
Photoperiodic regulation of melatonin membrane receptor (MT1R) expression and steroidogenesis in...
32
Accepted Manuscript Photoperiodic regulation of melatonin membrane receptor (MT1R) expression and steroidogenesis in testis of adult golden hamster, Mesocricetus auratus Arun Mukherjee, Chandana Haldar PII: S1011-1344(14)00273-5 DOI: http://dx.doi.org/10.1016/j.jphotobiol.2014.08.022 Reference: JPB 9825 To appear in: Journal of Photochemistry and Photobiology B: Bi- ology Received Date: 1 July 2014 Revised Date: 27 August 2014 Accepted Date: 28 August 2014 Please cite this article as: A. Mukherjee, C. Haldar, Photoperiodic regulation of melatonin membrane receptor (MT1R) expression and steroidogenesis in testis of adult golden hamster, Mesocricetus auratus, Journal of Photochemistry and Photobiology B: Biology (2014), doi: http://dx.doi.org/10.1016/j.jphotobiol.2014.08.022 This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Transcript of Photoperiodic regulation of melatonin membrane receptor (MT1R) expression and steroidogenesis in...

Accepted Manuscript
Photoperiodic regulation of melatonin membrane receptor (MT1R) expressionand steroidogenesis in testis of adult golden hamster, Mesocricetus auratus
Arun Mukherjee, Chandana Haldar
PII: S1011-1344(14)00273-5DOI: http://dx.doi.org/10.1016/j.jphotobiol.2014.08.022Reference: JPB 9825
To appear in: Journal of Photochemistry and Photobiology B: Bi-ology
Received Date: 1 July 2014Revised Date: 27 August 2014Accepted Date: 28 August 2014
Please cite this article as: A. Mukherjee, C. Haldar, Photoperiodic regulation of melatonin membrane receptor(MT1R) expression and steroidogenesis in testis of adult golden hamster, Mesocricetus auratus, Journal ofPhotochemistry and Photobiology B: Biology (2014), doi: http://dx.doi.org/10.1016/j.jphotobiol.2014.08.022
This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customerswe are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, andreview of the resulting proof before it is published in its final form. Please note that during the production processerrors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.

1
Photoperiodic regulation of melatonin membrane receptor (MT1R) expression and
steroidogenesis in testis of adult golden hamster, Mesocricetus auratus.
Arun Mukherjee and Chandana Haldar*
Pineal Research Lab., Department of Zoology, Banaras Hindu University, Varanasi-221005,
India.
E-mail ID of the authors:
Arun Mukherjee: [email protected],
Chandana Haldar: [email protected],
*Address for correspondence:
Prof. Chandana Haldar,
Pineal Research Lab.
Department of Zoology,
Banaras Hindu University,
Varanasi-221005, India
Ph: 91-542-6702535 Ext. 209; M. 91-9415222261
Fax: 91-542-2368174
E-mail: [email protected]

2
Abstract
Photoperiodic modulation of melatonin membrane receptor (MT1R) expression in testis has never
been reported for any seasonal breeder. Thus, the aim of the present study was to investigate the
expression dynamics of MT1R in testis and its interaction with testicular steroidogenesis in a long-
day breeder, Mesocricetus auratus. Hamsters were exposed to different photoperiodic conditions
i.e. critical- (CP; 12.5L:11.5D); short-day- (SD; 8L:16D) and long-day- (LD; 16L:8D) for 10 weeks
wherein testicular steroidogenesis, local melatonin synthesis and the expression of MT1R were
analyzed. SD induced melatonin suppressed testicular steroidogenesis as evident from regressed
testicular histoarchitecture, decreased expression of AR, StAR, LH-R, P450SCC and enzyme
activities of 3β- and 17β-HSD. Differential photoperiodic regulation of MT1R expression in testis
suggests its involvement in photoperiodic signal transduction for seasonal adjustment of
reproduction. Increased S-NAT (Serotonin N-acetyl transferase) activity and local testicular
melatonin under SD condition suggest an inhibitory effect of the local melatonergic system on
testicular steroidogenesis. Completely opposite responses were recorded for all the parameters
analyzed when hamsters were exposed to CP or LD conditions. In conclusion, we may suggest that
photoperiod via regulating circulatory and local melatonin level as well as MT1R expression in
testes fine tunes the steroidogenesis and thereby, the reproductive status of male golden hamster.

3
1. Introduction
Melatonin, the principal indoleamine secreted from the pineal gland of vertebrates,
synchronizes the seasonal and circadian physiological functions, including reproduction [1,
2]. It has been established that melatonin regulates reproduction in phylogenetically distant
seasonal breeders in a pro- or anti-gonadotropic manner [3, 4] depending on the species.
Being exposed to different photoperiodic and climatic conditions throughout the year,
seasonal breeders breed at a particular time of the year to avoid infant mortality arising due to
adverse climatic conditions. Herein, melatonin acts as a principal transducer of the
photoperiodic stimuli and provides the animal with the ‘time-of-year signal’, restricting the
delivery of the offspring to the most favourable period of the year [5], which helps in
minimizing the maternal cost to benefit ratio and maximizes the chances of offspring survival
[6]. Melatonin modulates seasonality in reproductive status by influencing the hypothalamo-
hypophysial-gonadal (H-P-G) axis [7]. Photoperiodically modulated peripheral melatonin via
MT1R influences the GnRH neurons in the hypothalamus and pars tuberalis of the pituitary,
thereby inhibiting the secretion of the gonadotropin releasing hormone (GnRH) from the
hypothalamus [8] and the gonadotropins (LH and FSH) from the gonadotrophs of the anterior
pituitary [9]. Till date, two high affinity G-protein coupled melatonin membrane receptors
(MT1R and MT2R) have been cloned and characterized and are reported to be expressed
ubiquitously in mammalian tissues [10, 11]. Despite the presence of both the receptor
subtypes in the testis [12], the major points of cross-talk of the melatonergic system with the
reproductive system are mainly mediated via the MT1R subtype [13, 14]. The presence of
two nonsense mutations in the coding frame of the MT2R subtype renders it inactive in the
transduction of photoperiodic stimuli in hamster [15,16] to an extent that transduction of
photoperiodic stimuli occurs principally via the MT1R [13]. The sexual cycle of the golden
hamster, M. auratus, depends on the environmental photoperiod, wherein, the durations of

4
the dark and the light periods affect its sexual activity [17]. Being a long-day breeder, it
remains sexually active throughout the year when exposed to ≥ 12.5 hours of light (critical
photoperiod) under laboratory conditions [18]. The presence of MT1R has been reported in
rat Leydig cells [19] and it has been suggested that melatonin may have a local inhibitory
action on the testicular steroidogenic pathway [4]. Previous studies established that, exposure
to inhibitory short photoperiods [4] or exogenous administration of melatonin [20] inhibits
testicular testosterone production. With the aforementioned reports, it is imperative that
photoperiodic stimulus is an important modulator of the testicular steroidogenic pathway.
Since, melatonin is the principal transducer of such stimuli it can be presumed that the local
melatonergic system may have substantial cross-talk with the testicular steroidogenic
pathway, via the MT1R. Based on the above hypothesis, we attempted to explicate the
photoperiodic regulation of the expression of MT1R and its interaction with testicular
steroidogenic pathways in adult male golden hamsters, M auratus.
2. Materials and methods
All the experiments were conducted in accordance with Institutional practice and within the
framework of experimental animals (Scientific Procedure) Act 2007, of the Committee for
the Purpose of Supervision and Control on Experiments on Animals (CPSCEA), Government
of India, on animal welfare.
2.1.Animal procurement and maintenance
Golden hamsters were procured from Central Drug Research Institute (CDRI), Lucknow,
India and colonies were developed and maintained in the departmental animal house facility.
Hamsters were kept under constant temperature (25±2°C) and light/dark cycle (Critical
photoperiod; 12.5 h light, 11.5 h dark; i.e. lights on at 07:00 a.m. and lights off at 07:30 p.m.).

5
Animals were maintained in polypropylene cages of equal sizes and provided with
commercial rodent pellet and tap - water ad libitum.
2.2.Photoperiodic treatments
Adult male golden hamsters (average weight 125 g, 90-100 days old) were randomly selected
and divided into three experimental groups (N=5/group) and exposed to different
photoperiodic regimes for 10 weeks:
Group I: Critical photoperiod (CP; 12.5 Light: 11.5 Dark),
Group II: Short-day photoperiod (SD; 8Light: 16Dark) and
Group III: Long day photoperiod (LD; 16Light: 8Dark)
Hamsters were exposed to photostimulatory long day condition and photoinhibitory short day
condition for 10 weeks to achieve maximum testicular growth and regression respectively.
The additional group of hamsters exposed to the critical photoperiod, served as control.
2.3.Tissue collection and processing
At the end of the treatment, hamsters were weighed and sacrificed under deep ether
anaesthesia. Right and left testes were immediately removed, blotted dry and weighed. Testes
were then either quickly fixed in 10% neutral formalin for histological and
immunohistochemical analyses or kept at -80°C for biochemical estimations and western blot
analyses.
2.4.Histology
For histological studies, testes were cut into two halves through its short axis and immersed
in 10% neutral formalin for 24 hours and then cut into small slices of 1-2 mm thickness and

6
kept in fresh 10% neutral formalin for 24 hours. After fixation, testes slices were washed
under running tap-water to remove extra fixative and sequentially dehydrated. After clearing
in xylene, tissues were embedded in paraffin and cut into 5µm sections. The sections were
stretched on clean glass slides pre-coated with 1% gelatine. Deparaffinised sections were
stained using hematoxylin and eosin stain. Testicular morphology was observed under
research microscope (Nikon, E 200, Japan) in randomly selected sections. Seminiferous
tubular diameter and area were measured from 100 round seminiferous tubules from the
observed cross-sections.
2.5.Immunohistochemical localization
For immunohistochemical localization, stretched sections on glass slides were sequentially
rehydrated. Endogenous peroxidase activity was blocked by 0.1 % H2O2 in methanol for 30
min. at room temperature (RT). Sections were washed thrice with phosphate buffered saline
(PBS; 0.1 M NaH2PO4, Na2HPO4, NaCl; pH 7.4) and incubated with horse blocking serum
(1:100 in PBS; PK-6200, Vector laboratories, Burlinghame, CA) for 2 hours. Then sections
were incubated with primary antibodies against AR (AR, N-20, sc-816, Lot # E0412, Santa
Cruz Biotech, USA, dilution 1:50) and MT1 (MT1R, R-18, sc- 13186, Lot # L1812, Santa
Cruz Biotech, USA, dilution 1:100) overnight at 40C. Sections were washed thrice with PBS
and incubated with biotinylated secondary antibody (Vectastain ABC Universal kit; PK-
6200, Vector laboratories, Burlinghame, CA, dilution 1:50). Sections were then washed with
PBS and a pre-formed ABC reagent was conjugated to the free biotin of the secondary
antibody. The antigens were visualized using the 0.03% peroxidase substrate 3,3-
diaminobenzidine (DAB, Sigma chemicals, St. Louis, USA) in 0.01M Tris-HCl (pH 7.6) and
0.1% H2O2 and counterstained with Ehrlich’s hematoxylin. Counterstaining with hematoxylin

7
was avoided in case of nuclear antigen (i.e. AR). Sections were dehydrated, mounted with
DPX and observed under research microscope (Nikon, E 200, Japan).
2.6.Western blot analyses
For western blot analyses, testes were homogenized and lysed in RIPA buffer containing
aprotinin, sodium orthovanadate and phenyl methylsulphonyl fluoride (PMSF) protease
inhibitor cocktail and was centrifuged at 12,000×g for 30 min, at 4ºC. The protein content of
the lysates was estimated [21]. The aliquots containing 100 µg protein was resolved on 12%
SDS-polyacrylamide gel (PAGE) for MT1R, P450SCC and StAR, , proteins and on 10% SDS-
PAGE for AR and LH-R, followed by electrotransfer (Biometra, Germany) on nitrocellulose
membrane (Boiscience, Keene, NH, USA) for 1 hour. Nitrocellulose membranes were
blocked for 60 min in Tris-buffered saline (TBS; Tris-HCl 50 mM, pH 7.5, NaCl 150 mM)
containing 5% fat free dry milk and were incubated with primary antibodies against AR (AR,
N-20, sc-816, Lot # E0412, Santa Cruz Biotech, USA, dilution 1:250); StAR (StAR, Rabbit
anti-mouse, dilution 1: 250); P450SCC (CYP11A1, H-165, sc-292456, Lot # J2511, Santa
Cruz Biotech, USA, dilution 1:250) and LH-R (LH-R, Rabbit anti-human, dilution 1:100),
MT1 (MT1R, R-18, sc- 13186, Lot # L1812, Santa Cruz Biotech, USA, dilution 1:200).
Antibodies against StAR and LH-receptor were kind gifts from D.M. Stocco (Texas Tech
University, Health Sciences Center, Lubbock, TX, USA) and Craig S. Atwood (William S.
Middleton Memorial Veterans Hospital, Madison, WI, USA) respectively. After overnight
incubation at 4°C, the membranes were washed thrice with TBS. The immunodetection was
carried out using horseradish peroxidase (HRP) conjugated secondary antibody (Goat anti-
rabbit, 1:1000; Donkey anti-goat, 1: 1000). Finally, the blots were washed thrice with TBS
and developed with Super Signal West Pico Chemiluminescent substrate (34080, Thermo
Scientific, USA). Further, the nitrocellulose membranes were then stripped with stripping

8
buffer (10% sodium azide) and were immunostained with β-actin antibodies at 1:5,000
dilution (A-2228; Sigma–Aldrich, USA) as internal loading control. Immunodetection of β-
actin was performed with HRP conjugated anti-mouse IgG (1:10,000). Band intensities were
quantified by densitometry using the Image-J analysis software. Values were expressed as the
ratio of the density of the specific signal to that of β-actin.
2.7.Testicular enzyme assay
To observe the effect of different photoperiodic conditions on the steroidogenic pathway,
testicular Δ5 3β-Hydroxysteroid Dehydrogenase (3β-HSD) and 17β-Hydroxysteroid
Dehydrogenase (17β-HSD) activity were measured according to the method of Talalay
(1962) and Jarabek et al. (1962) [22, 23], respectively with minor modifications. Briefly,
testes were homogenized in 20% (v/v) spectroscopic grade glycerol (Sisco Research
Laboratories Pvt. Ltd, India) containing 5 mM potassium phosphate and 1 mM EDTA at a
tissue concentration of 100 mg/ml to prepare 10% (w/v) homogenate and was centrifuged at
12,000 g for 30 min, at 4ºC. The supernatant (approx. 950 µl) was used for enzyme assay.
3β-HSD was measured by mixing 250 µl of the supernatant with 250 µl of 100 µM sodium
pyrophosphate buffer, (pH 8.9), 10µl ethanol containing 30 µg DHEA (Sigma, St. Louis,
USA) and 240 µl of BSA (Sigma, St. Louis, USA) at a concentration of 0.25 mg/ml. Enzyme
activity was measured after addition of 50 µl of 0.5 µM NAD to the mixture in a
spectrophotometer at 340 nm against a blank (without NAD). One unit of enzyme activity is
the amount causing a change in absorbance of 0.001/min at 340 nm. 17β-HSD was measured
by mixing 250 µl of the supernatant with 250 µl of 440 µM sodium pyrophosphate buffer
(pH 10.2), 10 µl ethanol containing 0.3 µM testosterone (Sigma, St. Louis, USA) and 240 µl
of BSA at a concentration of 0.25 mg/ml. Enzyme activity was measured after addition of 50
µl of 0.5 µM NAD to the mixture in a spectrophotometer at 340 nm against a blank (without

9
NAD). One unit of enzyme activity is the amount causing a change in absorbance of
0.001/min at 340 nm.
2.8.Plasma testosterone assay
The level of testosterone in serum was measured by using a highly sensitive and specific
commercial ELISA kit (DiaMetra, Italy-DKO002) as per the manufacturer’s instructions. All
samples were quantified in a single assay. The sensitivity of kit was 0.07ng/ml and the intra
and inter-assay variations were < 5.8% and < 10.5% respectively-as per the manufacturer.
2.9.Plasma melatonin assay
The assay was performed according to the manufacturer's protocol provided with the kit
(Uscn Life Science Inc. USA). The sensitivity of the kit was 4.68 pg/ml––as per the
manufacturer. All samples were quantified in a single assay. The intra and inter- assay
variation was <10% and <12% respectively and the recovery percentage was between 90 and
115.
2.10. Testicular S-NAT activity assay
Testicular S-NAT activity was measured following the radioisotopic method of Chae et al.,
(1999) [24]. Briefly, testes were dissected out under red light and quickly frozen in liquid
nitrogen. Testis was homogenized in 0.2M phosphate buffer pH 7.4 at a tissue concentration
of 100 mg/ml to prepare 10% (w/v) homogenate and was centrifuged at 12,000 g for 30 min,
at 4ºC. 10µl of homogenates were mixed in a reaction mixture containing 6 mM tryptamine
hydrochloride, 0.5mM Acetyl-coA and 3H Acetyl-coA (0.2µCi/assay). The reaction mixture
was incubated at 370C for 20 minutes and the reaction was stopped by addition of 180µl
0.2M phosphate buffer. The mixtures were then transferred to scintillation vials containing
3ml of scintillation fluid (PPO and POPOP in toluene). After 24 hours of incubation at room

10
temperature in dark, the radioactivity was recorded in a beta scintillation counter (Beckman
Coulter, USA). The activity of the enzyme was expressed as nmoles of N-acetyl tryptamine
formed /min/gm tissue weight.
2.11. Testicular melatonin concentration
Testicular melatonin concentration was determined with an ELISA kit (IBL, Hamburg,
Germany). Testes were homogenized in PBS with 0.1% ethanol in a sonicator at a tissue
concentration of 100 mg/ml to prepare 10% (w/v) homogenate and centrifuged at 3000 g for
10 min. Standards, controls and testicular homogenates were extracted using C18 extraction
columns (IBL, Hamburg, Germany) according to manufacturer’s protocol. The extract was
evaporated to dryness in an evaporator centrifuge (Speed-Vac®, SC110, Savant Instruments,
INC). The dried extracts were used for ELISA. Signals were analyzed at 405 nm using a
microplate reader (Multiskan Ex, Thermo Electron Corporation). Melatonin concentration in
testis was expressed as femtomole (fmol)/gm tissue weight.
2.12. Statistical analyses
Statistical analysis of the data was performed using SPSS 17.0 (SPSS Corp., USA) with one-
way ANOVA followed by Tukey’s multiple range tests for multiple comparisons. The
differences were considered statistically significant when p < 0.05.
3. Results:
3.1.Effect of different photoperiodic regimes on body weight and relative testes
weight
Variations were observed in body weight and relative testes weight in the hamsters exposed
to different photoperiodic conditions. A significant (p<0.01) increase in body weight (Table
1) with a significant (p<0.01) decrease in relative testes weight (Table 1) was recorded in

11
hamsters exposed to SD as compared to CP, whereas a significant (p<0.01) decrease in body
weight with a concomitant increase in paired testes weight was observed under LD condition
when compared with SD exposed hamsters.
3.2.Effect of different photoperiod on morphometry and testicular histoarchitecture
Morphometric analyses of seminiferous tubules revealed a significant (p<0.01) decrease in
seminiferous tubular diameter (Table 2) and seminiferous tubular area (Table 2) under SD
condition when compared with CP, whereas significant (p<0.01) increase in diameter and
area of seminiferous tubules was noted in LD group as compared to CP and SD exposed
hamsters. Marked differences in testicular histoarchitecture were noted in hamsters exposed
to SD and LD photoperiodic conditions. However, no such alterations were observed when
comparison was made between CP and LD group (Data not shown). Under LD condition,
normal histoarchitecture of the testis with sequential arrangement of germ cells at different
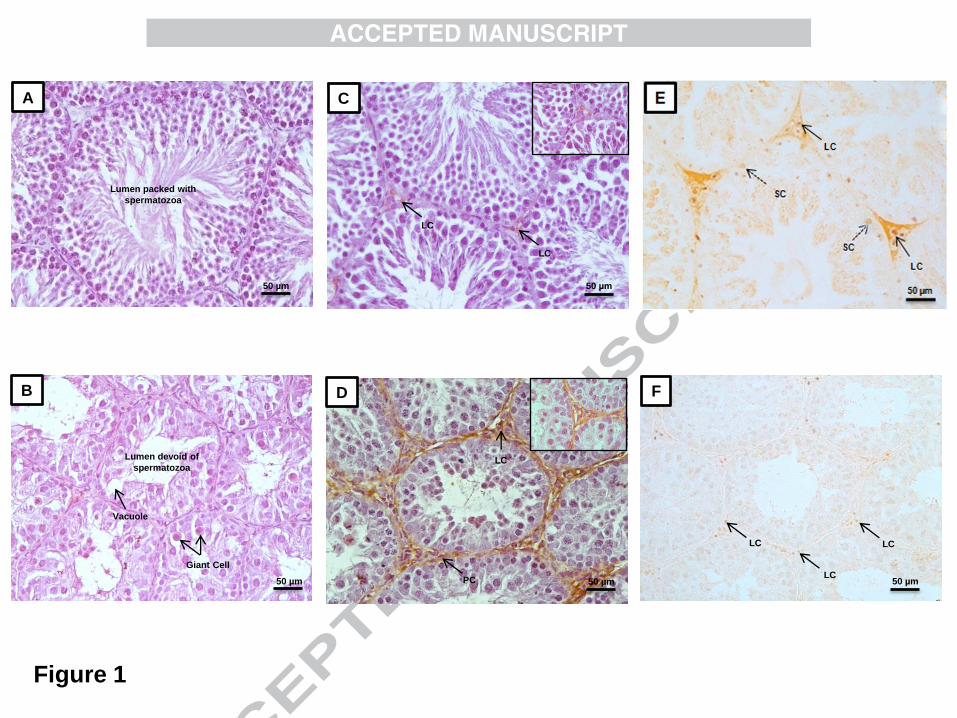
stages of development and lumen filled with sperm was observed (Figure 1A). Histologically,
the testis of SD exposed hamsters showed signs of atrophy with vacuolization and presence
of large number of giant cells in the seminiferous tubules with the lumen being completely
devoid of sperm (Figure 1B).
3.3. Immunohistochemical localization of MT1R and AR in testis
The product of immune reactions for MT1R was observed in the interstitial compartment, in
general and specifically on the Leydig cells and peritubular cells. Weak immunoreactivity for
MT1R was observed in testis under LD condition (Figure 1C) as compared to strong
immunoreactivity under SD condition (Figure 1D). Strong nuclear immunoreactivity for AR
was observed in the Leydig cells and Sertoli cells of hamsters exposed to LD (Figure 1E) as
compared to weak nuclear immunoreactivity for AR in the Leydig cells under SD condition
(Figure 1F).

12
3.4.Effect of different photoperiod on testicular steroidogenesis
Photoperiodic modulation of testicular steroidogenesis was assessed through western blot
analyses of key regulators i.e. LH-R (Lutinizing hormone receptor), StAR (Steroidogenic
acute regulatory protein), P450SCC (P450 Side-chain cleavage) enzyme, AR and estimations of
activity for the key enzymes i.e. 3β-HSD and 17β- HSD of steroidogenesis. Western blot
analyses revealed a significant (p<0.01) decrease in the expression of AR (Figure 2A), StAR
(Figure 2A), P450SCC (Figure 2B) and LH-R (Figure 2B) in hamsters exposed to SD
condition as compared to CP and LD hamsters. However, a significant (p<0.01) increase in
the expression of all the key regulators of steroidogenesis was noted in hamsters exposed to
LD condition when compared to CP and SD groups. Estimations of enzyme activities for 3β-
HSD and 17β- HSD followed the same pattern with a significantly (p<0.01) decreased
enzyme activity under SD condition and vice-versa (Table 3). Significant (p<0.01) decrease
in plasma level of testosterone was noted in SD exposed hamsters as compared to CP and LD
groups (Figure 2A). Taken together, an inhibitory action of SD exposure and a stimulatory
action of LD exposure on testicular steroidogenesis might be suggested.
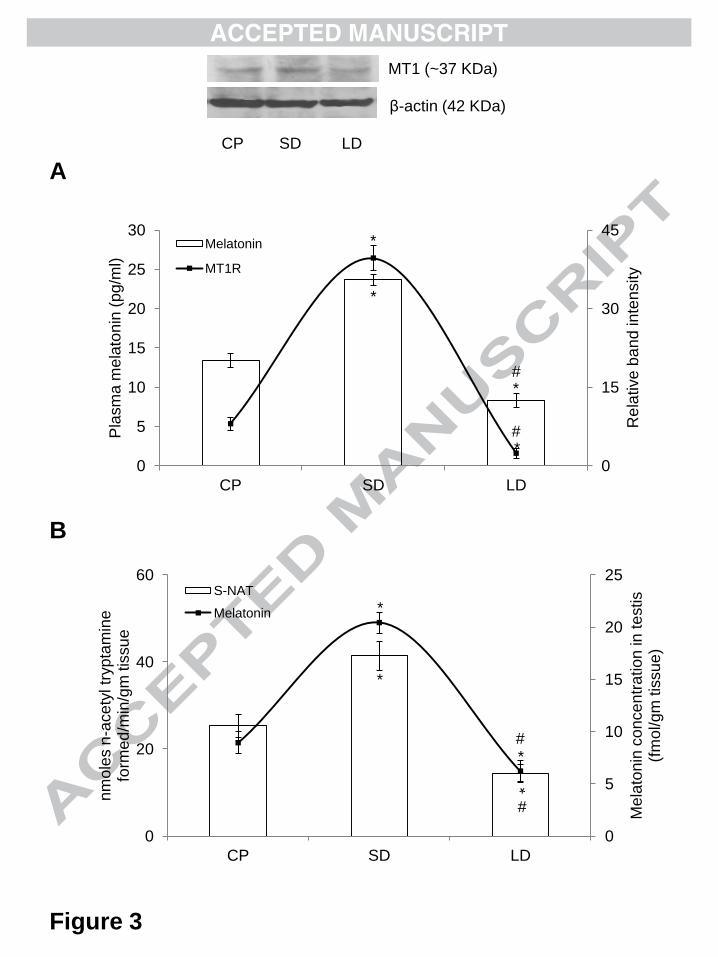
3.5.Effect of different photoperiod on plasma melatonin level and testicular MT1R
expression
Marked alterations in plasma levels of melatonin and testicular MT1R expression was
observed in hamsters exposed to different photoperiodic conditions. A significant (p<0.01)
elevation in the plasma level of melatonin was observed under SD condition as compared to
CP and LD exposed hamsters (Figure 3A). Western blot analysis for MT1R expression in
testis showed a significant (p<0.01) increase under SD condition as compared to CP and LD
hamsters with a significantly (p<0.01) decreased expression under LD exposure when
compared with SD and CP exposed hamsters (Figure 3A).

13
3.6.Effect of different photoperiod on testicular melatonin synthesis
Photoperiodic variation in testicular melatonin production was assessed through measurement
of enzyme activity for the S-NAT, the rate-limiting enzyme for melatonin biosynthesis, and
testicular melatonin level. A significant (p<0.01) increase in S-NAT enzyme activity (Figure
3B) and testicular melatonin concentration (Figure 3B) was recorded in hamsters exposed to
SD condition as compared to CP and LD groups. LD exposure significantly (p<0.01)
abolished the S-NAT enzyme activity and testicular melatonin concentration when compared
with CP and SD hamsters.
4. Discussion:
The essential role played by melatonin in regulation of seasonal reproductive rhythms has
been known for quite some time [25, 26]. The golden hamster, M. auratus is a long-day
seasonal breeder with its neuroendocrine axis being highly sensitive to the changing
photoperiod [25]. It is an excellent animal model for the study of photoperiodic modulation
of reproduction [27] and immunity [28]. Testis is the prime source of circulatory level of
testosterone and the unique site for spermatogenesis in mammals. Hence, detailed
understanding of the photoperiodic modulation of testicular steroidogenesis is of great
significance. The question of whether photoperiodic stimuli influence testicular steroidogenic
pathways via modulation of the melatonin-induced MT1R has never been explored.
In the present study, a significant increase in body mass was noted in hamsters exposed to SD
condition along with a concomitant decrease in paired testes weight. The increased body
weight in the hamsters is due to the increased carcass lipid content, adapted as a means of
successful hibernation strategy [29]. Further, the decreased testes weight in hamsters exposed
to SD photoperiod may be attributed to the inhibitory effect of SD on testicular growth which
thereby minimizes reproductive efforts under photoinhibitory SD condition [1]. The decrease

14
in testicular weight and in the diameter and area of seminiferous tubules was accompanied by
the presence of giant cells and intra-epithelial vacuolization in the seminiferous tubules with
lumen being completely devoid of spermatozoa as evidenced from the histological analyses
of testis under SD photoperiod. Our findings receive substantial support from previous
studies [30] suggesting that the testicular regression is due to androgen deficiency under SD
condition. Further, the presence of intraepithelial vacuoles and giant cells in the tubules can
be regarded as a hallmark of decreased plasma levels of testosterone as observed during the
normal process of ageing [31] or during exposure to immunosupessive drugs [32]. The
significant decrease in the level of plasma testosterone under SD condition complies with the
observed histological alterations in the seminiferous tubules. Therefore, it can be suggested
that a trade-off relationship exists between testosterone and melatonin to provide the best
possible way for the seasonal adjustment of reproduction [33].
The present study provides evidence for the photoperiodic variation in MT1R expression in
testis, being higher during the SD condition and lower during LD condition in golden
hamster, M. auratus. Till now, reports suggesting the photoperiodic regulation of MT1R
expression in testis are unavailable for any seasonal mammal, in general, and specially for
golden hamster. Melatonin is classically known to inhibit testicular androgen biosynthesis by
acting at multiple levels of the H-P-G axis through MT1R [7]. In the present study, an
elevated level of peripheral melatonin was observed under SD condition, implying that the
increased melatonin titre inhibits testicular steroidogenesis by up-regulating MT1R in the
Leydig cells.
Further, immunohistochemical (IHC) studies revealed the presence of MT1R in the
interstitial Leydig cells with strong immunoreactivity under SD condition and weak
immunoreactivity under LD condition suggesting a direct action of photoperiodically

15
modulated peripheral melatonin on Leydig cell steroidogenesis. To confirm our IHC data, we
quantitatively assessed the expression of MT1R through western blot analyses. Our
immunoblot analyses revealed highest expression of MT1R under SD condition and lowest
under the LD condition. The high circulatory level of melatonin paralleled the increased
expression of MT1R under SD condition whereas decreased expression of MT1R was
accompanied by low circulatory level of melatonin. Our results are in agreement with earlier
reports demonstrating increased expression of MT1R coincident with high circulatory level
of melatonin [34]. Analyses of AR expression showed an opposite pattern as compared to
MT1R expression i.e. minimal expression of AR under SD exposure and vice versa. Thus,
decreased AR expression in testis parallels the decreased plasma level of testosterone. Our
results mimic earlier reports in the bank vole [35] implying that the stabilization and function
of AR protein is dependent on prolonged receptor occupancy with androgen [36]. Thus,
under LD condition the decreased level of melatonin and MT1R expression coincide with the
increased level of testosterone and AR expression and vice versa suggesting an antagonistic
relationship between melatonin and testosterone and their receptors.
To further elucidate the mechanism of decreased androgen biosynthesis under SD exposure,
we delved into the testicular steroid biosynthetic pathway. Although many enzymes are
involved, it is believed that the rate-limiting step in the steroidogenesis is the movement of
cholesterol across the mitochondrial membrane so that it can be acted upon by the
cytochrome P450 side chain cleavage enzyme (P450SCC) [37]. In mammals, the movement has
been shown to be mediated by the steroidogenic acute regulatory protein (StAR) [38]. StAR
transcripts are rapidly synthesized in response to luteinizing hormone and cAMP [39]. A
significant decrease in StAR, P450SCC and LH-R expression with a simultaneous decrease in
3β- and 17β-HSD enzyme activity was observed under SD condition as compared to LD and
CP exposed hamsters, proposing an inhibitory action of SD condition on testicular

16
steroidogenic machinery. The increased levels of StAR P450SCC and LH-R expression and
enzyme activities of 3β- and 17β-HSD under LD condition concord with the elevated plasma
level of testosterone and are supported by earlier studies in golden hamster [4]. LH is known
to up-regulate the expression of StAR and P450SCC in a cAMP mediated pathway and thereby
stimulate the synthesis of testosterone [37, 40]. On the other hand, binding of melatonin to
MT1R decreases the cAMP level [41] and thus may down-regulate LH-R signaling in Leydig
cells and decreases the expression of steroidogenic genes. The decreased expression LH-R is
concurrent to the decreased expression of StAR, P450SCC and plasma level of testosterone
and is in accordance to the previous reports [4]. Further, evidences suggest local synthesis of
melatonin in rat testis [42]. To check, whether melatonin is being produced in hamster testis,
we measured the enzyme activity for S-NAT-rate limiting enzyme in melatonin synthesis.
Our observations confirmed testicular synthesis of melatonin and also in the variation of
testicular melatonin concentration in a photoperiod dependent manner. Therefore, an elevated
S-NAT enzyme activity and testicular melatonin level under SD condition suggested a local
inhibitory action of melatonin on testicular steroidogenesis.
Conclusion:
The present study demonstrates that the changing photoperiod causes alterations not only in
the peripheral levels of melatonin and androgen but also in the expression pattern of MT1R
and AR in the testis of a long-day breeder, M. auratus. Photoperiodic modulation of plasma
level of melatonin and expression of MT1R demonstrate a reciprocal relationship with the
plasma level of testosterone and expression of AR implying a trade-off relationship between
melatonin and testosterone in the seasonal regulation of testicular steroidogenesis. Further,
alterations in the S-NAT activity and local melatonin level in hamster testis suggest a direct
effect of photoperiodically modulated peripheral and local melatonin levels on testicular

17
steroidogenesis establishing a local microcircuit of this indoleamine in the regulation of
testicular androgen biosynthesis. In the light of the aforementioned, it can be concluded that
seasonality in reproductive status in golden hamster might be modulated by photoperiodic
moderation of the local melatonergic system in the testis.
Declaration of Interest
Authors would like to acknowledge the Council of Scientific and Industrial Research, New
Delhi, government of India (grant scheme number: 37/1462/11/EMR-II) for providing
financial support and Senior Research Fellowship (SRF) to Mr. Arun Mukherjee. Generous
gift of antibodies against LH-receptor and StAR from Dr. Craig S. Atwood (William S.
Middleton Memorial Veterans Hospital, Madison, WI, USA) and Dr. D.M. Stocco (Texas
Tech University, Health Sciences Center, Lubbock, TX, USA) respectively is gratefully
acknowledged. Instrument gift from the Alexander von Humboldt Foundation, Bonn,
Germany to Professor C. Haldar is gratefully acknowledged. The authors report no conflicts
of interest either financial or personal.
References
[1] R.J. Reiter, The pineal gland and seasonal reproductive adjustments, Int J
Biometeorol 19 (1975) 282-8.
[2] J Arendt, Melatonin and the pineal gland: influence on mammalian seasonal and
circadian physiology, Rev Reprod 3 (1998) 13-22.
[3] P Chemineau, L Bodin, M Migaud, J.C. Thiery, B Malpaux, Neuroendocrine and
genetic control of seasonal reproduction in sheep and goats, Reprod Domest Anim
45 (2010) 42-9.
[4] M.B. Frungieri, A Mayerhofer, K Zitta, O.P. Pignataro, R.S. Calandra, S.I.
Gonzalez-Calvar, Direct effect of melatonin on Syrian hamster testes: melatonin
http://www.ncbi.nlm.nih.gov/pubmed?term=Frungieri%20MB%5BAuthor%5D&cauthor=true&cauthor_uid=15550508
http://www.ncbi.nlm.nih.gov/pubmed?term=Mayerhofer%20A%5BAuthor%5D&cauthor=true&cauthor_uid=15550508

18
subtype 1a receptors, inhibition of androgen production, and interaction with the
local corticotropin-releasing hormone system, Endocrinology 146 (2005) 1541-52.
[5] R.J. Reiter, The melatonin rhythm: Both a clock and a calendar, Expenientia 49
(1993) 654-664.
[6] H Dardente, Melatonin-dependent timing of seasonal reproduction by the pars
tuberalis: pivotal roles for long daylengths and thyroid hormones, J
Neuroendocrinol 24 (2012) 249-66.
[7] V Aleandri, V Spina, A Morini, The pineal gland and reproduction, Hum Reprod
Update 2 (1996) 225-35.
[8] D Roy, D.D. Belsham, Melatonin receptor activation regulates GnRH gene
expression and secretion in GT1-7 GnRH neurons. Signal transduction
mechanisms, J Biol Chem 277 (2002) 251-8.
[9] V Filippa, A Penissi, F Mohamed, Seasonal variations of gonadotropins in the
pars distalis male viscacha pituitary. Effect of chronic melatonin treatment, Eur J
Histochem 49 (2005) 291-300.
[10] S.M. Reppert, D.R. Weaver, T Ebisawa, Cloning and characterization of a
mammalian melatonin receptor that mediates reproductive and circadian
responses, Neuron 13 (1994) 1177-85.
[11] M.L. Dubocovich, M Markowska, Functional MT1 and MT2 melatonin
receptors in mammals, Endocrine 27 (2005) 101-10.
[12] G Izzo, A Francesco, D Ferrara, M.R. Campitiello, I Serin, S Minucci, M
d'Istria, Expression of melatonin (MT1, MT2) and melatonin-related receptors in
the adult rat testes and during development, Zygote 18 (2010) 257-64.

19
[13] S Yasuo, T Yoshimura, S Ebihara, H.W. Korf, Melatonin transmits
photoperiodic signals through the MT1 melatonin receptor, J Neurosci 29 (2009)
2885-9.
[14] B.J. Prendergast, MT1 melatonin receptors mediate somatic, behavioral, and
reproductive neuroendocrine responses to photoperiod and melatonin in Siberian
hamsters (Phodopus sungorus), Endocrinol 151 (2010) 714-21.
[15] D.R. Weaver, C Liu, S.M. Reppert, Nature's knockout: the Mel1b receptor is
not necessary for reproductive and circadian responses to melatonin in Siberian
hamsters, Mol Endocrinol 10 (1996) 1478-87.
[16] X Jin, C von Gall, R.L. Pieschl, V.K. Gribkoff, J.H. Stehle, S.M. Reppert,
D.R. Weaver, Targeted disruption of the mouse Mel (1b) melatonin receptor, Mol
Cell Biol. 23 (2003) 1054-60.
[17] R.A. Hoffman, R.J. Reiter, Pineal gland: influence on gonads of male
hamsters, Science 148 (1965) 1609-11.
[18] R.W. Steger, K Matt, A Bartke, Neuroendocrine regulation of seasonal
reproductive activity in the male golden hamster, Neurosci Biobehav Rev 9 (1985)
191-201.
[19] S Valenti, M Giusti, R Guido, G Giordano, Melatonin receptors are present in
adult rat Leydig cells and are coupled through a pertussis toxin-sensitive G-
protein, Eur J Endocrinol 136 (1997) 633-9.
[20] R.J. Reiter, P.K. Rudeen, J.W. Sackman, M.K. Vaughan, L.Y. Johnson, J.C.
Little, Subcutaneous melatonin implants inhibit reproductive atrophy in male
hamsters induced by daily melatonin injections, Endocr Res Commun (1977)
4:35-44.

20
[21] M.M. Bradford, A rapid and sensitive method for the quantitation of
microgram quantities of protein utilizing the principle of protein-dye binding, Anal
Biochem 72 (1976) 248-254.
[22] P Talalay, Hydroxysteroid dehydrogenase. In: Colowick, S.P., Kalplan, N.O.
(Eds.), Methods in Enzymology, Elsevier, New York, 5 (1962) pp. 512–526.
[23] J Jarabak, J.A. Adams, H.G. Williams-Ashaman, P Talalay, Purification of 17
β-hydroxysteroid dehydrogenase function, J Biol Chem 237 (1962) 345-357.
[24] H.D. Chae, T.J. Park, Y.K. Lee, T.G. Lee, K.T. Kim, Rapid and simple
measurement of serotonin N-acetyltransferase activity by liquid biphasic diffusion
assay, Neurochem Int 35 (1999) 447-51.
[25] R.J. Reiter, Pineal control of reproduction, Prog Clin Biol Res 59 (1981) 349-
55.
[26] R.J. Reiter, D.X. Tan, L.C. Manchester, S.D. Paredes, J.C. Mayo, R.M. Sainz,
Melatonin and reproduction revisited, Biol Reprod 81 (2009) 445-56.
[27] A Bartke, Male hamster reproductive endocrinology, In: Siegel HI, ed, The
Hamster, New York: Plenum, (1985) pp. 73-98.
[28] D.K. Vishwas, A Mukherjee, C Haldar, Melatonin improves humoral and cell-
mediated immune responses of male golden hamster following stress induced by
dexamethasone, J Neuroimmunol 134 (2013) 23-36.
[29] T.J. Bartness, G.N. Wade, Photoperiodic control of seasonal body weight
cycles in hamsters, Neurosci Biobehav Rev 9 (1985) 599-612.
[30] M.W. Hance, J.I. Mason, S.M. Mendis-Handagama, Effects of photo
stimulation and nonstimulation of golden hamsters (Mesocricetus auratus) from
birth to early puberty on testes structure and function, Histol Histopathol 24
(2009) 1417-24.

21
[31] E Morales, R Horn, L.M. Pastor, L Santamaria, J Pallares, A Zuasti, C Ferrer,
M Canteras, Involution of seminiferous tubules in aged hamsters: an
ultrastructural, immunohistochemical and quantitative morphological study, Histol
Histopathol 19 (2004) 445-55.
[32] B.H. Caneguim, P.S. Cerri, L.C. Spolidario, S.M. Miraglia, E Sasso-Cerri,
Structural alterations in the seminiferous tubules of rats treated with
immunosuppressor tacrolimus, Reprod Biol Endocrinol 7 (2009) 19.
[33] R Ahmad, C Haldar, Photoperiod-testicular-immune interaction in a seasonal
breeder Indian palm squirrel Funambulus pennanti during the reproductively
inactive and active phases, J Neuroendocrinol 21(2009) 2–9.
[34] R Ahmad, C Haldar, Photoperiodic regulation of MT1 and MT2 melatonin
receptor expression in spleen and thymus of a tropical rodent Funambulus
pennanti during reproductively active and inactive phases, Chronobiol Int 27
(2010) 446-62.
[35] K.M. Tahka, Y.H. Zhuang, S Tahka, P Tuohimaa Photoperiod-induced
changes in androgen receptor expression in testes and accessory sex glands of the
bank vole, Clethrionomys glareolus, Biol Reprod 56 (1997) 898-908.
[36] Z.X. Zhou, M.V. Lane, JA Kemppainen, F.S. French, E.M. Wilson,
Specificity of ligand-dependent androgen receptor stabilization: receptor domain
interactions influence ligand dissociation and receptor stability, Mol Endocrinol 9
(1995) 208-18.
[37] A.H. Payne, D.B. Hales, Overview of steroidogenic enzymes in the pathway
from cholesterol to active steroid hormones, Endocr Rev 25 (2004) 947-70.
[38] D.M. Stocco, StAR protein and the regulation of steroid hormone
biosynthesis, Annu Rev Physiol 63 (2001) 193-213.

22
[39] S.R. King, P.R. Manna, T Ishii, P.J. Syapin, S.D. Ginsberg, K Wilson, L.P.
Walsh, K.L. Parker, D.M. Stocco, R.G. Smith, and D.J. Lamb, An essential
component in steroid synthesis, the steroidogenic acute regulatory protein, is
expressed in discrete regions of the brain, J Neurosci 22 (2002) 10613-10620.
[40] M Ascoli, F Fanelli, D.L. Segaloff, The lutropin/choriogonadotropin receptor,
a 2002 perspective, Endocr Rev 23 (2002) 141-74.
[41] C von Gall, JH Stehle, DR Weaver, Mammalian melatonin receptors:
molecular biology and signal transduction, Cell and Tissue Research 309 (2002)
151-162.
[42] M Tijmes, R Pedraza, L Valladares, Melatonin in the rat testis: evidence for
local synthesis, Steroids 61 (1996) 65-8.
Figure legends
Figure 1
Effect of different photoperiodic conditions (SD and LD) on histoarchitecture and
immunohistochemical localization of MT1R and AR in testes of golden hamster,
Mesocricetus auratus. SD = short-day photoperiod and LD = long-day photoperiod. A)
Showing normal histo-architecture of seminiferous tubule with compact germinal epithelium,
sequentially arranged germ cells at different stages of differentiation with the lumen packed
with spermatozoa under LD condition. B) Showing histological alterations in the
seminiferous tubule under SD condition with signs of testicular atrophy by the presence of
vacuoles and large number of giant cells in the seminiferous tubules with the lumen
completely devoid of sperm. C) Showing weak immunoreactivity for MT1 on the Leydig
cells under LD condition. D) Showing strong immunoreactivity for MT1 on Leydig cells

23
(LC) and peritubular cells (PC) under SD condition. E) Showing strong nuclear
immunoreactivity for AR in the Leydig cells (arrow) and Sertoli cells (dashed arrow) under
LD condition. F) Showing weak nuclear immunoreactivity for AR in the Leydig cells under
SD condition. Scale bar= 50μM.
Figure 2
Effect of different photoperiodic conditions (CP, SD and LD) on testicular steroidogenesis in
golden hamster, Mesocricetus auratus. CP = critical photoperiod, SD = short-day photoperiod
and LD = long-day photoperiod. A) The histogram represents densitometric analyses of AR
and StAR expression in testes. Line diagram on secondary axis shows plasma level of
testosterone. B) The histogram represents densitometric analyses of P450SCC and LH-R
expression in testes. β-actin expression was used as loading control. Values are expressed as
mean ± SEM, N = 5 for each group. Significance of difference; *p < 0.01, CP vs. all
experimental groups and #p < 0.01, SD vs. LD.
Figure 3
Effect of different photoperiodic conditions (CP, SD and LD) on melatonin concentration
(peripheral and local) and MT1R expression in testes of golden hamster, Mesocricetus
auratus. CP = critical photoperiod, SD = short-day photoperiod and LD = long-day
photoperiod. A) The histogram represents plasma level of melatonin and line diagram on
secondary axis shows densitometric analyses of MT1R expression in testes. β-actin
expression was used as loading control. B) The histogram represents testicular S-NAT
enzyme activity and line diagram on secondary axis shows melatonin concentration in testes.
Values are expressed as mean ± SEM, N = 5 for each group. Significance of difference; *p <
0.01, CP vs. all experimental groups and #p < 0.01, SD vs. LD.

24
Table 1
Effect of different photoperiodic conditions (CP, SD and LD) on body weight and relative
testes weight of golden hamster, Mesocricetus auratus. CP = critical photoperiod, SD =
short-day photoperiod and LD = long-day photoperiod. Values are expressed as mean ±SEM.
N = 5 for each group. Significance of difference; *p < 0.01, CP vs. all experimental groups
and #p < 0.01, SD vs. LD.
Table 2
Effect of different photoperiodic conditions (CP, SD and LD) on diameter and area of
semniferous tubule of golden hamster, Mesocricetus auratus. CP = critical photoperiod, SD =
short-day photoperiod and LD = long-day photoperiod. Values are expressed as mean ±SEM.
N = 5 for each group. Significance of difference; *p < 0.01, CP vs. all experimental groups
and #p < 0.01, SD vs. LD.
Table 3
Effect of different photoperiodic conditions (CP, SD and LD) on 3β-HSD and 17β-HSD
enzyme activity in testes of golden hamster, Mesocricetus auratus. CP = critical photoperiod,
SD = short-day photoperiod and LD = long-day photoperiod. Values are expressed as mean
±SEM. N = 5 for each group. Significance of difference; *p < 0.01, CP vs. all experimental
groups and #p < 0.01, SD vs. LD.
Giant Cell
Vacuole
Lumen devoid of
spermatozoa
B
50 µm
Lumen packed with
spermatozoa
50 µm 50 µm
50 µm
Figure 1
LC
C
LC
PC
D
LC
50 µm
50 µm 50 µm
F
LC
LC
LC
A

0
0.5
1
1.5
2
0
20
40
60
80
100
120
CP SD LD
Pla
sm
a te
sto
ste
ron
e (
ng/m
l)
Rela
tive
ba
nd
inte
nsity
AR
STAR
Testosterone
0
10
20
30
40
50
60
70
80
CP SD LD
Rela
tive
ba
nd
inte
nsity
P450SCC
LHR
P450SCC (49 KDa)
LH-receptor (85 KDa)
β-actin (42 KDa)
AR (110 KDa)
StAR (30 KDa)
CP SD LD
β-actin (42 KDa)
Figure 2
A B
*
* #
*
*
* #
* #
* *
* #
* #
CP SD LD
0
5
10
15
20
25
0
20
40
60
CP SD LD
Me
lato
nin
co
nce
ntr
ation
in te
stis
(fm
ol/gm
tis
su
e)
nm
ole
s n
-ace
tyl tr
yp
tam
ine
fo
rme
d/m
in/g
m tis
su
e
S-NAT
Melatonin
0
15
30
45
0
5
10
15
20
25
30
CP SD LD
Re
lative
ba
nd
in
ten
sity
Pla
sm
a m
ela
ton
in (
pg/m
l)
Melatonin
MT1R
β-actin (42 KDa)
CP SD LD
MT1 (~37 KDa)
Figure 3
A
B
*
* #
*
# *
*
* #
*
* #

Groups CP SD LD
Body weight (g)
135.33±4.62
170.34±5.33 *
123.33±4.72 *#
Relative testes weight (g/100g body wt.) 1.665±0.15
0.4095±0.083 *
1.852±0.108 *#
Table 1
Effect of different photoperiodic conditions (CP, SD and LD) on body weight and relative testes weight of
golden hamster, Mesocricetus auratus. CP = critical photoperiod, SD = short-day photoperiod and LD =
long-day photoperiod. Values are expressed as mean ±SEM. N = 5 for each group.
* Significance of difference; p < 0.01, CP vs. all experimental groups. # Significance of difference; p < 0.01, SD vs. LD.

Table 2
Effect of different photoperiodic conditions (CP, SD and LD) on diameter and area of semniferous tubule
of golden hamster, Mesocricetus auratus. CP = critical photoperiod, SD = short-day photoperiod and LD =
long-day photoperiod. Values are expressed as mean ±SEM. N = 5 for each group.
Groups CP SD LD
Seminiferous tubular diameter (µm)
179..43±8.42
89.22±6.34 *
239.75±8.88 *#
Seminiferous tubular area (µm2×103) 51.34±2.08
26.73±0.88 *
64.66±1.45 *#
* Significance of difference; p < 0.01, CP vs. all experimental groups. # Significance of difference; p < 0.01, SD vs. LD.

Groups YH AH OH
3β-HSD activity (U/mg tissue/hr.) 30.63±1.11
16.54±0.50 *
35.70±0.82 *#
17β-HSD activity (U/mg tissue/hr.) 24.59±0.58
11.09±0.48 *
36.44±1.45 *#
Table 3
Effect of different photoperiodic conditions (CP, SD and LD) on 3β-HSD and 17β-HSD enzyme activity in
testes of golden hamster, Mesocricetus auratus. CP = critical photoperiod, SD = short-day photoperiod
and LD = long-day photoperiod. Values are expressed as mean ±SEM. N = 5 for each group.
* Significance of difference; p < 0.01, CP vs. all experimental groups. # Significance of difference; p < 0.01, SD vs. LD.

Highlights
Present study established photoperiodic modulation of MT1R expression in testes for
the first time.
Presence of a local melatonergic system and its photoperiodic modulation is being
proposed.
Photoperiodically modulated peripheral and local melatonin might regulate testicular
steroidogenesis via MT1R.
Photoperiod via melatonin modulate testicular steroidogenesis to restrict male fertility
to a particular time of year.




















