Hysterectomy and uterine artery embolisation for the treatment ...
Upload
independentCategory
view
0download
0
Journal of Leukocyte Biology Volume 61, April 1997 427
Unique CD8� T cell-rich lymphoid aggregates inhuman uterine endometrium
Grant R. Yeaman,* Paul M. Guyre,t Michael W. Fanger,* Jane E. Collins,* Hillary D. White,*Wayne Rathbun� Kenneth A. Orndorff,t Jorge Gonzalez,t Judy E. Stern,� andCharles R. Wirat*Depajrtment� of Microbiology, t Physiology, 1 Pathology, and § Obstetrics & Gynecology, Dartmouth Medical School,
Lebanon, New Hampshire
Abstract: Using confocal scanning laser microscopy of
viable tissue sections, we have demonstrated organized
lymphoid aggregates (LA), that have a unique struc-
hire, in the stratum basalis of uterine endometrium.
These LA consist of a core of B cells surrounded by
more numerous T cells and an outer halo of monocytes/
macrophages. The T cells in the LA were almost ex-
clusively CD8’CD4 . These CD8� LA, in terms of
both their T cell and B cell components, were either
small or absent during the early proliferative stage
of the menstrual cycle, significantly larger in size at
mid-cycle and during the secretory phase, and absent
in post-menopausal women, suggesting that their de-
velopment is hormonally influenced. This new finding
ofa menstrual cycle-dependent, phenotypically unique,
organized immune cell structure may lead to new in-
sights into the mechanisms by which the endometrium
accepts a semiallogeneic graft while providing resis-
tance to infectious organisms. J. Leukoc. Rio!. 61:
427-435; 1997.
Key Words: repmductive immunology . mucosa � Tlymphocytes.
monocytes/macrophages . confocal scanning Inser micmscopy
INTRODUCTION
The mucosal immune system within the female reproduc-
tive tract is of particular importance because it is the first
site of immunological contact with many potentially patho-
genic bacteria and viruses. To be compatible with the repro-
ductive functions of these tissues, a local immune response
must protect against an infectious challenge, while at the
same time not mount a deleterious response to allogeneic
spermatozoa or an immunologically foreign fetal placental
unit [1]. The human non-pregnant uterus contains myeloid
and lymphoid cells that have the potential to function in
ways similar to their counterparts at other mucosal surfaces
and in the peripheral blood [2-7]. However, the distri-
bution of these cells within the female reproductive tract
appears to be uniquely regulated by estradiol and proges-
terone [8, 9]. Endometrial macrophages are distributed
diffusely throughout the uterine stroma and are also found
in small lymphoid aggregates that appear to increase in
number during the late secretory phase [3, 4, 7]. Macro-
phage numbers are surprisingly high, accounting for 5-
15% of the endometrial stromal cells [3, 5, 10]. A natural
killer (NK) cell population, phenotypically distinct from pe-
ripheral blood NK cells in that it is CD56 bright/CD16 dim,
is present in the uterine endometrium in large numbers
and increases markedly during the secretory phase and
during the first trimester of pregnancy [11-15; reviewed in
ref. 7]. The precise function of these cells within the fe-
male reproductive tract remains to be established. T cells
are found throughout the endometrium as isolated stromal
cells, intraepithelial lymphocytes, and in discrete lymphoid
follicles [2, 3, 6, 7, 10, 13, 16-18]. Our own work [Givan
et a!., unpublished resuitsi and that of others, show B cells
to be present in low but measurable numbers in the human
uterus [3, 7, 15, 19, 20]. The number of T cells relative
to other lymphoid cells does not increase significantly dur-
ing the menstrual cycle. However, the relative distribution
of T cells does change over the course of the menstrual cy-
cle. Discrete lymphoid follicles, or lymphoid aggregates, in
the uterine endometrium have long been recognized by his-
topathologists [reviewed in ref. 2] and were thought to oc-
cur in response to infection. More recently, Tabibzadeh
and others have presented evidence that these structures
develop during the menstrual cycle [17] and are composed
predominantly of T cells and some macrophages. It should
be noted that these structures are distinct from the NK cell
aggregates described by a number of authors [reviewed in
ref. 7J. To date, most immunophenotypic studies of human
uterine tissue have used single-color immunohistochemi-
cal staining of sequential frozen sections and have resulted
in little information about the extent of T cell aggregates
or the spatial relationships of the various cell types that
comprise them.
Abbreviations: LA, lymphoid aggregates; CSLM, confocal scanning
laser microscopy; NK, natural killer; PBS, phosphate-buffered saline;
Flit, fluorescein isothiocyanate; BSA, bovine serum albumin; PR Peyer’s
patches.
Correspondence: Dr. Grant R. Yeaman, Department of Microbiology,
Dartmouth Medical School, 1 Medical Center Drive, Lebanon, NH
03756.
Received November 6, 1996; revised January 6, 1997; accepted Jan-
uary 7, 1997.

428 Journal of Leukocyte Biology Volume 61, April 1997
In this study we report the cellular composition and dy-
namic changes in lymphoid aggregates (LA) located in the
stratum basalis of the human uterus throughout the men-
strual cycle by employing three-color immunofluorescent
staining of 30- to 70-�.tm-thick sections of freshly obtained
viable tissue analyzed by confocal scanning laser micros-
copy (CSLM). This approach permitted three-dimensional
modeling of these LA. Our results identify a unique popu-
lation of LA with a highly organized structure that includes
a B cell core surrounded by more numerous CD3+ T cells
and an outer mantle of macrophages. The size of these LA
fluctuated with the phase of the menstrual cycle, and with
few exceptions, the T cells in the aggregates were exclu-
sively CD8 �.
MATERIALS AND METHODS
Preparation of vibratome sections
Uterine endometrial tissue was obtained (with Institutional Review Board
approval) with informed consent from patients who had previously been
scheduled to undergo hysterectomy. For LA analysis, uterine endome-
trium were obtained from 9 postmenopausal women (average age 56,
range 42 to 70) and 23 cycling premenopausal women (average age 39.
range 25 to 52). One other patient included in the study was premeno-
pausal but not cycling (no. 707). This patient population is detailed in
Table 1 . Of the patients included in the study. none had a post-operative
diagnosis of malignant uterine disease. One patient (no. 445) had ovarian
cancer that had not spread to the Fallopian tube or the uterine endo-
metrium and two women had cervical carcinoma (nos. 707 and 724).
Single 150- to 300-mg samples of tissue were dissected out from equiv-
alent sites of the fundus in each uterus, distal to any gross pathology,
and placed immediately in sterile ice-cold phosphate-buffered saline
(PBS). The blocks of tissue were trimmed of excess myometrium, and
30- to 70-�.tm sections were cut using a vibratome (Vi000, Technical
Products International Inc. , St. Louis, MO). Sections were maintained
in ice-cold PBS throughout processing to prevent internalization of sur-
face markers. The stage of the menstrual cycle of the endometria was
determined in accordance with accepted histological practice using
hemotoxylin/eosin-stained paraffin sections. More precise evaluations of
proliferative phase endometria were carried out independently by two
pathologists who scored the degree of stromal edema and the relative
frequency of glandular and stromal mitoses; samples were classified as
early, middle, or late proliferative phase 1211. Secretory phase endo-
metria were not sub-staged.
Monoclonal antibodies
A panel of monoclonal antibodies, listed in Table 2, was used for direct
and indirect immunofluorescent staining: antibodies purified from hy-
bridoma cell culture supernatants (cell lines from American Type Cul-
ture collection, Rockville, MD) using HiTrap protein G superose col-
umns (Pharmacia Biotech Inc. , Piscataway, NJ) were labeled, where
indicated, with Cy3 or Cy5 Fluorolink protein labeling kits (Amersham
Life Sciences, Arlington Heights, IL) according to the manufacturer’s
TABLE 1. Summary of the Patient Population
Patient no. Age Uterine menstrual stage Uterine pathology Diagnosis
707 35 premenopausal (inactive) cervical cancer, HPV
477
358
380
445
502
522
526
578
627
60
57
56
57
53
42
70
45
63
postmenopausal
postmenopausal
postmenopausal
postmenopausal
postmenopausal
postmenopausal
postmenopausal
postmenopausal
postmenopausal
polyp
leiomyomata/adenomyoma
leiomyomata
leiomyomata
leiomyomata
leiomyomata/adenomyoma
benign
benign
benign
ovarian cancer
endometriosis
benign
other-appendicitis
endometriosis
benign
368
373
382
424
470
475
506
5 1 5
529
534
704
724
728
736
775
41
37
25
47
41
52
44
41
29
31
3 1
35
41
38
37
proliferative
proliferative
proliferative
proliferative
proliferative
proliferative
proliferative
proliferative
proliferative
proliferative
proliferative
proliferative
proliferative
proliferative
proliferative
leiomyomata
leiomyomata
hyalin sclerosis myometrium
leiomyomata
leiomyomata/adenomyoma
adenomyosis
leiomyomata
adenomyosis
leiomyomata
benign
benign
benign
benign
benign
benign
benign
benign
benign-inflammation
benign
endometriosis
cervical cancer
benign
benign
benign
387
446
484
563
564
678
697
698
37
40
48
39
44
3 1
41
43
secretory
secretory
secretory
secretory
secretory
secretory
secretory
secretory
leiomyomata
leiomyomata
leiomyomata
leiomyomata
leiomyomata/adenomyoma
inflammation
adenomyoma
uterus - premalignant
benign
benign
benign
benign
benign - inflammation
benign
benign

aCalt� Laboratories. San Francisco, CA.
b Antibodies were purified from cell culture supernatants of ATCC cell lines lAmerican Type Culture collection. Rockville, MD) and labeled with Cy3 or Cy5.
( Exalpha Corp. . Boston. MA.
‘1AMAC Inc.. Westbrook, ME.
�DAKO Corp.. Carpinteria. CA.
TABLE 2. Monoclonal Antibodies Used for Immunophenotyping
Yeaman et al. Unique human uterine lyniphoid aggregates 429
CD no.
CD3
CD3
CD4
CD4
CD8
CD8
CD14
CD14
CD I 9
CDI9
CD37
CD56
CD66b
Clone
�4.1
OKT3
S3.5
OKT4
3B5
OKT8
TUK 4
AML-223
SJ25-Ci
FMC63
WR17
562
531
Ber EP4
F8i 7
IVA12
P3
Supplier
Caltag#{176}
ATCCb
Caltag
ATCC
Caltag
ATCC
Caltag
ATCC
Caltag
ATCC
ATCC
Exalphac
AMACd
DAKOC
DAKO
ATCC
ATCC
Specificity
Pan T cell
Pan T cell
T helper subset
T helper subset
T suppressor/cytotoxic
T suppressor/cytotoxic
Monocyte/macrophage
Monocyte/macrophage
B cells
B cells
B cells
NK cell
Granulocytes
Epithelial cells
Pan HLA class II
Pan HLA class II
Mouse IgG1 isotype
Fluorochrome
FITC
Cy3, Cy5
FITC
Cy3, Cy5
FITC
Cy3, Cy5
FITC
Cy3, Cy5
FITC
Cy5
Cy5
FITC
FITC
FITC
FITC
Cy5
FITC, Cy3, Cy5
recommendations. Fluorescein isothiocyanate (FITC) -conjugated mouse
monoclonal antibodies were obtained from commercial suppliers as de-
noted in Table 2.
Three-color immunophenotyping
Three-color immunofluorescent staining of tissue sections was carried
out immediately after cutting. For direct staining. 2 �tg/IOO p1 each of
Flit-, Cy3-. and Cy5-labeled antibodies in PBS/1% bovine serum al-
bumin (BSA)/0.I% aside-containing human immunoglobulin (6 mg/mL
to block nonspecific and Fe-mediated binding) were added to sections
in 96-well plates and incubated overnight at 4#{176}Cin the dark with con-
tinuous gentle agitation. Unbound antibody was removed from the sec-
tions by aspiration followed by four 20-mm washes in PBS/1% BSA/0.I%
azide. Washed sections were then fixed overnight in the same buffer con-
taming 1% paraformaldehyde. Stained sections were wet-mounted in
anti-fade (Molecular Probes Inc. . Eugene, OR), sealed with nail varnish.
and stored at 4#{176}Cin the dark for up to 10 days before confocal imaging.
CSLM
Immunofluorescently labeled sections were optically sectioned using a
Bio-Rad MR1000 Confocal Scanning Laser Microscope system equipped
with a krypton/argon laser. Laser power, PMT gain, and enhancement
factors were then determined for the Flit, Cy3, and CyS channels by
use of the single fluorochrome-stained sections to ensure effective cross-
channel compensation. Three-color fluorescent sections were then eval-
uated for the presence of LAs. LAs were subjected to z sectioning using
a I- to 2-�tm z step at an appropriate confocality (typically an iris setting
between I and 2). The resulting image files were then used to create
three-dimensional reconstructions of the lymphoid aggregates.
RESULTS
Presence of basal LA at different stages of themenstrual cycle
Endometria were evaluated for the presence of basal LA
in sections stained with CD3. The presence of LA was
scored, on the basis of CD3 reactivity, as negative ( - ) if
there were no T cell clusters (i.e., accumulations ofT cells)
with a diameter of more than 2 T cells ( � 20 jim), small
( + ) if observed clusters had a diameter of 3-4 T cells
( � 20-40 �.tm), medium (+ +) if their diameter was be-
tween 5 and 10 T cells ( � 40-100 p.tm), and large (+ + +)
if their diameter was greater than 10 T cells ( 100 Rm).
The largest aggregates exceeded 20 cell diameters. Consid-
eration of the diameter of the T cell region in scoring the
size is important because the range of 0 to + + + suggests
a linear progression. However, the geometric nature of this
scale is emphasized by calculations of the number of cells
present in the different sizes of LA based on an average
cell diameter of 10.5 �im and assuming cell-cell contact.
Hence a score of zero represents fewer than 4 cells, where-
as large LA (+ + +) contain a minimum of 300 cells and
as many as 2400 cells.
The presence of CD3 � LA in postmenopausal, prolifer-
ative phase, and secretory phase endometna are summa-
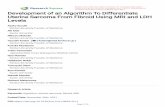
rized in Table 3. Significant differences in the presence
and size of these LA were observed during the menstrual
cycle. During the secretory phase of the cycle, when an mi-tial rise in estradiol is followed by a concomitant rise in
both estradiol and progesterone, all tissues examined con-
tamed large LA. As indicated in Table 3, LA in uteri from
women at the secretory stage were significantly larger than
those seen in uteri from women at the proliferative stage
of the menstrual cycle (P = 0.009, Fisher’s exact test). In
contrast, aggregates of T cells were conspicuously absent
in sections from postmenopausal women compared with
premenopausal women of either secretory phase (P = 2 x
10-u, Fisher’s exact test) or in proliferative phase (P =
0.01 1 , Fisher’s exact test). Proliferative phase endometria
exhibited a high degree of variability in both presence and
size of LA, suggesting that LA develop during this stage
of the menstrual cycle. To further define this heterogeneity
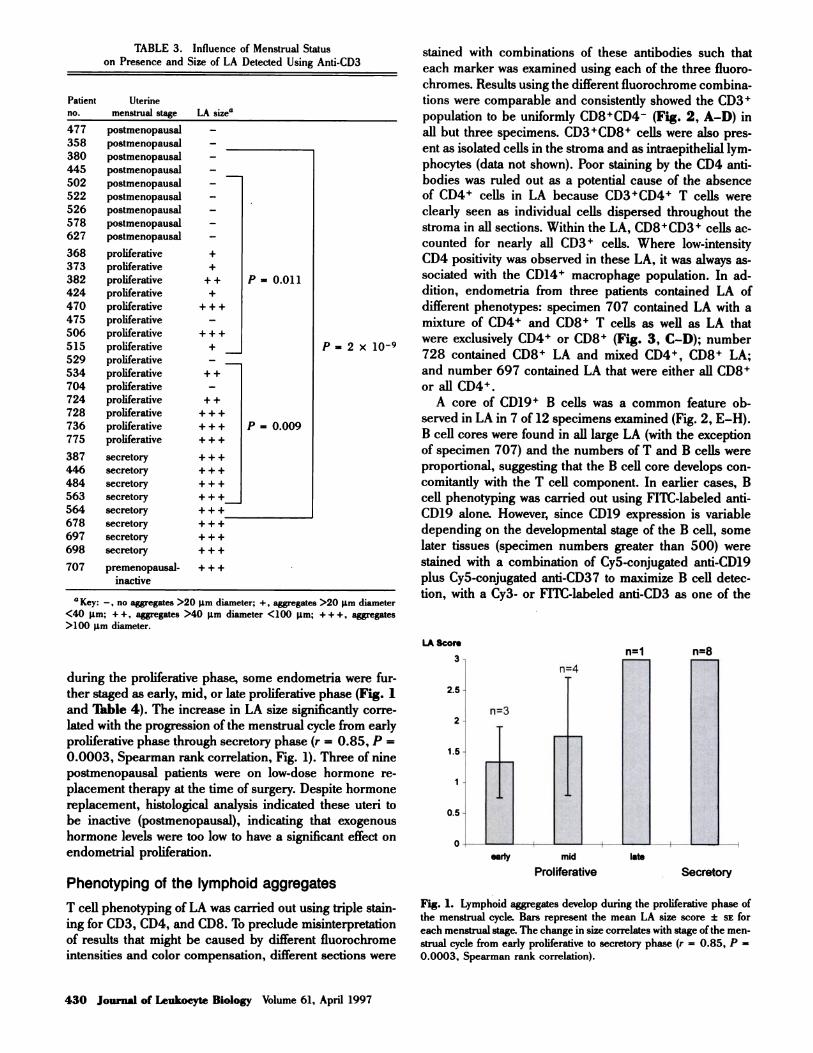
TABLE 3. Influence of Menstrual Status
on Presence and Size of LA Detected Using Anti-CD3
LA Score
3 n=1 n=8
n=4
2.5
n=32
I .5
0.5
0
early mid late
Proliferative Secretory
Fig. 1. Lymphoid aggregates develop during the proliferative phase of
the menstrual cycle. Bars represent the mean LA size score ± SE for
each menstrual stage. The change in size correlates with stage ofthe men-
strual cycle from early proliferative to secretory phase (r = 0.85, P =
0.0003, Spearman rank correlation).
430 Journal of Leukocyte Biology Volume 61, April 1997
Patient
no.
Uterine
menstrual stage LA sizea
477
358
380
445
502
522
526
578
627
postmenopausal
postmenopausal
postmenopausal
postmenopausal
postmenopausal
postmenopausal
postmenopausal
postmenopausal
postmenopausal
-
-
-
-
-
-
-
-
-
368
373
382
424
470
475
506
5 1 5
529
534
704
724
728
736
775
proliferative
proliferative
proliferative
proliferative
proliferative
proliferative
proliferative
proliferative
proliferative
proliferative
proliferative
proliferative
proliferative
proliferative
proliferative
+
+
+ +
+
+ + +
-
+ + +
+
-
+ +
-
+ +
+ + +
+ + +
+ + +
P = 0.011
P = 0.009
P = 2 x 10�
387
446
484
563
564
678
697
698
secretory
secretory
secretory
secretory
secretory
secretory
secretory
secretory
+++
+ + +
+ + +
+ + +
+ + +
+ + +
+ 4- +
+ + +
707 premenopausal-
inactive
+ + +
a Key: - , no aggregates >20 �sm diameter; + . aggregates >20 �tm diameter
<40 lim; + + , aggregates >40 �im diameter <100 �tm; + + + . aggregates
>100 �.tm diameter.
during the proliferative phase, some endometria were fur-
ther staged as early, mid, or late proliferative phase (Fig. 1
and Table 4). The increase in LA size significantly corre-
lated with the progression of the menstrual cycle from early
proliferative phase through secretory phase (r = 0.85, P =
0.0003, Spearman rank correlation, Fig. 1). Three of nine
postmenopausal patients were on low-dose hormone re-
placement therapy at the time of surgery. Despite hormone
replacement, histological analysis indicated these uteri to
be inactive (postmenopausal), indicating that exogenous
hormone levels were too low to have a significant effect on
endometrial proliferation.
Phenotyping of the lymphoid aggregates
T cell phenotyping of LA was carried out using triple stain-
ing for CD3, CD4, and CD8. To preclude misinterpretation
of results that might be caused by different fluorochrome
intensities and color compensation, different sections were
stained with combinations of these antibodies such that
each marker was examined using each of the three fluoro-
chromes. Results using the different fluorochrome combina-
tions were comparable and consistently showed the CD3�
population to be uniformly CD8�CD4 (Fig. 2, A-D) in
all but three specimens. CD3�CD8� cells were also pres-
ent as isolated cells in the stroma and as intraepithelial lym-
phocytes (data not shown). Poor staining by the CD4 anti-
bodies was ruled out as a potential cause of the absence
of CD4� cells in LA because CD3�CD4� T cells were
clearly seen as individual cells dispersed throughout the
stroma in all sections. Within the LA, CD8�CD3� cells ac-
counted for nearly all CD3� cells. Where low-intensity
CD4 positivity was observed in these LA, it was always as-
sociated with the CDi4� macrophage population. In ad-
dition, endometria from three patients contained LA of
different phenotypes: specimen 707 contained LA with a
mixture of CD4� and CD8� T cells as well as LA that
were exclusively CD4� or CD8� (Fig. 3, C-D); number
728 contained CD8� LA and mixed CD4�, CD8� LA;
and number 697 contained LA that were either all CD8�
or all CD4�.
A core of CD19� B cells was a common feature ob-
served in LA in 7 of 12 specimens examined (Fig. 2, E-H).
B cell cores were found in all large LA (with the exception
of specimen 707) and the numbers of T and B cells were
proportional, suggesting that the B cell core develops con-
comitantly with the T cell component. In earlier cases, B
cell phenotyping was carried out using Flit-labeled anti-
CD19 alone. However, since CD19 expression is variable
depending on the developmental stage of the B cell, some
later tissues (specimen numbers greater than 500) were
stained with a combination of Cy5-conjugated anti-CD19
plus Cy5-conjugated anti-CD37 to maximize B cell detec-
tion, with a Cy3- or FITC-labeled anti-CD3 as one of the

Patient
no. Histological stage Size score T cell phenotype
T cells
HLA-II B cell score
Macrophage
halo
Macrophage
HLA-II
736 proliferative (undetermined) + + + all CD8� - + + ±
775 proliferative (undetermined) + + + all CD8� ND + + ND
728 proliferative (undetermined) + + + all CD8�,
mixed CD4�
and CD8�
ND + + ND
368 proliferative (early) + all CD8� ND ND ND ND
515 proliferative (early) + all CD8� - - + ±
534 proliferative (early) + + all CD8� - - + ±
373 proliferative (mid) + all CD8� - - + ±
424 proliferative (mid) + all CD8� - ND + ±
382 proliferative (mid) + + all CD8� - - + ±
470 proliferative (mid) + + + all CD8� - + + ±
506 proliferative (late) + + + all CD8� - + + ±
697 secretory (early) + + + all CD8� and
all CD4�
ND ND ND ND
387 secretory + + + all CD8� - ND + ±
563 secretory + + + all CD8� - ND + ±
564 secretory + + + all CD8� - ND + ±
678 secretory + + + all CD8� ND ND ND ND
698 secretory + + + all CD8� ND ND ND ND
446 secretory (late) + + + all CD8� - + + ±
484 secretory (late) + + + all CD8� - + + ±
707 Inactive premenopausal. Active + + + all CD8�, ND - ND ND
papilloma virus infection. all CD4�,
Combined estrogen and mixed CD4�
progesterone therapy. and CD8�
Cervical cancer.
TABLE 4. Cellular Composition and HLA-II Expression of LA
Yeaman et al. Unique human uterine lymphoid aggregates 431
ND = not done.
other two colors. Sections stained in this way still showed
variable sizes of B cell cores, with two specimens (775 and
728) showing extensive B cell cores similar in extent to
germinal centers (Fig. 3B). No CD3�, CD37� cells were
observed.
CD14� monocyte/macrophage cells were also common
to all LA examined. As seen in Figure 2, G and H, these
cells always formed a halo around the T cell mantle (Table
4). On the other hand, polymorphonuclear leukocytes, ex-
amined using anti-CD66b, were not found within the LA.
LA were located in the stratum basalis, at a depth ap-
proximating the base of the uterine epithelial glands. No
direct contact with the glandular epithelium could be iden-
tilled even for large LA in five of five specimens: two from
late proliferative, and three from secretory phase (Fig. 3A).
This is in contrast to aggregated NK cells that we and others
find predominantly located in the upper layers of the endo-
metrium in close contact with both the glandular and lu-
minal epithelium [7] and as scattered individual cells in
the stratum basalis.
MHC class II expression
Using IVA12, a pan HLA class II marker (HLA-DR, DP,
DQ), we evaluated the class II expression of LA cells. In
all cases examined (n = 13), the T cell population was low
or negative for class II expression. In contrast, in 13 of 13
specimens, approximately half of the macrophages form-
ing the halo around the T cells were class II bright (Fig.
3, E-H). The question of B cell HLA class II expression
was specifically addressed for only three of the specimens.
Surprisingly, only in one (736) were B cells intensely pos-
itive for HLA class II, in another (506) they were clearly
negative, whereas in the third (534) there was a mixture
of class II positive and negative B cells within the same LA.
More comprehensive analysis of class II and other mole-
cules involved in antigen presentation will clearly be impor-
tant for future studies.
DISCUSSION
Using triple-color immunofluorescent staining and CSLM
of viable tissue sections, we have defined the characteris-
tics of immune cells in unique lymphoid aggregates pres-
ent in the human uterine stratum basalis. Previously, im-
munophenotyping of the reproductive tract has largely been
limited to either single-color immunohistochemical tech-
niques, which are generally inadequate for the detection
of dual expression of antigens on individual cells, or flow-
cytometric analysis of mechanically or enzymatically dis-
rupted tissue. Our approach presents three major advan-
tages over more conventional techniques. First, no current
tissue fixation technique uniformly preserves all epitopes
recognized by monoclonal antibodies, leading to a restric-
tion in antigens that can be detected; use of fresh unfixed
tissue sections largely avoids this problem. Second, this ap-
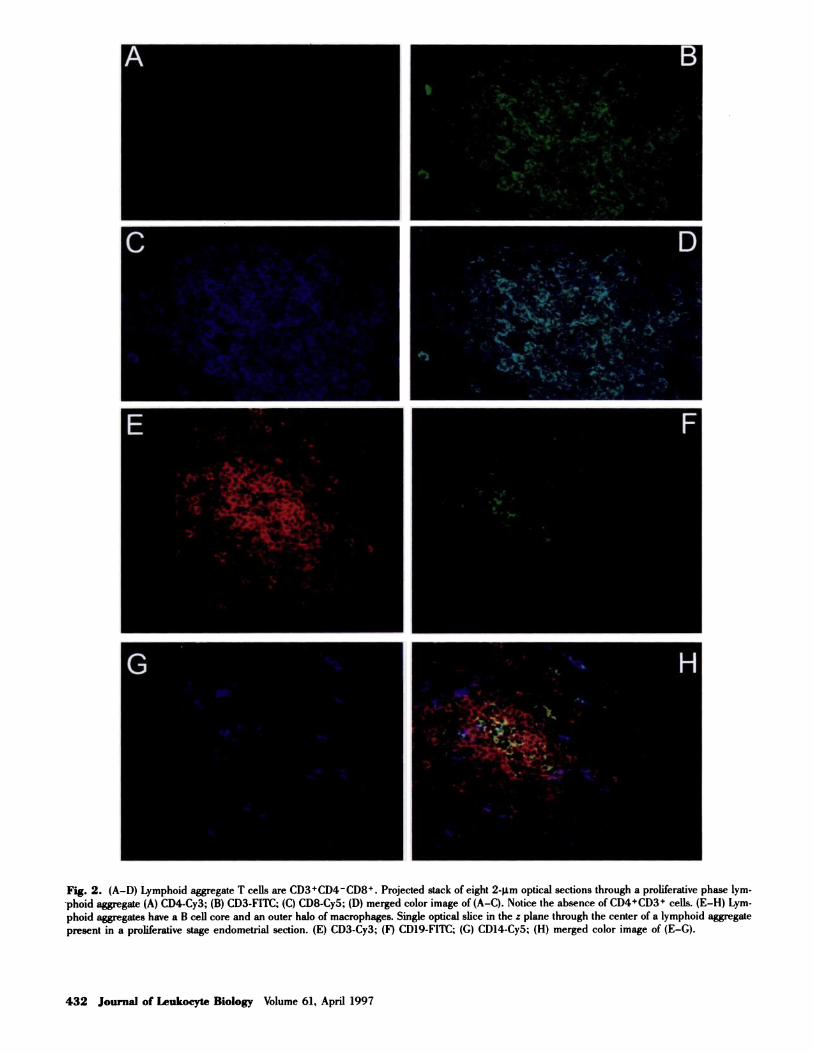
Fig. 2. (A-D) Lymphoid aggregate T cells are CD3�CD4CD8�. Projected stack of eight 2-ptm optical sections through a proliferative phase lym-
phoid aggregate (A) CD4-Cy3; (B) CD3-FITC; (C) CD8-Cy5; (D) merged color image of (A-C). Notice the absence of CD4�CD3� cells. (E-H) Lym-
phoid aggregates have a B cell core and an outer halo of macrophages. Single optical slice in the z plane through the center of a lymphoid aggregate
present in a proliferative stage endometrial section. (E) CD3-Cy3; (F) CD19-FITC; (G) CD14-Cy5; (H) merged color image of (E-G).
432 Journal of Leukocyte Biology Volume 61, April 1997
Fig. 3. (A) LA are located between the
bases of the endometrial glands. Projected
stack ofeight 2-�.tm optical sections through
a dual-color image of a CD8� LA in a Se-
cretory phase endometrium showing LA
CD8 cells (Cy5. blue) and glandular epithe-
hum stained with Ber-EP4 (Flit, green).
(B) B cell cores have a germinal center
appearance in some large LA. Single low-
confocality ( = 10 rim) slice through the cen-
ter of a large LA in a proliferative phase tissue
showing a large B cell core (CD19�CD37
both labeled with Cy5. blue) surrounded
by T cells (CD3-FITC, green) and macro-
phages (CD14-Cy3, red). (C) A CD4� LA
in a premenopausal non-cycling endome-
trium. Three-color single slice through the
center of a LA from endometria no. 707:
CD3-Cy3 (red); CD4-FITC (green); CD8-
Cy5 (blue). Notice the predominance of
CD4�CD3� cells (yellow/orange). (D) A
mixed CD8�/CD4� LA in a premenopausal
non-cycling endometrium. Three-color sin-
gte slice through the center of a LA from
the same vibratome section as in (C): CD3-Cy3 (red); CD4-FITC (green); CD8-Cy5 (blue). Notice the presence of both CD3�, CD8� cells (purple)
and CD3�, CD8� (yellow). (E-H) Macrophages but not T cells are class II positive in LA. Single optical slice through the center of a proliferative
phase lymphoid aggregate. (E) CDI4-Cy3; (F) CD3-FITC; (G) pan HLA class II-Cy5; (H) merged color image of (E-H). Notice the lack of CD3/pan
HLA class II dual-positive cells (turquoise color in H) and the relative abundance of CD14/pan HLA class II dual-positive cells (purple color in H).
Yeaman et al. Unique human uterine lymphoid aggregates 433

434 Journal of Leukocyte Biology Volume 61. April 1997
proach permits co-localization of antigens on individual
cells but without the tissue disruption required for flow cy-
tometry. Third, z sectioning of thick vibratome sections a!-
lows three-dimensional reconstruction, giving more de-
tailed spatial information, including location of cell types
and cell-cell interactions within the tissue.
The CD8� LA described in this study are distinct from
large granular lymphocyte aggregates described by others
[reviewed in ref. 7]. They appear to be spread between
endometrial glands approximately equidistant from the
base of the glands in the stratum basalis, and in accor-
dance with previously published micrographs [16, 22],
often seem to be associated with blood vessels, as evi-
denced by phase-contrast appearance and smooth muscle
autofluorescence in the Flit channel. These CD8� LA
develop during the proliferative phase of the menstrual cy-
cle, reach their largest cell mass at mid-cycle (i.e., late pro-
liferative phase) and persist throughout the secretory phase.
This, taken in conjunction with their absence from uterine
endometrium of post-menopausal women, suggests that
their development is under the influence of sex hormones.
During the proliferative phase LA often have a central B
cell core that is surrounded by a mass of (usually) exclu-
sively CD8� T cells, surrounded in turn by an outer halo
of macrophages.
Our finding of LA development during the course of the
menstrual cycle extends the previous finding of Tabibzadeh
[17] who reported that BrDU incorporation by CD3� LA
cells in vibratome sections increased during the menstrual
cycle, particularly following ovulation. This proliferation ap-
peared to be interleukin-2 independent and may depend
on steroid hormones because lymphoid aggregate T cells
express estrogen receptors [23]. Progesterone has previ-
ously been shown to increase T cell numbers throughout
the uterus during the luteal phase of the cycle [24]. Our
findings show that LA seen in uteri during the secretory
phase of the menstrual cycle are significantly greater in size
than those seen in the early proliferative phase. This sug-
gests that estradiol and progesterone, both known to be el-
evated during this time, may act either directly or indirectly
to increase T cell numbers in the lymphoid aggregates. Al-
though others have noted a predominance of CD8� T cells
in both endometrial stroma and LA [3, 6, 10], our studies
show that in nearly all cases the T cell lymphoid aggregates
in the basalis region are exclusively of the CD3 �CD8�CD4
phenotype. CD3�CD8�CD4� cells were observed in the
stroma of the same tissues but not within the lymphoid ag-
gregates. All CD4 � reactivity within LA in these speci-
mens was confined to the CD14�, monocyte/macrophage
population.
Three of the tissues did not follow the above pattern in
that they displayed either exclusively CD4� and/or mixed
CD4�/CD8� LA in addition to the exclusively CD8� LA.
One patient (707) with this phenotype had both an active
human papilloma virus infection and an endometrium his-
tologically classified as inactive. The latter probably re-
sulted from concomitant high-dose estrogen and progester-
one therapy. This patient also had extensive cervical cancer.
No overt endometrial infections were noted for the other
two patients. One patient (728) was receiving progesterone
therapy and the other (697) had ovarian cysts. It is not
possible with only three cases to determine the underlying
differences leading to the formation of CD4� LA, but they
may result from an immunological response to infection or
neoplasia or from disruption of the endocrine balance.
The presence ofa defined B cell core in large LA demon-
strates a precise structural relationship between B cells and
CD8� T cells in the uterus. Although scattered B cells
have been noted in the same region of the stratum basalis
in two previous studies [3, 10], the findings presented here
demonstrate that the B cell core is a defined structural fea-
ture of the larger lymphoid aggregates. In contrast small
LA, identified most commonly in tissues from women at
the early proliferative stage of the menstrual cycle, lack B
cell cores. Whether this reflects a sequential construction
of LA in which chemokines initially attract CD8� cells fol-
lowed by B cell migration remains to be established. Macro-
phages form a halo around the periphery of the LA and
are seldom seen deep within the T cell or B cell regions.
In 13 of 13 cases, a high proportion CD14� cells, both sur-
rounding the LA and distributed throughout the stroma,
were found to be strongly HLA class II positive. In con-
trast, lymphoid aggregate T cells were uniformly negative
for class II expression. HLA class II expression in the B
cell core has not, as yet, been definitively examined. HLA
class II expression in human endometrium has previously
been examined by a number of investigators who uniformly
reported stromal HLA-DR positivity to have the same stain-
ing pattern as the macrophage population [3, 5, 16, 18,
25-27]. Two reports have specifically shown cells surround-
ing the basal LA to be strongly HLA-DR positive [3, 25].
Both of these studies also showed some cells within LA to
be HLA-DR positive, however, it is not possible to identify
the phenotype of the positive cells from these studies.
Uterine CD8� LA are also unique and distinct from
those found in secondary lymphoid tissues and at other
mucosal surfaces [28, 29]. For example, whereas primary
follicles in the periarteriolar lymphoid sheaths of the spleen
contain a B cell core surrounded by CD4� T cells, LA in
the uterus contain CD8�CD4 cells. The gastrointestinal
tract Peyer’s patches (PP) contain both CD4� and CD84
zones that contain B cells. Because PP are anatomically ad-
jacent to epithelial cells that line the intestinal lumen, they
are exposed to luminal antigens that pass through M cells
directly into the PP. In contrast, LA in the uterine stratum
basalis usually lack CD4� T cells and have no direct con-
tact with the uterine epithelial cells, as we find them equi-
distant between the bases of the epithelial glands. To date,
despite extensive investigation, M cells have not been found
in reproductive tract tissues. Also, in contrast to PP that
have macrophages distributed throughout, uterine LA are
surrounded by a halo of macrophages.
The functions of LA are presently unknown but their de-
velopment during the menstrual cycle and their absence
in postmenopausal women suggests that they are hormon-
ally influenced. It is possible that as LA these cells play an

Yeaman et a!. Unique human uterine lymphoid aggregates 435
immunoregulatory role, perhaps suppressing cell-mediated
immune responses during the postovulatory implantation
window to prevent immune responses against the semi-
allogeneic conceptus. In support of this hypothesis, our
group has recently shown that cytotoxic T cell function,
present throughout the female reproductive tract (Fallopi-
an tube, uterus, cervix, and vagina) is specifically sup-
pressed only in the endometrium during the secretory stage
of the menstrual cycle [301. Whether localized suppression
of cytotoxic T cell function is a consequence of LA forma-
tion remains to be established. Alternatively, LA may play
a central role in immune protection at the time of menstru-.
ation (5-7 days) when the outer third of the endometrium
is shed. During this time the protective barrier of epithelial
cells and the stromal immune cells of the decidual stroma
are lost, leaving the uterus uniquely vulnerable to infection
by viral and bacterial pathogens. We postulate that the
growth of LA during the cycle in the basalis stroma, which
is not shed during menses, is consistent with a consolida-
tion of T and B cells to prevent loss during menstruation.
Overall, our results indicate that LA in the uterus are
unique, as judged by their structure, location, and potential
endocrine control. Further studies are needed to identify
the events responsible for LA formation and the physiolog-
ical role these cells play in contributing to maternal im-
mune protection and fetal survival.
ACKNOWLEDGM ENTS
This work was supported by National Institutes of Health
Grant AI34478 (C. R. W.). Confocal scanning laser micros-
copy was performed in the Herbert C. Englert Cell Analy-
sis Laboratory, which was established with a grant from the
Fannie E. Rippel Foundation, and is supported in part by
the core grant of the Norris Cotton Cancer Center (CA-
23108).
The authors would like to thank the following individu-
als for their technical and clinical support: Dr. Alice Given,
Dr. Vincent Memoli, Dr. John Currie, Dr. Stephen Andrews,
Dr. Joan Barthold, Dr. Jackson Beecham, Dr. John Ket-
terer, Dr. Eileen Kirk, Dr. Benjamin Mahlab, Dr. Paul Man-
ganiello. Dr. Eric Sailer, Dr. Barry Smith, Dr. William
Young, Jaclyn Logren, Fran Reinfrank, Jeannette Sawyer,
Tracy Stokes, Joanne Lavin, Nancy Leonard, Kris Ramsey,
Tamara Krivit, Laura Wolf, Peter Seery, Maryalice Ach-
bach, Judy Rook, and Esther Colby.
REFERENCES
I . Harbour, D. V.. Blalock. J. E. (1989) Lymphocytes and lymphocyti’ hor.
mones in pregnancy. Prog. Neuroendocrinol. Immunol. 2. 55-63.2. Dallenbach-Hellweg. G. (1975) Hi.stopathology of the Endometnum, New
York: Springer-Verlag.3. Kamat. B., lsaacson. P. (1987)The immunocytochemical distribution of leo.
kocytic subpopulations in human endometrium. Am. J. Pat/wI. 127.66-73.
4. Bonatz, G., Hansmann, M., Buchholz, F., Mettler, L., Radzun, H., Semm,K. (1992) Macrophage. and lymphocyte-subtypes in the endometrium dur-
ing different phases of the ovarian cycle. mt. J. Gynecol. Obstet. 37, 29-36.
5. Laguens, G., Goni, J., Jr., Laguens, M., Goni, J., Laguens, R. (1990)Demonstration and characterization of HLA-DR positive cells in the stromaof human endometrium. J. Reprod. Immunol. 18. 179-186.
6. Morris, H., Edwards, J., Tiltman, A., Emms, M. (1985) Endometrial lym-phoid tissue: An immunohistological study. 2 Clin. Pa.thol. 38, 644-652.
7. Loke, Y. W., King, A. (1996) Human Implantation: Cell Biology and Im-
munology, New York: Cambridge University Press, 102-129.
8. Wira, C., Stern, J. (1992) Endocrine regulation ofthe mucosal immune sys-
tem in the female reproductive tract: Control of IgA, IgG, and secretory com-ponent during the reproductive cycle, at implantation and throughout preg-
nancy. In Hormones and Fetal Pathophysiology (J. Pasqualini and R.
Scholler, eds.), New York: Decker, 343-368.9. Wira, C.. Sandoe. C. (1977) Sex steroid hormone regulation of immuno-
globulin G (IgG) and A (IgA) in rat uterine secretions. Nature 268,534-536.
10. Marshall, R., Jones, D. (1988) An immunohistochemical study of lymphoid
tissue in human endometrium. hit. J. Gynecol. Pat/wI. 7. 225-235.1 1 . Hunt, J. S. (1994) Immunologically relevant cells in the uterus. Biol. Re-
prod. 50, 461-466.12. Chen, C-K.. Huang. S. C.. Chen, C-L., Yen, M-R., Hsu, H-C., Ho, H-N.
(1995) Increased expression of CD69 and HLA-DR but not of CD2S or
CD71 on endometrial T lymphocytes of nonpregnant women. Human Im-munol. 42, 227-232.
13. Lachapelle, M-H., Miron. P.. Hemmings, R., Baron, C., Roy, D. C. (1996)Flow-cytometric characterization o hematopoietic cells in non-pregnant ho-
man endometrium. Am. J. Repmd. Immunol. 35, 5-13.14. Lachapelle, M. W., Miron, P., Hemmings, R., Roy, D. C. (1996) Endome-
trial T, B, and NK cells in patients with recurrent spontaneous abortion.J. ImmunoL 156, 4027-4034.
15. King, A., Wellings, V., Gardner, L., Loke. Y. W. (1989) Immunocytochemi-
cal characterization of the unusual large granular lymphocytes in human
endometrium throughout the menstrual cycle. Human Immunol. 24, 195-205�
16. Tabibzadeh, S. S.. Poubouridis, D. (1990) Expression ofleukocyte adhesionmolecules in human endometrium. Am. .� Clin. Pathol. 93, 183-189.
1 7. Tabibzadeh, S. (1990) Proliferative activity of lymphoid cells in the humanendometrium throughout the menstrual cycle. J. Clin. Endocrinol. Metab.
70. 437-443.18. Bulmer, J., Lunny, D.. Hagin, S. (1988) Immunohistochemical characteriza-
tion of stromal leukocytes in nonpregnant human endometrium. Am. J. Re-
prod. Immunol. Microbiol. 17. 83-90.19. Witz, C. A., Montoya. I. A.. Dey. T. D.. Sehenken. R. S. (1994) Character-
ization of lymphocyte subpopulations and T cell activation in endometriosis.
Am. J. Reprod. Immunol. 32. 173-179.20. Fernandez-Shaw. S.. Clarke, M. T.. Hicks, B. Naish, C. E.. Barlow, D. H..
Starkey, P. M. (1995) Bone marrow-derived cell populations in uterine andectopic endometrium. Human Reprod. 10, 2285-2289.
21. Noyes, R. W.. Hertig. A. T. (1955) Dating the endometrial biopsy. Fertil.
Steril. 1. 3-25.22. Tabibzadeh, S. (1990) Immunoreactivity of human endometrium: correla-
tion with endometrial dating. Fertil. Steril. 54. 624-631.23. Tabibzadeh. S. S.. Satyaswaroop. P. G. (1989) Sex steroid receptors in lym-
phoid cells of human endometrium. Am. J. Pat/wI. 91, 656-663.24. Booker, S. S.. Jayanetti. C.. Karalak. S.. Hsiu. J. G., Archer. D. F. (1994)
The effect of progesterone on the accumulation of leukocytes in the humanendometrium. Am. J. Obstet. Gynecol. 171, 139-142.
25. Tabibzadeh, S. (1990) Evidence of T-cell activation and potential cytokine
action in human endometrium. J. Clin. Endocrinol. Metab. 71, 645-649.26. Tabibzadeh, S. (1991) Distinct subsets of stromal cells confined to unique
microenvironments in human endometrium throughout the menstrual cv-cle. Am. J. Repmd. Immunol. 26. 5-10.
27. Bjercke. S.. Brandtzaeg. P. (1993) Glandular distribution of immunoglob-
ulins, J chain, secretory component, and HLA-DR in the human endome-trium throughout the menstrual cycle. Human Reprod. 8. 1420-1425.
28. Picker. L. J., Siegelman. M. H. (1993) Lymphoid tissues and organs. InFundamental Immunology. 3rd ed. (W. E. Paul. ed). New York: Raven
Press, 145-197.29. Griebel. P. J.. Hem, W. R. (1996) Expanding the role of Peyer’s patches
in B-cell ontogeny. Immunol. Today 17. 30-39.30. White, H. D.. Crassi. K. M.. Givan, A. L.. Stern. J. E.. Green, W. R.. Wira.
C. R. (1997) CTL activity within the human female reproductive tract: influ-efl(’e of stage of the menstrual cycle and menopause. J. ImmunoL. in press.
Copyright © 2022 FDOKUMEN