
Determinants of Mating Success in the Golden Hamster (Mesocricetus auratus ): I. Male Capacity
19
Anim. Behav., 1986, 34, 971-989 Determinants of mating success in the golden hamster (Mesocricetus auratus): Social dominance and mating tactics under seminatural conditions U. WILLIAM HUCK*, ROBERT D. LISK*, JON C. ALLISON* & C. G. VAN DONGENt *Department of Biology, Princeton University, Princeton, N.J. 08544, U.S.A. tBio-Research Institute, 9 Commercial Avenue, Cambridge, MA 02141, U.S.A. Abstract. We tested the hypothesis that high social status leads to high levels of copulatory behaviour and increased number of offspring in hamsters. When paired with albino female golden hamsters, males of three genotypic strains showed differences in the frequency and temporal patterning of some measures of copulatory behaviour but did not differ in general fertilization success. The results of two-male, one- female tests in which mating order and ejaculation frequencies were controlled indicated only relatively small differences in differential fertilizing capacity among these genotypes. Males of the genotypic strains displayed an equal probability of achieving alpha status in three-male, one-female competitive mating tests in which animals were habituated to semi-natural enclosures. Alpha males often slept in the female's nest box the night before she became receptive, were the first to mate on the following morning, achieved higher scores for all measures of sexual behaviour including ejaculation frequency and subsequently sired more young than did lower ranking males. However, the differential reproductive success of alpha males was not simply a function of higher ejaculation scores since they obtained more than twice as many young per ejaculation as subordinate males. Alpha males continued to copulate with long intromissions after exhausting their supply of ejaculates. Although females mated with several males, mating preferences were evidenced by their flank-gland and vaginal marking patterns on the day before receptivity and in their initial Choice of mates during tethered male tests. The behaviour of alpha males is consistent with previous findings regarding the effects of mating order and prolonged copulation on sperm competition and suggests that sperm utilization patterns and mating tactics have coevolved. The assertion that a male's dominance status is correlated with copulatory behaviour and differen- tial production of offspring is firmly embedded in the literature (e.g. Zuckerman 1932; Maslow 1936; Wilson 1975; Barash 1982). However, in a recent review of the subject, Dewsbury (1982) pointed out that three fundamental steps are required to demonstrate this hypothesis: (1) dominance must be defined and measured; (2) individual differences in copulatory behaviour must be measured and related to differences in dominance status; and (3) differential reproduction must be measured and related to an individual's social status and copula- tory behaviour. Although much research has focused on the relationship between dominance status and copulatory behaviour (cf. Dewsbury 1982; Huck & Banks 1982a, b; Chapais 1983; Lisk & Baron 1983) or status and reproductive success (cf. Dewsbury 1982), few researchers have attempted to demonstrate all three steps in a single study. In fact, of the nearly 100 studies included in his review, Dewsbury (1982) found only two in which dominance status, copulatory behaviour and differential reproduction were reported. Guhl & Warren (1946) found a clear association among all three factors in farm-yard flocks of white leghorn chickens and Dewsbury (1981) found that in large laboratory enclosures, dominant male deer mice (Peromyscus maniculatus) sired more off- spring than subordinates, although the statistical significance of this effect was marginal. The relation between dominance status and reproductive success may be complicated in situa- tions where females mate with more than one male during a single mating period. Under such condi- tions, two or more males may be represented in a given litter and the effect of social status may be reflected in the relative proportion of young sired rather than as an all-or-none phenomenon. Among mammals, field observations indicate that multi- ple-male matings occur in species as diverse as lions (Bertram 1976) and Norway rats (Telle 1966; Robitaille & Bovet 1976). Furthermore, electro- phoretic analysis of blood proteins has confirmed 971
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Determinants of Mating Success in the Golden Hamster (Mesocricetus auratus ): I. Male Capacity

Anim. Behav., 1986, 34, 971-989
Determinants of mating success in the golden hamster (Mesocricetus auratus): Social dominance and mating tactics under seminatural conditions
U. W I L L I A M H U C K * , R O B E R T D. LISK*, JON C. A L L I S O N * & C. G. VAN D O N G E N t
*Department of Biology, Princeton University, Princeton, N.J. 08544, U.S.A. tBio-Research Institute, 9 Commercial Avenue, Cambridge, MA 02141, U.S.A.
Abstract. We tested the hypothesis that high social status leads to high levels of copulatory behaviour and increased number of offspring in hamsters. When paired with albino female golden hamsters, males of three genotypic strains showed differences in the frequency and temporal patterning of some measures of copulatory behaviour but did not differ in general fertilization success. The results of two-male, one- female tests in which mating order and ejaculation frequencies were controlled indicated only relatively small differences in differential fertilizing capacity among these genotypes. Males of the genotypic strains displayed an equal probability of achieving alpha status in three-male, one-female competitive mating tests in which animals were habituated to semi-natural enclosures. Alpha males often slept in the female's nest box the night before she became receptive, were the first to mate on the following morning, achieved higher scores for all measures of sexual behaviour including ejaculation frequency and subsequently sired more young than did lower ranking males. However, the differential reproductive success of alpha males was not simply a function of higher ejaculation scores since they obtained more than twice as many young per ejaculation as subordinate males. Alpha males continued to copulate with long intromissions after exhausting their supply of ejaculates. Although females mated with several males, mating preferences were evidenced by their flank-gland and vaginal marking patterns on the day before receptivity and in their initial Choice of mates during tethered male tests. The behaviour of alpha males is consistent with previous findings regarding the effects of mating order and prolonged copulation on sperm competition and suggests that sperm utilization patterns and mating tactics have coevolved.
The assertion that a male's dominance status is correlated with copulatory behaviour and differen- tial production of offspring is firmly embedded in the literature (e.g. Zuckerman 1932; Maslow 1936; Wilson 1975; Barash 1982). However, in a recent review of the subject, Dewsbury (1982) pointed out that three fundamental steps are required to demonstrate this hypothesis: (1) dominance must be defined and measured; (2) individual differences in copulatory behaviour must be measured and related to differences in dominance status; and (3) differential reproduction must be measured and related to an individual's social status and copula- tory behaviour. Although much research has focused on the relationship between dominance status and copulatory behaviour (cf. Dewsbury 1982; Huck & Banks 1982a, b; Chapais 1983; Lisk & Baron 1983) or status and reproductive success (cf. Dewsbury 1982), few researchers have attempted to demonstrate all three steps in a single study. In fact, of the nearly 100 studies included in his review, Dewsbury (1982) found only two in
which dominance status, copulatory behaviour and differential reproduction were reported. Guhl & Warren (1946) found a clear association among all three factors in farm-yard flocks of white leghorn chickens and Dewsbury (1981) found that in large laboratory enclosures, dominant male deer mice (Peromyscus maniculatus) sired more off- spring than subordinates, although the statistical significance of this effect was marginal.
The relation between dominance status and reproductive success may be complicated in situa- tions where females mate with more than one male during a single mating period. Under such condi- tions, two or more males may be represented in a given litter and the effect of social status may be reflected in the relative proportion of young sired rather than as an all-or-none phenomenon. Among mammals, field observations indicate that multi- ple-male matings occur in species as diverse as lions (Bertram 1976) and Norway rats (Telle 1966; Robitaille & Bovet 1976). Furthermore, electro- phoretic analysis of blood proteins has confirmed
971

972 Animal Behaviour, 34, 4
multiple paternity in litters of deer mice (Birdsall & Nash 1973; Merritt & Wu 1975) and Belding's ground squirrels (Hanken & Sherman 1981).
Most laboratory studies relating dominance status to differential reproduction have employed males of strains differing in some marker gene, usually one for coat colour (e.g. Levine 1958; Levine & Lascher 1965; DeFries & McClearn 1970, 1972; Horn 1974; Dewsbury 1981). While this procedure facilitates determination of paternity patterns, it also necessitates adequate measures to control for potential strain differences in general fertilization success and differential fertilizing capacity. General fertilization success refers to a male's fertility in non-competitive situations (Dewsbury 1982). For example, Horn (1974) noted that when males of some strains of house mice were placed alone with females, they sired fewer off- spring than males of other strains. Lanier et al. (1979) defined differential fertilizing capacity as 'the relative ability of males of a given strain to produce offspring when both their sperm and those of another strain are deposited in the same female reproductive tract' (page 783). For example, when ejaculations from male hamsters of an agouti a n d cream strain were alternated (thus minimizing mating order effects), agouti males sired over 90~ of the young (Huck et al. 1985). Strain differences in differential fertilizing capacity have also been reported in insects (e.g. Lefevre & Jonsson 1962; Prout & Bundegaard 1977), chickens (Martin et al. 1974) and in several species of mammals, including rabbits, Oryctolagus euniculus (Dziuk 1965), house mice, Mus musculus (Edwards 1955; Levine 1967), Norway rats, Rattus norvegicus (Adler & Zoloth 1970; Lanier et al. 1979; Dewsbury & Hartung 1980), deer mice, Peromyscus maniculatus (Dewsbury & Baumgardner 1981), and prairie voles, Microtus ochrogaster (Dewsbury & Baumgardner 1981).
In the present study we examined the relation between dominance status, patterns of copulatory behaviour and differential reproduction in the golden hamster (Mesocricetus auratus). This spe- cies seemed particularly appropriate for such a study. Based on a review of available field studies on this genus, Murphy (1977) concluded that golden hamsters live alone in simple burrow sys- tems. Social contact among adults occurs primarily during the breeding season when the female allows the male around or in her burrow for a brief period of mating. A trapping study of the Rumanian hamster (M. neutoni) indicated that the home
ranges of several males might overlap that of a single female by 50~ or more (see Murphy 1977). This suggests that several males will be familiar with the burrow of a single female and, thus, male competition for access to the female may occur. Studies in large laboratory enclosures indicate that sexually receptive females which have actively terminated mating with one male will respond in succession to two or three additional males (Lisk & Baron t982, 1983).
Male golden hamsters readily establish stable dominance relationships even when interactions are limited to relatively brief (15-30 min) daily encounters (Whitsett 1975). Male dominance sta- tus did affect mating success in a three-male, one- female competitive mating situation in large enclo- sures, although the alpha males did not maintain exclusive control of the females and one or both of the subordinate males also copulated (Lisk & Baron 1983). Paternity patterns were not deter- mined in that study, however.
In this study three strains of males were used, each homozygous for a different coat colour gene. Thus, in a three-male, one-female competitive mating situation, the relative reproductive success of each genotype could be determined within 48 h of the birth of offspring. In experiment 1 we determined whether males of the three genotypes differed in their copulatory behaviour or general fertilization success in one-male, one-female mat- ing tests. We next examined differential fertilizing capacity in two-male, one-female tests in which the order of mating and number of ejaculations were controlled (experiment 2). Although some differen- tial fertilizing capacity was detected, it was of a relatively small magnitude, allowing us to proceed to the main question in experiment 3, namely: is there a significant reproductive advantage which accrues to the alpha male in a competitive mating situation? In order to more closely approximate natural conditions, each member of an experimen- tal group (three males and one female) was adapted to individual areas within a large, complex enclo- sure before the males were allowed to interact and establish dominant-subordinate relationships. In a subsequent mating test, copulatory behaviour was continuously recorded from the onset of the female's period of sexual receptivity until she had repeatedly attacked all three males. In experiment 4 we focused our attention on male and female scent- marking patterns and their possible role in mate selection.

Huck et al.: Dominance and differential reproduction 973
GENERAL M E T H O D S
Animals and Husbandry
All subjects were obtained from Bio-Research Consultants, Cambridge, Massachusetts and were maintained in our laboratory for at least 6 weeks before testing. A total of 37 females of the BIO- 86-93N (albino) strain, 90585 days old 0 ( = 157-6) and weighing 98-162 g ()(= 126.1), were used. Each was allowed at least 40 days between testing. Stage of the oestrous cycle was determined by extravagi- hal examination using procedures described by Orsini (1961). Each of the three male strains exhibited a distinctive coat colour and, when mated with albino females, produced correspondingly distinctive young. Specifically, males of the BIO- 1-5N strain exhibited white coat colour with black ears and produced young of the same colour. BIO- 7-88 males exhibited beige coat colour and pro- duced beige young. Lastly, males of the BIO- 15.16 strain exhibited brown coat colour and produced brown male and tawny female offspring when mated to albino females. A total of 111 males (37 of each strain), 128-290 days of age ( 2 = 173-7) and weighing 94--157 g ( 2 = 122.7), were used. Males were allowed at least 2 weeks between tests.
Animals were maintained on a reversed 14-h- L:10-h-D photoperiod with the dark (red light) phase beginning at 1200 hours. When not being tested, each subject was individually housed in a 35 x 20 x 17.5-cm rack-mounted stainless steel cage. Pregnant females were transferred to 38 x 30 x 17-cm plastic breeding boxes with wood shavings for bedding. Commercial laboratory chow and water were continuously available.
Recording Techniques
During each test, the time of onset and termina- tion of each behavioural act was recorded on magnetic tape using a microswitch keyboard con- nected to a timer which interfaced with a Hewlett- Packard 9815A calculator. At the termination of testing, the data were printed and retained for use in statistical analysis.
EXPERIMENT 1
Copulatory Behaviour and General Fertilizing Ability
Quantitative descriptions of the male copulatory
pattern are available for several species of rodents (reviewed by Dewsbury 1972, 1975, 1979a). While stereotypy of the copulatory pattern among con- specific individuals is usually high, quantitative examination of the copulatory behaviour of sub- species (Dewsbury 1979b) or inbred strains within a species (e.g. McGill 1962; Levine et al. 1966; Oglesby et al. 1981) often reveals significant differ- ences in the temporal relationships between most parameters of the copulatory pattern. Insofar as such differences may affect mating success, it is clear that before one can employ genetic strains as a convenient way to label paternity in studies designed to test for the effects of social dominance on reproductive success, one must first determine the quantitative mating pattern of each of the strains as well as their respective general fertilizing abilities with females of the strain used in the study. In experiment 1, such data were obtained for males of the genotypes used in the present study by pairing males with albino females in one-male, one- female non-competitive mating tests.
Methods
Subjects were 36 males (12 of each strain) and 36 albino females. Each female had one litter before being used in this study. The sexual experience of the male consisted of a single mating (to five ejaculations) with an ovariectomized, LVG-LAK (an outbred agouti strain) female in hormone- induced oestrus. To facilitate observation, tests were conducted in a circular Plexiglas arena, 35 cm in diameter. Males were allowed 5 rain to habituate before the introduction of a female and tests were terminated either after the attainment of a 30-min satiety criterion, during which no copulatory beha- viour was observed, or after the female attacked the male. All tests occurred between 1300 and 1500 hours. After testing, females were placed into breeding boxes and litters checked at parturition.
Copulatory behaviour in the hamster consists of mounts (without intromissions), short intromis- sions (2-3 s vaginal penetration without sperm transfer) and ejaculations. These are organized into discrete series, with each series terminating in an ejaculation. After 9-14 ejaculatory series, males show a modified copulatory pattern which consists of prolonged intromissions lasting 10-30 s during which the male shows intravaginal thrusting but no sperm transfer (Bunnell et al. 1977; Huck & Lisk 1985a, b). The following standard measures of

974 Animal Behaviour, 34, 4
copula tory behaviou_r (defined by Bunnell et al. 1977) were recorded: moun t latency and frequency, shor t in t romiss ion latency and frequency, ejacula- t ion latency, mean in ter- in t romiss ion interval, post-ejaculatory interval, e jacula t ion frequency, and long in t romiss ion frequency.
Results
Sexual behav iour scores for males of the three strains are summarized in Table I. Appreciable strain differences are apparent . Whi te and beige males ini t iated copulat ion significantly faster than b rown males. Brown males also required more shor t int romissions to achieve successive ejacula-
t ions and obta ined longer ejaculat ion latencies t han ei ther white or beige males. Mean inter- in t romiss ion intervals were shorter for white males than for males of the other strains, and post- e jaculatory intervals tended to be shor tes t for white males, in termediate for beige males, and longest for the b rown strain. Whi te males achieved signifi- cantly higher scores for total n u m b e r of long in t romiss ions but lower total short in t romiss ion scores than males of the other strains. However, there were no significant differences in the n u m b e r of females successfully impregnated (all females subsequent ly delivered young) or in mean litter sizes.
Table i. Mean copulatory behaviour scores of males of three genetic strains mated to satiety with albino females
White Beige Brown Ejaculatory
Measure series Mean ,+ SEM F-test SNK-test~
Short intromission latency - - 76.9+6-7 111-8-+12-1 165-3-+20-I 12.34"** B r > B e > W
Shortintromission 1 7-7+0-5 11.1__+ 0.7 14-8-+ 0.9 23.68*** B r > B e > W 2 2.6__+0.3 3-4_+ 0-3 5'0_+ 0.5 10.32"** Br>Be, W 3 2.8-+0.3 3.7_+ 0.3 4.4_+ 0.5 3.93* B r > W 4 2.9-+0.3 2.8_+ 0.3 4.9_+ 0.9 4.19" Br>Be, W 5 2.7_+0.3 2.5+ 0.3 4.0_+ 0.7 2.99Ns - -
Ejaculationlatency 1 75.6_+6.6 154.6_+14.2 224.3+13.1 39.93*** B r > B e > W 2 21.8-+4.2 57-9_+ 7.7 66.8-+ 8.0 12.02"** Br, Be>W 3 30.6_+3.1 50.2__+ 3.8 67.7___li.4 6.70** B r > W 4 32.5-+3.8 45.8-+ 5-2 78.5_+15.8 5.75** Br>Be, W 5 32.3-+3.5 44.9-+34.9 84.5___18.6 5.86** Br>Be, W
Mean inter-intromission 1 10.3_+0.6 14.7-/- 1-3 16.9_+ 0.8 13.11"** Br, B e > W interval 2 7.9+0.6 12-7-+ 0.8 13.8-+ 0.8 18.95"** Br, Be>W
3 11.5-+0.8 14-3-+ 0-4 15.8_+ 0-9 9.47*** Br, Be>W 4 12.3_+0.7 16-9_+ 0.7 17.6+ 0.6 19.21"** Br, B e > W 5 13.4_+1.1 19.1_+ 1.1 20.5_+ 0.8 13.28"** Br, Be>W
Post-ejaculatory 1 34.3+1.7 41.3_+ 2'0 44.2+ 1.6 7"96** Br, Be>W interval 2 38.2_+ 1.8 45.1__ 3.0 69.8_+ 4.0 29.62*** Br>Be, W
3 41.5+4.1 48.5_+ 2.9 67.8_+ 3.3 15.50"** Br>Be, W 4 44.4_+2.7 58.2-+ 4-3 63.4_+ 4.5 6.22** Br,Be>W 5 48-1-+2.3 55.8+ 6.0 66.3-+ 4.2 4.24* B r > W
Total mounts - - 21.8_+2.5 25.2-+ 2.4 22.5+ 2.3 0.54Ns - - Totalsfiortintromissions - - 33-6_+2.1 42.2_+ 2.5 53-5_+ 2.2 19.32"** B r > B e > W Total ejaculations - - 10.9_+0.1 9.8+ 0.4 9.8_+ 0.3 2.30Ns - - Total long intromissions - - 20.8+2.8 14.9_+ 2.l 13,3_+ 1.3 3-40* W>Be, Br Littersize - - 8.8+_0.8 8.8-+ 0.5 8-4_+ 0-8 0.11Ns - -
t Br = brown, Be = beige, W = white. * P<0.05; ** P<0.01; *** P<0"001; NS: not significant.

Huck et al.: Dominance and differential reproduction 975
Discussion
While males of the three genetic strains showed significant differences in the frequencies of specific behavioural measures as well as in their temporal patterning, there were no genotype-related effects on reproductive success. All females produced litters when mated with males of the various genotypes and mean litter sizes were similar for each of the strains. Therefore, the general fertilizing ability of the three strains appears equivalent, making them useful for further study of the relationship between copulatory behaviour and differential reproduction.
EXPERIMENT 2
Sperm Competition and Differential Fertilizing Capacity
Males of genotypes which show no differences in general fertilizing ability may still differ with respect to differential fertilizing capacity (Dews- bury 1982). In order to test directly for differential fertilizing capacity, it is necessary to minimize, as much as possible, the potential confounding effects of mating order and ejaculation frequency. To achieve this, ejaculations from two males of differ- ent genotype were alternated until each male had achieved five ejaculatory series.
Methods
Two test arenas, similar to those used in the previous experiment and each containing a male of a different strain, were placed next to each other. After allowing the males to habituate for 5 min, an albino female was moved from one male to the other after each had ejaculated once. This was continued until the female had received five ejacu- lations from each male. To control further for mating order effects in each treatment condition (see Table II), half o f the females received their first ejaculation from the male of one of the strains while the other half began with a male of the other strain. For example, of the 20 females in the W/Be x 5 condition, 10 received their first ejaculation from a white male and the other 10 began with a beige male. A total of 30 females (each had one or two previous litters) and 60 males (20 of each strain) were employed. Test order was random and each subject was used twice.
Results
Mean litter size for each treatment condit ion and the mean number and percentage of offspring sired by males of each genotype are summarized in Table 1I. The latter variable served as the basis for statistical comparisons of litter composit ion and was obtained by calculating the percentage of each litter sired by the male of a particular genotype and
Table II. Results of tests of differential fertilizing capacity among white (W), beige (Be), and brown (Br) males when mated to albino females
Treatment condition
Measure W/Be x 5* W/Br x 5 Be/Br x 5
No. litters 20 Mean litter size _+SEM 8"9_+0'5 Mean no. young sired by (W)5.4+0.6
male in parentheses Mean no. young sired by (Be)3-5 _ 0.5
male in parentheses Mean % young sired by (W)60.2_+5.4
male in parenthesest
20 20 8.8_+0.4 8.7_+0.4
(W)5.3 _ 0.5 (Be)4-4 _ 0-5
(Br)3.5 • 0.4 (Br)4.3 _+ 0.5
(W)61.2 +4.7 (Be)50-6• 5.2
* White and beige males alternated ejaculations until each had ejaculated five times; half of the females received their first ejaculation from a white male and half began with a beige male.
t White males obtained a significantly greater percentage of young when paired with beige males (Wilcoxon signed-ranks test, Z = 1.96, P < 0.05) or brown males (Z = 2.03, P < 0.05). The percentage of young obtained by beige and brown males in the Be/ Br x 5 condition did not differ significantly (Z= 0'67).

976 Animal Behaviour, 34, 4
then taking the mean ~f those values. Ejaculation order did not significantly affect litter composition in any of the treatment conditions and, thus, mating order subgroups were combined before further analysis. White males obtained a signifi- cantly greater percentage (about 60%) of young when competing with males of the beige or brown strain. However, there was no evidence for the existence of differential fertilizing capacity between beige and brown males since each strain sired about half of the young in the Be/Br x 5 treatment condition.
Discussion
Comparison of pregnancy rates and litter sizes in experiments 1 and 2 indicate that neither variable is affected when females receive alternate ejaculations from two males of different genotypes. However, the results reported here do indicate that the differential fertilizing capacity of males of the three strains is not equal. Males of the white genotype sired a significantly greater proportion of young when paired with males of the other two strains. Findings of unequal differential fertilizing capacity among genetic strains have been reported in several species (see above) and previous studies suggest that such differences are not simply a function of unequal sperm numbers (Edwards 1955; Beatty 1960; Dewsbury & Baumgardner 1981). While the strain differences in differential fertilizing capacity reported in this experiment must be taken into account in any subsequent discussion of differen- tial representation of genotypes following multiple male matings, they are not so large as to preclude the use of these strains in the following experiment.
E X P E R I M E N T 3
Copulatory Behaviour and Differential Reproduc- tion in a Three-male, One-female Competitive Situa- tion
Since the female golden hamster is more aggres- sive than the male and usually dominant in most social situations (Dieterlen 1959; Lawlor 1963; Payne & Swanson 1970; Takahashi & Lisk 1983), one might expect her to play an active role in mating activities. Indeed, we have already men- tioned that females which have actively terminated mating with one male will solicit additional copula-
tions fi'om different males (see above). When a female and several males are adapted to large enclosures, the female regulates not only the amount of mating with each male but also the location in which it occurs (Lisk & Baron 1982). Furthermore, Lisk & Baron (1983) found that male dominance status could affect mating success but only if interactions between the males and female occurred before the onset of receptivity. In four- male, one-female tests, establishment of a domi- nance hierarchy in the female's absence did not result in any competitive mating advantage for the dominant male. However, establishment of the dominance hierarchy in the presence of a female, with interactions between males and female occur- ring throughout the 4-day oestrous cycle prior to the female's receptive period, resulted in a clear mating priority for the dominant male.
These observations suggest that copulatory behaviour in a competitive mating situation is best investigated in large enclosures in which subjects are habituated to respective 'home areas' and allowed to establish social relationships for several days before the onset of sexual receptivity. The semi-natural testing paradigm described below meets these criteria and, along with the use of males with genetic markers, allows examination of the relation between dominance status, patterns of copulatory behaviour and differential reproduc- tion.
Methods
Behaviour tests were conducted in two identical 251 •215 150-cm concrete enclosures with a front wall of glass. Each enclosure was partitioned to accommodate four hamsters, one female and three males (one of each strain). The general arrangement of the enclosure was similar to that described by Lisk & Baron (1983). Masonite boards, 30.5 cm high, were used to separate the three parallel areas (160x73 cm) in which the males were confined. A clear Plexiglas divider formed the front wall of the male areas. Gates operated by wire and cord allowed access between male and female areas. Straw was scattered over the floor throughout the enclosure and the female's area also contained a clear plastic nest box.
Subjects were 75 males (25 of each strain) and 25 virgin albino females. The pre-testing sexual exper- ience of the males was identical to that described for male subjects in experiment 1. Three males, one

Huck et al.: Dominance and differential reproduction 977
of each strain matched for body weight (+ 3 g), and a single female were placed into the enclosure on pro-oestrus (the female's day of receptivity). The gates remained closed during the ensuing 4-day habituation period (one oestrous cycle). On the day following pro-oestrus (day 5 of the experiment), the female was removed from her area at 1300 hours and kept in a plastic box for 1 h. During this time the gates were opened and the males allowed to interact. The following agonistic behaviours were recorded: attack, chase, flee, tail-up, on-back, and fight (described by Grant & Mackintosh 1963). Chi-squared analyses of combined chase-attack scores were used to determine dominanWsubordi- nate relationships among the males. After 1 h, the males and females were returned to their respective areas and the gates closed. At 1300 hours on the following day, the gates were opened for the remainder of the experiment. Interactions between the four animals were observed for the next 1 h and the occurrence of agonistic behaviours recorded. The animals were similarly observed between 1300 and 1400 hours on days 7 and 8.
The female was again receptive on day 9 and behavioural observations began at 0700 hours and continued uninterrupted until 30 min had passed without copulatory behaviour (usually by 1200- 1400 hours). In addition to the previously listed measures of aggression, the following sexual beha- viours were recorded: the frequency, duration, and longest bouts of lordosis with each of the three males, and the frequency of mounts, short intromissions, long intromissions, and ejacula- tions. After testing, females were placed into plastic breeding boxes where they remained undisturbed until parturition.
Resul ts
Establishment o f dominance relationships In all 25 replicates, the males established domi-
nant-subordinate relationships during their initial encounter which remained stable over the remain- ing 4 days. The mean (-I-SEM) chase-attack scores of males that achieved alpha status on day 5 remained high through day 8 (range = 16.2+2.8 on day 5 to 14-8+0-9 on day 8). In contrast, the chase-attack scores of beta and omega males declined sharply during the same period; in fact, none of the omega males engaged in either beha- viour after day 5. Fights were observed only on
days 5 and 6 and occurred only between alpha and beta males. On-backs were also observed only on days 5 and 6 and only by defeated males immedia- tely following an attack. After day 6, beta and omega males always responded to an approach or attack by the alpha male by fleeing with their tails up.
There was no significant correlation between dominance status and strain of the males (Pearson rank correlation = 0.042, P>0.40). Overall, 10 white, seven beige, and eight brown males emerged as alphas. In 19 of the 25 replicates, females dominated all three males and in the remaining six replicates they dominated all but the alpha male.
Mating test: 1. Strain versus dominance status effects
Although there was little correlation between a male's strain and any of the behaviours recorded during the mating tests, there was a significant correlation between genotype and number of young sired (Table III). Further analysis revealed that white males sired significantly more young than beige or brown males (Table IV). While the strains did not differ in the mean number of ejaculations delivered to females, white males obtained a significantly higher mean number of young per ejaculation.
In contrast to the relatively minor strain effects,
Table III. Pearson's rank coefficients for correlations between strain and sexual behaviour and dominance status and sexual behaviour
Measure Strain Dominance status
Behaviour of males Attack 0.001 Ns (75) 0.867** (75) Chase 0.033 Ns (75) 0.870** (75) 'Induced' tail-up 0.044 NS (75) 0"834** (75) Mount 0.046 Ns (75) 0.551"* (75) Short intromission 0.076 Ns (75) 0.681"* (75) Ejaculation 0.114 Ns (75) 0.666** (75) Long intromission 0"105 NS (75) 0"665** (75) Ejaculation order 0.132 Ns (58) 0.729** (58) Number of young 0.296* (58) 0.685** (58)
Behaviour of females Lordosis frequency 0.116 NS (75) 0'639** (75) Longest bout oflordosis 0.110 Ns (75) 0.423** (75) Total lordosis duration 0.125 Ns (75) 0.629** (75)
The number of cases is given in parentheses after each correlation coefficient and its associated probability. * P< 0.05, ** P<0.001, NS: not significant.
978 Animal Behaviour, 34,4
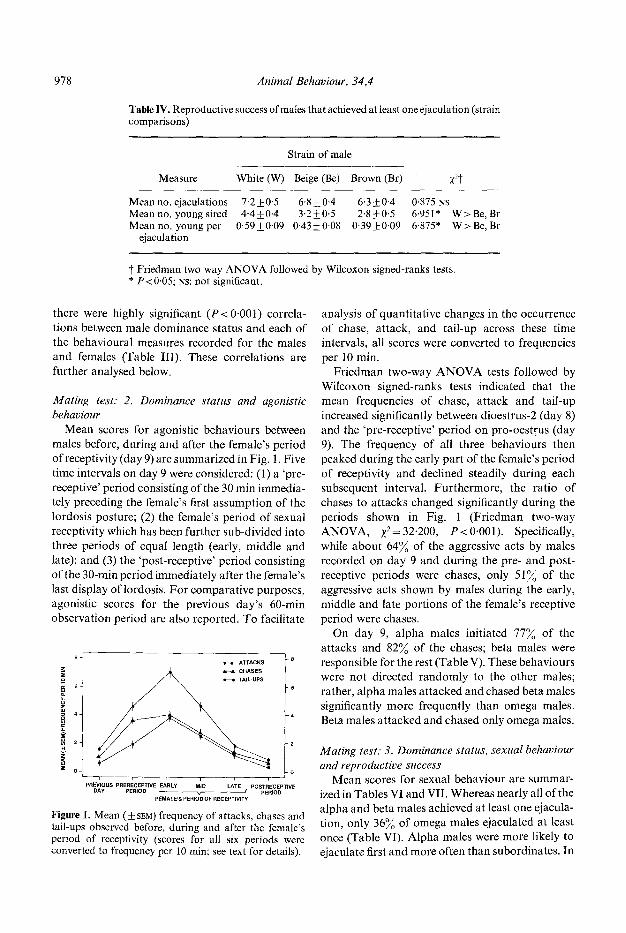
Table IV. Reproductive success of mates that achieved at least one ejaculation (strain comparisons)
Strain ofmale
Measure White (W) Beige (Be) Brown (BO :~2t
Meanno. ejaculations 7.2_+0.5 6.8_+0.4 6.3+0-4 0.875 NS Mean no. young sired 4.4_+0.4 3.2__+0.5 2.8_+_+0.5 6.951" Mean no. young per 0.59-+0.09 0.43_-t-0.08 0.39_+0.09 6.875*
ejaculation
W > Be, Br W>Be, Br
t Friedman two-way ANOVA followed by Wilcoxon signed-ranks tests. * P<0.05; Ns: not significant.
there were highly significant (P<0.001) correla- tions between male dominance status and each of the behavioural measures recorded for the males and females (Table III). These correlations are further analysed below.
Mating test: 2. Dominance status and agonistic behaviour
Mean scores for agonistic behaviours between males before, during and after the female's period of receptivity (day 9) are summarized in Fig. 1. Five time intervals on day 9 were considered: (1) a 'pre- receptive' period consisting of the 30 min immedia- tely preceding the female's first assumption of the lordosis posture; (2) the female's period of sexual receptivity which has been further sub-divided into three periods of equal length (early, middle and late); and (3) the 'post-receptive' period consisting of the 30-rain period immediately after the female's last display of lordosis. For comparative purposes, agonistic scores for the previous day's 60-rain observation period are also reported. To facilitate
8 - a ATTACKS
z ~ CHASES
~ 6- E
~ 2 -2
- o
PREVIOUS PRERECEPTWE EARLY MID LATE POSTRECEPTIVE DAY PERIOD v I PERrOD
FEMALE'S PERIOD OF RECEPTIVITY
Figure 1. Mean (_-+ SEM) frequency of attacks, chases and tail-ups observed before, during and after the female's period of receptivity (scores for all six periods were converted to frequency per 10 min; see text for details).
analysis of quantitative changes in the occurrence of chase, attack, and tail-up across these time intervals, all scores were converted to frequencies per 10 min.
Friedman two-way ANOVA tests followed by Wilcoxon signed-ranks tests indicated that the mean frequencies of chase, attack and tail-up increased significantly between dioestrus-2 (day 8) and the 'pre-receptive' period on pro-oestrus (day 9). The frequency of all three behaviours then peaked during the early part of the female's period of receptivity and declined steadily during each subsequent interval. Furthermore, the ratio of chases to attacks changed significantly during the periods shown in Fig. 1 (Friedman two-way ANOVA, )~2=32.200, P<0-001). Specifically, while about 64~ of the aggressive acts by males recorded on day 9 and during the pre- and post- receptive periods were chases, only 51~ of the aggressive acts shown by males during the early, middle and late portions of the female's receptive period were chases.
On day 9, alpha males initiated 77~ of the attacks and 82~ of the chases; beta males were responsible for the rest (Table V). These behaviours were not directed randomly to the other males; rather, alpha males attacked and chased beta males significantly more frequently than omega males. Beta males attacked and chased only omega males.
Mating test." 3. Dominance status, sexual behaviour and reproductive success
Mean scores for sexual behaviour are summar- ized in Tables VI and VII. Whereas nearly all of the alpha and beta males achieved at least one ejacula- tion, only 36~o of omega males ejaculated at least once (Table VI). Alpha males were more likely to ejaculate first and more often than subordinates. In

Huck et al., Dominance and differential reproduction 979
Table V, Mean 4- SEM frequency and target of agonistic behavieur by alpha and beta males during mating tests
Male initiating behaviour: Alpha Beta
Male receiving behaviour: Beta Omega Zt Alpha Omega Z
Behaviour: Attack 20.5• 11.7• 2.296* 0.0• 9.7-t-1-3 6.795** Chase 23.5+2.8 14.9_+1.9 2.445* 0.0_+0.0 8,3+1.1 6.015"*
t Mann-Whitney two-sample U-tests. * P < 0 . 0 5 ; ** P < 0 , 0 0 1 .
Table VL Dominance status and ejaculatory success
Male dominance status
Measure Alpha Beta Omega
Number (~o) of males which: Ejaculated at least once 23 (92) 24 (96) 9 (36) Ejaculated first 19 (76) 6 (24) 0 (0) Ejaculated most 20 (80) 5 (20) 0 (0) Ejaculated last 6 (24) 14 (56) 5 (20)
contrast, of the nine omega males that ejaculated, five ejaculated last. When all males (inclading those that failed to ejaculate) are considered, alpha males achieved significantly higher mount , intromission
and ejaculation scores than lower ranking males (Table VII). In general, beta males achieved higher sexual behaviour scores than omega males. Alpha males sired significantly more offspring than beta or omega males.
Even when only those males that achieved at least one ejaculation are considered, the reproduc- tive success of alpha males was significantly greater than that of either group of subordinates (Table VIII). The mean ejaculation frequency of alpha males was significantly higher than that of beta or omega males. Further analysis revealed that alpha males also obtained more than twice as many young per ejaculation as beta males and more than three times as many young per ejaculation as omega males.
Table VIL Mean_+ SEM sexual behaviour scores in three-male, one-female competitive tests
Male dominance status
Behaviour Alpha (A) Beta (B) Omega (O) Z2t
M a l e behaviours Mounts 151.4_+12.9 131.9• 10 ,3 71.4_+13.7 11.100" A > B > O Short intromissions 39.4_+ 3.8 29.4• 2.3 9.9• 2.8 22.800**A>B>O Ejaculations 8.8_+ 0.8 5.3_+ 0.5 1.8+ 0.7 27.300**A>B>O Longintromissions 12-4_+ 1.8 2.1_+ 1.4 1-4_+ 1.4 17.100" A > B , O No. youngsired 6.1• 0.6 2-0+_ 0.4 0.5• 0-3 23.175**A>B,O
Female behaviours Lordosis frequency 75.6+ 6.6 52-4+ 5.2 29.4_+ 6.3 18.300"* A> B>O Longest bout oflordosis 660.0+122-6 366.4+ 64-0 183.7+ 55.0 17.200" A > B > O Total lordosis duration 3044-8• 1808.3• 187.0 1048,I • 24-t00"* A > B > O
Scores represent the average of 25 replicates. t Friedman two-way ANOVA tests followed by Wilcoxon signed-ranks tests. * P < 0 . 0 1 ; ** P < 0 " 0 0 1 .

980 Animal Behaviour, 34,4
Table VIII. Reproductive success of males that achieved at least one ejaculation (dominanee status comparisons)
Male dominance status
Measure Alpha (A) Beta (B) Omega (0) Z2t
Mean no. ejaculation 10-0 4- 0.6 Mean no. young sired 7.0_+0.5 Mean no. young/ejaculation 0.76-+0.09
6.0_+0.7 4.4_+1.3 25.763" A>B, O 2-1_+0.5 1.3-+0.6 31.238" A>B,O
0.31-+0.06 0.20-+0.10 20.050*A>B,O
I" Friedman two-way ANOVA followed by Wilcoxon signed-ranks test. * P < 0.001.
Males that ejaculated first engaged in consider- able amounts of copulatory behaviour before another male gained an ejaculation (Table IX). However, on average, the 19 alpha males that ejaculated first, mounted, intromitted and ejacu- lated significantly more frequently before another male ejaculated than did the six beta males that ejaculated first.
Copulatory behaviour occurred predominantly in the female's home area, especially in her nest box. In fact, 74~ of total lordosis duration and 85~ of ejaculations occurred in the female's area, Several alpha males (N= 10) and a few beta males (N = 4) slept in the female's nest box on the evening of dioestrus-2 (day 8) and began mating with the female in her nest box on the following morning (pro-oestrus). Females frequently attacked males with which they had mated while subsequently mating with one of the other males.
D i s c u s s i o n
These results indicate a strong correlation between male dominance status, copulatory beha-
Table IX. Amount of copulatory behaviour achieved by the male that ejaculated first before another male gained an ejaculation
Behaviour
Male ejaculating first
Alpha (N = 19) Beta (N= 6) Z t
Mounts 132.6_+13.5 46-7_+12.6 3.024** Short intromissions 32.8+ 2.9 13'3_+ 1.7 3.405*** Ejaculations 7.0_+ 0.7 2.8-+ 0.6 3.040** Long intromissions 6.2-+ 1.8 0.1_+ 0.2 2.096*
I" Mann-Whitney two-sample U-tests. * P < 0 - 0 5 ; ** P < 0 . 0 1 ; *** P < 0 . 0 0 1 .
viour and differential reproduction. Alpha males achieved significantly higher scores for all measures of sexual behaviour including ejaculation fre- quency (Table VII). However, their differential reproductive success was not simply a function of higher ejaculation frequencies since they obtained more than twice as many young per ejaculation as did either class of subordinates.
Many of the genotype-related differences in sexual behaviour frequencies noted in one-male, one-female tests (Table I) were not observed in the present experiment. However, the results of this experiment did reveal a significant correlation between strain and the number of young sired (Table III). Further analysis indicated that this effect was largely due to males of the white strain obtaining more young per ejaculation than either beige or brown males (Table IV). Since strain was not correlated significantly with either social rank or any behavioural measure recorded during the mating tests, the most likely explanation for this effect is the enhanced differential fertilizing capa- city of the white genotype vis-fi-vis males of the other strains (experiment 2). In any event, this strain effect is rather small when compared with the effects of social status (see Table III).
Why does the dominant male sire significantly more young per ejaculation than beta or omega males? The strong correlation between dominance status and mating order (Table III) may provide an answer. Of the 23 alpha males that ejaculated, 19 ejaculated first and the rest ejaculated second. Furthermore, when ejaculating first, alpha males engaged in significantly more copulatory beha- viour than did beta males that ejaculated first (Table IX). Given our previous demonstration of a first male mating advantage in a recent study of sperm competition in this species (Huck et al. 1985), the pronounced tendency of alpha males to

Huck et al.: Dominance and differential reproduction 981
copulate first can be viewed as a highly adaptive mating tactic. Furthermore, in that earlier study it was also shown that prolonged copulatory activity by the first male could provide protection against subsequent sperm displacement through two mechanisms: (1) a delay in introduction of compet- ing sperm and (2) an increase in the relative concentration of the first male's sperm. In species in which there is a first male mating advantage (e.g. locusts, Parker & Smith 1975; silver perch, Shaw & Allen 1977), males typically engage in pre-copula- tory guarding of the female or engage in other behaviours that increase the likelihood that they will mate first (Parker 1970, 1978). The observa- tions, in the present study, that alpha males (1) often slept in the female's nest box the night before she became receptive, (2) engaged in particularly high levels of inter-male aggression early during the female's receptive period, and (3) continued to copulate with long intromissions even after their supply of ejaculate was exhausted, are consistent with the foregoing discussion of mating tactics as they relate to sperm competition.
The increased frequency of agonistic interactions between males observed at the onset of the female's period of sexual receptivity has also been noted among rhesus monkeys (Wilson & Boelkins 1970), rams (Lincoln & Davidson 1977), boars (Liptrap & Raeside 1978), rats (Thor & Carr 1979), mice (de Catanzaro 1981) and gerbils (Roper & Polioudakis 1977) and may be a general phenomenon (Taylor et al, 1984). Aggression by the alpha male, however, was not directed randomly; rather, most attacks and chases were directed toward beta males (Table V). Although consistent with the idea that beta animals posed a greater threat to the alpha male's mating priority, this finding is more likely to be the result of the omega males' tendency to avoid the other males. Interestingly, the two alpha males that failed to ejaculate spent most of their time attack- ing and chasing the other males. Similar observa- tions have been reported in competitive mating tests with deer mice (Dewsbury 1981) and brown lemmings (Huck & Banks 1982a). The concept of 'aggressive neglect' may appropriately describe the reduced copulatory activity of these dominant males.
The finding that the ratio of chases to attacks decreased significantly during the female's period of receptivity may aIso be interpreted as an adapt- ive behavioural tactic. Most aggressive interactions between males during this time occurred near the
female and were often preceded and/or followed by bouts of sexual behaviour. A reduced tendency to chase defeated competitors would facilitate the alpha male's remaining in the vicinity of the female and, thus, diminish the risk that a third male will gain a mating.
E X P E R I M E N T 4
Scent Marking and Female Selection of a Mating Partner in Tethered-male Tests
In the previous experiment, females engaged in significantly more sexual behaviour with alpha males than with subordinates. Although this result is probably due, in part, to aggressive interactions between the males, female mating preferences may also have played a role. Several previous studies have shown that female rodents are capable of ascertaining the dominance status of males by olfactory cues (e.g. Jones & Nowell 1973; Huck et al. 1981; Huck & Banks 1982b; White et al. 1984). The finding that dominant males achieved a mating priority only when the females and males have interacted before the female became receptive (Lisk & Baron 1983) further suggests that female prefer- ences were an important factor in the differential reproductive success of alpha males in experiment 3.
Female golden hamsters deposit vaginal secre- tions which have an excitatory effect on the male (Lisk el al. 1972; Murphy 1973) and the peak level of vaginal marking occurs during the 24-b period before sexual receptivity (Johnston 1977, 1979). Previous observations of heterosexual pairs in large enclosures indicated that females prefer to mate in or near their burrows and that on the day preceding sexual receptivity the female makes vaginal marks in a manner which appears to result in a 'trail ' between the male's home area and her burrow entrance (Lisk et al. 1983). In the present experi- ment we examined the possibility that vaginal marking might be used by the female to attract a preferred male to her nest box before or at the onset of sexual receptivity. Since secretions of the flank gland have been implicated in individual discimina- tion of females by males (Johnston & Rasmussen 1984), the location of scent marks from this gland were also recorded.
Preliminary observations indicated that brief periods of interaction were sufficient for males to

982 Animal Behaviour, 34,4
establish stable dominance relationships and for both sexes to engage in high levels of scent marking. Therefore, contact between the animals in the present experiment was limited to 30 min per day. In addition to the agonistic behaviour of the males, the frequency and location of scent marks by both sexes were recorded. Finally, to determine whether preferences in marking patterns were correlated with a mating preference, the males were tethered in their home areas on the female's day of recepti- vity. This prevented aggressive interactions between males and forced the female to approach the males in order for mating to occur.
Methods
Tests were conducted in a 244 x 122 x 40-cm Plexiglas enclosure similar in design to those described in experiment 3. Plexiglas partitions separated the parallel male areas which were connected to the larger female area by guillotine gates (see Fig. 2). The walls of the male compart- ments extended 57 cm past the doors and into the female area. In each compartment a short Plexiglas tube (10 cm in length and 6.5 cm in diameter) permitted access to a sunken Plexiglas nest box filled with wood shavings.
As in the previous experiment, three males and a single female were placed into the enclosure on the
day ofpro-oestrus and allowed to habituate for one complete oestrous cycle (4 days). On the following pro-oestrus (day 5 of the experiment), the female was removed from her area at 1200 hours and kept in a plastic box for 30 min. During this time the gates were opened and the males allowed to interact. In addition to the agonistic behaviours listed in experiment 3, the incidence and location of flank marking were also recorded. Determinations of social status were based on relative chase-attack scores. After 30 min, the males and female were returned to their respective compartments and the gates closed. On the following 3 days, all four animals were allowed to interact freely for 30 min beginning at 1200 hours. In addition to the above behaviours the frequency and location of flank and vaginal marks by the female were recorded during these observation periods.
The female was again receptive on day 9. At 0800 hours the males were tethered to the distal end of their compartments with a 40-cm wire leash. After 30 min the gates were opened giving the female access to the three male compartments. In all but one test the door was opened before the female had emerged from her nest box. Interactions between the males and female were observed for 1 h, beginning at the time the female first entered one of the male compartments. In addition to the sexual behaviours listed in experiment 3, the latency,
E
oAA
FEMALE AREA
2.44m
�9 �9 %
; 1 _. .
o
BETA MALE
o
ALPHA MALE
~o ooo
o
l�9 o ,~,~o
O NEST BOX ENTRANCE
�9 VAGINALMARK
o FEMALE FLANK MARK
A ALPHA MALE FLANK MARK
ooj OMEGA MALE
Z~Z~A o~
i 1.22m
Figure 2. Diagram of enclosure used in scent marking and tethered-male tests (experiment 4). Also shown are scent- marking patterns by a female and an alpha male (beta and omega males did not mark) recorded on day 8 of the experiment (24 h prior to the onset of the female's receptive period).

Huck et al.: Dominance and differential reproduction 983
frequency and duration of entries into each male's compartment were recorded.
Results
Establishment of dominance relationships Stable dominant-subordinate relationships were
established by the males in all 12 replicates. Overall, four white, five beige, and three brown males achieved alpha status. This pattern of domi- nance did not differ significantly from that found in the previous experiment (Z2(2)=1.12, P>0-50). Females dominated all three males in 10 of the 12 tests and all but the alpha male in the remaining two tests.
Flank marking by males Male flank marking was dependent both on
social rank and test day (Table X; Fig. 2). Alpha males flank marked significantly more than either beta or omega males on each of the test days. The frequency of flank marking by alpha males was almost three times higher on dioestrus-1 and dioestrus-2 than on the preceding days. In contrast, flank marking by beta males decreased after pro- oestrus while only three omega males marked on any of the test days. The majority of flank marks by alpha males were made near the entrances to the home areas and nest boxes of the other males (see Fig, 2). In contrast, beta and omega males placed all of their flank marks in their own home areas.
Flank and vaginal marking by females Flank and vaginal marks by females were ana-
lysed according to the location of their occurrence.
For statistical purposes, 'male area' was defined as the compartment in which a particular male was housed (his 'home area') plus the area immediately outside his gate between the Plexiglas walls that extended into the female's area for 57 cm (see Fig. 2).
In general, females were more likely to flank and vaginal mark in and around the alpha male's area than in the areas of subordinate males (Tables XI and XII; Fig. 2). This pattern was particularly prominent on dioestrus-2. Both types of scent marking increased over test days. In fact, vaginal marking increased more than three-fold between dioestrus-1 and dioestrus-2.
Female choice of mates in tethered-male tests Although females demonstrated a strong initial
preference for the alpha male in the tethered-male tests these initial preferences were often 'washed out' during the latter part of the 1-h test. For example, although females tended to avoid omega males, the overall time spent in the home areas of alpha and beta males did not differ significantly (Table XIII). However, when scores for time spent in a male's area are plotted in 5-min intervals, a different pattern emerges (Fig. 3). A two-way repeated measures ANOVA indicated a significant interaction between male preference and time interval (F= 1.95, P<0.01). Further analysis with Student-Newman-Keuls multiple-range compari- sons tests (P<0.05) indicated that females spent significantly more time with alpha males during the first 5 rain of the 1-h test than with either beta or omega males. Time spent in alpha and beta male areas did not differ significantly during the second
Table X. Mean -+ SEM frequency of flank marking by males
Male social status
Test day Alpha Beta Omega X2t
Pro-oestrus(P)w 7.3+1.6 2.7_+0.6 0.5_+0-3 10.292"* A>B>O Oestrus (O) 7.8_+3-0 0.6_+0.2 0.6_+0.4 5.950* A>B, O Dioestrus-1 (DI) 20.6_+3.5 1.6_+0-5 0.8_+0.4 18.667"** A>B, O Dioestrus-2(D2) 24.8+2.1 0.6_+0.3 0.4+0.3 18.167"** A>B, O X2t 9-250* 8.500* 0.250Ns
D1, D2>P,O P>O, D2
t Friedman two-way ANOVA followed, where appropriate, by Wilcoxon sigaled-ranks tests.
w Female was absent during pro-oestrus and present thereafter. * P< 0.05; ** P<0.01; ***P< 0.001; Ns: not significant.

984 Animal Behaviour, 34,4
Table XI. Mean ___ SEM frequency of flank marking by females
Location of scent marks
Test day Alpha (A) Beta (B) Omega (O) Female (F) z2t
Oestrus (O) 0.9_+0-6 0.6+0.3 0.5-+0.4 0.7-+0.3 Dioestrus-1 (DI) 7.5_.+3.3 2-3+ 1.2 1-0-+0.7 3-5• Dioestrus-2 (D2) 9.8 _+ 3-0 4.1 _+ 1.8 3.8 _+ 1.6 4.9 _+ 1.0 X2"l" 13"500"* 7"042* 2"013NS 6"912"
D1, D 2 > O D 2 > O D 1 , D 2 > O
0.167NS 7"167" A > B,O 6"792* A > B, O
t Friedman two-way ANOVA followed, where appropriate, by Wilcoxon signed- ranks tests.
* P<0.05; ** P<0.01; Ns: not significant.
Table XII. Mean _+ SEM frequency of vaginal marking by females
Location of scent marks
Test day Alpha (A) Beta (B) Omega (O) Female (F) ;('~
Oestrus (O) 3.8_+2.4 2.2_+0.8 0.6_+0.4 3.9-+ 1.8 0.292ys Dioestrus-I (D1) 4.9+ 1.9 2.0_+0.4 0.4_+0.2 2.7__+0.3 6-167" A > O Dioestrus-2 (D2) 15.4__+3.6 5.9:/:2.2 5.4-+1.7 4.7_+1.9 10.292"* A > B,O, F g2~ 15.167"* 6.667* 6.500* 2.012NS
D2>O, DI D2>O, D1 D2>O, D1
t Friedman two-way ANOVA followed, where appropriate, by Wilcoxon signed-ranks tests. * P<0.05; ** P<0.01; NS: not significant.
Table XlII. Mean + SEM frequency of female behaviours during tethered-male tests
Dominance status of male
Measure Alpha (A) Beta (B) Omega (O) F-testt
Number of entries into male areas 14.4_+ 1-0 13.4+ 1.0 8.8_+2-3 4.167" A, 13>O
Total time in male area (s) 792.9+52.8 689.3_+57.7 546.0_+67.9 5.837"* A , B > O
Lordosis frequency 11.9 _+ 1.0 10.3 __+ 1-1 6.4 + 1.6 7.130** A, B > O Total lordosis
duration (s) 225.8_+30.2 181.4+28.1 90.0_+29.1 7.402"* A, B > O
Means represent average of 12 tests. t One-way ANOVA followed by Student-Newman-Keuls multiple range compari- sons test. * P < 0 . 0 5 ; ** P < 0 . 0 1 .
5-min interval but bo th scores were significantly higher than for omega males. In subsequent 5-rain intervals, female preferences were less p ronounced . Lordosis dura t ion scores showed a similar pat tern .
Of the 12 females tested, eight ( 6 7 ~ ) first entered the a lpha male 's area while the remain ing four
(33~) first entered the beta male 's area. Similarly, nine (75~) of the females first engaged in lordosis in an a lpha male 's area while the remaining three (25~) first engaged in lordosis with a beta male. Females typically assumed a lordosis posture near one of the males and then moved toward the gate

Huek et al.: Dominance and differential reproduction 985
g ,~, lOO-
75 " z
5o -
*2
2 5 -
ALPHA MALE H BETA MALE L E ~
-125
-100
�9 75
m , , , i , J ,
CONSECUTIVE 5 "M IN INTERVALS
"50
�9 25
Figure 3. Mean (+ sere) time spent by females in the home areas of alpha, beta and omega males during 1-h tethered- male tests (in 5-min intervals).
leading to her home area as soon as the male mounted her. When the male did not follow, the female returned to his vicinity and assumed the lordosis posture again. After engaging in this behaviour repeatedly with one of the males, females then entered the home area of one of the other males. During the course of the 1-h test, all 36 tethered males mounted a female at least once. Not one of them achieved ejaculation, however.
Discussion
Females displayed the highest frequencies of vaginal and flank gland marking on the day before sexual receptivity and both types of marking behaviour occurred more frequently in the alpha male's area than in other parts of the enclosure. Furthermore, the pattern of vaginal marks within the female's home area would tend to lead the male investigating these marks to the entrance of her burrow (Fig. 2). This trail-like deposition of vagi- nal scent marks has been observed previously (Lisk et al. 1983); however, mate selection was not involved in that earlier study since only a single male was present. The results reported here suggest that when more than one male is present, the female displays a preference for a particular male (the alpha animal) by depositing vaginal and flank gland marks in and about the entrance to his home area.
Females are highly antagonistic toward adult conspecifics and will attack and chase males during the early part of their oestrous cycle (Takahashi & Lisk 1983). Since vaginal secretions are highly attractive to males (Johnston 1974; Darby et al. 1975) and their attractant quality does not depend
on the presence of the female (Johnston 1974, 1975), they appear well suited to communicate to the male both the female's location and her readi- ness to engage in copulatory behaviour in the near future.
Recently, Johnston & Rasmussen (1984) reported that individual discrimination of females by males is based on secretions of the flank gland and not on vaginal secretions. It is likely, therefore, that females are also able to discriminate between individual males on the basis of the latter's flank gland secretions. In the present experiment, the high rate of flank marking by both the alpha male and the female on the day preceding sexual recepti- vity suggests that these scent marks may aid in individual identification and help bring the pair together for mating.
When males were tethered within their respective home areas on pro-oestrus and the females had to seek them out in order for mating to occur, the overall scores for number of entries, time in male home area, and lordosis duration indicated that there is no difference between alpha and beta males for any of these measures, although omega males obtained significantly lower scores. Carr et al. (1965) suggested that experimental groups showing no reliable preferences at the end of a test may have exhibited preferences at shorter test durations. In the absence of reinforcement (i.e. copulatory sti- muli), the animals may subsequently mask their original preferences by investigating other stimuli. Such an effect has been documented in olfactory preference tests involving rats (Carr et al. 1965) and a similar phenomenon may have occurred in this experiment. Indeed, when the l-h test was divided into consecutive 5-min intervals females displayed a clear and significant preference for the alpha male during the initial 5 min of the test (Fig. 3).
Throughout the tethered-male tests, it appeared to observers that the female was attempting to lead the male toward her burrow. After several unsuc- cessful attempts to have a male follow her, she entered the home area of another male. This observation suggests that although tethered animal tests may be useful in examining initial mate preferences, they may be less suitable for studying the complete mating pattern.
GENERAL D I S C U S S I O N
In the golden hamster, high social status facilitated

986 Animal Behaviour, 34,4
establishment of a mating priority and led to increased reproductive success. Males of the three genotypes employed in this study did not differ in their general fertilization success (experiment 1), displayed only moderate differences in differential fertilizing capacity (experiment 2), and had an equal probability of achieving alpha status (experi- ments 3 and 4). Thus, unlike most previous studies attempting to relate social rank to differential reproduction (cf. Dewsbury 1982), we have demon- strated both the effect of rank on the frequency and timing of copulatory behaviour and the effect of status on reproductive success.
Both inter-male agonistic interactions as well as female choice behaviours were related to the differential copulatory success of the alpha males. The frequency of attacks and chases increased immediately before the female became sexually receptive and peaked during the early part of sexual receptivity. Female preferences were evidenced in scent-marking patterns on the day before recepti- vity and in initial choice of mates during the tethered-male tests (experiment 4).
Alpha males slept in the female's nest box the night before she became sexually receptive and competed vigorously for the opportunity to mate first on the following morning. These behaviours are similar to precopulatory guarding in insects (e.g. Parker 1970) and, in view of our earlier finding of a first male mating advantage in this species (Huck et al. 1985), suggest that sperm utilization patterns and male mating tactics have coevolved.
After exhausting their supply of ejaculate, alpha males continued to copulate with long intromis- sions. Although females subsequently mated with one or both of the subordinate males, the latter typically achieved fewer ejaculations than the alpha males. Elsewhere (Huck & Lisk 1986) we have shown that prolonged copulation (including long intromissions) by the first male effects an imme- diate but temporary reduction of female receptivity as well as a significant shortening of the receptive period. Since the risk of sperm displacement by a second male is inversely proportional to the length of the delay in the introduction of the second male's sperm (Huck et al. 1985), it appears that long intromissions would tend to protect the first male's ejaculatory investment at the time when his sperm are maximally vulnerable to displacement by those of a rival.
Although choices must be made by both sexes, in most species of mammals one would expect females
to show greater selectivity in choosing mates because they invest a greater proportion of their total parental effort in each litter and have more to lose as a result of an inappropriate mating (Trivers 1972). Countless observations and many empirical studies since the time of Darwin have confirmed this expectation (Dawkins 1976; Janetos 1980; Daly & Wilson 1983). Although female choice often takes the form of an exclusive preference for a particular male (e.g. Burley 1981), mating prefer- ences may also occur when females mate with several males. For example, a female may ensure that a preferred male sires her progeny by mating with him at the time in her sexual cycle that conception is likely to occur and restricting mating with non-preferred males to other times (Halliday t983). Indeed, just such an argument has been advanced to explain mating patterns in many social primates (Bernstein 1976).
In species that exhibit multiple-matings, female choice may also be correlated with patterns of sperm competition. For example, Halliday (1983) hypothesized that, in some species where the last male's ejaculate has precedence over the ejaculates of previous males, a female could ensure that her eggs are fertilized by the best available male by continuing to mate with additional males as long as each succeeding partner is of a higher quality than the last. Similarly, in species in which the first male's sperm takes precedence, females would be expected to mate first with a preferred male. Although speculative, the observed tendency for female golden hamsters to scent mark in and around the alpha male's area on the day before sexual receptivity, as well as their initial mating preferences in the tethered-male tests (experiment 4), suggest that female choice is expressed in which male the female mates with first and in the relative amount of mating with each of several males. Clearly, promiscuity does not preclude selectivity.
A C K N O W L E D G M E N T S
This study was supported by National Science Foundation grant BNS-8300892. We thank Doug Mills for statistical advice.
R E F E R E N C E S
Adler, N. T. & Zoloth, S. R. 1970. Copulatory behavior can inhibit pregnancy in female rats. Science, N.Y., 168, 1480-1482.

Huck et al.: Dominance and differential reproduction 987
Barash, D. P. 1982. Soeiobiology and Behavior. 2nd edn. New York: Elsevier.
Beatty, R. A. 1960. Fertility of mixed semen from different rabbits. J. Reprod. Fert., 1, 52-60.
Bernstein, I. S. 1976. Dominance, aggression and repro- duction in primate societies. J. theor. Biol., 60, 493-497.
Bertram, B. C. R. 1976. Kin selection in lions and in evolution. In: Growing Points in Ethology (Ed. by P. P. G. Bateson & R. A. Hinde), pp. 281-301. Cambridge: Cambridge University Press.
Birdsall, D. A. & Nash, D. 1973. Occurrence of successful multiple insemination of females in natural popula- tions of deer mice (Peromyscus manieulatus). Evolution, 27, 106-110.
Bunnell, V. N., Boland, B. D. & Dewsbury, D. A. 1977. Copulatory behaviour of golden hamster (Mesoerice- tus auratus). Behaviour, 61, 180-206.
Burley, N. 1981. Mate choice by multiple criteria in a monogamous species. Am. Nat., 117, 515-528.
Carr, W. J., Loeb, L. S. & Dissinger, M. L. 1965. Responses of rats to sex odors. J. comp. Psyehol., 59, 370-377.
de Catanzaro, D. 1981. Facilitation of intermale aggres- sion in mice through exposure to receptive females. J. eomp. physiol. Psyehol., 95, 638-645.
Chapais, B. 1983. Reproductive activity in relation to male dominance and the likelihood of ovulation in rhesus monkeys. Behav. Ecol. Sociobiol., 12, 215-228.
Daly, M. & Wilson, M. 1983. Sex, Evolution, and Behavior. 2nd edn. Boston: Willard Grant Press.
Darby, E. M., Devore, M. & Chorover, S. L. 1975. A presumptive sex pheromone in the hamster: some behavioral effects. J. eomp. physiol. Psyehol., 88, 496-502.
Dawkins, R. 1976. The Selfish Gene. New York: Oxford University Press.
DeFries, J. C. & McClearn, G. E~ 1970. Social dominance and Darwinian fitness in the laboratory mouse. Am. Nat., 104, 408-411.
DeFries, J. C. & McClearn, G. E. 1972. Behavioral genetics and the fine structure of mouse populations: a study in microevolution. In: Evolutionary Biology, Vol. 5 (Ed. by T. Dobzhansky, M. K. Hecht & W. C. Steere), pp. 179 291. New York: Appleton-Century-Crofts.
Dewsbury, D. A. 1972. Patterns of copulatory behavior in male mammals. Q. Rev. Biol., 47, 1-33.
Dewsbury, D. A. 1975. Diversity and adaptation in rodent copulatory behavior. Science, N.Y., 190, 947- 954.
Dewsbury, D. A. 1979a. Description of sexual behavior in research on hormone-behavior interactions. In: Endo- crine Control of Sexual Behavior (Ed. by C. Beyer), pp. 3-32. New York: Raven Press.
Dewsbury, D. A. 1979b. Copulatory behavior of deer mice (Peromyscus maniculatus): I. Normative data, subspecific differences, and effects of cross-fostering. J. comp. physiol. Psychol., 93, 151 160.
Dewsbury, D. A. 1981. Social dominance, copulatory behavior and differential reproduction in deer mice (Peromyscus rnaniculatus). J. comp. physiol. Psychol., 95, 880 895.
Dewsbury, D. A. 1982. Dominance rank, copulatory behavior, and differential reproduction. Q. Rev. Biol., 57, 135-159.
Dewsbury, D. A. & Baumgardner, D. L 1981. Studies on
sperm competition in two species of muroid rodents. Behav. Ecol. Sociobiol., 9, 121 133.
Dewsbury, D. A. & Hartung, T, G. 1980. Copulatory behaviour and differential reproduction of laboratory rats in a two-male, one-female competitive situation. Anim. Behav., 28, 95-102.
Dieterlen, F. 1959. Das Derhalten des Syrischen Gold- hamsters. Z. Tierpsychol., 16, 47-103.
Dziuk, P. J. 1965. Double mating of rabbits to determine capacitation time. J. Reprod. Fert., 10, 389-395.
Edwards, R. D. 1955. Selective fertilization following the use of sperm mixtures in the mouse. Nature, Lond., 175, 215-216.
Grant, E. C. & Mackintosh, J. H. 1963. A comparison of the social posture of some common laboratory rodents. Behaviour, 21, 246-259.
Guhl, A. M. &Warren, D. C. 1946. Numbers of offspring sired by cockerels related to social dominance in chickens. Poult. Sei., 25, 460~72.
Halliday, T. R. 1983. The study of mate choice. In: Mate Choice (Ed. by P. Bateson), pp. 3-32. Cambridge: Cambridge University Press.
Hanken, J. & Sherman, P. W. 1981. Multiple paternity in Belding's ground squirrel litters. Science, N.Y., 212, 351-352.
Horn, J. M. 1974. Aggression as a component of relative fitness in four inbred strains of mice. Behav. Genet., 4, 373-381.
Huck, U. W. & Banks, E. M. 1982a. Male dominance status, female choice and mating success in the brown lemming, Lemrnus trimucronatus. Anirn Behav, 30, 665-675.
Huck, U. W. & Banks, E. M. 1982b. Differential attraction of females to dominant males: olfactory discrimination and mating preference in the brown lemming (Lemrnus trimucronatus). Behav. Ecol. Socio- biol., 11,217-222.
Huck, U. W., Banks, E. M. & Wang, S.-C. 1981. Olfactory discrimination of social status in the brown lemming. Behav. Neural Biol., 33, 364371.
Huck, U. W. & Lisk, R. D. 1985a. Determinants of mating success in the golden hamster. I. Male capacity. J. comp. Psychol., 99, 98 107.
Huck, U. W. & Lisk, R. D. 1985b. Determinants of mating success in the golden hamster. II. Pregnancy initiation. J. eomp. Psyehol., 99, 231-239.
Huck, U. W. & Lisk, R. D. 1986. Mating induced inhibition of receptivity in the female golden hamster: I. Short-term and long-term effects. Behav. Neural Biol., 45, 107-119.
Huck, U, W., Quinn, R. P. & Lisk, R. D. 1985. Determinants of mating success in the golden hamster: IV. Sperm competition. Behav. Ecol. Soeiobiol., 17, 239~52.
Janetos, A. C. 1980. Strategies of female mate choice: a theoretical analysis. Behav. Ecol. Soeiobiol., 7, 107- 112.
Johnston, R. E. 1974. Sexual attraction function of golden hamster vaginal secretion. Behav. Biol., 12, 111- 117.
Johnston, R. E. 1975. Scent marking by male hamsters. 1. Effects of odors and social encounters. Z. Tierpsyehol., 37, 75-98.
Johnston, R. E. 1977. The causation of two scent-

988 Animal Behaviour, 34,4
marking behaviours in female golden hamsters, Anim. Behav., 25, 317-327.
Johnston, R. E. 1979. Olfactory preferences, scent mark- ing and 'proceptivity' in female hamsters. Horm. Behav., 13, 21 39.
Johnston, R. E. & Rasmussen, K. 1984. Individual recognition of female hamsters by males: role of chemical cues and of the olfactory and vomeronasal systems. Physiol. Behav., 33, 95-104.
Jones, R. D. & Nowell, N. W. 1973. Aversive and aggression-promotion properties of urine from domi- nant and subordinate male mice. Anim. Learn. Behav., 1,207-210.
Lanier, D. L., Estep, D. Q. & Dewsbury, D. A. 1979. Role of prolonged copulatory behavior in facilitating repro- ductive success in a competitive mating situation in laboratory rats. J. comp. physiol. Psyehol., 93, 781-792.
Lawlor, M. 1963. Social dominance in the golden hams- ter. Bull. Br. Psychol. Soe., 16, 25-38.
Lefevre, G. & Jonsson, U. B. 1962. Sperm transfer, storage, displacement and utilization in Drosophila melanogaster. Genetics, 47, 1719-1736.
Levine, L. 1958. Studies on sexual selection in mice. I. Reproductive competition between albino and black- agouti males. Am. Nat., 92, 21-26.
Levine, L. 1967. Sexual selection in mice. IV. Experimen- tal demonstration of selective fertilization. Am. Nat., 10I, 289-294.
Levine, L., Barsel, G. E. & Diakow, C. A. 1966. Mating behaviour of two inbred strains of mice. Anita. Behav., 14, 1-6.
Levine, L. & Lascher, B. 1965. Studies on sexual selection in mice. II. Reproductive competition between black and brown males. Am. Nat., 99, 67-72.
Lincoln, G. A. & Davidson, W. 1977. The relationship between sexual and aggressive behaviour, and pituitary and testicular activity during the seasonal cycle of rams, and the influence of photoperiod. J. Reprod. Fert., 49, 267-278.
Liptrap, R. M. & Raeside, J. E. 1978. A relationship between plasmic concentrations of testosterone and corticosteroids during sexual and aggressive behaviour in the boar. J. Endocrinol., 76, 75-85.
Lisk, R. D. & Baron, G. 1982. Female regulation of mating location and acceptance of new mating partners following mating to sexual satiety: the Coolidge effect demonstrated in the female golden hamster. Behav. Neural Biol., 36, 416-421.
Lisk, R. D. & Baron, G. 1983. Conditions necessary to the establishment of mating dominance by the male hams- ter. Behav. Neural Biol., 39, 105 115.
Lisk, R. D., Ciaceio, L. A. & Catanzaro, C. 1983. Mating behaviour of the golden hamster under seminatural conditions. Anita. Behav., 31, 659-666,
Lisk, R. D., Zeiss, J. & Ciaccio, L. A. 1972. The influence of olfaction on sexual behavior in the male golden hamster (Mesoerieetus auratus). J. exp. Zool., 181, 69- 78.
McGill, T. E. 1962. Sexual behaviour in three inbred strains of mice. Behaviour, 19, 341-350.
Martin, P. A., Reimers, T. J., Lodge, J. R. & Dziuk, P. J. 1974. The effect of ratios and numbers of spermatozoa mixed from two males on properties of offspring. J. Reprod. Fert., 39, 251-258.
Maslow, A. H. 1936. The role of dominance in the social and sexual behaviour of infra-human primates. II. A theory of sexual behaviour ofinfra-human primates. J. genet. Psychol., 48, 310-338.
Merritt, R. B. & Wu, B. J. 1975. On the quantification of promiscuity (or Promyscus maniculatus?). Evolution, 29, 575-578.
Murphy, M. R. 1973. Effects of female hamster vaginal discharge on the behavior of male hamsters. Behav. Biol., 9, 367-375.
Murphy, M. R. 1977. Intraspecific sexual preferences of female hamsters. J. comp. physiol. Psychol., 91, 1337- 1346.
Oglesby, J. M., Lanier, D. L. & Dewsbury, D. A. 1981. The role of prolonged copulatory behavior in male Syrian golden hamsters (Mesocrieetus auratus) in a competitive mating situation. Behav. Ecol. Sociobiol., 8, 47-54.
Orsini, M. W. 1961. The external vaginal phenomena characterizing the stages of the estrous cycle, preg- nancy, pseudopregnancy, lactation, and the anestrous hamster, Mesocrieetus auratus. Proc. Anita. Care Panel, 11, 193-206.
Parker, G. A. 1970. Sperm competition and its evolution- ary consequences in insects. Biol. Rev., 45, 525-567.
Parker, G. A. 1978. Evolution of competitive mate searching. A. Rev. EntomoL, 23, 173-196.
Parker, G. A. & Smith, G. L. 1975. Sperm competition and the evolution of precopulatory passive phase behavior in Loeusta migratoria migratoriodes. J. Ento- mol., 49, 155-171.
Payne, A. P. & Swanson, H. H. 1970. Agonistic behaviour between pairs of hamsters of the same and opposite sex in a neutral observation area. Behaviour, 36, 259- 269.
Prout, T. & Bundegaard, J. 1977. The population genetics of sperm displacement. Genetics, 85, 95-124.
Robitaille, J. A. & Bovet, J. 1976. Field observations on the social behaviours of the Norway rat, Rattus norvegieus (Barkenhout). Biol. Behav., 1, 289-308.
Roper, T. J. & Polioudakis, E. 1977. The behaviour of Mongolian gerbils in a semi-natural environment, with special reference to ventral marking, dominance, and sociability. Behaviour, 61, 207-237.
Shaw, E. & Allen, J. 1977. Reproductive behavior in the silver perch (Cymatogaster aggregate). Mar. Biol., 40, 81-86.
Takahashi, L. K. & Lisk, R. D. 1983. Organization and expression of agonistic and socio-sexual behavior in golden hamsters over the estrous cycle and after ovariectomy. Physiol. Behav., 31, 477~,82.
Taylor, G. T., Hailer, J., Rupich, R. & Weiss, J. 1984. Testicular hormones and intermale aggressive beha- viour in the presence of a female rat. J. Endocrinol., 100, 315-321.
Telle, H. J. 1966. Contribution to the knowledge of behavioral patterns in two species of rats, Rattus norvegieus and Rattus rattus. Z. Angew. Zool., 53, 129- 196.
Thor, D. H. & Carr, W. J. 1979. Sex and aggression: competitive mating strategy in the male rat. Behav. Neural Biol., 26, 261565.
Trivers, R. L. 1972. Parental investment and sexual selection. In: Sexual Selection and the Descent of Man

Huck et aL: Dominance and differential reproduction 989
(Ed. by B. G. Campbell), pp. 136-179. Chicago: Aldine.
White, P. J., Fischer, R. B. & Meuier, G. F. 1984. Female discrimination of male dominance by urine odor cues. Psych. Rep., 55, 487-491.
Whitsett, J. M. 1975. The development of aggressive and marking behavior in intact and castrated male hams- ters. Horm. Behav., 6, 47-57.
Wilson, A. P. & Boelkins, C. 1970. Evidence of seasonal
variation in aggressive behaviour by Macaca mulatta. Anita. Behav., 18, 719-724.
Wilson, E~ O. 1975. Sociobiology: The New Synthesis. Cambridge, Massachusetts: Belknap.
Zuckerman, S. 1932. The Social Life of Monkeys and Apes. New York: Harcourt, Brace.
(Received 26 February 1985; revised 21 May 1985; MS. number:A4497)




















