
Quantitative Image Analysis of HIV-1 Infection in Lymphoid Tissue

Journal of Photochemistry and Photobiology B: Biology 142 (2015) 141–153
Contents lists available at ScienceDirect
Journal of Photochemistry and Photobiology B: Biology
journal homepage: www.elsevier .com/locate / jphotobiol
Photoperiodic regulation of nuclear melatonin receptor RORa inlymphoid organs of a tropical rodent Funambulus pennanti: Role inseasonal oxidative stress
http://dx.doi.org/10.1016/j.jphotobiol.2014.11.0131011-1344/� 2014 Elsevier B.V. All rights reserved.
⇑ Corresponding author. Tel.: +91 542 2307149x209, +91 542 6702535, mobile:+91 9415222261; fax: +91 542 2368174, +91 542 2575093.
E-mail addresses: [email protected] (C. Haldar), [email protected](R. Ahmad).
1 Present address.
Sameer Gupta a, Chandana Haldar a,⇑, Raise Ahmad b
a Pineal Research Laboratory, Department of Zoology, Banaras Hindu University, Varanasi 221 005, Indiab Department of Endocrinology, Metabolism and Diabetes (EMD), Institut Cochin, INSERM U1016, CNRS 8104, Université Paris Descartes, 22 rue Méchain, 75014 Paris, France1
a r t i c l e i n f o a b s t r a c t
Article history:Received 10 July 2014Received in revised form 16 October 2014Accepted 3 November 2014Available online 11 December 2014
The orphan nuclear receptor RORa of RZR/ROR family has been suggested to mediate the genomic actionsof melatonin on the expression of antioxidant enzymes. However, no direct evidences exist. In the pres-ent study we explored the role of photoperiod (natural and artificial) in regulation of RORa and its asso-ciation with the photoperiod induced antioxidant defense system in the lymphoid organs (spleen andthymus) of seasonally breeding, tropical squirrels, Funambulus pennanti. The photoperiod mediated reg-ulation of antioxidant status was checked along with the RORa expression and circulatory melatoninlevel in the squirrels. The enhancement of the antioxidant capacity of serum and lymphoid organ wasconcomitant with the short photoperiod (10L:14D) induced high levels of plasma melatonin. Further,peripheral melatonin level enhanced the AANAT activity as well as the melatonin synthesis in the lym-phoid tissues. RORa expression presented an inverse correlation with the plasma level of melatonin aswell as the short day induced antioxidant enzyme activity in the lymphoid organs. The results suggestthat for reduction of seasonal oxidative stress melatonin might not be utilizing the nuclear receptor RORapathway; rather the rise in circulatory melatonin collectively with tissue specific melatonin might beprotecting the splenic and thymic lymphocytes from the seasonal oxidative stress.
� 2014 Elsevier B.V. All rights reserved.
1. Introduction
Substantial research has demonstrated that changes in daylength not only influence reproduction, but also the immunity[1,2]. In recent years, interaction of melatonin with the immunecomponents of the body has gained much attention which hasestablished that changes in melatonin level acts as a critical mod-ulator of hematopoiesis and immune cell functions [3,4]. Melato-nin influences the immunocompetent cells by regulating thecytokine production and thereby its activity and functions [5].Low molecular weight, lipophilic nature and ubiquitous presenceof melatonin raise the possibility for its intracellular and genomicactions. The reports of Becker-Andre et al. [6] and Weisenberg et al.[7] hypothesized that melatonin specifically binds and activates aspecific orphan nuclear receptor belonging to the RZR/RORsubfamily. Due to the influence of melatonin on these nuclear
transcription factors, members of this subfamily are consideredas nuclear melatonin receptor. Among them, RORa is known topotentially mediate the genomic actions of melatonin [7–10].Binding of melatonin to the purified nuclear fractions of spleenand thymus cells further strengthened the idea for existence of afunctional nuclear receptor for melatonin [11], hence, a direct generegulatory effect of melatonin has been proposed. The transcriptionfactor RORa, whose binding sites have been reported to exist in thepromoter region of a number of genes appear to mediatethese genomic actions of melatonin [12–14]. However, no directevidences are available.
By their virtue to respond to the changing ambient daylength/photoperiod, the seasonal breeders experiences the fluctuationsin internal physiological and hormonal milieu and are thus moreprone to seasonally occurring opportunistic diseases and stressors.Lymphoid organs in photo-responsive, seasonally breeding mam-mals are also known to exhibit seasonal variations reflected inactivity and functions of immunocompetent tissues [1,2]. Stressfulenvironment can contribute to increase the free radical loadthroughout the body resulting in altered immune responses.Seasonality thus, is an aspect where the survival and well-beingof the animal depends largely upon the management of stress

142 S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 142 (2015) 141–153
and free radical load generated due to changing environmentalconditions. Due to its free radical scavenging attributes and abilityto induce antioxidant enzyme activity, melatonin has been recog-nized as an endogeneous protective agent [15]. There are reportswhich suggest increased mRNA expression and activity of antioxi-dant enzymes like superoxide dismutase, catalase and glutathioneperoxidase [16,17] following melatonin administration.
However, the role of photoperiod in regulation of genomicactions of melatonin via modulation of its nuclear binding sites/receptors in lymphoid tissues has never been studied in any sea-sonal breeder. In the present study we hypothesized that photope-riod, which is responsible for modulating circulatory level ofmelatonin in Funambulus pennanti can also influence the expres-sion of its nuclear receptor RORa, thereby affecting transactivationof its molecular targets particularly expression/activity of anti-oxi-dant enzymes as a function of photoperiod.
Thus, we explored the role of photoperiod (seasonal and artifi-cial) in regulation of the expression of RORa and its correlationwith the photoperiod/melatonin induced antioxidant defense sys-tem (antioxidant enzymes) in the lymphoid organs (spleen andthymus) of a tropical, seasonally breeding rodent F. pennanti. Theexpression of RORa was assessed at protein and mRNA level byimmunoblot and semiquantitative RT-PCR analyses. To decipherthe effect of plasma melatonin on the tissue specific melatonin bio-synthesis and thereby on the modulation of photoperiod depen-dent antioxidant capacity, we also checked the AANAT activityand intra lymphoidal melatonin content. Total antioxidant status(TAS) of lymphoid tissues as well as blood plasma was further cor-related with circulatory melatonin level under different photoperi-odic conditions to elaborate the significance of photoperiod inregulation of RORa and seasonal/photoperiodic oxidative load.
2. Materials and methods
All the experiments were conducted in accordance with Institu-tional practice and with in the framework of experimental animals(Scientific Procedure) Act 2007, of the Committee for the Purposeof Control and Supervision of Experiments on Animals (CPCSEA),Government of India, on animal welfare.
2.1. Animals and maintenance
The Indian palm squirrel, F. pennanti is a seasonal (long-day)breeder of tropical origin. It is a diurnal rodent and semi-domesticin nature. Details of its breeding pattern, habit and habitat hasbeen published elsewhere [19]. Adult squirrels of average weight100 ± 10 g were collected from the vicinity of Varanasi, India (lati-tude 25�180 N; longitude 83�10 E) and were maintained in an open-air wire net enclosures, fully exposed to ambient conditions oftemperature, daylength and humidity. Squirrels were fed withsoaked gram seeds (Cicer arientium), nuts seasonal fruits/vegeta-bles and water ad libitum.
2.2. Experimental plan
2.2.1. Study under ambient photoperiodic conditionsThe Indian palm squirrels F. pennanti are vegetable crop pests
and can be easily maintained in captivity. Long day length of sum-mer marks the onset of reproductive activity in this species and ismentioned as reproductively active phase (RAP) while short-day-length signals cessation of reproductive activity and is describedas reproductively inactive phase (RIP) in the text further. The malesquirrels were collected from our own colony during summer(RAP; May: temp max. 47 �C and min. 30 �C, Humidity-65.39%and day length-13.42 h) and winter (RIP; Dec: temp max. 15 �C
and min 2 �C, Humidity-82.14% and day length-10.35 h). After aweek of acclimatization in the experimental wire net cages(25’’ � 25’’ � 30’’), the animals were sacrificed during the darkphase of the light–dark cycle (night time; 22:00 h IST) under deepether anesthesia by decapitation in dim red light.
2.2.2. Photoperiodic exposureThe adult male squirrels were exposed to experimental photo-
periodic regimens in the last week of August. During this time ofthe year ambient photoperiod is �12L:12D in the Indian tropicalzone. Following 2 weeks of acclimatization, the squirrels were ran-domly assigned into three groups. Group I consisted of five adultmale squirrels (body weight 100 ± 10 g range) that were exposedto a light condition equivalent to natural daylength (i.e. 12L:12D;lights-on at 08:00 h and lights-off at 20:00 h) and were treatedas controls. Group II comprising of five male squirrels was exposedto short-day length conditions (10L:14D; lights-on at 10:00 h andlights-off at 20:00 h). Group III consisted of five male squirrelsexposed to long-day photoperiodic conditions (16L:08D; lights-on at 04:00 h and lights-off at 20:00 h). Squirrels were maintainedin wire net cages in the respective photoperiodic conditions for10 weeks. At the end of each experiment, the animals wereweighed, deeply anesthetized by ether anesthesia and were sacri-ficed by decapitation. The squirrels were sacrificed at 22:00 h IST;when both the photoperiodic Groups II and III along with controlsquirrels experienced the dark phase.
2.2.3. In vitro experimentTo study the role of melatonin in regulation of RORa expression,
an in vitro melatonin supplementation experiment was performed.Seven squirrels (n = 7) exposed to 12L:12D photoperiodic regimenswere sacrificed at 22:00 h. The lymphoid tissues (spleen and thy-mus) were aseptically dissected out in chilled phosphate bufferedsaline (PBS) and processed for cell culture followed by receptorexpression studies as described in the following sections.
2.3. Sample collection and tissue processing
Following decapitation, trunk blood was collected in a heparin-ized tube. Plasma was separated by centrifugation at 3000 rpm for20 min at 4 �C and stored at �20 �C until radioimmunoassay (RIA)of melatonin and TAS assay. The spleen and thymus were dissectedout in chilled PBS and were stored at �80 �C for enzymatic estima-tions and Western blot analyses. For in vitro study, the lymphoidorgans were aseptically removed and processed for culture asdescribed in the following sections. For semi-quantitative RT-PCR, spleen and thymus were collected in RNA later (R0910,Sigma–Aldrich, St. Louis, Missouri, USA). A part of the lymphoid tis-sues were also fixed in neutral formalin for immunohistochemicallocalization of RORa.
2.4. Melatonin assays
The plasma level of melatonin was estimated by radioimmuno-assay (RIA) performed following the method of Rollag andNiswender [20] using Guildhey antisera (Guildhey, Surrey, UK).Details of the method and validation have been publishedelsewhere [21]. The intra- and inter-assay variation for melatoninwas 9% and 15% respectively. The sensitivity for melatonin RIAwas 18–20 pg/ml. The recovery of melatonin RIA was 92%.
For estimation of melatonin content in the lymphoid tissuescommercial melatonin ELISA kit (RE54021, IBL, Hamburg,Germany) was used as per manufacturer’s instruction. The spleenand thymus were homogenated in PBS with 0.1% ethanol andcentrifuged. The supernatants were used for determinations ofmelatonin. Standards, controls and tissue homogenates were

S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 142 (2015) 141–153 143
extracted using C18 reverses phase extraction columns accordingto the protocol of the manufacturer. The reaction was developedusing p-nitrophenyl phosphate and optical densities were deter-mined at 405 nm in an automatic microplate reader (Biotek,USA). The concentration of melatonin was expressed as pg/mg ofprotein measured in the tissue sample. The sensitivity of the mel-atonin assay was 1.6 pg/ml. Both the intra- and inter-assay coeffi-cients of variation (CV) were 11.4% and 19.3% respectively.
2.5. Arylalkylamine N-acetyltransferase (AANAT) assay
Arylalkylamine N-acetyltransferase (AANAT; EC 2.3.1.87) is thepenultimate enzyme of melatonin biosynthetic pathway. AANATactivity was measured following the method of Chae et al. [22]with modifications as published elsewhere [23]. Briefly, the spleenand thymus were disrupted using a cell sonicator in 100 ll ice-cold50 mM phosphate buffer (16 mM Na2HPO4, 40 mM NaH2PO4, pH6.8). Aliquots of 10 ll of the sonicate were added to reactionmixtures containing 1.5 mM tryptamine-HCl, 0.1 mM acetyl coen-zyme A and 0.01 mM of 3[H]-acetyl coenzyme A (0.2 lCi/assay)and the total volume was adjusted to 20 ll with phosphate buffer.The reaction mixture was then incubated at 37 �C for 20 min andthe reaction was stopped by dilution with 180 ll ice-coldphosphate buffer. 3 ml of scintillation fluid (containing 0.4% PPO,0.01% POPOP mixed with toluene) was rapidly added to the reac-tion mixture and the amount of radio labeled acetyltryptaminewas determined using a liquid scintillation counter (BeckmanInstruments). NAT activity was expressed as nmol N-acetyltrypta-mine produced/h/mg protein. Protein content was measuredfollowing the Bradford protocol [24].
2.6. Total antioxidant status (TAS)
The scavenging activity or capacity of antioxidants for ABTSradical cations was measured based on the method of Re et al.[25] as published elsewhere [26]. A stock solution of ABTS radicalcations was prepared 1 day before the assay, by mixing 5 ml of7 mM ABTS with 1 ml of 14.7 mM potassium persulfate and wasstored in dark at room temperature. The stock solution of ABTSwas diluted with water. ABTS radical cation was generated by oxi-dation of ABTS with potassium persulfate. ABTS solution of volume2.95 ml was mixed with 50 ll of 10% homogenate of spleen, thy-mus and blood plasma. The decrease in absorbance was monitoredfor 10 min at 734 nm.
2.7. Immunohistochemical localization of RORa in spleen and thymusof F. pennanti
Immunohistochemical localization of nuclear melatonin recep-tor RORa was performed according to method of Ahmad and Hal-dar [27]. The spleen and thymus were dissected out and fixed inneutral formalin. After dehydration, paraffin blocks were preparedand 7 lm-thick sections were cut and mounted on gelatin (1%)coated slides and deparaffinized. Endogenous peroxide activitywas blocked by 0.3% H2O2 in methanol after which the slides werepreincubated with horse blocking serum (1:100 in PBS; PK-6200,Vector Laboratories, Burlingame, CA) for 2 h. Sections were incu-bated with primary antibody (RORa, C-16, sc-6062; dilution1:250, Santa Cruz Biotech) overnight at 4 �C. Sections were washedand then incubated with biotinylated secondary antibody (Vecta-stain ABC Universal kit; PK 6200, dilution 1:50, Vector Laborato-ries). Again after washing with, a pre-formed ABC reagent wasadded conjugated to the free biotin of the secondary antibody.The antigens were visualized using the 0.03% peroxidase substrate3, 3-diaminobenzidine (DAB, Sigma Chemicals, St. Louis, Missouri,USA) in 0.01 M Tris–Cl (pH 7.6) and 0.1% H2O2. Finally, the sections
were sequentially dehydrated to mounting and were observed andphotographed under Leitz-MPV-3 microscope and documented.
2.8. In vitro study: effect of melatonin supplementation on RORaexpression in splenocytes and thymocytes
The splenocytes and thymocytes were cultured following themethod of Ahmad et al. [28]. Briefly, the spleen and thymus wereremoved aseptically and cells were separated by compressing thetissue between sterile frosted glass slides. The separated cells weresuspended in ice cold culture medium (RPMI-1640 plus Hepessupplemented with 1% penicillin (100 IU/ml) streptomycin(100 lg/ml), 1% L-glutamine (2 mM/ml), 0.1% 2-mercaptoethanol(5 � 10�2 M/ml), and 10% heat-inactivated fetal calf serum). Theresulting single cell suspension was washed thrice with PBS(0.1 M) and cell counts were adjusted to 1 � 107 cells/ml followingviability check (number of viable cells exceeding 95%) using trypanblue exclusion method. The 100 ll of cell suspension and 100 ll ofmelatonin (500 pg/ml) [18,28] were added to each well of 24-wellculture plate, and the final volume was adjusted to 1 ml byRPMI-1640. The control cells were incubated without melatoninsupplementation. After incubation at 37 �C in 5% CO2 humidifiedatmosphere for 48 h the cells were harvested and centrifuged at2500 rpm to collect cell pellet. Cell pellets were washed inphosphate buffer-saline (0.1 M; pH 7.4) and homogenate wasprepared with lysis buffer (1% NP40 (W/V), 0.1% SDS, 0.1%Aprotinin, 20 ll of 50 mM sodium orthovanadate, 1% PMSF inPBS) and mixed well, centrifuged at 12,000g for 10 min at 4 �C.The protein content of the supernatant was determined usingBradford method [24]. 40 lg of protein was resolved on 10%SDS–PAGE for RORa with prestained marker (Fermentas, MD,USA). Rest of the methodology and immunodetection was carriedout in a similar way as explained in Western blot analysis section.
2.9. Reverse transcriptase-PCR of nuclear melatonin receptor RORa
2.9.1. Total RNA isolationExpression of the RORa gene was studied by semi-quantitative
RT-PCR using C1000 thermal cycler (BioRad, USA), as publishedelsewhere [29]. Total RNA from the tissues and cultured cells wereisolated using TRI-reagent (Sigma–Aldrich, St. Louis, Missouri, USA)in accordance with the manufacturer’s protocol. The RNA wasquantified by spectrophotometry. The integrity of the RNA sampleswas verified by 1% formaldehyde agarose gel electrophoresis con-taining 20 mM MOPS, 8 mM Na-acetate, 1 mM EDTA and 2.2 Mformaldehyde.
2.9.2. Reverse transcriptionFor reverse transcription reaction 4 lg of RNA was used in total
30 lL reaction mixture containing 5X MMuLV buffer, 10 mM dNTPmix, 0.5 lg random hexamer primer, 40 U of RNasin ribonucleaseinhibitor and incubated at 25 �C for 10 min. 1 ll of MMuLV reversetranscriptase (Merck, 200 units) was added and the tube was thenincubated at 42 �C for 1 h. The reaction was terminated by heatingat 70 �C for 10 min, and after chilling on ice the tube was stored at�70 �C or directly used for the PCR reaction.
2.9.3. Polymerase chain reactionAs the DNA sequence for RORa in squirrel has not yet been
determined, those used by Carrillo-Vico et al. [9] for Swissmice were used. The oligonucleotide primers used for RORaPCR were 50-GGAAGAGCTCCAGCAGATAAC G30 (forward) and 50-GCTGACATCAGTACGAATGCAG-30 (reverse) which were reportedto amplify a single 417-bp band [9,30]. b-actin primers (forward50ATCGTGGGC CGCTCTAGGCACC-30, reverse 50 CTCTTTGATGT-CACGCACGATTTC-30) used were of mice [31] and amplified

144 S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 142 (2015) 141–153
543 bp. All the primers were synthesized by Metabion Interna-tional, Germany. The final PCR reaction was performed in 25 lLof reaction mixture containing 10X Taq buffer, 10 pM of each ofthe primers, 10 mM dNTPs mix using 2.5 U of Taq-DNA polymerasecontaining 5 lL of RT product. The samples were denatured at94 �C for 5 min and were further amplified using the followingspecifications: (RORa, 35 cycles were performed 94 �C, 1 min;60 �C, 1 min; 72 �C, 1 min) and (b-actin, 26 cycles were performed,94 �C, 30 s; 57 �C, 1 30 s; 72 �C, 30 s) with a final extension of 5 minat 72 �C. The size of the amplification product on gel was assessedby comparison with 100 bp DNA ladder (StepUp™ 100 bp DNA lad-der, MBD 13 J, Merck-Millipore, Bangalore, India). Further, theidentity the amplification product of the RORa primer wassequenced the with AB I3730 genetic analyzer (Merck-Millipore,Merck Specialties Pvt. Ltd., Bangalore, India) and submitted it toGene bank NCBI (accession no. JX227944). The sequence showed88% identity with Rattus norvegicus and Mus musculus nuclear mel-atonin receptor RORa (Blastn, NCBI, USA).
All PCR experiments included negative controls, in which tem-plate cDNA was omitted. The PCR product was electrophoresedon a 2% agarose gel containing ethidium bromide. The bands werevisualized in a UV-transilluminator and photographed (Gel Doc.Alpha Innotec., USA). The relative amount of the RORa wasexpressed as optical density relative to that of the b-actin (GelDoc. Alpha Innotec., USA).
2.10. Western blot analysis of melatonin nuclear receptor RORa
The Western blot analysis for nuclear melatonin receptor RORawas carried by following the method of Ahmad and Haldar [27].Briefly the lymphoid tissue (spleen and thymus) as well as the cellsharvested from the in vitro experiment were homogenized andlysed in lysis buffer (RIPA buffer) and quantified by the Bradfordmethod [24]. Aliquots containing 70 lg protein (40 lg protein forcells) were resolved with 10% SDS–polyacrylamide gel electropho-resis and followed by electrotransfer (Trans-blot Turbo, Bio-Rad,USA) to nitrocellulose membrane (Bioscience, Keene, New Hamp-shire, USA). A pre-stained multicolor broad range marker (Spec-tra™ multicolor broad range marker, 10–260 kDa, SM-1841;Fermentas, Glen Burnie, Maryland, USA) was run along with sam-ple proteins to confirm the position of the band. The nitrocellulosemembrane was blocked for 60 min with Tris-buffered saline con-taining 5% fat-free dry milk, and incubated with nuclear melatoninreceptor antibodies (RORa, C-16, sc 6062; in 1:200 dilution, SantaCruz Biotech). Immunodetection was performed with horseradishperoxidase conjugated secondary antibody (donkey anti-goatIgG-HRP diluted 1:1000). Finally, the blot was washed and devel-oped with Super Signal West Pico Chemiluminescent Substrate(Cat no. 34,080, Thermo Scientific, Rockford, Maryland, USA). Sim-ilarly, the blot was developed for b-actin (A-2228, Sigma–AldrichChemicals) in 1:1000 dilution as a loading control. Immunodetec-tion of b-actin was performed with anti-mouse IgG-HRP (1:1000).Bands were quantified by measurement of optical density usingScion Image Analysis Software (Scion Corporation, USA). Valueswere expressed as the ratio of the density of the specific signal tothe b-actin signal [27].
2.11. Antioxidant enzyme activity assay
2.11.1. Superoxide dismutase (SOD) assay10% homogenates of the lymphoid tissues were prepared in
150 mM phosphate buffered saline (PBS, pH 7.4) and centrifugedfor 30 min at 12,000g at 4 �C. The supernatant was again centri-fuged for 60 min at 12,000g at 4 �C and then processed for enzy-matic activity based on a modified spectrophotometric methodusing nitrite formation by superoxide radicals [32] and published
elsewhere [26]. One unit of enzyme activity is defined as theamount of SOD inhibiting 50% of nitrite formation under assay con-ditions and was expressed as SOD activity in U/g tissue weight.
2.11.2. Catalase (CAT) assayCatalase activity was measured following the procedure of
Sinha [33]. Immediately after sacrifice, 10% homogenate of spleenand thymus was prepared in PBS (10 mM; pH = 7.4) and thencentrifuged and supernatant was used for enzyme estimation.The supernatant was processed for the assay in a reaction mixturecomprising 0.8 mM H2O2 and potassium dichromate in glacialacetic acid heated in a water bath for 10 min and then OD wastaken at 570 nm and the decrease in H2O2 content was calculated.The standard curve was calibrated with varying concentrations of0.2 mM H2O2 in PBS. The activity of CAT was expressed as amountof H2O2 degraded per minute.
2.11.3. Glutathione peroxidase (GPx) assayGPx activity was assayed in spleen and thymus as described by
Mantha et al. [34]. The reaction mixture (1 ml) contained 50 llsample, 398 ll of 50 mM phosphate buffer (pH 7.0), 2 ll of 1 mMEDTA, 10 ll of 1 mM sodium azide, 500 ll of 0.5 mM NADPH,40 ll of 0.2 mM GSH and 1 U glutathione reductase. The reactionmixture was allowed to equilibrate for 1 min at room temperature.After this, the reaction was initiated by addition of 100 mM H2O2.The absorbance measured kinetically at 340 nm for 3 min. TheGPx activity was expressed as lmol of NADPH oxidized to NADP+
per min per mg of protein using an extinction coefficient(6.22 mM�1 cm�1) for NADPH.
2.12. Statistical analyses
Statistical analysis of the data was performed by one-wayANOVA followed by the Student Newman–Keuls multiple rangetest using software SPSS 17 Inc., USA. Further, to show comparisonbetween two groups of animals exposed to ambient photoperiodicconditions student-t test was performed in Microsoft-Excel sheet.The differences were considered significant when p was found tobe 60.05. Correlation analysis was performed to determine thepossible linear relationship among nuclear melatonin receptorand other parameters studied including plasma melatonin concen-tration and were expressed as Pearson coefficient (r).
3. Results
3.1. Melatonin concentration
Plasma melatonin concentration was significantly (p < 0.01)higher in the squirrels during the reproductively inactive phase(RIP) compared to the reproductively active phase (RAP; Fig. 1a).The plasma melatonin concentration was found to be significantly(p < 0.01) increased in the squirrels exposed to short-daylength(10L:14D), while long-day (16L:8D) exposure showed significant(p < 0.01) decrease in plasma melatonin concentration comparedto 12L:12D control group (Fig. 1b).
Thymus always presented higher melatonin content comparedto spleen. The melatonin content of the lymphoid tissues (spleenand thymus) was found to be significantly (p < 0.01) higher duringthe reproductively inactive phase (Fig. 3c). Exposure to short-day-lengths resulted in significant (p < 0.01) increase of the lymphoidtissue melatonin content compared to 12L:12D exposed controlgroup while long-daylengths significantly lowered the in situ mel-atonin content (Fig. 3d).
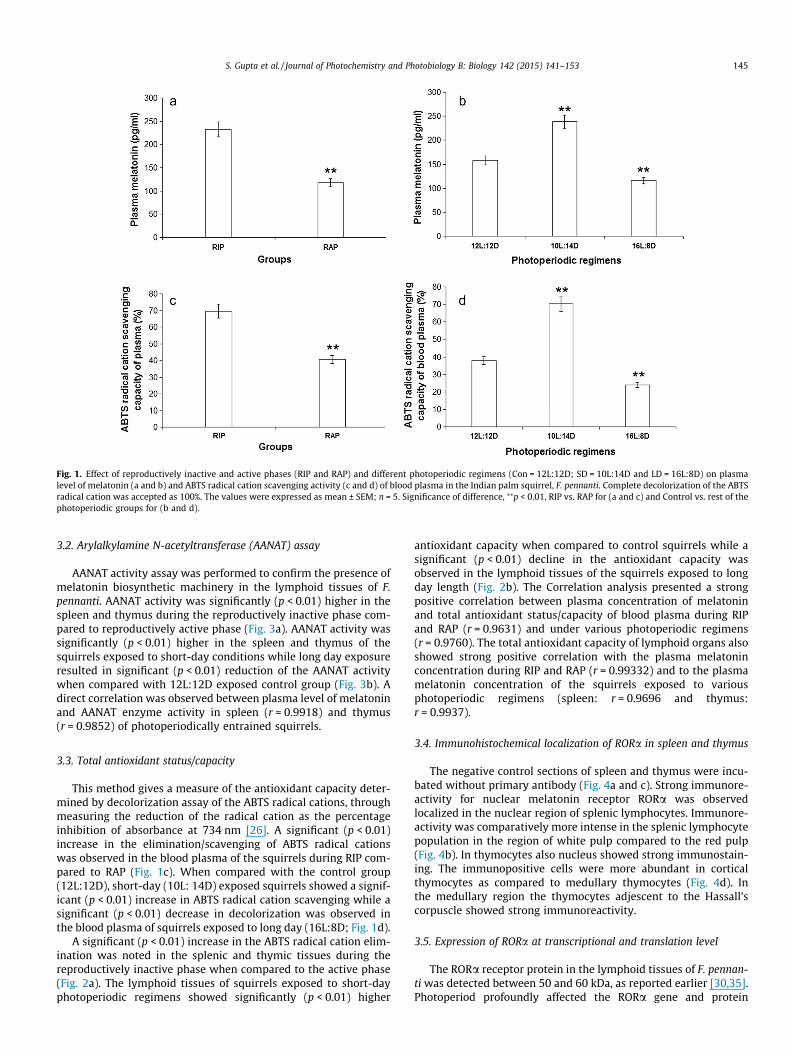
Fig. 1. Effect of reproductively inactive and active phases (RIP and RAP) and different photoperiodic regimens (Con = 12L:12D; SD = 10L:14D and LD = 16L:8D) on plasmalevel of melatonin (a and b) and ABTS radical cation scavenging activity (c and d) of blood plasma in the Indian palm squirrel, F. pennanti. Complete decolorization of the ABTSradical cation was accepted as 100%. The values were expressed as mean ± SEM; n = 5. Significance of difference, ⁄⁄p < 0.01, RIP vs. RAP for (a and c) and Control vs. rest of thephotoperiodic groups for (b and d).
S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 142 (2015) 141–153 145
3.2. Arylalkylamine N-acetyltransferase (AANAT) assay
AANAT activity assay was performed to confirm the presence ofmelatonin biosynthetic machinery in the lymphoid tissues of F.pennanti. AANAT activity was significantly (p < 0.01) higher in thespleen and thymus during the reproductively inactive phase com-pared to reproductively active phase (Fig. 3a). AANAT activity wassignificantly (p < 0.01) higher in the spleen and thymus of thesquirrels exposed to short-day conditions while long day exposureresulted in significant (p < 0.01) reduction of the AANAT activitywhen compared with 12L:12D exposed control group (Fig. 3b). Adirect correlation was observed between plasma level of melatoninand AANAT enzyme activity in spleen (r = 0.9918) and thymus(r = 0.9852) of photoperiodically entrained squirrels.
3.3. Total antioxidant status/capacity
This method gives a measure of the antioxidant capacity deter-mined by decolorization assay of the ABTS radical cations, throughmeasuring the reduction of the radical cation as the percentageinhibition of absorbance at 734 nm [26]. A significant (p < 0.01)increase in the elimination/scavenging of ABTS radical cationswas observed in the blood plasma of the squirrels during RIP com-pared to RAP (Fig. 1c). When compared with the control group(12L:12D), short-day (10L: 14D) exposed squirrels showed a signif-icant (p < 0.01) increase in ABTS radical cation scavenging while asignificant (p < 0.01) decrease in decolorization was observed inthe blood plasma of squirrels exposed to long day (16L:8D; Fig. 1d).
A significant (p < 0.01) increase in the ABTS radical cation elim-ination was noted in the splenic and thymic tissues during thereproductively inactive phase when compared to the active phase(Fig. 2a). The lymphoid tissues of squirrels exposed to short-dayphotoperiodic regimens showed significantly (p < 0.01) higher
antioxidant capacity when compared to control squirrels while asignificant (p < 0.01) decline in the antioxidant capacity wasobserved in the lymphoid tissues of the squirrels exposed to longday length (Fig. 2b). The Correlation analysis presented a strongpositive correlation between plasma concentration of melatoninand total antioxidant status/capacity of blood plasma during RIPand RAP (r = 0.9631) and under various photoperiodic regimens(r = 0.9760). The total antioxidant capacity of lymphoid organs alsoshowed strong positive correlation with the plasma melatoninconcentration during RIP and RAP (r = 0.99332) and to the plasmamelatonin concentration of the squirrels exposed to variousphotoperiodic regimens (spleen: r = 0.9696 and thymus:r = 0.9937).
3.4. Immunohistochemical localization of RORa in spleen and thymus
The negative control sections of spleen and thymus were incu-bated without primary antibody (Fig. 4a and c). Strong immunore-activity for nuclear melatonin receptor RORa was observedlocalized in the nuclear region of splenic lymphocytes. Immunore-activity was comparatively more intense in the splenic lymphocytepopulation in the region of white pulp compared to the red pulp(Fig. 4b). In thymocytes also nucleus showed strong immunostain-ing. The immunopositive cells were more abundant in corticalthymocytes as compared to medullary thymocytes (Fig. 4d). Inthe medullary region the thymocytes adjescent to the Hassall’scorpuscle showed strong immunoreactivity.
3.5. Expression of RORa at transcriptional and translation level
The RORa receptor protein in the lymphoid tissues of F. pennan-ti was detected between 50 and 60 kDa, as reported earlier [30,35].Photoperiod profoundly affected the RORa gene and protein
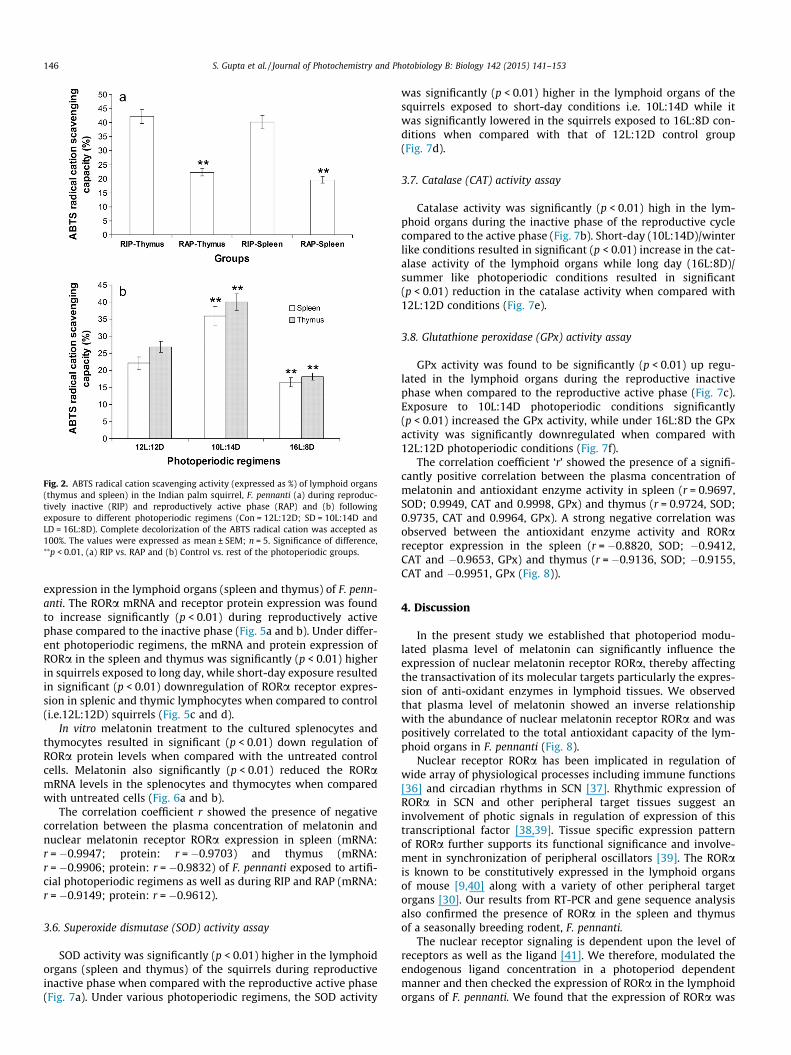
Fig. 2. ABTS radical cation scavenging activity (expressed as %) of lymphoid organs(thymus and spleen) in the Indian palm squirrel, F. pennanti (a) during reproduc-tively inactive (RIP) and reproductively active phase (RAP) and (b) followingexposure to different photoperiodic regimens (Con = 12L:12D; SD = 10L:14D andLD = 16L:8D). Complete decolorization of the ABTS radical cation was accepted as100%. The values were expressed as mean ± SEM; n = 5. Significance of difference,⁄⁄p < 0.01, (a) RIP vs. RAP and (b) Control vs. rest of the photoperiodic groups.
146 S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 142 (2015) 141–153
expression in the lymphoid organs (spleen and thymus) of F. penn-anti. The RORa mRNA and receptor protein expression was foundto increase significantly (p < 0.01) during reproductively activephase compared to the inactive phase (Fig. 5a and b). Under differ-ent photoperiodic regimens, the mRNA and protein expression ofRORa in the spleen and thymus was significantly (p < 0.01) higherin squirrels exposed to long day, while short-day exposure resultedin significant (p < 0.01) downregulation of RORa receptor expres-sion in splenic and thymic lymphocytes when compared to control(i.e.12L:12D) squirrels (Fig. 5c and d).
In vitro melatonin treatment to the cultured splenocytes andthymocytes resulted in significant (p < 0.01) down regulation ofRORa protein levels when compared with the untreated controlcells. Melatonin also significantly (p < 0.01) reduced the RORamRNA levels in the splenocytes and thymocytes when comparedwith untreated cells (Fig. 6a and b).
The correlation coefficient r showed the presence of negativecorrelation between the plasma concentration of melatonin andnuclear melatonin receptor RORa expression in spleen (mRNA:r = �0.9947; protein: r = �0.9703) and thymus (mRNA:r = �0.9906; protein: r = �0.9832) of F. pennanti exposed to artifi-cial photoperiodic regimens as well as during RIP and RAP (mRNA:r = �0.9149; protein: r = �0.9612).
3.6. Superoxide dismutase (SOD) activity assay
SOD activity was significantly (p < 0.01) higher in the lymphoidorgans (spleen and thymus) of the squirrels during reproductiveinactive phase when compared with the reproductive active phase(Fig. 7a). Under various photoperiodic regimens, the SOD activity
was significantly (p < 0.01) higher in the lymphoid organs of thesquirrels exposed to short-day conditions i.e. 10L:14D while itwas significantly lowered in the squirrels exposed to 16L:8D con-ditions when compared with that of 12L:12D control group(Fig. 7d).
3.7. Catalase (CAT) activity assay
Catalase activity was significantly (p < 0.01) high in the lym-phoid organs during the inactive phase of the reproductive cyclecompared to the active phase (Fig. 7b). Short-day (10L:14D)/winterlike conditions resulted in significant (p < 0.01) increase in the cat-alase activity of the lymphoid organs while long day (16L:8D)/summer like photoperiodic conditions resulted in significant(p < 0.01) reduction in the catalase activity when compared with12L:12D conditions (Fig. 7e).
3.8. Glutathione peroxidase (GPx) activity assay
GPx activity was found to be significantly (p < 0.01) up regu-lated in the lymphoid organs during the reproductive inactivephase when compared to the reproductive active phase (Fig. 7c).Exposure to 10L:14D photoperiodic conditions significantly(p < 0.01) increased the GPx activity, while under 16L:8D the GPxactivity was significantly downregulated when compared with12L:12D photoperiodic conditions (Fig. 7f).
The correlation coefficient ‘r’ showed the presence of a signifi-cantly positive correlation between the plasma concentration ofmelatonin and antioxidant enzyme activity in spleen (r = 0.9697,SOD; 0.9949, CAT and 0.9998, GPx) and thymus (r = 0.9724, SOD;0.9735, CAT and 0.9964, GPx). A strong negative correlation wasobserved between the antioxidant enzyme activity and RORareceptor expression in the spleen (r = �0.8820, SOD; �0.9412,CAT and �0.9653, GPx) and thymus (r = �0.9136, SOD; �0.9155,CAT and �0.9951, GPx (Fig. 8)).
4. Discussion
In the present study we established that photoperiod modu-lated plasma level of melatonin can significantly influence theexpression of nuclear melatonin receptor RORa, thereby affectingthe transactivation of its molecular targets particularly the expres-sion of anti-oxidant enzymes in lymphoid tissues. We observedthat plasma level of melatonin showed an inverse relationshipwith the abundance of nuclear melatonin receptor RORa and waspositively correlated to the total antioxidant capacity of the lym-phoid organs in F. pennanti (Fig. 8).
Nuclear receptor RORa has been implicated in regulation ofwide array of physiological processes including immune functions[36] and circadian rhythms in SCN [37]. Rhythmic expression ofRORa in SCN and other peripheral target tissues suggest aninvolvement of photic signals in regulation of expression of thistranscriptional factor [38,39]. Tissue specific expression patternof RORa further supports its functional significance and involve-ment in synchronization of peripheral oscillators [39]. The RORais known to be constitutively expressed in the lymphoid organsof mouse [9,40] along with a variety of other peripheral targetorgans [30]. Our results from RT-PCR and gene sequence analysisalso confirmed the presence of RORa in the spleen and thymusof a seasonally breeding rodent, F. pennanti.
The nuclear receptor signaling is dependent upon the level ofreceptors as well as the ligand [41]. We therefore, modulated theendogenous ligand concentration in a photoperiod dependentmanner and then checked the expression of RORa in the lymphoidorgans of F. pennanti. We found that the expression of RORa was
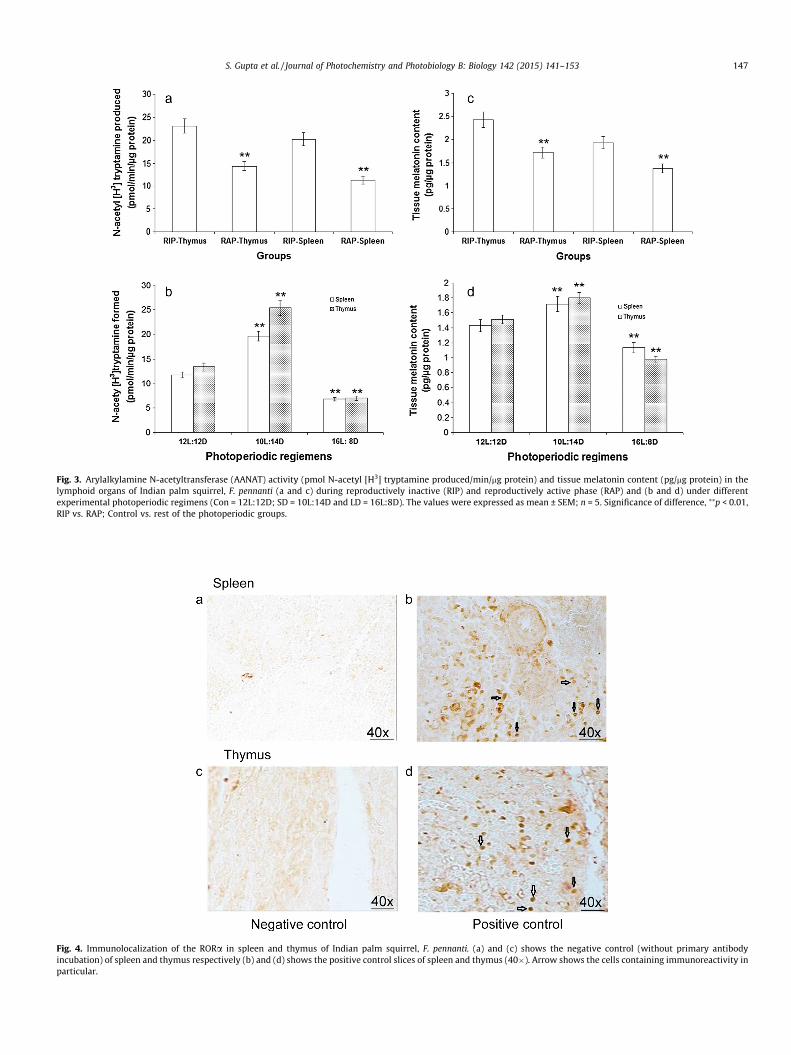
Fig. 3. Arylalkylamine N-acetyltransferase (AANAT) activity (pmol N-acetyl [H3] tryptamine produced/min/mg protein) and tissue melatonin content (pg/mg protein) in thelymphoid organs of Indian palm squirrel, F. pennanti (a and c) during reproductively inactive (RIP) and reproductively active phase (RAP) and (b and d) under differentexperimental photoperiodic regimens (Con = 12L:12D; SD = 10L:14D and LD = 16L:8D). The values were expressed as mean ± SEM; n = 5. Significance of difference, ⁄⁄p < 0.01,RIP vs. RAP; Control vs. rest of the photoperiodic groups.
Fig. 4. Immunolocalization of the RORa in spleen and thymus of Indian palm squirrel, F. pennanti. (a) and (c) shows the negative control (without primary antibodyincubation) of spleen and thymus respectively (b) and (d) shows the positive control slices of spleen and thymus (40�). Arrow shows the cells containing immunoreactivity inparticular.
S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 142 (2015) 141–153 147
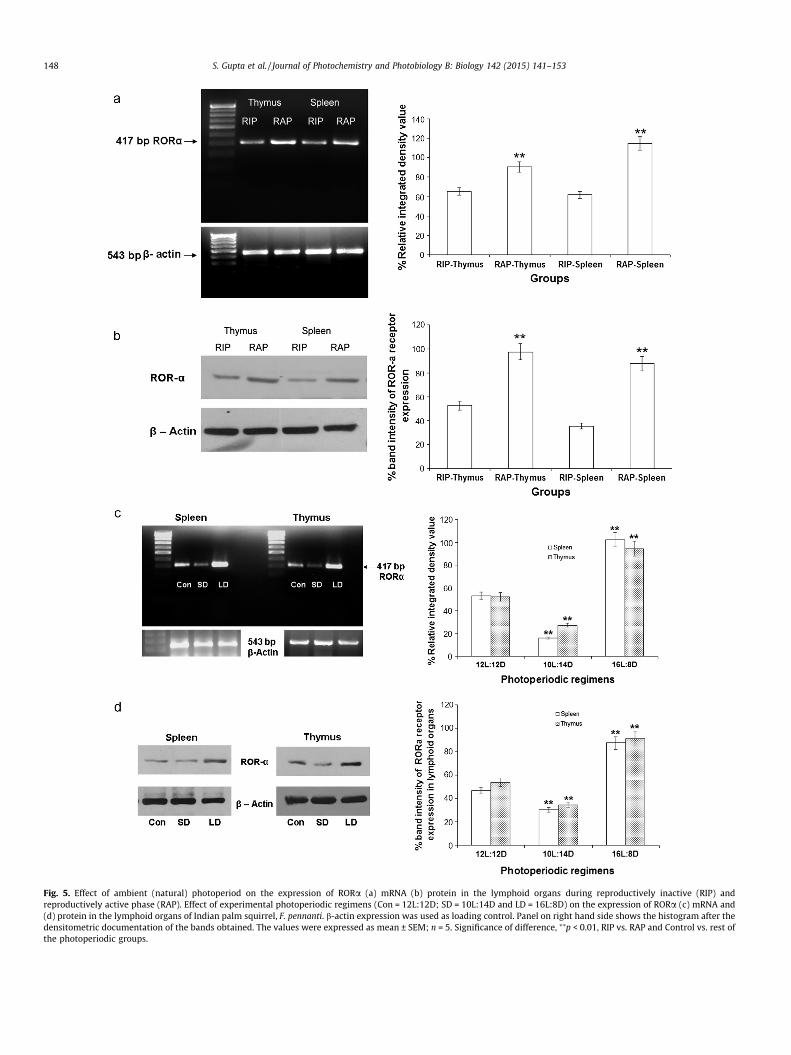
Fig. 5. Effect of ambient (natural) photoperiod on the expression of RORa (a) mRNA (b) protein in the lymphoid organs during reproductively inactive (RIP) andreproductively active phase (RAP). Effect of experimental photoperiodic regimens (Con = 12L:12D; SD = 10L:14D and LD = 16L:8D) on the expression of RORa (c) mRNA and(d) protein in the lymphoid organs of Indian palm squirrel, F. pennanti. b-actin expression was used as loading control. Panel on right hand side shows the histogram after thedensitometric documentation of the bands obtained. The values were expressed as mean ± SEM; n = 5. Significance of difference, ⁄⁄p < 0.01, RIP vs. RAP and Control vs. rest ofthe photoperiodic groups.
148 S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 142 (2015) 141–153
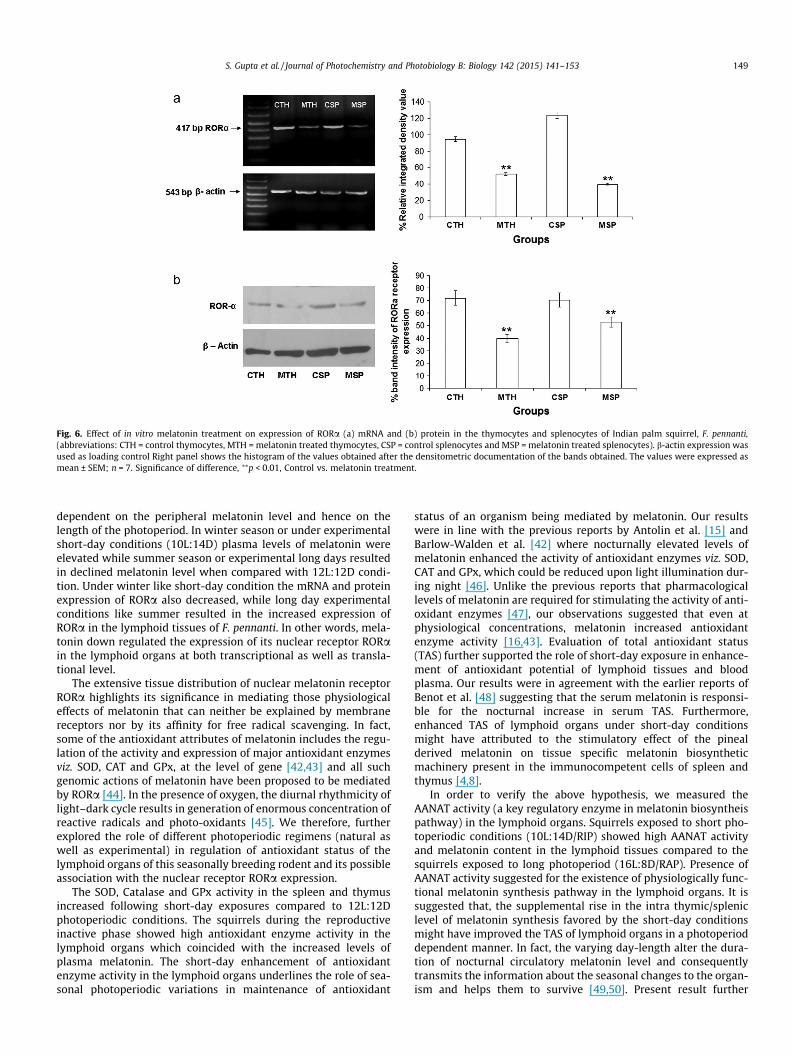
Fig. 6. Effect of in vitro melatonin treatment on expression of RORa (a) mRNA and (b) protein in the thymocytes and splenocytes of Indian palm squirrel, F. pennanti,(abbreviations: CTH = control thymocytes, MTH = melatonin treated thymocytes, CSP = control splenocytes and MSP = melatonin treated splenocytes). b-actin expression wasused as loading control Right panel shows the histogram of the values obtained after the densitometric documentation of the bands obtained. The values were expressed asmean ± SEM; n = 7. Significance of difference, ⁄⁄p < 0.01, Control vs. melatonin treatment.
S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 142 (2015) 141–153 149
dependent on the peripheral melatonin level and hence on thelength of the photoperiod. In winter season or under experimentalshort-day conditions (10L:14D) plasma levels of melatonin wereelevated while summer season or experimental long days resultedin declined melatonin level when compared with 12L:12D condi-tion. Under winter like short-day condition the mRNA and proteinexpression of RORa also decreased, while long day experimentalconditions like summer resulted in the increased expression ofRORa in the lymphoid tissues of F. pennanti. In other words, mela-tonin down regulated the expression of its nuclear receptor RORain the lymphoid organs at both transcriptional as well as transla-tional level.
The extensive tissue distribution of nuclear melatonin receptorRORa highlights its significance in mediating those physiologicaleffects of melatonin that can neither be explained by membranereceptors nor by its affinity for free radical scavenging. In fact,some of the antioxidant attributes of melatonin includes the regu-lation of the activity and expression of major antioxidant enzymesviz. SOD, CAT and GPx, at the level of gene [42,43] and all suchgenomic actions of melatonin have been proposed to be mediatedby RORa [44]. In the presence of oxygen, the diurnal rhythmicity oflight–dark cycle results in generation of enormous concentration ofreactive radicals and photo-oxidants [45]. We therefore, furtherexplored the role of different photoperiodic regimens (natural aswell as experimental) in regulation of antioxidant status of thelymphoid organs of this seasonally breeding rodent and its possibleassociation with the nuclear receptor RORa expression.
The SOD, Catalase and GPx activity in the spleen and thymusincreased following short-day exposures compared to 12L:12Dphotoperiodic conditions. The squirrels during the reproductiveinactive phase showed high antioxidant enzyme activity in thelymphoid organs which coincided with the increased levels ofplasma melatonin. The short-day enhancement of antioxidantenzyme activity in the lymphoid organs underlines the role of sea-sonal photoperiodic variations in maintenance of antioxidant
status of an organism being mediated by melatonin. Our resultswere in line with the previous reports by Antolin et al. [15] andBarlow-Walden et al. [42] where nocturnally elevated levels ofmelatonin enhanced the activity of antioxidant enzymes viz. SOD,CAT and GPx, which could be reduced upon light illumination dur-ing night [46]. Unlike the previous reports that pharmacologicallevels of melatonin are required for stimulating the activity of anti-oxidant enzymes [47], our observations suggested that even atphysiological concentrations, melatonin increased antioxidantenzyme activity [16,43]. Evaluation of total antioxidant status(TAS) further supported the role of short-day exposure in enhance-ment of antioxidant potential of lymphoid tissues and bloodplasma. Our results were in agreement with the earlier reports ofBenot et al. [48] suggesting that the serum melatonin is responsi-ble for the nocturnal increase in serum TAS. Furthermore,enhanced TAS of lymphoid organs under short-day conditionsmight have attributed to the stimulatory effect of the pinealderived melatonin on tissue specific melatonin biosyntheticmachinery present in the immunocompetent cells of spleen andthymus [4,8].
In order to verify the above hypothesis, we measured theAANAT activity (a key regulatory enzyme in melatonin biosyntheispathway) in the lymphoid organs. Squirrels exposed to short pho-toperiodic conditions (10L:14D/RIP) showed high AANAT activityand melatonin content in the lymphoid tissues compared to thesquirrels exposed to long photoperiod (16L:8D/RAP). Presence ofAANAT activity suggested for the existence of physiologically func-tional melatonin synthesis pathway in the lymphoid organs. It issuggested that, the supplemental rise in the intra thymic/spleniclevel of melatonin synthesis favored by the short-day conditionsmight have improved the TAS of lymphoid organs in a photoperioddependent manner. In fact, the varying day-length alter the dura-tion of nocturnal circulatory melatonin level and consequentlytransmits the information about the seasonal changes to the organ-ism and helps them to survive [49,50]. Present result further
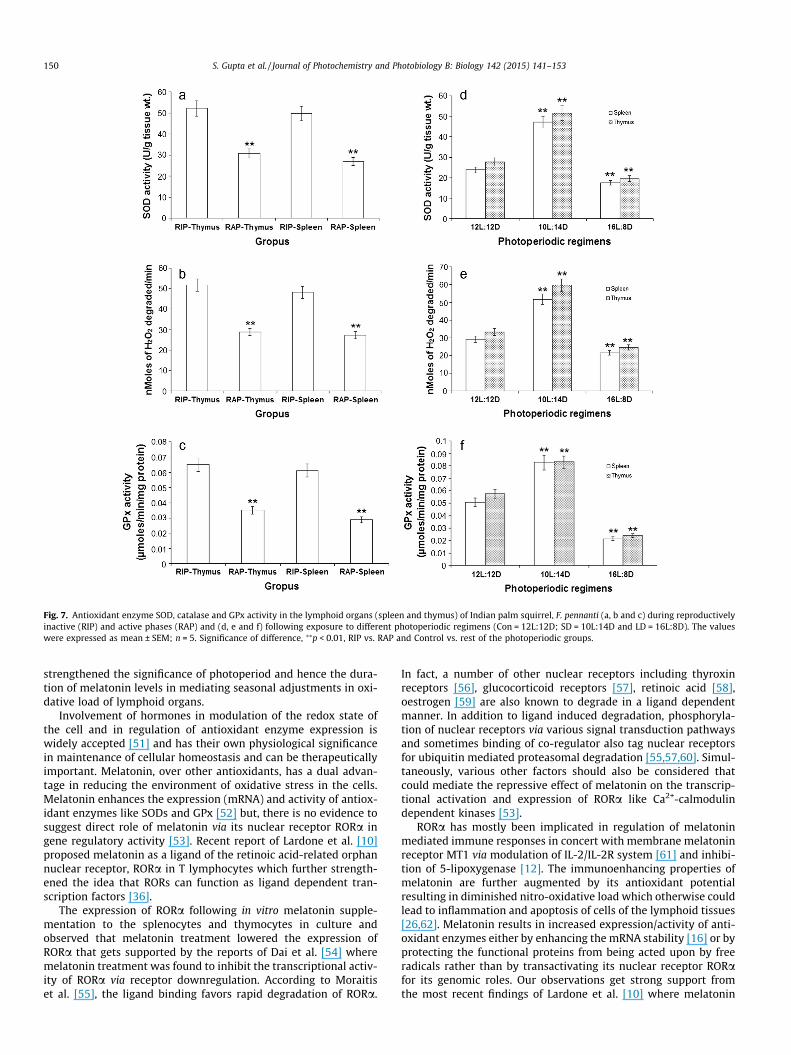
Fig. 7. Antioxidant enzyme SOD, catalase and GPx activity in the lymphoid organs (spleen and thymus) of Indian palm squirrel, F. pennanti (a, b and c) during reproductivelyinactive (RIP) and active phases (RAP) and (d, e and f) following exposure to different photoperiodic regimens (Con = 12L:12D; SD = 10L:14D and LD = 16L:8D). The valueswere expressed as mean ± SEM; n = 5. Significance of difference, ⁄⁄p < 0.01, RIP vs. RAP and Control vs. rest of the photoperiodic groups.
150 S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 142 (2015) 141–153
strengthened the significance of photoperiod and hence the dura-tion of melatonin levels in mediating seasonal adjustments in oxi-dative load of lymphoid organs.
Involvement of hormones in modulation of the redox state ofthe cell and in regulation of antioxidant enzyme expression iswidely accepted [51] and has their own physiological significancein maintenance of cellular homeostasis and can be therapeuticallyimportant. Melatonin, over other antioxidants, has a dual advan-tage in reducing the environment of oxidative stress in the cells.Melatonin enhances the expression (mRNA) and activity of antiox-idant enzymes like SODs and GPx [52] but, there is no evidence tosuggest direct role of melatonin via its nuclear receptor RORa ingene regulatory activity [53]. Recent report of Lardone et al. [10]proposed melatonin as a ligand of the retinoic acid-related orphannuclear receptor, RORa in T lymphocytes which further strength-ened the idea that RORs can function as ligand dependent tran-scription factors [36].
The expression of RORa following in vitro melatonin supple-mentation to the splenocytes and thymocytes in culture andobserved that melatonin treatment lowered the expression ofRORa that gets supported by the reports of Dai et al. [54] wheremelatonin treatment was found to inhibit the transcriptional activ-ity of RORa via receptor downregulation. According to Moraitiset al. [55], the ligand binding favors rapid degradation of RORa.
In fact, a number of other nuclear receptors including thyroxinreceptors [56], glucocorticoid receptors [57], retinoic acid [58],oestrogen [59] are also known to degrade in a ligand dependentmanner. In addition to ligand induced degradation, phosphoryla-tion of nuclear receptors via various signal transduction pathwaysand sometimes binding of co-regulator also tag nuclear receptorsfor ubiquitin mediated proteasomal degradation [55,57,60]. Simul-taneously, various other factors should also be considered thatcould mediate the repressive effect of melatonin on the transcrip-tional activation and expression of RORa like Ca2+-calmodulindependent kinases [53].
RORa has mostly been implicated in regulation of melatoninmediated immune responses in concert with membrane melatoninreceptor MT1 via modulation of IL-2/IL-2R system [61] and inhibi-tion of 5-lipoxygenase [12]. The immunoenhancing properties ofmelatonin are further augmented by its antioxidant potentialresulting in diminished nitro-oxidative load which otherwise couldlead to inflammation and apoptosis of cells of the lymphoid tissues[26,62]. Melatonin results in increased expression/activity of anti-oxidant enzymes either by enhancing the mRNA stability [16] or byprotecting the functional proteins from being acted upon by freeradicals rather than by transactivating its nuclear receptor RORafor its genomic roles. Our observations get strong support fromthe most recent findings of Lardone et al. [10] where melatonin
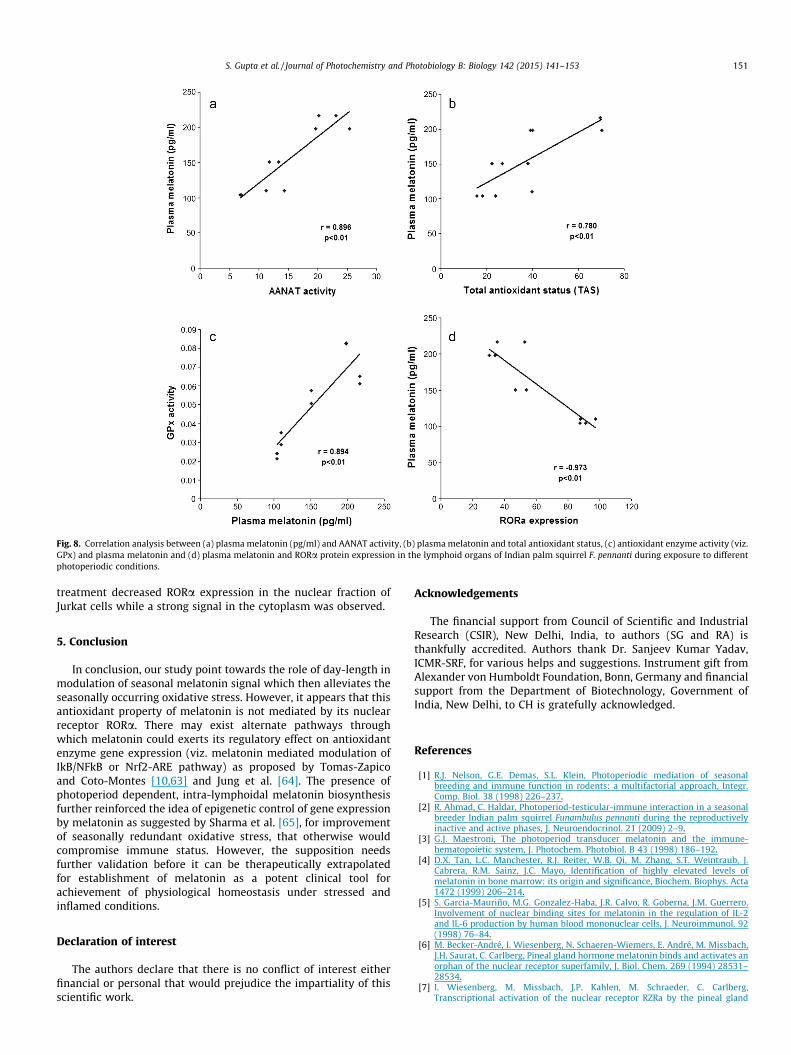
Fig. 8. Correlation analysis between (a) plasma melatonin (pg/ml) and AANAT activity, (b) plasma melatonin and total antioxidant status, (c) antioxidant enzyme activity (viz.GPx) and plasma melatonin and (d) plasma melatonin and RORa protein expression in the lymphoid organs of Indian palm squirrel F. pennanti during exposure to differentphotoperiodic conditions.
S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 142 (2015) 141–153 151
treatment decreased RORa expression in the nuclear fraction ofJurkat cells while a strong signal in the cytoplasm was observed.
5. Conclusion
In conclusion, our study point towards the role of day-length inmodulation of seasonal melatonin signal which then alleviates theseasonally occurring oxidative stress. However, it appears that thisantioxidant property of melatonin is not mediated by its nuclearreceptor RORa. There may exist alternate pathways throughwhich melatonin could exerts its regulatory effect on antioxidantenzyme gene expression (viz. melatonin mediated modulation ofIkB/NFkB or Nrf2-ARE pathway) as proposed by Tomas-Zapicoand Coto-Montes [10,63] and Jung et al. [64]. The presence ofphotoperiod dependent, intra-lymphoidal melatonin biosynthesisfurther reinforced the idea of epigenetic control of gene expressionby melatonin as suggested by Sharma et al. [65], for improvementof seasonally redundant oxidative stress, that otherwise wouldcompromise immune status. However, the supposition needsfurther validation before it can be therapeutically extrapolatedfor establishment of melatonin as a potent clinical tool forachievement of physiological homeostasis under stressed andinflamed conditions.
Declaration of interest
The authors declare that there is no conflict of interest eitherfinancial or personal that would prejudice the impartiality of thisscientific work.
Acknowledgements
The financial support from Council of Scientific and IndustrialResearch (CSIR), New Delhi, India, to authors (SG and RA) isthankfully accredited. Authors thank Dr. Sanjeev Kumar Yadav,ICMR-SRF, for various helps and suggestions. Instrument gift fromAlexander von Humboldt Foundation, Bonn, Germany and financialsupport from the Department of Biotechnology, Government ofIndia, New Delhi, to CH is gratefully acknowledged.
References
[1] R.J. Nelson, G.E. Demas, S.L. Klein, Photoperiodic mediation of seasonalbreeding and immune function in rodents: a multifactorial approach, Integr.Comp. Biol. 38 (1998) 226–237.
[2] R. Ahmad, C. Haldar, Photoperiod-testicular-immune interaction in a seasonalbreeder Indian palm squirrel Funambulus pennanti during the reproductivelyinactive and active phases, J. Neuroendocrinol. 21 (2009) 2–9.
[3] G.J. Maestroni, The photoperiod transducer melatonin and the immune-hematopoietic system, J. Photochem. Photobiol. B 43 (1998) 186–192.
[4] D.X. Tan, L.C. Manchester, R.J. Reiter, W.B. Qi, M. Zhang, S.T. Weintraub, J.Cabrera, R.M. Sainz, J.C. Mayo, Identification of highly elevated levels ofmelatonin in bone marrow: its origin and significance, Biochem. Biophys. Acta1472 (1999) 206–214.
[5] S. Garcia-Mauriño, M.G. Gonzalez-Haba, J.R. Calvo, R. Goberna, J.M. Guerrero,Involvement of nuclear binding sites for melatonin in the regulation of IL-2and IL-6 production by human blood mononuclear cells, J. Neuroimmunol. 92(1998) 76–84.
[6] M. Becker-André, I. Wiesenberg, N. Schaeren-Wiemers, E. André, M. Missbach,J.H. Saurat, C. Carlberg, Pineal gland hormone melatonin binds and activates anorphan of the nuclear receptor superfamily, J. Biol. Chem. 269 (1994) 28531–28534.
[7] I. Wiesenberg, M. Missbach, J.P. Kahlen, M. Schraeder, C. Carlberg,Transcriptional activation of the nuclear receptor RZRa by the pineal gland

152 S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 142 (2015) 141–153
hormone melatonin and identification of CGP 52608 as a synthetic ligand,Nucl. Acids Res. 23 (1995) 327–333.
[8] A. Carrillo-Vico, J.R. Calvo, P. Abreu, P.J. Lardone, S. García-Mauriño, R.J. Reiter,J.M. Guerrero, Evidence of melatonin synthesis by human lymphocytes and itsphysiological significance: possible role as intracrine, autocrine, and/orparacrine substance, FASEB J. 18 (2004) 537–539.
[9] A. Carrillo-Vico, A. García-Pergañeda, L. Naji, J.R. Calvo, M.P. Romero, J.M.Guerrero, Expression of membrane and nuclear melatonin receptor mRNA andprotein in the mouse immune system, Cell. Mol. Life Sci. 60 (2003) 2272–2278.
[10] P.J. Lardone, J.M. Guerrero, J.M. Fernández-Santos, A. Rubio, I. Martín-Lacave, A.Carrillo-Vico, Melatonin synthesized by T lymphocytes as a ligand of theretinoic acid-related orphan receptor, J. Pineal Res. 51 (2011) 454–462.
[11] M. Rafii-El-Idrissi, J.R. Calvo, A. Harmouch, S. García-Mauriño, J.M. Guerrero,Specific binding of melatonin by purified cell nuclei from spleen and thymus ofthe rat, J. Neuroimmunol. 86 (1998) 190–197.
[12] D. Steinhilber, M. Brungs, O. Werz, I. Wiesenberg, C. Danielsson, J.P. Kahlen, S.Nayeri, M. Schraeder, C. Carlberg, The nuclear receptor for melatonin represses5-lipoxygenase gene expression in human B lymphocytes, J. Biol. Chem. 270(1995) 7037–7040.
[13] M. Schräder, C. Danielsson, I. Wiesenberg, C. Carlberg, Identification of naturalmonomeric response elements of the nuclear receptor RZR/ROR. They alsobind COUP-TF homodimers, J. Biol. Chem. 271 (1996) 19732–19736.
[14] C. Carlberg, Gene regulation by melatonin, Ann. N. Y. Acad. Sci. 917 (2000)387–396.
[15] I. Antolín, C. Rodríguez, R.M. Saínz, J.C. Mayo, H. Uría, M.L. Kotler, M.J.Rodríguez-Colunga, D. Tolivia, A. Menéndez-Peláez, Neurohormone melatoninprevents cell damage: effect on gene expression for antioxidant enzymes,FASEB J. 10 (1996) 882–890.
[16] J.C. Mayo, R.M. Sainz, I. Antolin, F. Herrera, V. Martin, C. Rodriguez,Melatonin regulation of antioxidant enzyme gene expression, Cell Mol. LifeSci. 59 (2002) 1706–1713.
[17] C. Tomás-Zapico, A. Coto-Montes, J. Martínez-Fraga, M.J. Rodríguez-Colunga, R.Hardeland, D. Tolivia, Effects of delta-aminolevulinic acid and melatonin in theharderian gland of female Syrian hamsters, Free Radic. Biol. Med. 32 (2002)1197–1204.
[18] S. Gupta, C. Haldar, Physiological crosstalk between melatonin andglucocorticoid receptor modulates T-cell mediated immune responses in awild tropical rodent, Funambulus pennanti, J. Steroid Biochem. Mol. Biol. 134(2013) 23–36.
[19] C. Haldar, N. Saxena, Pineal gland and humidity effects on testicular function ofthe Indian palm squirrel, Funambulus pennanti, J. Pineal Res. 5 (1988) 411–418.
[20] M.D. Rollag, G.D. Niswender, Radioimmunoassay of melatonin in sheepexposed to different light regimes, Endocrinology 98 (1976) 482–488.
[21] C. Haldar, S. Dubey, Diurnal variations in circulating estradiol, testosterone,melatonin and harderian gland porphyrin concentration in Indian palmsquirrel, Funambulus pennanti, Indian J. Exp. Biol. 34 (1997) 695–697.
[22] H.D. Chae, T.J. Park, Y.K. Lee, T.G. Lee, K.T. Kim, Rapid and simple measurementof serotonin N-acetyltransferase activity by liquid biphasic diffusion assay,Neurochem. Int. 35 (1999) 447–451.
[23] A. Piesiewicz, E. Podobas, U. Kedzierska, E. Joachimiak, M. Markowska, P.Majewski, K. Skwarlo-Sonta, Season-related differences in the biosyntheticactivity of the neonatal chicken pineal gland, Open Ornithol. J. 3 (2010) 134–140.
[24] M.M. Bradford, A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dyebinding, Anal. Biochem. 72 (1976) 248–254.
[25] R. Re, N. Pellegrini, A. Proteggente, A. Pannala, M. Yang, C. Rice-Evans,Antioxidant activity applying an improved ABTS radical cation decolorizationassay, Free Radic. Biol. Med. 26 (1999) 1231–1237.
[26] S. Sharma, C. Haldar, Comparative effect of melatonin and vitamin E onphenylhydrazine-induced toxicity in the spleen of Funambulus pennanti,Environ. Toxicol. 24 (2009) 1–9.
[27] R. Ahmad, C. Haldar, Photoperiodic regulation of melatonin receptor MT1 &MT2 expression dynamics in spleen and thymus of a tropical rodentFunambulus pennanti during reproductively active and inactive phases,Chronobiol. Int. 27 (2010) 446–462.
[28] R. Ahmad, C. Haldar, S. Gupta, Melatonin membrane receptor type MT1modulates cell mediated immunity in seasonally breeding tropical rodentFunambulus pennanti, Neuroimmunomodulation 19 (2012) 50–59.
[29] S. Lahiri, C. Haldar, Response of melatonin receptor MT1 in spleen of a tropicalIndian rodent, Funambulus pennanti, to natural solar insolation and differentphotoperiodic conditions, Chronobiol. Int. 26 (2009) 1559–1574.
[30] L. Naji, A. Carrillo-Vico, J.M. Guerrero, J.R. Calvo, Expression of membrane andnuclear melatonin receptors in mouse peripheral organs, Life Sci. 74 (2004)2227–2236.
[31] P.K. Modi, M.S. Kanungo, Age-dependent expression of S100beta in the brain ofmice, Cell. Mol. Neurobiol. 30 (2010) 709–716.
[32] K. Das, L. Samanta, G.B.N. Chainy, A modified spectrophotometric assay ofsuperoxide dismutase using nitrite formation by superoxide radicals, Indian J.Biochem. Biophys. 37 (2000) 201–204.
[33] A.K. Sinha, Colorimetric assay of Catalase, Anal. Biochem. 47 (1972) 389–394.[34] S.V. Mantha, M. Prasad, J. Kalra, K. Prasad, Antioxidant enzymes in
hypercholesterolemia and effects of vitamin E in rabbits, Atherosclerosis 101(1993) 135–144.
[35] K. Bordji, J.P. Grillasca, J.N. Gouze, J. Magdalou, H. Schohn, J.M. Keller, A.Bianchi, M. Dauça, P. Netter, B. Terlain, Evidence for the presence of
peroxisome proliferator-activated receptor (PPAR) alpha and gamma andretinoid Z receptor in cartilage. PPARgamma activation modulates the effectsof interleukin-1beta on rat chondrocytes, J. Biol. Chem. 275 (2000) 12243–12250.
[36] A.M. Jetten, Recent advances in the mechanisms of action and physiologicalfunctions of the retinoid related orphan receptors (RORs), Curr. Drug TargetsInflamm. Allergy 3 (2004) 395–412.
[37] M. Akashi, T. Takumi, The orphan nuclear receptor RORalpha regulatescircadian transcription of the mammalian core-clock Bmal1, Nat. Struct. Mol.Biol. 12 (2005) 441–448.
[38] H.R. Ueda, W. Chen, A. Adachi, H. Wakamatsu, S. Hayashi, T. Takasugi, M.Nagano, K. Nakahama, Y. Suzuki, S. Sugano, M. Iino, Y. Shigeyoshi, S.Hashimoto, A transcription factor response element for gene expressionduring circadian night, Nature 418 (2002) 534–539.
[39] F. Guillaumond, H. Dardente, V. Giguère, N. Cermakian, Differential control ofBmal1 circadian transcription by REV-ERB and ROR nuclear receptors, J. Biol.Rhythms 20 (2004) 391–403.
[40] I. Dzhagalov, V. Giguere, Y.W. He, Lymphocyte development and function inthe absence of retinoic acid-related orphan receptor, J. Immunol. 173 (2004)2952–2959.
[41] X. Yang, A wheel of time: the circadian clock, nuclear receptors, andphysiology, Genes Dev. 24 (2010) 741–747.
[42] L.R. Barlow-Walden, R.J. Reiter, M. Abe, M. Pablos, A. Menendez-Pelaez, L.D.Chen, B. Poeggeler, Melatonin stimulates brain glutathione peroxidase activity,Neurochem. Int. 26 (1995) 497–502.
[43] T.W. Fischer, K. Kleszczynski, L.H. Hardkop, N. Kruse, D. Zillikens, Melatoninenhances antioxidative enzyme gene expression (CAT, GPx, SOD), preventstheir UVR-induced depletion, and protects against the formation of DNAdamage (8-hydroxy-2’-deoxyguanosine) in ex vivo human skin, J. Pineal Res.54 (2012) 303–312.
[44] C. Rodriguez, J.C. Mayo, R.M. Sainz, I. Antolín, F. Herrera, V. Martín, R.J. Reiter,Regulation of antioxidant enzymes: a significant role for melatonin, J. PinealRes. 36 (2004) 1–9.
[45] R. Hardeland, B. Poeggeler, I. Balzer, G. Behrmann, in: C. Guttenbrunner, G.Hidebrant, R. Moog (Eds.), A hypothesis on the evoulutionary orgins ofphotoperiodism based on circadian rhythmicity of melatonin inphylogenetically distant organisms, Chronobiology and Chronomedicine.Frankfurt: Peter Long, Verlag, 1993, pp. 113–120.
[46] M.I. Pablos, R.J. Reiter, G.C. Ortiz, M.T. Agapito, J.I. Chuang, E. Sewerynek,Rhythms of glutathione peroxidase and glutathione reductase in the brain ofchick and their inhibition by light, Neurochem. Int. 32 (1998) 69–75.
[47] M.I. Pablos, J.M. Guerrero, G.C. Ortiz, M.T. Agapito, R.J. Reiter, Both melatoninand a putative nuclear melatonin receptor agonist CGP52608 stimulateglutathione peroxidase and glutathione reductase activities in mouse brainin vivo, Neuroendocrinol. Lett. 18 (1997) 49–58.
[48] S. Benot, R. Goberna, R.J. Reiter, S. Garcia-Mauriño, C. Osuna, J.M. Guerrero,Physiological levels of melatonin contribute to the antioxidant capacity ofhuman serum, J. Pineal Res. 27 (1999) 59–64.
[49] H. Illnerová, J. Vancek, Pineal rhythm in N-acetyltransferase activity in ratsunder different artificial photoperiods and in natural daylight in the course ofa year, Neuroendocrinology 31 (1980) 321–326.
[50] T.J. Bartness, B.D. Goldman, Mammalian pineal melatonin: a clock for allseasons, Experientia 45 (1989) 939–945.
[51] A.D. Bolzán, O.A. Brown, R.G. Goya, M.S. Bianchi, Hormonal modulation ofantioxidant enzyme activities in young and old rats, Exp. Gerontol. 30 (1995)169–175.
[52] M. Kotler, C. Rodriquez, R.M. Sainz, I. Antolin, A. Menendez-Pelaez, Melatoninincreases gene expression for antioxidative enzymes in rat brain cortex, J.Pineal Res. 24 (1998) 83–89.
[53] C.D. Kane, A.R. Means, Activation of orphan receptor-mediated transcriptionby Ca(2+)/calmodulin-dependent protein kinase IV, EMBO J. 19 (2000) 691–701.
[54] J. Dai, P.T. Ram, L. Yuan, L.L. Spriggs, S.M. Hill, Transcriptional repression ofRORalpha activity in human breast cancer cells by melatonin, Mol., CellEndocrinol. 176 (2001) 111–120.
[55] A.N. Moraitis, V. Giguere, The corepressor hairless protects RORa orphannuclear receptor from proteasome-mediated degradation, J. Biol. Chem. 278(2003) 2511–52518.
[56] A. Dace, L. Zhao, K.S. Park, T. Furuno, N. Takamura, M. Nakanishi, B.L. West, J.A.Hanover, S. Cheng, Hormone binding induces rapid proteasome-mediateddegradation of thyroid hormone receptors, Proc. Natl. Acad. Sci. U.S.A. 97(2000) 8985–8990.
[57] A.D. Wallace, J.A. Cidlowski, Proteasome-mediated glucocorticoid receptordegradation restricts transcriptional signaling by glucocorticoids, J. Biol. Chem.276 (2001) 42714–42721.
[58] M. Boudjelal, Z. Wang, J.J. Voorhees, G.J. Fisher, Ubiquitin/proteasome pathwayregulates levels of retinoic acid receptor gamma and retinoid X receptor alphain human keratinocytes, Cancer Res. 60 (2000) 2247–2252.
[59] D.M. Lonard, Z. Nawaz, C.L. Smith, B.W. O’Malley, The 26S proteasome isrequired for estrogen receptor-alpha and coactivator turnover and for efficientestrogen receptor-alpha transactivation, Mol. Cell 5 (2000) 939–948.
[60] E. Kopf, J.L. Plassat, V. Vivat, H. de Thé, P. Chambon, C. Rochette-Egly,Dimerization with retinoid X receptors and phosphorylation modulate theretinoic acid-induced degradation of retinoic acid receptors alpha and gammathrough the ubiquitin-proteasome pathway, J. Biol. Chem. 275 (2000) 33280–33288.

S. Gupta et al. / Journal of Photochemistry and Photobiology B: Biology 142 (2015) 141–153 153
[61] P.J. Lardone, A. Carrillo-Vico, P. Molinero, A. Rubio, J.M. Guerrero, A novel interplaybetween membrane and nuclear melatonin receptors in human lymphocytes:significance in IL-2 production, Cell Mol. Life Sci. 66 (2009) 516–525.
[62] R. Ahmad, S. Gupta, C. Haldar, Age dependent expression of melatoninmembrane receptor (MT1, MT2) and its role in regulation of nitrosative stressin tropical rodent Funambulus pennanti, Free Radic. Res. 46 (2012) 194–203.
[63] C. Tomas-Zapico, A. Coto-Montes, A proposed mechanism to explain thestimulatory effect of melatonin on antioxidative enzymes, J. Pineal Res. 39(2005) 99–104.
[64] K.H. Jung, S.W. Hong, H.M. Zheng, H.S. Lee, H. Lee, D.H. Lee, S.Y. Lee, S.S. Hong,Melatonin ameliorates cerulein-induced pancreatitis by the modulation ofnuclear erythroid 2-related factor 2 and nuclear factor-kappaB in rats, J. PinealRes. 48 (2010) 239–250.
[65] R. Sharma, T. Ottenhof, P.A. Rzeczkowska, L.P. Niles, Epigenetic targets formelatonin: induction of histone H3 hyper-acetylation and gene expression inC17.2 neural stem cells, J. Pineal Res. 45 (2008) 277–284.
Copyright © 2022 FDOKUMEN




















