
Characterization of virulence of Leptospira isolates in a hamster model
Upload
independentCategory
view
0download
0
Available online at www.sciencedirect.com
4 (2008) 412–421www.elsevier.com/locate/phb
Physiology & Behavior 9
Testosterone, androstenedione, and 5α-dihydrotestosterone on male sexualbehavior and penile spines in the hamster
M. Arteaga-Silva a,⁎, R.M. Vigueras-Villaseñor b, S. Retana-Márquez a, M. Hernández-González c,C. Chihuahua-Serrano a, H. Bonilla-Jaime a, J.L. Contreras a, G. Moralí d
a Departamento de Biología de la Reproducción, Universidad Autónoma Metropolitana, Iztapalapa, P.O. Box 55 535, México 09340, D.F., Mexicob Lab. Histomorfología, Instituto Nacional de Pediatría, México, D.F., Mexico
c Instituto de Neurociencias, U. de G., Guadalajara, Jalisco, Mexicod Unidad de Investigación Médica en Farmacología, CMN Siglo XXI, IMSS, México, D.F., Mexico
Received 8 June 2007; received in revised form 10 February 2008; accepted 12 February 2008
Abstract
Testosterone, androstenedione, and 5α-dihydrotestosterone on male sexual behavior and penile spines in the hamster. PHYSIOL BEHAV XX(6) 000-000, 2007. — The expression of masculine sexual behavior (MSB) in male hamsters is optimally stimulated by aromatizable androgenslike androstenedione (AD) and testosterone (T), while the non-aromatizable androgen, 5α-dihydrotestosterone (DHT), exerting potent androgenicperipheral effects, only in high doses maintains MSB after castration. No data exist on the ability of these androgens to restore long intromissionsafter castration. In this study, AD, T, and DHT were administered to four-week gonadectomized, sexually experienced male hamsters, for threeweeks, in doses of 25 μg/day or up to 1000 μg/day to compare their potency in restoring MSB, penile size, and penile spines growth. Plasmalevels of these steroids and the metabolites estrone and estradiol, were determined at the end of the treatment period. Gonadectomy completelysuppressed MSB and induced a regression of penile spines. AD was more potent than T in restoring MSB, ejaculatory behavior being displayed bymost castrated subjects with a lower dose of AD (50 μg/day) than of T (300 μg/day), and long intromissions being shown by all AD-treatedcastrated hamsters but only by 20% of T-treated ones, when doses of 1000 μg/day were given. DHT did not stimulate any copulatory response.The three androgens, even at the lowest dose, partially stimulated penis and penile epithelium growth, DHT showing the highest potency.Treatment of castrated hamsters with AD (50 μg/day), restored steroid levels to similar values as those of intact animals. These results show thatAD and T restored MSB even with a partial stimulation of penile spines growth, AD being more potent than T. In contrast, DHT did not restoreMSB in the hamster in spite of its peripheral androgenic potency.© 2008 Elsevier Inc. All rights reserved.
Keywords: Golden hamsters; Androgens; Androstenedione; Male sexual behavior; Long intromissions; Penile spines
1. Introduction
Gonadal hormones play an important role for a variety ofreproductive functions in male mammals, such as the onset andmaintenance of masculine sexual behavior (MSB), and thedevelopment, growth, and function of the seminal vesicles,prostate, and penis among other tissues. In some mammals
⁎ Corresponding author.E-mail address: [email protected] (M. Arteaga-Silva).
0031-9384/$ - see front matter © 2008 Elsevier Inc. All rights reserved.doi:10.1016/j.physbeh.2008.02.007
including rodents, carnivores, and primates, the penis is coveredby a stratified epithelium having follicles from which cornifiedpapillae or spines emerge [1–3].
Two hypothesis have been proposed on the role of penilespines in rats: 1) they contribute to vaginal stimulation providedby the male at copulation, promoting neuroendocrine responsesthat prepare the uterus for pregnancy; 2) penile spines maycontribute to the removal of a seminal plug from a formercopulation, thus allowing the male to deposit a new plug into thevaginal tract [4].

413M. Arteaga-Silva et al. / Physiology & Behavior 94 (2008) 412–421
In hamsters [5], as in rats [2,3], keratinized penile spines arefound along the penile epithelium, being more densely locatedat the distal than at the proximal region of the penis. Penilespines are androgen-dependent structures, castration leading totheir regression and to follicular atrophy, while their growth iscompletely restored by treatment with testosterone (T) or withits peripherally active metabolite, 5α-dihydrotestosterone(DHT) [6–10]. However, development of the penile spinesdoes not correlate with the display of MSB, since systemicallyadministered estrogen [4], or intracerebral implants of T [11],may restore this behavior even in the absence, or after only apartial stimulation, of penile spines growth.
As in rats, several ejaculatory series are displayed byhamsters at copulation, and three types of stereotypedcopulatory responses may be recognized: mount, intromission,and ejaculation patterns. However, after several ejaculatoryseries have occurred, hamsters shift from intromission to longintromission patterns [12]. These responses appear as the maleapproaches sexual exhaustion, and are characterized by alengthening in intravaginal penile insertion [12–14]. Theseresponses have been only scarcely analyzed, and are thought toplay a role in triggering the neuroendocrine responsesassociated to pregnancy in females [15,16].
The effect of diverse androgens: T, DHT, androstenedione(AD), and androsterone, as well as that of estradiol (E2), onmale sexual behavior of hamsters, has been studied. T and ADin doses of 100 μg/day or up to 800 μg/day, maintain malesexual behavior in hamsters when given for several weeksstarting soon after castration [17–19]. E2 treatment onlystimulates mounting but neither intromission nor ejaculatorybehavior in castrated hamsters [19,20]. Results concerningDHT treatment are controversial. DHT in a dose of 1 mg/day,but not at lower doses, has been shown to be as effective as T tomaintain but not to restore male sexual behavior in adulthamsters when this behavior ceased to be displayed aftercastration [17–19,21]. This dose maintains the copulatorybehavior in adult (60 days-old), but not in juvenile (30 days-old)hamsters, though with few ejaculatory responses [21]. Otherauthors have found that the intracerebral administration of DHTfails to reverse the effects of castration, only restoring mounts[22,23]. On the other hand, stimulation of peripheral tissues bythe subcutaneous administration of DHT increases the beha-vioral effects of either E2 or T when implanted into the brain[24], supporting the idea that the peripheral, rather than centraleffects of DHT are involved in the complete stimulation ofcopulatory behavior in the male hamster. All these studies haveonly assessed the effects of hormone replacement on the first, orup to the third ejaculatory series, without analyzing their abilityto restore long intromissions.
The role of AD for the expression of MSB in hamsters hasbeen suggested by recent findings of our laboratory [20]. Thus,significantly higher plasma levels of AD than of T are present inadult intact hamsters. On the other hand, AD, at a daily dose of1 mg, is more effective than T or the combined treatment withDHT and E2, to restore MSB in castrated hamsters, includinglong intromissions. No reports exist neither on the effects oflower doses of AD on male sexual behavior, nor on the plasma
levels of T, AD, and its metabolites after its administration indifferent dose levels, as compared to those of intact hamsters. Inparticular, since aromatization of androgens plays an importantrole for the display of MSB in a number of species [25], andsince estrogen has been proposed to participate on a cerebralmechanism for stimulating mounting behavior in hamsters [24],it was considered of interest to determine the plasma levels ofestrone (E1) and estradiol (E2) as estrogenic metabolites of ADand T, respectively. On the other hand, no information exists onperipheral effects of AD on penile spines growth, and nocorrelations have been described between the behavioral and theperipheral effects of AD, T, and DHT after castration.
The aim of the present study was to compare the potency ofT, AD, and DHT given in various doses, to restore MSB andpenile spines in the castrated golden hamster.
2. Material and methods
2.1. Animals
Adult male and female hamsters (Mesocricetus auratus)weighing between 100 to 120 g were purchased from HarlanLaboratories. Housing and experimental procedures werecarried out in accordance with the Standard guidelines setforth in the NIH Guide for the Care and Use of LaboratoryAnimals, in groups of six per cage under a reversed light–darkcycle (L:14 h; D:10 h) with Chow Purina food and water adlibitum.
2.2. Screening tests
Three screening tests were done to each male hamster, atweekly intervals, to assess their copulatory behavior and toselect those sexually active. Tests were conducted during thedark phase of the light–dark cycle, under dim red light. Eachmale was allowed to adapt to the testing arena (1/8 in. Plexiglas,60 cm diameter, 45 cm high) for 5 min before a receptive femalewas presented. Receptive females were treated with 3 μgestradiol benzoate three times per week, and 500 μg progester-one 4 h before testing, both hormones from Sigma ChemicalCo. St. Louis. MO, USA, in 50 μl corn oil, sc. Only animalsdisplaying three successive copulatory series in no more than15 min in each test, were included in the study.
2.3. Experimental design
Animals included in the study were randomly assigned todifferent groups. All hamsters excepting those in one group(Sham), were castrated as is described below.
All subjects were tested for sexual behavior each week aftersurgery, until this behavior ceased to be displayed in castratedanimals (four weeks). Tests were carried out under the sameconditions as the screening tests, but lasted 1 h in order to besure that the behavior was completely eliminated aftercastration. After this period, males were randomly assigned tothe various groups (n=15 each) and received one of thedifferent daily doses of the androgens for three weeks. The

414 M. Arteaga-Silva et al. / Physiology & Behavior 94 (2008) 412–421
Sham group and a group of castrated animals were treated withcorn oil, 50 μl/day sc. Masculine sexual behavior testscontinued once per week (at days 7, 14, and 21) until the endof the treatment period. These tests also lasted 1 h to allow theoccurrence of long intromission responses. Numbers of mountsand intromissions displayed in the first copulatory series of thelast behavioral test, and the total number of ejaculations andlong intromissions displayed in that test, were recorded forcomparisons among groups. Immediately after the last beha-vioral test, animals were sacrificed by decapitation to obtainblood samples for sex hormone assays, and the penis for itsmeasurement and histological assessment.
2.4. Surgeries
Males in all groups were anesthetized with xylazine(10 mg/kg) and ketamine (90 mg/kg) ip, and subjected to amidline scrotal incision and their testicles were removed.Males in the Sham group were anesthetized in a similar wayand subjected to a scrotal incision without removing anytissue, followed by suture.
2.5. Steroids and hormonal treatment
The following steroid hormones and doses were used:Androstenedione (4-Androstene-3,17-dione, Fluka, ChemikaCo. Switzerland); Testosterone (4-Androstene-17-β-ol-3-one,Fluka, Chemika Co. Switzerland); 5α-Dihydrotestosterone (5α-Androstane-17-β-ol-3-one, Fluka, Chemika Co. Switzerland) indaily doses of 25, 50, 100, 300, 500, or 1000 μg in a volume of50 μl, sc. All steroids were dissolved in corn oil.
2.6. Collection of plasma and steroid hormone assay
Immediately after the last behavioral test, animals weresacrificed by decapitation and blood (1.5 ml samples) wascollected from the trunk. Plasma was obtained by centrifugationat 1500 rpm for 30 min. Regardless of hormonal treatment, ineach plasma sample the levels of T, AD, DHT, E2, and estrone(E1) were determined. Steroid hormones were extracted andquantified by HPLC following a modification of the proceduredescribed elsewhere [26]. Briefly, 100 μl of a 19-nortestoster-one solution (5 μg/ml in methanol) were added as internalstandard to 500 μl of plasma. Steroids were extracted byagitation with 5 ml of a diethyl ether-dichloromethane mixture(60:40 v/v) followed by centrifugation. One ml HPLC degreewater was added to the organic phase and centrifuged. Theorganic phase (3 ml) was evaporated under nitrogen at roomtemperature. Residue was dissolved in 100 μl of a methanol–water (60:40 v/v) mixture. The HPLC column and precolumn(Waters Corp., Milford, MA) were equilibrated with a water–acetonitrile (65:35 v/v) mixture, at a 0.4 ml/min flow. Steroidseparation was carried out at 40 °C. A system controller(Waters, 600-MS) was used to pump the mobile phase andsteroids were identified by an UV detector (Waters, Mod. 486)at a 250 nm wavelength. Results were analyzed by a datahandling station (Waters, Millennium32 Workstation). Intra-
and inter-assay variation coefficients for steroid determinationswere 2.93% and 7.12% respectively.
2.7. Collection and processing of penile samples
After collection of blood, the animals were placed in supineposition, the penis was completely protruded, and 1 ml of a 10%glutaraldehyde in paraformaldehyde solution was injectedthrough the penile vein until the penis appeared erect [27].The organ was cut and post-fixed by immersion in 10 ml of thesame fixative solution for 24 h. Tissues were then dehydratedand processed until embedded in paraffin. Longitudinal serialsections (10 μm) were obtained from each penis and stainedwith hematoxylin-eosin.
Sections of the midline portion of the penis were examinedunder the microscope at a 100× magnification, and theformation of penile follicles as well as the number of penilespines in a 500 μm length of both the basal and the distal regionof the penis, were recorded.
Some of the castrated animals in each group (n=6 pergroup), were deeply anesthetized after the last behavioral testand their penis were removed to assess the fresh weight (mg),and to measure their width and length (mm).
2.8. Statistical analysis
For each parameter, the mean and SEM in each experimentalgroup were calculated. Comparisons of the plasma levels ofsteroids, fresh weight, width, and length of the penis, thenumber of penile follicles and spines both in the basal and thedistal region of the organ, as well as the numbers of mounts,intromissions, ejaculations, and long intromissions performedon the last behavioral test by males receiving each treatment(Vehicle, AD, T, DHT) were done by a one-way ANOVAfollowed by post-hoc Tukey tests. For mounts and intromis-sions, only those performed in the first copulatory series, wereanalyzed. In addition comparisons of the effect of differenthormones, for a given dose level, were also done by Tukey tests.Tests were performed by using the GB-stat program (SchoolPark Dynamic Microsystems, Inc. 1997). Proportions of malesshowing mount, intromission, ejaculation, and long intromis-sion responses in the last behavioral test under the differentexperimental conditions were compared between groups by X2
followed by the Fisher Exact Probability test. A pb0.05 valuewas considered statistically significant.
3. Results
3.1. Male sexual behavior
Fig. 1 shows the percentages of hamsters displaying mounts,intromissions, ejaculations, and long intromissions in the lastbehavioral test under the various experimental conditions(intact, castration, and hormone treatment). The display ofcopulatory responses was not modified after sham surgicalprocedures, the animals continuing copulating four weeks later,and for three weeks of treatment with vehicle. On the other

Fig. 1. Percent animals showing mount, intromission, ejaculation, and long intromission responses, in the last test under the various experimental conditions: Sham,castrated and daily treatment with vehicle (C+VEH), or with the various doses of AD or T. Daily treatment (three weeks) with 1000 μg of AD fully restored the displayof copulatory responses in all animals. The same dose of T only stimulated the display of long intromissions in 20% of subjects. ⁎pb0.01 as compared to the Shamgroup; °pb0.01 as compared to AD (Fisher exact probability test).
415M. Arteaga-Silva et al. / Physiology & Behavior 94 (2008) 412–421
hand, castration resulted in a total decline of copulatoryresponses in all hamsters, four weeks after surgery.
Treatment with AD to castrated animals restored theirdisplay of copulatory responses in a dose-related manner: 40%of the hamsters receiving 25 µg/day and 80% of thosereceiving 50 µg/day, showed mounts, intromissions andejaculations along the three-week treatment period, and thepercentages of animals showing MSB in the last behavioraltest, after 21 days of treatment, increased with the androgendose, so that 100% of the animals treated with 500 or with1000 µg/day displayed ejaculations. All castrated hamstersreceiving AD at the dose of 1000 µg/day displayed longintromissions, in a similar way to the Sham group (Fig. 1).Numbers of mounts and intromissions displayed by hamstersin the first copulatory series at the last behavioral test, and thetotal numbers of ejaculations and long intromissions dis-played in that test, were compared among the various groupsof hamsters receiving AD. Numbers of copulatory responsesdisplayed by hamsters receiving AD differed amongthe various doses and as compared to the Sham group(Mounts: F(6,89)=64.05, pb0.001; intromissions: F(6,89)=24.68, pb0.001; ejaculations: F(6,78)=331.63, pb0.001).Hamsters receiving AD at 25, 50, 100 and 300 µg/day showedmore mounts (mean±SEM: 9.2±1.3; 8.1±0.3; 7.6±0.4; 7.5±0.33, respectively) and intromissions (14.2±0.9; 13.5±0.7;12.9±0.5; 12.1±0.6, respectively) (pb0.001) to achieve thefirst ejaculation, and less ejaculations per test (only one or two,pb0.001) than the Sham group (2.2±0.2 mounts, 7.2±0.3intromissions and 10.1±0.2 ejaculations). However, animals
receiving 1000 µg/day showed similar numbers of copulatoryresponses (mean±SEM: 2.5±0.2 mounts, 7.0±0.3 intromis-sions and 9.6±1.0 ejaculations), including long intromis-sions (mean±SEM: 3.9±0.4) as the Sham group (mean±SEM:4.5±0.4). Animals receiving 500 µg/day had a similarly lownumber of mounts (mean±SEM: 2.8±0.2) as the Sham group,lower than that displayed under the lower doses (pb0.01).They displayed intermediate numbers of intromissionsto ejaculate (10.7±0.7) and a higher number of ejaculations(5.9±0.3, pb0.01) than those of animals treated with thelower doses, but not reaching the values of the Sham group.
Treatment with T also stimulated the display of copulatoryresponses in a dose-related manner, though at a lower levelthan AD. Thus, a dose of 300 µg/day was required to stimulateejaculatory behavior in 73% of castrated animals, and a 100%of hamsters receiving 500 or 1000 µg/day did; however only20% of the hamsters receiving T at a dose of 1000 µg/daydisplayed long intromissions, lower doses being ineffective(Fig. 1). Numbers of copulatory responses displayed in the lastbehavioral test by hamsters receiving T differed among thevarious doses (Mounts: (F(6,80)=75.45, pb0.001; intromis-sions: F(6,80)=29.92, pb0.001; and ejaculations: F(6,63)=219.44, pb0.001). Hamsters receiving T at 25, 50, 100, and300 µg/day displayed more mounts (mean±SEM: 9.7±1.4;8.3±0.6; 9.1±0.4; 7.9±0.5, respectively) and intromissions(mean±SEM: 14.5±1.4; 15.1±0.4; 11.6±0.6; 12.2±0.4,respectively) (pb0.001) to achieve ejaculation, and fewerejaculations per test (1.0±0.1; 1.5±0.2; 1.3±0.2; 1.8±0.2,respectively) (pb0.001), than the Sham group, only those
416 M. Arteaga-Silva et al. / Physiology & Behavior 94 (2008) 412–421
receiving 500 or 1000 µg/day exhibiting similar numbers ofmounts and intromissions, but still fewer ejaculations (mean±SEM: 2.7±0.2; 5.9±0.3, respectively) (pb0.01), as comparedto the Sham group. The only three animals displaying longintromissions under treatment with 1000 µg/day only showed1 or 2 of these responses.
DHT in daily doses of 25 to 1000 µg, failed to stimulateMSB in the castrated hamsters, animals only showing persistentfemale's anogenital inspection and olfaction, without achievingany mount.
3.2. Penile growth and histology
Castration induced a significant reduction of the penileweight (pb0.01), width (pb0.01), and length (pb0.01), ascompared to the Sham group. Treatment with the variousandrogens induced dose-dependent increases on these para-meters (Table 1).
Penile weight, width and length of animals treated with ADat the various doses, were higher than those of vehicle-treatedanimals (weight: F(7,40)=61.20, pb0.01; width: F(7,40)=84.79, pb0.01; length: F(7,40)=53.22, pb0.01). Weight valuessimilar to those of the Sham group were only reached with 500and 1000 µg/day (Table 1). Penile width of castrated hamsterswas restored even with the lowest dose of AD, while penilelength, though higher than that of vehicle treated animals, didnot reach the values of the Sham group (Table 1).
Penile parameters of animals treated with T had alsohigher values than those of vehicle-treated hamsters (weight:F(7,40)=42.24, pb0.01; width: F(7,40)=48.11, pb0.01;length: F(7,40)=35.19, pb0.01). Castrated subjects treated
Table 1Penile parameters in intact (Sham) and castrated (C) hamsters after 3 weeks of treat
Doses Penile weight Penile width
(μg/day) (mg) (mm)
Sham 50.98±6.89y 2.30±0.06y
C+VEH 20.01±1.60b 0.95±0.01b
C+AD 25 30.98±3.03b,x 2.33±0.06y
50 29.49±3.13b,x 2.26±0.06y
100 30.24±3.33b,x 2.36±0.06y
300 40.48±5.25a,y 2.36±0.06y
500 50.74±4.89y 2.40±0.05y
1000 50.80±4.96y 2.50±0.03y
C+T 25 20.21±4.09b 1.95±0.01a,y
50 20.79±2.97b 2.16±0.06y
100 31.82±2.93b,x 2.26±0.06y
300 49.92±6.23y 2.56±0.14y
500 49.30±5.18y 2.63±0.13y
1000 51.97±6.42y 2.76±0.07b,y
C+DHT 25 30.30±2.89b,x 2.36±0.06y
50 31.09±3.94b,x 2.43±0.05y
100 50.07±5.28y 2.46±0.03y
300 51.10±4.13y 2.56±0.06y
500 53.25±6.28y 3.00±0.05y
1000 55.84±4.73y 3.00±0.05b,y
Data correspond to mean±SEM.apb0.05 bpb0.01 vs Sham.xpb0.05 ypb0.01 vs castrated+VEH.
with T at the doses of 100 µg/day or more, had similar penileweights to those of animals receiving AD, and statisticallyhigher (pb0.01) than those of vehicle-treated hamsters(Table 1). Animals treated with 500 or with 1000 µg/day hadsimilar penile weights to those of the Sham group (Table 1).Treatment with T stimulated the increase of penile width andlength in a dose-related manner (Table 1), 100 µg/day andhigher doses restoring these parameters to values similar or evenhigher to those of the Sham group.
Penile parameters of animals treated with DHT had alsohigher values than those of vehicle-treated hamsters (weight: F(7,40)=74.38, pb0.001; width: F(7,40)=142.15, pb0.001;length: F(7,40)=37.23, pb0.01). On the other hand, castratedhamsters receiving DHT showed a better restoration of penilegrowth than that of AD- or T-treated animals (Table 1). Thus,treatment with 25 µg/day restored penile widths, and 100 µg/day restored penile weights and lengths, to similar values tothose of the Sham group. Animals receiving 50 to 1000 µg/dayhad even larger penile widths than those of the Sham group, thisdifferences being statistically significant (pb0.01), at the dosesof 500 and 1000 µg/day.
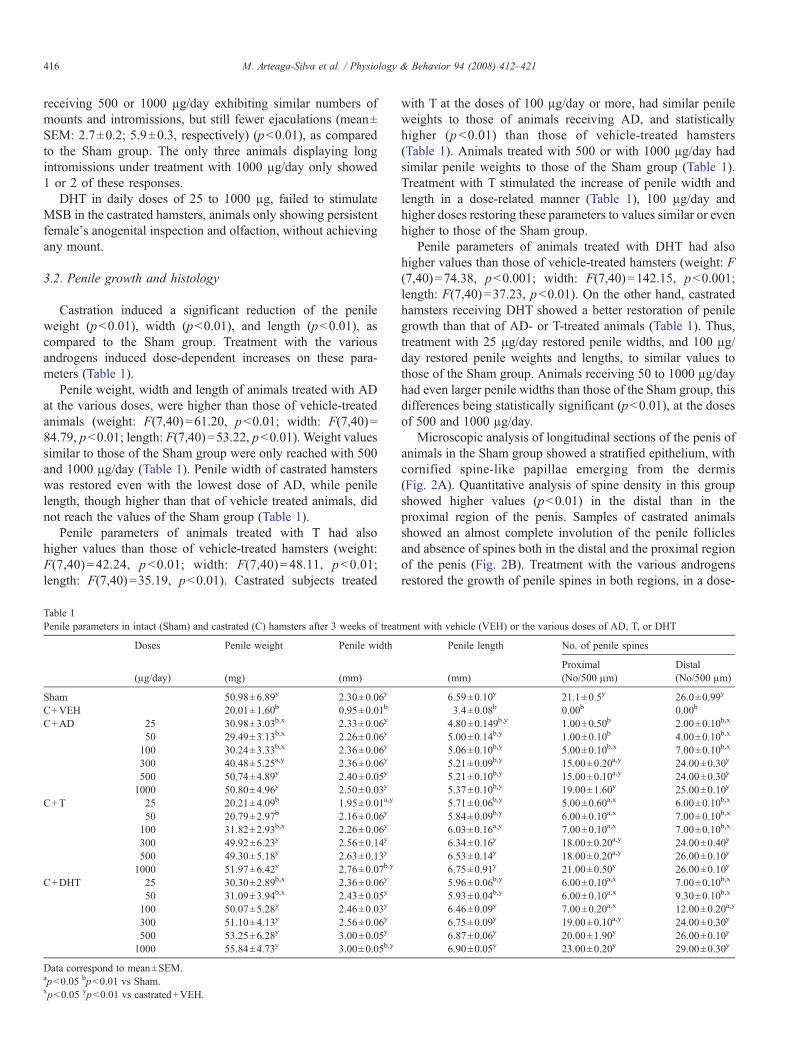
Microscopic analysis of longitudinal sections of the penis ofanimals in the Sham group showed a stratified epithelium, withcornified spine-like papillae emerging from the dermis(Fig. 2A). Quantitative analysis of spine density in this groupshowed higher values (pb0.01) in the distal than in theproximal region of the penis. Samples of castrated animalsshowed an almost complete involution of the penile folliclesand absence of spines both in the distal and the proximal regionof the penis (Fig. 2B). Treatment with the various androgensrestored the growth of penile spines in both regions, in a dose-
ment with vehicle (VEH) or the various doses of AD, T, or DHT
Penile length No. of penile spines
Proximal Distal(mm) (No/500 μm) (No/500 μm)
6.59±0.10y 21.1±0.5y 26.0±0.99y
3.4±0.08b 0.00b 0.00b
4.80±0.149b,y 1.00±0.50b 2.00±0.10b,x
5.00±0.14b,y 1.00±0.10b 4.00±0.10b,x
5.06±0.10b,y 5.00±0.10b,x 7.00±0.10b,x
5.21±0.09b,y 15.00±0.20a,y 24.00±0.30y
5.21±0.10b,y 15.00±0.10a,y 24.00±0.30y
5.37±0.10b,y 19.00±1.60y 25.00±0.10y
5.71±0.06b,y 5.00±0.60a,x 6.00±0.10b,x
5.84±0.09b,y 6.00±0.10a,x 7.00±0.10b,x
6.03±0.16a,y 7.00±0.10a,x 7.00±0.10b,x
6.34±0.16y 18.00±0.20a,y 24.00±0.40y
6.53±0.14y 18.00±0.20a,y 26.00±0.10y
6.75±0.91y 21.00±0.50y 26.00±0.10y
5.96±0.06b,y 6.00±0.10a,x 7.00±0.10b,x
5.93±0.04b,y 6.00±0.10a,x 9.30±0.10b,x
6.46±0.09y 7.00±0.20a,x 12.00±0.20a,y
6.75±0.09y 19.00±0.10a,y 24.00±0.30y
6.87±0.06y 20.00±1.90y 26.00±0.10y
6.90±0.05y 23.00±0.20y 29.00±0.30y
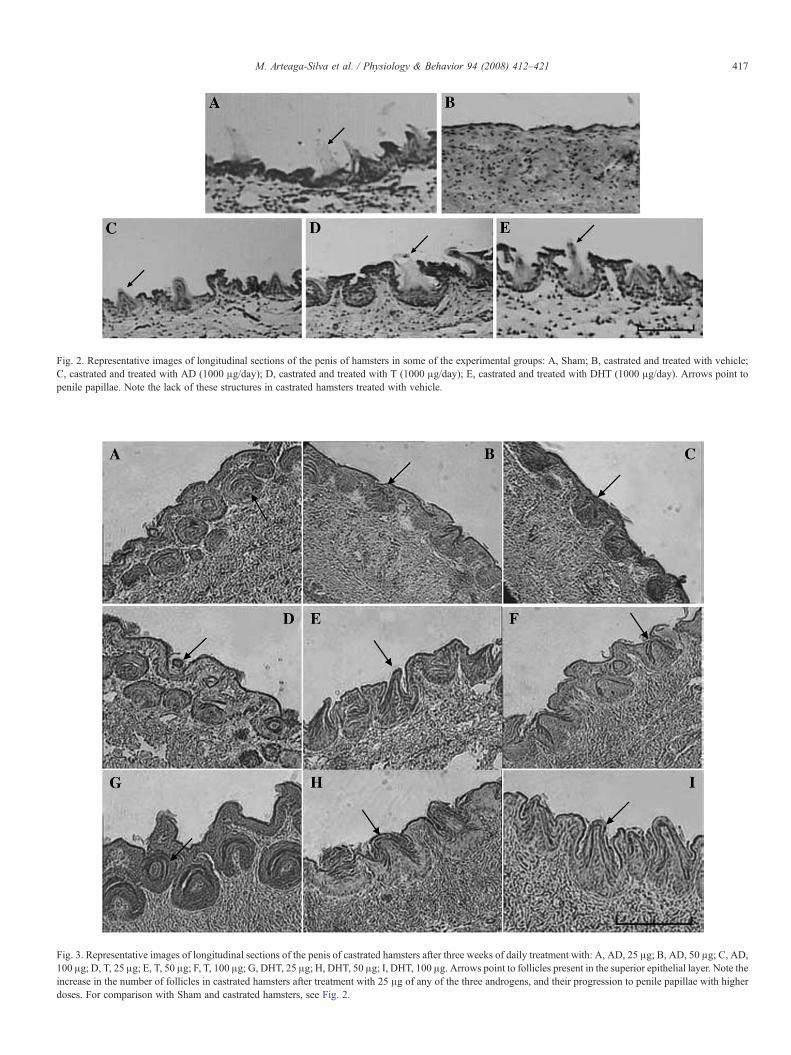
Fig. 2. Representative images of longitudinal sections of the penis of hamsters in some of the experimental groups: A, Sham; B, castrated and treated with vehicle;C, castrated and treated with AD (1000 μg/day); D, castrated and treated with T (1000 μg/day); E, castrated and treated with DHT (1000 μg/day). Arrows point topenile papillae. Note the lack of these structures in castrated hamsters treated with vehicle.
Fig. 3. Representative images of longitudinal sections of the penis of castrated hamsters after three weeks of daily treatment with: A, AD, 25 μg; B, AD, 50 μg; C, AD,100 μg; D, T, 25 μg; E, T, 50 μg; F, T, 100 μg; G, DHT, 25 μg; H, DHT, 50 μg; I, DHT, 100 μg. Arrows point to follicles present in the superior epithelial layer. Note theincrease in the number of follicles in castrated hamsters after treatment with 25 μg of any of the three androgens, and their progression to penile papillae with higherdoses. For comparison with Sham and castrated hamsters, see Fig. 2.
417M. Arteaga-Silva et al. / Physiology & Behavior 94 (2008) 412–421

418 M. Arteaga-Silva et al. / Physiology & Behavior 94 (2008) 412–421
dependent manner (Table 1), differences among groups withineach androgen tested, being statistically significant (for distalspines, AD: F(6,56)=142.90, pb0.0001; T: F(6,56)=485.17,pb0.0001; DHT: F(6,56)=427.09, pb0.0001) (for proximalspines, AD: F(6,56)=316.46, pb0.0001; T: F(6,56)=375.03,pb0.0001; DHT: F(6,56)=224.68, pb0.0001). A completerestoration occurred after daily treatment with 500 or 1000 μg ofT or DHT, and with 1000 μg of AD, to similar values as in theSham group (Fig. 2 C, D, E, Table 1).
Administration of any of the three androgens at the lowestdose (25 μg/day) stimulated the formation of follicles in thepenile epithelium, many of them located deep into the dermis,without emerging to the surface (Fig. 3A, D, G). Higher dosesof the androgens resulted in the complete development of penilefollicles, including their emergence to the surface and theformation of penile spines, DHT being more effective than ADor T, since doses of DHT as low as 50 or 100 μg/day wereeffective to promote penile spines formation (Fig. 3).
3.3. Plasma levels of steroid hormones
Table 2 shows the values of T, AD, DHT, E2, and E1 asassayed in plasma samples of animals in the Sham group, aswell as in the castrated groups receiving the various treatments.In the Sham group, AD showed the highest levels (pb0.01) ascompared to the other hormones, but E1 was undetectable.Castrated hamsters had undetectable levels of the varioushormones. AD administration to castrated hamsters causeddose-dependent increases in the plasma levels of this hormone(F(6,98)=210.12, pb0.0001), as well as those of T, DHT, E2,and E1 that were in most cases significantly higher than those ofthe Sham group (Table 2).
Table 2Plasma levels of steroid hormones in intact (Sham) and castrated (C) male hamsters
Doses T AD(μg/day) (ng/ml) (ng/ml)
Sham 2.02±0.16y 4.08±0.24y
C+VEH 0.00b 0.00b
C+AD 25 0.65±0.06a,x 1.33±0.06b,y
50 1.66±0.14y 4.31±0.30y
100 2.68±0.31y 7.56±0.40a,y
300 5.35±0.24y 10.54±0.22b,
500 16.70±1.04y 31.28±3.00b,
1000 30.44±1.29y 101.11±5.00b
C+T 25 1.50±0.11y 0.51±0.07b
50 2.37±0.20y 1.26±0.10b,y
100 4.06±0.31y 1.86±0.12b,y
300 13.80±0.50b,y 4.00±0.81y
500 40.35±0.01b,y 6.12±0.57a,y
1000 78.20±7.5b,y 13.05±0.00b,
C+DHT 25 Nd Nd50 Nd Nd100 Nd Nd300 Nd Nd500 Nd Nd1000 Nd Nd
Data correspond to mean±SEM.apb0.05 bpb0.01 vs Sham.xpb0.05 ypb0.01 vs castrated+VEH.
Similarly, administration of T to castrated hamstersresulted in dose-dependent increases of this hormone inplasma (F(6,98)=316.72, pb0.0001), as well as of AD,DHT, and E2, in most cases significantly higher than levelsfound in the Sham group. DHT administration to castratedhamsters only resulted in highly significant (F(6,98)=266.24,pb0.0001) dose-related increases of plasma levels of DHTas compared to the Sham group, but not of the other hormones(Table 2).
4. Discussion
To our knowledge, this is the first study in which acomprehensive assessment of the effects of different doses ofAD, T, and DHT, on penile growth and male sexual behavior ofcastrated male hamsters, including the display of longintromission patterns, has been done. Several studies exist onthe comparative effect of different hormones to maintaincopulation in male hamsters after castration [17–20]. On theother hand, very few, if any, exist in which the ability ofdifferent androgens to restore male sexual behavior in hamstersis compared, as in the present study, once this behavior hasdisappeared after castration. This paradigm represents astronger challenge, as evidenced by the lesser effectiveness ofvarious androgens including T, to restore rather than to maintainMSB after its decline, in several species [25,28]. Under theseconditions, a dose–response, comparative assessment of theeffectiveness of androgens to restore MSB, reveal more precisedifferences in their potency.
Under a restoration paradigm, duration of treatment withsteroid hormones, as well as the number of tests performedduring the treatment period, has been shown to significantly
after treatment with vehicle (VEH) or the various doses of AD, T , or DHT
DHT E2 E1
(ng/ml) (ng/ml) (ng/ml)
0.21±0.02y 0.08±0.07y Nd0.00b 0.00b 0.000.16±0.01 0.15±0.02a,y 0.25±0.00y
0.23±0.07y 0.17±0.02a,y 0.25±0.05y
0.37±0.06a,y 0.23±0.06b,y 0.24±0.04yy 0.55±0.04b,y 0.23±0.04b,y 0.53±0.09yy 0.75±0.05b,y 0.33±0.10b,y 1.03±0.12y,y 0.84±0.02b,y 1.26±0.13b,y 2.26±0.14y
0.24±0.06y 0.25±0.04by Nd0.33±0.03a,y 0.38±0.08b,y Nd0.54±0.02b,y 0.42±0.16b,y Nd1.09±0.07b,y 0.52±0.07b,y Nd1.86±0.92b,y 0.53±0.00b,y Nd
y 2.12±0.00b,y 1.83±0.17b,y Nd1.11±0.02b,y Nd Nd3.34±0.30b,y Nd Nd5.14±0.33b,y Nd Nd11.60±0.40b,y Nd Nd29.30±0.80b,y Nd Nd54.0±4.50b,y Nd Nd

419M. Arteaga-Silva et al. / Physiology & Behavior 94 (2008) 412–421
affect copulatory behavior. Thus, castrated hamsters treated for17 days with T display a higher level of MSB as compared tomales treated for only 7 days, and sexual performance of malesreceiving T for 17 days is better if tested three times during thisperiod, than only once at the end of treatment [29]. Inaccordance to those observations, treatment period in thepresent study lasted three weeks, and subjects were tested atweekly intervals but results were based on their display at thelast behavioral test.
Present results show that AD was more potent than T torestore MSB in castrated hamsters, since, though bothandrogens stimulated similar numbers of mounts and intromis-sions at comparable dose levels, 50 μg/day of AD stimulatedejaculatory behavior in 80% hamsters while 300 μg/day of Twere required to obtain a similar effect. In addition, longintromissions were displayed by all AD-treated hamstersreceiving 1000 μg/day while only 20% hamsters treated withthe same dose of T did. This seems to have resulted from thehigher number of ejaculations (10 as an average) displayed byAD-treated hamsters, as compared to those displayed by T-treated animals (6 as an average). These results are consistentwith those of other authors who have found AD more effectivethan T in maintaining ejaculation [17,19].
DHT in the present study, even at the highest dose,completely failed to stimulate MSB in castrated hamsters.Previous studies have shown that, while physiological doses ofDHT do not maintain MSB in castrated hamsters [17,30], highdoses (1 mg/day) are able to maintain this behavior for severalweeks after castration [21,31]. Differences between these dataand present results, showing the ability of high doses of DHT tomaintain MSB but not to restore it, may result from the decreasein density of androgen receptors occurring after castration inbrain structures related to MSB in hamsters, such as that seen inthe medial preoptic area and the bed nucleus of the striaterminalis of rats [32]. Castration has been shown to cause ashort term (four days) increase of the androgen receptor and itsmRNA, while they decrease after a long period (two months).These effects are reversed by treatment with 2 mg of DHT [32].The doses of DHT administered in the present study, and theduration of the treatment period (three weeks) may have notbeen enough to reverse the cellular changes as for MSB to bedisplayed.
A possible explanation for the effects of high doses of DHTin maintaining MSB, may be its bioconversion to non-phenoliccompounds such as 3β,5α-androstandiol able to exert estro-genic effects [33], and 3α,5α-androstandiol as an androgenicmetabolite, which may synergize and exert behavioral effects incastrated animals, as has been reported in rats [34]. Thesepossible effects of DHT seemed not to have been able to restorecopulation in hamsters three weeks after castration.
The ability of different hormones to restore long intromis-sions in castrated hamsters had not been previously evaluated.These responses appear in intact hamsters as sexual exhaustionapproaches, after displaying several ejaculations, so theseresponses allow predicting sexual satiety [12,14]. In behavioraltests only lasting for 10 to 15 min, T at a daily dose of 500 μghas been said to be as potent as AD to restore [18] and maintain
[17] MSB in hamsters after castration. However, in these shorttests only few ejaculations occur and long intromission patternsare not displayed. Present results showed AD to be moreeffective than T, since all hamsters performed long intromis-sions after treatment with 1000 μg/day AD while only 20% ofthem showed these responses after T treatment at the same dose.
The behavioral effects of AD and T may result from theirbioconversion both to 5α-reduced, and to phenolic metabolites(5α-androstanedione and DHT, as well as E1 and E2,respectively). AD and T may also interconvert to each otherand the same holds true for E1 and E2 [35,36]. These severalbiotransformations are reflected in the present study in theplasmatic levels of the various hormones assayed, in addition tothe particular hormone administered. Since DHT is convertedneither to delta-4 androgens nor to phenolic steroids, only DHTwas detected after administration of this steroid. DHT resultingeither from DHT administration or from the bioconversion ofAD and/or T, seems not to be, per se, enough to restore MSB inhamsters, since very high DHT levels were not related to thedisplay of this behavior, and even the highest dose of DHTwasunable to stimulate it. On the other hand, biotransformation of Tto AD, seems to play a significant role for stimulation of MSBin hamsters, plasma levels of AD being more related to theexpression of MSB than T levels. Indeed, doses of AD (50 μg/day or more), and doses of T (300 μg/day or more), able torestore AD levels to those of Sham animals, were also effectiveto stimulate ejaculatory behavior in most castrated subjects.
It is interesting to emphasize that, in the present study,administration of 50 μg/day of AD, which was effective torestore mounting and intromission in all castrated Ss andejaculation in most of them, resulted in plasma levels of AD, T,and DHT, similar to those of the Sham group. The display oflong intromissions, on the other hand, seems to require a higherstimulus, since this behavior was only stimulated by the highestdose (1000 μg/day) of AD and of T, eliciting a sufficientnumber of ejaculations as to result in long intromissions. Again,plasma levels of AD rather than those of T, seem to be related tothe expression of this behavioral pattern, since, in a previousstudy [20] as in the present one, higher plasma levels of AD thanT, resulting from AD administration, were related to a completerestoration of male copulation to similar levels than those ofintact animals, including long intromissions. Thus, thoseresults, as well as present ones, show AD as the most effectiveandrogen for stimulating full copulation including achievementof long intromissions and satiety.
However, not only AD, but several hormones, may play arole in the display of MSB in hamsters, since the simultaneoustreatment with T and DHT is more effective than T alone torestore MSB in castrated hamsters (M. Arteaga-Silva and G.Moralí, unpublished observations). In addition, both AD and Ttreatment in the present study resulted in higher levels of E2 thanthose of the Sham group, and AD administration, even at thelowest dose, increased E1 levels above those of Sham, intactanimals. Although some studies have explored the effects ofeither E1 or E2 alone or in combination with DHT to stimulateMSB in hamsters [19,20,24], the precise role of E1 in MSB inhamsters has not been systematically explored. Thus, the

420 M. Arteaga-Silva et al. / Physiology & Behavior 94 (2008) 412–421
possibility exists that several steroids interact with the neuralsubstrate for the expression of MSB in hamsters, as has beensuggested for the integrative hormonal regulation of MSB in therat and other species [25].
As reported in many species, and also in hamsters by otherauthors [5], castration resulted in the present study in areduction of penile size and weight, and in penile epitheliumatrophy with regression of follicles and spines. As has beendescribed in the rat [4], the behavioral effects of AD, T, andDHT did not correlate in the present study, with their potency tostimulate penile growth and penile epithelium in castratedhamsters. Thus, though DHTwas the most effective androgen torestore penile growth and penile spines development, it did notstimulate MSB. On the other hand, AD had the higher potencyto restore the occurrence of ejaculations and long intromissions,though having the lowest potency to stimulate penile weight,length, and spines formation.
However, though DHTwas more effective than AD and T torestore the number of penile spines to values similar to those ofthe Sham group, all androgens, even at the lowest dose level,stimulated the development of penile follicles and at higherdoses induced the formation of spines. The histological analysisof penile samples in tissue sections instead of their observationunder the scanning electron microscope, made possible torecognize the presence of follicles in hamsters receiving thelowest dose of the different androgens, even when located deepinto the dermis, without emerging to the surface. T and AD perse or through its conversion to other metabolites may induce theformation of spines. However, their bioconversion to DHT maybe responsible for this effect, as pointed out by other authors[25], and suggested by the plasma levels of DHT found after Tor AD administration.
It is interesting to note that the effects of the variousandrogens on penile weight differed according to their effectson penile length and width. Thus, AD was as affective as DHTto stimulate penile width, but it did not restore, even at thehighest dose, penile length to the level of the Sham group. Onthe other hand, T was less effective than AD and DHT inrestoring penile width, but more effective than AD to restorepenile length. This observation may represent differentialeffects of these androgens on the various cellular componentsof the penis.
It has been suggested for other species [4] that penile spinescontribute to vaginal stimulation provided by the male atcopulation, promoting neuroendocrine responses that preparethe uterus for pregnancy. However, in hamsters the occurrenceof long intromissions has been interpreted as relevant forvaginal stimulation to initiate these neuroendocrine responsesallowing ovulation and pregnancy to occur [15,16], besidesother sensory modalities provided by the male. The relativecontribution of penile spines for these processes in hamsters hasnot been elucidated.
In summary, the main contributions of the present study are:1) The dose–response comparative assessment of the effective-ness of AD, T, and DHT for reinstating MSB, revealing thehigher potency of AD as compared to T; 2) The assessment ofthe behavioral effects of AD, T, or DHT under the restoration
paradigm, once MSB has disappeared several weeks aftercastration, rather than the maintenance paradigm, as has beenmore often tested; 3) The dose–response comparative assess-ment of the effects of the androgens on the measure of thepenis and in the number of spines, showing that doses as low as25 µg/day stimulated the formation of penile follicles.
The relative contribution of AD and its various metaboliteson the restoration of MSB in castrated hamsters deserves furtherinvestigation.
References
[1] Aronson LR, Cooper, M.L.. Penile spines of the domestic cat: theirendocrine-behavior relations. Anat Rec 1967;157(1):71–8.
[2] Johnson RD, Halata Z. Topography and ultrastructure of sensory nerveendings in the glans penis of the rat. J Comp Neurol 1991;312(2):299–310.
[3] Kanagawa H, Hafez ES. The penis. In: Hafez ES, editor. Scanning ElectronMicroscopic Atlas of Mammalian Reproduction. Stuttgart: Thieme; 1975.
[4] O'Hanlon JK, Sachs BD. Fertility of mating in rats (Rattus norvegicus):contributions of androgen-dependent morphology and actions of the penis.J Comp Psychol 1986;100(2):178–87.
[5] Whitsett JM, Ayer ML, Muse KE. Androgenic control of phallic papillae inhamsters: a quantitative analysis using the scanning electron microscope.Biol Reprod 1980;23:669–76.
[6] Feder HH. The comparative actions of testosterone propionate and 5-androstan-17-ol-3-one propionate on the reproductive behaviour, physiol-ogy and morphology of male rats. J Endocrinol 1971;51(2):241–52.
[7] Phoenix CH, Copenhaver KH, Brenner RM. Scanning electronmicroscopy of penile papillae in intact and castrated rats. Horm Behav1976;7(2):217–27.
[8] Taylor GT, Weiss J, Komitowski D. Reproductive physiology andpenile papillae morphology of rats after sexual experience. J Endocrinol1983;98(2):155–63.
[9] Taylor GT, Komitowski D, Weiss J. Light and scanning electronmicroscopic study of testosterone-restored penile papillae in castratedrats. Anat Rec 1983;205(3):277–86.
[10] Sachs BD, Glater GB, O'Hanlon JK. Morphology of the erect glanspenis in rats under various gonadal hormone conditions. Anat Rec1984;210(1):45–52.
[11] Kierniesky NC, Gerall AA. Effects of testosterone propionate implants inthe brain on the sexual behavior and peripheral tissue of the male rat.Physiol Behav 1973;11(5):633–40.
[12] Bunnell BN, Boland BD, Dewsbury DA. Copulatory behavior of goldenhamster (Mesocricetus auratus). Behaviour 1976;61:180–206.
[13] Arteaga M, Moralí G. Characteristics of the motor and genital copulatoryresponses of the male hamster. J Physiol (París) 1997;91(6):311–6.
[14] Arteaga M, Motte-Lara J, Velázquez-Moctezuma J. Temporal pattern ofrecovery from sexual satiety in male golden hamster (Mesocricetusauratus). Physiol Behav 2000;68:591–4.
[15] Huck UW, Lisk RD. Determinants of mating success in the goldenhamster (Mesocricetus auratus): I. Male capacity. J Comp Psychol1985;99(2):98–107.
[16] Huck UW, Lisk RD. Determinants of mating success in the goldenhamster (Mesocricetus auratus): II. Pregnancy initiation. J Comp Psychol1985;99(2):231–9.
[17] Whalen RE, DeBold JF. Comparative effectiveness of testosterone,androstendione and dihydrotestosterone in maintaining mating behaviorin the castrated male hamster. Endocrinology 1974;95:1674–9.
[18] Christensen LW, Coniglio LP, Paup DC, Clemens LG. Sexual behavior ofmale golden hamsters receiving diverse androgen treatments. Horm Behav1973;4:223–9.
[19] Tiefer L. Gonadal hormones and mating behavior in the adult goldenhamster. Horm Behav 1970;1:189–202.
[20] Arteaga-Silva M, Márquez-Villanueva Y, Martínez-García R, Hernández-González M, Bonilla-Jaime H, Retana-Márquez S. Effects of hormonalreplacement with androgens and estrogens on male sexual behavior and

421M. Arteaga-Silva et al. / Physiology & Behavior 94 (2008) 412–421
plasma levels in gonadectomized golden hamster (Mesocricetus auratus).Physiol Behav 2005;85:571–80.
[21] Romeo RD, Cook-Wiens E, Richardson HN, Sisk CL. Dihydrostestoster-one activates sexual behavior in adult male hamsters but nut in juveniles.Physiol Behav 2001;73:579–84.
[22] Lisk RD, Bezier JL. Intrahypothalamic hormone activation of sexualbehavior in the male hamster. Neuroendocrinology 1980;30:220–7.
[23] Wood RI. Estradiol, but not dihydrotestosterone, in the medial amygdalafacilitates male hamster sex behavior. Physiol Behav 1996;59:833–41.
[24] Lisk RD, Greenwald DP. Central plus peripheral stimulation by androgenis necessary for complete restoration of copulatory behavior in the malehamster. Neuroendocrinology 1983;36:211–7.
[25] Luttge WG. Endocrine control of mammalian male sexual behavior: ananalysis of the potential role of testosterone metabolites. In: Beyer C,editor. Endocrine Control of Sexual Behavior. New York: Raven Press;1979.
[26] Woodward C, Emery P. Determination of plasma corticosterone using highperformance liquid chromatography. J Chromatogr 1987;419:280–4.
[27] Sachs BD. Role of striated penile muscles in penile reflexes, copulation,and induction of pregnancy in the rat. J Reprod Fertil 1982;66(2):433–43.
[28] Davidson JM. Characteristics of sex behavior in male rats followingcastration. Anim Behav 1966;14:266–72.
[29] Schulz KM, Richardson HN, Zehr JL, Osetek AJ, Menard TA, Sisk ChL.Gonadal hormones masculinize and defeminize reproductive behaviorsduring puberty in the male Syrian hamster. Horm Behav 2004;45:242–9.
[30] DeBold JF, Clemens LG. Aromatization and the induction of male sexualbehavior in male, female, and androgenized female hamsters. Horm Behav1978;11:401–13.
[31] Payne AP, Bennett NK. Effects of androgens on sexual behaviourand somatic variables in the male golden hamster. J Reprod Fertil1976;47:239–44.
[32] Handa RJ, Kerr JE, DonCarlos LL, McGivern RF, Hejna G. Hormonalregulation of androgen receptor messenger RNA in the medial preopticarea of the male rat. Brain Res Mol 1996;39:57–67.
[33] Thieulant ML, Benie T, Michaud S, Klein H, Vessieres A. Binding andeffects of 5α-androstane-3,17-β-diol in the male rat pituitary. J SteroidBiochem 1983;19:241–6.
[34] Moralí G, Oropeza MV, Lemus AE, Perez-Palacios G. Mechanismsregulating male sexual behavior in the rat: role of 3 alpha- and 3 beta-androstanediols. Biol Reprod 1994;51:562–71.
[35] Oshima H, Ochi AI. On testicular 17β-hydroxysteroid oxidoreductase:product activation. Biochim Biophys Acta 1973;306:227–35.
[36] Bogovich K, Payne AH. Purification of rat testicular microsomal 17β-ketosteroid reductase. J Biol Chem 1980;255:5552–8.
Copyright © 2022 FDOKUMEN




















