant from the closest im~ munoreacted perikarya [see Fig. 2(C)].
The mapping of these immunoreactive fibers is still in progress but it is already estab- lished that these can be consistently found in the periventricular region throughout most of the hypothalamus, in the tuberal hypothala- mus and in the mesencephalic central gray (Balthazart and Dupiereux, unpublished data). These fibers and punctate structures presumably represent the signature at the light level of the immunoreactive axons and pre-synaptic boutons that have been observed with the elec- tron microscope. They reveal that aromatase can be present in brain areas that are distinct from the regions containing the immuno-- reactive perikarya and produce estrogens in the vicinity of many different structures. The functional significance of this production of" estrogenic metabolites remains to be estab- lished.
A
i i!!~ ~I
!~ii~! ~ ?
Aromatase and sexual behavior 527
(3
Fig. 2. ARO-ir cells and fibers in the quail brain. (A) Group of immunoreactive cells at the level of the bed nucleus striae terminalis (notice the large number of immunoreactive processes). (B) Immunoreactive fibers and punctate structures at the level of the paraventricular organ. (C) One immunoreactive fiber (arrowheads) and associated punctate structures in the tuberal hypothalamus. Magnification
bar = 200/~m in A and 50 #m in B and C.
ANATOMICAL RELATIONSHIPS BETWEEN AROMATASE AND ER
The distribution of ER in the quail had been previously established both by autoradiog- raphy [33] and by ICC [34] and these receptors had been biochemically characterized by bind-
ing assays [36]. These studies clearly established that ER were present in all brain areas contain- ing ARO-ir cells. Such a co-existence was ex- pected from a functional point of view for two reasons: (a) aromatase produces estrogens which presumably bind to ER in order to activate behavioral and physiological processes

528 J. BALTHAZART and A. FOIDART
and (b) it is clear that estrogens, in birds, increase brain aromatase activity [23, 64].
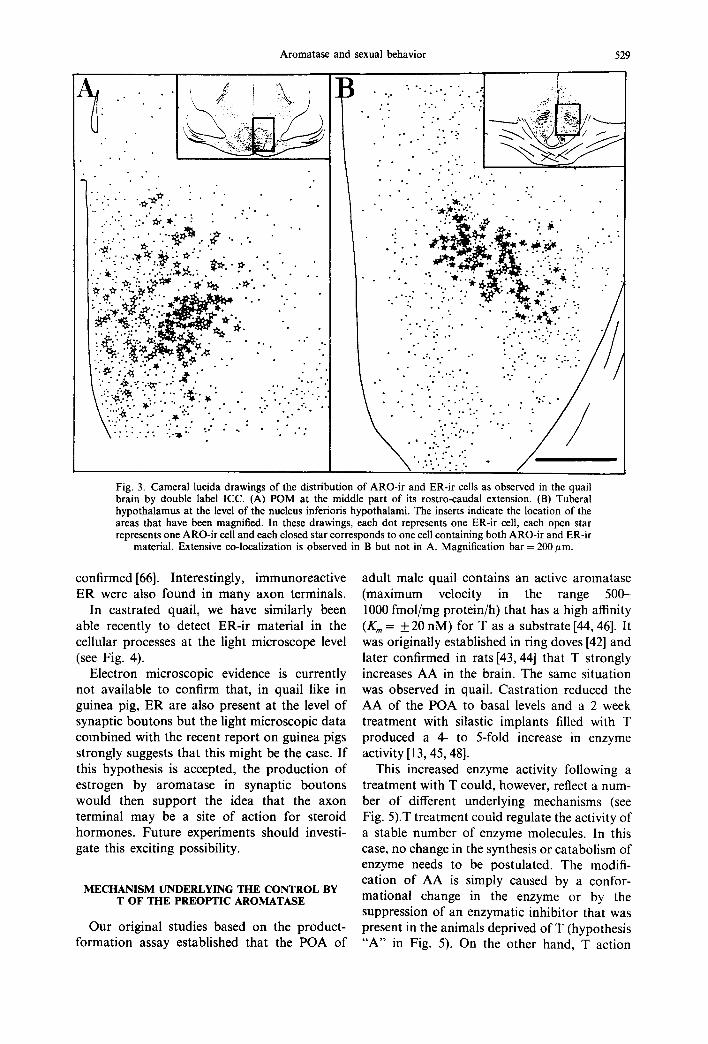
The study of the distribution of ARO-ir cells and of those containing ER immunoreactivity (ER-ir) however suggested that at a finer level of resolution, some differences in the distribution of these two antigens might be present [53]. In particular we found that in the POM, the distribution of these two markers was quite different. ER were not limited to POM. The highest density of ER-cells was observed in the medial portion of the nucleus (near the third ventricle). The density of these cells progress- ively decreased in the more lateral aspects of POM but ER-ir cells we still found lateral to the nucleus. By contrast, ARO-ir cells specifically outlined the borders of POM and were not found outside of its boundaries. The density of ARO-ir cells was higher in the lateral aspects of POM and lower in its medial part. Quantitative evaluations suggested that about 80-90% of the cells in POM contained ER-ir while only 10 15% of them were aromatase-immuno- reactive. Based on these data, it was therefore impossible to ascertain whether ARO-ir cells contained ER or not. A double-label ICC label- ling was therefore performed to answer this question [65].
Since the antiaromatase antibody had been raised in rabbit while antibody H222SP?, used to label ER was a rat monoclonal antibody, a double-label ICC procedure using a PAP system and diaminobenzidine as chromogen (brown reaction product) for the first sequence and an alkaline phosphatase system and fast blue as chromogen (blue reaction product) for the sec- ond sequence could easily be obtained. Unex- pectedly, we demonstrated in this way that a large proportion of the ARO-ir cells in the quail brain do not contain immunoreactive ER. Like in the previous studies using single label tech- niques, ARO-ir cells were found in the POM, in the septal region, in the nucleus striae terminals and in a large cell cluster extending from the dorso-lateral aspects of the ventromedial nu- cleus of the hypothalamus to the tuber at the level of the nucleus inferiors hypothalami. Im- munoreactive ER were also found in each of these brain areas but their distribution was much broader and included larger parts of the preoptic, septal and tuberal regions. In the ventromedial and tuberal hypothalamus, the majority of the ARO-ir cells (over 75%) also contained immunoreactive ER. By contrast, very few of the ARO-ir cells were double labeled
in the POA, in the septum and in the nucleus striae terminals. Less than 20% (17% in POM, 4% in the septum) of the aromatase-containing cells contained ER-ir in these regions (see Fig. 3).
Although this situation may have resulted from artifactual false negative results, the in+ ternal controls carried during the study suggest that this is probably not the case (see [65] for a more detailed discussion of this aspect). Fhese data therefore also suggest that the estrogens which are formed centrally by aromatization of testosterone might not exert their biological effects through binding with the classical nu- clear ER. The facts that significant amounts ot aromatase activity are found in synaptosomes purified by differential centrifugation[59, 60i and that aromatase immunoreactivity is ob. served at the electron microscope level in synap tic boutons or at the light microscopic level in fibers and punctate structures (see above) point to the possibility that aromatase might produce estrogens which act at the synaptic level ~t~, neurohormones or neuromodulators. It is oi course possible that estrogens formed in the pre-synaptic boutons diffuse to adjacent cells and bind, there, with nuclear ER but, at present, this interpretation does not seem to make much sense from a functional point of view.
It is also important to mention, in this con- text, that the subcellular distribution of ER is still controversial at present. Until recently it was commonly believed that the unoccupied ER are located in the cytoplasm of cells. However. based on ICC and cell fractionation studies, it was suggested in the mid-eighties that both occupied and unoccupied ER are essentially if not exclusively located in the cell nucleus (see [66] for additional detail). These ICC detec. tions of ER reporting exclusive localization in the cell nucleus used, however, relatively insensi- tive techniques. By increasing the sensitivit~ of the ICC procedure, namely through the use of the multiple bridge technique, Blaustein and collaborators were able to detect ER-ir m the cytoplasmic processes of many neurons in owtri- ectomized guinea pigs [67] and similar obser- vations have now been reported for many other mammalian species (see[66] for details). By combining electron microscopy with a pro++ embedding immunostaining of" ER, the presence of ER-ir in the cell nucleus but also in proximal and distal dendrites and in the rough endo- plasmic reticulum of neurons in the brain of ovariectomized guinea pigs was recent!~
Aromatase and sexual behavior 529
.1 , 1
t . . . • • . , • • • • • ~ . ,
.• o . + • • ...'..~ .::..'~.,,.~ . : • . .
+ • " . ! ? . . i . . ~ . " ~ " + ' . " ' ~ . . • •
• ' " " " . ~ I ( ~ ' ' ~ " "" " • " • . + . . . "- : . . . . . . ~ . : . ~ . ~ . . . . . ." . . . •
i ; ~ ! + ~ ' * ~ . ' ~ ' ~ - '+ ' . " " " • ' . . . " • . " "
:.-•'- ~_ "~e..-~." • . .
- " - .
~......:...'~ --~." ~ . ~ . . : . • ... . . "." . . ..
i ~ - ~ : : i ~ ; . . . . . + ' . . . : \ : i :~i '- .*. " : : " ' . ' " ~ ' : , " ." . . ' " - " ' : ."
o o
• .• " ' - : - . : - . --'. i::.ii[:,: .i~:: " " ' • • ' • " ...... ~ + . _ L .
• • • / i • " " " "" - - - - - - - . . . . ~ \ ' ~ . ~ ' ~ - ~ + ~ S ' ~ , '
- .
.° . • " . ' .
• , " • . • • . . . ' • . , " ,
. . . " • + •
• ° • . • , . ' ~ + o ° . ' . • • • . • . o
• :..: , ~ , ~ . ~ . " : : : - . : ~ • • ... - . . - . : / . . . , . ~ . : :. . . "+~/ .. • ._
• " . . . - ~ , ~ . ~ : " - ~ , . a ~ . . . : . -
. . .-.: .:-.i-. .: /
!i !!ill Fig. 3. Cameral lucida drawings of the distribution of ARO-ir and ER-ir cells as observed in the quail brain by double label ICC. (A) POM at the middle part of its rostro-caudal extension• (B) Tuberal hypothalamus at the level of the nucleus inferioris hypothalami. The inserts indicate the location of the areas that have been magnified• In these drawings, each dot represents one ER-ir cell, each open star represents one ARO-ir cell and each closed star corresponds to one cell containing both ARO-ir and ER-ir
material• Extensive co-localization is observed in B but not in A. Magnification bar = 200/~m.
confirmed[66]. Interestingly, immunoreact ive E R were also found in many axon terminals.
In castrated quail, we have similarly been able recently to detect ER-ir material in the cellular processes at the light microscope level (see Fig. 4).
Electron microscopic evidence is currently not available to confirm that, in quail like in guinea pig, E R are also present at the level o f synaptic bou tons but the light microscopic data combined with the recent report on guinea pigs strongly suggests that this might be the case. I f this hypothesis is accepted, the product ion o f estrogen by aromatase in synaptic boutons would then suppor t the idea that the axon terminal may be a site o f action for steroid hormones . Future experiments should investi- gate this exciting possibility.
M E C H A N I S M U N D E R L Y I N G T H E C O N T R O L B Y
T O F T H E P R E O P T I C A R O M A T A S E
Our original studies based on the product- format ion assay established that the P O A of
adult male quail contains an active aromatase (maximum velocity in the range 500- 1000 fmol/mg protein/h) that has a high affinity (Kin = _ 20 nM) for T as a substrate [44, 46]. It was originally established in ring doves [42] and later confirmed in rats [43, 44] that T strongly increases A A in the brain. The same situation was observed in quail• Castrat ion reduced the A A of the P O A to basal levels and a 2 week treatment with silastic implants filled with T produced a 4- to 5-fold increase in enzyme activity [13, 45, 48].
This increased enzyme activity following a treatment with T could, however, reflect a num- ber o f different underlying mechanisms (see Fig. 5).T treatment could regulate the activity o f a stable number o f enzyme molecules• In this case, no change in the synthesis or catabolism of enzyme needs to be postulated• The modifi- cation o f A A is simply caused by a confor- mational change in the enzyme or by the suppression of an enzymatic inhibitor that was present in the animals deprived of T (hypothesis " A " in Fig. 5). On the other hand, T action

530 J. BALTHAZART and A. FOIDART
Fig. 4. Microphotograph illustrating the cellular distribution of estrogen receptors (ER) in the hypothala- mus of castrated male quail. ER were detected by ICC using antibody H222SP 7. The multiple bridge technique was applied to increase the sensivity of the detection. Immunoreactive material is essentially located in the cell nuclei but cytoplasmic staining is clearly visible in many perikarya and cell processes.
Magnification bar = 50 ~m,
could result from a modification of aromatase concentration (presumably reflecting a de vovo synthesis). The concentration of aromatase in the positive cells could be increased (hypothesis "B") or the number of aromatase-containing cells could be changed (hypothesis "C" ) or both (hypothesis "D") .
Detailed kinetic experiments carried out in quail and in doves suggested that the first of these hypotheses (A) was not correct. By incu- bating equal amounts of enzyme with increasing concentrations of the substrate, it was indeed shown that an in vivo treatment with T increases the maximum velocity of the aromatase (Vmax) but does not change the affinity of the enzyme for its substrate [42, 69, 70]. In fact it has been shown in quail that the changes in AA related to the age, sex or hormonal condition (e.g. castrate versus hormone treated castrate) are always obtained without any significant modifi- cation of the Km of the enzyme [45, 70.71]. It is very likely that a regulation of aromatase due to conformational changes would produce a modification in enzyme affinity. On the other hand, amongst the three major types of enzyme inhibition (competitive, non-competitive and uncompetitive), only one (the non-competitive) involves no change in the apparent Km. If T was regulating the AA through a change in the concentration of an enzymatic inhibitor, this inhibitor should therefore be of the non-
competitive type (competitive or uncompetitive inhibitors would change the apparent K,,~) These kinetic data were therefore only compat- ible with an action of T mediated by either an increase of aromatase concentration or a repres- sion of a non-competitive enzyme inhibitor. A similar conclusion was reached independently for the ring dove[69] and it was therefore impossible to discriminate between these two possible interpretations.
Additional kinetic experiments were carried out in quail to collect data relevant to this question. It was reasoned that if an enzyme inhibitor was present in birds having a low AA (castrates vs T-treated birds, females vs males or young chicks vs adults [45, 70, 71]) then the en- zymatic velocity measured in a mixture of brains with high and low activities would be lower than the arithmetic mean of the activities measured separately. Alternatively, if the difference in velocities resulted only from a difference in enzyme concentration, then the velocity in ~ pool of brains with low and high activity would be the arithmetic mean of the velocities measured in separate incubations. All data col- lected during these experiments were consistent with the later interpretation (see [72] for example). However the nature of these exper- iments always made it difficult to make absol- utely sure that no inhibitor was present: the observed velocity in the pool of brains was

Aromatase and sexual behavior 531
Fig. 5. Schematic illustration of the mechanisms that could potentially explain an increase in AA in the POM following a treatment with T. At the top, the situation in the POM (ovoid nucleus) of a castrated male is shown: a few cells contain few aromatase molecules ([D. Aromatase activity can be increased by T without any change in concentration of the enzyme [regulation of the activity in the molecules that are already present; (A)] or by an increase in concen- tration of the enzyme in a constant number of cells (B) or by an increase in the number of cells expressing the enzyme
(C) or by a combination of these two mechanisms (D).
645% as compared to untreated castrates (see Fig. 6).
The number o f ARO- i r cells was also counted in 4 adjacent sections that had been collected in the medial par t o f the P O A in the second subset o f birds that had received the same hormona l treatments. All ARO- i r cells were located within the cytoarchitectonic boundaries o f the POM. These were very rare in castrates but their number was about 5 times higher in T-treated males. Given that the classification o f a cell as immunoposi t ive or immunonegat ive largely de- pends on the concentra t ion o f the antigen in this cell, these data strongly suggested that T in- creases aromatase concentra t ion in the POA. Since the percentage increase in the number o f ARO- i r cells and in A A were relatively similar, this also suggested that the increased enzyme concentra t ion was fully responsible for the ob- served change in AA. It must however be ac- cepted that this conclusion had two significant limitations. First ICC is certainly not a tech- nique o f choice for quantitative studies and the fact that the number o f ARO- i r cells shows a 5-fold increase after t reatment with T does not necessarily mean that the antigen concentra t ion has been multiplied by five. Second, the in- creased immunoreact ivi ty in T-treated birds may be due not to an increase in antigen concentra t ion but rather to a conformat ional change (e.g. phosphoryla t ion) in a constant amoun t o f protein which would then enhance its immunoreactivity. This interpretation is some- how unlikely since this conformat ional change
sometimes slightly different f rom the calculated value and it was difficult to decide whether this only reflected experimental errors or true en- zyme inhibition.
The availability o f an ICC procedure to visu- alize aromatase in the quail brain provided us with a mean to evaluate, in a semi-quantitative manner , the concentra t ion o f the enzyme and to study whether T was able to modula te this concentrat ion. Dur ing a first experiment, the brain o f castrated males and of castrates that had been treated with T-filled silastic implants (40 m m total length) were studied in parallel by in vitro produc t - fo rmat ion assay to measure A A and by ICC to visualize the immunoreact ive aromatase and determine the number o f ARO- i r cells in the P O A [50]. After a 5 day exposure to physiological levels o f T as provided by the silastic implants, the A A had increased by about
800
Effect of T on preoptic aromatase
600 ¢d
..~ 400
e~ ¢D
200 gh
0 AA ARO.ir cells A R O m R N A
Fig. 6. Effect of testosterone treatment in castrated quail on AA, on the number of ARO-ir cells and on the concen- tration of the aromatase mRNA in the quail POA. In each experiment 7-week-old castrated quail were exposed for 5 days to silastic capsules (40 mm in length) filled with T (CX + T) or left empty (CX) as control. All data are expressed as percent of values obtained in CX birds (AA): 226.5 + I. 1 fmol/mg protein/h; number of ARO-ir cells in 4 sections: 229 + 95 and concentration of aromatase mRNA:
0.270 + 0.078 amol/#g RNA).

532 J. BALFHAZART and A. FOIDART
should increase the activity of the enzyme with- out changing its affinity for the substrate (Kin), a pair of conditions that is unlikely to be fulfilled simultaneously.
More recently, additional evidence for a T- induced change in AA caused by variations in the enzyme concentration has been obtained by measuring the concentration of the aromatase mRNA in the quail POA [73]. The aromatase cDNA was first identified by homologies with the human aromatase and isolated from a brain cDNA library of Japanese quail. A quantitative PCR method of RNA transcripts was then developed to permit the determination of trace amounts of aromatase mRNA in the brain. The total brain RNA was isolated, subjected to reverse transcription and then quantified by PCR from cDNA with fluorescent primers. A modified aromatase mRNA was used as internal standard to calibrate quantitative measures. This newly developed method revealed that in agreement with previous product-formation as- says and ICC results, the brain aromatase was primarily transcribed in the POA-hypothalamus with negligible traces of transcript being present in the rest of the brain. It was also demonstrated that the concentration of aromatase mRNA was low in castrates by comparison with intact sexually mature males but was increased 4-fold following a 5-day exposure to physiological levels of T produced by subcutaneous silastic implants (see Fig. 6).
These results therefore demonstrate that the control by T of AA takes place at least in part at the pre-translational level. Although these data do not permit to exclude a regulation based on an increase in the stability of the aromatase mRNA, it is extremely likely that T enhances the transcription of the aromatase messenger. Data summarized in Fig. 6 clearly indicate that a same treatment with T (5-day exposure to 40mm silastic implants) produces changes in AA, in the number of ARO-ir cells and in the concentration of aromatase mRNA that are in the same order of magnitude. The observed percentage increase in AA was however slightly higher (645%) than the increase in the number of immunoreactive cells (497%) which was itself larger than the increase in the concentration of ARO mRNA (372%). These differences may of course reflect experimental errors finding their origin, for example, in the different techniques employed, in individual differences (different groups of birds had obviously to be used for the different measures) or in subtle neuroanatomi-
cal differences (AA and ARO mRNA were measured on the preoptic-anterior hypothala- mic block obtained by microdissections while the number of ARO-ir cells was evaluated in a sub-sample of POM). Alternatively, it is interesting to notice that the percentage increase plotted in Fig. 6 becomes higher when the dependent variable is functionally more distant from the original DNA expression. Based on this trend, it can therefore be hypothesized that besides regulating aromatase transcription (372% increase). T also enhances the translation of the ARO mRNA (this justify- ing the difference between 372 and 495%) or even modulates the activity of the aromatase enzyme (difference between 495 and 645%t. With the available evidence, it is however clem that the magnitude of these effects is small compared to the pre-translational regulation which explains about two thirds of the variation in AA.
ANDROGENS VERSUS ESTROGENS IN TI-tE C O N T R O L OF A R O M A T A S E
T is extensively metabolized in the brain of all vertebrate species that have been studied so far and many effects of T in the brain are actually caused by the action of androgenic or estrogenic metabolites of this steroid (see [6] for review). This conclusion also applies to quail. Numerous studies have shown that in the quail brain, T can be transformed into estrogens (e.g. E2) or an- drogens (e.g. 5e-DHT) and these steroids, alone or in combination, activate male copulatory behavior. This conclusion is supported by a host of experiments in which the effects on behavior of aromatizable or non-aromatizable andro- gens, of estrogens, of aromatase inhibitors, of antiandrogens or antiestrogens were tested (see above). Because the induction by T of AA in the POA appears to be a limiting step in the pro- duction of the behavioral effects of T [13], we researched whether the induction of AA dis- played the same hormonal specificity as the activation of copulatory behavior [23].
Castrated male quail were injected with '1- (1 mg/day), or the synthetic non-aromatizable androgen R1881 (methyltrienolone, 1 rag/day), the synthetic estrogen, diethylstilbestrol (DES, 200 #g/day) or received both compounds simul- taneously (R1881 +D ES ) . All treatments in- creased AA activity in the POA however those providing both androgenic and estrogenic stimulation (T or R1881 + D E S ) were clearly

Aromatase and sexual behavior 533
more efficient than the androgens or estrogens in isolation [see Fig. 7(A)].
In fact, the combined treatment with R1881 + DES was the only one which signifi- cantly increased AA by comparison with cas- trates (the dose of T used in this experiment was rather low so that it only had marginal effects). These data clearly demonstrated the synergism between both classes of steroids in the induction of AA but they provided little information on the possible anatomical specificity of these effects. In addition, the assay data did not allow to discriminate between increases in enzyme activity that would be caused by the regulation in the activity of a constant amount of enzyme from changes due to a real enzymatic induction (increase in enzyme concentration). All these questions were studied more recently by the ICC approach.
In a first study [54], we attempted to disasso- ciate the effects of androgens and estrogens in aromatase induction by comparing ARO-ir neurons of quail treated with T alone or T in the presence of a potent aromatase inhibitor
A Effects of synthetic androgens or estrogens on aromatase
200'
~. lOO
B
3OO
"9 200
o
CX CX+T CX+RI881 CX+DES CX+R18$1 +DES
Effects of androgens and estrogens on ARO-ir cells in P O M
CX CX+T CX+DHT CX+E2 CX+DHT+E2
Fig. 7. Effects of androgen and estrogens on the preoptic aromatase in quail. (A) Effects o f treatment with synthetic androgens and estrogens on the AA measured by in vitro product-formation assay. (B) Effect o f D H T and/or E2 on the number of ARO-ir cells in the POM (see text for details).
(R76713), which had been shown to depress AA levels and to suppress T-activated copulatory behavior [14]. As expected based on our pre- vious work, T activated an intense sexual behav- ior in this experiment and this effect was almost completely blocked by the concurrent adminis- tration of the aromatase inhibitor. In parallel, T increased the number of ARO-ir cells in POA, bed nucleus striae terminals (BNST) and tuberal hypothalamus (Tu). The T effect was inhibited by the concurrent treatment with R76713 in Tu, but not in POA and BNST. This differential effect of the aromatase inhibitor fitted in very well with our previous studies of the co-localiz- ation of aromatase and estrogen receptors [65]. The T effect was blocked by R76713 in areas where ARO-ir and ER-ir were generally co- localized (Tu) and was not affected in areas with mainly ARO-ir positive, ER-ir negative cells (POA, BNST) (see above). Taken together, these data suggested that estrogens were playing a major role for aromatase induction in the tuberal but not in the rostral hypothalamus and POA. This also pointed to possible anatomical differences in the expression or clearance of aromatase which could be differentially sensitive to androgens and estrogens depending on the brain area considered.
In a second experiment, the relative role of androgens and estrogens in the induction of aromatase was studied in a more direct way by counting ARO-ir cells in the brain of castrated quail that had been treated with silastic implants filled with T, E2, DHT or DHT + E2. Two weeks after the implantation of the silastic capsules, birds were tested for copulatory be- havior. As expected, high levels of male sexual behavior were activated by T and by the com- bined DHT + E2 treatment. Moderate behav- ioral effects only were observed in birds that received DHT or E2 alone. The numbers of ARO-ir cells were then counted in two sections through the POM and in two sections through the Tu. This demonstrated in each case an extremely significant effect of the treatments on the numbers of immunoreactive cells (P < 0.0001). In each area, the treatment with either T or DHT + E2 produced a major in- crease in the number of labeled cells. DHT alone had no significant effect at this level while E2 produced a significant increase in three out of the four sections that were counted [see Fig. 7 (B) for representative data in the anterior POM]. The increase observed in D H T + E2 birds always had a larger magnitude than in E2

534 J. BALTHAZART and A. FOIDART
birds but this difference only reached signifi- cance in the Tu and not in the POM.
The data of this second experiment therefore confirmed the existence of a synergism between androgenic and estrogenic metabolites of T in the activation of copulatory behavior. They also suggested that estrogens could play a major role in the induction of ARO-ir cells in both the POA and the Tu, D H T alone had little or no effect but this androgen seemed to synergize with E2 to enhance aromatase immunostaining in all brain areas although this synergism was especially prominent in the tuber.
These two ICC experiments therefore suggest that estrogens and androgens are able to in- crease the number of ARO-ir cells in the quail brain so that the increase in AA that has been described in birds exposed to treatments with these steroids presumably results from a change in aromatase concentration rather than fi'om the modulation of the activity in a constant amount of enzyme. They do not provide however a clear picture of the respective role of androgens and estrogens in this process since the two exper- iments lead to fairly different conclusions. Such a discrepancy was already present in the litera- ture devoted to the control of AA by androgens and estrogens. In ring doves, it has been suggested that the induction by T of the preoptic AA results mainly if not exclusively from the action of the estrogenic metabolites of this steroid. It was shown that the synthetic estro- gen, DES strongly increased the enzyme activity in castrated doves while non-aromatizable an- drogens such as 5~-DHT or R1881 had no detectable effects at this level [64, 74]. These two non-aromatizable androgens, however, signifi- cantly increased AA in the anterior hypothala- mus [74]. In rat, by contrast, the major role in aromatase induction appears to be played by androgens. Both T and 5~-DHT stimulate AA in the rat POA and the non-steroidal antiandro- gen, flutamide, strongly reduces the induction of AA by T in castrated males [43, 75]. In addition, rats that are genetically deficient for androgen receptors show in parallel a reduced level of AA[76]. In a recent study, it was however demonstrated that estradiol benzoate (EB), even if it has no detectable effect by itself, synergizes with 5~-DHT to enhance AA in the rat POA (but not in the hypothalamus)[77]. These data suggest that there are fundamental differences in the regulation of brain AA according to the species and maybe the brain region con- sidered (see [77] for additional discussion on
this aspect) but that a model based on a syner- gism between androgens and estrogens with a major role played by one or the other com- pound depending on the species and/or the brain area should be able to integrate most if not all data.
The product-formation assays also suggested a synergism between androgens and estrogens in the control of AA in quail. By ICC, we showed that the inhibition of aromatase decreases the induction of ARO-ir cells in the tuber but not in the POA suggesting that estrogens are more important for this process in the latter than in the former area. By contrast, the treatment with natural steroids points to a key role fbr estrogens acting possibly in synergy with andro- gens. Several factors could contribute to explain this discrepancy. It can be assumed that the aromatase inhibition induced by R76713 in the POA, if sufficient to block behavioral acti~ vation, was not complete and therefore that minimal amounts of estrogens were still pro- duced and could, in synergy with the androgenic metabolites of T, induce new ARO-ir cells. Alternatively, it is well established that E2 binds to some extent with the androgen receptor [78] and this cross-reaction may confound the in- terpretation of the aromatase induction ob- served after E2 treatment. The lack of effect of 5~-DHT could also be due to the fact that this steroid is very rapidly metabolized in the quail brain [18] as well as in mammals [79-81] which could prevent its accumulation in sufficient con- centration to induce a clear effect. These hy- potheses are currently tested by studying the effects on ARO-ir cells of the synthetic estrogen, DES that does not bind to the androgen recep- tor [78] and of the highly potent synthetic an- drogen, R1881.
Independent of the results of these additional studies, it is already clear that both androgens and estrogens, when acting in synergy, contrib- ute to the induction of AA in the quail brain and that the increased enzyme activity reflects an increased enzyme concentration as revealed by the ICC studies. A more recent experiment also demonstrates that 5~-DHT and E2 synergize to increase the ARO mRNA in the quail brain which provides further support to the notion that these steroids actually regulate the enzyme concentration [82].
The mechanism underlying this synergism is nevertheless unclear. Given that steroids regu- late the actual concentration of aromatase, it could be suspected, on a priori bases, that they

Aromatase and sexual behavior 535
act at the genomic level after binding to their classical nuclear receptors. Such an interpret- ation is, in fact, supported by the observation that in rats, the androgen receptor antagonist (antiandrogen), flutamide strongly inhibits the induction of AA by T [43]. The data available concerning the distribution of estrogen and androgen receptors in the quail brain, in par- ticular in the POA, bring, however, little sup- port to this speculation. Autoradiographic and ICC studies indicate that a large proportion of cells in the POA and in POM in particular contain ER [33, 34]. However, we showed by double label ICC (see above) that only a very small fraction (< 20%) of the ARO-ir cells in POM contain immunoreactive ER [65].
On the other hand, if autoradiography reveals a large number of cells accumulating tritiated T in the POA, very few of them (in fact almost none) assumulate tritiated 5~-DHT which leads to the conclusion that these cells actually con- tain ER (that bind E2 derived from T aroma- tization) but no androgen receptors [33]. We recently identified an ICC procedure that per- mits the visualization of androgen receptors in the avian brain. A high density of androgen receptor-immunoreactive cells was identified by this technique in the quail nucleus intercollicu- laris (which also accumulates tritiated 5~-DHT) but, in agreement with autoradiographic data, very few androgen receptor immunoreactive cells were identified in the POA[83]. This suggests that presumably the majority of the ARO-ir cells do not contain at the same time androgen receptors. This apparent lack of co- localization between aromatase and androgen or estrogen receptors may result from technical artifacts associated with the specific techniques used to visualize these receptors (false negative results). However given the number of positive controls that were included in these studies, this interpretation appears somehow unlikely (see [65, 83] for additional discussion). It must also be considered that if androgens and estro- gens synergize in the same cells to induce aromatase synthesis, both types of steroid recep- tors must be co-localized with aromatase and this situation appears even less likely to occur. No study has analyzed at the cellular level the relative distribution of androgen and estrogen receptors but in the canary hypothalamus, it has been shown that only a small percentage (< 20%) of the ER-ir cells accumulate tritiated 5~-DHT and no such cell could be found in the POA [84].
It seems therefore possible that the cells which produce aromatase are not the same as those that bind steroids. Transynaptic mechanisms of regulation for aromatase synthesis and/or ac- tivity need therefore to be contemplated. This idea is further supported by a recent study analyzing steroid control of AA in preoptic- hypothalamic blocks maintained in organotypic culture. These experiments showed that, in rats, regulations of AA normally taking place in vivo are not observed when the POA-hypothalamus is disconnected from adjacent tissue and main- tained in vitro [85]. In particular, no increase in AA could be observed in tissue blocks cultivated for 2 days in presence of T, 5~-DHT or E2. Similarly, we haved consistently failed to ob- serve any increase of the AA after treatment with T of embryonic quail POA-hypothalamus maintained in organotypic or dissociated in vitro cultures (Balthazart-Raze and Balthazart, un- published data).
It must nevertheless be reminded that in the live animal, T induces aromatase, at least in part, through a direct action in or around POA because stereotaxic implants of T aimed at this region increase AA in doves [86] and in- crease the number of ARO-ir cells in quail [40]. These results do not however exclude the contri- bution of multicellular transynaptic regulations possibly including extrahypothalamic inputs. Recent anatomical and pharmacological studies from our and other laboratories suggest that the catecholaminergic system may be part of neuroanatomical network regulating brain aromatase.
CATECHOLAMINES AND THE CONTROL OF AROMATASE
Evidence from different biological systems suggest that neurotransmitters are able to modulate the intracellular steroid metabolism and in particular aromatase. Two receptors subtypes for dopamine (DA) are classically rec- ognized (D1, D2) and have been characterized on the basis of their pharmacological and physiological properties [87]. The D2 subtype has now been divided into a pre- and a post- synaptic receptor. Additional subtypes (D3, D4 and D5) are present in the brain as recently demonstrated by molecular cloning and their pharmacological and functional characteriz- ation is currently in progress [88-90].
The D1 dopaminergic receptor subtype is classically linked to the enzyme adenylate
SB 44-4/6~p

536 J. BALTHAZART and A. FOIDART
cyclase by a stimulatory G-protein (Gs). Occu- pation of this receptor results in an increased intracellular cAMP production [87, 90]. This second messenger has been identified as a chemical regulator of AA in several tissues including the brain [91-94]. Since the cAMP- induced increase in AA only develops gradually over a 2- to 4-day period[91,94], it can be assumed that it results from an increase in enzyme concentration rather than from a modu- lation in the activity of a constant number of molecules. This fits in well with the fact that cAMP is known to modulate gene expression through the phosphorylation of transcriptional activator proteins [90] (for review in a behav- ioral endocrinology context see [95]). It is there- fore very tempting to hypothesize that DAergic activity significantly affects AA in the brain through the modulation of adenylate cyclase by the DI receptor subtype.
On the other hand, several in ritro prep- arations using tissue derived from the pineal or the testis, respectively point to a role for norepi- nephrine (NE) in the regulation of T metaboliz- ing enzymes. Data from the pineal studies show that stimulation of NE increases AA and de- creases 5c¢-reductase[92]. Similar stimulatory effects were observed for aromatase in the testis derived preparation [93, 96]. These effects seem to result from an interaction with fl-adrenergic receptors. In contrast, studies in fetal and neo- natal rat hypothalamus suggest that alpha and/or beta adrenergic stimulation might inhibit aromatization [97-100]. In support of this ob- servation, it has also been reported that in rats during ontogeny, the developmental decline of AA is paralleled by an increase in catechol- amine levels in the brain [97]. It appears that this inhibition acts through a modification of adenylate cyclase activity, the second messen- ger associated with many catecholamine recep- tors.
In quail, a few preliminary pharmacological studies also suggest that DA and/or NE may regulate AA. The injection of the noradrenergic neurotoxin, DSP4 which significantly depletes NE levels in the POA enhances the induction by T of AA in the POA of T-treated male quail [101]. This pharmacological treatment in- creases at the same time the activating effects of T on copulatory behavior [101,102] and it can be speculated that this behavioral effect is in- deed mediated by the change in AA.
On the other hand, a chronic treatment with the tyrosine hydroxylase inhibitor, a-methyl-
para tyrosine (teMPT) has been shown to de- plete DA and to a lesser extent NE levels in the quail brain [103, 104]. We have shown that such a treatment also slightly decreases (34%) AA in the POA (Balthazart, unpublished data). Since a chronic treatment with c¢-MPT decreases both NE (which presumably inhibits AA in the quail POA, see above) and DA, it can be expected that a selective DAergic inhibition would lead to a larger decrease in enzymatic activity. Pre- viously published data and these preliminary results are therefore consistent with the notion that DA regulates brain aromatase. Recent neu- roanatomical data give additional support to this idea.
M O R P H O L O G I C A L INTERACTIONS BETWEEN C A T E C H O L A M I N E R G I C AND THE STEROID-
SENSITIVE SYSTEMS
The availability of an ICC procedure to visu- alize ARO-ir cells [50, 51] has recently allowed us to study at the cellular level the anatomical relationships between the aromatase-containing cells and the catecholaminergic system charac- terized by the synthesizing enzymes, tyrosine hydroxylase (TH) and dopamine fl-hydroxyl- ase[105]. In sections stained by double label ICC TH-ir fibers and punctate structures were found in all areas containing ARO~ir cells and especially in the POM and in the septal region around the nucleus accumbens-bed nucleus striae terminals (Ac-nST complex). In these two areas, a close association between TH-ir fibers and ARO-ir cells was observed. Many ARO-ir cells were entirely surrounded by TH-ir fibers and punctate structures forming a sort of basket around the aromatase celt. This anatomical association was especially frequent in the Ac- nST complex. These basket-like DA projections surrounding ARO-ir cells could represent a morphological substrate for the regulation of aromatase activity by DA that is discussed above. This could also be one of the ways through which DA controls male sex behavior. Similar anatomical structures were found in sections double-labeled for aromatase and DBH. Some of the ARO-ir cells in the POA, hypothalamus and in the Ac-nST complex were closely associated with DBH-ir fibers and punc- tate structures. Some of the cells were also completely surrounded by these DBH-ir struc- tures.
These data suggest that catecholaminergic projections make close contact (synapse?) with

Aromatase and sexual behavior 537
ARO- i r cells and these connect ions may rep-
resent the ana tomica l substrate for the regu- lat ion of AA and aromatase concent ra t ion by
D A and /o r NE. Fur ther pharmacological studies should be carried out to test this possi- bility.
CONCLUSION
It is now well established that the aromatiza- t ion of T represents a l imiting step in the act ion
of this steroid on behavior. T aromat iza t ion takes place in the sexually d imorphic P O M and the locally produced estrogens also act in this nucleus to activate behavior. Product - format ion assays have shown that aromatase activity in the POA is regulated by steroids in a m a n n e r con- sistent with the l imiting role played by the enzyme in the act ivat ion of behavior. A recently developed ICC procedure allows the visualiza- t ion of ARO- i r cells in the quail bra in so that
the dis t r ibut ion and regulat ion of this enzyme can now be studied at a cellular level of resol- ut ion. The preoptic ARO- i r cells are specifically
located in the POM and outl ine its cytoarchitec- tonic boundaries . The control by T or AA
implies a true enzymatic induct ion (de novo
synthesis of enzyme) since the concent ra t ion of aromatase increases after T t rea tment as well as the concent ra t ion of the aromatase m R N A . These regulat ions involve both androgenic and estrogenic metaboli tes of the steroid but the exact con t r ibu t ion of each type of metab- olite remains to be established. This regulat ion may in par t implicate t ransynapt ic control mechanisms and, based on neuroana tomica l and prel iminary pharmacological data, it ap- pears that catecholamines may be involved at this level.
Acknowledgements--The recent work described in this re- view was supported by grants from the Belgian FRFC (Nbr. 2.9003.91 and 9.4601.90) and the University of Liege (Fonds Sp6ciaux pour la Recherche). I thank Professor E. Schoffe- niels for his continued interest and support of my research. I also would like to thank all my collaborators in Li6ge and in other laboratories who have been of an invaluable help in collecting the data presented here, namely C. Surlemont, M. Schumacher, A. De Clerck, L. Evrard and T. Bailhache (Laboratory of General and Comparative Biochemistry, University of Liege), F. Naftolin and C. Leranth (Depart- ment of Obstetrics and Gynecology, Yale University), G. C. Panzica, C. Viglietti-Panzica and N. Aste (Department of Human Anatomy and Physiology, University of Turin), N. Harada (Molecular Genetics, Fujita Health University, Toyoake) and G. F. Ball (Department of Psychology, Johns Hopkins University). Collaboration with G. F. Ball was supported by a NATO Collaborative Research Grant (CRG910526).
REFERENCES
1. Naftolin F., Ryan K. J. and Petro Z.: Aromatization of androstenedione by the anterior hypothalamus of adult male and female rats. Endocrinology 90 (1972) 295-298.
2. Naftolin F., Ryan K. J., Davies I. J., Reddy V. V., Flores F., Petro Z., Kuhn M., White R. J., Takaoka Y. and Wolin L.: The formation of estrogens by central neuroendocrine tissues. Recent Prog. Horm. Res. 31 (1975) 295-319.
3. Callard G. V., Petro Z. and Ryan K. J.: Conversion of androgen to estrogen and other steroids in the ver- tebrate brain. Am. Zoo. 18 (1978) 511-523.
4. Callard G. V., Petro Z. and Ryan K. J.: Phylogenetic distribution of aromatase and other androgen- converting enzymes in the central nervous system. Endocrinology 103 (1978) 2283-2290.
5. Callard G. V.: Aromatization in brain and pituitary: an evolutionary perspective. In Metabolism of Hor- monal Steroids in the Neuroendocrine Structures (Ed- ited by F. Celotti, F. Naftolin and L. Martini) Raven Press, New York (1984) pp. 79-102.
6. Balthazart J.: Steroid metabolism and the activation of social behavior. In Advances in Comparative and En- vironmental Physiology, Vol 3 (Edited by J. Balthazart) Springer Verlag, Berlin (1989) pp. 105-159.
7. Yahr P.: Data and hypotheses in tales of dihy- drotestosterone. Horm. Behav. 13 (1979) 92-96.
8. McEwen, B. S.: Neural gonadal steroid actions, Sci- ence 211 (1981) 1303-1311.
9. Adkins E. K. and Pniewski E. E.: Control of repoduc- tive behavior by sex steroids in male quail. J. Comp. Physiol. Psychol. 92 (1978) 1169-1178.
10. Adkins E. K., Bop J. J., Koutnik D. L., Morris J. B. and Pniewski E. E.: Further evidence that androgen aromatization is essential for the activation of copu- lation in male quail. Physiol. Behav. 24 (1980) 441- 446.
11. Schumacher M. and Balthazart J.: The effects of testosterone and its metabolites on sexual behavior and morphology in male and female Japanese quail. Phys- iol. Behav. 30 (1983) 335-339.
12. Balthazart J., Schumacher M. and Malacarne G.: Interaction of androgens and estrogens in the control of sexual behavior in male Japanese quail. Physiol. Behav. 35 (1985) 157-166.
13. Balthazart J., Foidart A. and Hendrick J. C.: The induction by testosterone of aromatase activity in the preoptic area and activation of couplatory behavior. Physiol. Behav. 47 (1990) 83-94.
14. Balthazart J., Evrard L. and Surlemont C.: Effects of the non-steroidal aromatase inhibitor R76713 on testosterone-induced sexual behavior in the Japanese quail (Coturnix coturnix japonica). Horm. Behav. 24 (1990) 510-531.
15. Adkins E. K. and Nock B. L.: The effects of the antiestrogen CI-628 on sexual behavior activated by androgen and estrogen in quail. Horm. Behav. 7 (1976) 417-429.
16. Alexandre C. and Balthazart J.: Effects of metabolism inhibitors, antiestrogens and antiandrogens on the androgen and estrogen induced sexual behavior in Japanese quail. Physiol. Behav. 38 (1986) 581-591.
17. Deviche P. and Schumacher M.: Behavioural and morphological dose-responses to testosterone and to 5ct-dihydrotestosterone in the castrated male Japanese quail. Behav. Proc. 7 (1982) 107-121.
18. Deviche P., Delville Y. and Balthazart J.: Central and peripheral metabolism of 5~-dihydrotestosterone in the male Japanese quail: biochemical characterization and relationship with reproductive behavior. Brain Res. 421 (1987) 106-116.

538 J. BALTHAZART and A. FOIDART
19. Bonne C. and Raynaud J. P.: Methyltrienolone, a specific ligand for cellular androgen receptors. Steroids 26 (1976) 227 232.
20. Bonne C. and Raynaud J. P.: Assay of androgen binding sites by exchange with methyltrienolone. Ster- oids 27 (1976) 449 507.
21. Doering C. H. and Leyra P. T.: Methyltrienolone (R1881) is not aromatized by placental microsomes or rat hypothalamic homogenates. J. Steroid Biochem. 20 (1984) 1157 1162.
22. Doering C. H. and Leyra P. T.: Lack of aromatization of methyltrienolone (RI181). In Metabolism of Hor- monal Steriods in the Neuroendocrine Structures (Edited by F. Celotti, F. Naftolin and L. Martini) Raven Press, New York (1984) pp. 139-148.
23. Schumacher M., Alexandre C. and Balthazart J.: Inter- actions des androg+nes et des oestrog6nes dans le contr61e de la reproduction. C.R. Acad. Sci. Paris, Serie 111 305 (1987) 569-574.
24. Morrell J. I., Kelley D. B. and Pfaff D. W.: Sex steroid binding in the brain of vertebrates. In Brain-Endocrine Interactions H (Edited by K. M. Knigge, D. E. Scott, H. Kobayashi, S. Muira and S. Ishii) Karger, Basel (1975) pp. 230 256.
25. Pfaff D. W.: The neuroanatomy of sex hormone receptors in the vertebrate brain. In Neuroendocrine Regulation of Fertility (Edited by T. C. Anand Kumar) Karger, Basel (1976) pp. 30-45.
26. Morell J. I. and Pfaff D. W.: A neuroendocrine approach to brain function: localization of sex steroid concentrating cells in vertebrate brains. Am. Zoo. 18 (1978) 447-460.
27. StumpfW. E. and Sar M.: Anatomical distribution of estrogen, androgen, progestin, corticoid and thyroid hormone target sites in the brain of mammals: phy- logeny and ontogeny. Am. Zoo. 18 (1978) 435 445.
28. Kelley D. B. and Pfaff D. W.: Generalizations from comparative studies on neuroanatomical and endo- crine mechanisms of sexual behaviour. In Biological Determinants of Sexual Behaviour (Edited by J. B. Hutchison) John Wiley & Sons, Chichester (1978) pp. 225 254.
29. Viglietti-Panzica C., Panzica G. C., Fiori M. G., Calcagni M., Anselmetti G. C. and Balthazart J.: A sexually dimorphic nucleus in the quail preoptic area. Neurosci. Lett. 64 (1986) 129-134.
30. Adkins-Regan E. and Watson J. T.: Sexual dimor- phism in the avian brain is not limited to the song control system of birds: a morphometric analysis of the brain of the quail (Coturnix.japonica). Brain Res. 514 (1990) 320-326.
31. Panzica G. C., Viglietti-Panzica C., Calcagni M., Anselmetti G. C., Schumacher M. and Balthazart J.: Sexual differentiation and hormonal control of the sexually dimorphic preoptic medial nucleus in quail. Brain Res. 416 (1987) 59~8.
32. Panzica G. C., Viglietti-Panzica C., Sanchez F., Sante P. and Balthazart J.: Effects of testosterone on a selected neuronal population within the preoptic sexu- ally dimorphic nucleus of the Japanese quail J. Comp. Neurol. 303 (1991) 443-456.
33. Watson J. T. and Adkins-Regan E.: Neuroanatomical localization of sex steroid-concentrating cells in the Japanese quail (Coturnix japonica): Autoradiography with [3H]-testosterone, [3H]-estradiol, and [3H]-dihy- drotestosterone. Neuroendocrinology 49 (1989) 51~4.
34. Balthazart J., Gahr M. and Surlemont C.: Distribution of estrogen receptors in the brain of the Japanese quail: an immunocytochemical study. Brain Res. 501 (1989) 205-214.
35. Schumacher M. and Balthazart J.: Neuroanatomical distribution of testosterone metabolizing enzymes in the Japanese quail. Brain Res. 422 (1987) 137 148.
36. Schlinger B. A. and Callard G. V: Estrogen receptors in quail brain: A functional relationship to aromatase and aggressiveness. Biol. Reprod. 40 (1989) 268-275.
37. Watson J. T. and Adkins-Regan E.: Activation of sexual behavior by implantation of testosterone propi- onate and estradiol benzoate into the preoptic area of the male Japanese quail (Coturniv japonica). Horm. Behav. 23 (1989) 251 268.
38. Watson J. T. and Adkins-Rcgan E.: Testosterone implanted in the preoptic area of male Japanese quail must be aromatized to activate copulation. Horm Behav. 23 (1989) 432~,47.
39. Balthazart J. and Surlemont C.: Copulatory behavior is controlled by the sexually dimorphic nucleus of the quail POA. Brain Res. Bull 25 (1990) 7-14.
40. Balthazart J., Surlemont C. and Harada N.: Aroma- tase as a cellular marker of testosterone action in the preoptic area. Physiol. Behav. 51 (1992) 395~,09.
41. Balthazart J. and Surlemont C.: Androgen and estro- gen action in the preoptic area and activation of copulatory behavior in quail. Physiol. Behav. 48 (1990) 599~609.
42. Steimer T. and Hutchison J. B.: Androgen increases formation of behaviorally effective oestrogen in dove brain. Nature 292 (1981) 345 34Z
43. Roselli C. E. and Resko J. A: Androgens regulate brain aromatase activity in adult male rats through a receptor mechanism. Endocrinoh~gy 114 (1984) 2183 -2189.
44. Roselli C. E., Horton L. E. and Resko L A.: Distri- bution and regulation of aromatase activity in the rat hypothalamus and limbic system Endocrinology 117 (1985) 24712477.
45. Schumacher M. and Balthazart J.: Testosterone-in- duced brain aromatase is sexually dimorphic. Brain Res. 370 (1986) 285-293.
46. Schlinger B. A. and Callard G. V.: Aromatase activity in quail brain: Correlation with aggressiveness. Endo- crinology 124 (1989) 437--443.
47. Palkovits M.: Isolated removal of hypothalamic or other brain nuclei of the rat. Brain Res. 59 (1973) 449J,50.
48. Balthazart J., Schumacher M. and Evrard L.: Sex differences and steroid control of testosterone- metabolizing enzyme activity in the quail brain, J. Neuroendocr. 2 (1990) 675-683.
49. Blaustein J. D. and Olster D H.: Gonadal steroid hormone receptors and social behaviors. In Advances in Comparative and Environmental Physiology, Vol 3 (Edited by J. Balthazart) Springer Verlag, Berlin (1989) pp. 31 104.
50. Balthazart J., Foidart A. and Harada N.: Immuno- cytochemical localization of aromatase in the brain. Brain Res. 514 (1990) 327-333.
51~ Balthazart J., Foidart A., Surlemont C., Vockel A. and Harada N.: Distribution of aromatase in the brain of the Japanese quail, ring dove, and zebra finch: An immunocytochemical study. J. Comp. Neurol. 301 (1990) 276-288.
52. Harada N.: Novel properties of human placental aromatase as cytochrome P-450: purification and characterization of a unique form of aromatase. J. Biochem. 103 (1988) 106-113.
53. Balthazart J., Foidart A., Surlemont C. and Harada N.: Preoptic aromatase in quail: behavioral, biochemi- cal and immunocytochemical studies. In Hormones, Brain and Behavior in Vertebrates. 2. Behavioural Acti- vation in Males and Females--Social Interactions and Reproductive Physiology. Comp. Physiol. Vol 9, (Edited by J. Balthazart) Karger, Basel (1990) pp. 45-62.
54. Balthazart J., Foidart A., Surlemont C., Harada N, and Naftolin F.: Neuroanatomical specificity in the autoregulation of aromatase-immunoreactive neurons

Aromatase and sexual behavior 539
by androgens and estrogens: An immunocytochemical study. Brain Res. 574 (1992) 280-290.
55. Ryan K. J.: Biological aromatization of steroids. J. Biol. Chem. 234 (1959) 268-272.
56. Fournet-Dulguerov., MacLusky N. J., Leranth C., Todd R., Mendelson C. R., Simpson E. R. and Naftolin F.: Immunohistochemical localization of aromatase cytochrome P-450 and estradiol dehydro- genase in the syncytiotrophoblast of the human pla- centa. J. Clin. Endocr. Metab. 65 (1987) 757-764.
57. Naftolin F., Leranth C. and Balthazart J.: Ultrastruc- rural localization of aromatase immunoreactivity in hypothalamic neurons. The Endocr. Soc. Abstr. 669 (1990) 192.
58. Balthazart J., Foidart A., Surlemont C., Harada N., Leranth C. and Naftolin F.: Immunocytochemical localization of aromatase and estrogen receptors in the brain. Soc. Neurosci. 16 (1990) 1313.
59. Steimer Th.: Aromatase activity in rat brain synapto- somes. Is an enzyme associated with the neuronal cell membrane involved in mediating non-genomic effects of androgens. Eur. J. Neurosci. (Suppl.) 0988) 9.
60. Schlinger B. A. and Callard G. V.: Localization of aromatase in synaptosomal and microsomal subfrac- tions of quail (Coturnix coturnix japonica) brain. Neuroendocrinology 49 (1989) 434-441.
61. Schumacber M.: Rapid membrane effects of steroid hormones: an emerging concept in neuroendocrin- ology. Trends Neurosci. 13 (1990) 359-362.
62. Wouters W., De Coster R., Krekels M., Van Dun J., Beerens D., Haelterman C., Raeymaekers A., Freyne E., Van Gelder J., Venet M. and Janssen P. A. J.: R76713, a new specific non-steriodal aromatase inhibi- tor. J. Steroid Biochem. 32 (1989) 781-788.
63. De Coster R., Wouters W., Bowden C. R., Vanden Bossche H., Bruynseels J., Tuman R. W., Van Ginckel R., Snoeck E., Van Peer A. and Janssen P. A. J.: New non-steroidal aromatase inhibitors: Focus on R76713. J. Steroid Biochem. Molec. Biol. 37 (1990) 335-341.
64. Hutchison J. B. and Steimer T.: Formation of behav- iorally effective 17fl-estradiol in the dove brain: steroid control of preoptic aromatase. Endocrinology 118 (1986) 2180-2187.
65. Balthazart J., Foidart A., Surlemont C. and Harada N.: Neuroanatomical specificity in the co-localization of aromatase and estrogen receptors. J. Neurobiol. 22 (1991) 143-157.
66. Blaustein J. D., Lehman M. N., Turcotte J. C. and Greene G.: Estrogen receptors in dendrites and axon terminals in the guinea pig hypothalamus. Endocrin- ology 131 (1992) 281-290.
67. Blaustein J. D. and Turcotte J. C.: Estrogen receptor- immunostaining of neuronal cytoplasmic processes as well as cell nuclei in guinea pig brain. Brain Res. 495 (1989) 75-82.
68. Schumacher M., Contenti E. and Balthazart J.: Partial characterization of testosterone-metabolizing enzymes in the quail brain. Brain Res. 305 (1984) 51-59.
69. Steimer T. and Hutchison J. B.: Is the androgen- dependent increase in preoptic estradiol-17fl formation due to aromatase induction. Brain Res. 480 (1989) 335-339.
70. Schumacher M. and Hutchison J. B.: Testosterone induces hypothalamic aromatase during early develop- ment in quail. Brain Res. 377 (1986) 63-72.
71. Hutchison J. B. and Schumacher M.: Development of testosterone-metabolizing pathways in the avian brain: enzyme localization and characteristics. Dev. Brain Res. 25 (1986) 33-42.
72. Balthazart J.: Testosterone metabolism in the avian hypothalmus. J. Steroid Biochem. Molec. Biol. 40 (1991) 557-570.
73. Harada N., Yamada K., Foidart A. and Balthazart J.: Regulation of aromatase cytochrome P-450 (estrogen synthetase) transcripts in the quail brain by testoster- one. Molec. Brain Res. 15 (1992) 19-26.
74. Hutchison J. B., Joris S., Hutchison R. E. and Steimer T.: Steroid control of sexual behavior and brain aromatase in the dove: Effects of nonaromatizable androgens, methyltrienolone (R1881), and 5ct-dihy- drotestosterone. Horm. Behav. 23 (1989) 542-555.
75. Roselli C. E., Horton L. E. and Resko J. A.: Time- course and steroid specificity of aromatase induction in rat hypothalamus-preoptic area. Biol. Reprod. 37 (1987) 628~33.
76. Roselli C. E., Salisbury R. L. and Resko J. A.: Genetic evidence for androgen-dependent and independent control of aromatase activity in the rat brain. Endo- crinology 121 (1987) 2205-2210.
77. Roselli C. E.: Synergistic induction of aromatase ac- tivity in the rat brain by estradiol and 5ct-dihy- drotestosterone. Neuroendocrinology 53 (1991) 79-84.
78. Sheridan P. J.: Androgen receptors in the brain: what are we measuring? Endocrine Rev. 4 (1983) 171-178.
79. S6derstein P. and Gustafsson J. A.: A way in which estradiol might play a role in the sexual behavior of male rats. Horm. Behav. 14 (1980) 271-274.
80. S6dersten P., Eneroth P., Hansson T., Mode A., Johansson D., Naslund B., Liang B., Liang T. and Gustafsson J. A.: Activation of sexual behaviour in castrated rats: the role of oestradiol. J. Endocrinology 111 (1986) 455-462.
81. Sbdersten P., Eneroth P. and Hansson T.: Oestradiol synergizes with 5~t-dihydrotestosterone or 3~t- but not 3fl-androstanediol in inducing sexual behaviour in castrated rats. J. Endocr. 119 (1988) 461-465.
82. Balthazart J., Foidart A., Loeffen R., Dohmae S. and Harada N.: Synergism between androgen and estrogen in the induction of aromatase in the quail brain: immunocytochemical and RT-PCR studies. Soc. Neurosci. Abstr. 18 (1992) 894.
83. Baltbazart J., Doidart A., Wilson E. M. and Ball G. F.: Immunocytochemical localization of androgen recep- tors in the male songbird and quail brain. J. Comp. Neurol. 317 (1992) 407-420.
84. Gahr M.: Localization of androgen receptors and estrogen receptors in the same cells of the songbird brain. Proc. Nam. Acad. Sci. U.S.A. 87 (1990) 9445-9448.
85. Lephart E. D., Simpson E. R. and Ojeda S. R.: Effects of cyclic AMP and androgens on in vitro brain aroma- tase enzyme activity during prenatal development in the rat. J. Neuroendocr. 4 (1992) 29-35.
86. Hutchison J. B., Steimer Th. and Hutchison R. E.: Formation of behaviorally active estrogen in the dove brain: induction of preoptic aromatase by intracranial testosterone. Neuroendocrinology 43 (1986) 416-427.
87. Kebabian J. W. and Calne D. B.: Multiple receptors for dopamine. Nature 277 (1979) 93-96.
88. Sokoloff P., Giros B., Martres M. P., Bouthenet M. L. and Schwartz J. C.: Molecular cloning and character- ization of a novel dopamine receptor (D3) as a target for neuroleptics. Nature 347 (1990) 146-151.
89. Snyder S. H.: The dopamine connection. Nature 347 (1990) 121-122.
90. Kandel E. R., Schwartz J. H. and Jessell T. M.: Principles of Neural Science. Elsevier, New York 3rd Edn (1991).
91. Callard G. V.: Aromatization is cyclic AMP- dependent in cultured brain cells. Brain Res. 204 (1981) 461-464.
92. Cardinali D. P., Ritta M. N. and Gejman P. V.: Norepinephrine stimulates testosterone aromatization and inhibits 5~-reduction via fl-adrenoreceptors in the

540 J. BALTHAZART and A. FO1DART
rat pineal gland. Molec. Cell. Endcocr. 2,8 (1982) 199-209.
93. Verhoeven G. P., Dierckx P. and de Moor P.: Stimu- lation effect of neurotransmitters on the aromatization of testosterone by Sertoli cell-enriched cultures. Molec. Cell. Endocr. 13 (1979) 241 253.
94. Mendelson C. R., Smith M. E., Cleland W. H. and Simpson E. R.: Regulation of aromatase activity of cultured adipose stromal cells by catecholamines and adrenocorticotropin. Molec. Cell Endocr. 37 (1984) 61-72.
95. Nock B. and Feder H. H.: Neurotransmitter modu- lation of steroid action in target cells that mediate reproduction and reproductive behavior. Neurosci. Biobehav. Rev. 5 (1981) 437-447.
96. Verhoeven G.: Effect o f neurotransmitters and follicle- stimulating hormone on the aromatization of andro- gens and the production of 3 ' ,5 '-monophosphate by cultured testicular cells. J. Steroid Biochem. 12 (1980) 315-322.
97. Canick J. A., Tobet S. A., Baum M. J., Vaccaro D. E., Ryan K., J., Leeman S. E. and Fox T. O.: Studies on the role of catecholamines in the regulation of the developmental pattern of hypothalamic aromatase. Steroids 50 (1987) 510-521.
98. Marcano M., Raum W. J. and Swerdloff R. S.: fl- adrenergic inhibition of aromatization in the female neonatal rat. Clin. Res. 29 (1981) 84A.
99. Raum W. J. and Swerdloff R. S.: The role of hypo- thalamic adrenergic receptors in preventing testoster-
one-induced androgenization in the female rat bra in Endocrinology 109 (1981) 273 278.
100. Raum W. J., Marcano M. and Swerdloff R. S.: Nu- clear accumulation of estradiol derived from the arom- atization of testosterone is inhibited by hypothalamic beta-receptor stimulation in the neonatal female rat. Biol. Reprod. 30 (1984) 388-396.
101. Balthazart J. and Ball G. F.: Effects of the noradren- ergic neurotoxin DSP-4 on luteinizing hormone levels, catecholamine concentrations, ~-adrenergic receptor binding, and aromatase activity in the brain of the Japanese quail. Brain Res. 492 (1989) 163-175.
102. Balthazart J., Libioulle J. M. and Sante Ph.: Stimu- latory effects of the noradrenergic neurotoxin DSP4 on sexual behavior in male quail. Behav. Proc. 17 (1988) 27~14.
103. El Halawani M. E., Burke W. t-I. and Ogren L. A~ Involvement of catecholaminergic mechanisms in the photoperiodically induced rise in serum luteinizing hormone of Japanese quail (Coturnix coturnix japon- ica). Gen. Comp. Endocr. 41 (1980) 14-21.
104. El Halawani M. E., Burke W H. and Ogren L. A: Effects of drugs that modify brain monoamine concen. trations on photoperiodically-induced testicular growth in coturnix quail (Coturnix coturnix japonica L Biol. Reprod. 18 (1978) 198--203.
105. Bailhache T., Foidart A., Surlemont C., Harada N. and Balthazart J.: Catechotaminergic innervation of aromatase and estrogen receptorqmmunoreactive cells in the quail brain. Soc. Neurosci~ Abstr. 17 (1991) 269.




















