
Lymphocide: cytokines and the control of lymphoid homeostasis
Upload
independentCategory
view
3download
0
J. Comp. Path. 1991 Vol. 105
Pathogenesis of Lymphoid Lesions in Murine Experimental Listeriosis
A.J. Marco*, M. Domingo*, M. Prats*, V. Briones~, M. Pumarola* and L. Dominguez~
Dept. de Analom{a Patoldgica,* Facultad de Veterinaria, Universidad Autdnoma de Barcelona, 08193 (Bellaterra) Barcelona, Spain and Dept. de Microbiologia,t Facultad de Veterinaria,
Universidad Complutense de Madrid, 28040 Madrid, Spain
S u m m a r y
Adult female Swiss albino mice were infected intraperitoneally or subcuta- neously with Listeria monocytogenes Serovar 4b or 1/2a and killed at intervals. Thymus, spleen, Peyer's patches and a variety of lymph nodes, including the jejunal (mesenteric), mediastinal, lumbar, mandibular and superficial in- guinal, were examined by histopathology and by immunocytochemistry for detection of L. monocytogenes antigen. Similar results were obtained with both Serovars and by both routes of inoculation used. In the spleen, L. monocytogenes was detected, by immunoperoxidase staining, as soon as 4 h after inoculation, inside phagocytic cells located predominantly in the marginal zone of the white pulp. This was Followed by inflammation, necrosis and depletion of lymphoid cells, which extended in extreme cases to the whole organ. Inflam- matory lesions diminished progressively at 5 to 6 days after inoculation. In animals dying of" the infection, a severe necrotizing splenitis was present. Depletion of lymphoid cells and inflammatory changes were widespread in the lymph nodes and to a lesser extent in the Peyer's patches. An extensive necrotizing lymphadenitis was the prominent lesion in severely affected nodes. Inflammatory lesions and detection of L. monocytogenes antigen started around the venules of high endothelium. A thymus depletion, not associated with the multiplication of bacteria in the organ, was also a constant feature of the infection. This study suggests that L. monocytogenes (1) is transported to the spleen and to the lymph nodes by phagocytes, entering the organs by the marginal sinus in the spleen and by the venules of high endothelium in the lymph nodes; (2) multiplies in these cells as well as in neutrophilic granulo- cytes (the latter rapidly migrate to the affected zgnes); and (3) induce a splenitis and lymphadenitls, involving predominantly T cell-dependent areas, with a necrotizing component in severe cases.
From our observations it is concluded that infection of the lymphoid system is a major feature in the pathogenesis of murine listeriosis.
Introduction
Experimental infection of mice with Lisleria monocytogenes is a useful model for s tudying cell-mediated immuni ty against intracellular pathogens (Mackaness, 1962; North, 1975; Jungi and Jungi , 1981; Chen-Woan and McGregor, 1984;
Reprint requests to: Dr M, Domingo, Dept. Anatomla Patol6gica, Facultad de Veterinaria, Universidad Aut6noma de Barcelona, 08193 Bellaterra (Barcelona), Spain.
0021-9975/91/050001 + 15 $03.00/0 © 1991 Academic Press Limited

2 A.J. Marco e t al.
Kaufmann, 1987). A T cell response plays a fundamental role in controlling the multiplication of L. monocylogenes and the suppression of'infection (Kostiala and McGregor, 1975; North and Deissler, 1975; Jungi and McGregor, 1977; Ziegler and Unanue, 1981; Handa, Mitsuyama, Watanabe, Koga and Nomoto, 1987; Kaufmann, 1987). Immunosuppression also has long been recognized as being able to precipitate systemic human listeriosis (Fleming, Cochi, MacDonald, Brondum, Hayes, Plikaytis, Holmes, Audurier, Broome and Reingold, 1985; Ho, Shands, Friedland, Eckind and Fraser, 1986; McLauchlin, 1987), and several forms of experimental immunosuppression or overload of the mononuclear phagocytic system markedly reduce the LDa0 of L. monocytogenes for the mouse.
In spite of the widespread use of this model, little is known about the pathogenesis of murine experimental listeriosis. Inoculation of mice by several routes leads to a systemic infection, with invariable involvement of liver and spleen (Gray and Killinger, 1966; Mandel and Cheers, 1980; Domingo, Ramos, Dominguez, Ferrer and Marco, 1986; McLauchlin, 1987) and, less frequently, other organs such as the heart, lungs, brain and, in pregnant mice, the uterus (Gray and Killinger, 1966; Pohjanvirta and Huttunen, 1985). Detailed descriptions of lesions in lymphoid organs are scarce. Thymic cortical lymphoid depletion associated with increased adrenal corticosteroid secretion has been described in the early phase of the infection (Mandel and Cheers, 1980). Depletion of lymphoid cells and inflammation have also been docu- mented in the spleen, but the relationship of the lesions to the infection with L. monocytogenes remains to be clarified (Mandel and Cheers, 1980; Domingo el al., 1986).
We report a sequential study of infection with L. monocylogenes and present immunocytochemical evidence of the implication of the bacteria in the causation of the lesions.
Material and Methods
Bacterial Cultures
L. monocylogenes Serovar 4b (NCTC 10527) and Serovar 1/2a (NCTC 7973) used for inoculation were passaged twice through sucking mice and were then grown on BHI agar (Difco). Suspensions of bacteria were prepared in sterile 0'9 per cent saline and injected i.p. or s.c. The number of bacteria inoculated was estimated by viable counts of cultures in BHI agar of serial dilutions of the suspension. Previous experiments with L. monocjtogenes Serovar 4b showed that the median lethal dose 50 (LDs0) for S.wiss mice was 10 ~ and 4"2 x 1 0 7 bacteria by the i.p. and the s.c. route, respectively.
Plan of Experiment Forty female Swiss albino mice (Interfauna, Spain) weighing 18 to 20 g at the time of infection, were divided into two equal groups and inoculated with L. monocylogenes Serovar 4b. The first group was infected by the intraperitoneal (i.p.) route, by giving 1.2 x 106 live L. monocytogenes cells in 250 gl &saline to each mouse. Each mouse in the second group received 6.2x 107 live bacteria in 250gl of saline, inoculated subcutaneously (s.c.) in the lumbar area. An additional 40 mice were divided into two groups of 20 as in the previous experiment and one group was infected i.p. with L.

Lymphoid Lesions in Listeriosis
rnonocytogenes Serovar 1/2a (3"0 x l0 s bacteria) and the other group by the s.c. route (8"2 x 107 bacteria). As control animals, six uninoculated mice of the same age were employed. They were killed before the inoculation was performed and served as a reference image of the normal structure of lymphoid tissues. Mice were killed with ether at intervals beginning at 4 h post-infection (h.p.i.) and every 8 h until 36 h.p.i. Subsequently, mice were killed at 48 h.p.i., and then daily until the 6th day post-infection (d.p.i.). Two mice were killed at each time interval. Mice that died from the infection were added to the mice killed at the following time interval. The lymphatic drainage of the peritoneal cavity was explored by indian ink injection in additional mice and subsequent euthanasia, following the same time schedule (up to 36 h) as for the L. monocytogenes infected mice.
Histopathology and Immunocylochemistry
Mice were necropsied and samples of" thymus, spleen, lymph nodes, Peyer's patches and other non-lymphoid organs were fixed tbr 48 h in 10 per cent neutral buffered formalin and embedded in paraffin wax following routine procedures. After embedding, tissues were sectioned 5 to 6 gm thick and stained with haematoxylin and eosin (HE), or processed lbr immunoeytochemistry. A standardized necropsy technique directed to investigate jejunal (mesenteric), mediastinal, mandibular, lumbar and superficial inguinal lymph nodes was applied. The number of lymph nodes which were examined from the 20 mice of" the group infected with Serovar 4b ranged between 10 and 18 for each location. Similar figures apply to Serovar 1/2a.
The peroxidase-antiperoxidase (PAP) technique used has been previously described (Domingo el al., 1986). Briefly, serum against L. monocytogenes was produced in rabbits following several intravenous inoculations, at 10-day intervals, of 106 to 10 a live L. monocytogenes cells. Serum titre was measured 7 days after each immunization by plate micro-agglutination (Gaultney, Wende and Williams, 1971). Immunization was considered adequate when the titre was 1 in 10 240 for 1/2a and 1 in 20 480 for 4b. For immunocytochemistry, the antiserum was used at a dilution of 1 in 1000. As a control of the first antiserum, serum from a normal, non-immunized rabbit was used at the same dilution. Reaction of this serum with L. monocytogenes by plate micro-agglutination was negative.
R e s u l t s
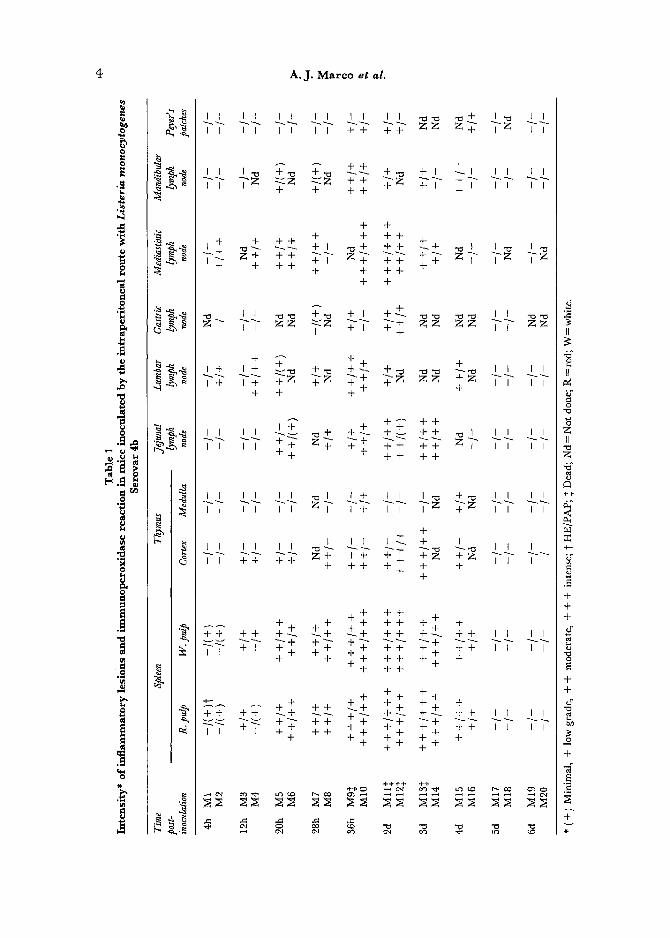
The presence and intensity of inf lammatory lesions and L. monocytogenes antigen after infection with Serovar 4b by the i.p. and s.c. routes are shown in Tables 1 and 2, respectively. Similar results, with only subtle differences, were observed by inoculation of Serovar 1/2a. Lesions in the liver in both experiments were similar to those reported by other authors (North, 1970; Mandel and Cheers, 1980; Wirsing von Koenig, Heymer , Hof and Finger, 1983; Domingo et al., 1986), and they will not be further described. The following descriptions, if not otherwise stated, refer to infection with L. monocytogenes Serovar 4b.
Histopathological and Immunocytochemical Findings in Mice Following Intraperitoneal Inoculation
Spleen. In the spleen, L. monocylogenes antigen was detected as early as 4 h.p.i. inside macrophages (M0) at the marginal zone by the PAP technique. At this time p.i., the immunoperoxidase (IP) reaction was localized to a few small bacillary forms in the cytoplasm of phagocytic cells. Lesions could not
4 A . J . M a r c o e t a l .
I.
@
]
] @
~.~
@
i
O
e~
I I I I I I I I I I I I I ~ ~ 1 ,, ,, ,, ,, ++ 77 ~ ~<+ Tz ,7
,, Tz <z ++ ~ +, + + + + +
+ + + + ++
~ + + + + j ~ + + + + + ~ I ~ ~ ~ ~ ~ . . . . ~ ~T T z ~ z I+ ~++ ++++ +I + ++
+ + ++ + +
I +I ++
+ ~ + +
, + , + ; - z ; - + ; ' ~ z z , , , , + + ++ +
J+ +~ +~ ÷+ i II II
++ ++ r II II
ii If ~ +
II il ++ ++ ++
f I I I I I ~ 1 I + I I I ~ + ~ I I I I I I ~ I i I z ~ i + I i ~ z ~ z t i i i
+ I II I~ + I
I I + + + + Z + + + + + I I I I + ++ ++ + + +
+ ++
++ ++ ++ ++ ++ ++ ++ I I I i .... +~ ++~ ~ ~+ ...... +t + + + ++ +~ +
+ ++
++ ++ --++ +÷- +÷ +_++ +++ ++_÷
+~ + ~+ ~+ T? +~ + ++ ++ ++ ++
+ ++ II II
++ II il +
÷÷ ++
II
b ~ o
Z II
m
.~=
+ +
F
+ +
] +
+ v
Lymphoid Lesions in Listeriosis 5
0
0
+ +++ +, +
[ + + L + ++ ++ +
+++ + +
"-< ~]- i, z, , I --~- 7z z yTz z z ]-z~
+
+ + -{-~- + <, ~, , I I I ~ ++~ +< +++ I
+ +~ + ~- +
+ +++ i I I I I I I I I +++~ + + +-b + +
I I I I + ++ ++ +++ + + +++
II I I I I III << '~ ~ ÷ ~ ~ +++
<< ~, , , ,, i+ II I II IL ++
+ +++
i +
+++ + ++÷
~ <÷+÷~ ++ ~, ~, , , ~, ÷+ ÷7+÷ +÷ ++÷
+++
÷+ ++ +~ ++
II
IL
Z II
Z
+÷
.=. + + +
o
+ +
o
+
+ v 4~

6 A.J. M a r c o e t al.
be observed at this early phase of invasion of the organ by the bacteria. Lesions, first detected at 12 h.p.i., appeared as depletions of the peri~arteriolar lymphoid sheaths (PALS), clearly observable as diminished nuclear staining, around the central vessels and extending to the external zones of the PALS. The marginal zone remained unchanged. Simultaneously, a moderate, multi- focal, mixed inflammatory infiltration of neutrophilic granulocytes (PMN) and M0, with some cells staining positively for anti-L, monocytogenes serum, became observable in the PALS, and, to a lesser extent, in the red pulp. L. monocytogenes antigen inside phagocytic cells was seen as diffuse or fine granular cytoplasmic deposits. Both depletion of lymphoid cells and inflammatory lesions, with many cells containing L. monocytogenes antigen, were well de- veloped by 28 h.p.i. (Figs 1 and 2). Inflammation progressively expanded to the white as well as to the red pulp. Karyorhexis in the zone around the central arterioles of the PALS was the first indication of necrosis (Fig. 3). In animals dying at 36 h.p.i., a severe diffuse necrotizing splenitis was observed. Similar lesions were observed at 48 h.p.i, in two animals (M11 and 12). Thrombosis of medium to large vessels was evident in these cases. In the following two days, both a necrotizing or an intense diffuse pyogranulomatous splenitis, devoid of necrosis, could be observed. The IP reaction in the spleen showed a tendency to be more localized and of lower intensity than in previous cases. Necrotizing inflammation was associated with spontaneous death or with animals killed when moribund, whereas pyogranulomatous reactions normally were observed in animals appearing healthy at the time of killing. From 4 d.p.i, onwards, lesions clearly diminished in intensity, and had disappeared by 6 d.p.i. At this interval, the residual inflammatory lesions gave way to an active multiplication oflymphoblastoid cells in the white pulp, as well as to formation of prominent secondary fbllicles. An increased number of mitotic figures per high magnifica- tion field was associated with this regenerative activity.
Lymph Nodes. In most lymph nodes studied, lesions consisted of a depletion of" lymphoid cells in the paracortical zone, observed in some of them as early as 4 or 12h.p.i. This was followed by a mixed inflammatory reaction in the interfollicular cortex and paracortex (20 h.p.i.), with PMN and M0 staining positively by the IP method. Lesions were oriented around high endothelium venules (HEV), and L. monocylogenes was first detected in phagocytic cells close to these endothelial cells (Fig. 4).
At 36 and 48 h.p.i., a necrotizing lymphadenitis, more severe in animals that died, was the prominent feature. This pattern of lesions was observed for all the lymph nodes studied, except for the mediastinal and jejunal (mesenteric) lymph nodes. In the mediastinal node, inflammatory lesions and specific IP reaction first appeared related to the subcapsular and medullary sinuses (Fig. 7). As this is consistent with an afferent spreading of the infection from the peritoneal cavity to the node, the lymphatic drainage of the peritoneal cavity was explored by indian ink injection in mice and subsequent necropsy. Indian ink was visualized as soon as 4 h.p.i, in the subcapsular sinus of the media~tinic lymph node (Fig. 8), but could not be detected at any time in the jejunal or in the lumbar lymph nodes.
Lymphoid Lesions in Listeriosis 7
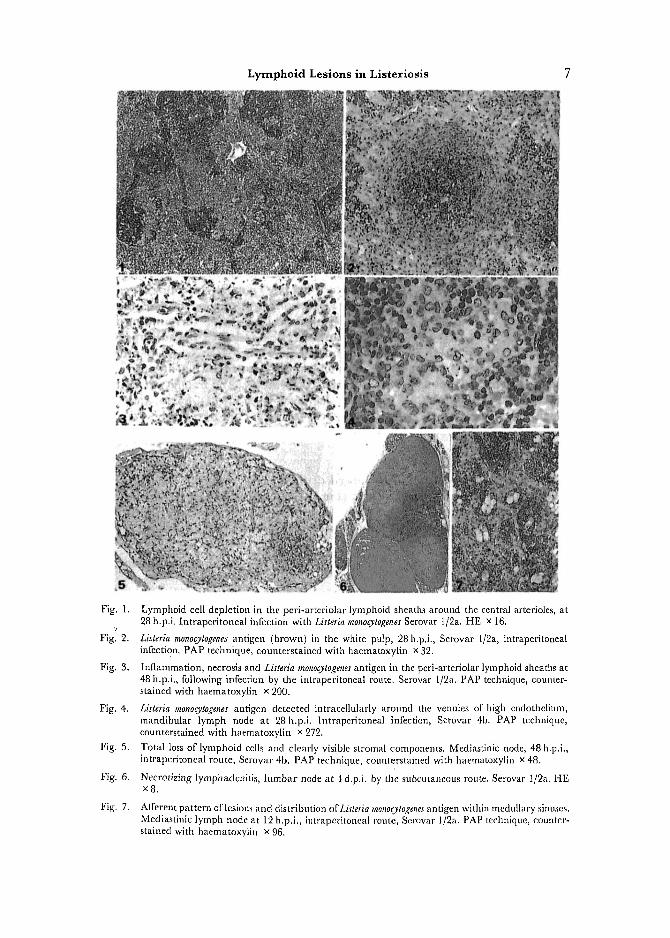
Fig. 1.
Figl 2.
Fig. 3.
Fig. 4.
Fig. 5.
Fig. 6.
Fig. 7.
Lymphoid cell depletion in the peri-arteriolar lymphoid sheaths around the central arterioles, at 28 h.p.i. [ntraperitoneal infection with Listeria monocytogenes Serovar 1/2a. HE × 16.
Listeria monocytogenes antigen (brown) in the white pulp, 28 h.p.i., Serovar l/2a, intraperitonea[ infection. PAP technique, counterstained with haematoxylin x 32.
Inflammation, necrosis and Listeria rnonocytogenes antigen in the peri-arteriolar lymphoid sheaths at 48 h.p.i., following infection by the intraperitoneal route. Serovar l/2a. PAP technique, counter- stained with haematoxylin x 200.
Listeria monocytogenes antigen detected intracellularly around the venules of high endothellum, mandibular lymph node at 28 h.p.i, lntraperitoneal infection, Serovar 4b. PAP technique, counterstained with haematoxylin x 272.
Total loss of lymphoid cells and clearly visible stromal components. Mediastinic node, 48 h.p.i., intraperitoneal route, Serowtr 4b. PAP technique, counterstained with haematoxylin x 48.
Necrotiziug lymphadcnitis, lumbar node at 4 d.p.i, by the suheutaneous route. Serovar 1/2a. HE xg.
Afferent pattern oflesion.s and distribution ofListeria monocytogenes antigen within medullary sinuses. Mediastinic lymph node at 12 h.p.i., intraperitoneal route, Serovar 1/2a. PAP technique, counter- stained with haematoxylin x 96.

8 A.J. Marco e t al.
Sporadically, the jejunal lymph node also showed a similar afferent pattern, but it was always of minor importance compared to the paracortical and interfollicular cortical involvement. The afferent infection in this node was interpreted as a result of inflammatory lesions in the wall of the organs of the abdominal cavity, particularly of the gastrointestinal t ract
Severely affected mediastinal lymph nodes appeared empty, apparently due to a massive destruction and/or depletion of lymphoid cells. The stromal components of the organ were clearly visible in these cases (Fig. 5).
Almost complete resolution occurred by 6 d.p.i. As in the spleen, secondary lymphoid follicles were prominent in the last two days of the experiment.
Thymus. A cortical lymphoid depletion was present in the thymus by 12 h.p.i., and was more marked by the 2nd and 3rd d.p.i. (Fig. 9). I n a few cases, a discrete focal pericapsular inflammatory reaction, with phagocytic cells show- ing cytoplasmic L. monocytogenes antigen appeared by 36 h.p.i, surrounding the organ. From the thymic capsule, inflammation extended to the subcapsular cortical parenchyme, as foci of PMN and M0 showing L. monocytogenes antigen in their cytoplasm. Thymic pericapsular lesions were similar in intensity to those observed in the mediastinal serosa. Nevertheless, the magnitude of the inflammatory changes was low. The degree of cellular depletion and the magnitude of the inflammatory lesions were markedly reduced on the 4 d.p.i., and on the 5 and 6 d.p.i., the thymus exhibited a normal appearance, with a well-developed cortical zone.
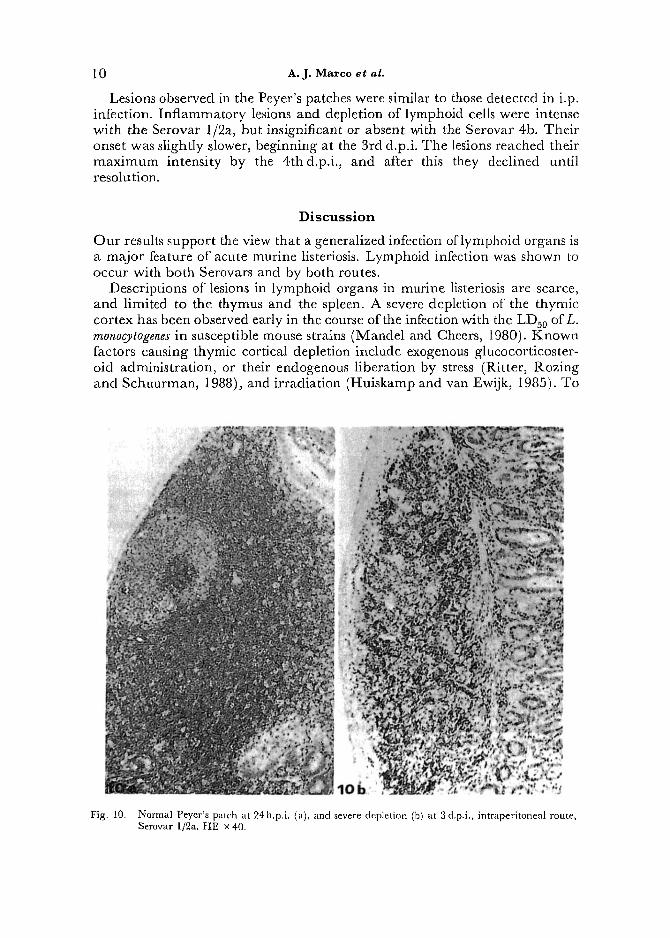
Peyer's Patches. Peyer's patches showed depletion of lymphoid cells following injection of either Serovar of Listeria (Fig. 10); however, inflammatory lesions with IP positive reaction were only significant in mice infected with the Serovar 1/2a. Inflammatory changes reached their peak at 36 h.p.i.
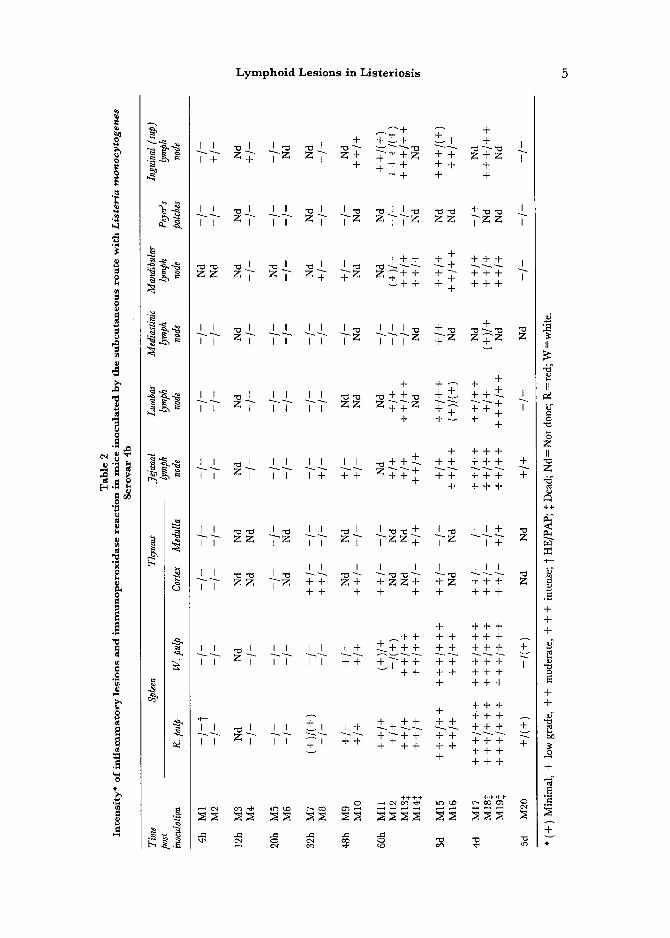
Histopathological and Immunocytoehemical Findings in Mice Following Subcutaneous Inoculation
The lesions observed after s.c. inoculation were similar in character to those observed after i.p. infection, but differed in distribution and intensity, and were also slightly slower in onset (Table 2). In the spleen, after initial depletion, a well-developed necrotizing or pyogranulomatous splenitis was observed at 3 d.p.i. Severe necrotizing lesions were predominantly recorded in animals that died spontaneously. Fibrinous thrombi were particularly evident inside splenic vessels in these animals.
There was an absence of pericapsular inflammatory lesions in the thymus. Depletion of the thymic cortex was first observed by 32 h.p.i and followed a similar course to the i.p. infection. Inflammation and loci of necrosis were also
L y m p h o i d L e s i o n s i n L i s t e r i o s i s
8
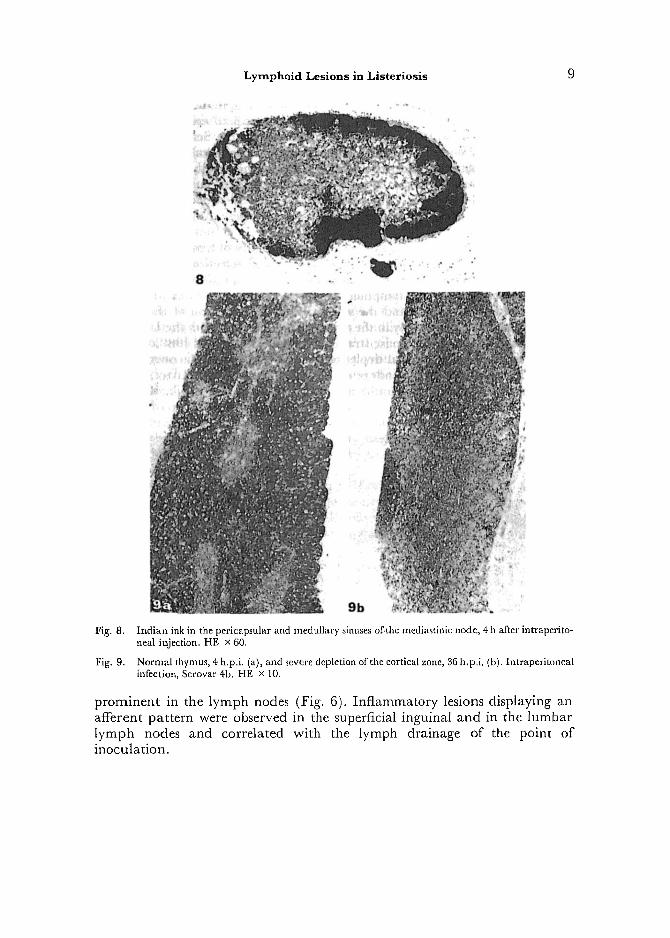
Fig. 8.
Fig, 9.
Indian ink in the pericapsular and medullary sinuses of.the mediastinic node, 4 h after intraperito- neal injection. HE x 60.
Normal thymus, 4 h.p.i. (a), and severe depletion of the cortical zone, 36 h.p,i. (b). Intraperitoneal infection, Serovar 4-b. HE x 10.
prominent in the lymph nodes (Fig. 6). Inflammatory lesions displaying an afferent pattern were observed in the superficial inguinal and in the lumbar lymph nodes and correlated with the lymph drainage of the point of inoculation.
1 0 A . J . M a r c o at al.
Lesions observed in the Peyer's patches were similar to those detected in i.p. infection. Inflammatory lesions and depletion of lymphoid cells were intense with the Serovar 1/2a, but insignificant or absent with the Serovar 4b. Their onset was slightly slower, beginning at the 3rd d.p.i. The lesions reached their maximum intensity by the 4thd.p.i,, and after this they declined until resolution.
D i s c u s s i o n
Our results support the view that a generalized infection of lymphoid organs is a major feature of acute murine listeriosis. Lymphoid infection was shown to occur with both Serovars and by both routes.
Descriptions of lesions in lymphoid organs in murine listeriosis are scarce, and limited to the thymus and the spleen. A severe depletion of the thymic cortex has been observed early in the course of the infection with the LDs0 of L. rnonocytogenes in susceptible mouse strains (Mandel and Cheers, 1980). Known factors causing thymic cortical depletion include exogenous glucocorticoster- oid administration, or their endogenous liberation by stress (Ritter, Rozing and Schuurman, 1988), and irradiation (Huiskamp and van Ewijk, 1985). To
Fig. 10. Normal Peyer's patch at 24 h.p.i. (a), and severe depletion (b) at 3 d.p.i., intraperitoneal route, Serovar 1]2a. HE x 40.

Lymphoid Lesions in Listeriosis [ 1
elucidate the role of glucocorticosteroid secretion in the production of thymic cortical lymphoid cell depletion, experimental infections with L. monoc_ytogenes in adrenalectomized mice or in Listeria-resistant mouse strains have been carried out (Mandel and Cheers, 1980). It was observed that adrenalectomy prevented the depletion caused by infection with L. monocytogenes, and deple- tion was not detected after infection in Listeria-resistant mouse strains (Mandel and Cheers, 1980). We have observed that subclinical, non-lethal infection in susceptible strains, demonstrated by the presence of necrosis and pyogranulo- matous inflammation with IP reaction in the PALS, failed to produce ~h~,mus depletion (unpublished results).
Thymic cortical depletion was also a prominent lesion in our experiments, observed following i.p. as well as s.c. infection, confirming previous reports. These observations lead to the conclusion that L. monocytogenes might cause cortical depletion of the thymus in an indirect way, mediated by the secretion of adrenal glucocorticosteroid hormones resulting from the acute stress of the infection, which produces lysis of cortical thymocytes. We could not detect L. monocytogenes antigen in severe atrophic thymuses of mice infected by the s.c. route, which further supports the exclusion of the bacteria as a direct cause of the lesion.
The thymic cortex was of normal appearance at 5 and 6 d.p.i., a time which is comparable to the time needed for regeneration of cortical thymocytes after sublethal irradiation (Huiskamp and vail Ewijk, 1985) or glucocorticosteroid treatment (van Ewijk, van Soest and van den Engh, 1981; Chan and Cheers, 1982b).
A depletion of lymphoid cells in the PALS and a concomitant inflammatory reaction in the form of micro-abscesses have been observed in the spleen; however, the pathogenesis of spleen lesions is unknown (Mandel and Cheers, 1980; Domingo et al., 1986). Available data indicate that the mechanisms underlying lymphoid depletion in the PALS are different from those causing thymus depletion. In contrast to the thymus, depletion in the PALS did not appear to be influenced by adrenal glucocorticosteroid secretion. Infection of both susceptible and resistant strains of mice with the LDs0 for susceptible strains, can induce depletion of the PALS (Mandel and Cheers, 1980). We have also observed that L. monocytogenes at sublethal doses can also produce inflammation and lymphoid depletion in the PALS in susceptible strains (unpublished results). However, inoculation of a large number of apathogenic Listeria or killed L. monocytogenes cells was unable to induce lesions in lymphoid organs (Chan and Cheers, 1982a). We believe that the actual presence of L. monocSogenes and the subsequent inflammatory reaction is a major cause of destruction of lymphocytes and depletion of the PALS. This is supported by the early detection, by the PAP technique, of L. monocylogenes located intracel- lularly in cells resembling maerophages in the white pulp of the spleen. This is followed by an influx of PMN and, shortly thereafter, by karyorhexis and necrosis in the PALS, suggesting that these lesions are associated with tile breakdown of inflammatory cells, mainly PMN, and the resultant liberation of hydrolytic enzymes. Such a mechanism could magnify the inflammatory lesions, despite the low numbers of L. monocytogenes demonstrated in them. The

12 A.J. Marco e t al .
extensive necrotic zones that then occur could, therefore, be a consequence of the extension of inflammation in the PALS and in the red pulp. Widespread thrombosis of medium size vessels has been observed by us, and it probably represents an additional cause of necrosis. An almost total necrosis of the spleen was a common finding in animals that succumbed to the infection, suggesting that the magnitude of the necrotizing splenitis is an indication of the severity of the disease. In contrast to this, pyogranulomatous inflammatory reactions appeared to represent an initial phase of repair, which was characterized by numerous activated macrophages and PMN, infiltrating the organ in a diffuse form. In parallel with this, the amount ofL. mono~togenes antigen, as shown by IP, decreases in these lesions, which can be interpreted as an effective elimination of bacteria by macrophages. This view is supported by observa- tions based on serial counts of L. monocytogenes and M0 in the spleen of Listeria- infected mice (Mackaness, 1962). In these experiments it was shown that an inverse relationship exists between the counts of bacteria in the spleen and the number of M0 entering the organ, from approximately 3 or 4 d.p.i.
Signs of repair are also co-incident with the establishment of a specific immunological response against L. monocytogenes (Mackaness, 1962; Mandel and Cheers, 1980; Sadarangani, Skamene and Kongshavn, 1980).
To our knowledge, lesions in lymph nodes in murine experimental listeriosis have not been previously described. This fact is surprising, if it is thought that Listeria are intracellular pathogenic bacteria, and that systemic lymph node involvement occurs in septicaemic listeriosis in other species (Gray and Killinger, 1966). In our experiments, both a granulomatous-necrotizing lym- phadenitis and lymphoid depletion were significant lesions in murine listeriosis.
From our observations, the spread ofL. rnonocylogenes to the lymphoid organs can be depicted as follows: (1) Initial dissemination of the bacteria to the lymph nodes occurs via the lymphatic drainage. Depending on the point of inoculation, the mediastinal lymph nodes (draining the peritoneal cavity, as shown by indian ink resorption experiments) and the superficial inguinal lymph nodes (draining the lumbar point of inoculation) are rapidly affected in the course of infection, showing an afferent pattern of distribution of inflam- matory cells and antigens. (2) Soon after, bacteria-laden macrophages transfer the infection haematogenously to the spleen and lymph nodes and, with the Serovar 1/2a, also the Peyer's patches. Entry of bacteria to these organs is through the marginal sinus in the spleen and through the HEV in the other lymphoid organs, and (3) the presence of bacteria in these locations triggers an inflammatory reaction, which severely damages the PALS and the paracortical and cortical zones in the lymph nodes, causing depletion and necrosis.
The dissemination of L. monocytogenes to the lymphoid organs probably indicates the normal pathways of migration of phagocytes to these organs. Macrophages are demonstrated in low numbers in the T cell dependent areas of the spleen of normal animals; however, their number increases significantly after administration of Bacillus of Calmette-Guerin (Hume, Robinson, Macpher- son and Gordon, 1983). A similar response could explain the selective localization of L. monocytogenes antigen in these zones. The point of entry of circulating macrophages to the lymph nodes is not clearly established. Our

Lymphoid Lesions in Listeriosls 13
observations show that Lisleria-laden phagocytes are first detected around and in the lumen of the HEV. Recently, receptors equivalent to the "lymphoid homing" receptors of lymphocytes have been demonstrated in macrophages and dendritic cells, supporting a receptor-mediated entry in the lymph nodes through the HEV (Pals, Horst, Scheper and Meijer, 1989).
Our observations point to L. monocytogenes as the direct cause of the lesions in the spleen, lymph nodes and Peyer's patches, whereas for the thymus, a glucocorticosteroid-mediated lysis of thymocytes could be the responsible mechanism. However, other factors such as a decrease in the circulating pool of lymphoid cells or modification of its normal migration pattern to the lymphoid organs (Chung, Samlowski and Daynes, 1986) could also be partly responsible for the observed changes.
We have shown that the interaction of L. monocylogenes with the lymphoid organs is a complex phenomenon, leading to a selective destruction/depletion of lymphoid cells, particularly in the T cell-dependent areas. The implication of this selective damage to T cells for the development of an effective immune response remains to be established. In fact, a deficient T cell response to mitogens has been observed shortly after Listeria infection in mice (Brunner and Kroll, 1989).
Whatever the exact role of the different factors implicated, it is concluded that, in the course of the infection by L. monocSogenes , significant damage to the lymphoid organs occurs. Pathological lesions in recent human cases of acute lethal septicaemic infection with L. monocytogenes (Fleming el al., 1985; Ho el al., 1986) have not yet been described. Lymphoid lesions have been observed by us in mice in the course of a septicaemic in~i~ction, and therefore, we suggest that similar lymphoid lesions could be expected to occur in human cases of acute listeriosis.
The lymphoid system of mice in~cted with L. monocylogenes is profoundly altered, and this fact should be taken into consideration when studying the immune response to Listeria and the factors affecting or modifying it.
Acknowledgments
The authors thank Francisco Perez and Pedro Losada for technical assistance. This work was supported by the Comisi6n Interdepartamental de Ciencia y Tecnologia (CYCIT), PB88-0231.
References
Brunner, H. and Kroll, H. P. (1989). Reduced proliferative response of mouse spleen cells to mitogens during infection with Salmonella typhimurium or Listeria monocytogenes. Microbial Pathogenesis, 6, 265-276.
Chan, Yu Yu and Cheers, C. (1982a). Mechanism of depletion ofT lymphocytes from the spleen of mice infbcted with Listeria monocytogenes. Infection and Immunity, 38, 686-693.
Chan, Yu Yu and Cheers, C. (1982b). Recovery ~?om T cell depletion during murine listeriosis and effects on a T-dependent antibody response. Infection and Immunity, 38, 694-698.
Chung, H.T., Samlowski, W.E. and Daynes, R.A. (1986). Modification of the murine immune system by glucocorticosteroids: alteration of" the tissue

14 A.J. Marco e t al.
localization properties of circulating lymphocytes. Cellular Immunology, 101, 571-585.
Domingo, M., Ramos, J.A., Dominguez, L., Ferret, L. and Marco, A. (1986). Demonstration of Listeria monoo, togenes with the PAP technique in formalin fixed and paraffin embedded tissues of experimentally infected mice. Journal of Veterinary Medicine, 33, 537-542.
Fleming, D. W., Coehi, S. L., MacDonald, K. L., Brondum, J., Hayes, P. S., Plikaytis, B.D., Holmes, M.B., Audurier, A., Broome, C.V. and Reingold, A. L. (1985). Pasteurized milk as a vehicle of infection in an outbreak of listeriosis. New England Journal of Medicine, 312, 404-407.
Gaultney, J.D., Wende, R.D. and Williams, R.P. (1971). Microagglutination procedures for febrile agglutination test. Applied Microbiology, 22, 635-640.
Gray, M. L. and Killinger, A. H. (1966). Listeria monocytogenes and listerie infections. Bacteriological Review, 30, 309-382.
Handa, T., Mitsuyama, M., Watanabe, Y., Koga, T. and Nomoto, K. (1987). A significant role of the macrophage accumulation induced by MCF in the protection of mice against Listeria monocytogenes in vivo. Cellular hnmunology, 106, 330-342.
Ho, J.J., Shands, K.N., Friedland, G., Eckind, P. and Fraser, D.W. (1986). An outbreak of type 4b Listeria monocylogenes infection involving patients li'om eight Boston hospitals. Archives of Internal Medicine, 146, 520-524.
Huiskamp, R. and van Ewijk, W. (1985). Repopulatlon of the mouse thynaus aller sublethal fission neutron irradiation. I. Sequential appearance of thymocyte subpopulations. Journal of Immunology, 134, 2 l 61-2169.
Hume, D.A., Robinson, A.P., Macpherson, G.G. and Gordon, S. (1983). The mononuclear phagocyte system of the rnouse defined by immunohistochemical localization of antigen F4/80. Relationship between macrophages, Langerhans cells, reticular cells, and dendritic ceils in lymphoid and hematopoietic organs. Journal of Experimental Medicine, 158, 1522-1536.
Jungi, T. W. and Jungi, R. (1981). Immunological memory to Listeria monocytogenes in rodents. IV. Studies on origin and fate of tissue positioned T memory cells. Immunology, 44, 789-798.
Jungi, T.W. and McGregor, D.D. (1977). Generation of macrophage chemotactic activity in situ in Listeria-immune rats. CelMar Immunology, 33, 322-339.
Kaufmann, S. H. E. (1987). Possible role of helper and cytolytic T lymphocytes in antibacterial defense: conclusions based on a murine model oflisteriosis. Review oJ Infectious Diseases, 9, 5650-5659.
Kostiala, A. A. I. and McGregor, D.D. (1975). The mediator of cellular immunity: IX. The ~ relationship between cellular hypersensitivity and acquired cellular resistance in rats infected with Listeria mon ocytogenes. Journal of Experimenlal Medicine, 141, 1249-1260.
Mackaness, G.G. (1962). Cellular resistance to infection. Journal of Experimental Medicine, 116, 381-406.
Mandel, T. E. and Cheers, C. (1980). Resistance and susceptibility of mice to bacterial infection: histopathology of listeriosis in resistant and susceptible strains. Infection and Immunity, 30, 851-861.
McLauchlin, J. (1987). Listeria monocytogenes: recent advances in the taxonomy and epidemiology of listeriosis in humans. Journal of Applied Bacteriology, 63, 1-11.
North, R.J. (1970). The relative importance of blood rnonocytes and fixed macrophages to the expression of cell-mediated immunity to infection. Journal Oat Experimental Medicine, 132, 521-534.
North, R.J. (1975). Nature of"memory" in T-cell-mediated antibacterial immunity: anamnestic production of mediator T cells. Infection and Immunity, 12, 754-760.
North, R.J. and Deissler, J .F. (1975). Nature of "memory" in T-cell-mediated antibacterial immunity: cellular parameters that distinguish between the active immune response and a state of "memory". Infection and Immunity, 12, 761-767.

Lymphoid Lesions in Listeriosis 15
Pals, S.T., Horst, E., Scheper, R.J. and Meijer, C.J .L.M. (1989). Mechanisms of human lymphocyte migration and their role in the pathogenesis of disease. Immunological Reviews, 108, 111-133.
Pohjanvirta, R. and Huttunen, T. (1985). Some aspects of murine experimental listeriosis. Acta Veterinaria Scandinavica, 26, 563-580.
Ritter, M.A., Rozing, J. and Schuurman, H.J. (1988). The true function of the thymus? Immunology Today, 9, 189-193.
Sadarangani, C., Skamene, E. and Kongshavn, P. A. L. (1980). Cellular basis for genetically determined enhanced resistance of certain mouse strains to listeriosis. Infection and Immunity, 28, 381-386.
Van Ewijk, W., van Soest, P. L. and van den Engh, G.J. (1981). Fluorescence analysis and anatomic distribution of mouse T lymphocyte subsets defined by monoclonal antibodies to the antigens Thy- 1, Lyt- 1, Lyt-2, and T-200. Journal of Immunology, 127, 2594-2604.
Wirsing yon Koenig, C.H., Heymer, B., Hof; H. and Finger, H. (i983). Course of infection and development of immunity in experimental infection of mice with Listeria serotypes. Infection and Immunity, 40, 1170-1177.
Chen-Woan, M. and McGregor, D. D. (1984). The mediators of acquired resistance to Listeria mono~ytogenes are contained within a population ofcytotoxic T cells. Cellular Immunology, 87, 538-545.
Ziegler, K. and Unanue, E.R. (1981). Identification of a macrophage antigen-processing event required for I region-restricted antigen presentation to T lymphocytes. Journal oj" Immunology, 127, 1869-1875.
E Received, August 24th, 1990 7 Accepted, October 31st, 1990]
Copyright © 2022 FDOKUMEN




















