
The Rio Chico Paleozoic crystalline complex and the evolution of Northern Patagonia

T H E B O T A N I C A L R E V I E W VOL. 45 J U L Y - S E P T E M B E R , 1 9 7 9 No. 3
PALEOZOIC SEED FERN POLLEN ORGANS
M I C H A E L A . M I L L A Y A N D T H O M A S N . T A Y L O R
D e p a r t m e n t o f B o t a n y
T h e O h i o S t a t e U n i v e r s i t y
C o l u m b u s , O h i o 4 3 2 1 0
Abstrac t . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 301
Introduct ion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 306
Early Pter idosperm Microsporangia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 307
Origin of Synangia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 307
A l c i c o r n o p t e r i s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 310
G e m i n i t h e c a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 311
P r o t o p i t y s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 311
S t a p h y l o t h e c a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 313 P a r a e a l a t h i o p s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 313
S i m p l o t h e e a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 313
Z i m m e r m a n n i t h e c a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 314
Lyginopter idaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 314
Branching Patterns . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 315
Permineral ized Specimens . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 319
Prepollen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 322
Evolut ionary Trends . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 322
Medul losaceae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 323 Permineral ized Specimens . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 325
Simple Synangia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 326
H a l l e t h e c a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 326 S t e w a r t i o t h e c a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 326
S u l l i t h e c a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 328
Aggregate Synangia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 331
R h e t i n o t h e c a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 331
P a r a s p o r o t h e c a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 332 Compound Synangia . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 334
P o t o n i e a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 334
D o l e r o t h e c a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 336
Impress ion-Compress ion Forms . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 341
A u l a c o t h e e a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 341
B o u l a y a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 4 4
C o d o n o t h e e a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3 4 4
D e l t e n r e a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 346
G i v e s i a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 346
G o l d e n b e r g i a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 346
S e h o p f i t h e c a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 347
W h i t t l e s e y a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 347
Prepollen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 348
The Botanical Review 45: 301-375. July-September, 1979. 301 �9 1979 The New York Botanical Garden

302 THE BOTANICAL REVIEW
C a l l i s t o p h y t a c e a e . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 349
P e r m i n e r a l i z e d S p e c i m e n s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 351
Pol len . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 352
P rob l em a t i ca l F o r m s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 353
D i c t y o t h a l a m u s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 353
P s a l i a n g i u m . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 353
S c h u e t z i a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 353
T h u r i n g i a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 356
U n g u i t h e c a . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 356
Di scuss ion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 357
P h y l o g e n e t i c T r e n d s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 357
B r a n c h i n g Pa t t e rns . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 363
P repoUen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 367
S u m m a r y . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 370
A c k n o w l e d g m e n t s . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 371
L i t e r a t u r e Ci ted . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 371
A B S T R A C T
Paleozoic pollen organs exhibit numerous morphological forms that have been arranged in categories based on their probable organization. Progymnosperm ancestors are characterized by three dimensional branching systems bearing pairs of terminal sporangia. Early Mississip- plan examples of seed fern fertile branches appear little modified from the progymnosperms. These pteridosperm microsporangia are nonsynan- giate and thin walled with longitudinal dehiscence. By Upper Mississip- pian time all forms show sporangial clustering into large or small groups, with several taxa exhibiting radially symmetrical synangia. In the Lower Pennsylvanian all pollen organs are synangiate and appear to consist of a uniseriate ring of sporangia that either surround a central hollow, or are bilaterally flattened. Sporangial dehiscence in all forms is longitudinal and toward the center of the synangium. In bilateral synangia with no central hollow, the sporangia either separate laterally or effective dehiscence areas are restricted to the free apical portions of the sporangia. Callis- tophytacean synangia resemble the lyginopterid type, but are abaxial on laminar foliage. This family is thought to have evolved from the lyginop- terids during the Early Pennsylvanian. Middle Pennsylvanian medullosan pollen organs are all radial and may be solitary, aggregated into groups, or fused into a large compound synangium. Several pollen organ types are reinterpreted, and the possible evolutionary relationships among the various Paleozoic pollen organ forms are discussed based on synangial organization, patterns of frond branching, and pollen or prepollen mor- phology.

PALEOZOIC SEED FERNS 303
RI~SUME
Les organes b. pollen de l'~re pal6ozoique manifestent nombreuses formes morphologiques qui permettent une cat6gorisation bas6e sur leur organisation probable. Les ascendants sont des progymnospermes et sont caract6risent par un syst~me de branchement/~ trois dimensions qui se termine par des sporanges dispos6s en paires. Les microsporanges des pt6ridospermes du d6but de la p6riode Mississippienne r6vblent peu de diff&ence d'avec ceux des progymnospermes. Les microsporanges des pt6ridospermes nonsynangiates (pas fusionn6s) pr6sentent des parois minces et une d6hiscence longitudinale. A partir de la deuxi~me moiti6 de la p6riode Mississippienne routes les formes manifestent un regroupe- ment de sporanges enlarges ou petits groupes et plusieurs taxons mani- festent des synanges a sym6trie rayonn6e. Dans la premibre p6riode Pennsylvanienne, tous les organes ~t pollen sont du genre synange et semblent se composer d'un anneau unis6riel de sporanges qui soit entoure un creux central, on soit s'aplatit de fa~on bilat6rale. La d6hiscence spo- rangiale sous routes ses formes est longitudinale et orient6e vers le centre du synange. Parmi les synanges de type bilat6ral, d6pourvu de creux central, les sporanges soit se divisent bilat6ralement ou manifestent des lieux de d6hiscence qui sont limit6s h la partie apicale des sporanges. Les synanges de la famille Callistophytac6enne ressemblent au type lyginop- t6ride, mais sont abaxiaux sur feuillage laminaire. Cette famille est sup- posse avoir 6volu6e h partir des lyginopt~rides au d6but de la periode Pennsylvanienne. Vers le milieu de cette m~me p6riode, les organes pollen genre Medullosa sont tous radiaires et peuvent ~tre solitaires, regroup6s, ou fusionn6s en un large synange compos6. Plusieurs types d'organes 5. pollen sont r6examines, et les rapports 6volutionnaires pos- sibles parmi les formes d'organes ~t pollen de l'~re Pal6ozoique sont dis- cut6s, bas6s sur l'organisation du synange, sur le branchement des frondes, enfin sur la morphologie du pollen ou pr6pollen.
ZUSAMMENFASSUNG
Paleoz~ine Blfitenstauborgane weisen zahlreiche morphologische For- men auf, die auf Grund ihrer wahrscheinlichen Organisation in Katego- rien aufgeteilt worden sind. Die Urtypen heiBen Progymnospermen und sind durch sich in drei Dimensionen abzweigende Systeme gekennzeich- net, die paarweise Endsporangien tragen. Bei Proben der Samenfarn- pflanzen aus der frfihen Mississippi-Zeitepoche scheinen fruchtbare Zweige von denen der Progymnospermen wenig abmodifiziert. Diese Mikrosporangien der Pteridospermen sind nichtsynangiatisch und haben diinne Wfinde mit L~ingsaufspringen. Bis zur Ober-Mississippi-Zeite-

304 THE BOTANICAL REVIEW
poche sammeln sich die Sporangien aller Formen in grol3e oder kleine Gruppen; die Sporangien mehrerer Taxus weisen eine strahlenfrrmige Symetrie auf. In der Nieder-Pennsylvania-Zeitepoche sind alle Blfiten- stauborgane synagiatisch: sie bestehen anscheinend aus einem einreihi- gen Kreis von Sporangien, die entweder eine zentrale Hrhle einkreisen oder sich beiderseitig abflachen. Das Aufspringen der Sporangien in allen Formen ereignet sich der L/inge nach und auf die Mitte des Synangiums zu. Bei zweiseitigen Sporangien mit keiner Hrhle 16sen sich die Spor- angien zur Seite, oder die wirkenden Gegenden des Aufspringens sind auf die freien an der Spitze befindlichen Teile der Sporangien beschr~inkt. Kallistophytoz~ine Synangien gleichen den lyginopteridischen Typen, sie sind aber nichtaxial und befinden sich auf bl~ittrigem Laubwerk. Diese Familie, denkt man, habe sich aus den Lyginopteriden w~ihrend der frii- hen Pennsylvania-Zeitepoche entwickelt. Markige Bliitenstauborgane der Mittel-Pennsylvania-Zeitepoche sind alle strahlenfrrmig und diJrfen ein- zeln, in Gruppen aggregiert, oder in einem grol3en zusammengesetzten Synangium zusammengeschmolzen sein. Mehrere Typen der Bliitenstau- borgane werden neuerkl/irt, und mrgliche Evolutionsverh~iltnisse der ver- schiedenen Formen von paleoz/inen Bliitenstauborganen werden auf Grund der Synangienorganisation, der Muster von Bl/itterabzweigung und der Bl/itenstaub- oder Vorbliitenstaubmorphologie besprochen.
R E S U M E N
Los 6rganos Paleozoicos portadores de polen, exhiben una variada morfologia que ha sido ordenada en categorfas, bas~indose en su probable organizacirn. Los antecesores progimnosprrmicos estfin caracterizados por sistemas ramificados tridimensionales que portan pares de esporan- gios terminales. Muestras del Mississippiano temprano de ramificaciones fdrtiles de helechos con semillas, parecen poco modificadas en relaci6n con las Progimnospermas. Estos microsporangios pteridosprrmicos son nosinangiados y con paredes delagadas con dehiscencia longitudinal. En el Mississippiano superior todas las formas muestran esporangios agru- pados en grandes o pequefios grupos, con algunos taxa que exhiben si- nangios con simetria radial. En el Pennsylvaniano temprano, todos los 6rganos portadores de polen son sinangiados y parecen consistir en un anillo uniseriado de esporangios que rodean un hueco central o se dis- ponen en forma de anillo achatado. La dehiscencia de los esporangios en todas las formas es longitudinal y hacia el centro del sinangio. En los sinangios bilaterales sin hueco central, los esporangios se separan late- ralmente o las ~ireas efectivas de dehiscencia est~in restringidas a las por- ciones apicales libres de los esporangios. Los sinangios de las Callisto- phytaceas parecen de tipo lyginopteroideo pero est~in en la parte abaxial de la lamina foliar. Se cree que esta familia ha evolucionado de las Ly-

PALEOZOIC SEED FERNS 305
ginopteroideas durante el Pennsylvaniano temprano. Los 6rganos por- tadores de polen de las medullosas del Pennsylvaniano medio son todos radiales y pueden ser solitarios, agregados en grupos o fusionados en grandes sinangios compues tos . Basfindose en ia organizaci6n sinangial, los patrones de ramificaci6n de las frondes y la morfologia del polen o del prepolen, algunos tipos de organos por tadores de po|en son reinter- pre tados y se discute las posibles relaciones evolut ivas entre las formas de 6rganos por tadores de polen del paleozoico.
A B C T P A K T
HaJ1eo3oficKne nb~abueab~e opraHbl npe~cTaaneHbl MHoroqncneH- HblUn mopqboaor14qecgHun qbopuaun, rOTOphm u o r y T 6blTb pac- npe~eJIeHbl no r a T e r o p ~ a u a 3aancnuocTn ox nx n p e J m o n a r a e u o r o ycTpoficTaa. Hponcule~lmne OT npornMHOCnepMHh~X xapaKTepH- 3ylOTC~I TpeMrt TnnaMn C14CTeM aeTB~ennn, necyttmMn napbl Tep- MIJHaJIbHblX crIopaHrrlfi. PaHHe-MrlccricrlncKne o6pa3ttbl n~o~Ioce- MeHHblX IIaI-IOpOTHHKOBblX BeTBeH Ma310 qeM OT3114qalOTC~I OT npOrHMHOCnepMnblX. ~anHble nanopownnKOCeMennble MHKpO- cnopaHran He CHHaHVnpylOT H TOHKO neperopo~Keobl HpoJ1031bHbIM
pacTpeCK14BaoI4eM. K Haqa21y BepxHe-M14ccHcHncKoro nep~o~Ia Bce tl2OpMbl ~IeMOHCTpHpylOT crlopaHFHaJlbHble CKOILrleOI4~l B 6o~bmne r~ Ma.abie rpynnbl c OTJle31bHblMl, l IIOCTpOeHI4~IMI4, IlpejlCTaB31~IIOIUHM14 pa/irta_rtbHO CI4MMeTp14qHhle CrtHaHFHH. B HHXHe-I-IeHcH31bBaHCK14fi rlep140~t Bce Ilbl.llbReBble opFaHbl CI4HaHFHpyI~TCfl 14 rlpe~ICTaB31YlIOTC,q KaK co~lepxamne o~mocepnfiHmfi Kpyr cnopanrnfi ; OHn ml60 oKpy- ~Ka~OT lleHTpa~bHy~O rlO~OCTb, 31H60 ~ByCTOpOHHe Cn31rotUeHbL CI1opaHvna.rlbHOe pacTpecK14Bar114e BO Bcex qbopMax llpO~Oa-lbHO 14 Hatlpaa~eHo K ReHTpy CBHaHF1414. B ~IBycTOpOHHe ClI311OLUeHHblX ClelHaHF14~IX 6e3 lleHTpaJlbHO~l I1031OCTId cnopaHr1414 ~ri6o pa3~e- h~0OTC~ no 6OKaM, 31n60 3qbqbeKT14BHble pafiOHbl pacTpeCK14BaH14~ orpaHl4qeHbl CBO60~14blM14 BepxyttleqHblMH HOpR14~M14 cIlopaHrnfi. K aJ]21 ld CTOdIgMTO Bbl e CMHaHFFIFI HaI1OMMHalOT .rlld FFI HOIITepld~14 bl.~
TMI1, O)IHaKO HaIlpaBJleHbl B CTOpOHy OT OC14 Ha C31OHCTOfi 31HCTBe. Hpenno31araeTca, tITO 3TO ceMefiCTBO pa3sl4~OCb I43 31,rnHonTepnzos BO speM~ Panne-Henc1431bsancKoro nep14o~a. Cpense-Henc14at,- sancxne cep~ItteSnHHble Hbl.rlhlXeSble opraHbl MOryT 6b~Tb 31yqe- o6pa3nopacxo~,qlR14M14C$1 143114 me OJ1HHOqHbIM14, CO6paHHblM14 B rpynnb~, 311460 C31r~TbI B 6o~bmofi C310~Hhlfi cmmm14fi. OT~lem, m, le T141qhl IIblJIbl.~eBbIX opFaHOB pflCCMOTpeHbl 3aHOBO, BO3MO~KHble
3BO.rlIO~MOHHble OTHOIHeH14fl pa3.rllaqHblX (12OpM Ha.rleo30fiCKldX Hb131bl.l,e-
BblX opFaHOB o6cyTK~leHbl B 3aB14C14MOCT14 OT C14 HaHFHa.rlbHOFO
yCTpOfiCTBa, pHCyHKOB .rlIdCTOB nanopOTH14Ka, a TaK>Ke HbI31bUeBO14
t4 ~lOnhl.rlbUeBOfi MopqboYIor14n.

306 THE BOTANICAL REVIEW
I N TRO D U CTI O N
Despite the increasing importance the study of pollen has played in understanding the evolution of early seed plants, it is surprising that no definitive study has been directed at the morphology and evolution of the organs that produced these grains. This is in part due to the infrequent occurrence of well-preserved pollen organs in the fossil record, and the absence of a theoretical framework upon which to evaluate the morpho- logical complexities of the organs. Within recent years, however, there have been a number of anatomically preserved pollen bearing organs described of Paleozoic age that provide important transitional types use- ful in elucidating the morphological complexities as well as indicating homologies. Such homologies provide a basis for establishing the system- atic affinities of new specimens and problematical forms, a potential index of evolutionary advancement, and a starting point from which to consider early seed plant origins.
There have been numerous reviews that have traced the origin and theoretical development of the seed habit from probable Devonian ances- tors. Such studies have relied almost exclusively on the origin of the integumentary system from an enveloping branching system, and the modification of the distal end of the sporangium for pollen reception. Two important discoveries that establish a starting point in seed plant evolu- tion are the seedlike structures described from Upper Devonian rocks as Archaeosperma (Pettitt and Beck, 1968) and Spermolithus (Chaloner et al., 1977). Despite the potential uncertainty regarding the precise nature of the integument and distal end of the nucellus in these forms, the mor- phological organization of the fossils provides a sound basis for consid- ering them as seedlike in function.
In tracing the origin and development of seed plant microspores, in- vestigators have relied almost exclusively on sporae dispersae grains that in many instances have poorly understood biological affinities. In these studies pollen evolution has been based upon such parameters as size, haptotypic features, grain polarity and organization, and more recently on ultrastructural features. Even in those instances where pollen has been extracted from organs, a better understanding of the natural relationships of the parent plant will make significant contributions to our understand- ing of various trends in pollen evolution.
At present our understanding of Mississippian seed fern microsporan- giate organs is sketchy due to the few reports of anatomically preserved specimens. In general, the remains from this time period are not synan- giate pollen organs and tend to be more similar in superficial appearance to the sporangia of Devonian progymnosperms. Fossils of Mississippian age are useful in demonstrating several trends that lead to the diverse organizations evident in Pennsylvanian seed fern families.

PALEOZOIC SEED FERNS 3~
Advances in our understanding of lyginopterid pollen organs are pro- vided by the description of Feraxotheca (Millay and Taylor, 1977, 1978), which appears similar in form to the widespread compression genus Cros- sotheca, and the description of new Telangium specimens by Jennings (1976). At the present time these taxa encompass the known morpholog- ical variability for pollen organs of this family. In a recent study by Dennis and Eggert (1978) several new ideas are advanced that appear to satis- factorily explain the basic organization and homologies that exist among several anatomically preserved medullosan pollen organs. These ideas have served as a framework from which to evaluate the morphology of all pollen organs currently associated with this family. Although a rela- tively small family in numbers of species, the Callistophytaceae consti- tutes an interesting seed fern complex owing to the manner in which the pollen organs are borne, and the highly evolved nature of the pollen.
It is the intent of this paper to trace the development of microsporan- giate organs from probable Devonian ancestors to the three well-recog- nized families that attained prominence during the late Carboniferous. In so doing we will review in detail those Mississippian specimens that dem- onstrate transitional forms showing basic evolutionary trends in pollen organ phylogeny. In addition, the recent description of several permin- eralized forms of Pennsylvanian age provides an opportunity to reinter- pret the morphology of a number of genera based on compression remains lacking structure.
EARLY PTERIDOSPERM MICROSPORANGIA
The phylogenetic fusion of elongate sporangia into synangia is a pro- cess that probably occurred several times in the past. The possession of synangia is a diagnostic feature of both seed ferns and marattialean ferns, and it is probable that early members of both groups lacked synangia, but were in the process of aggregating sporangia into synangial complexes during the Late Mississippian. The stages involved in the origin of seed fern (Halle, 1933) and marattialean synangia (Millay, 1979) have been discussed to some extent in the literature, but warrant review in the light of recent fossil evidence.
Origin of Synangia
Pteridosperm pollen organs are generally borne on variously modified branching systems that may become quite frondlike. Rarely are synangia borne on laminar ultimate foliar segments. In marattialean ferns, synangia were probably associated with pinnulelike structures early in the evolu- tion of the group. In both orders the ancestors probably had branched fertile regions with sporangia occurring singly or in pairs at the apices of

308 THE BOTANICAL REVIEW
f Fig. 1. A-C, Psilophyton dawsonii sporangia in transverse section; A, dehisced pairs
showing inward facing dehiscence slits, • 17; B, immature sporangial pairs showing differ-

PALEOZOIC SEED FERNS 309
dichotomizing branches. There are numerous Devonian plants currently classified as progymnosperms and pre-ferns that show these early stages in sporangial aggregation. For example, the Upper Devonian taxon Rha- cophyton ceratangium Andrews and Phillips (1968) is characterized by a rachis bearing laterals alternately that in turn produce pairs of sterile and fertile pinnae. Fertile pinnae are profusely dichotomized, with the ultimate pinnae producing one to several sporangia. Sporangia are elon- gate, terminate in a narrow tip, and lack evidence of an annulus. Other possible progymnosperms from the Middle and Upper Devonian show more planation of the ultimate fertile pinnae. In Tetraxylopteris schmidtii (Beck) Bonamo and Banks'(1967) the primary fertile axis bears laterals oppositely-suboppositely that are twice dichotomized. Each resulting branch is three times pinnate in the same plane with the ultimate divisions bearing the sporangia. ReUimia thomsonii Bonamo (1977) is morpholog- ically similar except that the main axis bears secondary units helically. In both genera the elongate sporangia are borne in pairs because the last dichotomy produces two short sporangial stalks. In these taxa the spo- rangia exhibit longitudinal dehiscence and may be extended into a short apiculate tip. The aggregation of the sporangia into radial clusters, and ultimately into synangia, is easily envisioned by only a slight rearrange- ment of the branches. Telescoping of ultimate and penultimate branches, in combination with overtopping, would create either a monopodial or dichotomous system in which sporangia are clustered terminally on branches.
An important contribution by Banks et al. (1975) on the Lower De- vonian trimerophyte Psilophyton, demonstrates an interesting feature of these early sporangia. In P. dawsonii the sporangia are rather thick walled (Fig. 1C), and are borne in pairs at the tip of a branch dichotomy, with the dehiscence slits facing one another (Fig. 1A). In one example, four sporangia belonging to a slightly telescoped branch pair illustrated in transverse section (Fig. IB), show how a ring of sporangia with inter- nally directed dehiscence slits could have evolved. If dehiscence slits develop on the side of a sporangium that faces neighboring sporangia, dehiscence will be toward the center of the group when three or more sporangia are involved.
<--
ential thickness of walls; C, sporangial wall, x90. D, G, Protopitys scotica; D, sporangium in transverse section, • G, recurved fertile branches with terminal sporangia, x7. E, F, I-!, Alcicornopteris hallei; E, fertile branches x0.75; F, dehiscence area of sporangium wall, x 112: It, fertile branches with numerous sporangia in transverse section, x7.0 I, Crossotheca hughesiana, fertile pinna, x4.0. (Figures drawn from: A--C, Banks et al., 1975; D, G, Walton. 1957; E, F, H, Walton, 1949; I, Kidston, 1906.)

310 THE BOTANICAL REVIEW
Some early examples of possible seed fern microsporangiate organs from the Lower Carboniferous appear similar to some of the previously discussed progymnosperm and pre-fern types. These "pollen organs" are considered problematical for two reasons. First, they have not been found in connection with known seed fern stems, however, most anatomical evidence available suggests a pteridosperm affinity. Second, the micro- sporangiate fossils in most instances are not synangiate and therefore not pollen organs, but appear to consist of pairs of terminally borne sporan- gia. It should be noted that in the absence of organic attachment of these microsporangiate remains to known seed plants, some investigators still prefer to consider the plants as progymnosperms (e.g., Beck, 1976).
Alcicornopteris.--The genus Alcicornopteris Kidston (1887) was orig- inally established for distinctive appearing dichotomizing frond segments that were either sterile or fertile. Walton (1949) subsequently included A. hallei in the genus, based on an anatomically preserved specimen that showed a continuous series of curved dichotomies (Fig. 1 E) characteristic of the generitype A. convoluta. Sporangia are borne on stout pedicels that appear to be clustered on the sides or at the extremities of the rach- ides. Sporangia are elongate (possibly up to 1.0 cm), exannulate, and thin walled (Fig. 1H). The sporangial walls appear to be only a single cell in thickness with the outer and anticlinal cell wails considerably thickened (Fig. IF). Sporangia show a tendency to break longitudinally, and some demonstrate more than a single break in the wall. Walton (1949) acknowl- edges that little about the sporangia appears pteridospermous, and relies on anatomical features of the frond axes to support a seed fern affinity. Walton (1949) indicates that the presently accepted forms of Alcicornop- teris probably represent limited segments of the branching microspo- rangiate parts of different genera of early seed ferns. If this suggestion is accurate it is interesting that these early seed ferns possessed sporangia with very thin walls.
An additional species was added to Alcicornopteris (A. zeilleri) by Vaffier (1901) from Lower Carboniferous specimens from Scotland. In A. zeilleri the dichotomies are not closely spaced, and the sporangia (1.0 cm long) are regarded as borne on the periphery of an expanded pinna apex. Although referred to as a synangium (Kidston, 1924), this aggregation of free sporangia only simulates the appearance of a synangium. Because of the branching pattern this species was transferred to the pteridosperm Calathiops by Gothan (1927). The species should now probably be trans- ferred to Paracalathiops, since Calathiops is now restricted to ovulate fructifications (Benson, 1935).
The trilete, granulate spores of Alcicornopteris hatlei were described by Walton (1949) as variable in size depending on their location in the sporangial mass (62-91 /xm). Walton also noted that the spores tend to

P A L E O Z O I C S E E D FERNS 311
have an outer, easily separated wall layer comparable to a perine. Smith (1962a) studied these spores in detail and observed variability in size (50- 130 ~m) and ornamentation. Spores with a thick exine exhibit an irreg- ularly punctate ornamentation with the arms of the trilete suture quite long.
Geminitheca.--Both cupulate ovules and microsporangia are placed in Geminitheca scotica Smith (1959), a taxon known from the Lower Car- boniferous (Cementstone Group) of Scotland. Microsporangiate branches were correlated with the ovulate axes by the similarity of epidermal fea- tures including stomata and numerous hair bases. In addition, micro- spores present in the ovule pollen chambers are similar in size and mor- phology to those contained in the microsporangia. Sporangia are borne in terminal ~'bunches" on a dichotomizing axis. Each cluster was thought to contain about 16 elongate sporangia each approximately 1.5 mm long and 0.5 mm in diameter. Sporangia are free, and borne terminally in pairs due to the apical dichotomy of the branches. The sporangial walls are at least two cells thick and there is no indication of a dehiscence mode. Trilete microspores (45-58 ~m) are circular to roundly triangular in polar view and possess a narrow equatorial ridge (cingulum).
The microspores of Geminitheca show two rather consistent features present in, and possibly characteristic of, both progymnosperms and ear- ly seed ferns. These features include the trilete suture through which proximal germination presumably occurred, and the tendency of the exine to separate to form a girdling saccus. Presently known progymnosperm microspores are larger and have a more irregular separation of the exine (pseudosaccus).
The description of Geminitheca is based on both compression and permineralized specimens. The presence in the Lower Carboniferous of a seed fern bearing microsporangiate branching systems similar to those of Devonian progymnosperms is interesting, and supports previous spec- ulation concerning the origin and homologies of seed fern synangia.
Protopitys.--This genus is named for Lower Carboniferous stems of variable size that may include specimens with abundant secondary xylem (Goeppert, 1850). A small stem of Protopitys (P. scotica) bearing fertile branches alternately was described by Walton (1957). Each fertile branch dichotomizes several times (Fig. IG), with the ultimate branches arranged pinnately and bearing elongate sporangia (3.0 mm long). Sporangia exhibit longitudinal dehiscence and have a two-parted wall consisting of large epidermal cells and an inner layer one to two cells thick (Fig. 1D). Spo- rangia terminate in an elongate beak and possess stomata.
Spores of P. scotica are spherical, range from 82-163 kLm in diameter and have a symmetrical trilete suture, with the arms extending �89 the spore radius. Although the spores are described as smooth walled, a
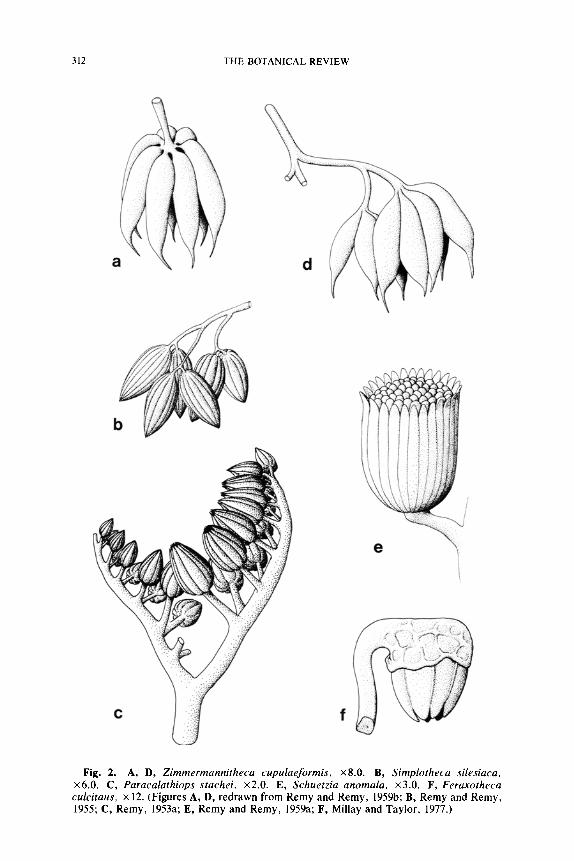
312 THE BOTANICAL REVIEW
Fig. 2. A, D, Zimmermannitheca cupulaeformis, x8.0. B, Simplotheca silesiaca, x6.0. C, Paracalathiops stachei, x2.0. E, Schuetzia anomala, x3.0. F, Feraxotheca culcitaus, x 12. (Figures A, D, redrawn from Remy and Remy, 1959b; B, Remy and Remy, 1955; C, Remy, 1953a; E, Remy and Remy, 1959a; F, Millay and Taylor, 1977.)

PALEOZOIC SEED FERNS 313
granular substance was present that appears similar to the perispore de- scribed in other early seed ferns with nonsynangiate fertile parts.
Staphylotheca.---Staphylotheca kilpatrikensis Smith (1962b) was de- scribed from fragmentary material in Lower Carboniferous rocks. The genus is represented by dichotomizing axes that have bunches of sterile linear structures (0.5 x 10.0 mm) borne on the lower surface. The short sporangia (0.7 x 1.0 mm) are apparently borne on these sterile structures. Little detail was preserved in this compression specimen and it has there- fore been difficult to determine the identity of the sterile structures as branches. Smith (1962b) suggests that the sterile processes may be anal- ogous to a rudimentary "cupule," which implies they functioned in pro- tecting the sporangia rather than merely bearing them. Such an organi- zation, if accura te , appears somewhat intermediate be tween Alcicornopteris which is completely fertile, and Paracalathiops which may have an outer zone of sterile protective units.
The spherical microspores of Staphylotheca are large (78-100 ~m, av. 87 ~m), thick walled, and have a symmetrical trilete suture. The exine is thick (5 ~m) over most of the spore surface, but thins at the proximal pole (3 ~m). The subtle exine sculpturing present consisted of internal punctation of the outer wall layer.
Paracalathiops.--Other types of possible pollen organs of Upper Mis- sissippian age (Namurian A) appear similar to the previously discussed nonsynangiate types, but are still highly problematical. One of these is Paracalathiops Remy (1953a). The morphological organization of this pollen organ is poorly understood because it is known only from compres- sions. It consists of a dichotomizing branching system, with the ultimate axes borne in pairs on the same side of the penultimate branch (Fig. 2C). The synangia are approximately 0.7 mm long, and appear to consist of an outer zone of sterile appendages that enclose numerous tubular spo- rangia. Microspores of Paracalathiops are large (210-255 ~m), saccate, and have a symmetrical trilete suture with arms extending �89 the radius of the central body. The air sac (saccus) is irregularly formed around the spherical corpus and lacks endoreticulations. Ornamentation is granu- lose. The taxon may represent either a cluster of elongate, basally fused sporangia or a hollow ring of sporangia.
Simplotheca.--Another problematical form from Namurian A sedi- ments is Simplotheca silesiaca Remy and Remy (1955). In Simplotheca the small (2.0-2.2 mm long), spindle-shaped organs are borne terminally on very narrow dichotomizing axes (Fig. 2B). The general morphology appears similar to Aulacotheca, however, in Simplotheca the internal organization in unknown. The genus was originally described as a simple sporangium, but the form is equally suggestive of a synangium. No spec- imens have been found that exhibit an apical opening to suggest the unit was hollow.

314 THE BOTANICAL REVIEW
Microspores of Simplotheca are saccate and appear similar to the dis- persed spore Schulzospora. They possess an elliptical saccus (up to 150 /~m long) that encloses a spherical central body 60-75 /~m in diameter. A symmetrical trilete suture is present, and endoreticulations of the sac- cus are absent (Fig. 18C).
Zimmermannitheca. Zimmermannitheca cupulaeformis Remy and Remy (1959b) is the binomial for a sporangial aggregation of lower Na- murian A age. In this genus the sporangia are aggregated at the apices of ultimate divisions of a dichotomizing branching system. Sporangia are elongate (2.5-5.0 mm long) with narrow attenuated apices (Fig. 2A, D). The number of sporangia in a cluster varies from 2-7, and the compact- ness of the unit is also variable. No mode of dehiscence has been iden- tified. Microspores of Zimmermannitheca are spherical to oval and thick walled. The large spores (65-125/~m) possess a prominent trilete suture and exhibit a granular to verrucate ornamentation.
The last three pollen organ types represent a closer and more regular association of the elongate sporangia into synangiumlike structures. Zim- mermannitheca is not synangiate, but in the character and arrangement of the sporangia (Fig. 2A) and branches it is very similar to Telangium. Only a small amount of basal sporangial fusion is required in Zimmer- mannitheca to simulate the Telangium appearance. Both Simplotheca and Paracalathiops are probably synangiate, but their detailed organi- zation is not known. Either taxon could be a closed ring of sporangia like the Pennsylvanian pollen organ Aulacotheca, and this interpretation is favored here because radial arrangements are widespread and functional in the medullosan seed ferns. A synangiate '~cluster" of sporangia, unless very lax, seems unnatural because any proposed arrangement must make provision for longitudinal dehiscence of the component sporangia. The morphology of the microspores also suggests that Paracalathiops and Simplotheca are early medullosan pollen organs, since prepollen with the same gross morphology and saccus type is also known in the medullosans (Parasporotheca). The microspores of Zimmermannitheca appear un- specialized and resemble those of Potoniea and most lyginopterid seed ferns.
L Y G I N O P T E R I D A C E A E
The iyginopterid seed ferns are probably the most poorly understood group of early seed plants despite their important position in the estab- lishment of the pteridosperms. Little has been accomplished in the reconstruction of whole plants in this family, and most information is based on an assortment of sterile and fertile organs. In addition, the familial concept is poorly defined and almost nothing is known of evo-

PALEOZOIC SEED FERNS 315
lution within this group. The paucity of Mississippian remains of seed fern families other than the Calamopityaceae and Lyginopteridaceae is suggestive that at this time the families represented an artificial complex that was ancestral to later seed fern families.
A number of plant organs have been described that together serve to characterize or typify the family. Only a single plant (Lyginopteris) has thus far been reconstructed in this family. Following is a listing of plant parts associated with the Pennsylvanian lyginopterid seed ferns.
Stems Petioles Foliage
Heterangiurn Lyginorachis Sphenopteris Lyginopteris Microsperrnopteris Schopfiastrurn
Seeds Pollen Organs Prepollen
Conostoma Canipa Cyclogranisporites Coronostoma Crossotheca Granulatisporites Lagenostoma Feraxotheca Physostoma Telangiopsis Tyliosperma Telangium
Branching Patterns
Until recently almost nothing was known about lyginopterid pollen organs. Although occasionally found in the compressed condition, ana- tomically preserved specimens are extremely rare. Compression speci- mens are valuable in showing the general morphology of the fertile pin- nae, but anatomical information is critical for making meaningful comparisons. Presently, only two types of pollen organs are known for this family, and are borne in distinctive ways on the plant. Genera such as Telangium Benson (1904) or Telangiopsis Eggert and Taylor (1971) are borne on monopodial branching systems. In some instances the entire branching system appears to be three-dimensional, and may represent a portion of an otherwise planated vegetative frond. In other cases it could conceivably replace an entire leaf on the stem. Jennings (1976) has shown that Mississippian specimens of Telangiurn consist of pollen organs that are terminal on small dichotomous branching systems that appear to re- place penultimate pinnae (Fig. 4). These three-dimensional units are borne alternately along otherwise planated frond units. In Telangiopsis Eggert and Taylor (1971), also of Mississippian age, no portion of the branching system appears to be planated (Fig. 15F). The fertile frond segments of
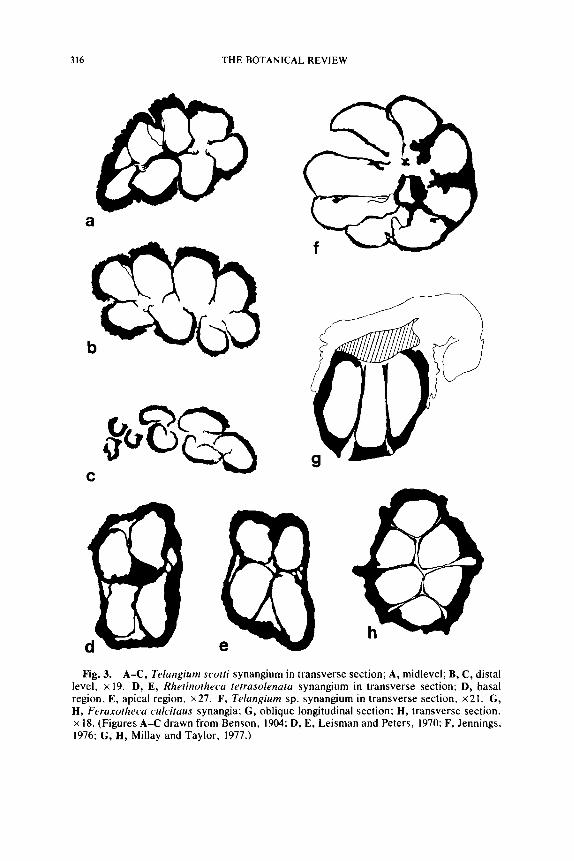
316 THE BOTANICAL REVIEW
a
C
Fig. 3. A-C, Telangium scotti synangium in transverse section; A, midlevel; B, C, distal level, • 19. D, E, Rhetinotheca tetrasolenata synangium in transverse section; D, basal region, E, apical region, x27. F, Telangium sp. synangium in transverse section, • G, H, Feraxotheca culcitaus synangia; G, oblique longitudinal section; H, transverse section. • 18. (Figures A-C drawn from Benson, 1904; D, E, Leisman and Peters, 1970: F, Jennings, 1976; G, H, Millay and Taylor, 1977.)
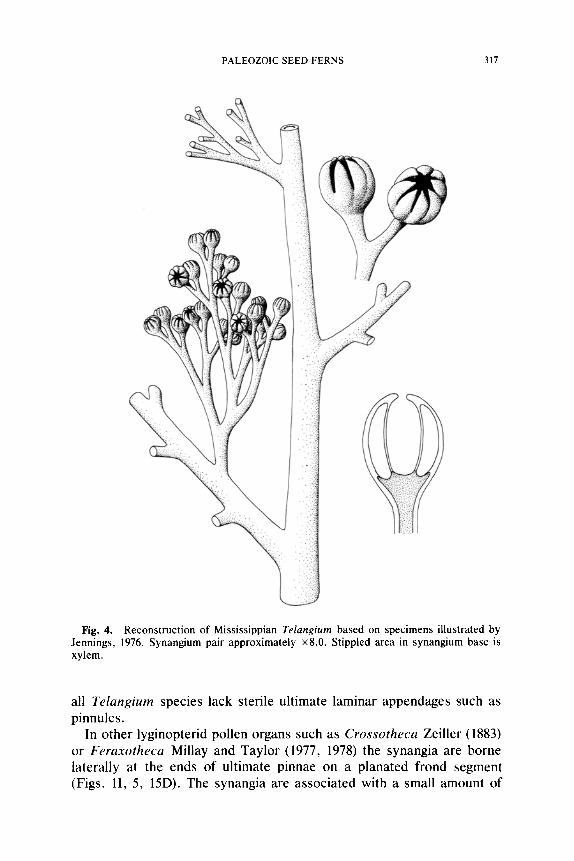
PALEOZOIC SEED FERNS 317
Fig. 4. Reconstruction of Mississippian Telangium based on specimens illustrated by Jennings, 1976. Synangium pair approximately x8.0. Stippled area in synangium base is xylem.
all Telangium species lack sterile ultimate laminar appendages such as pinnules.
In other lyginopterid pollen organs such as Crossotheca Zeiller (1883) or Feraxotheca Millay and Taylor (1977, 1978) the synangia are borne laterally at the ends of ultimate pinnae on a planated frond segment (Figs. lI, 5, 15D). The synangia are associated with a small amount of
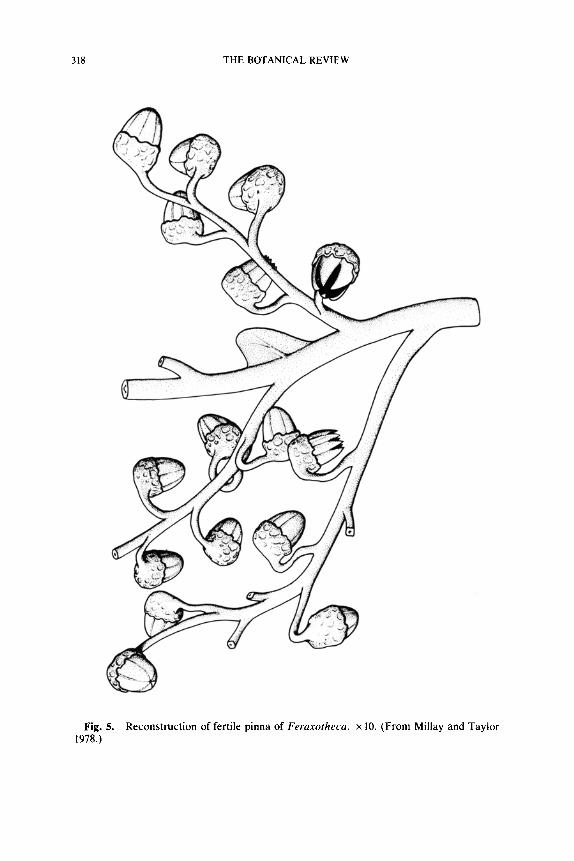
318 THE BOTANICAL REVIEW
Fig. 5. Reconstruction of fertile pinna of Feraxotheca. • 10. (From Millay and Taylor 1978.)

PALEOZOIC SEED FERNS 319
foliage at their bases, and normal pinnules and axial pinnules (=Zwi- schenfiedern) generally occur in nonfertile portions of the same frond. In Crossotheca the synangia may be located at or near the tip, in the middle, or near the base of otherwise vegetative pinnae. In general, the pollen organ location shows the same range of variability known for supposed lyginopterid ovules and cupules. Presumably, the three-dimensional branching systems represent the primitive condition, and increasing plan- ation is a later derived condition. It is interesting to note that the ultimate synangium-bearing axes in both Feraxotheca and Telangium are rect- angular in transverse section, possibly reflecting their original branchlike nature.
Permineralized Specimens
A complete analysis of the anatomically preserved specimens is espe- cially important in this group since the fossils are so rare. Specimens assigned to Telangium Benson (1904) are known from the Westphalian A (T. scotti Benson) of Great Britian and the Upper Mississippian (Te- langium sp.) of the United States (Jennings, 1976). These specimens ap- pear comparable in most basic respects, but differ in minor features that may be related to preservational phenomena. In Telangium scotti the synangia measure approx. 1.7 mm in maximum diameter and are com- posed of eight elongate, thick-walled sporangia that are basaily fused. Each synangium consists of two rows of four sporangia, that give this region of the organ a bilateral symmetry (Fig. 3A, B). The bilateral sym- metry of Benson's specimen is convincing, but because no additional specimens of this species have been studied in serial section it can not be determined that preservational factors were not the cause of the ap- parent bilateral configuration. An examination of isolated specimens sim- ilar to T. scotti (Fig. 17C) suggests that the sporangia are in fact arranged in two rows, and that the synangium is bilaterally symmetrical. The synangia in T. scotti, however, do not appear to differ in basic structure, or in the way they functioned, from the radially symmetrical synangia of Telangium described by Jennings (1976). In T. scotti the outward-facing sporangial walls appear thick (perhaps 5 or more cells thick) with an epidermis of large cells. A considerable amount of parenchyma was ap- parently present in the synangial base, however, judging from the avail- able illustrations, the tissue is histologically similar to that present in the sporangial walls. Each sporangium has ~'transfusion tissue" along the midline of the outer facing walls that may have functioned as vascular tissue.
It is important to determine the method of sporangial dehiscence in pollen organs. Dehiscence in Telangium is longitudinal along the midline

320 THE BOTANICAL REVIEW
of the inner facing sporangial walls (Figs. 3B, C, 17C). The dehiscence suture appears to extend for most of the length of the sporangium, even though the sporangial walls remain laterally continuous for V2 or more of the synangial length. If the sporangia do not surround a central hollow area this implies that the sporangia open into one another at basal levels (Fig. 3B). There is little evidence in T. scotti to suggest that the sporangia laterally break away from one another as a part of dehiscence such as is characteristic of some marattialean fern synangia. Some compression specimens of Telangiopsis, however, clearly give the impression that the sporangia spread widely apart on dehiscence (Eggert and Taylor, 1971).
Some anatomically preserved Mississippian specimens of Telangium appear to differ from T. scotti in several important details (Jennings, 1976). These new specimens are radially symmetrical with eight sporangia surrounding a large central hollow area (Fig. 4). Sporangia are up to 1.0 mm long and appear in most specimens to be laterally free for about z/3 of their length. Sporangia are attached to a vascularized parenchymatous core that Jennings (1976) compares favorably with the basal region in T. scotti. The sporangial walls are rather thin in the Mississippian Telan- gium, perhaps only two cells thick on the outer facing walls, and unise- riate on the inner facing walls (Fig. 3F). This feature, together with the probable difference in synangial symmetry, constitute the significant dif- ferences between these, the two most completely understood specimens of Telangium.
Features dictated by preservation and the techniques used to study the material of Telangium scotti make it difficult to compare the taxon in a meaningful way with the Mississippian specimens. The apparent bilateral symmetry, and extremely thick sporangial walls of T. scotti may be due to vagaries of preservation and illustration. It seems likely, however, that the delicately constructed, radially symmetrical synangia of Chester (Up- per Mississippian) age were antecedents to the thick-walled bilaterally symmetrical material from Westphalian A sediments (Lower Pennsylva- nian equivalent).
The other general type of lyginopterid pollen organ has been histori- cally described as associated with the reduced laminar tissue of a pinnate frond system. The common Pennsylvanian compression fossil Crosso- theca Zeiller (1883) represents this pollen organ type, and the genus Fer- axotheca Millay and Taylor (1977, 1978) may in fact represent the same pollen organ preserved structurally. Because of certain preservational parameters, specimens of Crossotheca have been interpreted in a variety of ways (Arnold and Steidtmann, 1937; Danzr, 1955, 1960; Kidston, 1923). Feraxotheca is valuable in demonstrating the probable anatomical construction of this lyginopterid pollen bearing organ. The synangia of Crossotheca are borne on the side of an expanded ultimate pinna, and

PALEOZOIC SEED FERNS 321
some irregular laminar tissue is produced by the pinna axis that is closely associated with each synangium base (Figs. 2F, 3G). This foliage is not very extensive in the permineralized specimens so it is surprising that a reduced lamina is frequently discerned as being present around the synan- gia in compression specimens of Crossotheca.
The synangia of Feraxotheca demonstrate a pattern of organization that is unlike other known synangia. The base consists of a hemispherical or cone-shaped structure composed of thick walled parenchyma (Fig. 3G). Sporangia arise from the flat surface of the pad and are fused to one another at the center and along the lateral surface (Fig. 3H). Outer facing sporangial walls are thick (more than 5 cells) and lack any indication of internal zonation. Inner facing walls are uniseriate, and longitudinal de- hiscence is effected by the rupture of smaller cells along the midline. Symmetry of the synangium may be radial or bilateral depending on the number of sporangia present (Millay and Taylor, 1978). In this paper we extended the known structure of Feraxotheca to the morphologically more problematical species of Crossotheca including C. sagittata and C. trisecta. In these species the synangia have the form of an arrowhead, with the three pointed segments attached basally (Fig. 13C). The distinc- tive shape of the C. sagittata synangium probably originated from the basal fusion of three oval synangia on a small pinnate frond segment, rather than from branching of the synangium. For this reason the pollen organ of C. sagittata should probably be considered a compound syn- angium.
The correct arrangement of sporangia in C. sagittata was first de- scribed by Arnold and Steidtmann (1937). Their reconstruction showing the arrangement of the sporangia is reproduced in Fig. 13G, where the thick outer facing walls of the sporangia are not so illustrated. This ar- rangement is like that present in Feraxotheca where the inner facing sporangial walls are closely appressed so that there is no open space within the synangium. The odd shape of the synangium apparently re- sulted in tightly packed sporangia that could have 3-5 sides depending on the number of sporangia in contact. Before sporangia became laterally free, cells between sporangia in the outer facing walls must break down. The rupture of the inner facing walls was induced by the outer sporangial walls pulling away due to drying.
Synangia of Feraxotheca are vascularized by a pinna vascular bundle that spreads out over the surface of the synangial pad. No vascular tissue is present in the pad or in the sporangial walls. The synangia of Ferax- otheca and Telangium are not similar and the uncertainty over some basic features of T. scotti make additional comparisons difficult. The basic organization of these pollen organs consists of a ring of sporangia with inwardly directed dehiscence slits, and a synangium base construct-

322 THE BOTANICAL REVIEW
ed of parenchyma. In Feraxotheca the parenchyma pad seems to form a structural entity on which the sporangia are attached, whereas the parenchyma in the other forms is difficult to distinguish from the sporan- gial bases. Vascularization seems to support this interpretation because the outer surface of the pad is vascularized in Feraxotheca, whereas in Telangium the vascular tissue passes centrally through the base, and in some forms, into the sporangia. Dehiscence in all taxa involves either partial or complete lateral separation from neighboring sporangia. This feature is poorly known for Telangium and is supported principally by the appearance of compressed dehisced synangia of Telangiopsis. Syn- angial dehiscence is considered to be an important criterion in assessing relationships among pollen organs in a similar way to that demonstrated for the synangia of fossil and extant marattialean ferns (Millay, 1976, 1978).
Prepollen 1
The microspores of all presently acknowledged lyginopterid seed ferns appear remarkably similar. All may be characterized as small, spherical- ovoid (41-71 ~zm diameter), and either trilete or monolete. Exine orna- mentation is simple, consisting of numerous, evenly distributed papillae, coni or grana (Fig. 18B). In one species of Crossotheca the grains possess a thin inner exine layer (nexine) and a thick (1.8/zm) outer zone (sexine). The suture represents an outfolding of a thin region in the sexine, and resembles closely the sutures in the isospores of homosporous ferns (Millay et al., 1978). The presence of an apparently functional proximal suture on these grains, together with the absence of a distal aperture, suggests that these seed plant microspores should be considered func- tionally as prepollen (Millay et al., 1978).
Evolutionary Trends
Evolutionary trends in the pollen organs of lyginopterid seed ferns are not clearly evident from the limited fossil evidence available. There would appear to be a progressive modification of the fertile branching system from the three-dimensional form present in Upper Mississippian Telan- giopsis to the partially planated type seen in Telangium (Jennings, 1976). It is important to emphasize that the synangia of Telangium are borne on a three-dimensional branching system that theoretically represents either a portion of an otherwise sterile planated frond, or a unit that occupies the place of a sterile frond. Pollen organs of this type are prob-
i Prepollen is present in those seed plants in which microgametophyte germination ap- pears to have been proximal, as is characteristic of isospores and microspores (Chaloner, 1970).

PALEOZOIC SEED FERNS 323
ably always borne on a frond or frond segment. Such a series might culminate in the regularly pinnate fertile fronds observed in Canipa Skog et al. (1969), and the partially sterile pinnate frond segments of Crosso- theca and Feraxotheca known from the Pennsylvanian.
Based on their geological occurrences and the frond system, one might speculate that the Feraxotheca synangium is derived from the Telangium type. If this series is accurate, sporangia of Telangium would have to become more closely appressed centrally (e.g,, T. scotti), with the syn- angium twisting basally in order to occupy the side of the pinna apex. It should be noted however, that the synangium position in both genera could reflect the orientation of the branching system. In an upright branching system the synangia are erect, whereas in a horizontal frond- like system the synangia are pendulous in orientation.
The presence of thick sporangial walls and a vascular system in T. scotti may be a response to the large size and exposed nature of the synangia, however, this sort of reasoning breaks down when applied to the slightly smaller Mississippian Telangium.
The uniform morphology of lyginopterid prepollen may accurately demonstrate the level of pollen evolution in this group, or may only reflect the fact that few types of pollen organs have been identified with this family. There are some previously discussed pollen organs of Mississip- plan age with saccate microspores (e.g., Paracalathiops, Simplotheca) that may represent lyginopterids, but most of these are more likely to have affinities with the medullosan seed ferns.
MEDULLOSACEAE
The Medullosaceae is a large and diverse family of seed ferns with a macrofossil record extending from the Upper Mississippian through the Permian. The various plant parts referable to this family are common fossils in the Late Paleozoic and have many characteristic anatomical features that make assignment to this family possible. Several well-known examples of plant organs belonging to this family are listed below.
Stems Petioles Foliage
Medullosa Myeloxylon Alethopteris Sutcliffia Neuropteris
Seeds Pollen organs Prepollen
Albertlongia Aulacotheca Parasporotheca Monoletes Hexapterospermum Boulaya Potoniea Parasporites Pachytesta Codonotheca Rhetinotheca Sarcospermum Dolerotheca Schopfitheca Stephanosperrnum Goldenbergia Stewartiotheca
Halletheca Sullitheca Whittleseya
324 THE BOTANICAL REVIEW
0
0
e,,,
_o
E
0
e ~
N
e-, <
X X X J X X X X X X X
O t ' 4 0
I ~"1',,., I ~ ~ I ~ x o
O
O
. h ~. . .

PALEOZOIC SEED FERNS 325
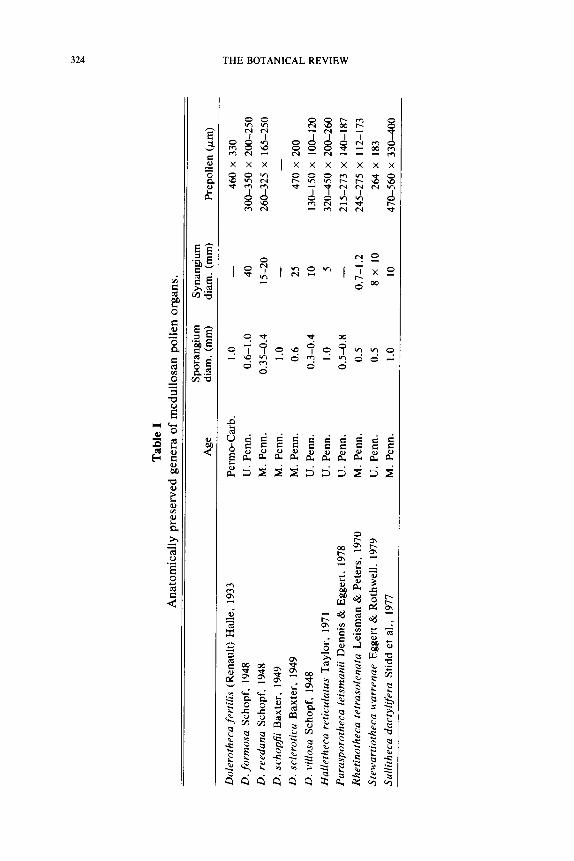
The number of compressed medullosan pollen organs is large, and their recognition is possible based primarily on their distinctive microspore type. There are presently six or seven anatomically preserved medul- losan pollen organs known, and it is clear that little of their complex in- ternal organization would be preserved in a compressed form. The com- pression fossil record has primarily been of importance in determining the manner in which the pollen organs were borne on the plant, and in showing variation in microspore form. It is the permineralized taxa that have provided the most insight into the sometimes complex construc- tion of medullosan pollen organs, and have demonstrated the consistency of most organizational designs. We will first discuss the anatomically pre- served genera so as to develop a framework that will aid in the interpreta- tion of the compression forms. Important features of the permineralized forms are summarized in Table 1.
Permineralized Specimens
Among the permineralized forms there is considerable diversity in pol- len organ morphology and anatomical complexity. This diversity includes small synangia that are solitary or aggregated, and large synangia that may be aggregated, or ontogenetically fused into compound synangia. One small radially symmetrical type that has been described under the name Halletheca Taylor (1971) was probably borne singly at the tips of branches. No specimens have been found with the epidermis intact, and therefore the expected presence of hairs on this synangium remains un- known. Synangia of Rhetinotheca Leisman and Peters (1970) are small and anatomically simple like those of Halletheca, but are characteristi- cally borne in small clusters. This mass of pollen organs was held together in early developmental stages by small, peglike, "interlocking" hairs pro- duced on the epidermis of each synangium. In the compressed condition such an aggregation of synangia would probably simulate the appearance of a larger pollen organ like Potoniea. Another example of aggregate synangia is provided by Parasporotheca Dennis and Eggert (1978). In this form the synangia do not appear radial, but consist of a curved sheet of. elongate sporangia. These fan-shaped synangia are wrapped around one another and are held together by interlocking hairs. Dolerotheca Halle (1933) and Potoniea (Stidd, 1978b) represent examples of com- pound synangia. Dolerotheca apparently represents a row of four, large, radial synangia that are highly convoluted and fused together early in development. On the other hand, permineralized Potoniea specimens consist of concentric rows of small radial pollen organs that have appar- ently grown together during development. It is clear that there may be simple, aggregate or compound synangia within this seed fern complex,

326 THE BOTANICAL REVIEW
and that except in one example (Parasporotheca), the smallest synangial unit is always clearly radial in symmetry. A detailed comparison of each genus is necessary to show the remarkable series of homologies that exist in the construction of the synangia, or synangial components, and the important features of dehiscence demonstrated by each taxon.
Simple Synangia
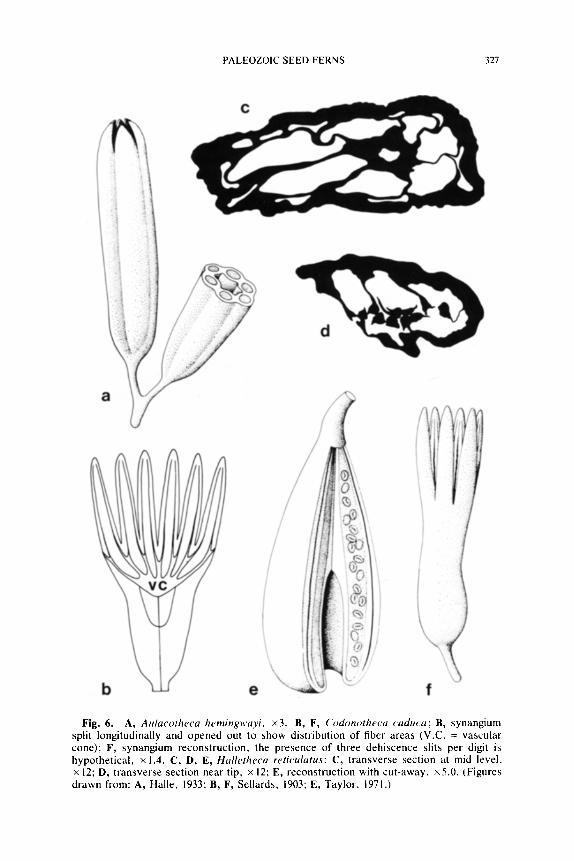
Halletheca.--Synangia of Halletheca reticulatus Taylor (1971) are pyr- iform and measure approximately 1.5 cm long and are up to 5.0 mm in diameter near the apex. Each synangium consists of five sporangia that surround a central zone of fibers near the base and a central hollow area near the distal end (Fig. 6E). The sporangial cavities alternate with large lacunae, and both are embedded in a thick ground tissue (Fig. 6C). Each sporangium is vascularized by a single bundle that traverses the ground tissue along the outer surface of the sporangium. The central fibrous zone of each synangium consists of elongate thick-walled cells. This fiber-area is five-sided in the synangium base, but becomes hollow at the midlevel. In transverse section each sporangium has a triangular-shaped fiber zone forming the inner facing wall of the sporangium. Dehiscence slits form along the midline of these triangular areas of fiber cells (Fig. 6D), and pollen is shed into the central hollow area of the synangium. Halletheca exhibits all of the features characteristic of medullosan synangia including alternating sporangia and lacunae in the ground tissue~ dorsally vascu- larized sporangia, and dehiscence through an internal or ventral area composed of fiber cells.
It would appear likely that Halletheca synangia were borne singly at the apex of small branches much as has been demonstrated in Aulaco- theca (Eggert and Kryder, 1969). The presence of a thick outer ground tissue suggests that synangia were solitary as in Goldenbergia, and the discovery of an epidermis with hairs would add considerable support to this idea. Halletheca lacks the distinctive interlocking pegs and thin outer ground tissue characteristic of aggregate synangia (e.g., Rhetinotheca). We believe the simple structure of the Halletheca synangium embodies the basic form of all medullosan pollen organs, and is probably closely related to the ancestral type.
Stewartiotheca.--The recently described pollen organ Stewartiotheca warrenae Eggert and Rothwell (1979) has an interesting internal construc- tion that appears to be a variation of the basic Halletheca-type of orga- nization. Stewartiotheca synangia are campanulate (approx. 8 • 10 mm in diameter) in form with an eccentrically placed dorsal peduncle. The peduncle is continuous with a core of fiber cells that is solid in the syn- angium base but becomes hollow near the distal face (Fig. 13E). The
PALEOZOIC SEED FERNS 327
Fig. 6. A, Aulacolheca hemingwayi, x3. B, F, Codonotheca caduca; B, synangium split longitudinally and opened out to show distribution of fiber areas (V.C. = vascular cone); F, synangium reconstruction, the presence of three dehiscence slits per digit is hypothetical, x 1.4. C, I), E, Halletheca reticulatus: C, transverse section at mid level. x 12; D, transverse section near tip, x 12; E, reconstruction with cut-away, x5.0. (Figures drawn from: A, Halle, 1933; B, F, Sellards, 1903; E, Taylor, 1971.)

328 THE BOTANICAL REVIEW
numerous (up to 80) tubular sporangia that surround the fiber core are arranged in a uniseriate ring that, during development, has been internally folded or plicated on a radial plan (Fig. 13E). The symmetry is imperfect only because of the eccentric location of the fiber core, that develop- mentally represents the center of the organ. The infolding of the ring of sporangia is therefore more shallow on the side of the synangium occu- pied by the fiber area.
Sporangial tubes alternate with narrow lacunae, and each sporangium was shown to possess a single dorsal vascular bundle (Eggert and Roth- well, 1979). The sporangial tubes appear to occur in pairs with the lon- gitudinal dehiscence slits facing one another across a shallow groove in the distal face of the synangium. The cells of the dehiscence zone resem- ble those of a palisade tissue and are very similar to those present in Dolerotheca (Schopf, 1948).
The general arrangement of structural features in Stewartiotheca is also present in other permineralized taxa of medullosan pollen organs. Stewartiotheca synangia closely resemble several species of Dolerotheca in overall form, dehiscence zone and ground tissue histology, and the similar covering of multiceilular hairs. The principal anatomical differ- ence between these two genera is the presence of a "central" fiber area in Stewartiotheca. A more profound difference is the simply synangiate condition of Stewartiotheca contrasting with the compound synangiate organization of Dolerotheca (Fig. I 1).
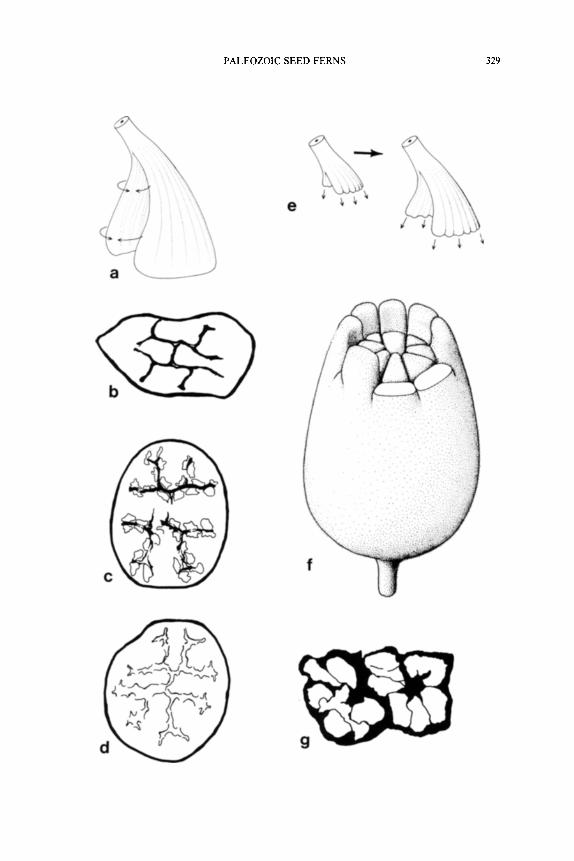
Sullitheca.--Synangia of Sullitheca dactylifera Stidd, Leisman, and Phillips (1977) are obpyriform and measure approximately 3.0 cm long and 1.0 cm in diameter basally where the synangia are largest. The apical one-third of each synangium forms a ring of solid fingerlike projections constructed primarily of the same thick-walled cells that form the thick outer tissues of the pollen organ (Fig. 7F). These projections do not appear to have anything to do with synangial function since the sporangial cavities end near their bases.
Figure 7B-D shows three silhouette diagrams of the pollen organ in transverse section; one through the base (Fig. 7B) and the others near the midlevel and beyond (Fig. 7C, D). Although the fact is not readily apparent from an examination of transverse sections, Sullitheca repre- sents a simple radial synangium. At basal levels the sporangia are ar-
Fig. 7. A, E, Parasporotheca synangia; A, possible origin of radial synangia; E, possible origin of bilateral synangia, x 3,0. B-i) , F, Sullitheca dactylifera; B-D, transverse sections at progressively higher levels, x4.0; F, reconstruction of synangium with sterile fingerlike processes, • G, Potoniea, two radial clusters of sporangia in transverse section, at a level near the distal surface of the organ, • (Figures drawn or redrawn from: A, Dennis and Eggert, 1978; B-D, Stidd et al., 1977; G, Stidd, 1978.)
PALEOZOIC SEED FERNS 329

330 THE BOTANICAL REVIEW
ranged around the periphery of a symmetrically branched area of scle- renchyma. The fiber area is H-shaped centrally in the base, but each of the four arms dichotomizes and extends to near the periphery of the organ (Fig. 7B). This fibrous central core of the synangium is homologous with the five-sided column of fibers present in the base of Halletheca. In Sullitheca, however, the fiber zone is more amplified in a symmetrical fashion to accomodate the large number of sporangia present in the syn- angium. The basic organization of this pollen organ becomes more ap- parent in transverse sections near the midlevel of the synangium (Fig. 7D). At these levels all sporangia dehisce into the common central cavity of the synangium (Fig. 17E). The synangium is convoluted symmetrically, with the sporangia clearly positioned around the periphery of a central cavity that is large in surface area. Stidd et al. (1977) describe the central cavity as originating from median divisions along the arms of the multi- branched sclerenchyma core. This is comparable to the organization of Halletheca in which the sclerenchyma central column becomes hollow and dehiscence areas form around the periphery. The synangium in Sul- litheca is structurally homologous to Halletheca; the more numerous sporangia present in Sullitheca are accommodated by internal convolu- tions on a radial plan. Internal fluting represents the only possible mod- ification for the enlargement of pollen organs in which each sporangium requires interior wall space for dehiscence.
Additional similarities between Sullitheca and Halletheca include the dorsal vascular strand associated with each sporangium, and the thick ground tissue of the synangium. Sullitheca is covered with hairs as would be expected of an exposed pollen organ borne on branches. One prob- lematical feature of Sullitheca is the peripheral ring of vascular bundles in the outer ground tissue of the synangium. Most of these bundles do not appear to be associated with sporangia, and this is unusual because all vascular tissue in other pollen organs (when known) appears to be related to sporangial position. The peripheral vascular system probably has its origin from repeated dichotomies of the inner, sporangial vascular system, and is no doubt necessary to vascularize the physiologically ac- tive outer zone of the organ. This suggestion is supported by the presence of stomata on the epidermis. The peripheral vascular bundles may phy- logenetically represent some secondary enclosing structure that originat- ed from an outer ring of sterile telomes that have fused to the outer surface of the pollen organ. This will have to remain in the realm of speculation until adequately preserved, stratigraphically older specimens are discovered.
It should be noted that the interpretation of Sullitheca as a simple synangium was proposed by Dennis and Eggert (1978), and is at variance with the compound synangium interpretation of Stidd et al. (1977). Ac-

PALEOZOIC SEED FERNS 331
cording to these latter authors each small cluster of sporangia (4-6) that is positioned around the periphery of the Sullitheca organ phylogenet- ically represents a solitary synangium. In each of these small "pollen organs" the sporangial vascular bundles are located in ground tissue on the inside of the "organ" and the sclerenchyma and dehiscence slits face outward. Such an organization is like a Halletheca synangium turned inside out, and is not regarded by us as plausible.
The sterile fingerlike projections of Sullitheca synangia are an inter- esting feature that does not have a parallel in other anatomically pre- served synangia. The fingers do not appear to represent the sterile rem- nants of once fertile regions, but may instead simply function to seal off the open system of grooves from desiccation during early developmental stages. They are fibrous extensions of both the outer synangial wall and the inwardly directed dehiscence areas. The fingers occur between de- hiscence grooves and represent extensions of a small group of sporangia with outward facing dehiscence slits (Fig. 7F). To some extent this is what led Stidd et al. (1977) to conclude that these sporangia were grouped naturally, each representing a synangium.
If found compressed, a Sullitheca synangium would not be interpreted as anything close to its true morphology. Rather, it would be considered a pyriform synangium with 9-10 large sporangia (2.0 mm in diameter) that were fused basally and free distally for a short distance. The presence of approximately 40 sporangia in a complex arrangement could scarcely be determined in a compression. Perhaps other pollen organ compression genera of similar form such as Codonotheca or Schuetzia are in need of a serious reevaluation with regard to the potential complexity of their morphology.
Aggregate Synangia
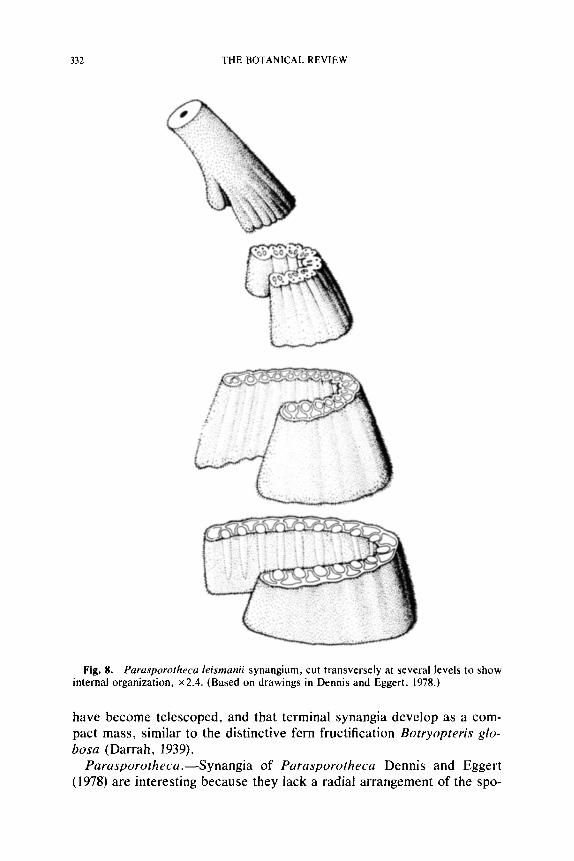
Rhetinotheca.--Synangia of Rhetinotheca tetrasolenata Leisman and Peters (1970) consist of four sporangia that surround an area of fiber cells at the base (Fig. 3D), and a central hollow region (Fig. 3E) at the distal end (Dennis and Eggert, 1978). Each synangium measures 2.0-3.6 mm in length and 0.7-1.2 mm in diameter, with the sporangial cavities alternat- ing with lacunae in a rather thin ground tissue (Fig. 3D, E). No vascular tissue has been identified in the outer sporangial walls. The sporangia appear to dehisce longitudinally and release prepollen into the distal hol- low region of the synangium. Preservation of the inner sporangial walls is not sufficient so as to demonstrate the cell arrangement that allows dehiscence. The epidermis of Rhetinotheca is covered with evenly spaced, peglike hairs that interdigitate with those on the surface of neigh- boring synangia. It appears clear that in Rhetinotheca the fertile branches
332 THE BOTANICAL REVIEW
Fig. 8. Parasporotheca leismanii synangium, cut transversely at several levels to show internal organization, • (Based on drawings in Dennis and Eggert, 1978.)
have become telescoped, and that terminal synangia develop as a com- pact mass, similar to the distinctive fern fructification Botryopteris glo- bosa (Darrah, 1939).
Parasporotheca.--Synangia of Parasporotheca Dennis and Eggert (1978) are interesting because they lack a radial arrangement of the spo-

PALEOZOIC SEED FERNS 333
Fig. 9. Parasporotheca leismanii, pinnate branching system bearing aggregate of synan- gia, • (Redrawn from Dennis and Eggert, 1978.)
rangia. Instead, the elongate sporangia fan out from the base into a curved scooplike synangium (Figs. 8, 9). As in most other medullosan pollen organs, the sporangial cavities alternate with elongate lacunae in the ground tissue, and a single vascular bundle is present on the outer (dorsal) surface of each sporangium. Like Rhetinotheca, these aggregate synangia have a very thin ground tissue and are covered with the distinctive in- terlocking peglike hairs on the outer surface. In the basal ~/3 of the syn- angium the ventral surface is composed of fibers, but at more distal levels, where dehiscence slits form, the wall is composed of thin walled cells. Dennis and Eggert (1978) note that the entire synangial mass was flattened and incomplete, however, the fructification measured 20.0 cm in length and 3.0 cm in width. No common ground tissue covered the mass that might suggest the aggregated synangia were part of a single unit. It is probable that individual synangia dried and separated from the aggrega- tion, and therefore could be preserved as a unit of the parent fructifica- tion. Dehiscence did not require that the synangial mass disaggregate since numerous examples of dehiscent sporangia were illustrated in in- timate contact with neighboring synangia.
Dennis and Eggert (1978) suggest that each laminate synangium of Parasporotheca may have originated from a laminar fertile telome system that underwent lateral fusion. This is in contrast to ~ther medullosan pollen organs that appear to be basically radial in construction (e.g., Sullitheca, etc.). These authors further suggest that the radial forms may also have had their origin from laminar fertile teiome systems through fusion of the two lateral margins of the synangium (Fig. 7A). Since we

334 THE BOTANICAL REVIEW
believe that all medullosan pollen organs are basically radial and not laminar, we offer the following alternate hypothesis. According to this hypothesis the laminate synangium of Parasporotheca is basically radial, although a small area of the enlarging cone-shaped synangium fails to develop (Fig. 7E). Such a region of abortive sporangia would form a slit up the side of the synangium and allow the organ to open out into the variously curved synangia seen in Parasporotheca. This hypothesis is supported by the increasingly radial aspect of these synangia near their base. A detailed study of the vascular system in the base of Parasporo- theca synangia might help to clarify the symmetry of the organ. Further evidence that suggests the ancestral medullosan pollen organ has small radial synangia is provided in the general discussion.
Compound Synangia
Potoniea.--The pollen organ Potoniea Zeiller (1899) is known primar- ily from compression fossils, but recently a permineralized specimen was reported (Stidd, 1978b). This discovery is extremely important because, like Dolerotheca, the morphology of Potoniea has been much debated (Bertrand, 1913; Carpentier, 1911, 1929; Halle, t933, 1942; Laveine, 1971). In addition, the absence of knowledge of the internal structure of Potoniea has resulted in the genus being placed in the Medullosaceae on the basis of gross appearance, and an association with medullosan foliage (Linopteris). The microspores of Potoniea are large (av. 90/zm), radial and trilete (Fig. 18A), and are completely unlike those of all other prob- able medullosan pollen organs (Fig. 18D, F). The nature of the micro- spores made the assignment of Potoniea to the medullosans questionable.
The overall form of Potoniea is generally described as campanulate (bell-shaped), with an abrupt constriction on the dorsal surface that forms a tapering pedicel (Fig. 10). The whole organ measures from 0.5-1.0 cm in diameter, and consists of numerous, parallel, elongate sporangia embedded in a ground tissue. One of the unusual features of Potoniea is the sporangial tips that protrude from the distal face of the pollen organ for approximately 1.0 ram. The compound nature of the synangium only becomes clear in paradermal sections of the face of the organ where sporangia may be viewed in transverse sections. Figure 17A is a para- dermal section of the face of the organ through the free tips of the spo- rangia. In this section the pollen organ appears to consist of perhaps five concentric rings of sporangia. Within each ring the sporangia are arranged in clusters of 4-6. These appear to represent small radial synangia that are organized into the larger, radial, compound synangium. Evidence for this is provided by the introrse dehiscence of the sporangiai groups sug- gesting they are functional units, and by the vascular system. Each spo-

PALEOZOIC SEED FERNS 335
Fig. 10. Potoniea synangium with cut-away to expose small radial clusters of elongate sporangial tubes arranged in concentric rings, x 12. (Redrawn from Stidd, 1978.)
rangium is vascularized by a bundle located toward the periphery of the sporangial group in a manner similar to a radial synangium like Hallethe-
ca. In addition, sporangia of each group surround a central cellular zone in the distal region of the pollen organ proximal to the level where the tips are free (Fig. 7G). The presence of flee sporangial tips appears unusual, however, it should be noted that each sporangium requires free wall space in order to dehisce. We believe that no Paleozoic pollen organs possessed true apical dehiscence; rather sporangial dehiscence was longitudinal even in instances where sporangia opened into juxtaposed pollen sacs, or was limited to reduced lateral slits.
Two other interpretations of the structure of Potoniea are based on compression specimens, and are not consistent with the observed com- plexity present in permineralized specimens. Halle (1933) macerated specimens and obtained spore masses of different lengths. From this he deduced that the organ consisted of a cuplike outer 'fintegument" from the bottom of which radiated numerous elongate sporangia. This as- sumption is accurate, but does not detail the grouping of sporangia, or the ground tissue in which sporangial cavities occur. Well-preserved specimens described by Carpentier (1929) and Laveine (1971) show the

336 THE BOTANICAL REVIEW
arrangement of the sporangial tips in fours. Laveine interprets the organ as a large, solid, bell-shaped structure with superficial sori composed of four short sporangia.
We are now in the interesting position of knowing a great deal about this pollen organ except for the group of plants to which it belongs. The early geological occurrence of this pollen organ type (Namurian; based on compression specimens of Deltenrea) does help to explain the pres- ence of decidedly nonmedullosan prepollen, but we believe the basic prepollen form was established very early in this family (e.g., Schulz- ospora). Creation of the Potoniea compound synangium would require the telescoping, aggregation, and fusion of a radial branching system bear- ing small radial synangia. If this occurred in the Namurian, it is interesting that most contemporaneous forms were still in the process of simple synangium formation. It is possible that Potoniea is an early medullosan offshoot or a lyginopterid, although the consistent association of this microsporangiate organ with medullosan foliage supports the former.
Dolerotheca.--One of the most interesting and complex seed fern pol- len organs is Dolerotheca Halle (1933). The genus is easily recognized because of the very large size of the campanulate organ (up to 4.0 cm in diameter), and the characteristic radiating pairs of sporangia (Schopf, 1948). Probable specimens of Dolerotheca are also known as isolated compression fossils (Gillespie and Clendening, 1966), and are distinguish- able from Potoniea by the arrangement of the sporangia and morphology of the prepollen. The structural arrangement of Dolerothecaformosa has recently been described in detail by Dennis and Eggert (1978), and com- pared with Parasporotheca. The interpretation of these authors regarding the morphology of Dolerotheca appears plausible, and we believe it is consistent with the recognized organization known for other structurally preserved pollen organs.
The basic shape of the Dolerotheca campanulum consists of an elon- gate-hemispherical unit with an eccentrically placed pedicel (Fig. 12). The outer surface is covered with numerous multiceUular hairs similar to those described for Sullitheca. A paradermal section of the face (bottom) of the organ is necessary to determine the arrangement of the sporangiai tubes as seen in transverse section. In a complete specimen there are four areas visible in the pollen organ (Fig. 11); each represents a single radial synangium that is folded in a symmetrical way. The two middle synangia are shaped like "bow ties" whereas the synangium at each end is roundly triangular and has the configuration of a bow tie folded along one side. The original radial nature of the component synangia, and the manner of internal folding is apparent in Fig. 11. In either of the end synangia the deepest fold divides the tubular synangium in half, while the next deepest fold divides each of the halves into quarters. This plication
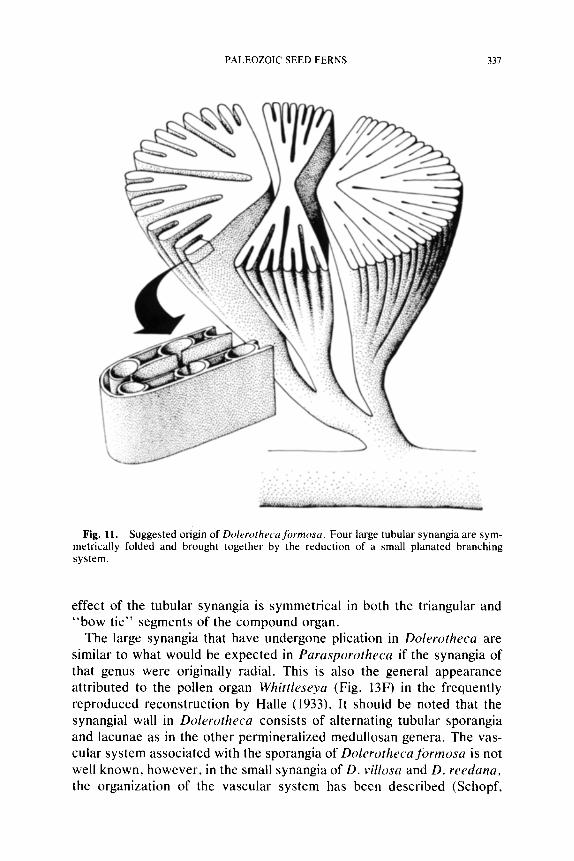
PALEOZOIC SEED FERNS 337
Fig. 11. Suggested origin of Dolerotheca formosa. Four large tubular synangia are sym- metrically folded and brought together by the reduction of a small planated branching system.
effect of the tubular synangia is symmetrical in both the triangular and "bow tie" segments of the compound organ.
The large synangia that have undergone plication in Dolerotheca are similar to what would be expected in Parasporotheca if the synangia of that genus were originally radial. This is also the general appearance attributed to the pollen organ Whittleseya (Fig. 13F) in the frequently reproduced reconstruction by Halle (1933). It should be noted that the synangial wall in Dolerotheca consists of alternating tubular sporangia and lacunae as in the other permineralized medullosan genera. The vas- cular system associated with the sporangia of Dolerothecaformosa is not well known, however, in the small synangia of D. villosa and D. reedana, the organization of the vascular system has been described (Schopf,
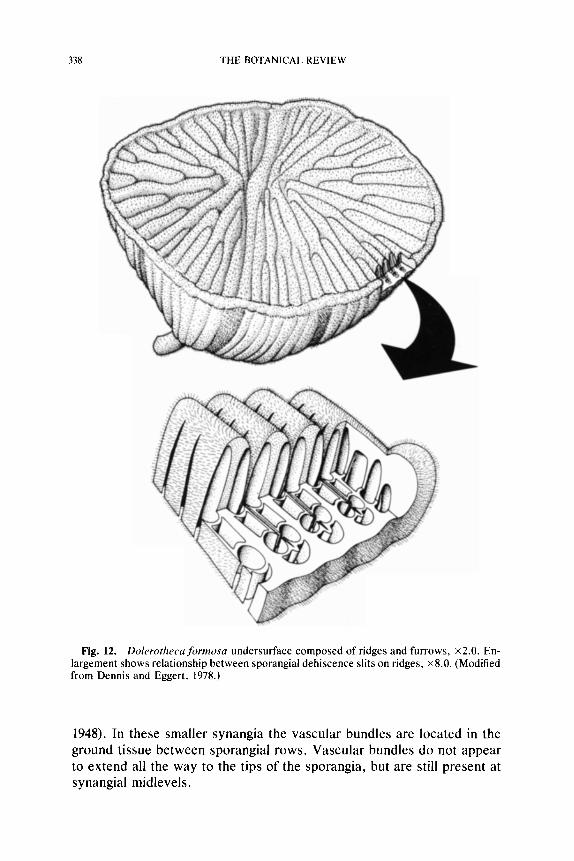
338 THE BOTANICAL REVIEW
Fig. 12. Dolerothecaformosa undersurface composed of ridges and furrows, • En- largement shows relationship between sporangial dehiscence slits on ridges, • 8.0. (Modified from Dennis and Eggert, 1978.)
1948). In these smaller synangia the vascular bundles are located in the ground tissue between sporangial rows. Vascular bundles do not appear to extend all the way to the tips of the sporangia, but are still present at synangial midlevels.

PALEOZOIC SEED FERNS 339
Sporangial dehiscence in a large compound pollen organ like Dolero- theca superficially appears inefficient because of the crowded nature of the sporangia. Folding of the individual synangia is organized in such a way so that dehiscence surfaces of two sporangia are opposite. This ar- rangement necessitates that sporangia dehisce into each other at levels still within the synangium. On the face of the synangium the surface of the organ is marked by radiating grooves (1.0-2.0 mm deep) that repre- sent the common dehiscence area of a row of paired sporangia (Fig. 12). In Dolerotheca therefore, sporangia do not dehisce through apical pores, but rather undergo longitudinal dehiscence so that prepollen is released into opposing sporangia and a shallow distal groove (Schopf, 1948; Stidd, 1978a).
Synangia that comprise the compound pollen organ Dolerotheca ap- pear similar to synangia of Sullitheca. Synangia in both genera consist of a hollow ring of sporangia that has been symmetrically plicated. This is suggested by the regular alternation of sporangial cavities and presum- ably lysigenous lacunae that surround a more or less continuous area of sclerenchyma in the synangium base, and a branched, but continuously open groove at distal levels (Fig. 7B-D). The position of sclerenchyma and dehiscence areas clearly coincides in Sullitheca, but is less obvious in sections of Dolerotheca formosa. In the detailed work of Schopf (1948, p. 696) it was noted: " . . . aside from the campanulary cover, supporting tissue of the large fleshy male frutification of D. formosa is limited to the fibrous sclerenchyma strands developing above the dehiscence grooves. As mentioned, cells of the dehiscence tissue merge with the sclerenchyma in these areas by a gradual transition." In D. sclerotica Baxter (1949) a thick band of sclerenchyma separates each of the paired sporangial tubes. Dehiscence slits must form through the fiber bands, similar to the situa- tion in Halletheca. The outer surfaces of both Sullitheca and Doler- otheca are covered with multicellular hairs that are also present in the dichotomous furrows of the complex dehiscence area. The princi- pal differences between these taxa involve the presence of sterile scler- enchymatous "fingers" in Sullitheca, and features of the dehiscence areas. In Sullitheca and Halletheca dehiscence occurs through a fibrous part of the inner facing wall, whereas Dolerotheca formosa has spe- cialized cells associated with the dehiscence areas. These cells are thin walled, rectangular, and uniform in height so that when viewed in longitudinal section they resemble a compact palisade tissue.
Among the six species of Dolerotheca presently recognized, only in D. formosa is the overall synangiai morphology clearly understood. The European species, D. fertilis, is known from fragmentary material, but appears to be about the same size as D.formosa. One apparent difference between the two is that D.fertilis apparently has little or no sclerenchyma
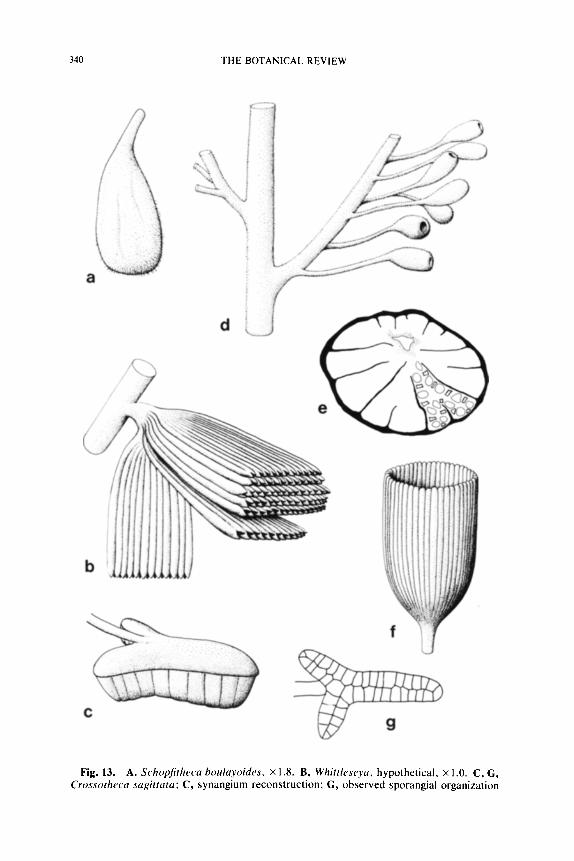
340 THE BOTANICAL REVIEW
Fig. 13. A. Schopfitheca boulayoides, • 1.8. B, Whittleseya, hypothetical. • 1.0. C, G, Crossotheca sagittata; C, synangium reconstruction; G, observed sporangial organization

PALEOZOIC SEED FERNS 341
in the synangium. Unfortunately sections through the dehiscence area are not available for this species. Both D. reedana and D. villosa are similar in histology to D. formosa, but their gross morphology is not known. Since D. reedana and D. villosa are considerably smaller than D. formosa, it is possible that these species are constructed of fewer synangial units. For example, D. villosa is only 1.0 cm in diameter, and like Sullitheca may consist of only a single radially plicated synangium. Specimens of D. reedana are incomplete, but are estimated to represent organs that are 1.5 to 2.0 cm in diameter. Conceivably a pollen organ of this size could be constructed of from 1-4 synangia. Appropriate sections of these other Dolerotheca species are not presently available to deter- mine synangial organization, but the detailed anatomy of these species is similar. That simple synangia should not be included in the same genus with those of a compound organization is evident when considering ad- ditional pollen organs of the Dolerotheca type such as Stewartiotheca.
Impression-Compression Forms
We have attempted to demonstrate in the previous section that there is considerable diversity in the overall organization of medullosan pollen organs, but uniformity in the structure of synangia or the synangial units. Even in the most complex arrangement of the sporangia (e.g., Potoniea) certain basic elements of construction are apparent. This consistency of design forms the basis for the following discussion of the morphology of a variety of compressed pollen organs. It is not our intention to try to fit the compression fossils into known morphological categories based on the permineralized forms. However, it would appear reasonable to favor the detailed and consistent plan established for permineralized forms in lieu of the speculation previously associated with the compression fossils. We will initially discuss forms that appear tO represent simple medullosan synangia borne on branching systems, and later focus on systematically more problematical types. Features of the better known compression forms are summarized in Table II.
Aulacotheca.--Aulacotheca is thought to represent a medullosan pol- len organ because it contains prepollen of the Monoletes type. Most specimens of Aulacotheca are isolated or consist of small clusters of
on lower surface of synangium, x3.5. D, Goldenbergia glomerata, x2.5. E, Stewartio- theca warrenae, synangium in transverse section. A single row composed of sporangia (circular) alternating with lacunae (rectangular) is folded radially around an off-center hol- low region, x4.5. F, Whittleseya elegans, x0.8. (Figures redrawn from: A, Delevoryas, 1964; C, Millay and Taylor, 1977; D, F, Halle, 1933; E, Eggert and Rothwell, 1979; G, Arnold and Steidtmann, 1937.)
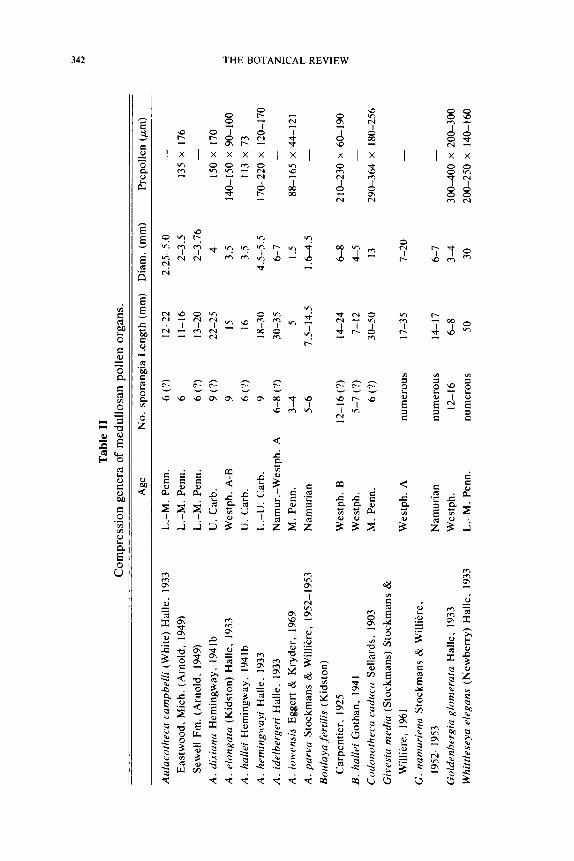
342 THE BOTANICAL REVIEW
0
"8
r
0
I--.-
X I x x x x I x I x I x I I x x
<
~ ~ ~ ~ . ~
I
�9 ~ ~ ~ ~ ~ ~ ~ ~ ~ . ~ ~. ~.. : .~ ~ . ~
o o ~ , o
e-
z ~

PALEOZOIC SEED FERNS 343
synangia (Table II). Eggert and Kryder (1969) describe a small frond segment of this genus as consisting of a delicate primary axis bearing six alternately arranged laterals. The synangia are borne in "tufts" along lateral branches, apparently at the tips of small dichotomizing axes.
The genus was established and intensively studied by Halle (1933) who embedded and sectioned the compressed synangia. He deduced that the synangia consisted of a closed, hollow ring of sporangia, and reconstruct- ed A. elongata as a large tubular synangium. This hollow ring interpre- tation was supported by Halle owing to the arrangement of the com- pressed sporangia and the small amount of tissue in the center of the synangium (Fig. 6A). Halle (1933, p. 38) does note, however, that in several specimens " . . . there is a narrow central area of black carbon- aceous matter which gradually widens from near the base to near the apex." He further suggests that " . . . in the concrescent sporangia of Aulacotheca hemingwayi the inner walls facing the central cavity were rather thick." We support Halle's interpretation of Aulacotheca, and note particularly that it compares closely with Halletheca. In the Hal- letheca synangium the sporangia surround a fiber core at the base, and the inner facing sporangial walls are composed of fibers in the distal half of the synangium. We suggest that Aulacotheca has an anatomy like Halletheca, but with varying numbers of sporangia per synangium. The size of the central hollow in the synangium of Aulacotheca probably varies considerably depending on the species. The internal dehiscence area could be restricted to as little as a third of the synangium length. Moreover, the core of fiber cells could be restricted to the base of the synangium in some species (e.g., A. elongata) in which the central hollow appears to be rather extensive.
One interesting feature of Aulacotheca that was given little attention by Halle is the serrated apex of the synangium. Halle (1933, p. 30) did observe that the longitudinal ridges show a tendency " . . . to become separated from one another and form free lappets at the ends of the specimens, especially at the apex." He believed this indicated ~ ' . . . that the specimens are formed by concrescent segments which may spring apart as valves." Some specimens illustrated by Halle clearly exhibit pointed lobes at the synangial apex that appear to correspond in number to the sporangia present. We interpret these as free sporangial tips sur- rounding the central hollow (Fig. 6A). It seems unlikely that synangia were closed at the apex by sporangial tips as Halle's reconstruction in- dicates, but rather were open to varying degrees throughout their devel- opment.
The toothlike sporangial tips in Aulacotheca are also present in other compression genera such as Boulaya and Whittleseya. If the apex of Halletheca were known it might help in further understanding the mor-

344 THE BOTANICAL REVIEW
phology of some compression genera such as Aulacotheca and Golden- bergia.
Boulaya.--The genus Boulaya was proposed by Carpentier (1925) for some isolated pollen organs that had earlier been referred to as Whittle- seya fertilis (Kidston) Carpentier (1914). Synangia of Boulaya differ from Aulacotheca only in their pyriform shape, since these specimens fall within the size range of AMacotheca (1.0-3.5 cm). Some specimens de- scribed by Carpentier (1914) are relatively large (1.4-2.4 cm long), while others are rather small (10-11 mm x 3.5-4.5 ram). Synangia assigned to B. fertilis by Halle (1933) measure 1.7 cm long and up to 0.6 cm wide. The synangium is believed to be roundly triangular in transverse section based on impressions left in part and counterpart specimens (Kidston, 1914).
Halle (1933) studied the genus and concluded that the synangium was hollow at the distal end because there was little tissue present in the center of the synangium near the apex. As noted in Aulacotheca, there is a central area of tissue that is narrow at the base and wider in the apex. This tissue was thought to represent the remains of the inner facing spo- rangiai walls by Halle, but we would suggest that the synangia were hollow only in the broad distal half. The distal margin of tht synangium has teeth that appear to correspond in position and number to the embed- ded sporangia and may in fact represent the sporangial apices. As in Aulacotheca, Halletheca, and others, Boulaya sporangia probably de- hisced into a central cavity. The absence of longitudinal ribs on the sur- face of Boulaya probably indicates that the outer synangial wall was thick like that in Halletheca. The apparent absence of epidermal hairs is another feature shared by these genera. Other freely borne pollen organs (e.g., Goldenbergia) generally have a dense covering of hairs.
Codonotheca.---Codonotheca caduca Sellards (1903, 1907) is a rela- tively common fossil in the American Carboniferous. The synangium con- sists of a simple ring of six spore bearing units that are basally fused but free for ~/3 to ~/2 of their length (Fig. 6F, 17D). One notable feature about the synangium is its large size (3.0-5.0 cm long), and enormous diameter of what are generally interpreted to be the component sporangia (ap- proximately 3.0 mm in diameter). Codonotheca has not been the subject of detailed research since the initial description by Seilards, who gave a rather complete discussion of the genus apparently based on numerous well-preserved specimens. Of the two interpretations concerning the mor- phology that were presented by Sellards, the one suggesting the synan- gium as a simple ring of six sporangia was not favored. Sellards was hesitant to identify the six, radially arranged, "spore bearing segments or divisions" as sporangia since he apparently believed that more than one sporangium was represented by each segment. It was Halle's (1933)

PALEOZO1C SEED FERNS 345
acceptance of these fertile segments as sporangia that established this interpretation in subsequent publications.
If Codonotheca does in fact represent a uniseriate whorl of 5-6 large sporangia it can be compared closely to the synangium of Halletheca. Sellards provides a diagram of the synangium as though it has been " . . . cut down the middle and laid open" (Fig. 6B). He indicates that a "vascular cone" was present in the synangium base that provides two vascular strands to each sporangium (Fig. 6B, VC). We interpret the supposed vascular tissue as representing fibers (Fig. 17D), since the cen- tral regions of other medullosan pollen organs are constructed in a similar manner. Further support for this suggestion concerns the size of vascular strands which would appear microscopical and not of the dimensions indicated by Sellards. Codonotheca synangia are probably solid in the base and constructed of a fiber core that becomes hollow toward the distal end of the organ. The pairs of elongate fiber areas (Sellard's vas- cular bundles) present on each spore bearing segment are clearly inter- nally located, and may delineate the dehiscence line of one to several sporangia. When compared to Halletheca (Fig. 6D) it is possible to en- vision the origin of paired fiber strands in Codonotheca as the free-ends of the sporangium wall along the dehiscence line.
Despite the apparent close similarity between Codonotheca and Hal- letheca, we do not believe that the two organs are of comparable mor- phological complexity. A comparison of Codonotheca with structurally preserved meduliosan pollen organs suggests some major inconsistencies. The sporangium diameter in permineralized synangia is relatively small, and ranges from 0.3 mm in Dolerotheca villosa to 1.0 mm in Sullitheca (Table I). The spore bearing segments of Codonotheca are many times the diameter of Sullitheca sporangia. We suggest that each supposed sporangium of Codonotheca may contain several sporangia facing the inner surface of the synangium (Fig. 6F), and more if the synangium were internally fluted as in Sullitheca. It may also be significant that the radial synangia of Sullitheca possess a number of essentially sterile, apical processes. Despite the above speculation, Codonotheca still has a totally problematical morphology and will hopefully become the topic of further research using partial petrifactions. We believe that Codonotheca rep- resents a poor example on which to base speculation about the origin of other synangium types since the morphology of this organ is at best poor- ly known.
Schopf (1948) has indicated that the outer surface of Codonotheca is covered with multicellular hairs similar to those that cover the synangium of Dolerotheca. Sellards (1907) describes a specimen in which the synan- gia appear to be paired and are attached to the main axis by slender petioles. Such a specimen (Fig. 15B) is illustrated by Darrah (1970), and

346 THE BOTANICAL REVIEW
suggests that the synangia are not mixed in with foliage on an otherwise sterile frond, but were probably borne on an entirely fertile frond or frond segment.
Deltenrea.--Deltenrea is a monotypic genus established by Stockmans and Willibre (1958) for small synangia borne terminally on stout branches. Branches bend abruptly at their apex and bear a single oval synangium surrounded by radiating sporangial tips. The fructifications have much the same morphology as a small Potoniea (8.0 mm diameter), but the free portions of the sporangia represent fully half the diameter of the synangium. The authors compare the general appearance of Deltenrea with the Mesozoic pollen organ Pteruchus. Knowledge of the microspore type would help to indicate if the affinities of this poorly known genus are indeed close to Potoniea.
Givesia.--The taxon Givesia namuriena Stockmans and Willi~re (1952-1953) was established for small synangia (14-17 mm long) similar to Whittleseya in morphology. A second species was transferred to Gi- vesia (G. media) from Whittleseya that is intermediate in morphology between the type species of these two genera. As in Boulaya, synangia of Givesia taper basally from a point near their apex, beyond which they are slightly constricted. Synangia of Whittleseya generally appear par- allel-sided and are not apically constricted. In addition, synangia of Gi- vesia have large " teeth" distally with correspondingly wider ridges along the sides of the organ. The teeth or sporangial tips are very pronounced in this form and may be 1.0 mm long and 1.0 mm in basal width.
Stockmans and Willi6re (1961) acknowledge that forms of Givesia may intergrade with some specimens of Whittleseya or Boulaya. The synangia of Givesia may have been an open cup, such as has been suggested for Whittleseya, or may have been flattened and produced in clusters. We do not support the open cup suggestion because of the large size of the fructifications, and correspondingly large internal surface area that would be open to dehydration and possible predation by insects.
Goldenbergia.----Goldenbergia glomerata Halle (1933) was established for small entirely fertile frond segments bearing stalked synangia (Fig. 13D). The synangia appear to be borne in pairs on the same side of pinna axes that are in turn alternately arranged on the penultimate pinna. The pinnae and synangia are covered with a dense pubescence. The ovoid synangia are small (6-8 • 3-4.5 mm), and taper basally to a thick stalk, while the apex is truncated. These synangia have no longitudinal ribbing and are believed to have a thick outer integument. Halle (1933) provides a reconstruction of Goldenbergia in which each synangium consists of a ring of 12 sporangia that surround a central hollow area. Halle believed that the synangia were closed structures, and further suggested that de- hiscence occurred along the inner side or top of each sporangium, or that

PALEOZOIC SEED FERNS 347
the capsule split lengthwise into individual sporangia. Our interpretation is that Goldenbergia synangia are cuplike and open apically. With this organization sporangia would dehisce so that pollen was shed toward the center of the synangium (Fig. 13D). The presence of a central fiber area is not known for Goldenbergia, however, in cuplike pollen organs such a tissue could be restricted to the base of the synangium.
Schopfitheca.--Schopfitheca boulayoides (Delevoryas, 1964) is the bi- nomial applied to clavate-pyriform microsporangiate pollen organs known as compression fossils of Middle Pennsylvanian age. The specimens mea- sure approximately 2.0 cm long and are up to 1.0 cm in diameter; the proximal end is elongate and probably represents a stalk, whereas the distal end is rounded and lacks any indication of dehiscence areas (Fig. 13A). The fructification is reconstructed with numerous, closely spaced hairs covering the surface of the organ as would be expected in the sol- itary synangium types. Because of the nature of preservation, nothing is known about the internal organization of the pollen organ. The presence of Monoletes pollen grains up to 480 ~m long and 390/xm wide constitutes the only conclusive evidence for assigning Schopfitheca with the med- ullosans.
Whittleseya.--When described by Newberry (1853a, b), the pollen or- gan Whittleseya elegans was initially regarded as a ginkgophyte leaf. Later, Halle (1933) established the synangial nature of this medullosan pollen organ. Although apparently of limited stratigraphic range, the basic morphology of this pollen organ is well understood. Each synangium is approximately 3.0 cm wide and 5.0 cm long, and tapers to an elongate pedicel (Fig. 13F). The apex of the synangium is dentate with the small teeth appearing to correspond in position to the numerous longitudinal striations (grooves) on the surface (Fig. 17F). Halle (1933) studied nu- merous specimens of Whittleseya synangia, and concluded that the organ consisted of a uniseriate ring of elongate sporangia surrounding a large central hollow. His reconstruction of the hollow campanulum is frequent- ly reproduced in textbooks (compare Fig. 13F). There has been some debate as to whether the synangium is naturally hollow, or represents a flattened uni- or biseriate structure (Schopf, 1948). The former viewpoint was supported (Halle, 1933) by the existence of specimens in which a small amount of rock matrix was observed to extend into the distal por- tion of the synangium, and partially fill the central hollow. Schopf (1948) did not accept the uniseriate whorl hypothesis offered by Halle because it was at variance with the paired sporangia he thought to be characteristic of Dolerotheca. Schopf (1948, p. 721) suggested that " . . . the very thin intervening shale layer Halle demonstrated at the distal edge of the lamina is not at all commensurate with the hollow cavity he postulated, and possibly it may be related to a dehiscence groove similar to those of

348 THE BOTANICAL REVIEW
Dolerotheca." He further quotes Whittlesey (1849) who had examined hundreds of specimens for evidence that it was a "hollow pod," but found no support for the idea.
We are inclined to accept Schopf's interpretation that these synangia were naturally flat. Halle demonstrated that Whittleseya consists of two rows of sporangia and if paired, the sporangia have facing dehiscence slits like those of Dolerotheca. A groove may have been present apically between the sporangial rows. For a number of reasons we find the idea of giant hollow campanula or pedicellate pollen producing shingles diffi- cult to accept. In both examples an enormous amount of surface area would be exposed, and tissue dehydration during some stages of devel- opment would be unavoidable. The sporangial walls in Whittleseya are not particularly thick and no hairs have been reported on these organs. In general, medullosan synangia that have been described in detail are quite compact and are protected from dehydration in a variety of ways, including the clumping together of organs, thick synangial walls and hairs. In other synangia with hollow central areas, the apical opening is rela- tively small, and the exposed dehiscence areas are composed of fibers. None of these structural features is known to be present in Whittleseya. We suggest that Whittleseya pollen organs were aggregated together into a stack of flat synangia (Fig. 13B). Such an organization is reminiscent of Parasporotheca, in which uniseriate synangia are aggregated into an irregular mass (Fig. 9). Synangia in Whittleseya would presumably de- velop in the stack, and at maturity shed pollen through the apical groove separating each paired row of sporangia (Fig. 13B). Synangia may have become disaggregated late in ontogeny and scattered over the ground like a fumbled deck of cards. Many investigators have commented on the unusual abundance of Whittleseya synangia on a given surface area of matrix. Evidence contrary to this interpretation involves the apparent absence of interlocking (peglike) hairs on the surface of Whittleseya synangia, and the occasional presence of a long pedicel on a synangium. Such pedicels might be flattened together along with the synangia in the aggregation.
Prepollen
The prepollen of Pennsylvanian medullosans shows considerable vari- ation in size (100-560/~m long), but at present only two morphological forms are associated with the family. The most common type is Mono- letes, an oval thick-walled grain possessing a prominent monolete suture with a median deflection (Fig. 18F). Specimens of Monoletes have a smooth outer surface and generally show two deep crescentic grooves in the sexine of the distal surface. Another microspore type (Parasporites) was isolated from Parasporotheca Dennis and Eggert (1978), that super-

PALEOZOIC SEED FERNS 349
ficially appears quite different from Monoletes. Specimens of Paraspo- rites (Fig. 18D) are characterized by a complex external ornamentation and flangelike sacci on each end of the central body. The construction of the exine is the principal feature that shows the close homology of these two microspore types. The thick inner nexine layer of both types supports a sexine that is constructed of small spherical chambers (Millay et al., 1978). This alveolate sexine infrastructure may consist of chambers that interconnect with one another, creating a matrix of anastomosing rods, or may consist of solitary chambers. Most of the medullosan pollen or- gans presently known have Monoletes with anastomosing rods, as does Parasporites. It may be significant that some genera lacking intercon- nected sexinous chambers (Codonotheca, Halletheca, Rhetinotheca) also appear to lack distal grooves, and have a broad, depressed monolete suture region (Millay et al., 1978). The likelihood that some of these features show variable expression (e.g., distal grooves) or are related to developmental changes (e.g., sexine infrastructure) was documented by Taylor (1978) in a broadly based study of Monoletes grain ultrastructure. Additional studies are needed at the ultrastructural level on Monoletes to determine the natural variation present in the microspores of certain pollen organ taxa, and features of sporoderm development.
It has been suggested that the organization of the nexine in the suture region of Monoletes and Parasporites is like that present in the isospores of certain homosporous ferns (Millay et al., 1978). The suture therefore represents an outfoiding of the nexine into the sexine. The presence of an apparently functional suture in these grains, along with the apparent absence of a preformed germination region on the distal surface, suggests that germination was proximal in these grains. It is for this reason that such grains have been referred to as prepollen.
C A L L I S T O P H Y T A C E A E
The most recently discovered, and most completely understood seed fern family is the Callistophytaceae (Stidd and Hall, 1970b). The scattered vegetative and fertile remains of Callistophyton are common in American coal balls of Middle and Late Pennsylvanian age, and numerous publi- cations have appeared detailing the sterile and fertile organs of this fam- ily. Below is a list of organs now believed to have affinities with this family.
Stems Petiole Foliage
Callistophyton Callistophyton Mariopteris ?
Seeds Pollen organs Pollen Callospermarion C allandrium Ve sic aspora Taxospermum Idanothekion

350 THE BOTANICAL REVIEW
Fig. 14. A-C, E, ldanothekion glandulosum; A-C, transverse sections of synangium moving toward sporangial tips showing loss of sterile tissue in center of synangium: E, reconstruction with cut-away, all x35. D, F, G, Callandrium callistophytoides; D, fertile pinnules, x8; F, G, transverse section of synangium at base and midlevels, x30. (Figures redrawn from: A-C, E, Millay and Eggert, 1970; D, Stidd and Hall, 1970a.)

PALEOZOIC SEED FERNS 351
Permineralized Specimens
At this time two genera of pollen organs are known for the callisto- phytacean seed ferns. Isolated specimens of Idanothekion Millay and Eggert (1970) were described from Middle Pennsylvanian sediments, whereas the Late Pennsylvanian Callandrium Stidd and Hall (1970a) was described on foliage of Callistophyton. As originally described these pol- len organ genera did not appear to be similar, but a reinvestigation by Rothwell (1972b) suggests that most of the supposed differences between the two genera represent different developmental stages and preserva- tional phenomena.
The synangia in both forms are radially symmetrical, sessile and borne abaxially on the foliage of Callistophyton. This foliage is still very poorly known, but appears to have a lobed margin and distantly spaced, dicho- tomizing lateral veins like pinnules of Mariopteris (Fig. 14D) (Stidd and Hall, 1970a). Callistophytacean synangia consist of a ring of elongate, exannulate sporangia that basally surround a solid parenchymatous area, and apically surround a central hollow. In Idanothekion the solid central tissue occupies V3 to �89 the synangium length (Fig. 14A, E), while in Callandrium the cellular region occupies �88 or less of the synangium base (Fig. 14F). Idanothekion synangia are larger (up to 1.4 • 1.7 mm) and contain 6-9 sporangia, while the smaller (1.1 z 0.7 mm) synangia of Cal- landrium are composed of 5-7 sporangia. The solid sporangial tips in both genera are curved and directed toward the synangium center (Fig. 14D, E), These distinctive tips are in some respects similar to the spo- rangial tips in Psilophyton and some Devonian progymnosperms (e.g., Tetraxylopteris ).
The outer facing sporangial walls appear to be slightly thicker (2-4 cells) and more complex in Idanothekion, although at maturity (postdis- persal) the sporangial walls of both forms may appear only 1-2 cells in thickness. In both Callandrium and Idanothekion the central portion of the synangium base is heavily vascularized, but only in Idanothekion are individual sporangia vascularized along the midline of the outer facing sporangial walls. Rothwell (1972b) considers this to be the most important difference between the two genera, but acknowledges that this character may not be of generic significance. Sporangial vascularization seems to be a character of only relative significance, that is easily lost through reduction in overall size.
In our opinion the only important difference between the Middle and Upper Pennsylvanian pollen organs of the Callistophytaceae concerns details of the structure of the inner facing sporangial walls relating to dehiscence. In Idanothekion only about �89 of the inner sporangiai walls are available for dehiscence, and much of this tissue is composed of thick

352 THE BOTANICAL REVIEW
walled elongate cells (Fig. 14B, C) like those occupying the center of the synangium base. At more distal levels the inner facing walls are thinner in the dehiscence zone. In Callandrium most of the length of the sporan- gium is involved in dehiscence, and this zone is marked by enlarged cells that ruptured to effect dehiscence (Fig. 14G, 17B). In general morphology these two taxa of pollen organs have the same basic organization. How- ever, the cellular construction of the dehiscence zone in these two taxa is quite different. In this feature, these pollen organs differ from one another to a greater degree than they differ from pollen organs in other families (e.g., Feraxotheca, Halletheca).
Pollen
The pollen of the Callistophytaceae has been extensively studied. In general organization these grains resemble the pollen of some extant co- nifers (e.g., Picea). This fossil pollen type, if found dispersed, would be included in the sporae dispersae genus Vesicaspora. Grains of this type are characterized by an endoreticulate girdling monosaccus that sur- rounds a central corpus (Fig. 18E). Since the air sac is laterally reduced, and the lobes of the saccus are distally inclined, this grain has a quite modern appearance. The saccus lobes distally delimit a germinal furrow (sulcus) that is bisected by a leptoma, providing the exit site for the pollen tube. A small branched pollen tube extending from the sulcus region of a Vesicaspora grain contained in the pollen chamber of a callistophyta- cean ovule has recently been reported (Rothwell, 1972a). This exciting find confirms the presence of true pollen and moreover the probable ex- istence of true siphonogamy in this seed fern group by the Middle Penn- sylvanian.
PROBLEMATICAL FORMS
Most of the problematical forms to be discussed come from Permian sediments of Europe. Some genera may exhibit a morphology interme- diate between Carboniferous types and pollen organs of the Mesozoic. Unfortunately, our current level of understanding Permian seed fern pol- len organs is incomplete due to their paucity in the fossil record and the general mode in which they are preserved. Numerous important biolog- ical changes appear to have taken place during the Permian, and it is quite possible that in this segment of geologic time Mesozoic pollen organ types were rapidly radiating. Pollen organs from Permian deposits resem- ble forms present in the lyginopterid seed fern complex in their general morphology, even though their affinities are probably among the medul- losan and callistophytacean genera. Because of this uncertainty we be-

PALEOZOIC SEED FERNS 353
lieve it unwise to speculate as to the possible relationships between the Mesozoic types and the Carboniferous seed ferns based on pollen organs alone.
Dictyothalamus.--Dictyothalamus schrollianus Goeppert (1864-1865) is a rather distinctive pollen organ from the Lower Permian of Germany. The fructification consists of a stout axis bearing laterals alternately that terminate in sporangial masses. The sporangia are 0.5 cm or more in length and appear to radiate in all directions from the tip of the ultimate pinna. Many of the sporangia appear to be apically pointed although many others are blunt.
The pollen of Dictyothalamus is large (80-160/~m long) and possesses a large girdling monosaccus. The central body is spherical to oval and appears thick walled. Some specimens appear to lack haptotypic features, while others have trilete sutures (Remy and Remy, 1958).
The general form of D. schrollianus is suggestive of a pollen organ that has undergone dehiscence, and it seems likely that the sporangia were more closely associated at an earlier developmental stage. This prede- hiscence stage would probably closely resemble the pollen organ Schuet- zia anomala which is known from the same stratigraphic level as Dic- tyothalamus. The presence of saccate pollen in Schuetzia (Remy and Rettschlag, 1954) of a similar size and morphology to that known from Dictyothalamus strengthens the likelihood that these two genera are closely related and possibly congeneric.
Psaliangium.--The pollen organ Psaliangium odontopteroides Remy (1953c) is known only from fragmentary remains from the Upper Ott- weiler of Germany. The pollen organs have short pedicels and are borne in a simple pinnate arrangement. Each fructification consists of an elon- gate tapering member (1.7 cm long and 0.2 cm wide) bearing microspo- rangia on the lower surface. Around the margin of the sporangium-bearing unit are pointed lobes that surround abaxially positioned sporangia. The overall morphology of this organ is comparable to an elongate Crosso- theca.
Spores isolated from the middle of the organ measure approximately 100/xm in diameter, and are spherical with a clearly defined trilete suture. Ornamentation is described as coarsely granular (Remy, 1953c).
The affinities of the taxon remain problematical at this time due to the paucity of specimens, however, the association of the fertile specimens with Reticulopteris (Linopteris) odontopteroides suggests a medullosan origin, although the general morphology of the fertile parts resembles the lyginopterid Crossotheca. It appears possible that Psaliangiura may be related to Potoniea since they have similar pollen, and both are thought to belong to plants producing "paripinnate" neuropterid foliage.
Schuetzia.--A number of pinnate fertile specimens have been assigned

354 THE BOTANICAL REVIEW
\
0
e
C

PALEOZOIC SEED FERNS 355
to Schuetzia anomala Geinitz (1863) from Lower Permian sediments of central Europe. Fertile branches are once pinnate with the short laterals borne alternately and in the same plane (Fig. 15A). Lateral branches are terminated by a tight cluster of elongate sporangia measuring 0.7-1.2 cm long.
In some specimens illustrated by Goeppert (1864-1865), the flattened sporangia appear like sterile "bracts" that are basally attached to a com- mon branch. Schuster (1911) describes some specimens as consisting of a "basket" of microsporophylls, and provides a line drawing portraying the sporophylls laterally fused at their bases into a cup. He indicated that a sporangium was present on the upper surface of each sporophyll. Remy and Remy (1959a) reconstruct fructifications of Schuetzia as a cluster of elongate sporangia surrounded by sterile bracts (Fig. 2E).
An axis bearing sporangial clusters of Visean age was described by Kidston (1884) as a new species of Schuetzia. Schuetzia bennieana differs primarily from the generitype in the arrangement of the sporangial clus- ters. Sporangia measure from 1.0-1.2 cm long and are fused laterally in the region of the bases, or perhaps were borne on a sterile peltate disk (Kidston, 1924). A specimen of S. bennieana was sectioned by Halle (1933), who found it to consist of i0 sporangia that were free distally and closely aggregated in the base. No distinctive arrangement of the sporan- gia was evident in these cross sections to help clarify the construction of the synangia, however, it is interesting that no sterile peripheral struc- tures were present. It has been suggested that S. bennieana should be transferred to another genus (Walton, 1931), however, the limited knowl- edge of the specimens precludes another assignment at this time.
Schuster (1911) obtained small (50 ~m diameter) radial, trilete micro- spores from specimens of S. anomala, while Kidston (1924) obtained similar spores (50-70 p,m) from S. bennieana. More recently, Remy and Rettschlag (1954) have isolated monosaccate pollen (130 x 100/zm) from specimens of S. anomala from the Rotliegenden of Germany.
Schuetzia is a generalized pollen organ type consisting of numerous elongate sporangia. In the Mississippian (equivalent) age specimens the synangia are composed of relatively few sporangia that may have been
<_....
Fig. 15. A-F, shadow diagrams of fertile branches; A, Schuetzia anomala, x0.5; B, Codonotheca caduca, x0.55; C, possible ovulate (left) and microsporangiate (right) branch- es of seed fern with erernopterid foliage, x 1.0; D, Crossotheca hughesiana, x0.8; E, Neu- ropteris heterophylla (?) pinnae with terminal synangia, x 1.0; F, Telangiopsis arkansanum, x3.0. (Figures drawn from illustrated specimens; A, Goeppert, 1864-1865; B, Darrah, 1970; C, Delevoryas and Taylor, 1969; D, Kidston, 1906; E, Halle, 1933; F, Eggert and Taylor, 1971 .)

356 THE BOTANICAL REVIEW
randomly arranged at the pinna apex. Such a random clustering of spo- rangia may have been quite functional if sporangia were able to separate laterally during dehiscence. Schuetzia specimens from the Permian ap- pear to contain more sporangia, and it is likely that some regular arrange- ment of the sporangia is present that is associated with dehiscence and has not yet been identified. Several authors (Goeppert, 1864-1865; Schim- per, 1870-1872), have suggested that Dictyothalarnus and Schuetzia an- omala are segments of the same biological species; one representing the pollen organ and the other containing seeds. The isolation of similar ap- pearing pollen from both organs confirms that both are microsporangiate, and further indicates that Dictyothalamus may represent a preservational type of Schuetzia in which the synangia are basally compressed, or in which sporangia became laterally disassociated prior to preservation.
Thuringia.--Thuringia callipteroides Remy (1953b) is a sausage-shaped pollen organ from the Lower Permian (Rotleigenden) of Thuringia. The synangia are cylindrical, measure 29.0 mm long and about 3.5 mm wide, and taper abruptly at each end. From transverse sections it was estimated that approximately nine sporangial tubes were present, but their arrange- ment and mode of dehiscence still remain in doubt. Each synangium apparently possessed a thick outer ground tissue with stomata. Some features of the cuticle compare closely with those of the sterile foliage Callipteris conferta, present in abundance at the same stratigraphic level as the pollen organ. The similarity of cuticular features and association of plant parts led Remy (1953b) to suggest that Thuringia is the pollen organ of C. conferta.
Pollen isolated from Thuringia is monosaccate with a spherical (30-35 /xm) central body. Grains measure approximately 50/zm in diameter, and all appear alete.
The morphology of Thuringia is somewhat similar to some of the small radial types in the Pennsylvanian (e.g., Aulacotheca). Remy suggests that the synangium of Thuringia was a solid unit with sporangial tubes embedded in a continuous ground tissue. Sporangial dehiscence would have to be apical if his reconstruction is accurate. The pollen is unlike most Permian forms because the saccus appears to lack an endoreticu- lum, and a germinal furrow (distal sulcus) is not present. Several impor- tant features of Thuringia need to be determined. These include the meth- od of sporangial dehiscence, details of the pollen, and the way in which the pollen organ was attached to the plant.
Unguitheca.--Unguitheca pecopteroides is the binomial applied to fragmentary fertile frond material from the Rotliegenden (Permian) of Ilfeld (Germany). As reconstructed by Remy (1953d), the specimens have a broad penultimate pinna that produces small ultimate pinnae alternate- ly. Synangia are terminal on short stalks, and borne alternately and in the

PALEOZOIC SEED FERNS 357
same plane on the ultimate pinnae. Each synangium consists of 5-7 spo- rangia measuring up to 1.0 mm long. The mode of sporangial dehiscence and spore type is not known. The overall appearance of the specimen as a planated branching system bearing small Telangium-like synangia was the basis used in suggesting affinities with the pteridosperms (Remy, 1953d).
Unguitheca was found attached to a poorly preserved fertile frond segment resembling a Pecopteris of the Unita group. With the possible exception of Zeilleria, the specimen does not appear similar to any mar- attialean fern. The foliage of Zeilleria (Pecopteris avoldensis) does re- semble P. unita in a number of features, and synangia of Zeilleria are borne in a marginal position at the terminus of lateral veins on the pin- nules. If the lamina were not preserved the synangia might appear to be terminal on lateral branches.
Because of the fragmentary nature of the specimens, Unguitheca will have to be considered problematical until additional, better preserved specimens are discovered. The possession of synangia would limit the taxon to either the seed ferns or marattialean ferns.
D I SCU SSI O N
Phylogenetic Trends
The microspore or pollen bearing organs of the four presently recog- nized Paleozoic seed fern families are not very similar when compared in detail. Certain general trends in sporangial arrangement are evident in these seed ferns that suggest a relationship to the relative age of the specimens (see Fig. 16). The earliest examples (Lower-Middle Missis- sippian) of probable seed ferns have elongate sporangia borne in clusters along dichotomizing axes. Sporangia are thin walled and in some taxa do not appear to have a specialized dehiscence mechanism (Fig. IH). The thin walled sporangia in these forms may be an adaptation to the clustered organizations, since a similar correlation is observable in Devonian spo- rangia that may be solitary and thick walled (e.g., Psilophyton), or clus- tered and thin walled (e.g., Rhacophyton). The sporangium-bearing branches found in the Early Mississippian forms may be quite three-di- mensional (e.g., Protopitys), or may appear planated to some degree (e.g., Alcicornopteris). This appears to be the general arrangement of the microspore-bearing axes of the Calamopityaceae, a poorly understood and somewhat artificial family of seed ferns.
There is some question as to whether the Calamopityaceae should be regarded as a natural taxonomic group, or simply an early stage in the evolution of other seed fern families. In our opinion, the possession of
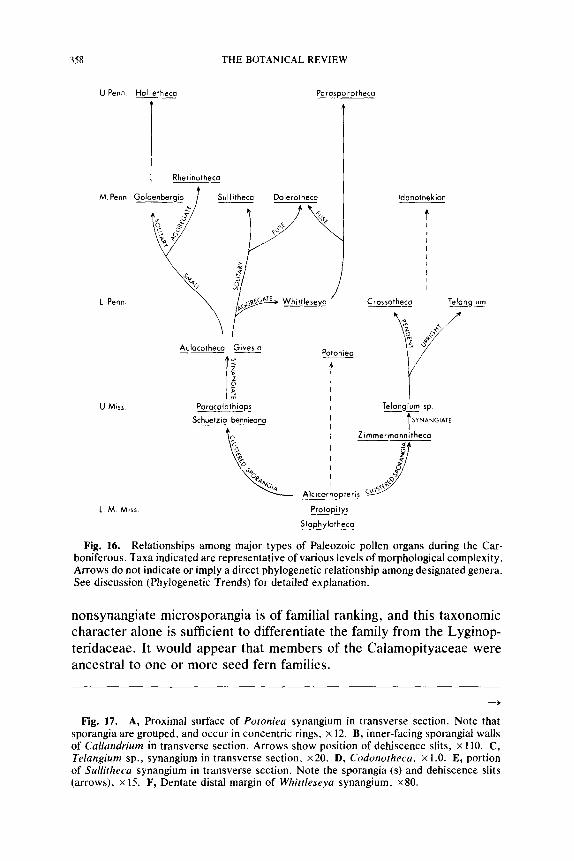
358 T H E BOTANICAL R E V I E W
UPenn Holletheca Paras ~orotheca
Rhetinotheca
M Penn Goldenbergio t Sullitheca Dolerotheca
L.Penn. o~-~- c~ ~'~E Whltll _
Aulacotheca Givesia Potoniea
U. Miss. Paracalathi_ops Schuetzia bennieana
J Alcicorn0pteris
p~oropitys S taphylotheca
L. M Miss.
Idanothekion
C ros sotheca Telangium
\ /
/ lela2n iu~m sp.
SYNANGIATE
Zimmermannitheca
Fig. 16. Relationships among major types of Paleozoic pollen organs during the Car- boniferous. Taxa indicated are representative of various levels of morphological complexity. Arrows do not indicate or imply a direct phylogenetic relationship among designated genera. See discussion (Phylogenetic Trends) for detailed explanation.
nonsynangiate microsporangia is of familial ranking, and this taxonomic character alone is sufficient to differentiate the family from the Lyginop- teridaceae. It would appear that members of the Calamopityaceae were ancestral to one or more seed fern families.
Fig. 17. A, Proximal surface of Potoniea synangium in transverse section. Note that sporangia are grouped, and occur in concentric rings, x 12. B, inner-facing sporangial walls of Callandrium in transverse section. Arrows show position of dehiscence slits, x 110, C, Telangium sp., synangium in transverse section, x20. D, Codonotheca, x l.0. E, portion of Sullitheca synangium in transverse section. Note the sporangia (s) and dehiscence slits (arrows), x 15. F, Dentate distal margin of Whittleseya synangium, x80.
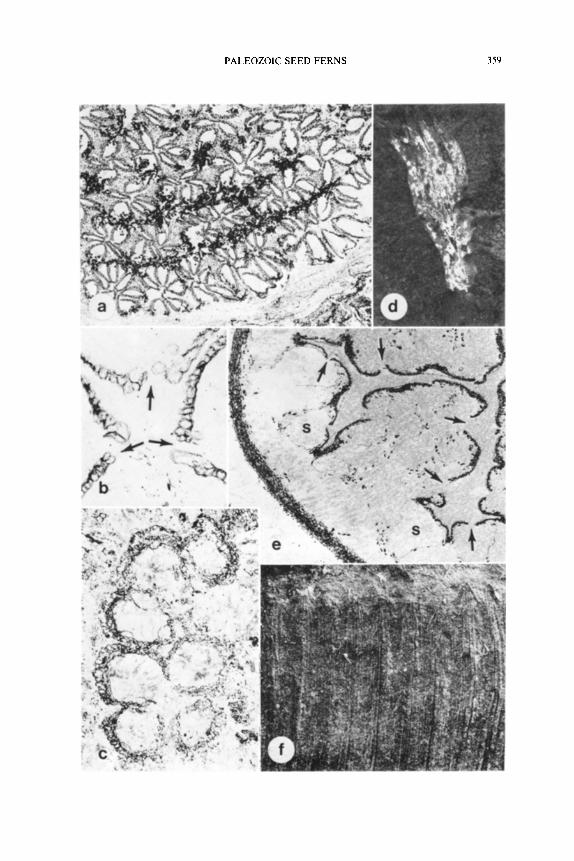
PALEOZOIC SEED FERNS 359
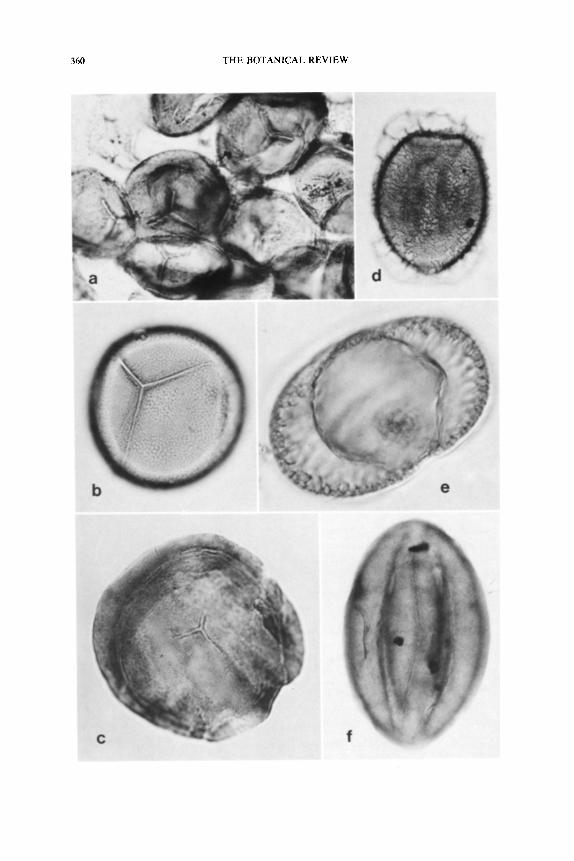
360 THE BOTANICAL REVIEW

PALEOZOIC SEED FERNS 361
In younger Mississippian rocks a mixture of microsporangiate struc- tures including both aggregations of sporangia and true synangia is pres- ent. In some cases a large number of sporangia are clustered in groups that lack discernible organization (e.g., Schuetzia bennieana), while in other examples a few sporangia may be arranged in a uniseriate whorl (e.g., Telangium, Zimmermannitheca). This suggests that within the seed fern complex, synangia were evolving and becoming established at this time. Other fossils from this period are still problematical; for example, little is known about the organization of Paracalathiops, other than it appears synangiate, and it is not certain whether Simplotheca represents synangia or simple sporangia. Anatomical information about these forms is necessary because some interesting types appear to be present that may incorporate presumably sterile peripheral elements. At this evolu- tionary stage it is especially difficult to discern the familial relationships of a given fossil. It seems likely, however, that early medullosans had aggregations of both large and small numbers of sporangia, while lygi- nopterids had only small sporangial aggregations.
By the end of the Mississippian it appears likely that most seed ferns probably had synangia that consisted either of a uniseriate ring of spo- rangia, or a poorly organized cluster of basally fused sporangia. Lower Pennsylvanian synangia all appear to be of the uniseriate ring type. In most cases the synangia have an open area distally that is delineated by the sporangial tips (e.g., Aulacotheca), but some may have become bi- laterally flattened in which case the central area is lost (Feraxotheca, Whittleseya). All forms are thought to have longitudinal dehiscence of the sporangia in which the contents are released toward the interior of the synangium.
The Mississippian specimens of Telangium described by Jennings (1976) consist of a uniseriate whorl of thin walled sporangia, and were probably derived from a terminal cluster of elongate sporangia (e.g., Zim- mermannitheca). Vascular tissue in these forms is restricted to the spo- rangium pedicel or base. Stratigraphically younger species of Telangium (Westphalian A) are thick walled with vascular tissue in each sporangium. Other lyginopterid pollen organs of a comparable age (e.g., Feraxotheca) are also thick walled, but vascular tissue is restricted to the synangium base. Evolution of lyginopterid pollen organs appears to be in the direc- tion of thicker walled sporangia with a corresponding reduction of the central open area of the synangium.
Fig. 18. A-F, Prepollen or pollen types; A, Potoniea, x800; B, Crossotheca, x800; C, Schulzospora, x800; D, Parasporotheca, x200; E, Idanothekion, x 1,500; F, Dolerotheca, • 180.

362 THE BOTANICAL REVIEW
With a few exceptions, Pennsylvanian pollen organs possess vascular tissue near the outer surface (away from the dehiscence area) of each sporangium. Such an arrangement might suggest that the sporangia were initially vascularized prior to the formation of synangia. Unfortunately, the absence of vascularized sporangia of Mississippian age detracts from this hypothesis. We would suggest that the vascularization of sporangia, at least in the seed ferns, is correlated with either the evolution of synan- gia, or the thickening of the outer facing sporangial walls.
In the lyginopterid pollen organ Crossotheca there is a more complete incorporation of the fertile branching system into the planated sterile frond. The primitive condition in this group may be demonstrated by the three-dimensional fertile branches of Telangium, that presumably occu- pied a portion of an otherwise sterile planated frond. Synangia of Cros- sotheca occupy the same position as they do in Telangium, but the synan- gia are pendent on a pinnate branching system that develops some pinnular foliage on the frond. Other than reduction of the fertile frond resulting in the basal fusion of several synangia (Fig. 13C), little modifi- cation appears to have occurred in Crossotheca synangia over a rather long geological history.
The pinnule borne synangia of the Callistophytaceae appear most closely related to the lyginopterid seed ferns. They have small radial synangia similar to early forms of Telangium, that are borne on the lower surface of apparently unmodified pinnules. We envision that the family originated from the lyginopterid complex sometime in the Early Penn- sylvanian. Most organs of the plant Callistophyton appear considerably advanced when compared to contemporaneous lyginopterids, and evo- lution must have occurred rapidly in order to achieve the clear delimi- tation of this family by the Middle Pennsylvanian. It is interesting that apparently little change took place in the pollen organs of this family from Middle to Late Pennsylvanian.
Within the medullosan seed ferns the vast array of pollen organ types appears to have been present by the Middle Pennsylvanian. New orga- nizations at this time (Fig. 16) include both clustering to form aggregations and fusion to form compound synangia. The aggregation of small Aula- cotheca-like synangia to form the Rhetinotheca cluster is probably re- lated to the reduction of supporting branches. The compound synangium characteristic of Potoniea probably had a similar developmental origin, but the microspore type and probable frond morphology (paripinnate neu- ropterids) suggests that the synangia comprising this unit had an early evolutionary origin. In addition, the radial nature of the concentric rings making up the pollen organ implies that the fertile frond unit was origi- nally radial. In Dolerotheca the four units of the compound synangium

PALEOZOIC SEED FERNS 363
are side by side, indicating that lateral fusion occurred on already pla- nated axes.
One of the more interesting evolutionary modifications seen in the large solitary medullosan pollen organs like Sullitheca and Stewartiotheca, is the convolution or plication of the internal dehiscence areas. Such a change increases the surface area available for sporangial dehiscence zones while keeping the external surface area of the synangium relatively small. Codonotheca is a large synangium that may also have been or- ganized in this manner, but is presently incompletely known.
The aggregation of large synangia is evident in Parasporotheca, in which large fan-shaped synangia are appressed into a mass. Distinctive peglike hairs cover the surface of the synangia and interdigitate with those of neighboring synangia to hold the individual units together. A similar arrangement has been hypothesized for Whittleseya, but the presence of interdigitating pegs has not yet been confirmed.
The recent discovery that Dolerotheca formosa has a compound cam- panulum consisting of four, large, internally convoluted synangia con- firms the hypothesis that fusion and internal elaboration are the primary modes of pollen organ evolution in the Pennsylvanian medullosans. No doubt many other medullosan pollen organs will be discovered that have one of the forms discussed here, but it is difficult to envision any new variations that are not already known for the family.
The manner in which medullosan pollen organs were borne is known in many cases to be on small entirely fertile fronds or frond segments. Where not known for certain (e.g., Dolerotheca, Whittleseya) there has been a tendency to assume that the pollen organs replaced a pinnule or pinna on an otherwise sterile frond such as has been suggested for the ovules. A revitalization of interest in compression floras holds promise of discovering specimens in which large pollen organs will be found at- tached to the frond segments.
Branching Patterns
Currently, our knowledge of how seed fern pollen organs were borne on the plant is in most instances incomplete. Certainly, the most reliable evidence has been generated from impression-compression remains, al- though compressed fertile remains of many seed fern groups constitute rare occurrences in the fossil record. Structurally preserved pteridosperm fertile parts, and the position these organs occupy on the parent plant, are known only for the Callistophytaceae, and to a limited extent for members of the Lyginopteridaceae.
Despite the numerous reconstructions of the Carboniferous seed fern Lyginopteris present in the literature, so far as we are aware the manner

364 THE BOTANICAL REVIEW
in which the fertile parts were borne has not been conclusively demon- strated in this genus. It has generally been assumed that synangia were borne in an upright manner at the apices of a naked branching system. Such a branching system could presumably demonstrate varying levels of planation, and might replace an entire leaf on the stem, or represent a fertile portion of an otherwise sterile planated frond. For example, relatively extensive specimens of the Telangiopsis branching system lack sterile foliage (Fig. 15F), and appear to have been three-dimensional in organization (Eggert and Taylor, 1971). On the other hand, in Telangium sp. (Fig. 4), also from the Upper Mississippian, the large frond axes of the plant show a distinct planation (Jennings, 1976).
Possibly the most frondlike organization of the fertile segments that exists among lyginopterids is present in Crossotheca and Feraxotheca. In these genera the synangia have reduced pinnular segments basally (Fig. 2F) and are positioned at the tips of the ultimate pinnae of the planated, pinnate frond segment (Figs. 1I, 5, 15D). These fertile pinnae have been described as occurring at the apex, near the middle, or at the base of otherwise sterile fronds in Crossotheca. A pinnate branching system like that of Crossotheca and Feraxotheca has been described in Canipa quadrifida (Skog et al., 1969), however, no laminar tissue is known to be associated with the synangia in this taxon.
In another possible lyginopterid seed fern, interesting because of its eremopterid foliage, the fertile frond is three times pinnate (Fig. 15C). The ultimate pinnae are thought to have produced either bicornute ovules or clusters of basally fused sporangia (Delevoryas and Taylor, 1969). Unlike the previously noted lyginopterids, the smaller fertile fronds of Eremopteris appear to be equivalent to entire sterile fronds.
In the Callistophytaceae the fronds are relatively small and pinnate (Rothwell, 1975). In this family the pollen organs have been described as occurring on the abaxial surface of pinnules in both Middle and Upper Pennsylvanian specimens (Stidd and Hall, 1970a; Rothwell, 1972b). Al- though the morphology of the pinnules is only partially known, synangia are positioned near the tips of widely spaced, dichotomizing veins (Fig. 14D). In a recent report (Rothwell, 1979) the Pennsylvanian ovules are thought to occupy a position on the lower pinnule surface. The position of fertile structures in this seed fern family appears to be consistent with the generalization that pteridosperm ovules and pollen organs tend to be borne in similar ways on the plant.
Although pollen organs and ovules appear to be produced on the same morphological structures in the Callistophytaceae (pinnules) and Lygi- nopteridaceae (pinnae), the Medullosaceae exhibit more variability in where fertile parts are borne. While the position that ovules occupy in this family continues to remain conjectural, there is an increasing con-

PALEOZOIC SEED FERNS 365
sistency with reference to where pollen organs are borne. In Aulacotheca the small synangia are borne in clusters on many times dichotomized distal branches that constitute segments of a large fertile frond system. To date, only one extensive branching system is known (Eggert and Kry- der, 1969), although clusters of synangia have been reported several times (Kidston, 1906). The consistently poor preservation of branches associ- ated with these synangia suggests that the frond segments bearing them were of a fragile nature and not easily preserved. The stalked synangia of Goldenbergia are terminal, on a several times pinnate fertile frond system (Fig. 13D). Halle (1933, Text-Fig. 4A) reconstructs Goldenbergia with a series of elongate stalks each of which bears a single pollen organ. Halle illustrates several specimens in which the stalks dichotomize and bear two synangia (1933, P1. 1, Figs. 1-3). This would suggest that at distal levels the tripinnate fertile frond is not planated.
A problematical branching system bearing possible pollen organs has been attributed to the medullosan foliage type Neuropteris heterophylla (Kidston, 1887, 1904). This specimen was later illustrated by Halle (1933, P1. 10, Figs. 3-5), and shown to consist of small campanulate structures borne terminally in pairs on a branching system (Fig. 15E). It is inter- esting to note that at several levels of the specimen ovate pinnules are borne on the fertile branches. Halle suggests that this specimen could be a cupule, and further notes that the affinities with N. heterophylla are unproven.
The presence of paired synangia on a branching system is a common morphological pattern exhibited by several groups of seed ferns. For example, in Codonotheca (Fig. 15B) several specimens have been de- scribed in which the paired synangia appear to be borne along the side of a common supporting branch (Sellards, 1907; Darrah, 1970). The pres- ence of paired synangia was also indicated for the Mississippian genus Paracalathiops (Fig. 2C). In Paracalathiops the paired synangia are adaxially directed and borne on dichotomizing axes; the abaxial surface of the pinna axis lacks appendages. In the Permian species of Schuetzia and Dictyothalamus the synangia are borne alternately on opposite sides of the fertile pinna (Fig. 15A). The overall morphology of these fertile fronds is not known since only the distal portions were preserved, and consequently important features such as the presence or absence of ster- ile foliage remains speculative.
It has been widely accepted that the larger medullosan pollen organ types (e.g., Dolerotheca, Whittleseya) were borne in the place of pinnae or pinnules on otherwise sterile fronds. Such suggestions have been sup- ported by the occasional discovery of compressed ovules occupying these positions, and by the occurrence of some problematical associations of pollen organs and sterile foliage. The occurrence of large ovules occu-

366 THE BOTANICAL REVIEW
pying the position of a terminal or lateral pinnule has been reported from Carboniferous deposits many times (Arnold, 1935; Bertrand, 1913; Hem- ingway, 1941a; Jongmans, 1954; Kidston, 1904, 1914). Ovules of the Rhabdocarpus and Trigonocarpus-type, have long been regarded as the megasporangiate unit of the medullosans. Unfortunately, no structurally preserved specimens have been discovered that confirm this ovule type as being borne in the place of a pinnule. In addition, there are several reports of large ovules borne at the tips of dichotomizing branches (Kid- ston and Jongmans, 1911; Stockmans and Willi~re, 1961). Although the precise position of the large Trigonocarpus ovule is still a matter of con- jecture, smaller Pennsylvanian seeds are known to have been borne on the lower surface of Alethopteris pinnules (Raymond Pheifer, personal communication). The confirmation of abaxially borne ovules is further supported by a number of similar forms reported by Halle (1927, 1929, 1932) from Permian deposits of China. We believe that these reports are the basis for homologizing pollen organ and ovule positions.
We are aware of no specimens that can be included in the Medullo- saceae based upon pollen type in which the position of the pollen organ is comparable to the previously discussed ovules. In 1932, Dix described a compression specimen consisting of an elongate seedlike body contain- ing spores borne on the upper surface of a pinna bearing pinnules. Later, Hemingway (1941b) demonstrated that this pollen organ (Aulacotheca dixiana) was not organically attached to the pinna, but rather was com- pressed on the surface and represented a chance juxtaposition of plant parts. One rarely cited pollen organ that has been described as occurring on pinnule bearing pinnae is Givesia (ex. Whittleseya) media described by Jongmans (1954) from the Limberg Coalfield of The Netherlands. Based on the available illustrations of the foliage and associated pollen organs, we are not convinced that these two plant parts are organically attached in the manner indicated. As we have already noted, Whittleseya- like synangia are generally found abundantly scattered in the matrix, and the chance for accidental superposition on foliage is great. A similar oc- currence of a campanulate structure closely associated with Alethopteris serli foliage (Buisine, 1961, PI. XI), is not well enough preserved to allow a final determination.
In a recent paper by Ramanujam et al. (1974) the frond position of Dolerotheca synangia is hypothesized based upon the vasculature in the penduncle of the pollen organ. Based on features of the vasculature these authors suggest that the pollen organs occupied positions equivalent to the ultimate pinnae. Moreover, their reconstruction suggests that Doler- otheca campanula replace ultimate pinnae on the sterile frond. Although the vasculature of the Myeloxylon petiole and the peduncle of the Do- lerotheca synangium may appear identical, there is no evidence at this

PALEOZOIC SEED FERNS 367
time that indicates these pollen organs were borne among the sterile fo- liage members. As the earlier discussion concerning a new interpretation of the Dolerotheca synangium suggests, the compound nature of the pol- len organ and vascular system may reflect the origin of the unit from a single pinna (Fig. 11). Such an ultimate pinna would bear simple synangia in the position of pinnules as is suggested for Codonotheca (Fig. 15B) and Goldenbergia (Fig. 13D). In light of our knowledge concerning the organization of other pollen organs, an alternative view regarding the manner in which Dolerotheca synangia are borne may be equally plau- sible. For example, synangia of Dolerotheca may have been borne on completely fertile pinnae in a manner similar to a number of compression forms. Support for such a morphological organization is suggested by the relatively large number of Paleozoic pollen organs that appear to be borne in this manner. Although such ~'negative" evidence should not bias pa- leobiologists in their attempts to accurately reconstruct entire fossil plants, the repeated occurrences of certain morphological arrangements in similar organisms can help to more clearly characterize natural familial units.
Prepollen
A survey of seed fern prepollen suggests the presence of four easily recognizable types. These include large pseudosaccate grains found in some Mississippian seed ferns, the small, radial trilete forms character- istic of the Lyginopteridaceae, the large Monoletes type found in most medullosan seed ferns, and the small, monosaccate, true pollen present in the Cailistophytaceae. A stratigraphic analysis of the Mississippian seed fern microspore types indicates that the various saccate forms were as widely distributed and abundant as the radial, trilete, nonsaccate forms. These characteristic microspores or prepollen types of the earliest seed ferns are morphologically similar to the grains of Devonian progym- nosperms. The microspores of most Devonian progymnosperms are large (100-200/~m), radial in organization, and characterized by a large, sym- metrical trilete suture. In these forms there may be a distinct separation of the exine that results in a spherical central body (corpus) surrounded by a thin, irregular envelope (pseudosaccus). Despite the importance of determining the developmental patterns and ultrastructural organization of grains of this type, it is unfortunate that detailed studies of grain ul- trastructure in these forms has not been undertaken. Such studies would be useful in determining ultrastructural homologies necessary in estab- lishing systematic affinities, and also in suggesting possible phylogenetic trends in grain organization and function. Because of these limitations it is necessary to rely on presumed homologies that have been formulated utilizing geologically younger prepollen grains of a similar morphological

368 THE BOTANICAL REVIEW
type. For example, in the grains of the medullosan seed fern Paraspo- rotheca (Fig. 18D), the two crescent-shaped sacci originate from a sep- aration within the outer exine layer (sexine). Therefore, the large central body (nexine) is covered by a layer of sexine. This sexinous layer imparts a granulose appearance to the corpus similar to that present on the sacci. We would suggest that this organization is both widespread and basic in the early seed fern groups in which saccate microspores are known. Such an organization has been suggested for Rhabdosporites (Millay and Tay- lor, 1974), the spore type of the progymnosperm Tetraxylopteris, and also appears probable for the saccate microspores of several Mississip- pian seed fern microsporangiate structures (e.g., Paracalathiops, Sim- plotheca).
Another spore type in which a superficial layer is separable from the spore body appears distinct from the previously discussed saccate types. For example, the spores of Psilophyton dawsonii possess a thin exter- nally ornamented layer that closely invests the smooth spore body (Banks et al., 1975). The origin of this layer is problematical. It may constitute an extra-exinous tapetal residue (perispore) or may result from some form of mechanical separation of the sporoderm occurring naturally during development. A similar appearing membranous layer has also been re- ported attached to the sporoderm of Alcicornopteris (Smith, 1962a), Pro- topitys (Walton, 1957), and Potoniea (Laveine, 1971; Stidd, 1978b).
A third spore type that characterizes some later seed ferns, particularly those of the Lyginopteridaceae, is spherical and trilete. These grains lack both an internally differentiated sexine and extra-exinous layer. Spores of this type are known from the Namurian A microsporangiate fructifi- cation Zimmermannitheca (Remy and Remy, 1959b), in which the ar- rangement and position of the sporangia suggest a phylogenetic aggre- gation of sporangia into a radial synangiate unit. The basic morphology of Zimmermannitheca spores is common in stratigraphically younger seed fern pollen organs such as Crossotheca, Feraxotheca, and Telan- giopsis. To date the ultrastructure of this grain type has been studied only in Crossotheca. These spores possess a thin homogeneous nexine that supports an appreciably thicker sexine lacking infrastructure. The surface of the sexine is ornamented by numerous, closely spaced papillae (Fig. 18B). Like Crossotheca, the spores of Telangiosis and Feraxotheca possess small, evenly spaced surface ornament.
At the present time the origin of the basic medullosan spore type is uncertain. Monoletes (Fig. 18F), the spore type characteristically asso- ciated with the family, circumscribes ellipsoidal monolete grains that range from 100-560/zm long. The sporoderm is constructed of two well- defined exine components. The inner nexine layer is thick (up to 3/zm)

PALEOZOIC SEED FERNS 369
and constructed of delicate, uniformly thickened and stratified lamellae (Taylor, 1978). The outer wall layer (sexine) is internally sculptured, be- cause the wall is filled with continuous (overlapping) or discontinuous iumina. Millay et al. (1978) indicate that the ultrastructure of the monolete suture of Monoletes and Crossotheca shows the same basic organiza- tional relationships between the nexine and sexine, as has been reported in the spores of homosporous pteridophytes (Lugardon, 1972). This sim- ilarity in wall organization in the region of the suture suggests a potential functional homology that has prompted some authors to apply the term prepollen to the grains of these early seed plants. That a large variety of medullosan prepollen exists is shown by the sporae dispersae grain type Nanoxanthipollenites (Clendening and Nygreen, 1976), which is now known to exhibit the alveolate exine infrastructure (Taylor, 1979). Al- though relatively small (88-135 ~m long) for a grain of this type, and characterized by coarse spines, this prepollen possesses the distinctive median deflection of the monolete suture.
Two additional prepollen types thought to have medullosan affinities appear to compare more favorably with the microspores of some early seed plants. In one example, Potoniea, the microspores are radial, trilete, and characterized by the presence of a delicate membrane surrounding a thick homogeneous exine (Fig. 18A). At the light microscopic level there is little to distinguish these grains from the "perinate" microspores of some early pteridosperms (e.g., Alcicornopteris and Staphlotheca).
In another form, the saccate medullosan grain Parasporites, the ultra- structure of the sporoderm is similar to that present in Monoletes (Millay et al., 1978), and the two crescent-shaped sacci lack endoreticulations on the inner walls. The absence of this structural feature provides a sug- gested evolutionary link between the Pennsylvanian Parasporites type, and earlier, monosaccate types known to have been produced by several seed fern pollen organs (e.g., Simplotheca, Paracalathiops).
Presently only one Paleozoic seed fern microspore type is known to have functioned like modern pollen. This form, unique to the Callisto- phytaceae, is characterized by a girdling monosaccus that contains well- developed reticulations lining the saccus. Stages in the evolution of pollen of this type from possible lyginopterid forms has been suggested (Millay and Taylor, 1976). In these grains the sporoderm of the central body is thin and laminated, and germination is known to have occurred via a pollen tube that developed from the distal pole of the grain (Rothwell, 1972a). On the basis of general grain organization, microgametophyte development, and the presence of the apparently siphonogamous pollen tube, callistophytacean pollen is regarded as the most evolutionarily ad- vanced pollen type found within the Paleozoic seed ferns.

370 THE BOTANICAL REVIEW
S U M M A R Y
1. The earliest probable pteridosperm microsporangiate structures are nonsynangiate and closely resemble those of presumed progymno- sperm ancestors.
2. Early Mississippian pteridosperm microsporangia are elongate, thin walled, unvascularized, and sometimes lack specific dehiscence areas.
3. Microsporangia and early synangia were borne on three-dimensional, entirely fertile branches. Later forms show more planation of the branching system and encroachment or incorporation of sterile foliar units. Abaxial synangia on pinnules is presumably the most advanced condition.
4. Several genera from Namurian A sediments show clustering of the microsporangia at stem tips but are not known to be synangiate.
5. Lyginopterid synangia consist of a simple ring of sporangia that usu- ally surround a hollow central area, and may be radial or bilateral in symmetry. No synangia in this family are presently known to be aggregated or compound.
6. Pennsylvanian medullosan pollen organs basically consist of a ring of sporangia that usually surround a central hollow, and may be radial or bilateral in symmetry.
7. Medullosan pollen organs may be simple and solitary, or clustered into aggregates. Some forms represent compound organs composed of several to many simple synangia developmentally fused together. Compound synangia may be radial or bilateral in symmetry.
8. Synangial variability in medullosan seed ferns includes: a simple ring of four to numerous sporangia (e.g., Aulacotheca); an incomplete ring or crescent of sporangia (e.g., Parasporotheca); an internally folded or plicated "ring" of sporangia (e.g., Sullitheca).
9. Permian pollen organs are still problematical with regard to basic morphology and systematic affinities. In situ pollen is of unknown significance taxonomically for these organs.
10. Initially only two microspore types are recognizable in pterido- sperms; both are radial and trilete but an exine separation (pseudo- saccus) is present in one form (e.g., Remysporites).
11. By Upper Mississippian time prepollen morphology was established in the Lyginopteridaceae, which include radial, trilete grains with delicate ornamentation. In the Medullosaceae a similar organization was present in the prepollen of one taxon (Potoniea), while other prepollen types are monolete with a reduced third laesura and an alveolate sexine (e.g., Monoletes). The small monosaccate pollen of the Callistophytaceae is morphologically and functionally the most advanced Pennsylvanian type.

PALEOZOIC SEED FERNS 371
ACKNOWLEDGMENTS
We wish to thank Dr. Benton M. Stidd for making specimens of Po- toniea and Sullitheca available for illustration, Dr. Robert M. Kosanke for providing specimens of Schulzospora, and Dr. Jorge V. Crisci for the Spanish translation of our abstract. We gratefully acknowledge Mr. David M. Dennis, staff artist, who rendered the many line drawings illustrating this paper. This research was in part funded by the National Science Foundation through grants DEB 76-01714 and DEB 77-02239.
LITERATURE CITED
A n d r e w s , H. N. and T. L. Philips. 1968. Rhacophyton from the Upper Devonian of West Virginia. J. Linn. Soc. (Bot.) 61: 37-64.
Arnold, C. A. 1935. The seeds of Alethopteris, and other pteridosperms from North America. Congr. Stratigr. Carb. Heerlen, pp. 41-45.
�9 1949. Fossil flora of the Michigan Coal Basin. Contrib. Mus. Paleontol. Univ. Mich. 7: 131-269.
- - a n d W. E. Steidtmann. 1937. Pteridospermous plants from the Pennsylvanian of Illinois and Missouri. Am. J. Bot. 24: 644-650.
Banks, H. P., S. Leelercq, and F. M. l-Iueber. 1975. Anatomy and morphology of Psilo- phyton dawsonii, sp. n. from the Late Lower Devonian of Quebec (Gasp6). and On- tario, Canada. Palaeontogr. Am. 8: 77-127.
Baxter, R. W. 1949. Some pteridosperm stems and fructifications with particular reference to the Medullosae. Ann. Mo. Bot. Gard. 36: 287-352.
Beck, C. B. 1976. Current status of the Progymnospermopsida. Rev. Palaeobot. Palynol. 21: 5-23.
Benson, M. 1904. Tc, langium Scotti, a new species of Telangium (Calymrnatotheca) show- ing structure. Ann. Bot. 18: 161-177.
�9 1935. The fructification, Calathiops bernhardti, n. sp. Ann. Bot. 49: 155-160. Bertrand, P. 1913. Les fructifications de Nevropt6rid6es recueillies dans le terrain houiller
du Nord de la France. Ann. Soc. Geol. Nord 42:113-144. Bonamo, P. M. 1977. Rellimia thomsonii (Progymnospermopsida) from the Middle De-
vonian of New York State. Am. J. Bot. 64: 1272-1285. - - a n d H. P. Banks. 1967. Tetraxylopteris schmidtii: its fertile parts and its relation-
ships within the Aneurophytales. Am. J. Bot. 54: 755-768. Buisine, M. 1961. Contributions a l'6tude de la flore du terrain houiller. Les Al6thopt6ridees
du nord de la France. Etudes geol. Atlas Top. sout., H.B.N.P.C., l.-Flore fossile, 4 ~ fasc., Texte, pp. 1-317, Atlas, PI. I-LXXIV.
Carpentier, A. 1911. Sur quelques fructifications et inflorescences du Westphalien du Nord de la France. Rev. Gen. Bot. 23: 441-458.
�9 1914. Note sur des empreintes de Whittleseya(?)fertilis Kidston sp. trouv6es dans le Houiller du Nord de la France. Bull. Soc. Bot. Ft. 14(ser. 4): 363-367.
�9 1925. Note sur quelques empreintes de graines et microsporanges de pteridosper- m6es provenant du Westphalien du Nord de la France. Rev. Gen. Bot. 37: 145-156.
�9 1929. Empreintes de fructifications trouv6es en 1929 dans le Westphalien du Nord de la France. Rev. G6n. Bot. 41: 469-473.
Chaioner, W. G. 1970. The evolution of miospore polarity. Geosci. Man 1: 47-56. , A. J. Hill, and W. S. Lacey. 1977. First Devonian platyspermic seed and its im-
plications in gymnosperm evolution. Nature 265: 233-235. C l e n d e n i n g , J. A. and P. W. Nygreen. 1976. Nanoxanthipollenites. a new monolete mio-
spore from the Upper Pennsylvanian of Kansas. Geosci. Man 15: 125-128. Danz/~, J. 1955. Le genre Crossotheca Zeiller. C. R. Acad. Sci. 241: 1616-1618.

372 THE BOTANICAL REVIEW
- - - . 1960. Remarques sur quelques fructifications de foug/~res Carbonif'eres. C. R. Quatr. Congr~s pour l 'Avancm, l~tudes Strat. Geol. Carbonif~re, Heerlen 1: 93-99.
Darrah, W. C. 1939. The fossil flora of Iowa coal balls. II. The fructification of Botryop- teris. Harvard Univ. Bot. Mus. Leafl. 7: 157-180.
�9 1970. A critical review of the Upper Pennsylvanian floras of eastern United States with notes on the Mazon Creek Flora of Illinois. W. C. Darrah (1969), Gettysburg, 300 PP.
Delevoryas, T. 1964. A probable pteridosperm microsporangiate fructification from the Pennsylvanian of Illinois. Palaeontology 7: 60-63.
and T. N. Taylor. 1969. A probable pteridosperm with eremopterid foliage from the Allegheny Group of northern Pennsylvania, Postilla 133: 1-14.
Dennis, R. L. and D. A. Eggert. 1978. Parasporotheca, gen. nov., and its bearing on the interpretation of the morphology of permineralized medullosan pollen organs. Bot. Gaz. 139: 117-139.
Dix, E. 1932. On a sporocarp probably attached to a frond of Neuropteris schlehani Stur. Ann. Bot. 46: 1064-1067.
Eggert, D. A. and R. W. Kryder. 1969. A new species of Aulacotheca (Pteridospermales) from the Middle Pennsylvanian of Iowa. Palaeontology 12: 414-419.
and G. W. Rothwell. 1979. Stewartiotheca gen. n. and the nature and origin of complex permineralized medullosan pollen organs. Am. J. Bot. 66: 851-866.
and T. N. Taylor. 1971. Tetangiopsis gen. nov., an Upper Mississippian pollen organ from Arkansas. Bot. Gaz. 132: 30-37.
Geinitz, H. B. 1863. Uber zwei neue dyadische Pflanzen. Neues Jahrb. Miner., Geol. Palaeon. pp. 525-530.
Gillespie, W. H. and J. A. Clendening. 1966. West Virginia plant fossils, 1. Dolerotheca and Daubreeia. Proc. W. Va. Acad. Sci. 38: 159-168.
Goeppert, H. R. 1850. Monographie der fossilen Coniferen. Natuurk. Verh. Holland. Maatsch. Wetens. Haarlem, G, Leiden. 286 pp.
- - . 1864-1865. Die fossile Flora der Permischen Formation. Palaeontographica 12: 1- 224, pls. 1-40 Q864); 225-316, pls. 41-64 (1865).
Gothan, W. 1927. Uber einige Kulmpflanzen vom Kossberg bei Plauen im Vogtlande. Abh. Sachs. Geol. Landesamts 5: I-8.
~ . 1941. /,3ber die Samen und Pollenorgane von Lonchopteris rugosa. Jahr. Reichs- stelle Bodenforsch. 1940. 61: 278-282.
Halle, T. G. 1927. Palaeozoic plants from Central Shansi. Palaeontol. Sin. 2A: 317 pp. ~ . 1929. Some seed-bearing pteridosperms from the Permian of China. K. Sven.
Vetenskapsakad. Handl. ser. 3, 6: 1-24. ~ . 1932. On the seeds of the pteridosperm Emplectopteris triangularis Halle. Bull.
Geol. Soc. China 11: 39-306. ~ . 1933. The structure of certain fossil spore-bearing organs believed to belong to
pteridosperms. K. Sven. Vetenskapsakad. Handl. ser. 3, 12:103 pp. ~ . 1942. Some specimens of Potoniea from the Carboniferous (Westphalian) of Bel-
gium. Bull. Mus. R. Hist. Nat. Belg. 18: 1-7. Hemingway, W. 1941a. Neuropteris tenuifolia with carports attached. Ann. Bot. N.S. 5:
193-196. ~ . 1941b. On the Coal-Measure plant, Aulacotheca. Ann. Bot. N.S. 5: 197-201. Jennings, J. R. 1976. The morphology and relationships of Rhodea, Telangium, Telan-
giopsis, and Heterangium. Am. J. Bot. 63:1119-1133. Jongmans, W . J . 1954. Contribution to the knowledge of the flora of the seam Girondelle
(lower part of the Westphalian A). Part I. Meded. Geol. Stichting-Ser. C, III-1: 5-16. Kidston, R. 1884. On a new species of Schutzia from the Calciferous Sandstone of Scot-
land. Ann. Mag. Nat. Hist. 13: 77-80. ~ . 1887. On the fructification of some ferns from the Carboniferous formation. Trans.
R. Soc. Edinburgh 33: 137-156. - - . 1904. On the fructification of Neuropteris heterophylta. Brongniart. Philos. Trans.
R. Soc. London 197B: 1-5. ~ . 1906. On the microsporangia of the Pteridospermeae with remarks on their rela-
tionship to existing groups. Philos. Trans. R. Soc. London 198B: 413-445.

PALEOZOIC SEED FERNS 373
�9 1914. On the fossil flora of the Staffordshire Coal Fields. Part IIl .--The fossil flora of the Westphalian Series of the South Staffordshire Coal Field. Trans. R. Soc. Edin- burgh 50: 73-190.
�9 1923. Fossil plants of the Carboniferous rocks of Great Britain. Mem. Geol. Surv. Great Britain, Palaeontology 2, part 4, pp. 275-376.
�9 1924. Fossil plants of the Carboniferous rocks of Great Britain. Mem. Geol. Surv. G.B. Palaeontology 2, part 5, pp. 377-522.
- - a n d W. J. Jongmans. 1911. Sur la fructification de Neuropteris obliqua Bgt. Arch. Neerl. Sci. Exactes Nat. Set. 3B, 1: 1-2.
L a v e i n e , J.-P. 1971. Sporomorphes in situ de quelques Parispermfes (Neuroptfridfes) du Carbonif~re. Ann. Soc. G~ol. Nord 91: 155-173.
L e i s m a n , G. A. and J. S. Peters. 1970. A new pteridosperm male fructification from the Middle Pennsylvanian of Illinois. Am. J. B o t . 5 7 : 867-873.
L u g a r d o n , B. 1972. La structure fine de l'exospore et de la pfrispore de filicinfes iso- sporfes. I--Gfnfrali tfs . Eusporangifes et Osmundales. Pollen Spores 14: 227-261.
Millay, M. A. 1976. Synangia of North American Pennsylvanian petrified marattialeans. Ph.D. Thesis, Univ. Illinois.
- - . 1978. Studies of Paleozoic marattialeans: The morphology and phylogenetic po- sition of Eoangiopteris goodii sp. n. Am. J. Bot. 65: 577-583.
- - . 1979. Studies of Paleozoic marattialeans: A monograph of the American species of Scolecopteris. Palaeontographica. 169B: 1-69.
- - a n d D. A. Eggert. 1970. ldanothekion gen. n., a synangiate pollen organ with saccate pollen from the Middle Pennsylvanian of Illinois. Am. J. Bot. 57: 50-61.
, - - , a n d R. L. Dennis. 1978. Morphology and ultrastructure of four Pennsyl- vanian prepollen types. Micropaleontology. 24:303-315.
- - a n d T. N. Taylor�9 1974. Morphological studies of Paleozoic saccate pollen. Palae- ontographica 147B: 75-99.
- - a n d - - . 1976. Evolutionary trends in fossil gymnosperm pollen. Rev. Palaeo- hot. Palynol. 21: 65-91.
- - and - - . 1977. Feraxotheca gen. n., a lyginopterid pollen organ from the Penn- sylvanian of North America. Am. J. Bot. 64: 177-185.
- - a n d . 1978. Fertile and sterile frond segments of the lyginopterid seed fern Feraxotheca. Rev. Palaeobot. Palynol. 25:151-162.
N e w b e r r y , J. S. 1853a. Fossil plants from the Ohio coal basin. Ann. Sci., Cleveland 1: 106-108.
�9 1853b. New fossil plants from Ohio. Ann. Sci., Cleveland 1:116-117. P e t t i t t , J. M. and C. B. Beck. 1968. Archaeosperma arnoldii--A cupulate seed from the
Upper Devonian of North America. Contrib. Mus. Paleontol. Univ. Mich. 22: 139- 154.
Ramannjam, C. G. K., G. W. Rnthweli, and W. N. Stewart�9 1974. Probable attachment of the Dolerotheca campanulum to a Myeloxylon-Alethopteris type frond. Am. J. Bot. 6 1 : 1057-1066.
Remy, R. and W. Remy. 1955. Simplotheca silesiaca n. gen. et sp. Abh. Dtsch. Akad. Wiss. Berlin K1. Chem. Geol. Biol. 2: 1-7.
Remy, W. 1953a. Untersuchungen fiber einige Fruktifikationen von Farnen und pterido- spermen aus dem Mitteleuropaischen Karbon und Perm. Abh. Dtsch. Akad. Wiss. Berlin Kl. Math. Allg. Naturwiss. 2: 4-38.
�9 1953b. Beitr~ige zur Kenntnis der Rotliegendflora Thiiringens. Sitzungsber. Dtsch. Akad. Wiss. Berlin KI. Math. Allg. Naturwiss. 1: 1-24.
�9 1953c. Reticulopteris ("Linopteris") odontopteroides n. sp. und die dazugehfrige Fruktifikation. Z. Geol. 2: 146-149.
�9 1953d. Unguitheca peeopteroides n. gen. et sp. sowie eine weitere Fruktifikation aus dem Rotliegenden von Ilfeld. Z. Geol. 2: 373-377.
- - a n d R. Remy. 1958. Die sporen von Dictyothalamus schrollianus G6ppert. Abh. Dtsch. Akad. Wiss. Berlin KI. Chem. Geol. Biol. 5: 1-6.
- - a n d . 1959a. Pflanzenfossilien. Akademie, Berlin. - - and - - . 1959b. Zimmerrnannitheca cupulaefi~rmis n. gen. n. sp. Monatsber.
Dtsch. Akad. Wiss. Berlin 1: 767-776.

374 THE BOTANICAL REVIEW
- - a n d R. Rettschlag. 1954. Neue Untersuchungen fiber die Pollen von Schuetzia anomala H. B. Geinitz. Geologie 3: 582-589.
RothweU, G. W. 1972a. Evidence of pollen tubes in Paleozoic pteridosperms. Science 175: 772-774.
- - . 1972b. Pollen organs of the Pennsylvanian Callistophytaceae (Pteridospermop- sida). Am. J. Bot. 59: 993-999.
- - . 1975. The Callistophytaceae (Pteridospermopsida): I. Vegetative structures. Pa- laeontographica 151B: 171-196.
- - . 1979. The Callistophytaceae (Pteridospermopsida): II. Reproductive features. Pa- laeontographica. (In press.)
Schimper, W.P. 1870-1872. Trait6 de pal6ontologie vdg6tale, ou la flore du monde primitif. J. B. Bailli~re et ills, Paris, 2: 1-522(1870), 523-968(1872).
Schopf, J. M. 1948. Pteridosperm male fructifications: American species of Dolerotheca, with notes regarding certain allied forms. J. Paleontol. 22: 681-724.
Schuster, J. 1911. Uber die Fruktifikation von Schuetzia anomala. Sitzungsber. Kais. Akad. Wiss. Wien, KI. Math.-Natur. 120:1125-1134.
Sellards, E. H. 1903. Codonotheca, a new type of spore-bearing organ from the Coal Measures. Am. J. Sci. 16: 87-95.
�9 1907. Notes on the spore-bearing organ Codonotheca and its relationship with the Cycadofilices. New Phytol. 6: 175-178.
Skog, J. E., H. N. Andrews, and S. H. Mamay. 1969, Canipa quadrifida, gen. et sp. nov., a synangial fructification from the Middle Pennsylvanian of West Virginia. Bull. Torrey Bot. Club 96: 276-287.
Smith, D. L. 1959. Geminitheca scotica gen. et sp. nov.: a pteridosperm from the Lower Carboniferous of Dunbartonshire. Ann. Bot. N.S. 23: 477-491.
- - . 1962a. The spores of Alcicornopteris hallei Walton. Ann. Bot. N.S. 26: 267-277. - - . 1962b. Three fructifications from the Scottish Lower Carboniferous. Palaeontology
5: 225-237. Stidd, B. M. 1978a. The synangiate nature of Dolerotheca. Am. J. Bot. 65: 243-245.
�9 1978b. An anatomically preserved Potoniea with in situ spores from the Penn- sylvanian of Illinois. Am. J. Bot. 65: 677-683.
- - a n d J. W. Hall. 1970a. Callandrium callistophytoides, gen. et sp. nov., the probable pollen-bearing organ of the seed fern, Callistophyton. Am. J. Bot. 57: 394--403.
- - a n d . 1970b. The natural affinity of the Carboniferous seed, Callospermarion. Am. J. Bot. 57: 827-836.
- - , G. A. Leisman, and T. L. Phillips. 1977. Sullitheca dactylifera gen. et sp. n.: A new medullosan pollen organ and its evolutionary significance. Am. J. Bot. 64: 994- 1002.
Stockmans, F. and Y. Willii~re. 1952-1953. V6getaux Namuriens de la Belgique. Assoc. I~tude Paldontol. Stratigr. Houill., Bruxelles. Publ. 13. Atlas (1952): 57 pl., Texte (1953): XI-382.
- - a n d - - . 1958. Un beau toit ~. vdg6taux au si/~ge 'Ties taux" des charbonnages de Monceau-Fontaine (zone de Genk, pattie sup6rieure). In B. M. Aderca, Y. Willi/~re, F. Demanet, A. Pastiels, J. Scheere, and R. van Tassel, l~tude geologique du Bassin Houiller de Charleroi. Assoc. l~tude Paldontol. Stratigr. Houill., Bruxelles, Publ. 34, 210 pp.
- - and . 1961. V6g6taux du Westphalien A de la Belgique. Graines, inflores- cences et synanges. Centre Nat. G6ol. Houill., Bruxelles. Publ. 4, 118 pp.
Taylor, T. N. 1971. Halletheca reticulatus gen. et sp. n.: A synangiate Pennsylvanian pteridosperm pollen organ. Am. J. Bot. 58: 300-308.
- - . 1978. The ultrastructure and reproductive significance of Monoletes (Pteridosper- males) pollen. Can. J. Bot. (In press.)
- - . 1979. Ultrastructural studies of pteridosperm pollen: Nanoxanthipollenites. Rev. Palaeobot. Palynol. 56:3105-3118.
Vaflier, A. 1901. Etude g6ologique et paleontologique du Carbonif/~re inf~rieur du Macon- nais, Ann. Univ. Lyon, Nouv. Ser., 1 (pt. 7): 159 pp.
Walton, J. 1931. Contributions to the knowledge of Lower Carboniferous plants. 3. On

PALEOZOIC SEED FERNS 375
the fossil-flora of the Black Limestones in Teilia Quarry, Gwaenysgor, near Prestatyn, Flintshire, with special reference to Diplopteridium teilianum Kidston sp. (gen. nov.) and some other fern-like fronds. Philos. Trans. R. Soc. London 219B: 347-379.
�9 1949. A petrified example of Alcicornopteris (A. Hallei sp. nov.) from the Lower Carboniferous of Dunbartonshire. Ann. Bot. N.S. 13: 445452.
�9 1957. On Protopitys (G6ppert): With a description of a fertile specimen -Protop- itys scotica'" sp. nov. from the Calciferous Sandstone Series of Dunbartonshire. Trans. R. Soc. Edinburgh 63: 333-340.
Whittlesey, C. 1849. Description of a coal plant supposed to be new. Am. J. Sci., ser. 2, 8: 375-377.
Zeiller, R, 1883. Fructifications de foug~res du terrain houiller. Ann. Sci. Nat. Bot. set. VI, 16: 177-207.
- - . 1899. I~tude sur la fiore fossile du bassin houiller d'Heracl6e (Asie Mineura). M6m. Soc. G6ol. Fr. Pal6ontol. 21:91 pp.
Copyright © 2022 FDOKUMEN




















