
REPRESENTACIÓN DE LENGUAJES DE PATRONES DE ANÁLISIS DE DOMINIO

248
Rev. Fac. Agron. (LUZ). 2012, 29: 248-272
Efecto de dietas proteicas a base de harina delombríz roja (Eisenia spp.) en el comportamiento del
hámster dorado (Mesocricetus auratus L.).I. Dinámica de peso y digestibilidad
Effect of protein diets based on red worm (Eisenia spp.)meal in the behavior of golden hamster (Mesocricetus
auratus L.). I. weight dynanic and digestibility
D.E. García1, L.J. Cova2, J.V. Scorza D.2, M.E. González3, P. Pizzani4,M. G. Medina1, F. Perea1 y D. González1
1Departamento de CienciasAgrarias, NúcleoUniversitario “RafaelRangel” (NURR), Universidad de Los Andes (ULA), estado Trujillo,Venezuela2Instituto Experimental “José WitremundoTorrealba”, ULA, estadoTrujillo, Venezuela3Departamento de Biología y Química, NúcleoUniversitario NURR,ULA, estado Trujillo, Venezuela4Área Agronomía, Universidad Nacional Experimental “RómuloGallegos”,estado Guárico, Venezuela.
Resumen
Se evaluó el consumo de agua, dinámica del peso vivo y metabólico, ydigestibilidad aparente de la materia seca (DAMS) en 72 Hámsters Dorados(Mesocrisetus auratus L.) que consumieron dietas (n=12 cada una) elaboradas abase de harina de Lombriz Roja (Eisenia spp.; HL) y descritas de la forma si-guiente: D1: PB=13,5% (HL: 21%); D2: PB=15,4% (HL: 24%); D3: PB=17,6% (HL:27%); D4: PB=19,6% (HL: 30%) y dos alimentos convencionales (D0: Conejarina®;PB11,4% y D5: Ratarina®; PB=21,1%). El estudio se realizó en el Vivario delNúcleo Universitario “Rafael Rangel” de la Universidad de Los Andes, Trujillo,Venezuela. Los datos se analizaron mediante un diseño totalmente aleatorizadopara medidas repetidas en el tiempo. El mayor consumo diario de agua se observóa los 9 días en el grupo alimentado con D4 (16,33 mL/animal), mientras que en elresto de los grupos la mayor ingestión de agua ocurrió el día 18 del experimento.Los hámsters del grupo D4 fueron luego retirados del ensayo y sacrificados porexhibir signos de intoxicación. La mayor ganancia de peso (>1g/animal/día) se
Recibido el 27-1-2010 Aceptado el 30-1-2012Autor de correspondencia e-mail: [email protected]

Rev. Fac. Agron. (LUZ). 2012, 29: 248-272
249
observó a partir de los 15 días de iniciado el experimento en los animales alimen-tados con D1 y D5, siendo estos mismos tratamientos lo que presentaron la mayorDAMS promedio (73,03 y 77,61%, respectivamente). La alimentación con D3 y D4afectó drásticamente la ganancia de peso y la DAMS en los animales (P<0,05).Con D1 se logró una ganancia de peso y una digestibilidad similar a la obtenidasólo con Ratarina®. La HL constituyó un suplemento proteico de buena calidadnutritiva para el hámster dorado cuando fue incluida en la dieta en una propor-ción del 21%.Palabras clave: Harina de lombriz, composición nutricional, Eisenia spp.,hámster dorado, digestibilidad.
Abstract
It was evaluated the water intake, dynamics of body and metabolic weightand apparent digestibility of dry matter (DAMS) in 72 Golden Hamster (Mesocrisetusauratus L.) that consumed diets (n=12) made with red worm (Eisenia spp.; HL)meal and describe as: D1: PB=13,5% (HL: 21%); D2: PB=15,4% (HL: 24%); D3:PB=17,6% (HL: 27%); D4: PB=19,6% (HL: 30%) and two commercial feeds (D0:Conejarina®; PB=11.1% and D5: Ratarina®; PB=21,1%). The study was conductedat the vivarium of the Núcleo Universitario “Rafael Rangel, Universidad de LosAndes, Trujillo, Venezuela. The data were analyzed by a completely randomizeddesign with repeated measures. The main water intake was observed in animalsfed with D4 (16.33 mL.animal-1) on day 9 of the experiment, while in the rest of thegroups the maximum water consumption was on day 18. The hamsters of D4group were later removed of the assay and sacrificed because exhibited foodintoxication symptoms. Starting from day 15 of the experiment, the animals fedwith D1 and D5 presented the greater weight gain (>1g.animal.day-1); also animalsfrom D1 and D5 groups had the greater DAMS average (73.03 and 77.61%,respectively). Animals that consumed D3 and D4 affected drastically the weightgain and DAMS in the animals (P<0.05). However, with D1 a similar weight gainand DAMS were obtained compared with Ratarina®. The HL represented a proteinsupplement of good nutritional quality for the Golden Hamster when was includedin a proportion of 21% in the diet.Key words: earthworm meal, nutritional composition, Eisenia spp., goldenhamster, digestibility.
Introducción
La evaluación de fuentes de ali-mentos no convencionales para anima-les es, en la actualidad, uno de los prin-cipales objetivos de los profesionalesque trabajan en el área de la nutrición
Introduction
The evaluation of nonconventional food sources for animalsis nowadays one of the main objectivesof professionals who work in thenutrition area in Latin America,

García et al.
250
en los países Latinoamericanos, don-de la disponibilidad de ingredientes deelevado valor nutricional es limitada.Si bien es conocido que existen nume-rosos forrajes y residuosagroindustriales que constituyen alter-nativas para la suplementación de losrumiantes (García et al., 2009), la eva-luación de alimentos no tradicionalesque sean factibles para no rumiantesconstituye un reto más difícil, consi-derando los elevados requerimientosque presentan estos animales y algu-nos aspectos intrínsecos asociados conlos sistemas intensivos de producción(Savón et al., 2008).
Una de las principales limitantesen la búsqueda de alimentos adecua-dos para estos animales radica en laconcentración y la calidad de la proteí-na presente en dichas fuentes, la cualen todos los casos no es adecuada paramantener los estándares de produccióna escala comercial. En este sentido, laharina de la Lombriz Roja (Eiseniaspp.) se perfila como una importantefuente de proteínas de calidad (>50%en base seca) y de aminoácidos de ele-vada disponibilidad, para ser utiliza-da en la nutrición animal (Morón-Fuenmayor et al., 2008). Sin embar-go, existen pocas evaluaciones en lascuales se ha estudiado su viabilidadcomo material suplementario (Cova etal., 2009).
Por otra parte, los animales delaboratorio constituyen no solo mode-los biológicos idóneos para realizar es-tudios de fisiología y farmacología apli-cada, sino también para desarrollarinvestigaciones relacionadas con otrasáreas del conocimiento (De Jesús yQuintero, 2001; De Jesús y Quintero,2008); prescindiendo del uso de otros
where the availability of ingredientswith high nutritional value is limited.It is well known that there arenumerous fodder and agro industrialresidues that are alternatives forfeeding the ruminants (García et al.,2009), the evaluation of nontraditionalfood feasible for monogastric constitutea difficult goal, considering the elevaterequirements that these animals have,and some intrinsic aspects related tointensive production systems (Savónet al., 2008).
One of the main limiting factorssearching the adequate food for theseanimals relies on the concentrationand quality of protein present in suchsources, which in all cases is notadequate to keep the standards ofproduction at a commercial scale. Inthe sense, flour of red worm (Eiseniaspp.) seems to be as an importantsource of high-quality protein (>50%in dry base) and of amino acids withhigh availability to be used in the ani-mal nutrition (Morón-Fuenmayor etal., 2008). However, there are fewresearches where has been studied itsvariability as supplementary material(Cova et al., 2009).
On the other hand, thelaboratory animals do not onlyconstitute suitable biological models tocarry physiology and appliedpharmacology researches, but also todevelop researches related to otherknowledge areas (De Jesús and Quin-tero, 2001; De Jesús and Quintero,2008); excluding the use of biggeranimals that require higher quantityof food in their diets for the realizationof nutritional tests. In this matter,Gold Hamster (Mesocricetus auratusL.) constitutes an adequate alternative

Rev. Fac. Agron. (LUZ). 2012, 29: 248-272
251
animales más grandes que requierenmayor cantidad de alimento en susdietas para la realización de pruebasnutricionales. En este sentido, elHámster Dorado (Mesocricetusauratus L.) constituye una alternati-va adecuada para evaluar preliminar-mente las potencialidades de alimen-tos experimentales por su fácil mane-jo, baja capacidad ingestiva y docili-dad (Llewellyn et al., 1985; Carr et al.,1996; Campbell et al., 2009).
Por tales motivos, el objetivo deesta investigación fue estudiar el usode dietas a base de harina deshidratadade Eisenia spp., y otras fuentes de ali-mentos convencionales, sobre el com-portamiento de M. auratus en térmi-nos de dinámica de peso vivo ymetabólico, consumo de agua ydigestibilidad aparente del alimento.
Materiales y métodos
Localización del ensayoEl estudio se desarrolló en el
Vivario del Núcleo Universitario “Ra-fael Rangel” (NURR) de la Universi-dad de Los Andes (ULA), Trujillo, es-tado Trujillo, Venezuela, que se en-cuentra localizado a 800 msnm, conuna temperatura media anual de25,2ºC y 69% de humedad relativa.
Condiciones de experimenta-ción
Se utilizaron 72 hámsters hem-bras (M. auratus) con un peso prome-dio de 81,45 ± 8,5 g, provenientes delBioterio de la Facultad de Ciencias dela ULA, Mérida, Venezuela; los cua-les fueron divididos en 12 grupos deseis animales y distribuidos de formaaleatoria en jaulas de acero inoxida-ble de 45,5 cm de largo, 40,0 cm de
for evaluating the potentialities of ex-perimental food by the easy handle, lowdigestive capacity and docility(Llewellyn et al., 1985; Carr et al.,1996; Campbell et al., 2009).
For such reasons, the objective ofthis research was to study the use ofdiets based on dehydrated flours ofEisenia spp., and other sources ofconventional food, on the behavior of M.auratus in terms of dynamic of alive andmetabolic weight, consumption of waterand apparent digestibility of the food.
Materials and methods
Location of the essayThe research was developed at
the vivarum of the University campus“Rafael Rangel” (NURR) of Universi-dad de los Andes (ULA), Trujillo,Trujillo state, Venezuela, located at800 masl with an annual meantemperature of 25.2°C and 69% ofrelatively humidity.
Experiment conditions72 female hamsters (M.
auratus) were used with an averageweight of 81.45±8.5 g, coming fromthe Botanic of the Science Faculty ofULA, Mérida, Venezuela; which weredivided in 12 group of six animalsdistributed at random in stainlesssteel cages of 45.5 cm of length and40.0 cm of width and 22 cm of heightwith interchangeable metallic traysin the inferior part for collecting theexcrements. During all the essayphase, the animals were submitted tolight-darkness cycles of 12 h, withsupply of water and daily offer of food20% superior to the indicated byhamsters using maintenance diets(NRC, 1995).

García et al.
252
ancho y 22 cm de altura, con bande-jas metálicas intercambiables en laparte inferior para la recolección deexcretas. Durante todas las etapas delensayo los animales fueron sometidosa ciclos de luz-oscuridad de 12 h, consuministro de agua a voluntad y unaoferta diaria de alimento de un 20%superior a lo indicado para hámsteresusando dietas de mantenimiento(NRC, 1995).
Para la adaptación a las condi-ciones de experimentación los anima-les fueron confinados una semana pre-via al inicio del experimento, en lascondiciones descritas anteriormente;en dicho periodo se les suministró dia-riamente una dieta peletizada consti-tuida por 11,4% de proteína bruta (PB);1,2% de grasas; 44,0% de fibra deter-gente neutro (FDN); 64,4% de extrac-to libre de nitrógeno (ELN), 8,6% decenizas y agua ad libitum. Posterior-mente al periodo pre-experimental, to-dos los días a las 7:00 am, y por 21días, se les ofertaron las dietas en loscomederos de cada jaula, después dela respectiva dosificación diaria; reti-rándose el remanente del día anterior,que fue pesado para determinársele elcontenido de materia seca (MS)(AOAC, 1990).
Ingredientes, formulación dedietas y tratamientos
Para la formulación de las racio-nes, de acuerdo a los tratamientos, seusaron varios ingredientes; estos fue-ron: pasto estrella (Cynodonnlemfluensis), granos de maíz (Zeamays L.), gelatina tratada (Pandock,Distribuidora Valera, Trujillo, Vene-zuela), carbonato de calcio (CaCO3),fosfato monocálcico (Ca(H2PO4)2) ymelaza. La composición química de los
For the adaptation to experimen-tal conditions, the animals wereconfined a week prior to theexperiment, in the conditionspreviously described, in such periodwere daily provided with a pelletingdiet constituted by 11.4% of rawprotein (PB); 1.2% of fats; 44.0% ofneutral detergent fiber (FDN); 64.4%of extract free of nitrogen (ELN), 8.6%of ashes and ad libitum water.Consequently to the pre experimentalperiod, all days at 7:00 am and for 21days, were given the diets on each cage,after their respective daily dose,retiring the remnant the day after,which was weighted to determine thecontent of dry matter (MS) (AOAC,1990).
Ingredients, formulation ofdiets and treatments
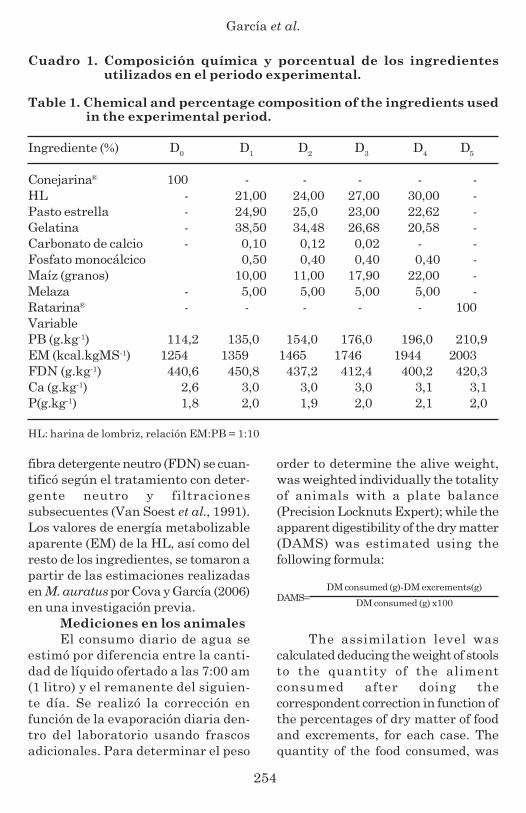
For formulating the portions,according to the treatments, differentingredients were used, these were:Cynodon nlemfluensis, grains of corn(Zea mays L.), treated gelatin (Pandock,Distributor Valera, Trujillo, Venezue-la), calcium carbonate (CaCO3),monocalcium phosphate (Ca(H2PO4)2)and molasses. The chemicalcomposition of the ingredients and theformulated diets are in table 1.
Restructuring of the foodand obtaining of the pellet
Considering that Ratarina® andConejarina® presented different pelletsizes, these food were initiallyrestructured with comparative purposes.Initially were grinded using a crown-typegrinder (14/249, Royal Triumph brand),which was also used for the rest of thecomponents of the diets.
The ingredients of each diet werehomogenized sequentially, employing

Rev. Fac. Agron. (LUZ). 2012, 29: 248-272
253
ingredientes y las dietas formuladasse muestra en el cuadro 1.
Restructuración del alimen-to y obtención de los pellet
Considerando que la Ratarina® yla Conejarina® presentaban tamañosde pellet diferentes, estos alimentosfueron inicialmente restructurados confines comparativos. Inicialmente semolieron usando un molino tipo coro-na (14/249, marca Royal Triumph),que también se utilizó para el resto delos componentes de las dietas.
Los ingrediente de cada dieta sehomogenizaron, de forma secuenciada,empleando un homogenizador eléctri-co (Equifarm, marcaULTRATURRAX). Para larestructuración se usó unapeletizadora de 8 cuchillas de corte convariador electrónico de velocidad ymotor de 3HP. Los pellet formados fue-ron secados en una estufa con ventila-ción forzada (YRH 02-3, marcaKaltein) durante 48 horas y despuésde enfriarse se mantuvieron a tempe-ratura ambiente (25°C). Se dosificaronlas cantidades a ofertar en los comede-ros según cada tratamiento.
Las características físicas de lospellet de cada dieta fueron las siguien-tes: peso = 1,40 ± 0,2 g/pellet; largo =1,83 ± 0,5 cm; ancho = 1,03 ± 0,2 cm.Parámetros de dureza: absorción deagua a los 15 minutos = 66,30 ± 0,2%(p/p); distancia de fracturabilidad =0,35 ± 0,02 cm y tiempo defracturabilidad = 0,08 ± 0,002s.
Mediciones analíticas en losalimentos
Se determinaron los contenidosde PB, calcio (Ca) y fósforo (P), mediantelas metodologías tradicionales paraanálisis de alimentos (AOAC, 1990). La
an electrical homogenizer (Equifarm,ULTRATURRAX brand). For therestructuration was used a pellet presswith 8-cut knives with electricalvariator of velocity and a motor of 3HP.The pellet forms were let dried in astove with forced ventilation (YRH 02-3, Kaltein brand) for 48 hours and oncecold were kept at environmenttemperature (25°C). The quantities tobe offered were dosed in the feed hopperaccording to each treatment.
The physical characteristics ofthe pellet of each diet were the following:weight= 1.40 ± 0.2 g/pellet; length =1.83±0.5 cm; width = 1.03±0.2 cm.Hardness parameters: absorption ofthe water at 15 minutes = 66.30±0.2%(p/p); fracturability distance = 0.35 ±0.02 cm and fracturability time =0.08±0.002s.
Analytical measures in foodThe contents of PB, calcium (Ca)
and phosphorus (P) were determined,using the traditional methodologies forthe analysis of food (AOAC, 1990). Theneutral detergent fiber (FDN)quantified according to the neutraldetergent and subsequent filtration(Van Soest et al., 1991). The apparentmetabolizable energy values (EM) ofHL, as well as the rest of theingredients, were taken after theestimations done in M. auratus porCova and García (2006) in a previousresearch.
Measures in animalsThe daily consumption was
estimated by difference between thequantity of liquid offered at 7:00 am (1liter) and the remnant the followingday. The correction was done infunction of the daily evaporation in thelaboratory using additional bottles. In
García et al.
254
fibra detergente neutro (FDN) se cuan-tificó según el tratamiento con deter-gente neutro y filtracionessubsecuentes (Van Soest et al., 1991).Los valores de energía metabolizableaparente (EM) de la HL, así como delresto de los ingredientes, se tomaron apartir de las estimaciones realizadasen M. auratus por Cova y García (2006)en una investigación previa.
Mediciones en los animalesEl consumo diario de agua se
estimó por diferencia entre la canti-dad de líquido ofertado a las 7:00 am(1 litro) y el remanente del siguien-te día. Se realizó la corrección enfunción de la evaporación diaria den-tro del laboratorio usando frascosadicionales. Para determinar el peso
Cuadro 1. Composición química y porcentual de los ingredientesutilizados en el periodo experimental.
Table 1. Chemical and percentage composition of the ingredients usedin the experimental period.
Ingrediente (%) D0 D1 D2 D3 D4 D5
Conejarina® 100 - - - - -HL - 21,00 24,00 27,00 30,00 -Pasto estrella - 24,90 25,0 23,00 22,62 -Gelatina - 38,50 34,48 26,68 20,58 -Carbonato de calcio - 0,10 0,12 0,02 - -Fosfato monocálcico 0,50 0,40 0,40 0,40 -Maíz (granos) 10,00 11,00 17,90 22,00 -Melaza - 5,00 5,00 5,00 5,00 -Ratarina® - - - - - 100VariablePB (g.kg-1) 114,2 135,0 154,0 176,0 196,0 210,9EM (kcal.kgMS-1) 1254 1359 1465 1746 1944 2003FDN (g.kg-1) 440,6 450,8 437,2 412,4 400,2 420,3Ca (g.kg-1) 2,6 3,0 3,0 3,0 3,1 3,1P(g.kg-1) 1,8 2,0 1,9 2,0 2,1 2,0
HL: harina de lombriz, relación EM:PB = 1:10
DAMS=DM consumed (g)-DM excrements(g)
DM consumed (g) x100
order to determine the alive weight,was weighted individually the totalityof animals with a plate balance(Precision Locknuts Expert); while theapparent digestibility of the dry matter(DAMS) was estimated using thefollowing formula:
The assimilation level wascalculated deducing the weight of stoolsto the quantity of the alimentconsumed after doing thecorrespondent correction in function ofthe percentages of dry matter of foodand excrements, for each case. Thequantity of the food consumed, was

Rev. Fac. Agron. (LUZ). 2012, 29: 248-272
255
vivo se pesó de forma individual latotalidad de los animales con unabalanza de plato (PrecisionLocknuts Expert); mientras que ladigestibilidad aparente de la mate-ria seca (DAMS) se estimó median-te la siguiente fórmula:
El nivel de asimilación se calcu-ló restando el peso de las heces a lacantidad de alimento consumido, lue-go de realizar la corrección correspon-diente en función de los porcentajes demateria seca del alimento y lasexcretas, en cada caso. La cantidad dealimento consumido se estimó por ladiferencia entre la oferta y el rechazoen cada jaula, dividiendo el valor re-sultante entre el número de animalespresentes al momento de cada evalua-ción. El manejo, los cuidados y el man-tenimiento de los hamsters se realiza-ron siguiendo las normas vigentes parael cuidado de animales de laboratorio(Home Office, 1989).
Diseño experimental y aná-lisis estadístico
Se empleó un diseño totalmentealeatorizado para medidas repetidas enel tiempo, en el cual las dietas evalua-das constituyeron los tratamientos.Para el procesamiento de la informa-ción se utilizó el paquete estadísticoSPSS versión 10.0 para Windows®
(Visauta, 1998). Para comparar lostratamientos en función de las varia-bles medidas se empleó la prueba decomparación de SNK (Student-Newman-Keuls) para un 5% de proba-bilidad.
estimated by the difference betweenthe offer and the rejection on each cage,dividing the resultant numberbetween the numbers of animalspresent at the moment of eachevaluation. The handle, cares andmaintenance of the hamster were donefollowing the current norms for thehandle and care of laboratory animals(Home Office, 1989).
Experimental design andstatistical analysis
A completely randomized designwas applied for repeated measures,where the evaluated diets constitutedthe treatments. For processing theinformation, statistical software SPP,version 10.0 for Windows® was used(Visauta, 1998). In order to comparethe treatments in function of the va-riables measured, the SNKcomparison test (Student-Newman-Keuls) was used, for 5% of probability.
Results and discussion
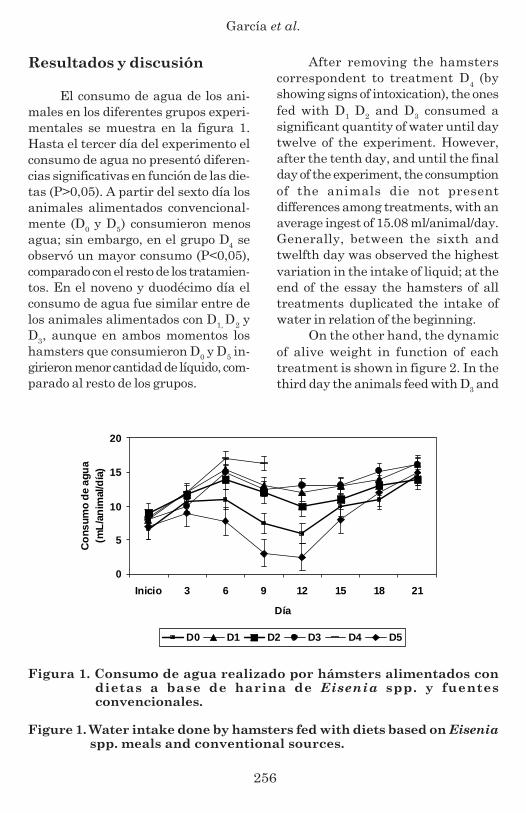
The water consumption by handsof animals in the different experimen-tal groups is shown in figure 1. Untilthe third day of the experiment thewater consumption did not presentsignificant differences in function of thediets (P>0.05). After the sixth day, theanimals fed conventionally (D0 and D5)consumed less water, however, in groupD4 was observed a higher consumption(P<0.05), compare to the rest of thetreatments. In the ninth and twelfthday, the consumption of water was si-milar between the animals fed with D1D2 and D3, even though in bothmoments the hamsters that consumedD0 and D5 drank less quantity of liquidcompare to the rest of the groups.
DAMS=Consumo de MS(g)-peso seco de las heces(g)
Consumo de MS(g) x100
García et al.
256
Resultados y discusión
El consumo de agua de los ani-males en los diferentes grupos experi-mentales se muestra en la figura 1.Hasta el tercer día del experimento elconsumo de agua no presentó diferen-cias significativas en función de las die-tas (P>0,05). A partir del sexto día losanimales alimentados convencional-mente (D0 y D5) consumieron menosagua; sin embargo, en el grupo D4 seobservó un mayor consumo (P<0,05),comparado con el resto de los tratamien-tos. En el noveno y duodécimo día elconsumo de agua fue similar entre delos animales alimentados con D1, D2 yD3, aunque en ambos momentos loshamsters que consumieron D0 y D5 in-girieron menor cantidad de líquido, com-parado al resto de los grupos.
0
5
10
15
20
Inicio 3 6 9 12 15 18 21
Día
Con
sum
o de
agu
a (m
L/an
imal
/día
)
D0 D1 D2 D3 D4 D5
Figura 1. Consumo de agua realizado por hámsters alimentados condietas a base de harina de Eisenia spp. y fuentesconvencionales.
Figure 1. Water intake done by hamsters fed with diets based on Eiseniaspp. meals and conventional sources.
After removing the hamsterscorrespondent to treatment D4 (byshowing signs of intoxication), the onesfed with D1 D2 and D3 consumed asignificant quantity of water until daytwelve of the experiment. However,after the tenth day, and until the finalday of the experiment, the consumptionof the animals die not presentdifferences among treatments, with anaverage ingest of 15.08 ml/animal/day.Generally, between the sixth andtwelfth day was observed the highestvariation in the intake of liquid; at theend of the essay the hamsters of alltreatments duplicated the intake ofwater in relation of the beginning.
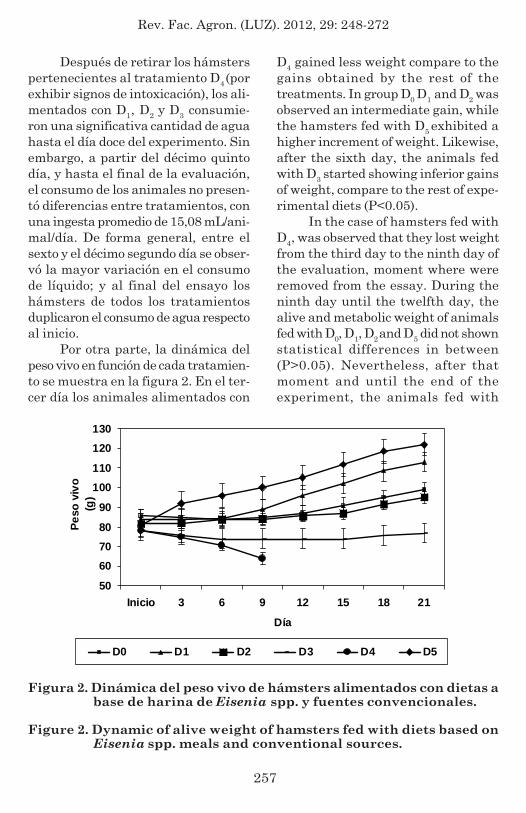
On the other hand, the dynamicof alive weight in function of eachtreatment is shown in figure 2. In thethird day the animals feed with D3 and
Rev. Fac. Agron. (LUZ). 2012, 29: 248-272
257
Después de retirar los hámsterspertenecientes al tratamiento D4 (porexhibir signos de intoxicación), los ali-mentados con D1, D2 y D3 consumie-ron una significativa cantidad de aguahasta el día doce del experimento. Sinembargo, a partir del décimo quintodía, y hasta el final de la evaluación,el consumo de los animales no presen-tó diferencias entre tratamientos, conuna ingesta promedio de 15,08 mL/ani-mal/día. De forma general, entre elsexto y el décimo segundo día se obser-vó la mayor variación en el consumode líquido; y al final del ensayo loshámsters de todos los tratamientosduplicaron el consumo de agua respectoal inicio.
Por otra parte, la dinámica delpeso vivo en función de cada tratamien-to se muestra en la figura 2. En el ter-cer día los animales alimentados con
50
607080
90
100110
120130
Inicio 3 6 9 12 15 18 21
Día
Peso
viv
o(g
)
D0 D1 D2 D3 D4 D5
Figura 2. Dinámica del peso vivo de hámsters alimentados con dietas abase de harina de Eisenia spp. y fuentes convencionales.
Figure 2. Dynamic of alive weight of hamsters fed with diets based onEisenia spp. meals and conventional sources.
D4 gained less weight compare to thegains obtained by the rest of thetreatments. In group D0 D1 and D2 wasobserved an intermediate gain, whilethe hamsters fed with D5 exhibited ahigher increment of weight. Likewise,after the sixth day, the animals fedwith D3 started showing inferior gainsof weight, compare to the rest of expe-rimental diets (P<0.05).
In the case of hamsters fed withD4, was observed that they lost weightfrom the third day to the ninth day ofthe evaluation, moment where wereremoved from the essay. During theninth day until the twelfth day, thealive and metabolic weight of animalsfed with D0, D1, D2 and D5 did not shownstatistical differences in between(P>0.05). Nevertheless, after thatmoment and until the end of theexperiment, the animals fed with

García et al.
258
D3 y D4 ganaron menos peso, compa-rado con las ganancias obtenidas conel resto de los tratamientos. En los gru-pos D0, D1 y D2 se observó una ganan-cia intermedia; mientras que loshámsters alimentados con D5 exhibie-ron un mayor incremento de peso. Asi-mismo, a partir del sexto día los ani-males alimentados con D3 comenzarona exhibir ganancias inferiores de peso,comparado con el resto de las dietasexperimentales (P<0,05).
En el caso de los hámsters alimen-tados con D4, se observó pérdida de pesodesde el tercer al noveno día de evalua-ción, momento en el cual fueron retira-dos del ensayo. Durante el noveno y elduodécimo día el peso vivo y metabólicode los animales alimentados con D0, D1,D2 y D5 no mostraron diferencias esta-dísticas entre sí (P>0,05). No obstante,a partir de ese momento, y hasta el fi-nal del experimento, los animales ali-mentados con Ratarina® (grupo D5) pre-sentaron mayor tasa de crecimiento,seguido por los hámsters que consumie-ron D1 (P>0,05). Las ganancias de pesode los animales alimentados con D0 yD2 no exhibieron variaciones significa-tivas entre sí (P>0,05), aunque fueroninferiores a los animales de los gruposD1 y D5.
En general, las tendencias indi-can una ganancia de peso superior enel grupo D5, secundado por D1, que fue-ron seguidos por los grupos D0 y D2.En el caso de D3, no se observó aumen-to sostenido de peso, mientras que losanimales alimentados con D4 tendie-ron a perder masa corporal desde elcomienzo del experimento. Asimismo,cuando los datos fueron corregidos apeso metabólico (kg0,75) se observó lamisma tendencia en cada punto de
Ratarina® (group D5) presented ahigher growth rate, followed byhamsters who consumed D1 (P>0.05).The weight gains of animals fed withD0 and D2 did not show significantvariations among them (P>0.05),though were inferior to animals ofgroup D1 and D5.
In general, the tendenciesindicate a superior weight gain ingroup D5, followed by D1, which werefollowed by D0 and D2. In the case ofD3, it was not observed a steadyincrement of weight, while the animalsfed with D4 tended to lose corporal massfrom the beginning of the experiment.Likewise, when the data was correctedto metabolic weight (kg0.75) wasobserved the same tendency on eachpoint of analysis, showing that the bestgains of weight were detected in theanimals of group D1 and D5.
DAMS of the diets throughoutthe experimental period are shown infigure 3. At the beginning of the essay,the digestibility of all diets were simi-lar in between (P0.05). However, thethird day of the evaluation D5 exhibitedhigher apparent digestibility compareto the rest of the formulas. In the sixthday, DAMS of groups D1 and D5 werestatistically superior to the rest of thediets (P<0.05); while in the fourth andfifth evaluation (days 9 and 12) thedigestibility with Conejarina® was in-ferior to the rest of the treatments.After the fifteenth day D0 D2 and D3exhibited less apparent digestibilityuntil the end of the experiment,without observing significantdifferences among them (P>0.05).Generally, was observed an incrementtendency of DAMS between the sixthand fifteenth day of the evaluation,

Rev. Fac. Agron. (LUZ). 2012, 29: 248-272
259
análisis, demostrándose que las mejo-res ganancias de peso se detectaron enlos animales de los grupos D1 y D5.
La DAMS de las dietas duranteel período experiental se muestra enla figura 3. Al inicio del ensayo ladigestibilidad de todas las dietas fue-ron similares entre sí (P>0,05). Sinembargo, al tercer día de evaluaciónD5 exhibió mayor digestibilidad aparen-te, comparada con el resto de lasformulaciones. En el sexto día, laDAMS de los grupos D1 y D5 fueronsuperiores estadísticamente al resto delas dietas (P<0,05); mientras que enla cuarta y quinta evaluación (días 9 y12) la digestibilidad de la Conejarina®
fue inferior a los demás tratamientos.A partir del décimo quinto día D0, D2 yD3 exhibieron la menor digestibilidadaparente hasta el final del experimen-to, sin observarse diferencias signifi-cativas entre sí (P>0,05). En sentido
Figura 3. Digestibilidad aparente de la materia seca de hámstersalimentados con dietas a base de harina de Eisenia spp. yfuentes convencionales.
Figure 3. Apparent digestibility of dry matter of hamsters fed withdiets based on Eisenia spp. meals and conventional sources.
0
20
40
60
80
100
Inicio 3 6 9 12 15 18 21
Día
DA
MS
(%)
D0 D1 D2 D3 D4 D5
moment where the percentages ofdigestible MS reduced for D2 and D3.The averages of DAMS for each dietduring the experimental period wereD0: 42.81 ± 6.5; D1: 73.03 ± 4.3; D2:54.55 ± 7.5; D3: 50.19 ± 9.7; D4: 42.68± 9.9; D3: 77.61 ± 6.5%.
Water is an essential element forliving beings, and in all the cases thepercentage of corporal composition issuperior to 56%, which might indicatethat the consumption level is relatedto a great quantity of vital activities.On this matter, is known that inanimals the highest intake of wateroccurs immediately after theconsumption of food, likewise, thevolume of water drank is determinedby the quality and quantity of thealiment eaten (De Vries, 1996). Onthis sense, different researches havebeen done in different types of experi-mental animals, where is informed an

García et al.
260
general, se observó una tendencia deincremento de la DAMS entre el sextoy el décimo quinto día de evaluación;momento en el cual los porcentajes deMS digestible disminuyeron para D2 yD3. Los promedios de DAMS para cadadieta durante el periodo experimentalfueron: D0: 42,81 ± 6,5; D1: 73,03 ± 4,3;D2: 54,55 ± 7,5; D3: 50,19 ± 9,7; D4:42,68 ± 9,9; D3: 77,61 ± 6,5%.
El agua es un elemento esencialpara los seres vivos, y en todos los ca-sos el porcentaje de composición cor-poral es superior al 56%, lo cual pre-supone que el nivel de consumo se en-cuentre relacionado con una gran can-tidad de actividades vitales. Al respec-to, se conoce que en los animales lamayor ingesta de agua ocurre inme-diatamente después del consumo delalimento; asimismo, el volumen deagua consumido está determinado porla calidad y la cantidad de alimentoingerido (De Vries, 1996). En este sen-tido, se han realizado numerosos es-tudios, en diferentes tipos de anima-les experimentales, en los cuales seinforma una relación importante en-tre la composición de las dietas y elconsumo de agua (De Vries, 1996;Rees y Watson, 2000; Ferreira et al.,2002). Sin embargo, algunas investi-gaciones han mostrado resultados di-vergentes; ya que el comportamientoanimal se encuentra influenciado porelementos innatos y aprendidos, porla preferencia específica por un tipode alimento y por el estrés fisiológicoy metabólico, tanto de animales enproducción como de laboratorio (Misray Singh, 2002). Por lo tanto, el com-portamiento en el consumo de líqui-dos cuando se suministran dietas ex-perimentales, podría estar sujeto a las
important relation between thecomposition of diets and the intake ofwater (De Vries, 1996; Rees andWatson, 2000; Ferreira et al., 2002).However, some researchers haveshown divergent results, since theanimal behavior is influenced byinnate and learned elements, by thespecific preference by a type of foodand by the physiological and metabolicstress in production animals andlaboratory animals (Misra and Singh,2002). Therefore, the behavior in theintake of liquids when are suppliedexperimental diets, might besubjected to learning experiences ofthe animal used on each essay(Campbell et al., 2009) or due toimmediate responses to regulate theosmotic balance in the intestine as aresult of the food eaten (Mora et al.,2009).
Additionally, the behavior ofanimals regarding the water and foodconsumption was strongly influencedby the breed and category of the ani-mal, the particularities of the digestivesystem and of the excretionmetabolism and, in a great way, bythe level and energy-protein relationof the portion (Ferreira et al., 2002).However, it was also proved that insome cases the water intake was notrelated to the voluntary consumption,the retention of nitrogen, thedigestibility and ingest capacity(Hadjigeorgiou et al., 2000).
From the biochemical point ofview, has been determined that inmany monogastric animals, there is adirect association between the leptinconcentration, hormone produced bythe adipocytes in proportion of its sizewith the water intake, which is

Rev. Fac. Agron. (LUZ). 2012, 29: 248-272
261
experiencias de aprendizaje del ani-mal utilizado en cada ensayo(Campbell et al., 2009) o debido a res-puestas inmediatas para regular elbalance osmótico en el intestino, comoresultado del alimento consumido(Mora et al., 2009).
Adicionalmente, el comportamien-to de los animales en cuanto al consu-mo de agua y de alimento se encuentrainfluenciado de forma notable por laraza y la categoría animal, las particu-laridades del sistema digestivo y delmetabolismo de excreción y, en granmedida, por el nivel y relación energía-proteína de la ración (Ferreira et al.,2002). Sin embargo, también fue de-mostrado que en algunos casos laingesta de agua no se asoció con el con-sumo voluntario, la retención de nitró-geno, la digestibilidad y la capacidadingestiva (Hadjigeorgiou et al., 2000).
Desde el punto de vistabioquímico, se ha determinado que enmuchos animales no rumiantes existeuna asociación directa entre la concen-tración de Leptina, hormona produci-da por los adipositos en proporción asu tamaño, con el consumo de agua, locual está regulado por el sistema ner-vioso (estimulación del centro de sa-ciedad), en respuesta al tipo de dieta yla proporción de los principalesnutrientes que la componen (Grunfeldet al., 1996; Kuo et al., 2005). En estosanimales también se relacionó el con-sumo de agua con la tonicidadplasmática, la osmolaridad, el efectode llenado del sistema digestivo, elestrés calórico, la concentración del iónsodio (Na+) y otros electrolitos presen-tes en el agua y la dieta, así como conla compatibilidad del tipo de alimentoy la bebida ofrecida.
regulated by the nervous system(stimulation of the satiety center) inresponse to the type of diet and theproportion of the main nutrients thatcompose them (Grunfeld et al., 1996;Kuo et al., 2005). In this animals, wasalso related the water intake to theplasmatic tonicity, osmolarity, thefilling effect of the digestive system,caloric stress, concentration of sodiumion (Na+) and other electrolytes presentin the water and the diet, as well asthe compatibility of the food’s type andthe beverage offered.
On the other hand, in adults ratsSprague-Dawley were observedsubstantial variations in the water intakerelated to physiological changes duringdifferent production cycles, in which case,the depression of the food and waterintake was related to neuro-endocrineevents (Tarttelin and Gorski, 1971).
However, in the Syrian hamster,was reported that the water intakewas not affected by the injection ofelectrolytes and other chemicalcompound (Na+ and Cl-; polyethyleneglycol, isoproterenol, serotonin andangiotensin II and III), which mightindicate that the variations in theconsumption of liquid in this speciemight be related to specific componentsof the diets (Neil, 1988). This behaviorwas also informed for rats and degus(Octodon degus), which might indicatethat in these rodents is a direct relationbetween the chemical characteristicsof the food consumed and theacceptance of liquids.
In this sense, the ingest behaviorof liquids in non ruminant animalsalso seems to be related specifically tothe protean content of the diets; sincethe elevated levels of proteins must be

García et al.
262
Por otra parte, en adultos de ra-tas Sprague-Dawley se observó varia-ciones sustanciales en el consumo deagua asociado a cambios fisiológicosdurante varios ciclos de producción; encuyo caso la depresión de la ingesta dealimentos y agua fue asociada con even-tos neuroendocrinos (Tarttelin yGorski, 1971).
Sin embargo, en el hámster Sirio,se reportó que el consumo de agua nofue afectado por la inyección deelectrolitos y otros compuestos quími-cos (Na+ y Cl-; polietilenglicol,isoproterenol, serotonina yangiotensina II y III), lo cual pareceindicar que las variaciones en el con-sumo de líquido en esta especie podríanestar asociados a componentes especí-ficos de las dietas (Neil, 1988). Estecomportamiento también se informópara ratones y Degús (Octodondegus), lo cual podría indicar que enlos roedores existe una relación direc-ta entre las características químicasdel alimento consumido y laaceptabilidad de líquidos.
En este sentido, también el com-portamiento ingestivo de líquidos enalgunos animales no rumiantes pare-ce relacionarse específicamente con elcontenido proteico de las dietas; ya quelos niveles elevados de proteínas de-ben ser catabolizados y excretados, enparte como urea y ácido úrico, víatransformaciones mediadas por el sis-tema renal. Este proceso hace que elanimal requiera mayor volumen deagua y que en algunas especies de la-boratorio, con requerimientosproteicos bajos, un incremento de 1%en el nivel proteico aumente el consu-mo de agua aproximadamente en 3%(De Vries, 1996).
catabolised and excreted, in part asurea and uric acid, via transformationsof the renal system. This processmakes that the animal requires highervolume of water and that somelaboratory species, with low proteanrequirements, an increment of 1% inthe protean level increased the waterintake in approximately 3% (De Vries,1996).
In this experiment, the volumeof the water consumed by hamsterswas more related to the presence of HLin the food portion than with the netprotean content of each diet, since theanimals that ate diets formulated withHL (D1, D2, D3 and D4) exhibited higheraverage intake of water (13.90mL.animal.day-1; PB: 14-22%) thanthose fed with Conejarina® (9.67mL.animal.day-1; PB: 11%) or withRatarina® (8.03 mL.animal.day-1; PB:21%). This behavior can be explainedconsidering the asseverations pointedby García et al., (2008) and Mora etal., (2009), who suggested that whenanimals are submitted to nutritionalregimes to which are not associativehistory at medium or long term, thehigher intake of water in the initialphase occurs due to the adaptationprocess, where the animal familiarizeswith the offer, not only from theorganoleptic point of view; but also, byemetic signs that derive from theinteraction of the animal with the food.
On the other hand, the proteanrequirements and of other nutrimentsfor M.auratus, were studied indifferent researches (Arrington et al.,1966; Arrington et al., 1979), wherewas proved that rodents might consu-me diets with a PB content from 11 to28%, as long as the ingredient do not

Rev. Fac. Agron. (LUZ). 2012, 29: 248-272
263
En este experimento el volumende agua consumida por los hámstersse relacionó más con la presencia deHL en la ración alimenticia que con elcontenido proteico neto de cada dieta;ya que los animales que consumieronlas dietas formuladas con HL (D1, D2,D3 y D4) exhibieron mayor consumopromedio de agua (13,90mL.animal.día-1; PB: 14-22%) que losalimentados con Conejarina® (9,67mL.animal.día-1; PB: 11%) o conRatarina® (8,03 mL.animal.día-1; PB:21%). Este comportamiento se puedeexplicar considerando las aseveracio-nes señaladas por García et al. (2008)y por Mora et al. (2009), quienes sugi-rieron que cuando los animales sonsometidos a regimenes nutricionalescon los cuales no existen antecedentesasociativos a mediano o largo plazo, elconsumo de agua mayor en la etapainicial ocurre debido al procesoadaptativo que tiene lugar, en el cualel animal se familiariza con la oferta,no solamente desde el punto de vistaorganoléptico; sino también por lasseñales eméticas que se derivan dela interacción del animal con el ali-mento.
Por otra parte, los requerimien-tos proteicos, y de otros nutrimentos,para M. auratus fueron estudiados ennumerosas investigaciones (Arringtonet al., 1966; Arrington et al., 1979), enlas cuales se demostró que estos roe-dores pueden consumir dietas con uncontenido de PB entre 11 y 28%; siem-pre que los ingredientes no contengancompuestos que perjudiquen los pro-cesos fisiológicos digestivos (De Vries,1996).
Asimismo, se demostró que cuan-do el alimento se encuentra compues-
have compounds that might harm thephysiological digestive process (DeVries, 1996)
Likewise, was proved that whenthe food is composed by proteins of si-milar nature, the requirements arecovered more effectively, due to thehigher efficiency in the utilization ofnitrogen, different when the hamstersare fed with portions containingproteins of different complexity anddifferent sources (Banta et al., 1975).
The weight lost (1.56g.animal.day-1) experimented by groupD4 might had occurred by different cau-ses. Such diet had the highestproportion of HL, which has beenindicated as an appropriate food sourcefor the animal alimentation due to itshigh content of amino acids (Morán-Fuenmayor et al., 2008; Cova et al.,2009). However, the dehydrated of theworm has toxic celomatic liquid causedby a high bacterial concentration in thedigestive tract of the annelid. On thismatter, has been informed that theseendogenous microorganisms of Eiseniaspp. generate a series of toxicmetabolites when the annelid dies,which affect numerous uni andmulticellular (Satchell et al.,1983); suchcompounds are steady and might keepalive during the dehydration process ofthe material, causing toxicity when theHL content in the diet is considerable.Sin However, the toxic effect mentioneddepends on the type and category of theanimal that consumes the flour, thequantity and the structure of the toxicpresent in the diet, the additive and foodcomponents used, and theparticularities of the digestive systemsof the anima that eat it (García et al.,2005).

García et al.
264
to por proteínas de naturaleza similar,los requerimientos son cubiertos deuna forma más efectiva, debido a lamayor eficiencia en la utilización delnitrógeno, a diferencia de cuando loshámsters son alimentados con racio-nes que contienen proteínas de diferen-te complejidad y de diversas fuentes(Banta et al., 1975).
La pérdida de peso (1,56g.animal.día-1) experimentada por elgrupo D4 puede haber ocurrido por di-ferentes causas. Dicha dieta conteníala mayor proporción de HL, la cual seha indicado como una fuente alimen-ticia apropiada para la alimentaciónanimal debido a su elevado contenidode aminoácidos (Morán-Fuenmayor etal., 2008; Cova et al., 2009). Sin em-bargo, el deshidratado de lombriz con-tiene líquido celomático tóxico, a cau-sa de una elevada carga bacteriana enel tracto digestivo del anélido. Al res-pecto, se ha informado que estosmicroorganismos endógenos de laEisenia spp. generan una serie demetabolitos tóxicos cuando el anélidomuere, los cuales afectan a numero-sos organismos uni y pluricelulares(Satchell et al.,1983); dichos compues-tos son estables y quizás podrían man-tenerse activos durante el proceso dedeshidratación del material, causan-do toxicidad cuando el contenido de HLen la dieta es considerable.
Sin embargo, el efecto tóxico men-cionado depende del tipo y la categoríadel animal que consume la harina, lacantidad y la estructura del tóxico pre-sente en la dieta, los aditivos y compo-nentes alimenticios usados y las par-ticularidades del sistema digestivo delanimal que los ingiere (García et al.,2005).
Likewise, Cova et al., (2009)based on the degradation dynamic ofthe lipid profile of HL, indicated thatduring its obtaining and storingspontaneous degradable processesoccurred, which affected the nutritiousquality of this food compound.Likewise, is known than in the storingof meat flours and derived for animalconsumption, generates a wide varietyof non protein nitrogen substancesnamed biogenic amines (such ashistamine, tyramine,phenylethylamine, putrescine,cadaverine, and tryptamine) which actas antinutritional factors in animalsand human beings (Izquierdo et al.,2006). Even though have not yet beenfound reports of the presence of thesemetabolites in HL, the biogenic aminesmight constitute a cause of theweight’s lost observed with D4 and D3(0.06 g.animal.day-1), and the low gainof hamsters fed with D2 (0.63g.animal.day-1), compare to theobtained with Conejarina®
, of lownutritional value, (0.64 g.animal.day-1),D1 (1.37 g.animal.day-1) and Ratarina®
(1,94 g.animal.day-1).Thus, the consumption of food
with high concentrations ofmonoamines represent an intoxicationrisk since the proteins and the aminoacids of meat flours might sufferbiochemical changes during thedehydration and storing, which makethem in harmful compounds. Thesecan be formed by the enzymatic activityof the dietary tissues of themicroorganisms that contaminatethem by means of enzymes thatdescarboxyle the precursor aminoacids (Izquierdo et al., 2006). Evenwhen is known that not all the

Rev. Fac. Agron. (LUZ). 2012, 29: 248-272
265
Asimismo, Cova et al. (2009) ba-sados en la dinámica de degradacióndel perfil lipídico de la HL, indicaronque durante su obtención y almacena-miento ocurrieron procesosdegradativos espontáneos que afecta-ron la calidad nutritiva de este com-puesto alimenticio. Asimismo, se co-noce que en el almacenamiento de ha-rinas de carnes y derivados para con-sumo animal, se genera una ampliavariedad de substancias nitrogenadasno proteicas denominadas aminasbiogénicas (tales como histamina,tiramina, feniletilamina, putrescina,cadaverina y triptamina) que actúancomo factores antinutricionales enanimales y humanos (Izquierdo et al.,2006). Aunque no se han encontradoreportes de la presencia de estosmetabolitos en la HL, las aminasbiogénicas pudiera constituir una cau-sa de la pérdida de peso observada conD4 y D3 (0,06 g.animal.día-1), y de labaja ganancia de los hámsters alimen-tados con D2 (0,63 g.animal.día-1), com-parado con la obtenida con Conejarina®
,de poco valor nutritivo, (0,64g.animal.día-1), D1 (1,37 g.animal.día-1)y Ratarina®
(1,94 g.animal.día-1).De manera que el consumo de
alimentos con elevadas concentracio-nes de monoaminas representa un ries-go de intoxicación debido a que las pro-teínas y los aminoácidos de las hari-nas de carne pueden sufrir cambiosbioquímicos durante la deshidratacióny almacenamiento, que las conviertenen compuestos nocivos. Éstas puedenformarse por la actividad enzimáticade los tejidos alimenticios o de losmicroorganismos que los contaminan,por medio de enzimas quedescarboxilan los aminoácidos pre-
microorganisms that pollute the foodmay form ammines, those with a highprotean concentration or elevatebacterial concentration, as in HL,might form these compounds.
In this sense, the symptomsobserved in the animals submitted toD4 during the essay are consistent tothe strong intoxication described inanimals submitted to diets withpresence of biogenic ammines (Fusi etal ., 2004). On this matter, hasfrequently been used M. auratus as amodel for studies of applied toxicology,since these animals rapidly developtoxic symptoms, depressing theconsumption of food and losing weightconsistently (Llewellyn et al., 1985;De Vries, 1996; García et al., 2005).
Considering that the productionof HL with supplementary purposes isdone in a craftwork way, the formationof biogens constitutes an importantaspect to study in this type of food toavoid risks at commercial scale. Thiswould make possible establishing via-ble strategies of preservation of the foodsource; for example, the storing at lowtemperature, that has proved mightconstitute an adequate option in somecases (Cova et al., 2009).
The lower weigh gain obtainedwith D0 might has been thatConejarina® had a low protean content(PB: 11-13%) and could not cover therequirements of adult hamsters atmedium term (NRC, 1995). Thehighest requirement was reached withD5 and D1; in the first case becauseRatarina constitute the most usedconventional food for the nourishmentand maintenance of these animals byits high content of PB (21-26%,secondly, because was used the HL

García et al.
266
cursores (Izquierdo et al., 2006). Aúncuando se conoce que no todos losmicroorganismos que contaminan losalimentos pueden formar aminas; losque presentan una elevada concentra-ción proteica o elevada cargabacteriana, como en el caso de la HL,pudieran ser más propensos a la for-mación de estos compuestos.
En este sentido, los síntomas ob-servados en los animales sometidos aD4 durante el ensayo son consistentescon la intoxicación aguda descrita enanimales sometidos a dietas con pre-sencia de aminas biogénicas (Fusi etal., 2004). Al respecto, se ha utilizadofrecuentemente a M. auratus comomodelo para estudios de toxicologíaaplicada, ya que estos animales desa-rrollan cuadros tóxicos rápidamente,deprimiendo el consumo de alimento yperdiendo peso de forma consistente(Llewellyn et al., 1985; De Vries, 1996;García et al., 2005).
Considerando que la producciónde HL con fines de suplementación serealiza de forma artesanal, la forma-ción de biogénos constituye un aspectoimportante a estudiar en este tipo dealimento para evitar riesgos a escalacomercial. Esto haría posible estable-cer estrategias viables de conservaciónde esta fuente alimentaria, como porejemplo el almacenamiento a baja tem-peratura, que se ha demostrado pudie-ra constituir una opción adecuada, enalgunos casos (Cova et al., 2009).
La menor ganancia de peso logra-da con D0 quizás se debió a que laConejarina® tenía un contenido bajo enproteínas (PB: 11-13%), y pudo no cu-brir los requerimientos de los hamstersadultos a mediano plazo (NRC, 1995).Mientras que el mejor comportamien-
proportion (21%) which is apparentlyless harmful, from the point of view ofthe content of biogenic substances andwith a protean content, though infe-rior to D5, appropriate to stimulate agrowth compare to the obtain withRatarina. This suggests that theinclusion of HL in the portion in aproportion of 21% is adequate forfeeding these rodents.
In this sense, the daily gains ofweight obtained with D1 (PB: 13.5%)and D5 (PB: 21.1%) are similar to theones informed by Banta et al., (1975),who after a food elaborated with corn,soy, casein and a content of 13.7% ofPB obtained gains from 1.8 to 2.0g.animal.day-1 in Syrian hamsters ofboth sexes, while, with 20% of PB thegain oscillated from 1.8 to 1.9g.animal.day-1. This emphasizes thatthe components of the diet are veryimportant to improve the corporalcondition of these rodents more thanthe net value of PB. This tendency wasalso described by Birt and Conrad(1981) who incremented from 18 to22% the PB content in the portion,without improving the weight gainsthat oscillated from 1.8-1.9g.animal.day-1.
Nevertheless, the resultsobtained with D1 and D5 were superiorto the reported by Feldman et al.,(1982), who using an essentiallyprotean diet (PB: 24%) achieved agrowth rate of 1.2 g/animal/day inadults, while with other of 18% of PBobtained gains of 2.0 g.animal.day-1 inweaned animals. These results showthat the requirements of theseanimals, in occasions, vary in functionto the physiological phase, the growthphase and sex; and the nature of the

Rev. Fac. Agron. (LUZ). 2012, 29: 248-272
267
to se alcanzó con D5 y D1; en el primercaso porque la Ratarina constituye elalimento convencional utilizado parala alimentación y el mantenimiento deestos animales por su elevado conteni-do de PB (21-26%), y en el segundo casoporque se utilizó la proporción de HL(21%) aparentemente menos nociva,desde el punto de vista del contenidopotencial de substancias biogénicas, ycon un contenido proteico que aunqueinferior a D5, fue apropiado para esti-mular un crecimiento comparable allogrado con la Ratarina. Esto sugiereque la inclusión de HL en la ración enun proporción de 21% es adecuado parala alimentación de estos roedores.
En este sentido, las gananciasdiarias de peso obtenidas con D1 (PB:13,5%) y D5 (PB: 21,1%) son similaresa las informadas por Banta et al.(1975), quienes a partir de un alimen-to elaborado con maíz, soya, caseína ycon un contenido de 13,7% de PB ob-tuvieron ganancias entre 1,8 y 2,0g.animal.día-1 en hamsters Sirios deambos sexos; mientras que con una de20% de PB la ganancia osciló entre 1,8y 1,9 g.animal.día-1. Esto enfatiza quelos componentes de la dieta son muyimportantes para mejorar la condicióncorporal de estos roedores, más que elnivel de PB neto. Esta tendencia tam-bién fue descrita por Birt y Conrad(1981) quienes incrementaron de 18 a22% el contenido de PB en la raciónsin mejorar las ganancias de peso queoscilaron entre 1,8-1,9 g.animal.día-1.
No obstante, los resultados obte-nidos con D1 y D5 fueron superiores alos reportados por Feldman et al.(1982), quien utilizando una dieta esen-cialmente proteica (PB: 24%) logró unatasa de crecimiento de 1,2
protean source is an important varia-ble to consider when formulating nonconventional diets for M. auratus.
On the other hand, Arrligton etal., (1966) with a diet of 18% of PBused to feed six-week adults of M.auratus, achieved a gain of 1.49g.animal.day-1, while with otherportion elaborated with casein or soyand 16% of PB, the weight gain wassimilar (1.40-1.50 g.animal.day-1).This indicates that not only the proteansource, but also the type anddegradation capacity of the protein, isof great importance in the weight gainof these rodents. Likewise with dietsbased on casein and growing contentsof PB (9, 18 and 25%), Horowitz andWaisman (1966) obtained a maximumweight gain of 1.4 g.animal.day-1 onlywith the portion that had 18% of PB.Thus, the results obtained in the ex-perimental conditions of this researchindicate that the requirements of adultfemale hamsters were covered with theproportion of nutriments presented inD1, and such diet constitutes the bestoption to be included in thealimentation of animals with a simi-lar digestive metabolism to M.auratus.
Additionally, is well known thehigh digestibility of HL when is usedas unique diet for ruminants as for nonruminants, aspect that has beenproved employing in vitro and in vivotechniques (García et al., 2009; Covaet al., 2009). However, when this isused as component of more complexfood, the digestibility of this fractionis influenced by the rest of thecomponents included in the portionand by the type of animal used on thetest. The digestibility of D1 (21% lower

García et al.
268
g.animal.día-1 en adultos; mientras quecon otra de 18% de PB logró ganan-cias de 2,0 g.animal.día-1 en animalesdestetados. Estos resultados ponen demanifiesto que los requerimientos deestos animales, en ocasiones, son va-riables en función del estado fisiológi-co, la etapa de crecimiento y el sexo; yque la naturaleza de la fuente proteicaes una variable importante a conside-rar cuando se formulan dietas no con-vencionales para M. auratus.
Por otra parte, Arrligton et al.(1966) con una dieta de 18% de PButilizada para alimentar adultos deM. auratus seis semanas de edad selogró una ganancia de 1,49g.animal.día-1, mientras que con otraración elaborada a base de caseína osoya y un 16% de PB la ganancia depeso fue similar (1,40-1,50g.animal.día-1). Lo cual indica queno sólo la fuente proteica, sino tam-bién el tipo y la capacidad de degra-dación de la proteína, es de vital im-portancia en la ganancia de peso enestos roedores. Asimismo, con die-tas a base de caseína y contenidocrecientes de PB (9, 18 y 25%),Horowitz y Waisman (1966) obtuvie-ron una ganancia de peso máximade 1,4 g.animal.día-1 solamente conla ración que tenía 18% de PB. Demodo que, los resultados obtenidosen las condiciones experimentales deeste estudio indican que los requeri-mientos de las hembras hámsteradultas quedaron cubiertos con laproporción de nutrimentos presentesen D1, y que dicha dieta constituyela mejor opción para ser incluida enla alimentación de animales conmetabolismo digestivo similar al deM. auratus.
proportion of HL) was similar to theobserved for the alone HL (García etal., 2009). While DAMS of D4 (30%higher proportion of HL) was muchinferior to the reported by suchauthors, which show that there is nota lineal behavior of the digestibility ofthe portion to the percentage of HLincluded in the diet, which is probablyrelated to the harmful effects of highproportions of HL in the portion and/or metabolic interactions of compoundsor components of it.
The digestibility of D1 consideredthe best diet formulated with HL, aswell as D5 included as conventionalfood, was similar to the informed byHasdai and Liener (1983) when feedingGold hamsters for 32 days with portionsof raw and treated soy. In suchexperiment, conditioned by the presenceof some toxic compounds in the diets,the digestibility oscillated from 77.1 to84.6% with a consumption of food infe-rior (5.5-7.6 g.animal.day-1) to theregistered in the current research andvariable weight gains (0.33-1.57g.animal.day-1). The inferior productivebehavior of hamsters fed with D3 andD4 in terms of weight and DAMS in thecurrent essay, was similar to theobtained by the last author with dietsthat had an elevated concentration ofanti nutritional metabolites, whichaffected the digestive physiology ofrodents; indicating the harmful effectin the development of M. auratus bypossible harmful compounds presentedin HL.
Conclusions
From all the diets formulatedwith red worm flour for the

Rev. Fac. Agron. (LUZ). 2012, 29: 248-272
269
Adicionalmente, es muy conoci-da la elevada digestibilidad de la HLcuando es utilizada como dieta únicatanto para rumiantes como para norumiantes, aspecto que se ha com-probado empleando técnicas in vitroe in vivo (García et al., 2009; Covaet al., 2009). Sin embargo, cuando esusada como componente de alimen-tos más complejos, la digestibilidadde esta fracción se encuentrainfluenciada por el resto de los com-ponentes incluidos en la ración y porel tipo de animal que se utiliza en laprueba. La digestibilidad de D1 (21%; menor proporción de HL) fue si-milar a la observada para la HL sola(por García et al., 2009). Mientrasque la DAMS de D4 (30%; mayor pro-porción de HL) fue muy inferior a lareportada por dichos autores, lo cualdemuestra que no existe un compor-tamiento lineal de la digestibilidad dela ración con el porcentaje de de HLincluida en la dieta; lo cual está pro-bablemente relacionado con los efec-tos perjudiciales de proporciones ele-vadas de HL en la ración y/o coninteracciones metabólicas de com-puestos o componentes de la misma.
La digestibilidad de D1, considera-da la mejor dieta formulada con HL, asícomo de D5, incluido como alimento con-vencional, fue similar a la informada porHasdai y Liener (1983) al alimentarhamsters dorados durante 32 días conraciones a base de soya cruda y tratada.En dicho experimento, condicionado porla presencia de algunos compuestos tóxi-cos en las dietas, la digestibilidad oscilóentre 77,1 y 84,6%, con un consumo dealimento inferior (5,5-7,6 g.animal.día-1)al registrado en este estudio y gananciasde peso variables (0,33-1,57
alimentation of M. auratus, the onethat included 21% of this componentin the portion was the best in terms ofweight’s dynamic and digestibility ofthe food. Likewise, the behavior ofanimals assigned to this diet was si-milar to those fed with a conventionaldiet (Ratarina) with a superior contentof PB, which emphasizes that theinclusion of HL in these proportionswas adequate as supplementaryprotean source for the growing of theseanimals.
In the diets where HL wasincluded in superior proportions (24,27 and 30%) the performance of thegold hamster was affected, maybebecause of the presence of compoundsthat were naturally in HL or by thebacterial degradation of somenutritional components of it, whichmight reduce the nutritional qualityof HL or generate toxic substances. Itis recommended to carry qualitativeand quantitative determinations ofbiogenic ammines in the annelid flour,with the aim of proving or not thistheory.
Acknowledgement
The authors want to thank theCouncil for Scientific, Humanistic,Technological and Arts Development(CDCHTA) of Universidad de los An-des, by financing this research(NURR-C-355-04-03-A). The Experi-mental station and AgricultureProduction “Rafael Rangel” by thesupport received for obtain the wormflour used in this research. TheNational Center laboratory ofAgriculture Researches (CENIAP-Maracay), by the collaboration in some

García et al.
270
g.animal.día-1). El comportamiento pro-ductivo inferior de los hámsters alimen-tados con D3 y D4 en términos de peso yDAMS en el presente ensayo, fue similaral obtenido por estos ultimo autores condietas que tuvieron una elevada concen-tración de metabolitos antinutricionalesque afectaron la fisiología digestiva de losroedores; indicando el efecto perjudicialen el desempeño del M. auratus por posi-bles compuestos nocivos presentes en laHL.
Conclusiones
De las dietas formuladas con ha-rina de lombriz para la alimentacióndel M. auratus, la que incluyo un 21%de este componente en la ración fue elmejor en términos de dinámica de pesoy digestibilidad del alimento. Asimis-mo, el comportamiento de los anima-les asignados a esta dieta fue similara los que se alimentaron convencional-mente (Ratarina®) con un contenido su-perior de PB, lo cual enfatiza que lainclusión de HL en estas proporcionesfue adecuada como fuente proteica su-plementaria para el crecimiento deestos animales.
En las dietas en las que la HL fueincluida en proporciones superiores (24,27 y 30%) el desempeño del hámsterdorado se vio afectado, quizás por la pre-sencia de compuestos que se encontra-ban de forma natural en la HL o por ladegradación bacteriana de algunos com-ponentes nutricionales de la misma, locual pudo reducir la calidad nutricionalde la HL o generar substancias toxicas.Se recomienda realizar determinacionescualitativas y cuantitativas de aminasbiogénicas en la harina del anélido, conel fin de comprobar o no esta teoría.
analytical determinations. TheVivarium of the University campus“Rafael Rangel” in Trujillo state, Ve-nezuela, especially Mr. JavierMoncayo, by his collaboration in themaintenance of the animals during theexperimental period.
End of english version
Agradecimientos
Los autores desean expresar suagradecimiento al Consejo de Desarro-llo Científico, Humanístico, Tecnológicoy de las Artes (CDCHTA) de la Uni-versidad de los Andes por elfinanciamiento de esta investigación(NURR-C-355-04-03-A). A la EstaciónExperimental y de Producción Agríco-la “Rafael Rangel” por el apoyo recibi-do para la obtención de la harina delombriz utilizada en el ensayo. Al la-boratorio del Centro Nacional de In-vestigaciones Agropecuarias (CENIAP-Maracay) por su colaboración en algu-nas de las determinaciones analíticas.Al Vivario del Núcleo Universitario“Rafael Rangel” del estado Trujillo,Venezuela, en la persona del Sr. Ja-vier Moncayo, por su colaboración enel mantenimiento de los animales du-rante el periodo experimental.
Literatura citada
AOAC. 1990. Official methods of analysis.15th ed. Association of Official. 500 p.
Arrington L.R., J.K. Platt y R.L. Shirley.1966. Protein requirements ofgrowing hamster. Lab. Anim. Sci. 16:492-496.

Rev. Fac. Agron. (LUZ). 2012, 29: 248-272
271
Arrington L.R., C.B. Anunerman y D.E.Franke. 1979. Protein requirementsof hamster fed a natural diet. Lab.Anim. Sci. 29: 469-471.
Banta C.A., R.G. Warner and J.B. Robertson.1975. Protein nutrition of the goldenhamster. J. Nutr. 150: 38-45.
Campbell M.A., M.S. Golub, P. Iyer, F.L.Kaufman, L. Ling-Hong, F. Moran-Messen, J.E. Morgan y J.M. Donald.2009. Review Article: Reduced waterintake: Implications for rodentdevelopmental and reproductivetoxicity studies. Birth DefectsResearch Part B: Developmental andReproductive Toxicology 86(3): 157-175.
Carr T.P., D.D. Gallaher, C.H. Yang y C. A.Hassel. 1996. Increased intestinalcontents viscosity reduces cholesterolabsorption efficiency in hamsters fedhydroxypropyl methylcellulose1’2’3.J. Nutr. 126: 1463-1469.
Cova L.J. y D.E. García. 2006. Evaluación deparámetros nutricionales utilizandoharina de lombriz (Eisenia spp.) enla alimentación del hamster(Mesocricetus auratus L.). JornadaTécnica, Estación Experimental y deProducción Agrícola “Rafael Rangel”(EEPARR)-Instituto Nacional deInvestigaciones Agrícolas (INIA),Trujillo, Venezuela 12 p.
Cova L.J., D.E. García, J.V. Scorza D., M.G.Medina, T. Clavero, F. Perea y D.González. 2009. Efecto de laestrategia de conservación en lacalidad nutritiva de la harina de lalombriz roja (Eisenia spp.) a medianoplazo. Rev. Fac. Agron. (LUZ) 26(1):107-128.
De Jesús R. y Z. Quintero. 2001. Influenciade diferentes niveles de intensidadde la luz sobre algunos parámetrosreproductivos en ratones NMRI. Rev.Cient. FCV. LUZ IX: 403-407.
De Jesús R. y Z. Quintero. 2008.Comparación del crecimiento,parámetros reproductivos y dehematología y glicemia en hámsters(Mesocricetus auratus) alojados adiferentes niveles de intensidad deluz. Zootecnia Trop. 26(1): 19-26.
De Vries. J. 1996. Food safety and toxicity.De Vries. J. (Ed) 338 p.
Feldman D.B., E.E. Mc Connell and J.J.Knapka. 1982. Growth kidneydisease and longevity of Syrianhamsters (Mesocricetus auratus) fedvarying levels of proteins. Lab. Anim.Sci. 32: 613-618.
Ferreira, V., L.C. Hoffman, S.J. Schoemanand R. Sheridan. 2002. Water intakeof Boer goats and Mutton merinosreceiving either a low or high energyfeedlot diet. Small Rum. Res. 43(3):245-248.
Fusi E., L. Rossi, R. Rebucci, F. Cheli, A. DiGiancamillo, C. Domeneghini, L.Pinotti, V. Dell’Orto and A. Baldi.2004. Administration of biogenicamines to Saanen kids: effects ongrowth performance, meat qualityand gut histology. Small Rum. Res.53(1): 1-7.
García D.E., L.J. Cova, A.R. Castro, M.G.Medina y J.R. Palma. 2009. Efecto delsustrato alimenticio en lacomposición química y el valornutritivo de la harina de la lombrizroja (Eisenia spp.). Revista CientíficaFCV-LUZ XIX(1): 55-62.
García D.E., M.G. Medina, L.J. Cova, J.Humbría, A. Torres y P. Moratinos.2008. Preferencia caprina porespecies forrajeras con ampliadistribución en el estado Trujillo,Venezuela. Arch. Zootecnia, 57(220):403-413.
García D.E., M.G. Medina, M. Soca y I.L.Montejo. 2005. Toxicidad de lasleguminosas forrajeras en laalimentación de animalesmonogástricos. Pastos y Forrajes 28:279-289.
Grunfeld C., C. Zhao, J. Fuller, A. Pollock,A. Moser y J. Friedman. 1996.Endotoxin and cytokines induceexpression of leptin, the ob geneproduct, in hamsters. Clin. Invest.97: 2152-2157.
Hadjigeorgiou I., K. Dardamani, C. Goulas yG. Zervas. 2000. The effect of wateravailability on feed intake anddigestion in sheep. Small Rum. Res.37(2): 147-150.

García et al.
272
Hasdai A. y I.E. Liener. 1983. Growth,digestibility and enzymatic activitiesin the pancreas and intestines ofhamsters fed raw and heated soyflour. J. Nutr. 113(3): 662-668.
Home Office. 1989. Code of practice for thehousing and care of animals used inscientific procedure. 2da ed. HM&O,Londres.
Horowitz I. y H.A. Waisman. 1966. Somebiochemical changes in the hamsterfed excess phenyl alanine diets. Proc.Soc. Exp. Biol. Med. 132: 750-755.
Izquierdo P., M. Allara, A. García, G. Torres,E. Rojas y M.Y. Piñero. 2006. Aminasbiógenas y bacterias en salchichóntipo milano: Efecto del tiempo dealmacenamiento. Rev. Cient. FCV-LUZ XVI(2): 186-194.
Kuo A., M.A. Cline, E. Werner, P.B. Siegel yD.M. Denbow. 2005. Leptin effects onfood and water intake in lines ofchickens selected for high or low bodyweight. Physiology & Behavior 84(3):459-464.
Llewellyn G.C., E.A. Floyd, G.D. Hoke,L.B. Weekley y T.D. Kimbrough.1985. Influence of dietary aflatoxin,zinc, and copper on bone size, organweight, and body weight in hamstersand rats. Bull. Environ. Contam.Toxicol. 35(1): 149-156.
Misra, A.K. y K. Singh. 2002. Effect of waterdeprivation on dry matter intake,nutrient utilization and metabolicwater production in goats undersemi-arid zone of India. Small Rum.Res. 46(3): 159-165.
Mora N., M. Casas, A. Marín, R.N. Águila, I.Sánchez, H. Hernández y L.Sanginés. 2009. The kelp Macrocystispyrifera as nutricional supplementfor goats. Revista Científica FCV-LUZ, XIX(1): 63-70.
Morón-Fuenmayor O.E., D. Diaz, S.Pietrosemoli, R. Barrera, N. Gallardo,J. Peña y M. Leal. 2008. Efecto de lainclusión de harina de lombriz sobre
el rendimiento en canal, en cortes ycalidad físico-química de la carne decodorniz (Coturnix coturnix japonica).Rev. Fac. Agron. (LUZ) 25: 674-685.
Neil, E.R. 1988. Water intake of Djungarianand Syrian hamsters treated withvarious dipsogenic stimuli. Physiology& Behavior 43(6): 851-854.
NRC. 1995. Nutrients Requirements ofLaboratory Animal. Hamster. Fourthrevised edition. National AcademyPress. 138 p.
Rees N. and D. Watson. 2000. Internationalstandards for food safety. AspenPublishers, Inc. 256 p.
Satchell J.E. 1983. Earthworm ecology fromDarwing to vermiculture. Chapman& Hall. Nueva York, EEUU. 485 pp.
Savón L., L.M. Mora, L.E. Dihigo, V.Rodriguez, Y. Rodriguez, I. Scull; Y.Hernández y T.E. Ruíz. 2008. EfectoDe la harina de follaje de Tithoniadiversifolia en la morfometría deltracto gastrointestinal de cerdos encrecimiento-ceba. Zootecnia Trop.,26(3): 387-390.
Tarttelin M.F. and R.A. Gorski. 1971.Variations in food and water intakein the normal and acyclic female rat.Physiology & Behavior 7(6): 847-852.
Van Soest, P.J., J. Robertson y B. Lewis.1991. Symposium: Carbohydrate,methodology, metabolism andnutritional implications in dairy cattle,Methods for dietary fiber, neutraldetergent fiber and nonstarchpolysaccharides in relation to animalnutrition. J. Dairy Sci., 74: 3583-3597.
Visauta B. 1998. Análisis Estadístico con SPSSpara Windows. En Visauta B (Ed)Estadística Multivariante. Mc-Graw-Hill-Interamericana. Madrid, España.200 pp.
Copyright © 2022 FDOKUMEN




















